 Kun Jia1
Kun Jia1 Huhu Wang
Huhu Wang- 1National Center of Meat Quality and Safety Control, Nanjing Agricultural University, Nanjing, China
- 2Jiangsu Collaborative Innovation Center of Meat Production and Processing, Quality and Safety Control, Nanjing Agricultural University, Nanjing, China
Salmonella has emerged as a well-recognized food-borne pathogen, with many strains able to form biofilms and thus cause cross-contamination in food processing environments where acid-based disinfectants are widely encountered. In the present study, RNA sequencing was employed to establish complete transcriptome profiles of Salmonella Enteritidis in the forms of planktonic and biofilm-associated cells cultured in Tryptic Soytone Broth (TSB) and acidic TSB (aTSB). The gene expression patterns of S. Enteritidis significantly differed between biofilm-associated and planktonic cells cultivated under the same conditions. The assembled transcriptome of S. Enteritidis in this study contained 5,442 assembled transcripts, including 3,877 differentially expressed genes (DEGs) identified in biofilm and planktonic cells. These DEGs were enriched in terms such as regulation of biological process, metabolic process, macromolecular complex, binding and transferase activity, which may play crucial roles in the biofilm formation of S. Enteritidis cultivated in aTSB. Three significant pathways were observed to be enriched under acidic conditions: bacterial chemotaxis, porphyrin-chlorophyll metabolism and sulfur metabolism. In addition, 15 differentially expressed novel non-coding small RNAs (sRNAs) were identified, and only one was found to be up-regulated in mature biofilms. This preliminary study of the S. Enteritidis transcriptome serves as a basis for future investigations examining the complex network systems that regulate Salmonella biofilm in acidic environments, which provide information on biofilm formation and acid stress interaction that may facilitate the development of novel disinfection procedures in the food processing industry.
Introduction
Salmonellosis is a worldwide public health issue due to its increasing hospitalization and mortality rates in many countries (Scallan et al., 2013; Ziebell et al., 2017). The genus Salmonella comprises more than 2,500 serovars that have been associated with animal or human infection, and Salmonella enterica subspecies enterica serovar Enteritidis is the most common serovar in food outbreaks and infection (Carrique-Mas et al., 2008; Vieira et al., 2009; Sangal et al., 2010; Ziebell et al., 2017). According to the 2016 EFSA-ECDC annual report, S. Enteritidis was confirmed in 31,829 (45.7%) cases of salmonellosis in humans in 2015 (EFSA-ECDC, 2016), and the majority of salmonellosis is caused by S. Enteritidis in most developed countries (Ashton et al., 2016).
Salmonella can colonize processing surfaces and equipment such as stainless steel, marble and granite, and traditional cleaning and sanitation procedures may not be able to eradicate Salmonella from such surfaces (Rodrigues et al., 2011; Arguello et al., 2012). Salmonella is usually present on the processing surfaces of food equipment in the form of a biofilm, which can enable cells to cope with harsh conditions, in particular the Typhimurium and Enteritidis serotypes (Simões et al., 2010; Finn et al., 2013; Wang et al., 2015), and cells present in the biofilm matrix are more resistant to routinely used disinfectants than their planktonic cells (Joseph et al., 2001; Soni et al., 2013). Recent studies have shown that Salmonella species are able to survive under various environmental conditions, being found on poultry, meat, melons and grains as well as in the food processing industry (Bower and Daeschel, 1999; Stepanovi et al., 2004).
Many studies have assessed the responses of planktonic bacteria or formed-biofilms to environmental stresses such as acid, sodium chloride, starvation, and heat (Scher et al., 2005; Rodrigues et al., 2011; O'Leary et al., 2015; Philips et al., 2017). Previous studies have shown that acid tolerance, an adaptive system organisms use to respond to mild acidic conditions, can enable Salmonella to survive more extreme acidic conditions (Foster and Hall, 1990; Foster, 1995). It has been demonstrated that Salmonella Typhimurium cells exposed to mild acidic conditions of pH 4.5–5.8 can survive more extreme acidic conditions through the acid tolerance response (ATR) as time goes by Foster (1995), Bang et al. (2000). Wang et al. (2016) found that long-term acid stress obviously inhibited the biofilm formation of S. Enteritidis within a pH range of 5.0–7.2 compared with a control group (pH 7.2); however, no difference was observed in planktonic cell numbers. Several studies have focused on the resistance of biofilm formation to acid stress at different time points, such as short-term or long-term acid stress (Kubota et al., 2008; Chen et al., 2015; Wang et al., 2016). However, the molecule mechanisms underlying the gene expression networks that function during biofilm formation remain unclear, in particular the small non-coding RNAs (sRNAs). Whether the sRNAs of S. Enteritidis were involved in the stress tolerance or the regulation of stress response is not clear. Therefore, a deeper understanding of these mechanisms is necessary for the development of effective biofilm removal methods in the food industry.
High-throughput deep sequencing, sequencing hundreds of thousands to millions of DNA molecules at one time, is one of the latest techniques used to investigate the global gene expression profiles of pathogenic bacteria in response to environmental stress (Karavolos et al., 2008; Deng et al., 2012; Amin et al., 2016; Beaubrun et al., 2017; Philips et al., 2017). A number of studies have evaluated the resistance of Salmonella planktonic cells to various stresses, in particular of Typhimurium serovar (Mühlig et al., 2014; Amin et al., 2016; Ryan et al., 2016), however, few study have focused on the transcriptome of biofilm formed by Enteritidis serovar exposed to acid stress due to the incomplete reference genome. Therefore, the goal of this study was to study the global gene expression profiles of mature biofilms formed by S. Enteritidis on a stainless steel surface by RNA-seq in acidic TSB (pH = 6.0 TSB) and regular culture medium (pH = 7.2 TSB) compared with planktonic cells.
Materials and Methods
Bacterial Strain and Culture Conditions
The S. Enteritidis (NMC1461) strain used in this study was isolated from meat processing surfaces in 2014 and was stored in National Center of Meat Quality and Safety Control in China. The strain was stored frozen (−80°C) in 40% glycerol before utilization and was twice resuscitated by cultivation in tryptic soy agar (TSA, Luqiao Technology Co. Ltd., Beijing, China). A single colony of the strain was then inoculated into Tryptic Soytone Broth (TSB, Luqiao Technology Co. Ltd., Beijing, China) at 37°C for 18 h, and cells from the stationary phase were harvested by centrifugation at 10,000 × g for 10 min at 4°C, washed twice with 10 mL of an 0.85% (w/v) NaCl solution and finally re-suspended in an 0.85% NaCl solution (ca. 108 CFU/ml) and used as the inoculum for the biofilm formation assay.
Biofilm Formation and Acid Treatment
Food grade stainless steel (GSS) coupons (75 × 25 × 1 mm, 304 type, 2b finish, Shunfen Stainless Steel Material Co. Ltd., Tongnan, China) were the non-biological substrates used for biofilm development. Prior to use, the coupons were washed with acetone (overnight) as previously described (Giaouris et al., 2013). To produce biofilms, 20 cleaned GSS coupons were placed in a glass box to ensure that the upper part of each coupon (ca. 12 mm) was exposed to the liquid-air interface, which provides a good environment for biofilm formation. The glass box containing the GSS coupons was then autoclaved at 121°C for 15 min. Sterilized and cooled TSB (pH = 7.2) or acidic TSB (pH = 6.0) (230 ml) with a 230 μl (108 CFU/ml) aliquot of a cell suspension, prepared as described above, was transferred into the cooling glass box containing 20 GSS coupons. The inoculated box was incubated at 20°C for 3 days without any dynamic conditions to facilitate biofilm development on the coupons, and the planktonic cells (grown without stainless steel in TSB or aTSB fluid) were incubated in the same manner. A 0.1 M phosphate buffer solution was added to the TSB to maintain a pH of 6.0 for the acid stress group. Non-attached cells were removed by rinsing the coupons three times with a 0.85% NaCl solution, whereas the attached biofilm cells were detached from the coupons using a violent water-flapping motion that has been proven to effectively remove cells attached to coupons (Wang et al., 2015). The coupons were transferred into a stomacher blender bag and shaken at a speed of 200 oscillations/min for 2 min using a bag mixer (BagMixer 400VW, Interscience) to detach the cells from the coupons. The planktonic cells were harvested by centrifugation (10,000 × g, 15 min) at 4°C, whereas the biofilm cells were harvested by passage through a filter with a pore diameter of 0.22 μm pore.
Total RNA Isolation, Library Construction, and Sequencing
Total RNA for Illumina sequencing was extracted from the biofilm cells (TB, aTB) and the planktonic cells (TF, aTF) prepared as described above using a Trizol Bacterial RNA Isolation Kit (Thermo Fisher) according to the manufacturer's protocol. During RNA isolation, DNA was removed by treating the samples with RNase-free DNase (Solarbio) for 30 min at 37°C to avoid DNA contamination. The purity, concentration and integrity of the RNA was examined using a NanoPhotometer spectrophotometer (IMPLEN, CA, USA), Qubit 2.0 fluorometer (Life Technologies, CA, USA) and an Agilent Bioanalyzer 2100 system (Agilent Technologies, CA, USA). The total RNA in each sample used for library construction had to pass quantity and quality control. The RNA integrity number (RIN) values of all the samples used for RNA-seq were >6.0 (Schroeder et al., 2006).
A total of 3 μg of each RNA sample was used as the input material for the RNA sample preparations, and three biological replicates were analyzed for a total of 12 (3 × 4) samples. The sequencing libraries, in which index codes were added to attribute sequences to each sample, were constructed and sequenced using an NEBNext Ultra™ Directional RNA Library Prep Kit for Illumina (NEB, USA) according to the manufacturer's recommendations. Briefly, mRNA was enriched from total RNA via poly-T oligo-conjugated magnetic beads by depleting rRNA and then fragmented into short fragments using an RNA fragmentation reagent (Life Technologies). First-strand cDNA was synthesized from these cleaved mRNA fragments using random hexamer primers and M-MuL V Reverse Transcriptase (RNaseH-), followed by the synthesis of second-strand cDNA using DNA Polymerase I and RNase H in the presence of dATP, dGTP, dCTP, and dTTP. Exonuclease and polymerase were used to blunt and adenylate the 3′ ends of DNA fragments, and NEBNext Adaptors with hairpin loop structures were ligated to the fragments in preparation for hybridization. The library fragments were purified using the AMPure XP system (Beckman Coulter, Beverly, USA) to preferentially select cDNA fragments 150–200 bp in length. Next, the purified cDNA fragments were enriched via PCR, the products were purified (AMPure XP system), and library quality was assessed on the Agilent Bioanalyzer 2100 system. Finally, the cDNA libraries were sequenced using the Illumina HiSeq 2500 platform, and 150 bp paired-end reads were generated.
Reads Alignment and Analysis of Differential Gene Expression
The raw reads were filtered by removing low-quality sequences to obtain clean reads according to the following criteria: (1) removing reads containing poly-N; (2) removing reads containing adapters; and (3) removing low-quality reads. At the same time, the Q20 and Q30 and the GC content of the clean data were calculated to evaluate the sequencing quality, and all the downstream analyses were performed using high-quality clean data. The clean reads were then mapped to the S. Enteritidis strain SEJ complete genome (NCBI reference sequence: NZ_CP008928.1) using Bowtie2-2.2.3 (Langmead and Salzberg, 2012). The mapping parameters were set as follows: (1) clean reads total mapped to the reference genome, >70%; (2) clean reads multiple mapped to the reference genome, <10%; and (3) clean reads uniquely mapped to the reference genome, >90%.
To estimate the abundance of each unigene, the FPKM (expected number of Fragments Per Kilobase of transcript sequence per Millions of base pairs sequenced) method was used, which considers the effect of gene length and sequencing depth for the read count at the same time and is currently the most common method used to calculate the log2 (FPKM biofilm/FPKM planktonic) fold change. Differential expression analysis of mature biofilm/planktonic cells (three biological replicates per condition) was performed using the DEGSeq R package (1.18.0), which provided statistical routines to identify differentially expressed genes (DEGs) between the mature biofilm and planktonic cells using a model based on the negative binomial distribution with an absolute value of log2 (fold change) >1 and an adjusted P-value threshold of <0.5. According to the gene expression profiles, the Pearson correlation coefficients among the 12 samples were calculated.
Go and KEGG Enrichment Analysis of Differentially Expressed Genes
The DEGs were used for Gene Ontology (GO) and KEGG (Kyoto Encyclopedia of Genes and Genomes) enrichment analyses to help further understand the biological functions of the identified genes and significantly enriched metabolic pathways. In this study, both GO terms and KEGG pathways with adjusted P-value < 0.05 were considered significantly enriched in DEGs.
Quantitative RT-PCR Analysis
Several genes that showed significant up- or down-regulation were selected for quantitative reverse transcription-PCR (qRT-PCR) validation to determine if gene expression was consistent between qRT-PCR and RNA-seq. Total RNA was isolated as described above, followed by cDNA synthesis using reverse transcriptase. Quantitative PCR (qPCR) was carried out for each sample using SYBR green (Prime Script RT Master Mix; TAKARA, China) methodology and the 7,500 Fast Real-time PCR System (Applied Biosystems, Foster, USA) with a total of 500 ng of RNA as the starting material. The 16S DNA of Salmonella was used as an internal gene to normalize the expression of the tested genes. The primers used in this study are listed in supporting information Table S1. Melting curve analysis was used to validate the specificity of primers, and the ΔΔCT method was used to calculate the relative gene expression. The expression level of each target gene was compared with the 16S DNA internal control gene using the 2−ΔΔCt method, and each gene was analyzed at least four times.
Results
RNA Sequencing and Preliminary Analysis of the Raw Data
To obtain a comprehensive overview of the transcriptome profile of Salmonella biofilms in response to acid stress, RNA was extracted from four different groups, TB, TF, aTB, and aTF, with three independent biological replicates per group. The RNA used for library construction had to pass quantity and quality control criteria. Twelve libraries were constructed from the four different groups, and raw reads were produced from the 12 libraries using the Illumina sequencing platform. A total of 13–28 million reads per sample were generated by RNA-seq (Table 1). Clean reads were harvested with strict quality control criteria and data filtration, and the average numbers of total clean reads in the four libraries were 22,568,103 (TB), 15,179,200 (aTB), 17,549,117 (TF), and 15532433 (aTF). These reads were mapped to the S. Enteritidis strain SEJ complete reference genome. Altogether, 91.41% (TB), 92.86 (aTB), 95.11% (TF), and 90.70% (aTF) of the clean reads were uniquely mapped to the reference genome. Under normal circumstances, if the reference genome is appropriate and there is no contamination in the relevant experiments, the percentage of mapped sequences generated by the experiment should be higher than 70%, with multiple mapped reads accounting for <10%. In this study, over 90% of the clean reads for each sample were mapped to the reference genome, with multiple mapped reads accounting for <5%. Additionally, biological repetition is essential to any biological experiment, and in high-throughput sequencing, the square of the Pearson correlation coefficient should be larger than 0.8; if this is not the case, the reason should be determined or the experiment should be repeated. As shown in supporting information Figure S1, the correlation coefficient diagram illustrates the relationship among the four treatments, with an average coefficient (R2) of 0.989, 0.981, 0.985, and 0.986 for TB, TF, aTF, and aTB, respectively. These results indicated that the reference genome in our study was very suitable and that the basic RNA-seq data could be used for subsequent bioinformatics analysis.
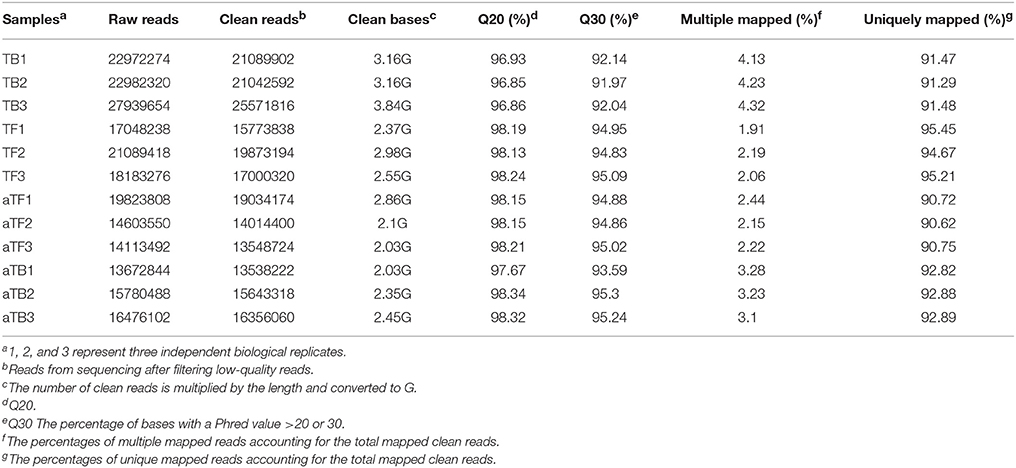
Table 1. RNA-seq data statistics.
Differentially Expressed Genes (DEGS) Under Acid Stress
The RNA-seq data were used to evaluate differences in gene expression, and the FPKM was calculated to quantify the expression levels of all genes in the four groups, TB, TF, aTB, and aTF. The DEGs were determined using the DESeq R package with the criteria of log2 (fold change) >1 and adjusted P-value <0.05. There were 3,162 genes that were significantly differentially expressed between aTB and aTF, including 1,546 up-regulated genes and 1,616 down-regulated genes (Figure 1A). A total of 2,992 genes were identified between TB and TF, with 1,457 genes up-regulated and 1,535 genes down-regulated (Figure 1B). After acid treatment, 2,303 DEGs were identified in aTFF (aTF vs. TF), and 1,831 DEGs were identified in aTBB (aTB vs. TB). The numbers of DEGs showing up or down-regulation are shown for aTFF (Figure 1C) and aTBB (Figure 1D). The DEGs were classified using a Venn diagram (Figure 1E) which indicated that of the 3,162 DEGs in aTBF and 2,992 DEGs in TBF, 535 genes were specifically altered in TBF, 703 genes were specifically altered in aTBF, and 2,445 genes were differentially expressed in both TBF and aTBF. The DEGs identified in both TBF and aTBF likely play an important role in biofilm formation. Furthermore, there were also 1,103 DEGs identified in aTFF and aTBB, which provided evidence that these DEGs may be induced in an acidic environment and may be essential for the response to acid stress in Salmonella.
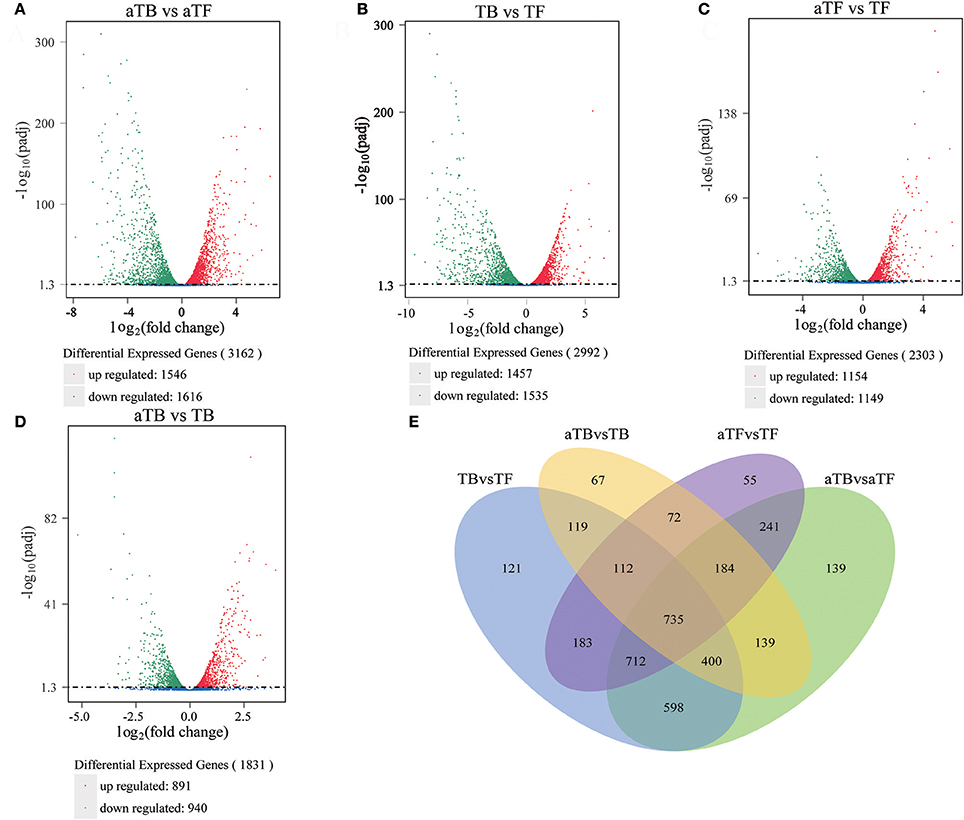
Figure 1. Global comparison of DEGs in different groups: (A) Volcano plot of DEGs in aTBF. (B) Volcano plot of DEGs in TBF. (C) Volcano plot of DEGs in aTFF. (D) Volcano plot of DEGs in aTBB. (E) Venn diagram of DEGs identified in different groups.
A heat map and hierarchical clustering were used to determine the expression patterns of different genes under different experimental conditions, and genes with similar expression patterns may have similar functions or may be involved in the same metabolic process or cellular pathway. Therefore, the genes with the same or similar expression patterns were clustered into classes to predict the function of unknown genes or new functions of known genes. In this study, FPKM was used to conduct the hierarchical clustering analysis of transcript abundance, and 3,877 DEGs across the four different groups were analyzed. The heat map and hierarchical clustering demonstrated that the TF groups showed similar transcriptome profiles to the aTF groups and that the TB groups were similar to the aTB groups (Figure 2). The biofilm-associated groups (TB, aTB) and planktonic cells groups (TF, aTF) were distinctly separated, and the differentially expressed mRNAs in each group were clustered into two types, one with higher expression in the biofilm-associated groups but lower expression in the planktonic cells groups, and the other with lower expression in the biofilm-associated groups but higher expression in the planktonic cells groups.
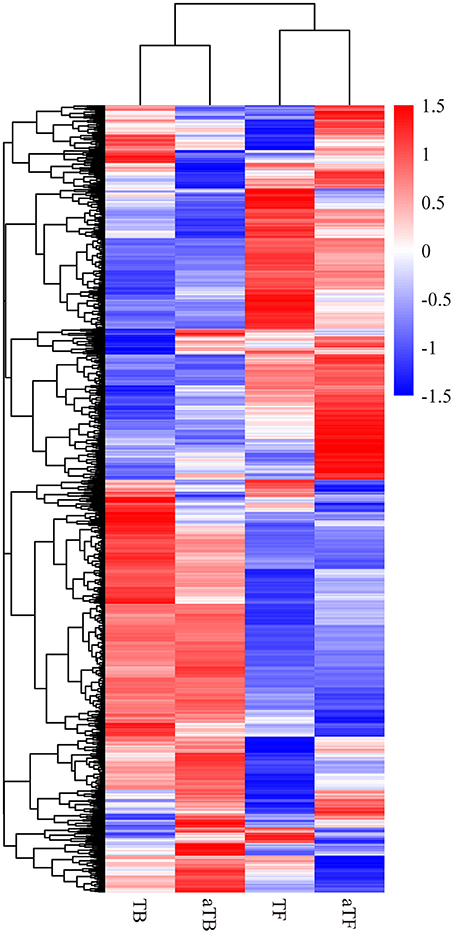
Figure 2. Heatmap and hierarchical clustering of the differentially expressed mRNAs in the four groups: Note: Red indicates a higher mRNA expression level, and blue indicates a lower mRNA expression level. The color from red to blue indicates log10 (FPKM+1) from large to small.
Functional Annotation of the Transcriptome
Gene Ontology (GO) enrichment analysis of DEGs was performed to functionally analyze the S. Enteritidis transcriptome according to NR annotation. The GO terms from hierarchical vocabularies describing biological processes, cellular components and molecular functions for analyzing transcripts were assigned using Blast2GO software (Conesa and Götz, 2008). Among the 3,697 genes in aTBF, 2,290 DEGs were assigned to the biological process, cellular component and molecular function categories. These DEGs were assigned to many GO terms, and we selected the 30 most enriched GO terms with p-values < 0.05, which are displayed in Figure 3A. The terms “amino acid transport,” “amino acid transmembrane transport,” “macromolecular complex,” “transferase activity, transferring one-carbon groups,” and “iron-sulfur cluster binding” were the most highly represented. Among the 3,697 genes in TBF, 2139 DEGs were assigned to the three GO categories (Figure 3B). The 30 most enriched functional categories included “nitrogen compound metabolic process,” “cellular aromatic compound metabolic process,” “signaling,” “macromolecular complex” and “sulfur compound binding.” There were three biological process terms (metabolic process, biosynthetic process and signaling), one cellular component term (macromolecular complex) and three molecular function terms (binding, methyltransferase activity and transferase activity) that exhibited alterations in both aTBF and TBF, and these categories are likely associated with biofilm formation. As shown in Figure 3C, regulation of biological process played a very important role in the response of S. Enteritidis to acid stress, while none of the cellular component terms were significant enriched. The molecular functions of binding, receptor activity and signal transducer activity were also associated with acid stress compared with TF. The top 30 functional categories included acid transport, carboxylic acid transport, membrane part, membrane and transporter activity (Figure 3D).
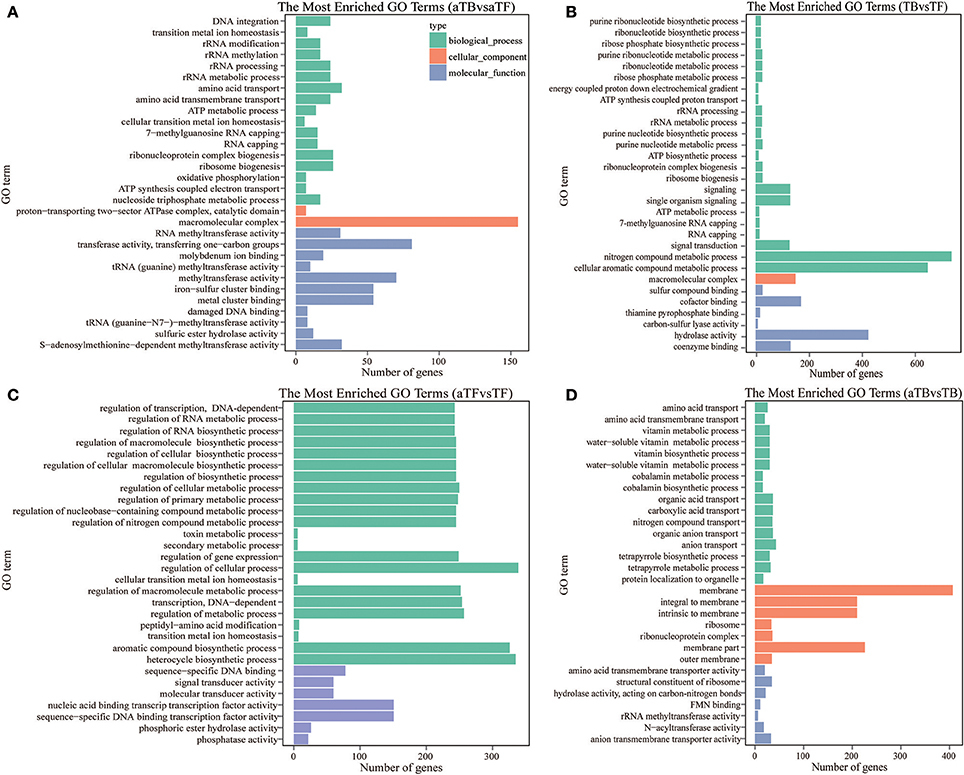
Figure 3. Blast2GO annotation of differentially expressed genes assigned to three GO categories (p-value below 0.05). (A) The Top 30 Most Enriched Go Terms in aTBF. (B) The Top 30 Most Enriched Go Terms in TBF. (C) The Top 30 Most Enriched Go Terms in aTFF. (D) The Top 30 Most Enriched Go Terms in aTBB.
The KOBAS software was used to test the statistical enrichment of DEGs in KEGG pathways to assess the reliability of our transcriptome results and our annotation process. The 2,139 DEGs in TBF and 2,290 DEGs in aTBF were mapped to 90 different KEGG pathways. However, no significantly enriched pathways were identified in the two groups by KEGG enrichment analysis (p < 0.05). As shown in Table 2, three significantly enriched pathways (p < 0.05) were identified in aTFF (1 pathway) and aTBB (2 pathways). In the aTBB group, there were a total of 28 DEGs involved in porphyrin and chlorophyll metabolism and 27 DEGs involved in sulfur metabolism. Additionally, a total of 21 DEGs in aTFF were significantly enriched in bacterial chemotaxis. These three significantly enriched pathways may be associated with the acid tolerance of S. Enteritidis.
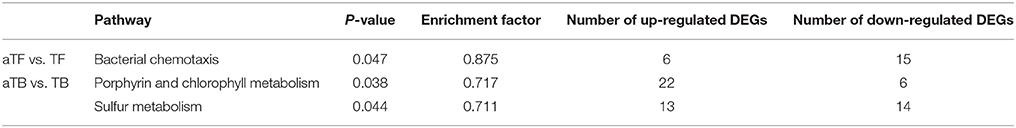
Table 2. The numbers of DEGs significantly enriched in KEGG pathways.
Fifteen differentially expressed small non-coding RNAs (sRNAs) were identified in the transcriptomes of the two biofilm-associated groups (Table 3); 11 sRNAs were identified in the TBF group, 10 sRNAs were identified in the aTBF group, and six sRNAs were identified in both groups. Most of the 15 sRNAs were located on the negative DNA strand (Minus), and only three sRNAs (sRNA314, sRNA282, and sRNA290) were located on the positive DNA strand (Plus). Additionally, 10 of the 15 sRNAs possessed a simple sRNA structure predicted viaMfold (HP), and only five possessed more than one hairpin (HS).
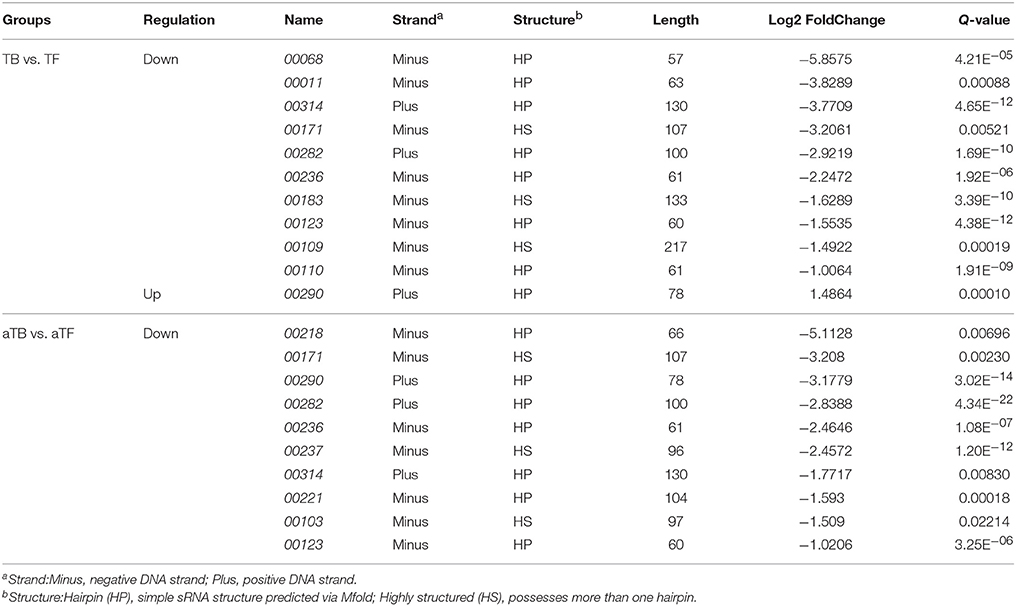
Table 3. Differently expressed novel non-coding small RNAs (sRNAs) in the transcriptomes of the two biofilm-associated groups.
Validation of DEGs
qRT-PCR was performed on 19 selected genes (8 up-regulated and 11 down-regulated) to verify the RNA-seq data in this study. As shown in Figure 4, the qRT-PCR data correlated well with the RNA-seq data in identifying gene up- or down-regulation (R2 = 0.936).
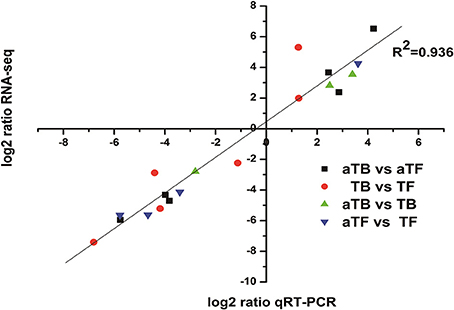
Figure 4. Correlation of gene expression measurements by RNA-seq and qRT-PCR for 19 selected genes. Fold change values were converted to log2 values for comparison.
Discussion
Biofilm formation and the bacterial response to acid stress are governed by many regulators, not just several specific genes (Quinn et al., 2014; Wang et al., 2015; Anbalagan et al., 2016). However, the transcriptome and regulatory network involved in biofilm formation or the bacterial stress response in S. Enteritidis are only partly known, as most recent studies have focused on the transcriptome of S. Typhimurium (Wang et al., 2015; Anbalagan et al., 2016), while few studies have focused on S. Enteritidis (Shah, 2014). Therefore, it is critical to identify the transcriptome profile of S. Enteritidis. In the present study, RNA-seq technology was used to obtain the transcriptome profile of mature S. Enteritidis biofilms on a stainless steel surface subjected to acid stress. Furthermore, we used comparative transcriptomics to compare biofilm cells with planktonic cells and found that thousands of genes were specifically up- or down-regulated. Additionally, we identified several novel sRNAs and performed functional analyses (KEGG, GO annotation). The expression levels determined by qRT-PCR were consistent with the results from RNA-seq. The following aspects associated with acid stress or biofilm formation were characterized from a global perspective.
DEGs Identified by Comparing Biofilm-Associated Cells with Planktonic Cells
Our results showed that thousands of genes were significantly up- or down-regulated in biofilm-associated cells compared with planktonic cells. Of these genes, 20 that were differentially expressed and likely had an impact on biofilm formation are shown in Table 4. The fimFH genes showed high levels of expression, and FimH protein in particular has been the focus of many studies. In a previous study, FimH protein was found to be the mannose-specific adhesin of Escherichia coli type 1 fimbriae (Krogfelt et al., 1990), and E. coli biofilm formation could be significantly reduced by inhibition of FimH (Sarkar et al., 2016). It has been shown that the minor fibril components FimF, FimG, and FimH are involved in receptor binding, which is important for the initial attachment. In the present study, the expression change (more than 3-fold) of fimFH in TBF was similar to that observed in aTBF, showing significant up-regulation in biofilm-associated cells. A similar finding has also been observed by Pratt and Kolter (1998), who presented evidence that FimH protein was required for initial surface attachment. Compared with planktonic cells, the prpDE gene involved in propanoate metabolism was highly expressed in biofilm cells in our study. Consistent with our result, prpBCDE expression was found to be higher in biofilm-associated cells of E. coli O157:H7 compared with planktonic cells (Lee et al., 2011). Additionally, Lee et al. (2007) suggested that prpCDE, which induces E. coli biofilm formation, is involved in propanoate metabolism. The regulation of the prp operon is complex due to the transcriptional regulators, such as PrpR, the σ54 factor RpoN, and IHF, that are required to activate the transcription of prpBCDE (Simonte et al., 2017). The RNA polymerase sigma factor RpoS, which is required for virulence, stress resistance and biofilm formation, was found to be up-regulated in biofilm-associated cells in this study. RpoS can bind to the csgD promoter and has an important impact on the transcription of csgD, which is required for the formation of biofilms and production of cellulose by regulating the aggregation and motility of cells (Gerstel et al., 2003; Jonas et al., 2007). In addition, hundreds of hypothetical proteins were differentially expressed in both TBF and aTBF, indicating that they are likely important in biofilm formation. The differentially expressed hypothetical proteins are shown in Table 4.
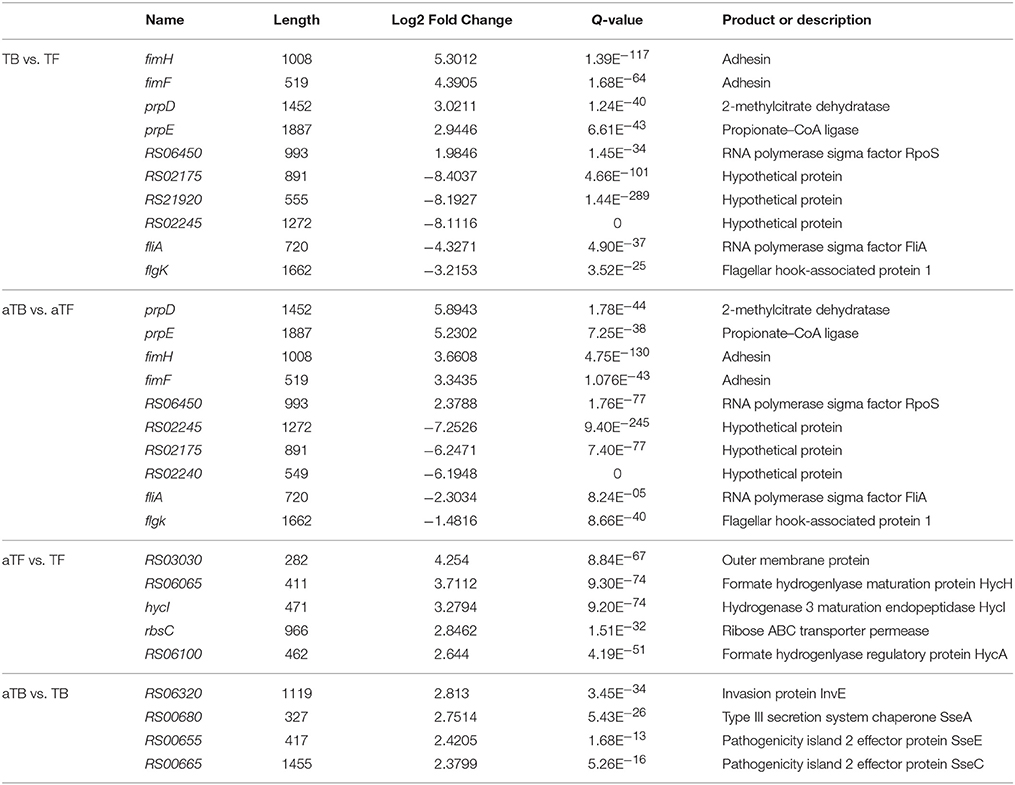
Table 4. DEGs (up- or down-regulated) in different groups.
Up-Regulated Genes in Response to Acid Stress
As shown in Table 4, genes associated with formate hydrogenlyase, (such as RS06065, HycI, RS06100, HycE, and HycE), ABC transporter permease (rbsC) and outer membrane proteins (RS03030) were up-regulated in aTF compared with TF. The HycADEHI genes were up-regulated, with a log2 fold change of more than 2 in aTF compared with TF. The Hyc genes, which are involved in the cellular metabolism of S. Typhimurium, can remove excess reductant generated during acid stress by producing H2 and consuming protons, so the expression of hydrogenase is likely involved in the pH response and regulation (Zbell and Maier, 2009; Noguchi et al., 2010). HycA is thought to regulate the Hyc operon, HycD is a transmembrane protein, HycE is the Ni-containing large subunit of Hyc-hydrogenase, and HycI is known to play an important role in the maturation of the formate hydrogenlyase (FHL) complex (Lamichhane-Khadka et al., 2015). In the present study, the HycADEHI genes were up-regulated in aTF, with log2 fold changes of 2.1 to 3.7, and all of these genes are associated with hydrogenase. Consistent with our results, Noguchi et al. (2010) observed that hydrogenase-3, known as a part of the FHL complex, contributed to acid resistance of anaerobic cultures by consuming protons. A similar result was found by Hayes et al. (2006), who suggested that hydrogenase plays an important role in acid resistance in partly aerated cultures. A number of genes associated with ABC transporters showed significant up- or down-regulation in aTF compared with TF, reflecting the changing requirements for cells when encountering an acidic environment. The D-ribose transporter gene rbsC is one such gene, showing significant up-regulation under aTF in this study. A previous study showed that molecular pumps, characterized as ATP-binding cassette (ABC) transporters, translocate solutes across the membrane by utilizing the energy of ATP hydrolysis (Oldham et al., 2013), and several other bacterial ABC transporters changed significantly after acid shock (Bore et al., 2007). Additionally, levels of the rbsABCD operon, which encodes D-ribose transporter components, were up-regulated (7.0–14.96-fold) during artificial gastric fluid stress (Sun et al., 2014).
Of particular interest were genes associated with virulence in the present study, whose levels were mostly up-regulated under aTB compared with TB. As shown in Table 4, the protein InvE, which is also encoded within SPI-1, is necessary for translocating bacterial proteins into host cells and dramatically up-regulated in aTB in this study, with a log2 fold change of 2.8. Quinn et al. (2014) found that when S. Typhimurium was subjected to acid stress, which is induced for optimal expression of its virulence genes. In contrast to our results, the invE gene was down-regulated in lysogenic S. Typhimurium treated at pH 3, 4, and 5 for a short time (Kim et al., 2014). In addition, several proteins associated with virulence, such as SseA, SseE and SseC, were over-expressed in aTB compared with TB. Several studies have shown that virulence factors (called effectors), which are the matrix of the secretory device, encoded by SPI-2 play an important role in the pathogenicity of Salmonella (Li et al., 2015; Sun et al., 2016). The secretion activity of SPI-2 is probably induced in response to an acidic environment, and Coombes et al. (2004) demonstrated that the expression of SPI-2-encoded effectors is acid-regulated. In our study, the expression of most effector proteins was up-regulated under aTB. Consistent with our results, higher expression of genes associated with virulence and invasion was found during biofilm formation at pH 5 compared with pH 7 (O'Leary et al., 2015).
Prediction of sRNAs Differentially Expressed in Biofilm-Associated Cells
According to our RNA-seq data, a total of 88 candidate sRNAs identified in our study need to be further investigated. Of note, only 15 of these sRNAs were differentially expressed (|log2 fold change| > 1) under TBF and aTBF, and only one of the sRNAs showed up-regulation in biofilm-associated cells compared with planktonic cells. Small RNAs (sRNAs), short non-coding RNA sequence with transcript sizes ranging from 50 to 500 nucleotides, are ubiquitous regulators that play an important role in many cellular activities, affecting the translation of mRNAs by imperfect base pairing with the help of Hfq (Huang et al., 2009; Storz et al., 2011). Large numbers of sRNAs acting as major posttranscriptional regulators are induced in different physiological environments. Jørgensen et al. (2013) observed that CsgD is linked to biofilm formation by regulating the transcription of curli biogenesis under the control of multiple sRNAs (GcvB, RprA, OmrA, OmrB). In our study, to determine whether our 88 predicted sRNAs resemble known bacterial sRNAs, we aligned the sequences of our sRNAs to sequences in 3 non-coding RNA databases: Rfam, sRNATarBase and BSRD (Amin et al., 2016). To our surprise, only 10 of our 88 candidate sRNAs aligned significantly with known sRNAs (supporting information: Table S2), including RyfA, GcvB, Sib, and GlmZ, via comparative analyses with the chromosomes of Salmonella, E. coli and Shigella flexneri with the help of Hfq RNA binding protein, and none of the 15 differentially expressed sRNAs exhibited significant identity to known sRNAs, likely because this is the first report to investigate the sRNAs associated with biofilm formation in S. Enteritidis. Importantly, this means that 78 of the 88 sRNAs in our study are novel and characteristic in S. Enteritidis. Additionally, six sRNAs were down-regulated in both TBF and aTBF, which indicated that these six sRNAs probably play prominent role in biofilm formation. However, this hypothesis needs to be further validated using gene knockout technology, such as λ-red homologous recombination and CRISPR-Cas9 (Song et al., 2015).
Conclusion
Salmonella biofilm is considered to be a common source of cross-contamination, posing a serious threat to food factories worldwide where various environmental stresses are encountered. The RNA-seq data in this study provide insight into the gene expression patterns, gene functional categories and significant pathways associated with acid stress or biofilm formation. Furthermore, the list of 15 novel differentially expressed sRNAs predicted in this study provides candidates for further studies. This global analysis of the S. Enteritidis transcriptome revealed new candidate genes that will increase our knowledge of the S. Enteritidis response to acid stress and may suggest ways to prevent biofilm cross-contamination, bringing us one step closer to uncovering the complex gene networks and molecular mechanisms that involved in the regulation of mature biofilms or acid stress.
Author Contributions
HW and XX designed research; KJ, GW, LL, HW, and MW performed research; KJ and HW wrote the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by the project funded by the grants from the National Natural Science Foundation of China (31501574), the Fundamental Research Funds for the Central Universities (KJQN201650) and the Natural Science Foundation of Jiangsu Province in China (BK20150678).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01861/full#supplementary-material
References
Amin, S. V., Roberts, J. T., Patterson, D. G., Coley, A. B., Allred, J. A., Denner, J. M., et al. (2016). Novel small RNA (sRNA) landscape of the starvation-stress response transcriptome of Salmonella enterica serovar typhimurium. RNA Biol. 13, 331–342. doi: 10.1080/15476286.2016.1144010
Anbalagan, K., Chinni, V., and Phua, K. (2016). Transcriptome analysis of Salmonella enterica subspecies enterica serotype Typhi Biofilm. Int. J. Infect. Dis. 45, 133. doi: 10.1016/j.ijid.2016.02.329
Arguello, H., Carvajal, A., Collazos, J. A., García-Feliz, C., and Rubio, P. (2012). Prevalence and serovars of Salmonella enterica on pig carcasses, slaughtered pigs and the environment of four Spanish slaughterhouses. Food Res. Int. 45, 905–912. doi: 10.1016/j.foodres.2011.04.017
Ashton, P. M., Nair, S., Peters, T. M., Bale, J. A., Powell, D. G., Painset, A., et al. (2016). Identification of Salmonella for public health surveillance using whole genome sequencing. PeerJ. 4:e1752. doi: 10.7717/peerj.1752
Bang, I. S., Kim, B. H., Foster, J. W., and Park, Y. K. (2000). OmpR regulates the stationary-phase acid tolerance response of Salmonella enterica serovar Typhimurium. J. Bacteriol. 182, 2245–2252. doi: 10.1128/JB.182.8.2245-2252.2000
Beaubrun, J. J.-G., Tall, B. D., Flamer, M., Patel, I., Gopinath, G., Auguste, W., et al. (2017). Increased secretion of exopolysaccharide and virulence potential of a mucoid variant of Salmonella enterica serovar montevideo under environmental stress. Microb. Pathog. 103, 107–113. doi: 10.1016/j.micpath.2016.12.005
Bore, E., Langsrud, S., Langsrud, Ø., Rode, T. M., and Holck, A. (2007). Acid-shock responses in Staphylococcus aureus investigated by global gene expression analysis. Microbiology 153, 2289–2303. doi: 10.1099/mic.0.2007/005942-0
Bower, C., and Daeschel, M. (1999). Resistance responses of microorganisms in food environments. Int. J. Food Microbiol. 50, 33–44. doi: 10.1016/S0168-1605(99)00075-6
Carrique-Mas, J., Papadopoulou, C., Evans, S., Wales, A., Teale, C., and Davies, R. (2008). Trends in phage types and antimicrobial resistance of Salmonella enterica serovar Enteritidis isolated from animals in Great Britain from 1990 to 2005. Vet. Rec. 162, 541–546. doi: 10.1136/vr.162.17.541
Chen, D., Zhao, T., and Doyle, M. P. (2015). Single-and mixed-species biofilm formation by Escherichia coli O157: H7 and Salmonella, and their sensitivity to levulinic acid plus sodium dodecyl sulfate. Food Control. 57, 48–53. doi: 10.1016/j.foodcont.2015.04.006
Conesa, A., and Götz, S. (2008). Blast2GO: a comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genomics 2008:619832. doi: 10.1155/2008/619832
Coombes, B. K., Brown, N. F., Valdez, Y., Brumell, J. H., and Finlay, B. B. (2004). Expression and secretion of Salmonella pathogenicity island-2 virulence genes in response to acidification exhibit differential requirements of a functional type III secretion apparatus and SsaL. J. Biol. Chem. 279, 49804–49815. doi: 10.1074/jbc.M404299200
Deng, X., Li, Z., and Zhang, W. (2012). Transcriptome sequencing of Salmonella enterica serovar Enteritidis under desiccation and starvation stress in peanut oil. Food Microbiol. 30, 311–315. doi: 10.1016/j.fm.2011.11.001
EFSA-ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control) (2016). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 14:e04634. doi: 10.2903/j.efsa.2016.4634
Finn, S., Condell, O., McClure, P., Amézquita, A., and Fanning, S. (2013). Mechanisms of survival, responses and sources of Salmonella in low-moisture environments. Front. Microbiol. 4:331. doi: 10.3389/fmicb.2013.00331
Foster, J. W. (1995). Low pH adaptation and the acid tolerance response of Salmonella typhimurium. Crit. Rev. Microbiol. 21, 215–237. doi: 10.3109/10408419509113541
Foster, J. W., and Hall, H. K. (1990). Adaptive acidification tolerance response of Salmonella typhimurium. J. Bacteriol. 172, 771–778. doi: 10.1128/jb.172.2.771-778.1990
Gerstel, U., Park, C., and Römling, U. (2003). Complex regulation of csgD promoter activity by global regulatory proteins. Mol. Microbiol. 49, 639–654. doi: 10.1046/j.1365-2958.2003.03594.x
Giaouris, E., Samoilis, G., Chorianopoulos, N., Ercolini, D., and Nychas, G.-J. (2013). Differential protein expression patterns between planktonic and biofilm cells of Salmonella enterica serovar Enteritidis PT4 on stainless steel surface. Int. J. Food Microbiol. 162, 105–113. doi: 10.1016/j.ijfoodmicro.2012.12.023
Hayes, E. T., Wilks, J. C., Sanfilippo, P., Yohannes, E., Tate, D. P., Jones, B. D., et al. (2006). Oxygen limitation modulates pH regulation of catabolism and hydrogenases, multidrug transporters, and envelope composition in Escherichia coli K-12. BMC Microbiol. 6:89. doi: 10.1186/1471-2180-6-89
Huang, H.-Y., Chang, H.-Y., Chou, C.-H., Tseng, C.-P., Ho, S.-Y., Yang, C.-D., et al. (2009). sRNAMap: genomic maps for small non-coding RNAs, their regulators and their targets in microbial genomes. Nucleic Acids Res. 37(Suppl. 1), D150–D154. doi: 10.1093/nar/gkn852
Jonas, K., Tomenius, H., Kader, A., Normark, S., Römling, U., Belova, L. M., et al. (2007). Roles of curli, cellulose and BapA in Salmonella biofilm morphology studied by atomic force microscopy. BMC Microbiol. 7:70. doi: 10.1186/1471-2180-7-70
Jørgensen, M. G., Thomason, M. K., Havelund, J., Valentin-Hansen, P., and Storz, G. (2013). Dual function of the McaS small RNA in controlling biofilm formation. Genes Dev. 27, 1132–1145. doi: 10.1101/gad.214734.113
Joseph, B., Otta, S., Karunasagar, I., and Karunasagar, I. (2001). Biofilm formation by Salmonella spp. on food contact surfaces and their sensitivity to sanitizers. Int. J. Food Microbiol. 64, 367–372. doi: 10.1016/S0168-1605(00)00466-9
Karavolos, M., Spencer, H., Bulmer, D., Thompson, A., Winzer, K., Williams, P., et al. (2008). Adrenaline modulates the global transcriptional profile of Salmonella revealing a role in the antimicrobial peptide and oxidative stress resistance responses. BMC Genomics 9:458. doi: 10.1186/1471-2164-9-458
Kim, S., Ryu, K., Biswas, D., and Ahn, J. (2014). Survival, prophage induction, and invasive properties of lysogenic Salmonella Typhimurium exposed to simulated gastrointestinal conditions. Arch. Microbiol. 196, 655–659. doi: 10.1007/s00203-014-1005-z
Krogfelt, K. A., Bergmans, H., and Klemm, P. (1990). Direct evidence that the FimH protein is the mannose-specific adhesin of Escherichia coli type 1 fimbriae. Infect. Immun. 58, 1995–1998.
Kubota, H., Senda, S., Nomura, N., Tokuda, H., and Uchiyama, H. (2008). Biofilm formation by lactic acid bacteria and resistance to environmental stress. J. Biosci. Bioeng. 106, 381–386. doi: 10.1263/jbb.106.381
Lamichhane-Khadka, R., Benoit, S. L., Miller-Parks, E. F., and Maier, R. J. (2015). Host hydrogen rather than that produced by the pathogen is important for Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 83, 311–316. doi: 10.1128/IAI.02611-14
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lee, J., Bansal, T., Jayaraman, A., Bentley, W. E., and Wood, T. K. (2007). Enterohemorrhagic Escherichia coli biofilms are inhibited by 7-hydroxyindole and stimulated by isatin. Appl. Environ. Microbiol. 73, 4100–4109. doi: 10.1128/AEM.00360-07
Lee, J.-H., Kim, Y.-G., Cho, M. H., Wood, T. K., and Lee, J. (2011). Transcriptomic analysis for genetic mechanisms of the factors related to biofilm formation in Escherichia coli O157: H7. Curr. Microbiol. 62, 1321–1330. doi: 10.1007/s00284-010-9862-4
Li, J., Overall, C. C., Nakayasu, E. S., Kidwai, A. S., Jones, M. B., Johnson, R. C., et al. (2015). Analysis of the Salmonella regulatory network suggests involvement of SsrB and H-NS in σE-regulated SPI-2 gene expression. Front. Microbiol. 6:27. doi: 10.3389/fmicb.2015.00027
Mühlig, A., Behr, J., Scherer, S., and Müller-Herbst, S. (2014). Stress response of Salmonella enterica serovar Typhimurium to acidified nitrite. Appl. Environ. Microbiol. 80, 6373–6382. doi: 10.1128/AEM.01696-14
Noguchi, K., Riggins, D. P., Eldahan, K. C., Kitko, R. D., and Slonczewski, J. L. (2010). Hydrogenase-3 contributes to anaerobic acid resistance of Escherichia coli. PLoS ONE 5:e10132. doi: 10.1371/journal.pone.0010132
Oldham, M. L., Chen, S., and Chen, J. (2013). Structural basis for substrate specificity in the Escherichia coli maltose transport system. Proc. Natl. Acad. Sci. U.S.A. 110, 18132–18137. doi: 10.1073/pnas.1311407110
O'Leary, D., McCabe, E. M., McCusker, M. P., Martins, M., Fanning, S., and Duffy, G. (2015). Acid environments affect biofilm formation and gene expression in isolates of Salmonella enterica Typhimurium DT104. Int. J. Food Microbiol. 206, 7–16. doi: 10.1016/j.ijfoodmicro.2015.03.030
Philips, J., Rabaey, K., Lovley, D. R., and Vargas, M. (2017). Biofilm formation by Clostridium ljungdahlii is induced by sodium chloride stress: experimental evaluation and transcriptome analysis. PLoS ONE 12:e0170406. doi: 10.1371/journal.pone.0170406
Pratt, L. A., and Kolter, R. (1998). Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 30, 285–293. doi: 10.1046/j.1365-2958.1998.01061.x
Quinn, H. J., Cameron, A. D., and Dorman, C. J. (2014). Bacterial regulon evolution: distinct responses and roles for the identical OmpR proteins of Salmonella Typhimurium and Escherichia coli in the acid stress response. PLoS Genet. 10:e1004215. doi: 10.1371/journal.pgen.1004215
Rodrigues, D., Cerca, N., Teixeira, P., Oliveira, R., Ceri, H., and Azeredo, J. (2011). Listeria monocytogenes and Salmonella enterica Enteritidis biofilms susceptibility to different disinfectants and stress-response and virulence gene expression of surviving cells. Microb. Drug Resist. 17, 181–189. doi: 10.1089/mdr.2010.0183
Ryan, D., Ojha, U. K., Jaiswal, S., Padhi, C., and Suar, M. (2016). The Small RNA DsrA influences the acid tolerance response and virulence of Salmonella enterica Serovar Typhimurium. Front. Microbiol. 7:599. doi: 10.3389/fmicb.2016.00599
Sangal, V., Harbottle, H., Mazzoni, C. J., Helmuth, R., Guerra, B., Didelot, X., et al. (2010). Evolution and population structure of Salmonella enterica serovar Newport. J. Bacteriol. 192, 6465–6476. doi: 10.1128/JB.00969-10
Sarkar, S., Vagenas, D., Schembri, M. A., and Totsika, M. (2016). Biofilm formation by multidrug resistant Escherichia coli ST131 is dependent on type 1 fimbriae and assay conditions. Pathog. Dis. 74:ftw013. doi: 10.1093/femspd/ftw013
Scallan, E., Mahon, B. E., Hoekstra, R. M., and Griffin, P. M. (2013). Estimates of illnesses, hospitalizations and deaths caused by major bacterial enteric pathogens in young children in the United States. Pediatr. Infect. Dis. J. 32, 217–221. doi: 10.1097/INF.0b013e31827ca763
Scher, K., Romling, U., and Yaron, S. (2005). Effect of heat, acidification, and chlorination on Salmonella enterica serovar Typhimurium cells in a biofilm formed at the air-liquid interface. Appl. Environ. Microbiol. 71, 1163–1168. doi: 10.1128/AEM.71.3.1163-1168.2005
Schroeder, A., Mueller, O., Stocker, S., Salowsky, R., Leiber, M., Gassmann, M., et al. (2006). The RIN: an RNA integrity number for assigning integrity values to RNA measurements. BMC Mol. Biol. 7:3. doi: 10.1186/1471-2199-7-3
Shah, D. H. (2014). RNA sequencing reveals differences between the global transcriptomes of Salmonella enterica serovar enteritidis strains with high and low pathogenicities. Appl. Environ. Microbiol. 80, 896–906. doi: 10.1128/AEM.02740-13
Simões, M., Simões, L. C., and Vieira, M. J. (2010). A review of current and emergent biofilm control strategies. LWT - Food Sci. Technol. 43, 573–583. doi: 10.1016/j.lwt.2009.12.008
Simonte, F. M., Dötsch, A., Galego, L., Arraiano, C., and Gescher, J. (2017). Investigation on the anaerobic propionate degradation by Escherichia coli K12. Mol. Microbiol. 103, 55–66. doi: 10.1111/mmi.13541
Song, C. W., Lee, J., and Lee, S. Y. (2015). Genome engineering and gene expression control for bacterial strain development. Biotechnol. J. 10, 56–68. doi: 10.1002/biot.201400057
Soni, K. A., Oladunjoye, A., Nannapaneni, R., Schilling, M. W., Silva, J. L., Mikel, B., et al. (2013). Inhibition and inactivation of Salmonella Typhimurium biofilms from polystyrene and stainless steel surfaces by essential oils and phenolic constituent carvacrol. J. Food Prot. 76, 205–212. doi: 10.4315/0362-028X.JFP-12-196
Stepanović, S., Ćirković, I., and Ranin, L. (2004). Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 38, 428–432. doi: 10.1111/j.1472-765X.2004.01513.x
Storz, G., Vogel, J., and Wassarman, K. M. (2011). Regulation by small RNAs in bacteria: expanding frontiers. Mol. Cell. 43, 880–891. doi: 10.1016/j.molcel.2011.08.022
Sun, H., Kamanova, J., Lara-Tejero, M., and Galán, J. E. (2016). A family of Salmonella type III secretion effector proteins selectively targets the nf-κb signaling pathway to preserve host homeostasis. PLoS Pathog. 12:e1005484. doi: 10.1371/journal.ppat.1005484
Sun, X., Liu, T., Peng, X., and Chen, L. (2014). Insights into Vibrio parahaemolyticus CHN25 response to artificial gastric fluid stress by transcriptomic analysis. Int. J. Mol. Sci. 15, 22539–22562. doi: 10.3390/ijms151222539
Vieira, A., Jensen, A., Pires, S. M., Karlsmose, S., Wegener, H. C., and Wong, D. L. F. (2009). “WHO global foodborne infections network country Databank–A resource to link human and non-human sources of salmonella,” in Proceeding of the 12th Symposium of the International Society for Veterinary Epidemiology and Economics (Durban).
Wang, H., Wu, N., Jiang, Y., Ye, K., Xu, X., and Zhou, G. (2016). Response of long-term acid stress to biofilm formation of meat-related Salmonella Enteritidis. Food Control. 69, 214–220. doi: 10.1016/j.foodcont.2016.04.055
Wang, H., Zhang, X., Dong, Y., Xu, X., and Zhou, G. (2015). Insights into the transcriptome profile of mature biofilm of Salmonella Typhimurium on stainless steels surface. Food Res. Int. 77, 378–384. doi: 10.1016/j.foodres.2015.08.034
Zbell, A. L., and Maier, R. J. (2009). Role of the Hya hydrogenase in recycling of anaerobically produced H2 in Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 75, 1456–1459. doi: 10.1128/AEM.02064-08
Ziebell, K., Chui, L., King, R., Johnson, S., Boerlin, P., and Johnson, R. P. (2017). Subtyping of Canadian isolates of Salmonella enteritidis using multiple locus variable number tandem repeat analysis (MLVA) alone and in combination with pulsed-field gel electrophoresis (PFGE) and phage typing. J. Microbiol. Methods 139, 29–36. doi: 10.1016/j.mimet.2017.04.012
Keywords: Salmonella Enteritidis, biofilm, acid stress, transcriptome, sRNAs
Citation: Jia K, Wang G, Liang L, Wang M, Wang H and Xu X (2017) Preliminary Transcriptome Analysis of Mature Biofilm and Planktonic Cells of Salmonella Enteritidis Exposure to Acid Stress. Front. Microbiol. 8:1861. doi: 10.3389/fmicb.2017.01861
Received: 12 July 2017; Accepted: 12 September 2017;
Published: 26 September 2017.
Edited by:
Marta López, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
Catherine Maeve Burgess, Teagasc, The Irish Agriculture and Food Development Authority, IrelandDario De Medici, Istituto Superiore di Sanità, Italy
Copyright © 2017 Jia, Wang, Liang, Wang, Wang and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huhu Wang, aHV1d2FuZ0BuamF1LmVkdS5jbg==