 Jan Dolfing
Jan Dolfing Casey R. J. Hubert
Casey R. J. Hubert- 1School of Civil Engineering and Geosciences, Newcastle University, Newcastle upon Tyne, United Kingdom
- 2Department of Biological Sciences, University of Calgary, Calgary, AB, Canada
Souring is the undesirable production of hydrogen sulfide (H2S) in oil reservoirs by sulfate-reducing bacteria (SRB). Souring is a common problem during secondary oil recovery via water flooding, especially when seawater with its high sulfate concentration is introduced. Nitrate injection into these oil reservoirs can prevent and remediate souring by stimulating nitrate-reducing bacteria (NRB). Two conceptually different mechanisms for NRB-facilitated souring control have been proposed: nitrate-sulfate competition for electron donors (oil-derived organics or H2) and nitrate driven sulfide oxidation. Thermodynamics can facilitate predictions about which nitrate-driven mechanism is most likely to occur in different scenarios. From a thermodynamic perspective the question “Which reaction yields more energy, nitrate driven oxidation of sulfide or nitrate driven oxidation of organic compounds?” can be rephrased as: “Is acetate driven sulfate reduction to sulfide exergonic or endergonic?” Our analysis indicates that under conditions encountered in oil fields, sulfate driven oxidation of acetate (or other SRB organic electron donors) is always more favorable than sulfide oxidation to sulfate. That predicts that organotrophic NRB that oxidize acetate would outcompete lithotrophic NRB that oxidize sulfide. However, sulfide oxidation to elemental sulfur is different. At low acetate HS− oxidation is more favorable than acetate oxidation. Incomplete oxidation of sulfide to S0 is likely to occur when nitrate levels are low, and is favored by low temperatures; conditions that can be encountered at oil field above-ground facilities where intermediate sulfur compounds like S0 may cause corrosion. These findings have implications for reservoir management strategies and for assessing the success and progress of nitrate-based souring control strategies and the attendant risks of corrosion associated with souring and nitrate injection.
Introduction
Souring, the undesirable production of hydrogen sulfide (H2S) in oil reservoirs by sulfate-reducing bacteria (SRB), is a common problem during secondary oil recovery when sea water is injected into the reservoir to maintain the high pressures required for oil extraction (Vigneron et al., 2017). The high concentration of sulfate in seawater (≈ 28 mM) promotes the growth and activity of sulfate reducing bacteria. Introducing sulfate to environments rich in reduced compounds such as hydrocarbons, organic acids and possibly H2 (from anaerobic metabolism; Muyzer and Stams, 2008) creates ideal conditions for SRB, hence souring poses ubiquitous challenges for oil producers, especially at offshore operations (Table 1, reactions R1 and R2). Nitrate injection into oil reservoirs can prevent and remediate souring by stimulating the growth and activity of nitrate-reducing bacteria (NRB) (e.g., Telang et al., 1997; Gittel et al., 2009). Two conceptually different mechanisms for NRB-facilitated souring control have been proposed (Hubert et al., 2009; Hubert, 2010). (i) Nitrate-sulfate competition: Nitrate is a better (energetically more favorable) electron acceptor than sulfate (Figure 1) (Table 1, e.g., reactions R3–R5 vs. R1, or R7 vs. R2); therefore NRB outcompete SRB, and consequently nitrate injection suppresses sulfate reduction. (ii) Nitrate driven sulfide oxidation: The sulfide produced by SRB during souring is re-oxidized with nitrate as electron acceptor (Table 1, R8). The latter mechanism potentially results in a cryptic sulfur cycle (i.e., regenerated sulfate can be re-used by SRB if electron donors are available; Hubert et al., 2003) but as long as nitrate is available sulfide will be essentially absent from the system. As such the two mechanisms are potentially stoichiometrically identical (nitrate-driven oxidation of electron donors either directly or via intermediate S cycling), with the essential difference between them being (Figure 2) that in pathway (i) organotrophic nitrate reducers dominate, while pathway (ii) hinges on the activity of sulfide oxidizing nitrate reducers.
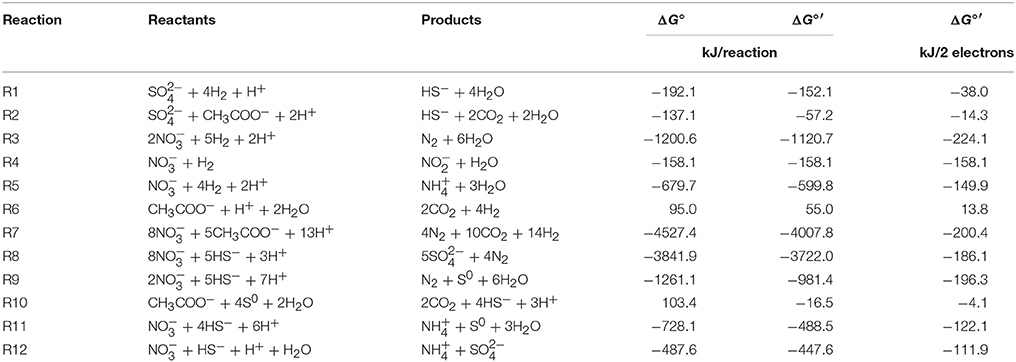
Table 1. Stoichiometry and change in Gibbs free energy values for reactions potentially involved in nitrate based souring control.
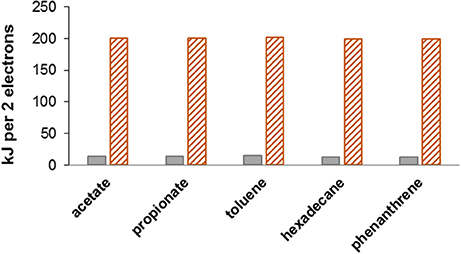
Figure 1. Nitrate (hatched orange bars) is a better electron acceptor than sulfate (gray bars). Energy yield (ΔG°′ in kJ/2electrons) is essentially independent of the type of organic electron donor used. In this example nitrate reduction is via denitrification to N2 (gas), while sulfate is reduced to sulfide. Partial reactions: acetate oxidation, CH3COO− + H+ + 2H2O → 2CO2 + 8[H], ΔG°′ = 14.3 kJ/2electrons; propionate oxidation, CH3CH2COO− + H+ + 4H2O → 3CO2 + 14[H], ΔG°′ = 14.2 kJ/2electrons; toluene oxidation, C6H5CH3 + 14H2O → 7CO2 + 36[H], ΔG°′ = 15.4 kJ/2electrons; hexadecane oxidation, C16H34 + 32H2O → 16CO2 + 98[H], ΔG°′ = 12.9 kJ/2electrons; phenanthrene oxidation, C14H10 + 28H2O → 14CO2 + 66[H], ΔG°′ = 13.4 kJ/2electrons; denitrification, 2H+ + 2 + 10[H] → N2 + 6H2O, ΔG°′ = −224.1 kJ/2electrons; sulfate reduction, 2H+ + + 8[H] → H2S + 4H2O, ΔG°′ = −38.0 kJ/2electrons; values for toluene and hexadecane are from Helgeson et al. (1998); for phenanthrene is from Dolfing et al. (2009).
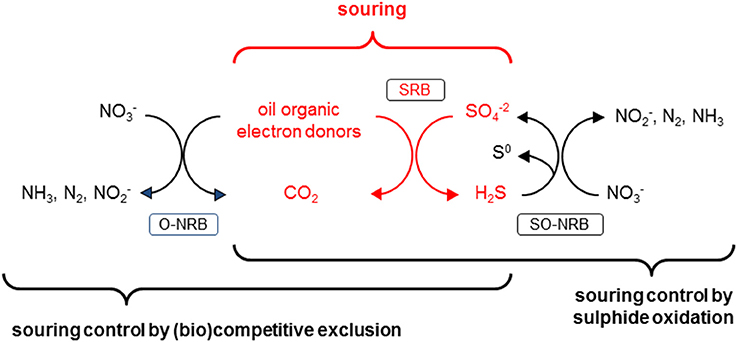
Figure 2. Schematic depicting reservoir souring (microbial production of H2S) in oil reservoirs due to the activity of sulfate reducing bacteria (SRB) that use organics present in the oil as electron donors. This souring scenario can interact with two potential mechanisms of nitrate-dependent souring control: (i) biocompetitive exclusion, where organotrophic nitrate-reducing bacteria (NRB) outcompete SRB for common electron donors (i.e., O-NRB), and (ii) sulfide oxidation, where sulfide-oxidizing NRB (i.e., SO-NRB) re-oxidize sulfide produced by SRB to S0 and/or .
Differentiating between causative agents of successful souring control following nitrate application is vital to better understand and further utilize nitrate injection as an emerging technology in the oil and gas sector. Here we use a thermodynamic approach to evaluate which nitrate reduction pathway is most likely to occur in different industrially encountered scenarios. Our considerations include the possibility of nitrate-driven sulfide oxidation to elemental sulfur (Table 1, R9), which has implications for corrosion of oil field infrastructure. The findings can contribute to predictive reservoir souring management strategies as well as assessments of souring- and nitrate-associated microbial-influenced corrosion risks.
Materials and Methods
Thermodynamic Calculations
The amount of free energy available from a reaction depends on the Gibbs free energies of formation of substrates and products as given by the relationship ΔG0 = Σ (products) - Σ (reactants). ΔG0 is the increment in free energy for the reaction under standard conditions: 25°C, 1 atm pressure for gases, 1 molal concentrations for solutes, pH = 0 (also 1 molal; Hanselmann, 1991). For biological systems the conventional standard conditions is as above, but with pH 7 (Thauer et al., 1977). This is considered in ΔG°′ values. ΔG°′ is identical with ΔG° except that the standard conditions of the H+ ion is that of pH 7, i.e., (H+) = −39.95 kJ/mol (Thauer et al., 1977; Dolfing et al., 2010).
Under environmentally relevant conditions the concentrations of reactants and products are generally not identical to those under standard conditions. This is considered in ΔG′ values. For a hypothetical reaction aA + bB → cC + dD, ΔG′ values were calculated by using the mass equation:
Gibbs free energy of formation () and enthalpy of formation () values (used to make temperature corrections for temperatures other than the standard temperature of 25°C) were taken from Hanselmann (1991) with gases (CO2 and H2) in the gaseous phase, and all other compounds except hexadecane in the aqueous phase. Values for hexadecane were for hexadecane in the liquid state (Helgeson et al., 1998).
Temperature corrections for ΔG° were made with the Gibbs-Helmholz equation according to:
with T in K; Tref = 298.15 K (Dolfing et al., 2008; Dolfing, 2015).
Results and Discussion
The Thermodynamic Approach
From a thermodynamic perspective the question “Which reaction yields more energy, nitrate driven oxidation of sulfide or nitrate driven oxidation of organic compounds (e.g., acetate)?” can be rephrased as: “What is the equilibrium constant for the acetate driven sulfate reduction?” or more to the point: “is acetate driven sulfate reduction exergonic or endergonic?” Figure 3 answers these questions and illustrates this line of reasoning by comparing ΔG°′ values for nitrate driven acetate oxidation (Table 1, R7) and nitrate driven sulfide oxidation (Table 1, R8), showing that the former is more exergonic than the latter (200 kJ/2 electrons transferred vs. 186 kJ/2 electrons transferred, under standard conditions). Figure 3 also depicts how the difference between the energetics of these two reactions (14 kJ/2 electrons transferred) represents the energetics of the sulfate driven oxidation of acetate (Table 1, R2). Thus, one can simply consider the energetics of the sulfate driven oxidation of acetate, and whether it is exergonic or endergonic under various conditions, to delineate under which conditions nitrate driven acetate oxidation is more favorable than nitrate driven sulfide oxidation.
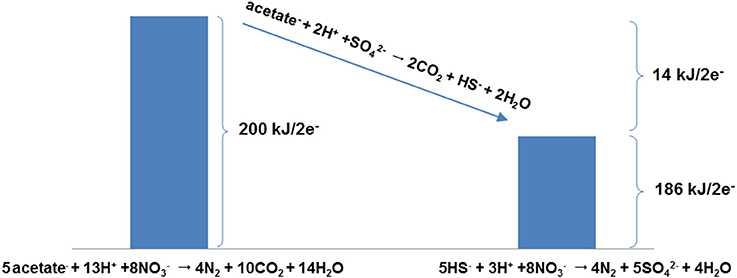
Figure 3. Energetics (ΔG°′ per 2 electrons) for the O-NRB reaction 5CH3COO− + 13H+ + 8 → 4N2 + 10CO2 + 14H2O and the SO-NRB reaction 5HS− + 3H+ + 8 → 4N2 + 5 + 4H2O, illustrating that the difference in ΔG°′ per 2 electrons between the two reactions corresponds to the ΔG°′ per 2 electrons for acetate-fuelled souring, i.e., for the reaction CH3COO− + 2H+ + ⇆ 2CO2 + HS− + 2H2O. This observation allows the acetate-driven sulfate reduction reaction to be used as a predictive tool for assessing whether acetate or sulfide is a better electron donor, i.e., depending on conditions driving the reaction to the left or the right.
Using the sulfate driven oxidation of acetate as a “tool” in this way, we can delineate under which conditions either sulfide or acetate as NRB electron donors are energetically favored. As an example, Figure 4 shows the “window of opportunity” for acetate driven sulfate reduction. The line depicts the combination of HS− and acetate concentrations where the ΔG of the reaction is zero, that is, where the reactants and products are in thermodynamic equilibrium (Dolfing et al., 2008). Above the line, the ΔG of the reaction is positive, i.e., under these conditions sulfide is the more exergonic electron donor for NRB. Below the line, acetate is the more exergonic electron donor for NRB. The predictive value and conclusion from this graph is that under realistic oil field conditions where the sulfide concentrations never exceeds or even approaches 1 M (34 g L−1) acetate oxidation is more favorable than sulfide oxidation.
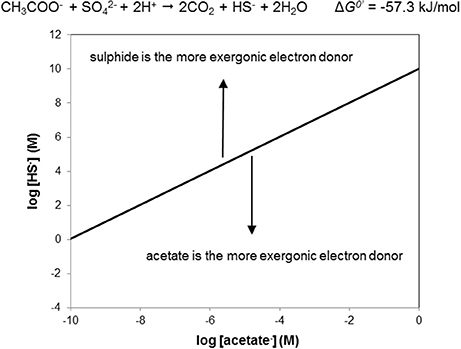
Figure 4. The window of opportunity for acetate driven denitrification (O-NRB) vs. sulfide driven denitrification (SO-NRB), illustrating at which combinations of sulfide and acetate either reaction is most exergonic. Above the black line sulfide is the most exergonic electron donor, below the black line acetate is the most endergonic electron donor. In oil fields the sulfide concentration is never higher than 1 M (34,000 mg L−1). Thus, acetate will always be the favored electron donor when nitrate is in large supply and assuming sulfide is completely oxidized to sulfate (as expected when nitrate is in large supply; cf. Figure 8). Calculations are for standard conditions (reactants and products at 1 M aqueous concentrations or at 1 atm partial pressure, at pH 7 and a temperature of 25°C). Reaction stoichiometries are as shown in Figure 3.
Figure 4 assumes conditions of pH 7 and 25°C. The conclusion that acetate is a more favorable electron donor for NRB than sulfide also holds for other pH values. Since protons are reactants in acetate driven sulfate reduction this statement seems trivial for pH < 7 if we employ the stoichiometry CH3COO− + + 2H+ → 2CO2 + HS− + 2H2O (Table 1, R2) on which Figure 4 is based. However, at pH < 7 H2S rather than HS− is the prevalent reduced inorganic species (Stumm and Morgan, 1996). Thus, the stoichiometry to be used is: CH3COO− + + 3H+ → 2CO2 + H2S + 2H2O, which implies that three rather than two moles of protons are consumed per mole of sulfate reduced. At pH > 7 HS− is the prevalent reduced inorganic sulfur species (Stumm and Morgan, 1996). Figure 5 shows the combinations of acetate and HS− concentrations at which the acetate driven sulfate reduction to sulfide is energy neutral (ΔG = 0) at higher pH. This reinforces the above conclusion that under in situ conditions in oil reservoirs and oil production waters at 25°C acetate oxidation is more favorable than sulfide oxidation.
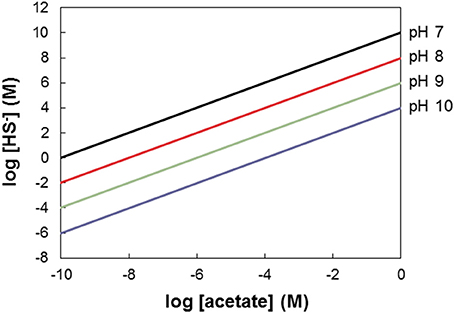
Figure 5. The window of opportunity for acetate driven denitrification (Table 1, R7) vs. sulfide driven denitrification (Table 1, R8), illustrating combinations of sulfide and acetate for which either reaction is most exergonic as function of pH. At pH = 7 sulfide is the most exergonic electron donor for combinations of [HS−] and [acetate] plotting above the black line. The black line thus indicates the combinations of acetate and sulfide where the energetics of the two denitrification reactions are equal (cf. Figure 4). At increasing pH values this line is lower on the graph (red, green, and blue lines give the boundaries at pH 8, pH 9, and pH 10, respectively).
Organic Electron Donors Other than Acetate
The above conclusion that under real world conditions at 25°C acetate oxidation is more favorable than sulfide oxidation can be generalized to other organic compounds. Figure 1 shows this for a variety of different organic electron donors typically encountered in oil field produced waters (e.g., Barth, 1991; Utvik, 1999), including acetate, propionate, toluene, hexadecane, and naphthalene. Expressed per two electrons transferred the change in standard Gibbs free energy for sulfate reduction to sulfide, for all electron donors, is essentially identical to the ΔG°′ values calculated for acetate driven sulfate reduction.
Incomplete Sulfide Oxidation
The above analysis indicates that under environmentally realistic conditions in oil fields oxidation of organics is always more thermodynamically favorable than sulfide oxidation to sulfate. However, for sulfide oxidation to elemental sulfur the picture changes. Consequences of elemental sulfur being present in the context of souring control are important to consider given that S0 and other intermediate sulfur compounds may play an aggressive role in oil field corrosion (Nemati et al., 2001; Hubert et al., 2005; Drønen et al., 2014), a phenomenon causing some operators to critically evaluate nitrate injection technology as a souring control mitigation option. Considering the single reaction “tool” presented above, the change in Gibbs free energy for the reaction CH3COO− + 4S0 + 2H2O ⇆ 2CO2 + 4HS− + 3H+ is −16.5 kJ/mol under biological standard conditions at pH 7 (Table 1, R10). Low acetate concentrations will drive this reaction to the left (i.e., make sulfide oxidation to S0 more favorable than acetate oxidation). At pH 7 the acetate concentration would need to be low enough for the critical ratio between acetate and sulfide would be about 1:1,000 (on a molar basis). Due to the speciation change of H2S and HS− at pH 7, low pH has less of an effect on this ratio than would be intuitively expected from the above equation where three moles of H+, and four moles of HS−, are produced per mole of acetate oxidized. In the range between pH = 4.5 and pH = 7 (the respective pK values for acetate and HS−) a lower pH slightly decreases the ΔG for the S0 driven acetate oxidation (Figure 6).
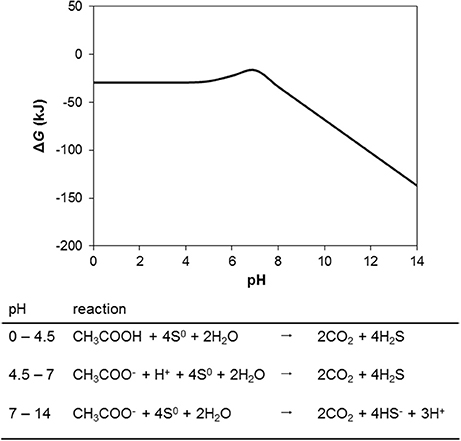
Figure 6. The effect of pH on the change in Gibbs free energy (ΔG) of S0 driven acetate oxidation under otherwise standard conditions.
Effect of Temperature
Acetate oxidation is more favorable than sulfide oxidation to sulfate in the temperature range between 2 and 100°C, and becomes less favorable with decreasing temperatures (Figure 7). For sulfide oxidation to elemental sulfur (S0) the effect of temperature is larger and more profound. As illustrated in Figure 7, the energetic advantage of acetate oxidation over sulfide oxidation to S0 strongly decreases when temperature decreases, to the extent that at low temperature (2°C) the energetic advantage of acetate disappears. The scenarios depicted for the lower temperatures in Figure 7 may be relevant at topsides oil field facilities in high latitude offshore environments such as the North Sea or farther north in the Barents Sea or the Arctic.
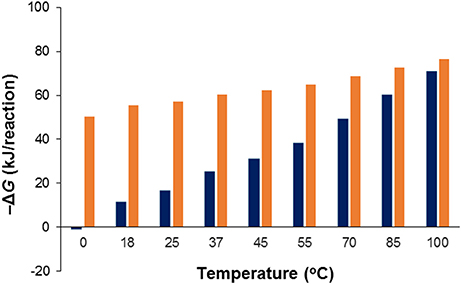
Figure 7. Effect of temperature on the change in Gibbs free energy (ΔG) for the acetate driven reduction of (Table 1, R1; red bars) and S0 (Table 1, R10; blue bars) to sulfide under otherwise standard conditions at pH 7. Note that at low temperature one of the reverse reactions, viz. the oxidation of sulfide to elemental sulfur coupled to CO2-based formation of acetate is slightly exergonic.
Denitrification vs. Dissimilatory Nitrate Reduction to Ammonia
Denitrification of nitrate to N2 is not the only potential nitrate conversion pathway catalyzed by nitrate-reducing microbial communities. Dissimilatory nitrate reduction to ammonia (DNRA) can also be envisaged (Kraft et al., 2014; van den Berg et al., 2015). While denitrifying NRB in oil fields are well known, such as organotrophic Thauera and Pseudomonas spp. (Agrawal et al., 2012; Fida et al., 2016) and lithotrophic Sulfurimonas spp. (e.g., strain CVO; Gevertz et al., 2000), some oil field O-NRB and SO-NRB (e.g., facultatively chemolithotrophic Sulfurospirillum spp.) have been shown to reduce nitrate to ammonia in pure culture (Hubert and Voordouw, 2007). Oil fields harboring organisms that reduce nitrate to ammonia are potentially amenable to souring control being achieved with addition of less nitrate, as DNRA is an 8 mol electron transfer reaction per mole of nitrate while denitrification of nitrate to N2 is a 5 mol electron transfer reaction per mole of nitrate (Table 1, R5 vs. R3). The lower cost to companies of using less nitrate may be attractive to operators, but this would depend on a knowledge of the ecophysiology of the NRB community present in a given oil production system, e.g., NRB metabolism and whether nitrate gets converted to fully reduced end products (Reinsel et al., 1996; Fida et al., 2016; Okpala et al., 2017).
There are two metrics that need to be assessed when evaluating the energetics of nitrate based sulfide oxidation: the ΔG per mole of nitrate used and the ΔG per mole of sulfide oxidized. Figure 8 illustrates that per mole of nitrate used incomplete sulfide oxidation to S0 with nitrate as the electron acceptor yields slightly more energy than complete oxidation to sulfate, independent of whether nitrate is reduced to N2 or ammonia (Table 1, reactions R8, R9, R11, and R12). When expressed in kJ/mol sulfide oxidized, the differences between the various scenarios are larger. Denitrification yields more energy than DNRA and complete oxidation to sulfate yields more energy than incomplete oxidation to S0. When nitrate is limiting, incomplete oxidation of sulfide to S0 is likely to prevail, whereas if nitrate is in excess (e.g., a high nitrate dose applied by operators) then complete oxidation to sulfate is expected to prevail.
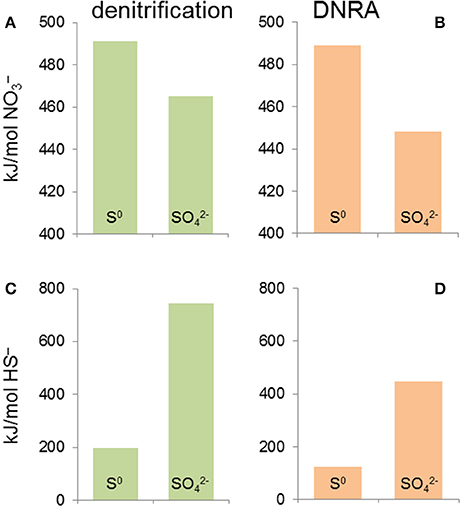
Figure 8. Change in Gibbs free energy values (ΔG°′) for sulfide-based nitrate reduction to N2 (denitrification) (A,C) and to ammonia (DNRA) (B,D). (A,B) show the ΔG°′ expressed per mole of nitrate reduced, whereas (C,D) show the ΔG°′ per mole of sulfide oxidized. This predicts that when nitrate is limiting, incomplete oxidation of sulfide to S0 is likely to prevail, whereas when sulfide is in short supply (i.e., application of a higher nitrate dose) complete oxidation to sulfate is expected to prevail. These predictions have important implications for managing corrosion risk in nitrate injection scenarios.
Perchlorate or Chlorate Based Souring Control Interventions
Perchlorate and chlorate have recently been proposed as alternative souring control interventions (Liebensteiner et al., 2014). These alternatives are posited to work via the same mechanisms as nitrate based control interventions: (bio)competitive exclusion and sulfide oxidation. If that assumption is valid, the same analysis as performed here for nitrate based interventions also applies to (per)chlorate based interventions. Perchlorate and chlorate are excellent electron acceptors, with redox potentials at least as favorable as nitrate (Liebensteiner et al., 2014). Therefore, the reasoning outlined in Figure 3 also applies to (per)chlorate: acetate based effects [(bio)competitive exclusion] vs. sulfide oxidation based effects can be rationalized by evaluating the energetics of the reactions CH3COO− + + 2H+ → 2CO2 + HS− + 2H2O (Table 1, R2) for compete sulfide oxidation when the electron acceptor (nitrate or (per)chlorate) is in large supply, and CH3COO− + 4S0 + 2H2O → 2CO2 + 4HS− + 3H+ (Table 1, R10) when the electron acceptor is limiting.
Implications for Souring Control and Corrosion Risk
These findings have implications for reservoir souring management and mitigation strategies. For example, there is an abundance of available electron donor in the reservoir, whereas topsides oil/water separation on surface platforms takes place in tanks where nitrate and sulfide may be present in the water but concentrations of acetate (and other organics) may be lower. In this latter context, sulfide oxidation could be most problematic (i) because it is more likely to be expected thermodynamically, and (ii) because of the potential for soNRB mediated corrosion in these topsides settings.
H2 as Source of Reducing Equivalents
Organics and reduced sulfur compounds are not the only potential electron donors in oil field systems. Hydrogen (H2) can also be envisaged as a by-product of anaerobic fermentative metabolism of crude oil compounds (Head et al., 2003), and is an excellent electron donor for both SRB and NRB. The (bio)competitive exclusion of H2-oxidizing SRB by H2-oxidizing NRB should therefore be considered. A thermodynamic evaluation of H2 vs. reduced sulfur compounds as electron donors for NRB in a souring control context follows the same line of reasoning as sketched above for acetate: whether H2 or reduced sulfur compounds are the most favorable electron donor can be evaluated based on the change in Gibbs free energy for the reaction 4H2 + + H+ → HS− + 4H2O (Table 1, R1). Under otherwise standard conditions at pH 7 H2 based sulfate reduction is exergonic (ΔG0′ = −152 kJ/mol sulfate): The equilibrium for the reaction 4H2 + + H+ ⇆ HS− + 4H2O (Table 1, R1) is to the right, which implies that H2 is energetically a more favorable electron donor than sulfide, and that hydrogenotrophic NRB would out-compete sulfide oxidizing NRB in a souring control setting. At equimolar concentrations of sulfate and sulfide the threshold H2 partial pressure below which sulfide is the most favorable electron acceptor is 0.02 Pa (Figure 9). In reality (i.e., in oil fields) the molar sulfide concentration will be orders of magnitude lower than the sulfate concentration, which further strengthens this assertion. Thus, H2 is an energetically more favorable electron acceptor than sulfide when sulfate is the oxidation product. This conclusion is slightly affected by temperature. The PH2 below which sulfide oxidation is more favorable than H2 oxidation decreases from 1.5 Pa at 85°C to an exceedingly low value of 0.003 Pa at 2°C. The sulfide oxidation product has a significant effect on the favorability of H2 vs. sulfide as reductant though. The PH2 below which sulfide oxidation to elemental sulfur is more favorable than H2 oxidation ranges from 0.7 Pa at 2°C to 3.5 Pa at 85°C. There are very few data in the literature on hydrogen gas concentrations in petroleum reservoirs (Dolfing et al., 2008). In methanogenic environments H2 levels are typically 1–2 Pa, vs. ~0.2 Pa under sulfate reducing conditions (Lovley and Goodwin, 1988; Hoehler et al., 1999). Thus, whether H2 or sulfide is the most favorable electron donor for NRB in oil fields will be strongly affected by the local conditions, and cannot be as easily predicted from first principles, compared to the situation for organic compounds such as acetate. It is therefore important to understand the anaerobic microbial communities and metabolic networks whereby oil-derived electron donors promote souring (sulfate reduction) in the subsurface, and the extent to which addition of nitrate as thermodynamically more favorable electron acceptor will divert these substrates to nitrate reduction pathways to control souring.
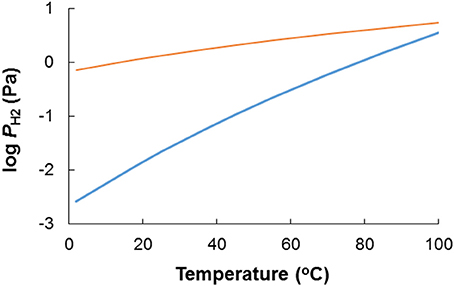
Figure 9. The effect of temperature on the H2 “threshold” concentration below which H2 is no longer a more favorable electron donor for nitrate reduction than sulfide. The blue line depicts the case where HS− is oxidized to (Table 1, R2), while the orange line depicts the case where HS− is oxidized to S0 (Table 1, R10).
Caveats
Our analyses are based on the assumption that all the reactions under consideration can be performed by the organisms present, i.e., that the O-NRB and SO-NRB in question are always found in oil field microbial communities, and would become activated under the chemical and environmental conditions described and assumed in the above scenarios. Obviously this is not necessarily the case in all instances or environments; it may well be that some of the organisms are absent, or are present but unable to be active, or to catalyse the reactions assumed above (e.g., complete reduction of nitrate to end products), or require a certain lag time to build up an effective population. Ongoing studies of subsurface microbial diversity and potential are continually assessing these parameters (Fida et al., 2016; Li et al., 2017; Okpala et al., 2017; Suri et al., 2017; Vigneron et al., 2017). Another caveat is that organisms may act as mixotrophs, for example use sulfide and nitrate and organics and nitrate simultaneously (Hubert and Voordouw, 2007). Another caveat is that other factors like kinetics may trump thermodynamic paradigms for predicting metabolic responses in mixed microbial systems (Chen et al., 2017). Hence thermodynamic calculations should be treated with caution when used as a predictive tool by operators in considering souring mitigation strategies and options. Furthermore, we are aware that introducing various partial pressures or concentrations of CO2 can have a profound influence on the reaction pathway in anaerobic ecosystems (cf. Mayumi et al., 2013). While those constraints are needed to make definite statements on the thermodynamics in the systems, we have worked with CO2 at atmospheric pressure, as this will provide a baseline for comparisons of reaction energetics.
Conclusion
The aim of this work is to provide a thermodynamic framework to evaluate the energetics of the various pathways potentially involved in nitrate-facilitated oil reservoir souring control. Nitrate injection technology is based on the textbook premise that nitrate is an energetically more favorable electron acceptor than sulfate. However, this does not necessarily imply that nitrate-facilitated souring control works via direct competition between NRB and SRB for organic electrons donors. An alternative mechanism in which the SRB use the reducing equivalents in the organics to reduce sulfate to sulfide followed by re-oxidation of the produced sulfide by NRB is also relevant, and has been interpreted in field settings following nitrate injection (Telang et al., 1997). The key observation put forward in the present work is that these alternative nitrate reduction mechanisms can be assessed by evaluating the thermodynamics of the difference between these two reactions. Our analysis indicates that, with acetate as a model organic electron donor (Figure 3) sulfate reduction to sulfide is always more energetically favorable than the reverse reaction under realistic oil field conditions. This approach thus predicts that acetate would be a more favorable electron donor than sulfide, e.g., for NRB and hence in a nitrate-based souring control context. Thus, to answer the question phrased in the introduction: nitrate-sulfate competition seems a more likely souring control mechanism than nitrate driven sulfide oxidation, with nitrate reduction fuelled by acetate and other organic compounds as the pathway of choice. However, sulfide is not necessarily oxidized fully to sulfate under all conditions. Incomplete oxidation of sulfide to elemental sulfur by SO-NRB can also be envisaged, especially under conditions where nitrate is limiting. This changes the essential thermodynamics of the reactions under consideration significantly; under nitrate limited conditions, where sulfide is oxidized to sulfur, this reaction can be the energetically more favorable outcome; this is especially true when acetate concentrations are low, and or at low temperatures—conditions that are relevant for oil field topsides facilities where corrosion, possibly accelerated by elemental sulfur, is a major concern. The work presented here offers a simple thermodynamic approach to rationalize the most likely potential outcomes of souring control interventions, and enable predictions around nitrate-based souring control management by oil producers and operators. An additional crucial implication of the considerations made in our study is that half-hearted measures with limited nitrate supplements can be counterproductive as they may be contribute to the formation of S0.
Author Contributions
JD: conceived the idea; JD and CH: designed the project, conducted the analyses, and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by the Engineering and Physical Sciences Research Council, UK (grant reference EP/J002259/1). The authors also wish to thank Gerrit Voordouw for valuable discussions.
References
Agrawal, A., Park, S., Nathoo, S., Gieg, L. M., Jack, T. R., Miner, K., et al. (2012). Toluene depletion in produced oil contributes to souring control in a field subjected to nitrate injection. Environ. Sci. Technol. 46, 1285–1292. doi: 10.1021/es203748b
Barth, T. (1991). Organic acids and inorganic ions in waters from petroleum reservoirs, Norwegian continental shelf: a multivariate statistical analysis and comparison with American reservoir formation waters. Appl. Geochem. 6, 1–15. doi: 10.1016/0883-2927(91)90059-X
Chen, J., Hanke, A., Tegetmeyer, H. E., Kattelmann, I., Sharma, R., Hamann, E., et al. (2017). Impacts of chemical gradients on microbial community structure. ISME J. 11, 920–931. doi: 10.1038/ismej.2016.175
Dolfing, J. (2015). “Protocols for calculating reaction kinetics and thermodynamics,” in Hydrocarbon and Lipid Microbiology Protocols – Springer Protocols Handbooks, eds T. J. McGenity, K. N. Timmis, and B. Nogales (Berlin: Springer), 155–163.
Dolfing, J., Larter, S. R., and Head, I. M. (2008). Thermodynamic constraints on methanogenic crude oil biodegradation. ISME J. 2, 442–452. doi: 10.1038/ismej.2007.111
Dolfing, J., Xu, A., Gray, N. D., Larter, S. R., and Head, I. M. (2009). The thermodynamic landscape of methanogenic PAH degradation. Microb. Biotechnol. 2, 566–574. doi: 10.1111/j.1751-7915.2009.00096.x
Dolfing, J., Xu, A., and Head, I. M. (2010). Anomalous energy yields in thermodynamic calculations: importance of accounting for pH-dependent organic acid speciation. ISME J. 4, 463–464. doi: 10.1038/ismej.2010.14
Drønen, K., Roalkvam, I., Beeder, J., Torsvik, T., Steen, I. H., Skauge, A., et al. (2014). Modeling of heavy nitrate corrosion in anaerobe aquifer injection water biofilm: a case study in a flow rig. Environ. Sci. Technol. 48, 8627–8635. doi: 10.1021/es500839u
Fida, T. T., Chen, C., Okpala, G., and Voordouw, G. (2016). Implications of limited thermophilicity of nitrite reduction for control of sulfide production in oil reservoirs. Appl. Environ. Microbiol. 82, 4190–4199. doi: 10.1128/AEM.00599-16
Gevertz, D., Telang, A. T., Voordouw, G., and Jenneman, G. E. (2000). Isolation and characterization of strains CVO and FWKO B, two novel nitrate-reducing, sulfide-oxidizing bacteria isolated from oil field brine. Appl. Environ. Microbiol. 66, 2491–2501. doi: 10.1128/AEM.66.6.2491-2501.2000
Gittel, A., Sorensen, K. B., Skovhus, T. L., Ingvorsen, K., and Schramm, A. (2009). Prokaryotic community structure and sulfate reducer activity in water from high-temperature oil reservoirs with and without nitrate treatment. Appl. Environ. Microbiol. 75, 7086–7096. doi: 10.1128/AEM.01123-09
Hanselmann, K. W. (1991). Microbial energetics applied to waste repositories. Experientia 47, 645–687. doi: 10.1007/BF01958816
Head, I. M., Jones, D. M., and Larter, S. R. (2003). Biological activity in the deep subsurface and the origin of heavy oil. Nature 426, 344–352. doi: 10.1038/nature02134
Helgeson, H. C., Owens, C. E., Knox, A. M., and Richard, L. (1998). Calculation of the standard molal thermodynamic properties of crystalline, liquid and gas organic molecules at high temperatures and pressures. Geochim. Cosmochim. Acta 42, 985–1081. doi: 10.1016/S0016-7037(97)00219-6
Hoehler, T. M., Alperin, M. J., Albert, D. B., and Martens, C. S. (1999). Acetogenesis from CO2 in an anoxic marine sediment. Limnol. Oceanogr. 44, 662–667. doi: 10.4319/lo.1999.44.3.0662
Hubert, C. (2010). “Chapter 26: Microbial ecology of oil reservoir souring control by nitrate injection,” in Handbook of Hydrocarbon and Lipid Microbiology, Vol. 4, ed K. N. Timmis (Berlin: Springer), 2753–2766.
Hubert, C., Nemati, M., Jenneman, G., and Voordouw, G. (2003). Containment of biogenic sulfide production in continuous up-flow packed-bed bioreactors with nitrate or nitrite. Biotechnol. Prog. 19, 338–345. doi: 10.1021/bp020128f
Hubert, C., Nemati, M., Jenneman, G., and Voordouw, G. (2005). Corrosion risk associated with microbial souring control using nitrate or nitrite. Appl. Microbiol. Biotechnol. 68, 272–282. doi: 10.1007/s00253-005-1897-2
Hubert, C., and Voordouw, G. (2007). Oil field souring control by nitrate-reducing Sulfurospirillum spp. that outcompete sulfate-reducing bacteria for organic electron donors. Appl. Environ. Microbiol. 73, 2644–2652. doi: 10.1128/AEM.02332-06
Hubert, C., Voordouw, G., and Mayer, B. (2009). Elucidating microbial processes in nitrate- and sulfate-reducing systems using sulfur and oxygen isotope ratios: the example of oil reservoir souring control. Geochim. Cosmochim. Acta 73, 3864–3879. doi: 10.1016/j.gca.2009.03.025
Kraft, B., Tegetmeyer, H. E., Sharma, R., Klotz, M. G., Ferdelman, T. G., Hettich, R. L., et al. (2014). The environmental controls that govern the end product of bacterial nitrate respiration. Science 345, 676–679. doi: 10.1126/science.1254070
Li, X.-X., Liu, J.-F., Zhou, L., Mbadinga, S. M., Yang, S.-Z., Gu, J.-D., et al. (2017). Diversity and composition of sulfate-reducing microbial communities based on genomic DNA and RNA transcription in production water of high temperature and corrosive oil reservoir. Front. Microbiol. 8:1011. doi: 10.3389/fmicb.2017.01011
Liebensteiner, M. G., Tsesmetzis, N., Stams, A. J. M., and Lomans, B. P. (2014). Microbial redox processes in deep subsurface environments and the potential application of (per)chlorate in oil reservoirs. Front. Microbiol. 5:428. doi: 10.3389/fmicb.2014.00428
Lovley, D. R., and Goodwin, S. (1988). Hydrogen concentrations as an indicator of the predominant terminal electron-accepting reactions in aquatic sediments. Geochim. Cosmochim. Acta 52, 2993–3003.
Mayumi, D., Dolfing, J., Sakata, S., Maeda, H., Miyagawa, Y., Ikarashi, M., et al. (2013). Carbon dioxide concentration dictates alternative methanogenic pathways in oil reservoirs. Nat. Commun. 4:1998. doi: 10.1038/ncomms2998
Muyzer, G., and Stams, A. J. M. (2008). The ecology and biotechnology of sulfate-reducing bacteria. Nat. Rev. Microbiol. 6, 441–454. doi: 10.1038/nrmicro1892
Nemati, M., Jenneman, G. E., and Voordouw, G. (2001). Impact of nitrate-mediated microbial control of souring in oil reservoirs on the extent of corrosion. Biotechol. Prog. 17, 852–859. doi: 10.1021/bp010084v
Okpala, G., Chen, C., Fida, T., and Voordouw, G. (2017). Effect of thermophilic nitrate reduction on sulfide production in high temperature oil reservoir samples. Front. Microbiol. 8:1573. doi: 10.3389/fmicb.2017.01573
Reinsel, M. A., Sears, J. T., Stewart, P. S., and McInerney, M. J. (1996). Control of microbial souring by nitrate, nitrite or glutaraldehyde injection in a sandstone column. J. Indus. Microbiol. 17, 128–136. doi: 10.1007/BF01570056
Suri, N., Voordouw, J., and Voordouw, G. (2017). The effectiveness of nitrate-mediated control of the oil field sulfur cycle depends on the toluene content of the oil. Front. Microbiol. 8:956. doi: 10.3389/fmicb.2017.00956
Telang, A. J., Ebert, S., Foght, J. M., Westlake, D. W. S., Jenneman, G. E., Gevertz, D., et al. (1997). Effect of nitrate injection on the microbial community in an oil field as monitored by reverse sample genome probing. Appl. Environ. Microbiol. 63, 1785–1793.
Thauer, R. K., Jungermann, K., and Decker, K. (1977). Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41, 100–180.
Utvik, T. I. R. (1999). Chemical characterisation of produced water from four offshore oil production platforms in the North Sea. Chemosphere 39, 2593–2606. doi: 10.1016/S0045-6535(99)00171-X
van den Berg, E. M., van Dongen, U., Abbas, B., and van Loosdrecht, M. C. M. (2015). Enrichment of DNRA bacteria in a continuous culture. ISME J. 9, 2153–2161. doi: 10.1038/ismej.2015.26
Keywords: thermodynamics, sulfate reduction, nitrate injection, petroleum reservoir, souring control, biocompetitive exclusion, sulfide oxidation, denitrification
Citation: Dolfing J and Hubert CRJ (2017) Using Thermodynamics to Predict the Outcomes of Nitrate-Based Oil Reservoir Souring Control Interventions. Front. Microbiol. 8:2575. doi: 10.3389/fmicb.2017.02575
Received: 26 September 2017; Accepted: 11 December 2017;
Published: 19 December 2017.
Edited by:
Kenneth Wasmund, University of Vienna, AustriaReviewed by:
Jeffrey M. Dick, Central South University, ChinaClemens Glombitza, Ames Research Center (NASA), United States
Copyright © 2017 Dolfing and Hubert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Dolfing, amFuLmRvbGZpbmdAbmNsLmFjLnVr