 Beatriz del Rio
Beatriz del Rio Begoña Redruello
Begoña Redruello Maria Fernandez
Maria Fernandez M. Cruz Martin
M. Cruz Martin Victor Ladero
Victor Ladero Miguel A. Alvarez
Miguel A. Alvarez- Department of Technology and Biotechnology of Dairy Products, Dairy Research Institute (IPLA-CSIC), Madrid, Spain
Lactic acid bacteria (LAB) are among the most widely used microorganisms in food fermentation. However, some LAB species can also be used as live vehicles for the in situ delivery of therapeutic molecules to the mucosa of the human gastrointestinal tract (GIT). Many LAB species have ‘qualified presumption of safety’ status and survive passage through the GIT. Indeed, some are part of the usual GIT microbiota. These are appropriate candidates for the in situ production of recombinant prophylactic and therapeutic proteins. Live recombinant LAB that produce microbial antigens have been shown to elicit an immune response that confers protection against the corresponding pathogens; these LAB could therefore be used as oral vaccines. In addition, some LAB have been genetically engineered to produce therapeutic, neutralizing antibodies. The variable domain of heavy-chain-only antibodies from camelids – known as VHH antibodies or nanobodies – has peculiar properties (nanoscale size, robust structure, acid resistance, high affinity and specificity, easily produced in bacteria, etc.) that make them ideal choices as LAB-produced immunotherapeutic agents. The present review examines the advantages offered by LAB for the in situ production of therapeutic proteins in the human GIT, discusses the use of in situ produced VHH antibody fragments, and assesses the usefulness of this strategy in the treatment of infectious and non-infectious gastrointestinal diseases.
Introduction
New therapeutic strategies are needed if we are to better face the challenges posed by cancer, resistance to antibiotics, and viral infections. The development of systems that allow drugs to be more precisely delivered to their target organs, and that better control their release, is a major goal (Wells, 2011; Hosseinidoust et al., 2016); non-specific drug delivery can be associated with toxic side effects in non-target tissues and organs.
It has been proposed that live bacteria be used as vectors for the in situ delivery of recombinant proteins for prophylactic and therapeutic purposes (Medina and Guzman, 2001; Wells and Mercenier, 2008; Cano-Garrido et al., 2015; Hosseinidoust et al., 2016; Ding et al., 2018). This strategy should be inexpensive since bacteria are easy to grow, the pharmaceutical production and purification of the active agent are avoided, and degradation problems (which are particularly severe in the gastrointestinal tract [GIT]) can be overcome (Wells, 2011; Wang et al., 2016). The producing bacteria can also be lyophilized, avoiding the need to maintain a cold chain (Pant et al., 2006). Attenuated pathogenic bacteria were originally proposed for use in such systems, but lactic acid bacteria (LAB) quickly became recognized as ideal candidates, especially for the prevention and treatment of mucosal diseases (Cano-Garrido et al., 2015; Wang et al., 2016).
Advantages of Lab as Live Vectors for the In Situ Production of Therapeutic Proteins
The LAB form a heterogeneous group of Gram-positive bacteria that include technologically important species of the genera Lactobacillus, Lactococcus, Streptococcus, Leuconostoc, Oenococcus, and Pediococcus. They are mainly characterized by their ability to produce lactic acid from sugar – an ability long exploited by humans to produce fermented foods such as cheese, yoghurt, sausage, wine, sauerkraut, and pickled vegetables. Such foods can be safely consumed long after they are made and may even be of enhanced nutritional value. The presence of LAB in foods and their lack of pathogenic traits or association with any foodborne disease have led to their being regarded as safe. In fact, most LAB species are ‘generally regarded as safe’ (GRAS) by the US Food and Drug Administration (FDA) and have been awarded ‘qualified presumption of safety’ (QPS) status by the European Food Safety Authority (EFSA).
LAB occupy a wide range of niches. Some species are found in milk and in plants, while others are common members of the endogenous microbiota associated with the oral, urogenital, and GIT mucosae in animals and humans (Klaenhammer et al., 2002). Certain LAB strains play a role in controlling the intestinal microbiota, restoring the intestinal barrier function, and alleviating the inflammatory responses associated with GIT disease. This has fomented their use in the treatment and prevention of various intestinal disorders (Ouwehand et al., 2002; Isolauri et al., 2004; Gareau et al., 2010). Certain LAB groups, such as the lactobacilli, are important members of the endogenous microbiota of human mucosal surfaces, where they play crucial roles in homeostasis, providing the protection against pathogenic bacteria and stimulating the immune system (Isolauri et al., 2004; Bernardeau et al., 2008). Given their beneficial nature, LAB species such as Lactobacillus acidophilus, Lactobacillus casei, and Lactobacillus rhamnosus, which meet the definition of probiotics (FAO/WHO, 2006), have been added to food to provide consumers with their health benefits (Saxelin et al., 2005; Gareau et al., 2010; Salminen et al., 2010). Indeed, LAB, and lactobacilli in particular, have become a focus of interest for use in human and animal biomedical applications (Wells and Mercenier, 2008; Behnsen et al., 2013; Cano-Garrido et al., 2015; Wang et al., 2016).
LAB are reported to be ideal candidates as live delivery vehicles for releasing therapeutic and prophylactic molecules directly at the oral, nasal, and genital mucosae and to be a realistic option for the treatment of human and animal diseases (Cano-Garrido et al., 2015; Piñero-Lambea et al., 2015; Wang et al., 2016). The safety status of the LAB, the ability of some strains to survive passage through the GIT (Vesa et al., 2000), and the capacity of some species (e.g., L. rhamnosus , Bifidobacterium animalis, and Lactobacillus plantarum) to remain viable in the GIT for a period of time render food-grade LAB ideal vehicles for delivering and even producing therapeutic molecules in situ at the GIT mucosa (Daniel et al., 2011; Wang et al., 2016). The absence of lipopolysaccharides (LPSs) in their cell walls (which is not the case in Gram-negative bacteria such as Escherichia coli ) is a further advantage, allowing them to be administered orally without the risk of endotoxic shock (Szatraj et al., 2017). In addition, the oral administration of therapeutic molecules via live recombinant LAB is a suitable alternative to invasive administration methods, for example, parenteral or subcutaneous injection, avoiding their potential side effects. Further, it circumvents the degradation of orally administered naked molecules in the digestive tract and ensures the production of the therapeutic protein at the GIT mucosa (Wang et al., 2016). Moreover, the in vivo synthesis of the therapeutic molecule reduces the dose required when compared to systemic or subcutaneous treatment (Steidler et al., 2000; Cano-Garrido et al., 2015).
In recent decades, much effort has gone into the genetic manipulation of LAB with the aim of producing recombinant therapeutic molecules (García-Fruitós, 2012; Cano-Garrido et al., 2015). Tools that allow cloning, the modulation of expression, and even the localization of recombinant proteins are now available (de Ruyter et al., 1996; Martin et al., 2000, 2011; Hanniffy et al., 2004; Benbouziane et al., 2013; Linares et al., 2014; Linares et al., 2015; Michon et al., 2016). Recombinant proteins can be engineered to be secreted into the extracellular environment or to be secreted and then anchored on the bacterial surface. Proteins to be secreted must have an N-terminus signal peptide recognized by the bacterial secretion machinery. One of the secretion mechanisms most studied in genetic engineering is the Sec-dependent pathway (Mathiesen et al., 2008). This drives the translocation of the precursor protein (i.e., the signal peptide plus the mature protein) across the plasma membrane. Either during or after translocation, a signal peptidase cleaves off the signal peptide and the mature protein is released into the extracellular environment (Schneewind and Missiakas, 2014). Different signal peptides have been exploited for engineered secretion in LAB, such as that associated with the major lactococcal secreted protein Usp45 (Dieye et al., 2001), the Lactobacillus brevis S-layer protein (SlpA) (Oh et al., 2007), the Streptococcus pyogenes M6 protein (Hols et al., 1997), and the Lactobacillus crispatus aggregation-promoting factor (APF) (Martin et al., 2011; Pant et al., 2011; Gunaydin et al., 2014), among others (Mathiesen et al., 2008). Secreted recombinant proteins can also be engineered by the translational fusion of an anchor peptide (displayed on the bacterial surface) via covalent or non-covalent bonding (Desvaux et al., 2006; Zadravec et al., 2015; Mao et al., 2016; Michon et al., 2016). Indeed, several anchoring peptides derived from surface-exposed proteins of Gram-positive bacteria have been used to bind secreted recombinant proteins to the surface of LAB – either to the plasma membrane or to the cell wall components (Michon et al., 2016). The signals used to anchor the protein to the cell membrane include N-terminal transmembrane helix anchors, for example, the transmembrane domain of the PgsA protein from Bacillus subtilis (Narita et al., 2006), and lipoprotein anchor domains, for example, the anchor domain of lactococcal basic membrane protein A (BmpA) (Zadravec et al., 2014). Recombinant proteins can be covalently anchored to the bacterial cell wall, which is usually achieved by fusing the target protein to an LPXTG-type cell wall-sorting signal. This consists of an LPXTG motif followed by a C-terminal hydrophobic domain and a positively charged tail that adequately orientates the protein in the membrane, allowing a membrane-anchored sortase to cut a peptide bond of the LPXTG motif and catalyze covalent bonding with the peptidoglycan (Schneewind et al., 1993; Siegel et al., 2017). The LPXTG anchors used include the cell-wall anchor of the S. pyogenes M6 protein (Dieye et al., 2001; Cortes-Perez et al., 2003), the anchor sequence of proteinase P of Lactobacillus zeae (Kruger et al., 2002; Martin et al., 2011), and the LPXTG anchor of the cell wall surface adherent protein Lp_2578 produced by L. plantarum WCFS1 (Fredriksen et al., 2010). Recombinant proteins may also be non-covalently attached to the cell wall of LAB by fusing them to different cell wall-binding domains such as the anchoring system of the APF of L. crispatus (Martin et al., 2011), the lysin motif (LysM) (Fredriksen et al., 2012), and the C-terminal region of S-layer proteins (Hu et al., 2011). Thanks to these genetic tools, a large number of recombinant LAB strains have been designed that produce therapeutic or prophylactic proteins (Wells and Mercenier, 2008; Bermudez-Humaran et al., 2011; Daniel et al., 2011; Cano-Garrido et al., 2015; Wang et al., 2016).
Recombinant Lab that Produce Therapeutic Molecules
Some LAB are natural components of the GIT microbiota. Strains such as L. rhamnosus GG and B. animalis BB12 have been used as probiotics for modulating the GIT microbiota and even to treat some intestinal disorders (Gerritsen et al., 2011). The earliest use of therapeutic molecule-producing recombinant LAB was in the treatment of GIT inflammatory diseases and bacterial and viral infections. The literature now contains many articles describing LAB genetically engineered to produce therapeutic proteins for use in the treatment of gastrointestinal disease (Bermudez-Humaran et al., 2011; Cano-Garrido et al., 2015; Mays and Nair, 2018).
Recombinant LAB for Use Against GIT Inflammatory Diseases
Many recombinant LAB have been developed to fight inflammatory bowel disease (IBD), a group of disorders that cause chronic inflammation in different parts of the GIT. Crohn’s disease and ulcerative colitis are the two most important types. Almost two decades ago, a recombinant strain of Lactococcus lactis was engineered to secrete the anti-inflammatory interleukin 10 (IL-10) (Steidler et al., 2000), and when administered orally, it reduced colitis in two mouse models (mice treated with dextran sulfate sodium [DDS] and IL-10 −/− mice). The success obtained led the authors to continue their research with the aim of performing clinical trials, and they developed a biocontained L. lactis producing IL-10 in which the host thymidylate synthase gene (thyA) was replaced by a synthetic human IL-10 gene. Since thyA is essential for the growth of L. lactis, the viability of the biocontained strain depended on the presence of thymidine or thymine (Steidler et al., 2003). A Phase I clinical trial involving patients with Crohn’s disease reported reduced disease activity and confirmed the strain’s biocontainment, safety, and tolerability (Braat et al., 2006).
Other recombinant LAB strains designed for the oral delivery of anti-inflammatory molecules to the gut have also been reported to prevent or reduce inflammation in different murine models of colitis (Cano-Garrido et al., 2015; Wang et al., 2016). For example, L. lactis strains were engineered to produce the immunomodulatory low calcium response V (LcrV) protein of Yersinia pseudotuberculosis (Foligne et al., 2007), the cytoprotective trefoil factor (TFF) (Vandenbroucke et al., 2004), the immunosuppressive cytokine interleukin 27 (IL-27) (Hanson et al., 2014), insulin-like growth factor I (IGF-I) (Liu et al., 2016), heat shock protein 65 (Hsp65) (Gomes-Santos et al., 2017), and serine protease inhibitors (Bermudez-Humaran et al., 2015). Recombinant L. casei producing superoxide dismutase has also been designed (Watterlot et al., 2010).
Celiac disease, an autoimmune disorder of genetically pre-disposed individuals, also causes bowel inflammation after the ingestion of gluten. The oral administration of live recombinant LAB producing prolyl endopeptidases – enzymes that degrade the immunotoxic peptides of gluten – might be a way of preventing an inflammatory response (Alvarez-Sieiro et al., 2014). A generated food-grade recombinant L. casei strain was reported to stably and constitutively secrete a prolyl endopeptidase able to break down the 33-mer peptide of gluten (one of the most immunotoxic). The strain also resisted deactivation under simulated gastrointestinal conditions in an in vitro model with its ability to secrete the enzyme intact (Alvarez-Sieiro et al., 2014).
Recombinant LAB as Live Mucosal Vaccines
One of the most extended uses of recombinant LAB that produce therapeutic or prophylactic molecules is as live mucosal vaccines (Wells and Mercenier, 2008; Wyszynska et al., 2015; Szatraj et al., 2017). In addition to the already mentioned advantages of LAB as live mucosal delivery vehicles, these microorganisms have additional properties that render them good choices as live vaccine vectors for mucosal vaccination. For instance, some LAB strains show innate immunoadjuvant properties that enhance the immune response provoked by the desired heterologous immunogenic molecule (Hanniffy et al., 2004; Mohamadzadeh and Klaenhammer, 2008; Bermudez-Humaran et al., 2011). In addition, they have the potential to elicit antigen-specific cellular, mucosal, and systemic humoral immune responses (del Rio et al., 2008, 2010; Wells and Mercenier, 2008; Bermudez-Humaran et al., 2011). Further, live recombinant LAB allow for immunization via the oral route, enabling the production in vivo of the therapeutic or prophylactic molecules at the GIT mucosa and avoiding the degradation of the treatment molecule in the harsh environment of the stomach (Wang et al., 2016). Moreover, administration is non-invasive and can be performed by personnel with relatively low-level training. There are neither any needles nor syringes to dispose of. It is therefore an ideal strategy for large-scale immunization programs (del Rio et al., 2014). Finally, the possibility of storing recombinant lyophilized LAB avoids the need for a cold chain, making these types of vaccine suitable for use in developing countries.
LAB for Active Immunization
Recombinant LAB that produce microbial antigens have been proposed as oral vaccines against a range of pathogens. Back in 1993, L. lactis was engineered to express fragment C of the tetanus toxin (TTFC) (Wells et al., 1993). Further studies showed that the administration of TTFC-expressing recombinant L. lactis via either the nasal (Norton et al., 1997) or the oral route (Robinson et al., 1997) protected mice against a lethal challenge with tetanus toxin. Since then, a large number of recombinant LAB strains have been generated as potential mucosal vaccines to prevent infections caused by other pathogens (Wells, 2011; Wang et al., 2016). Several recombinant LAB orally administered in appropriate animal models were reported to confer in vivo protection against infections of the GIT caused by Helicobacter pylori (Gu et al., 2009; Hongying et al., 2014; Li et al., 2014), enterohemorrhagic Escherichia coli (EHEC) (Ahmed et al., 2014), enterotoxigenic E. coli (ETEC) (Liu et al., 2009; Wei et al., 2010; Wen et al., 2012), Clostridium difficile (Yang et al., 2013; Guo et al., 2015), Salmonella enterica (Kajikawa et al., 2007), rotavirus (Marelli et al., 2011), the yeast Candida albicans (Shibasaki et al., 2014), and even parasites such as Plasmodium yoelii (Zhang et al., 2005) and Giardia lamblia (Lee et al., 2009). Peptide allergen-producing LAB have also been proposed as a therapeutic option for the treatment of peanut (Ren et al., 2014) and cow’s milk allergies (Adel-Patient et al., 2005).
LAB for Passive Immunization
LAB might also be used to produce antibodies to generate passive immunity. Passive immunity, which involves the transfer of ready-to-act antibodies, occurs naturally when maternal antibodies are transferred to the fetus through the placenta, and during lactation. However, it can be artificially induced by transferring large numbers of pathogen- or toxin-specific antibodies to a non-immune individual through blood products, for example, in immunoglobulin or antiserum therapy. LAB can be used as a vehicle for passive immunization by engineering them to produce specific antibodies. LAB producing single-chain variable fragment (scFv) antibodies (chimeric molecules that fuse two different regions of both heavy and light immunoglobulin chains) (Harmsen and De Haard, 2007) have been developed in an attempt to provide passive protection against different diseases of the GIT and other organs (Shiroza et al., 2003; Giomarelli et al., 2004; Monedero et al., 2004; Chancey et al., 2006; Marcotte et al., 2006; Yuvaraj et al., 2008; Wells, 2011; Thueng-in et al., 2014; Michon et al., 2015; Marcobal et al., 2016; Wang et al., 2016). After being orally administered in the corresponding animal model, passive immunoprotection developed against gastrointestinal infections caused by C. albicans (Oggioni et al., 2001), Streptococcus mutans (Kruger et al., 2002, 2005), norovirus (Hoang et al., 2015), and against anthrax edema toxin (Andersen et al., 2011).
VHH Antibody Fragments
In addition to conventional immunoglobulin G (IgG) antibodies, the serum of llamas, camels, and dromedaries has antibodies with two identical heavy chains but no light chains (Hamers-Casterman et al., 1993; Muyldermans, 2013). The variable domains of these heavy chains are recognized as a new class of antibody fragment: VHH antibody fragments (also known as nanobodies or single-domain antibodies [sdAbs]; hereafter referred to simply as VHH). These have many advantages over conventional monoclonal antibodies (mAb) and scFv, which has encouraged research into their use possible clinical agents, either in purified form or delivered by VHH-producing live systems.
Advantages of VHH Over Conventional Antibodies
Despite their small size – 12–15 kDa compared to the 150 kDa of traditional antibodies – VHH can recognize and bind to haptens, oligopeptides, and proteins. Indeed, they are the smallest fragments that retain the full antigen-binding capacity of antibodies. Their small size and structural properties allow them to behave like drugs and invest them with unique advantages over mAb for use in biotechnological and biomedical applications (Harmsen and De Haard, 2007; Goldman et al., 2017; Eden et al., 2018). For example, VHH are of low immunogenicity because of their notable sequence similarity with the VHIII subset family of human VH (Cortez-Retamozo et al., 2002; Su et al., 2002; Baral et al., 2006). They also show good solubility and little tendency to aggregate due to the presence of polar and charged amino acid residues (Siontorou, 2013); they exhibit potent-binding capacity (Muyldermans et al., 2001) and have a high affinity for their antigens (Muyldermans et al., 2001). VHH antibody fragments also show remarkably high-conformational stability and can fully reverse the unfolding process induced by denaturing agents and conditions (such as those encountered in the stomach). They can also resist high pressures and urea and guanidinium chloride concentrations (Dumoulin et al., 2002), are extremely stable at high temperatures (below 50 to over 80°C) (van der Linden et al., 1999; Dumoulin et al., 2002), and remain functional under low pH conditions (as low as pH 2) (van der Vaart et al., 2006). Protein engineering has improved the refolding capacity of VHH and therefore their stability under such denaturing conditions (Goldman et al., 2017).
In addition to the conformational stability of VHH, the paratope of these antibody fragments (i.e., the region of the antibody that binds to an epitope) occupies such a small part of the antibody that the recognition of hidden regions of epitopes, hardly accessible to mAb, becomes possible (Muyldermans et al., 2001). Further, and in contrast to mAb, VHH can act as potent and specific enzyme inhibitors since they can access protein cavities and bind to the active site (Lauwereys et al., 1998). Moreover, VHH penetrate tissues much better than mAb and can therefore target solid tumors and metastatic lesions much more effectively (Cortez-Retamozo et al., 2002).
The single-domain nature of VHH allows for the easy cloning of VHH-encoding genes, as well as the generation of multimeric VHH using the same or different building blocks. Multivalent VHH are created when two or more identical target-binding VHH are combined. In addition, multiparatopic or multispecific VHH can be created by combining different VHH targeting different antigens or different epitopes of the same antigen (Els Conrath et al., 2001; Harmsen and De Haard, 2007; Hultberg et al., 2011). Increasing the valency and specificity of VHH enhances their therapeutic efficacy, potency, and cross reactivity over that of monovalent and monospecific VHH (Hultberg et al., 2011; Huet et al., 2014).
These beneficial properties of VHH have boosted their use in the detection of toxins in foods (Xu et al., 2015; Wang et al., 2018), in disease diagnosis (De Meyer et al., 2014; Hu et al., 2017; Leow et al., 2017), in pathogen detection (Saerens et al., 2008; Abbady et al., 2012), and in the treatment of different diseases in humans (Hu et al., 2017) and animals (Harmsen et al., 2009; Liu et al., 2015). The following lines focus on their use in the treatment of non-infectious and infectious diseases of the human GIT.
VHH as Immunotherapeutic Agents Against Diseases of the GIT
VHH have been used to combat GIT inflammatory diseases, infections, and cancers (Steeland et al., 2016).
VHH Against Gastrointestinal Inflammatory Diseases
Over the last decade, interest has increased in developing VHH antibodies that target proteins involved in inflammation as a means of treating gastrointestinal inflammatory diseases, including IBD (Steeland et al., 2016). Bradley et al. (2015) generated a biparatopic VHH against the chemokine receptor CXCR2, a G-protein-coupled receptor that plays an important role in mediating the chemotaxis of neutrophils to sites of inflammation, which is involved in a variety of acute and chronic inflammatory diseases, including IBD, asthma, fibrosis, psoriasis, and rheumatoid arthritis (Stadtmann and Zarbock, 2012). This recognized two different epitopes, allowing it to act as an inverse agonist and inhibitor of CXCR2 signaling in vitro – a promising strategy for treating inflammatory diseases.
VHH Against Gastrointestinal Bacterial and Viral Infections
VHH have also been directed against bacterial pathogens and their toxins. The intensive clinical and agricultural use of antibiotics has favored the emergence of multi-resistant pathogens that cause difficult-to-treat or even un-treatable healthcare-associated and community-acquired infections (Watkins and Bonomo, 2016; Frieri et al., 2017). There is therefore an urgent need to develop non-antibiotic antimicrobial agents to combat these pathogens. VHH have been designed to target different bacterial pathogens of the gut (or their toxins) (Steeland et al., 2016), for example, against S. mutans, the main cause of dental caries (Nakano et al., 2010). The oral administration of VHH targeting streptococcal antigen I/II (SA I/II), a cell wall-anchored adhesin present on the bacterial surface, reduced the development of smooth surface caries in a rat-desalivated caries model, probably by blocking the binding of the bacteria to the enamel pellicle (Kruger et al., 2006). VHH have also been developed against Campylobacter jejuni, a zoonotic pathogen that causes foodborne gastroenteritis worldwide (Kaakoush et al., 2015). An orally administered pentameric VHH directed against the flagella of this pathogen (a major virulence factor) reduced its motility in vitro and significantly reduced colonization in infected chickens (Riazi et al., 2013). VHH have also been raised against H. pylori, a common pathogen that causes gastritis, gastric and duodenal ulcers, gastric cancer, and other gastrointestinal diseases (Venerito et al., 2015). Some VHH target an important virulence factor, in this case the pathogen’s urease enzyme (Ardekani et al., 2013; Hoseinpoor et al., 2014). This hydrolyses urea into ammonia, helping to neutralize the gastric acid in the bacteria’s environment and aiding their colonization of the epithelium (Marshall et al., 1990; Olivera-Severo et al., 2017). These neutralizing VHH were resistant to pepsin and trypsin, an important trait for any VHH to be used in the treatment of H. pylori infection.
Other VHH antibodies have also been generated to target the botulinum neurotoxins of Clostridium botulinum (Dong et al., 2010; Mukherjee et al., 2012; Bakherad et al., 2013; Yao et al., 2017), the shiga toxins produced by shiga toxin-producing E. coli strains (STEC) (Tremblay et al., 2013; Mejias et al., 2016), and toxin A (TcdA), toxin B (TcdB), and binary toxin (CDT) of C. difficile (Hussack et al., 2011; Yang et al., 2014; Unger et al., 2015).
VHH have also shown a promise against human viral diseases (Vanlandschoot et al., 2011; Wu et al., 2017). Some have been designed to block the entry of viruses into the host cells by targeting the viral envelope proteins, while others target essential intracellular viral proteins (Wu et al., 2017). These agents have shown a particular promise against rotavirus and norovirus (Ghosh et al., 2018), the most common causes of severe acute gastroenteritis in children (Riera-Montes et al., 2017). Several VHH generated against Group A rotavirus (RVA) strains were shown to neutralize rotavirus in vitro and provided different degrees of in vivo passive protection against rotavirus challenge when orally administered in a neonatal mouse model (van der Vaart et al., 2006; Garaicoechea et al., 2008; Gómez-Sebastián et al., 2012; Tokuhara et al., 2013; Maffey et al., 2015). They also provided protection against human RVA-induced diarrhea in gnotobiotic piglets (Vega et al., 2013). The therapeutic efficacy of an anti-rotavirus VHH known as ARP1 (ARP: anti-rotavirus protein) has been demonstrated in a randomized placebo-controlled clinical trial (
VHH as Anti-Tumoral Agents
VHH are gaining importance as anti-tumoral agents. Their small size, tumor penetrating capacity, and homogeneous distribution compared to mAb can lead to improved treatment results, and even tumor eradication (Bannas et al., 2015). In addition, the discovery and production costs for VHH are lower than those associated with mAb, perhaps allowing for more affordable and sustainable cancer treatments. The absence in VHH of the fragment crystallizable (Fc) region of IgG, which is present in mAbs, might also reduce the adverse immune responses associated with this domain (Steeland et al., 2016). However, its absence might also reduce the efficacy of VHH in cancer treatment (Steeland et al., 2016; Hu et al., 2017) since it triggers the antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) reactions upon antigen binding; both are important in tumor eradication (Weiner et al., 2010). These findings have together boosted the search for VHH anti-tumoral agents (Oliveira et al., 2013; Kijanka et al., 2015; Steeland et al., 2016; Hu et al., 2017; Iezzi et al., 2018).
The growth of certain gastrointestinal tumors has already been reported to be reduced by some VHH (Cortez-Retamozo et al., 2004; Rozan et al., 2013). VHH have been designed to elicit the ADCC and CDC responses, such as a Fab-like bispecific antibody that comprised a VHH targeting the carcinoembryonic antigen (CEA; a well-characterized tumor marker of interest for mAb-based cancer therapy) and a VHH that specifically bound and activated the FcγRIIIa receptor (an Fc-receptor class with important anti-tumoral effects) (Rozan et al., 2013). This antibody triggered potent in vitro cell-mediated cytotoxicity toward different CEA-positive human tumor cell lines (including human colon adenocarcinoma and primary pancreatic adenocarcinoma lines) in the presence of natural killer cells used as effector cells. Further, it showed potent in vivo activity, inhibiting tumor growth after being injected intraperitoneally together with human peripheral blood mononuclear cells in mice xenografted with a CEA-positive pancreatic cell line.
Other anti-tumoral VHH have been designed to block inhibitory immune checkpoints (Zhang et al., 2017). These are inhibitory pathways that under normal physiological conditions prevent autoimmunity and modulate immune response against microbial infections to prevent tissue damage (Tsai and Hsu, 2017). Some inhibitory immune checkpoints are upregulated in many cancers as a mechanism of tumor resistance to, and evasion of, the host immune system (Zou and Chen, 2008). Antibodies that block immunosuppressive checkpoints are therefore emerging as promising strategies (Pardoll, 2012; La-Beck et al., 2015; Singh et al., 2015; Shi et al., 2018). For example, immunotherapies based on the blockade of PD-1/PD-L1 (PD-1, or programmed death protein-1, is a surface cell receptor present in activated T-cells, and PD-L1, or ligand 1 of PD-1, is abundant in cells of colon cancer and other carcinomas) (Dong et al., 2002; Droeser et al., 2013) using anti-PD-1 (Lipson, 2013; Le et al., 2015, 2017; O’Neil et al., 2017) or anti-PD-L1 mAb (Herbst et al., 2014; Singh et al., 2015) have led to remarkable clinical responses in patients with colorectal cancer. In the context of VHH, Zhang et al. (2017) developed an intraperitoneally administered anti-PD-L1 nanobody fused to the Fc-domain of human IgG1 that efficiently inhibited the PD-1/PD-L1 interaction, induced the T-cell cytokine production in vitro, and caused the in vivo inhibition of tumor growth in a melanoma xenograft mouse model.
Another immunotherapeutic strategy aimed at impeding the growth of gastrointestinal and other tumors is the use of antibodies targeting angiogenic factors. Angiogenesis, a process whereby new blood vessels are formed from existing vessels, is over-induced in tumors. The new vessels that form in the vicinity of the tumor provide it with nutrients and oxygen (Kong et al., 2017). VHH that block overproduced pro-angiogenic factors have been shown effective in preventing tumor angiogenesis and therefore offer a promise as anti-tumoral agents (Arezumand et al., 2017). VHH have been developed that target key regulators of tumor angiogenesis, such as vascular endothelial growth factor (VEGF) (Farajpour et al., 2014; Ebrahimizadeh et al., 2015; Kazemi-Lomedasht et al., 2015), placental growth factor (PLGF) (Arezumand et al., 2016), and vascular endothelial growth factor receptor 2 (VEGFR-2) (Behdani et al., 2012; Ghavamipour et al., 2014; Ma et al., 2016), among others (Arezumand et al., 2017). In addition, they have been directed against hepatocyte growth factor (HGF) and its receptor c-Met since HGF/c-Met-signaling is involved in angiogenesis and the progression of different gastrointestinal cancers, including colon and gastric cancers (Birchmeier et al., 2003; Kijanka et al., 2015; Arezumand et al., 2017). A VHH targeting HGF reduced the growth of solid tumors in mice bearing glioblastoma xenografts (interestingly, some mice were even cured) (Vosjan et al., 2012), while an anti-c-Met VHH efficiently inhibited cell proliferation, adhesion, and in vitro migration in HGF-mediated multiple myeloma (Slordahl et al., 2013). VHH that target angiogenic factors may therefore have a bright future as anti-tumoral/anti-metastatic immunotherapeutic agents.
Lab that Produce VHH
The small size and the hydrophilic and single-domain nature of VHH allow for their easy production in different eukaryotic systems, including yeasts (e.g., Saccharomyces cerevisiae and Pichia pastoris), fungi (e.g., Aspergillus awamori and Aspergillus oryzae), plants (e.g., Nicotiana benthamiana and Arabidopsis thaliana), mammalian cell lines [e.g., Chinese hamster ovary (CHO) cells and mouse B cells], and insect cells (e.g., those of Spodoptera frugiperda and Trichoplusia ni). They can also be produced in prokaryotic microorganisms, including Gram-negative bacteria such as E. coli, Gram-positive bacteria such as Brevibacillus choshinensis, some species of LAB, and species belonging to the genus Bifidobacterium (Harmsen and De Haard, 2007; Liu and Huang, 2018). The eukaryotic systems allow for the production of soluble, functional, and N-glycosylated VHH, although degradation by proteases and inefficient secretion can reduce the yield obtained. It should be remembered, however, that some of these systems, for example, mammalian cells, are quite expensive for use in large-scale production (Liu and Huang, 2018). In addition, they cannot be used for the in situ delivery of the VHH produced. Prokaryotic systems are therefore preferable. These can be genetically manipulated with relative ease and are more economically viable as production systems. E. coli is one of the microbial hosts most widely used for VHH production (Liu and Huang, 2018). However, it has some limitations. The reducing environment of its cytoplasm, for example, drives the incorrect folding of VHH and the formation of insoluble and non-functional aggregates (Liu and Huang, 2018). In addition, the presence of LPS in the bacterial membrane – which is pyrogenic in humans and other mammals (Mamat et al., 2015) – limits its use as a host.
In contrast, Gram-positive bacteria, including the LAB, have no LPS in their cell membranes (Malanovic and Lohner, 2016). This, plus the other characteristics of LAB mentioned earlier, renders this group a better alternative when producing VHH for therapeutic use. This is particularly true when they are to be orally administered as live systems for delivering VHH to the GIT (Pant et al., 2006, 2011; Vandenbroucke et al., 2010; Martin et al., 2011; Alvarez et al., 2015; Andersen et al., 2015).
L. lactis is one of LAB species of choice for such a delivery system. This non-pathogenic bacterium can be safely consumed; indeed, it has been used for centuries in food fermentation and preservation without raising any safety concerns, and it has been granted QPS status by the EFSA (Ricci et al., 2018) and GRAS status by the FDA (Wells and Mercenier, 2008). In addition, it is a non-invasive, non-colonizing bacterium (Steidler et al., 2000), and many genetic engineering tools are available that can induce its production of a wide range of heterologous proteins (Song et al., 2017). These advantages make L. lactis a successful microbial cell factory and have prompted its use as a vehicle for delivering therapeutic molecules directly into the GIT (Song et al., 2017).
Other LAB – specifically the members of the genus Lactobacillus – have been engineered to produce VHH directly in the GIT. These have extra advantages as VHH delivery vehicles. In addition to being safe for consumption, many Lactobacillus strains are normal constituents of the human gut microbiota and can survive passage to the intestine since they are resistant to gastric and bile acids. They also transiently adhere to the mucosal epithelium and have intrinsic immunostimulatory properties (Alvarez-Sieiro et al., 2014; del Rio et al., 2014). Moreover, some Lactobacillus strains have probiotic characteristics and are beneficial to health (Azad et al., 2018). For instance, L. rhamnosus GG, L. casei Shirota YIT9029, and L. casei DN-114001, among others, have antimicrobial effects against gastric and enteric bacterial pathogens and rotavirus (Lievin-Le Moal and Servin, 2014). Moreover, many genetic engineering tools, such as plasmid-based cloning systems, are available to help generate recombinant Lactobacillus strains capable of producing heterologous proteins (Landete, 2017). Other technologies conferring selective advantages over the use of plasmids are also available, such as genome editing for the stable integration of expression cassettes in the bacterial chromosome (based on the integrative machinery of bacteriophages) (Martin et al.1, 2000, 2011; Lin et al., 2013; Alvarez-Sieiro et al., 2014), systems based on integration by double recombination (Gosalbes et al., 2000; Steidler et al., 2003; Douglas and Klaenhammer, 2011), and technologies based on clustered regularly interspaced short palindromic repeats (CRISPR) and associated (Cas) proteins (CRISPR/Cas) (Hidalgo-Cantabrana et al., 2017; van Pijkeren and Barrangou, 2017). Lactobacillus is currently regarded as the most suitable LAB genus for the oral delivery of VHH against diseases of the GIT. Indeed, Lactobacillus paracasei was the first LAB species engineered to produce VHH, which were termed as “lactobodies” (Pant et al., 2006).
The following lines provide a comprehensive review of all the recombinant LAB that have been genetically engineered to deliver VHH antibodies in situ at the GIT mucosa.
LAB That Produce VHH Against Gastrointestinal Inflammatory Diseases
Tumor necrosis factor alpha (TNFα) is one of the major cytokines involved in the pathogenesis of IBD (Billmeier et al., 2016), and immunotherapy with anti-TNFα antibodies is becoming recognized as an efficient treatment (Billmeier et al., 2016). Vandenbroucke et al. (2010) developed recombinant L. lactis strains that secreted monovalent (MT1) and bivalent (MT1-MT1; constructed using two MT1 and a llama IgG2a upper hinge) (Coppieters et al., 2006) VHH against murine TNFα (Figure 1 ). Both the MT1 and (especially) the MT1-MT1 VHH neutralized the soluble and the transmembrane forms of TNFα in vitro. The same study revealed the in vivo therapeutic efficacy of orally administered recombinant L. lactis secreting MT1-MT1 in terms of reducing the symptoms of chronic colitis in two mouse models (DSS-induced colitis and established colitis in IL-10−/− mice). Importantly, the study also showed that the production of MT1-MT1 at the site of inflammation avoided the increase in the incidence of systemic viral and bacterial infections sometimes observed after the intravenous administration of purified anti-TNFα IgG1 antibodies. This result is of great importance since current treatments for Crohn’s disease and ulcerative colitis involve the intravenous administration of anti-TNFα IgG1 antibodies (Colombel et al., 2004; Zabana et al., 2010). The oral administration of anti-TNFα VHH-producing L. lactis provides a way of treating chronic GIT inflammation locally, avoiding the adverse effects associated with systemically administered anti-TNFα antibodies.
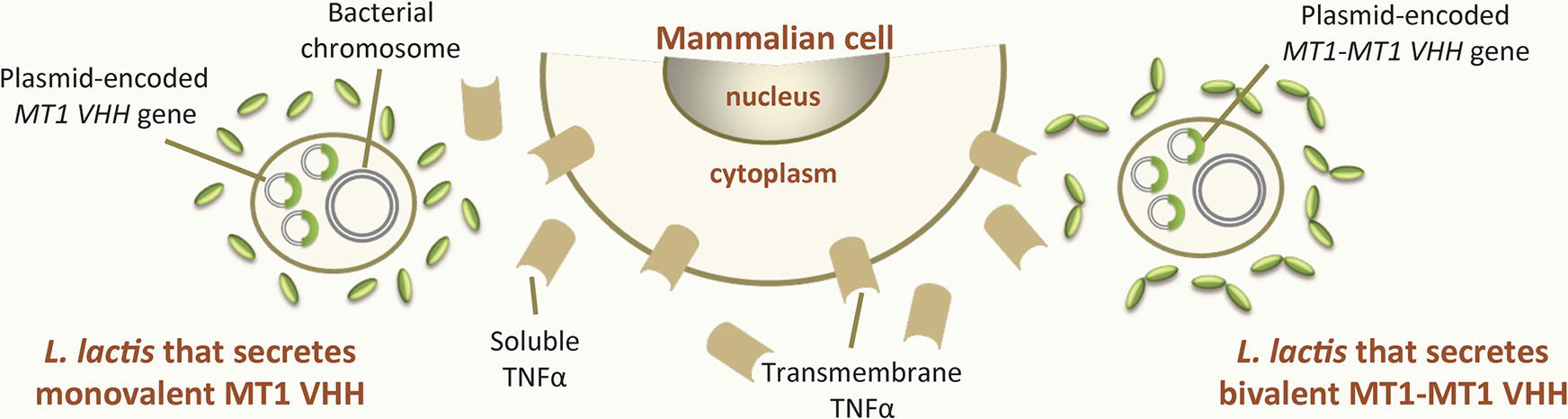
Figure 1. Recombinant LAB that produce VHH against intestinal inflammatory bowel diseases. Recombinant Lactococcus lactis strains secreting either monovalent MT1 or bivalent MT1-MT1 VHH against TNFα, an inflammatory cytokine involved in inflammatory bowel disease. The gene encoding MT1 VHH is constitutively expressed within a replicative plasmid either as a single copy (monovalent form, MT1 VHH) or as two copies in tandem (bivalent form, MTI-MT1 VHH).
LAB That Produce VHH Targeting Bacterial Toxins
Lactobacilli have been engineered to produce VHH to combat gastrointestinal infections caused by C. difficile, the leading cause of nosocomial infections in healthcare settings (Aktories et al., 2017), and of most antibiotic-associated diarrheal disease. It is also a causal agent of pseudomembranous colitis, megacolon, and chronic infections and can even cause death (Aktories et al., 2018). C. difficile produces two secreted enzymatic exotoxins TcdA and TcdB (Aktories et al., 2017); these are major virulence factors of this bacterium and are clearly responsible for associated diarrhea and colitis (Chandrasekaran and Lacy, 2017). TcdA and TcdB exert their effects by binding to surface receptors on intestinal epithelial cells (Aktories et al., 2017).
Andersen et al. (2015) proposed an oral antitoxin strategy for the prevention and treatment of C. difficile-associated diarrhea, based on the neutralization of TcdB using engineered L. paracasei BL23 that constitutively produced anti-TcdB VHH in the GIT (Figure 2 ). These authors generated several recombinant L. paracasei strains that produced neutralizing anti-TcdB VHH that targeted different epitopes of the exotoxin. The bacteria were engineered to either secrete anti-TcdB VHH into the extracellular medium or produce them anchored to the cell wall. Both types neutralized TcdB in vitro. Importantly, the oral administration of a combination of two strains displaying different cell wall-anchored anti-TcdB VHH delayed the development of C. difficile infection and provided partial protection in a hamster protection model (Andersen et al., 2015). These results confirmed the prophylactic effect of such VHH against this pathogen. It should also be noted, however, that the oral administration of purified anti-TcdB VHH produced by a yeast provided no such protection in vivo, probably due to the proteolytic degradation of the antibodies under the harsh conditions of the GIT (Vandenbroucke et al., 2010). These results highlight the advantages of the continuous production of neutralizing VHH anchored to the cell wall and acting in situ in the GIT: this system avoids the degradation suffered by purified VHH and might prevent the diffusion of the toxin by sequestering it on the bacterial surface. Engineered lactobacilli producing surface-exposed anti-TcdB VHH might be used to complement existing therapies designed to tackle C. difficile infection.
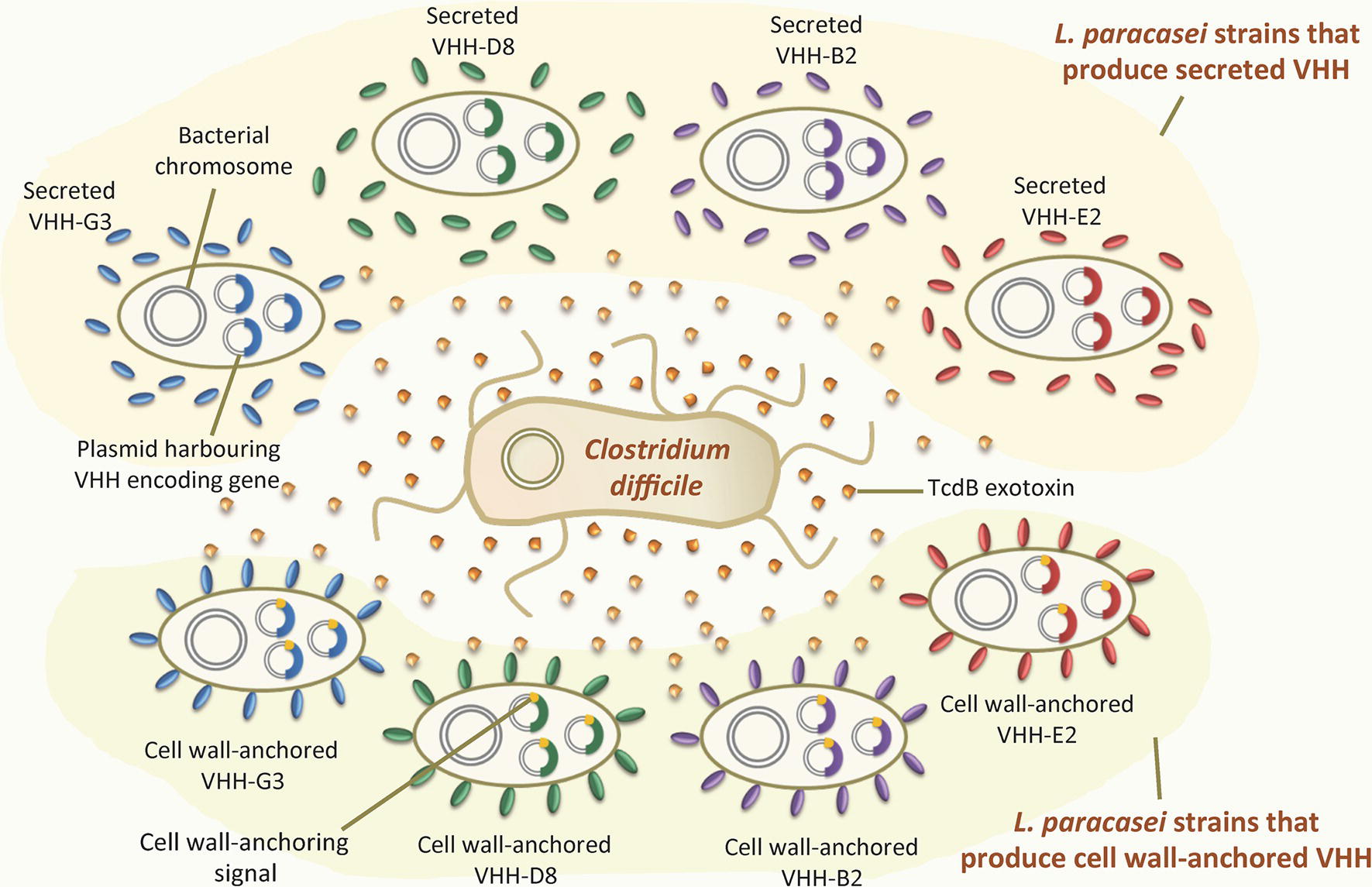
Figure 2. Recombinant LAB that produce VHH against the TcdB exotoxin of Clostridium difficile. Recombinant Lactobacillus paracasei strains producing four different VHH (G3, D8, B2, and E2) against the TcdB exotoxin of Clostridium difficile. Some of these strains secrete the VHH into the extracellular environment, while others produce it covalently attached to the cell wall. The VHH-encoding genes are constitutively expressed within replicative plasmids inside the L. paracasei cells.
LAB That Produce VHH Against Viruses
Most of the lactobacilli engineered to produce VHH have been developed to treat or provide protection against rotavirus infection (Figure 3 ). Pant et al. (2006) proposed an oral prophylactic therapy to prevent rotavirus-associated diarrhea based on engineered Lactobacillus that produced anti-rotavirus VHH in situ in the GIT. This was the first time that lactobacilli were successfully engineered to produce VHH. Using a plasmid expression system, L. paracasei was engineered to constitutively express secreted or cell wall-anchored VHH1 (referred to as anti-rotavirus proteins [ARP1] in the present work), a VHH antibody that targets the RRV strain of rhesus monkey rotavirus serotype G3 (RRV). The study showed that both VHH1-secreting lactobacilli and VHH1-anchored lactobacilli bound and neutralized rotavirus in vitro in a dose-dependent manner. Moreover, VHH1-anchored lactobacilli were able to survive and constitutively produce VHH1 antibodies in situ in the jejunum and ileum after being orally administered to both uninfected and RRV-infected mice. The results highlighted the better performance of VHH1-anchored lactobacilli in preventing rotavirus infection and rotavirus-associated diarrhea in a mouse pup model of rotavirus infection. The good prophylactic effect observed was probably due to the numerous anti-rotavirus VHH antibodies exposed on the bacterial surface; these likely increased affinity for the viral particles, enhanced their agglutination, and facilitated their subsequent clearance. The same study also showed that recombinant VHH1-anchored lactobacilli maintain their protective activity after they are lyophilized and reconstituted. They can therefore be stored in lyophilized format, eliminating the dependence on a cold chain for their preservation, with all the advantages this brings.
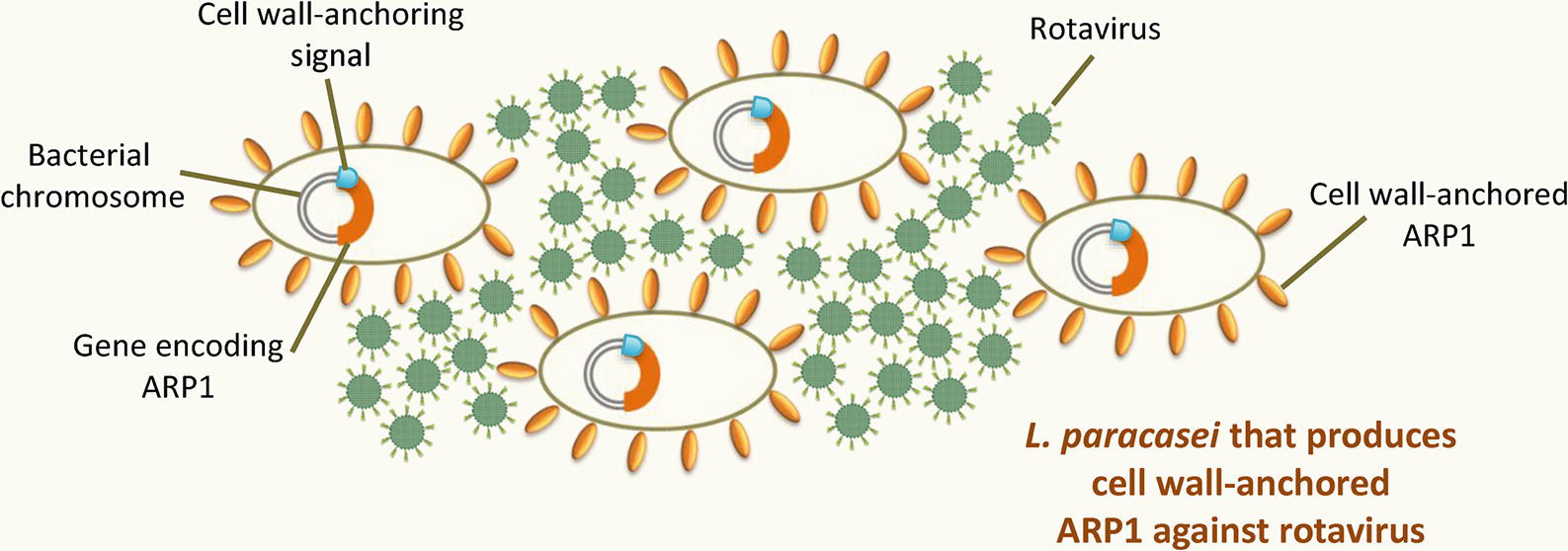
Figure 3. Recombinant LAB that produce VHH against rotavirus. A recombinant Lactobacillus paracasei strain with ARP1, targeting rhesus-monkey rotavirus serotype G3, anchored to its cell wall. The expression cassette harboring the gene encoding ARP1 is fused to a cell wall-anchoring signal sequence and is stably integrated within the bacterial chromosome.
In a later study, Pant et al. (2011) showed that increasing the specificity of anti-rotavirus VHH improves the prophylactic and therapeutic effects of orally administered recombinant lactobacilli. These authors generated different recombinant L. paracasei strains that produced anti-rotavirus VHH with different valency formats (monovalent and bivalent VHH) and specificity (monospecific and bispecific VHH) that recognized different epitopes of the viral particle and that cross-reacted with and cross-neutralized different rotavirus serotypes in vitro (Pant et al., 2011; Aladin et al., 2012). These strains produced either: 1) cell wall-anchored monovalent, monospecific ARP1 (referred to as VHH1 in the original study) or anti-rotavirus protein 3 (ARP3), 2) bivalent, monospecific ARP1-ARP1, ARP3-ARP3, or 3) bivalent, bispecific ARP1-ARP3 proteins. Although all the strains producing anchored VHH bound to the RRV strain in vitro, the bispecific ARP1-ARP3 format returned the best results. In addition, the latter neutralized RRV in vitro and reduced the infection rate. This strain showed the greatest in vivo prophylactic effect after being orally administered in a mouse pup model of rotavirus infection. In addition, in therapeutic use, it most strongly reduced the prevalence, severity, and duration of diarrheal disease. The generation of recombinant lactobacilli producing multivalent and multispecific VHH against rotavirus might improve their prophylactic and therapeutic efficacy against a broad range of viral serotypes.
More recently, Gunaydin et al. (2014) developed several expression cassettes that were cloned in tandem to allow the co-expression of ARP1 and ARP3 as either secreted or cell wall-anchored forms in L. paracasei BL23. Using these cassettes, they generated three recombinant strains: L. paracasei pAF1300 producing secreted ARP1 and anchored ARP3, L. paracasei pAF1400 producing anchored ARP1 and ARP3, and L. paracasei pAF1200 producing secreted ARP1 and ARP3. Those producing ARP1 and ARP3 in anchored form bound a broad range of human and simian rotavirus serotypes with great affinity. Secreted ARP1 produced by L. paracasei pAF1300 and L. paracasei pAF1200 and secreted ARP3 produced by L. paracasei pAF1200 bound to different human rotavirus serotypes and simian rotavirus RRV. Analysis of the mechanism of interaction between rotavirus and L. paracasei pAF1300 suggested that once the virus is captured by anchored ARP3, the bacterium produces secreted ARP1, which targets other rotavirus epitopes. This confirms the feasibility of engineering Lactobacillus to co-produce VHH with different specificities and cell locations, increasing the chances of neutralizing different rotavirus serotypes and reducing the chances of escape mutants appearing. Moreover, the simultaneous production of several VHH with different specificities in one engineered Lactobacillus would considerably reduce production costs compared to preparing cocktails of different recombinant lactobacilli, each producing different VHH.
The recombinant lactobacilli described in the above-mentioned studies relied on plasmid expression systems for the production and delivery of anti-rotavirus VHH. These systems have been widely used to overproduce heterologous proteins in LAB, but they suffer drawbacks that hamper their use as delivery systems for the in situ production of therapeutic proteins in the GIT. For instance, expression plasmids show structural and segregational instability, and their maintenance in cells therefore requires a selection pressure, commonly resistance to antibiotics. The presence of antibiotic resistance genes in therapeutic recombinant lactobacilli, however, is undesirable since antibiotics would be needed to select VHH-producing strains in the intestine. In addition, they would increase the risk of antibiotic resistance genes being horizontally transferred to other, perhaps pathogenic, GIT bacteria.
Given the above, different approaches have been proposed to maintain heterologous genes in recombinant LAB. A feasible alternative is the use of food-grade expression vectors with selection markers other than antibiotics that provide new phenotypes (e.g., immunity to bacteriocins, utilization of rare sugars, etc.) or complement specific mutations that restore impaired functions (e.g., complementation of auxotrophic mutants depending on threonine, thymidine, alanine, lactose, etc.) (Landete, 2017). But in any case, it is necessary to maintain a selective pressure in the GIT, which can be avoided by the chromosomal integration of the heterologous genes. Martin et al. (2011) developed a universal integrative expression system that allows the stable integration of expression cassettes for the production of scFv and VHH in the chromosome of Lactobacillus. This system is based on an ingenious genome editing technology, developed by Martin et al. (2000), that involves the site-specific integration of heterologous genes into the LAB chromosome, plus a purification system that – once the vector is integrated into the chromosome – leads to the elimination of all non-food-grade deoxyribonucleic acid, including the antibiotic selection genes. This system allowed the generation of different food-grade recombinant L. paracasei strains that stably produce VHH against rotavirus (Martin et al., 2011). The recombinant L. paracasei EM233 strain, which stably produces cell wall-anchored ARP1 (Figure 3 ), was shown to effectively bind rotavirus in vitro, and importantly, the integrated gene encoding APR1 VHH antibody was stable with no need for any selective pressure. After its oral administration, the recombinant L. paracasei strain producing anchored ARP1 showed prophylactic efficacy, reducing the duration and severity of diarrhea in a mouse model of rotavirus infection. More recently, L. rhamnosus GG was genetically engineered using this system to stably produce cell wall-anchored ARP1 antibody against rotavirus (Alvarez et al., 2015). L. rhamnosus GG is a well-known probiotic strain that, in addition to having intrinsic anti-rotavirus activity (Lievin-Le Moal and Servin, 2014; Guarino et al., 2015), helps reinforce the intestinal barrier, triggers the innate and adaptive immune responses (Sindhu et al., 2014), and transiently colonizes the intestine (Alander et al., 1999). It is therefore an ideal vehicle for the production of VHH against rotavirus in the GIT mucosa. Alvarez et al. (2015) tested the capacity of several L. rhamnosus GG strains to display anchored ARP1 on their surfaces for which they used the plasmid-based expression system described above (Martin et al., 2011). Although all these strains produced ARP1, only one, a naturally occurring exopolysaccharide-deficient mutant strain named L. rhamnosus GG (UT), efficiently produced anchored ARP1. Genomic analysis suggested that two genes (welE and welF) belonging to the exopolysaccharide (EPS) cluster were inactive in this strain, which impaired the production of cell-bound EPS. The EPS deficiency allowed the appearance of the anchored ARP1; in all the other strains tested the thick layer of EPS masked it. L. rhamnosus GG (UT) was further genetically engineered to produce ARP1 anchored to the cell wall through the use of the above-described integrative-expression system (Martin et al., 2011). This strain, known as L. rhamnosus GG EM233, bounded rotavirus particles and seemed to confer a protective effect in vivo; when it was orally administered to mice before they were infected with rotavirus, the prevalence, duration, and severity of diarrhea were reduced compared to the control group (Alvarez et al., 2015).
Thus, the oral administration of engineered lactobacilli that either secrete VHH or display cell wall-anchored VHH (with single or multiple valencies and specificities) may be a way of preventing or treating rotavirus infection and its associated diarrhea. The site-specific integration expression technology used in the above studies represents an important advance in the genetic manipulation of LAB for the generation of food-grade, live recombinant bacteria that produce prophylactic and therapeutic VHH in situ in the GIT.
Concluding Remarks and Future Perspectives
Over the last decade, LAB have emerged as powerful oral delivery vectors for the in situ production of VHH at the GIT mucosa where they may act as therapeutic agents or provide passive protection against different gastrointestinal diseases. Specific LAB strains have been genetically modified to either secrete VHH or produce them anchored to their surfaces. This has been achieved using expression systems based on replicative plasmids, which require a selection pressure such as the presence of antibiotic resistance genes for their maintenance in bacteria (Pant et al., 2006, 2011; Vandenbroucke et al., 2010; Gunaydin et al., 2014; Andersen et al., 2015) or by using other genetic tools that allow the integration of VHH-encoding genes into the bacterial chromosome. This provides stability to the integrated gene without the need for a selection pressure system (Martin et al., 2011; Alvarez et al., 2015). In the near future, other powerful chromosome editing technologies, such as those based on the CRISPR/Cas systems (Hidalgo-Cantabrana et al., 2017), might also be used to generate recombinant LAB that stably produce VHH against diseases of the GIT. At the time of writing, only L. lactis (Vandenbroucke et al., 2010), L. paracasei (Pant et al., 2006, 2011; Martin et al., 2011; Gunaydin et al., 2014; Andersen et al., 2015), and L. rhamnosus (Alvarez et al., 2015) have been engineered to produce VHH for treating or preventing GIT disease. Many LAB strains have been described as possessing probiotic characteristics of benefit against IBD (Saez-Lara et al., 2015), and against the bacteria (Goderska et al., 2018; Lin et al., 2018; McFarland et al., 2018) and viruses (Guarino et al., 2015) that cause gastrointestinal infections. Some of them even show anti-tumoral properties against colorectal cancer (Ambalam et al., 2016). Based on their intrinsic benefits, specific probiotic LAB strains could be selected and genetically modified to produce therapeutic or prophylactic VHH. This would result in a synergistic effect against the target disease. So far, recombinant LAB that produce VHH have been directed to prevent or treat IBD (Vandenbroucke et al., 2010) and infections caused by C. difficile (Andersen et al., 2015) and rotavirus (Pant et al., 2006, 2011; Martin et al., 2011; Gunaydin et al., 2014; Alvarez et al., 2015). Given the great potential of VHH as therapeutic and prophylactic agents, many other LAB might be engineered to produce VHH targeting other diseases of the GIT, such as infections caused by pathogenic bacteria (e.g., C. jejuni, H. pylori, ETEC, STEC, Salmonella, etc.), viruses (e.g., norovirus), fungi (e.g., C. albicans), and intestinal parasites and to provide relief from food allergies and intolerances, inflammatory diseases, and even gastrointestinal cancers. They might also be produced to treat disease in other mucous membranes inhabited by LAB.
Data Availability
No datasets were generated or analyzed for this study.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work was funded by the Spanish Ministry of Economy and Competitiveness (AGL2016-78708-R).
Acknowledgments
The authors thank Adrian Burton for linguistic assistance. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
References
Abbady, A. Q., Al-Daoude, A., Al-Mariri, A., Zarkawi, M., and Muyldermans, S. (2012). Chaperonin GroEL a Brucella immunodominant antigen identified using nanobody and MALDI-TOF-MS technologies. Vet. Immunol. Immunopathol. 146, 254–263. doi: 10.1016/j.vetimm.2012.01.015
Adel-Patient, K., Ah-Leung, S., Creminon, C., Nouaille, S., Chatel, J. M., Langella, P. , et al. (2005). Oral administration of recombinant Lactococcus lactis expressing bovine beta-lactoglobulin partially prevents mice from sensitization. Clin. Exp. Allergy 35, 539–546. doi: 10.1111/j.1365-2222.2005.02225.x
Ahmed, B., Loos, M., Vanrompay, D., and Cox, E. (2014). Oral immunization with Lactococcus lactis-expressing EspB induces protective immune responses against Escherichia coli O157:H7 in a murine model of colonization. Vaccine 32, 3909–3916. doi: 10.1016/j.vaccine.2014.05.054
Aktories, K., Papatheodorou, P., and Schwan, C. (2018). Binary Clostridium difficile toxin (CDT)—A virulence factor disturbing the cytoskeleton. Anaerobe 53, 21–29. doi: 10.1016/j.anaerobe.2018.03.001
Aktories, K., Schwan, C., and Jank, T. (2017). Clostridium difficile toxin biology. Annu. Rev. Microbiol. 71, 281–307. doi: 10.1146/annurev-micro-090816-093458
Aladin, F., Einerhand, A. W., Bouma, J., Bezemer, S., Hermans, P., Wolvers, D. , et al. (2012). In vitro neutralisation of rotavirus infection by two broadly specific recombinant monovalent llama-derived antibody fragments. PLoS One 7:e32949. doi: 10.1371/journal.pone.0032949
Alander, M., Satokari, R., Korpela, R., Saxelin, M., Vilpponen-Salmela, T., Mattila-Sandholm, T. , et al. (1999). Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl. Environ. Microbiol. 65, 351–354.
Alvarez, B., Krogh-Andersen, K., Tellgren-Roth, C., Martinez, N., Gunaydin, G., Lin, Y. , et al. (2015). An exopolysaccharide-deficient mutant of Lactobacillus rhamnosus GG efficiently displays a protective llama antibody fragment against rotavirus on its surface. Appl. Environ. Microbiol. 81, 5784–5793. doi: 10.1128/aem.00945-15
Alvarez-Sieiro, P., Martin, M. C., Redruello, B., del Rio, B., Ladero, V., Palanski, B. A. , et al. (2014). Generation of food-grade recombinant Lactobacillus casei delivering Myxococcus xanthus prolyl endopeptidase. Appl. Microbiol. Biotechnol. 98, 6689–6700. doi: 10.1007/s00253-014-5730-7
Ambalam, P., Raman, M., Purama, R. K., and Doble, M. (2016). Probiotics, prebiotics and colorectal cancer prevention. Best Pract. Res. Clin. Gastroenterol. 30, 119–131. doi: 10.1016/j.bpg.2016.02.009
Andersen, K. K., Marcotte, H., Alvarez, B., Boyaka, P. N., and Hammarstrom, L. (2011). In situ gastrointestinal protection against anthrax edema toxin by single-chain antibody fragment producing lactobacilli. BMC Biotechnol. 11:126. doi: 10.1186/1472-6750-11-126
Andersen, K. K., Strokappe, N. M., Hultberg, A., Truusalu, K., Smidt, I., Mikelsaar, R. H. , et al. (2015). Neutralization of Clostridium difficile toxin B mediated by engineered lactobacilli producing single domain antibodies. Infect. Immun. 84, 395–406. doi: 10.1128/iai.00870-15
Ardekani, L. S., Gargari, S. L., Rasooli, I., Bazl, M. R., Mohammadi, M., Ebrahimizadeh, W. , et al. (2013). A novel nanobody against urease activity of Helicobacter pylori. Int. J. Infect. Dis. 17, e723–728. doi: 10.1016/j.ijid.2013.02.015
Arezumand, R., Alibakhshi, A., Ranjbari, J., Ramazani, A., and Muyldermans, S. (2017). Nanobodies as novel agents for targeting angiogenesis in solid cancers. Front. Immunol. 8:1746. doi: 10.3389/fimmu.2017.01746
Arezumand, R., Mahdian, R., Zeinali, S., Hassanzadeh-Ghassabeh, G., Mansouri, K., Khanahmad, H. , et al. (2016). Identification and characterization of a novel nanobody against human placental growth factor to modulate angiogenesis. Mol. Immunol. 78, 183–192. doi: 10.1016/j.molimm.2016.09.012
Azad, M. A. K., Sarker, M., Li, T., and Yin, J. (2018). Probiotic species in the modulation of gut microbiota: an overview. Biomed. Res. Int. 2018:9478630. doi: 10.1155/2018/9478630
Bakherad, H., Mousavi Gargari, S. L., Rasooli, I., Rajabibazl, M., Mohammadi, M., Ebrahimizadeh, W. , et al. (2013). In vivo neutralization of botulinum neurotoxins serotype E with heavy-chain camelid antibodies (VHH). Mol. Biotechnol. 55, 159–167. doi: 10.1007/s12033-013-9669-1
Bannas, P., Lenz, A., Kunick, V., Well, L., Fumey, W., Rissiek, B. , et al. (2015). Molecular imaging of tumors with nanobodies and antibodies: timing and dosage are crucial factors for improved in vivo detection. Contrast Media Mol. Imaging 10, 367–378. doi: 10.1002/cmmi.1637
Baral, T. N., Magez, S., Stijlemans, B., Conrath, K., Vanhollebeke, B., Pays, E. , et al. (2006). Experimental therapy of African trypanosomiasis with a nanobody-conjugated human trypanolytic factor. Nat. Med. 12, 580–584. doi: 10.1038/nm1395
Behdani, M., Zeinali, S., Khanahmad, H., Karimipour, M., Asadzadeh, N., Azadmanesh, K. , et al. (2012). Generation and characterization of a functional nanobody against the vascular endothelial growth factor receptor-2; angiogenesis cell receptor. Mol. Immunol. 50, 35–41. doi: 10.1016/j.molimm.2011.11.013
Behnsen, J., Deriu, E., Sassone-Corsi, M., and Raffatellu, M. (2013). Probiotics: properties, examples, and specific applications. Cold Spring Harb. Perspect. Med. 3:a010074. doi: 10.1101/cshperspect.a010074
Benbouziane, B., Ribelles, P., Aubry, C., Martin, R., Kharrat, P., Riazi, A. , et al. (2013). Development of a stress-inducible controlled expression (SICE) system in Lactococcus lactis for the production and delivery of therapeutic molecules at mucosal surfaces. J. Biotechnol. 168, 120–129. doi: 10.1016/j.jbiotec.2013.04.019
Bermudez-Humaran, L. G., Kharrat, P., Chatel, J. M., and Langella, P. (2011). Lactococci and lactobacilli as mucosal delivery vectors for therapeutic proteins and DNA vaccines. Microb. Cell Fact. 10 (Suppl 1):S4. doi: 10.1186/1475-2859-10-S1-S4
Bermudez-Humaran, L. G., Motta, J. P., Aubry, C., Kharrat, P., Rous-Martin, L., Sallenave, J. M. , et al. (2015). Serine protease inhibitors protect better than IL-10 and TGF-beta anti-inflammatory cytokines against mouse colitis when delivered by recombinant lactococci. Microb. Cell Fact. 14:26. doi: 10.1186/s12934-015-0198-4
Bernardeau, M., Vernoux, J. P., Henri-Dubernet, S., and Gueguen, M. (2008). Safety assessment of dairy microorganisms: the Lactobacillus genus. Int. J. Food Microbiol. 126, 278–285. doi: 10.1016/j.ijfoodmicro.2007.08.015
Billmeier, U., Dieterich, W., Neurath, M. F., and Atreya, R. (2016). Molecular mechanism of action of anti-tumor necrosis factor antibodies in inflammatory bowel diseases. World J. Gastroenterol. 22, 9300–9313. doi: 10.3748/wjg.v22.i42.9300
Birchmeier, C., Birchmeier, W., Gherardi, E., and Vande Woude, G. F. (2003). Met, metastasis, motility and more. Nat. Rev. Mol. Cell Biol. 4, 915–925. doi: 10.1038/nrm1261
Braat, H., Rottiers, P., Hommes, D. W., Huyghebaert, N., Remaut, E., Remon, J. P. , et al. (2006). A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin. Gastroenterol. Hepatol. 4, 754–759. doi: 10.1016/j.cgh.2006.03.028
Bradley, M. E., Dombrecht, B., Manini, J., Willis, J., Vlerick, D., De Taeye, S. , et al. (2015). Potent and efficacious inhibition of CXCR2 signaling by biparatopic nanobodies combining two distinct modes of action. Mol. Pharmacol. 87, 251–262. doi: 10.1124/mol.114.094821
Cano-Garrido, O., Seras-Franzoso, J., and Garcia-Fruitos, E. (2015). Lactic acid bacteria: reviewing the potential of a promising delivery live vector for biomedical purposes. Microb. Cell Fact. 14:137. doi: 10.1186/s12934-015-0313-6
Chancey, C. J., Khanna, K. V., Seegers, J. F., Zhang, G. W., Hildreth, J., Langan, A. , et al. (2006). Lactobacilli-expressed single-chain variable fragment (scFv) specific for intercellular adhesion molecule 1 (ICAM-1) blocks cell-associated HIV-1 transmission across a cervical epithelial monolayer. J. Immunol. 176, 5627–5636. doi: 10.4049/jimmunol.176.9.5627
Chandrasekaran, R., and Lacy, D. B. (2017). The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 41, 723–750. doi: 10.1093/femsre/fux048
Colombel, J. F., Loftus, E. V. Jr., Tremaine, W. J., Egan, L. J., Harmsen, W. S., Schleck, C. D. , et al. (2004). The safety profile of infliximab in patients with Crohn’s disease: the Mayo clinic experience in 500 patients. Gastroenterology 126, 19–31. doi: 10.1053/j.gastro.2003.10.047
Coppieters, K., Dreier, T., Silence, K., de Haard, H., Lauwereys, M., Casteels, P. , et al. (2006). Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis. Arthritis Rheum. 54, 1856–1866. doi: 10.1002/art.21827
Cortes-Perez, N. G., Bermudez-Humaran, L. G., Le Loir, Y., Rodriguez-Padilla, C., Gruss, A., Saucedo-Cardenas, O. , et al. (2003). Mice immunization with live lactococci displaying a surface anchored HPV-16 E7 oncoprotein. FEMS Microbiol. Lett. 229, 37–42. doi: 10.1016/S0378-1097(03)00778-X
Cortez-Retamozo, V., Backmann, N., Senter, P. D., Wernery, U., De Baetselier, P., Muyldermans, S. , et al. (2004). Efficient cancer therapy with a nanobody-based conjugate. Cancer Res. 64, 2853–2857. doi: 10.1158/0008-5472.CAN-03-3935
Cortez-Retamozo, V., Lauwereys, M., Hassanzadeh Gh, G., Gobert, M., Conrath, K., Muyldermans, S. , et al. (2002). Efficient tumor targeting by single-domain antibody fragments of camels. Int. J. Cancer 98, 456–462. doi: 10.1002/ijc.10212
Daniel, C., Roussel, Y., Kleerebezem, M., and Pot, B. (2011). Recombinant lactic acid bacteria as mucosal biotherapeutic agents. Trends Biotechnol. 29, 499–508. doi: 10.1016/j.tibtech.2011.05.002
De Meyer, T., Muyldermans, S., and Depicker, A. (2014). Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 32, 263–270. doi: 10.1016/j.tibtech.2014.03.001
de Ruyter, P. G., Kuipers, O. P., and de Vos, W. M. (1996). Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol. 62, 3662–3667.
del Rio, B., Dattwyler, R. J., Aroso, M., Neves, V., Meirelles, L., Seegers, J. F. , et al. (2008). Oral immunization with recombinant Lactobacillus plantarum induces a protective immune response in mice with Lyme disease. Clin. Vaccine Immunol. 15, 1429–1435. doi: 10.1128/CVI.00169-08
del Rio, B., Fuente, J. L., Neves, V., Dattwyler, R., Seegers, J. F., and Gomes-Solecki, M. (2010). Platform technology to deliver prophylactic molecules orally: an example using the Class A select agent Yersinia pestis. Vaccine 28, 6714–6722. doi: 10.1016/j.vaccine.2010.07.084
del Rio, B., Martín, M. C., Sarmiento, M. E., Acosta, A., and Alvarez, M. A. (2014). “Lactobacilli as live vectors for oral vaccination against TB,” in The art and science of tuberculosis vaccine development . eds. Norazmi, M. N., Acosta, A., and Sarmiento, M. E. (Shah Alam, Selangor, Malaysia: Oxford University Press), 794–811.
Desvaux, M., Dumas, E., Chafsey, I., and Hebraud, M. (2006). Protein cell surface display in Gram-positive bacteria: from single protein to macromolecular protein structure. FEMS Microbiol. Lett. 256, 1–15. doi: 10.1111/j.1574-6968.2006.00122.x
Dieye, Y., Usai, S., Clier, F., Gruss, A., and Piard, J. C. (2001). Design of a protein-targeting system for lactic acid bacteria. J. Bacteriol. 183, 4157–4166. doi: 10.1128/jb.183.14.4157-4166.2001
Ding, C., Ma, J., Dong, Q., and Liu, Q. (2018). Live bacterial vaccine vector and delivery strategies of heterologous antigen: a review. Immunol. Lett. 197, 70–77. doi: 10.1016/j.imlet.2018.03.006
Dong, H., Strome, S. E., Salomao, D. R., Tamura, H., Hirano, F., Flies, D. B. , et al. (2002). Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat. Med. 8, 793–800. doi: 10.1038/nm730
Dong, J., Thompson, A. A., Fan, Y., Lou, J., Conrad, F., Ho, M. , et al. (2010). A single-domain llama antibody potently inhibits the enzymatic activity of botulinum neurotoxin by binding to the non-catalytic alpha-exosite binding region. J. Mol. Biol. 397, 1106–1118. doi: 10.1016/j.jmb.2010.01.070
Douglas, G. L., and Klaenhammer, T. R. (2011). Directed chromosomal integration and expression of the reporter gene gusA3 in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 77, 7365–7371. doi: 10.1128/aem.06028-11
Droeser, R. A., Hirt, C., Viehl, C. T., Frey, D. M., Nebiker, C., Huber, X. , et al. (2013). Clinical impact of programmed cell death ligand 1 expression in colorectal cancer. Eur. J. Cancer 49, 2233–2242. doi: 10.1016/j.ejca.2013.02.015
Dumoulin, M., Conrath, K., Van Meirhaeghe, A., Meersman, F., Heremans, K., Frenken, L. G. , et al. (2002). Single-domain antibody fragments with high conformational stability. Protein Sci. 11, 500–515. doi: 10.1110/ps.34602
Ebrahimizadeh, W., Mousavi Gargari, S. L., Javidan, Z., and Rajabibazl, M. (2015). Production of novel VHH nanobody inhibiting angiogenesis by targeting binding site of VEGF. Appl. Biochem. Biotechnol. 176, 1985–1995. doi: 10.1007/s12010-015-1695-y
Eden, T., Menzel, S., Wesolowski, J., Bergmann, P., Nissen, M., Dubberke, G. , et al. (2018). A cDNA immunization strategy to generate nanobodies against membrane proteins in native conformation. Front. Immunol. 8:1989. doi: 10.3389/fimmu.2017.01989
Els Conrath, K., Lauwereys, M., Wyns, L., and Muyldermans, S. (2001). Camel single-domain antibodies as modular building units in bispecific and bivalent antibody constructs. J. Biol. Chem. 276, 7346–7350. doi: 10.1074/jbc.M007734200
FAO/WHO. (2006). Probiotics in food. Health and nutritional properties and guidelines for evaluation . Rome: FAO Food Nutrition.
Farajpour, Z., Rahbarizadeh, F., Kazemi, B., and Ahmadvand, D. (2014). A nanobody directed to a functional epitope on VEGF, as a novel strategy for cancer treatment. Biochem. Biophys. Res. Commun. 446, 132–136. doi: 10.1016/j.bbrc.2014.02.069
Foligne, B., Dessein, R., Marceau, M., Poiret, S., Chamaillard, M., Pot, B. , et al. (2007). Prevention and treatment of colitis with Lactococcus lactis secreting the immunomodulatory Yersinia LcrV protein. Gastroenterology 133, 862–874. doi: 10.1053/j.gastro.2007.06.018
Fredriksen, L., Kleiveland, C. R., Hult, L. T. O., Lea, T., Nygaard, C. S., Eijsink, V. G. H. , et al. (2012). Surface display of N-terminally anchored invasin by Lactobacillus plantarum activates NF-κB in monocytes. Appl. Environ. Microbiol. 78, 5864–5871. doi: 10.1128/AEM.01227-12
Fredriksen, L., Mathiesen, G., Sioud, M., and Eijsink, V. G. H. (2010). Cell wall anchoring of the 37-kilodalton oncofetal antigen by Lactobacillus plantarum for mucosal cancer vaccine delivery. Appl. Environ. Microbiol. 76, 7359–7362. doi: 10.1128/AEM.01031-10
Frieri, M., Kumar, K., and Boutin, A. (2017). Antibiotic resistance. J. Infect. Public Health 10, 369–378. doi: 10.1016/j.jiph.2016.08.007
Garaicoechea, L., Aguilar, A., Parra, G. I., Bok, M., Sosnovtsev, S. V., Canziani, G. , et al. (2015). Llama nanoantibodies with therapeutic potential against human norovirus diarrhea. PLoS One 10:e0133665. doi: 10.1371/journal.pone.0133665
Garaicoechea, L., Olichon, A., Marcoppido, G., Wigdorovitz, A., Mozgovoj, M., Saif, L. , et al. (2008). Llama-derived single-chain antibody fragments directed to rotavirus VP6 protein possess broad neutralizing activity in vitro and confer protection against diarrhea in mice. J. Virol. 82, 9753–9764. doi: 10.1128/jvi.00436-08
García-Fruitós, E. (2012). Lactic acid bacteria: a promising alternative for recombinant protein production. Microb. Cell Fact. 11, 157–157. doi: 10.1186/1475-2859-11-157
Gareau, M. G., Sherman, P. M., and Walker, W. A. (2010). Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 7, 503–514. doi: 10.1038/nrgastro.2010.117
Gerritsen, J., Smidt, H., Rijkers, G. T., and de Vos, W. M. (2011). Intestinal microbiota in human health and disease: the impact of probiotics. Genes Nutr. 6, 209–240. doi: 10.1007/s12263-011-0229-7
Ghavamipour, F., Shahangian, S. S., Sajedi, R. H., Arab, S. S., Mansouri, K., and Aghamaali, M. R. (2014). Development of a highly-potent anti-angiogenic VEGF8-109 heterodimer by directed blocking of its VEGFR-2 binding site. FEBS J. 281, 4479–4494. doi: 10.1111/febs.12956
Ghosh, S., Malik, Y. S., and Kobayashi, N. (2018). Therapeutics and immunoprophylaxis against noroviruses and rotaviruses: the past, present, and future. Curr. Drug Metab. 19, 170–191. doi: 10.2174/1389200218666170912161449
Giomarelli, B., Maggi, T., Younson, J., Kelly, C., and Pozzi, G. (2004). Expression of a functional single-chain Fv antibody on the surface of Streptococcus gordonii. Mol. Biotechnol. 28, 105–112. doi: 10.1385/MB:28:2:105
Goderska, K., Agudo Pena, S., and Alarcon, T. (2018). Helicobacter pylori treatment: antibiotics or probiotics. Appl. Microbiol. Biotechnol. 102, 1–7. doi: 10.1007/s00253-017-8535-7
Goldman, E. R., Liu, J. L., Zabetakis, D., and Anderson, G. P. (2017). Enhancing stability of camelid and shark single domain antibodies: an overview. Front. Immunol. 8:865. doi: 10.3389/fimmu.2017.00865
Gomes-Santos, A. C., de Oliveira, R. P., Moreira, T. G., Castro-Junior, A. B., Horta, B. C., Lemos, L. , et al. (2017). Hsp65-producing Lactococcus lactis prevents inflammatory intestinal disease in mice by IL-10- and TLR2-dependent pathways. Front. Immunol. 8:30. doi: 10.3389/fimmu.2017.00030
Gómez-Sebastián, S., Nuñez, M. C., Garaicoechea, L., Alvarado, C., Mozgovoj, M., Lasa, R. , et al. (2012). Rotavirus A-specific single-domain antibodies produced in baculovirus-infected insect larvae are protective in vivo. BMC Biotechnol. 12, 59–59. doi: 10.1186/1472-6750-12-59
Gosalbes, M. J., Esteban, C. D., Galan, J. L., and Perez-Martinez, G. (2000). Integrative food-grade expression system based on the lactose regulon of Lactobacillus casei. Appl. Environ. Microbiol. 66, 4822–4828. doi: 10.1128/AEM.66.11.4822-4828.2000
Gu, Q., Song, D., and Zhu, M. (2009). Oral vaccination of mice against Helicobacter pylori with recombinant Lactococcus lactis expressing urease subunit B. FEMS Immunol. Med. Microbiol. 56, 197–203. doi: 10.1111/j.1574-695X.2009.00566.x
Guarino, A., Guandalini, S., and Lo Vecchio, A. (2015). Probiotics for prevention and treatment of diarrhea. J. Clin. Gastroenterol. 49 (Suppl 1), S37–45. doi: 10.1097/mcg.0000000000000349
Gunaydin, G., Alvarez, B., Lin, Y., Hammarstrom, L., and Marcotte, H. (2014). Co-expression of anti-rotavirus proteins (llama VHH antibody fragments) in Lactobacillus: development and functionality of vectors containing two expression cassettes in tandem. PLoS One 9:e96409. doi: 10.1371/journal.pone.0096409
Guo, S., Yan, W., McDonough, S. P., Lin, N., Wu, K. J., He, H. , et al. (2015). The recombinant Lactococcus lactis oral vaccine induces protection against C. difficile spore challenge in a mouse model. Vaccine 33, 1586–1595. doi: 10.1016/j.vaccine.2015.02.006
Hamers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hamers, C., Songa, E. B. , et al. (1993). Naturally occurring antibodies devoid of light chains. Nature 363, 446–448. doi: 10.1038/363446a0
Hanniffy, S., Wiedermann, U., Repa, A., Mercenier, A., Daniel, C., Fioramonti, J. , et al. (2004). Potential and opportunities for use of recombinant lactic acid bacteria in human health. Adv. Appl. Microbiol. 56, 1–64. doi: 10.1016/s0065-2164(04)56001-x
Hanson, M. L., Hixon, J. A., Li, W., Felber, B. K., Anver, M. R., Stewart, C. A. , et al. (2014). Oral delivery of IL-27 recombinant bacteria attenuates immune colitis in mice. Gastroenterology 146, 210–221.e213. doi: 10.1053/j.gastro.2013.09.060
Harmsen, M. M., and De Haard, H. J. (2007). Properties, production, and applications of camelid single-domain antibody fragments. Appl. Microbiol. Biotechnol. 77, 13–22. doi: 10.1007/s00253-007-1142-2
Harmsen, M. M., Fijten, H. P., Engel, B., Dekker, A., and Eble, P. L. (2009). Passive immunization with llama single-domain antibody fragments reduces foot-and-mouth disease transmission between pigs. Vaccine 27, 1904–1911. doi: 10.1016/j.vaccine.2009.01.110
Herbst, R. S., Soria, J.-C., Kowanetz, M., Fine, G. D., Hamid, O., Gordon, M. S. , et al. (2014). Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 515, 563–567. doi: 10.1038/nature14011
Hidalgo-Cantabrana, C., O’Flaherty, S., and Barrangou, R. (2017). CRISPR-based engineering of next-generation lactic acid bacteria. Curr. Opin. Microbiol. 37, 79–87. doi: 10.1016/j.mib.2017.05.015
Hoang, P. M., Cho, S., Kim, K. E., Byun, S. J., Lee, T. K., and Lee, S. (2015). Development of Lactobacillus paracasei harboring nucleic acid-hydrolyzing 3D8 scFv as a preventive probiotic against murine norovirus infection. Appl. Microbiol. Biotechnol. 99, 2793–2803. doi: 10.1007/s00253-014-6257-7
Hols, P., Slos, P., Dutot, P., Reymund, J., Chabot, P., Delplace, B. , et al. (1997). Efficient secretion of the model antigen M6-gp41E in Lactobacillus plantarum NCIMB 8826. Microbiologica 143, 2733–2741. doi: 10.1099/00221287-143-8-2733
Hongying, F., Xianbo, W., Fang, Y., Yang, B., and Beiguo, L. (2014). Oral immunization with recombinant Lactobacillus acidophilus expressing the adhesin Hp0410 of Helicobacter pylori induces mucosal and systemic immune responses. Clin. Vaccine Immunol. 21, 126–132. doi: 10.1128/cvi.00434-13
Hoseinpoor, R., Mousavi Gargari, S. L., Rasooli, I., Rajabibazl, M., and Shahi, B. (2014). Functional mutations in and characterization of VHH against Helicobacter pylori urease. Appl. Biochem. Biotechnol. 172, 3079–3091. doi: 10.1007/s12010-014-0750-4
Hosseinidoust, Z., Mostaghaci, B., Yasa, O., Park, B.-W., Singh, A. V., and Sitti, M. (2016). Bioengineered and biohybrid bacteria-based systems for drug delivery. Adv. Drug Deliv. Rev. 106, 27–44. doi: 10.1016/j.addr.2016.09.007
Hu, S., Kong, J., Sun, Z., Han, L., Kong, W., and Yang, P. (2011). Heterologous protein display on the cell surface of lactic acid bacteria mediated by the s-layer protein. Microb. Cell Fact. 10:86. doi: 10.1186/1475-2859-10-86
Hu, Y., Liu, C., and Muyldermans, S. (2017). Nanobody-based delivery systems for diagnosis and targeted tumor therapy. Front. Immunol. 8:1442. doi: 10.3389/fimmu.2017.01442
Huet, H. A., Growney, J. D., Johnson, J. A., Li, J., Bilic, S., Ostrom, L. , et al. (2014). Multivalent nanobodies targeting death receptor 5 elicit superior tumor cell killing through efficient caspase induction. MAbs 6, 1560–1570. doi: 10.4161/19420862.2014.975099
Hultberg, A., Temperton, N. J., Rosseels, V., Koenders, M., Gonzalez-Pajuelo, M., Schepens, B. , et al. (2011). Llama-derived single domain antibodies to build multivalent, superpotent and broadened neutralizing anti-viral molecules. PLoS One 6:e17665. doi: 10.1371/journal.pone.0017665
Hussack, G., Arbabi-Ghahroudi, M., van Faassen, H., Songer, J. G., Ng, K. K., MacKenzie, R. , et al. (2011). Neutralization of Clostridium difficile toxin A with single-domain antibodies targeting the cell receptor binding domain. J. Biol. Chem. 286, 8961–8976. doi: 10.1074/jbc.M110.198754
Iezzi, M. E., Policastro, L., Werbajh, S., Podhajcer, O., and Canziani, G. A. (2018). Single-domain antibodies and the promise of modular targeting in cancer imaging and treatment. Front. Immunol. 9:273. doi: 10.3389/fimmu.2018.00273
Isolauri, E., Salminen, S., and Ouwehand, A. C. (2004). Microbial-gut interactions in health and disease. Probiotics. Best Pract. Res. Clin. Gastroenterol. 18, 299–313. doi: 10.1016/j.bpg.2003.10.006
Kaakoush, N. O., Castaño-Rodríguez, N., Mitchell, H. M., and Man, S. M. (2015). Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 28, 687–720. doi: 10.1128/CMR.00006-15
Kajikawa, A., Satoh, E., Leer, R. J., Yamamoto, S., and Igimi, S. (2007). Intragastric immunization with recombinant Lactobacillus casei expressing flagellar antigen confers antibody-independent protective immunity against Salmonella enterica serovar Enteritidis. Vaccine 25, 3599–3605. doi: 10.1016/j.vaccine.2007.01.055
Kazemi-Lomedasht, F., Behdani, M., Bagheri, K. P., Habibi-Anbouhi, M., Abolhassani, M., Arezumand, R. , et al. (2015). Inhibition of angiogenesis in human endothelial cell using VEGF specific nanobody. Mol. Immunol. 65, 58–67. doi: 10.1016/j.molimm.2015.01.010
Kijanka, M., Dorresteijn, B., Oliveira, S., and van Bergen en Henegouwen, P. M. P. (2015). Nanobody-based cancer therapy of solid tumors. Nanomedicine 10, 161–174. doi: 10.2217/nnm.14.178
Klaenhammer, T., Altermann, E., Arigoni, F., Bolotin, A., Breidt, F., Broadbent, J. , et al. (2002). Discovering lactic acid bacteria by genomics. Antonie Van Leeuwenhoek 82, 29–58. doi: 10.1023/A:1020638309912
Kong, D.-H., Kim, M. R., Jang, J. H., Na, H.-J., and Lee, S. (2017). A review of anti-angiogenic targets for monoclonal antibody cancer therapy. Int. J. Mol. Sci. 18:1786. doi: 10.3390/ijms18081786
Koromyslova, A. D., and Hansman, G. S. (2015). Nanobody binding to a conserved epitope promotes norovirus particle disassembly. J. Virol. 89, 2718–2730. doi: 10.1128/JVI.03176-14
Koromyslova, A. D., and Hansman, G. S. (2017). Nanobodies targeting norovirus capsid reveal functional epitopes and potential mechanisms of neutralization. PLoS Pathog. 13:e1006636. doi: 10.1371/journal.ppat.1006636
Kruger, C., Hu, Y., Pan, Q., Marcotte, H., Hultberg, A., Delwar, D. , et al. (2002). In situ delivery of passive immunity by lactobacilli producing single-chain antibodies. Nat. Biotechnol. 20, 702–706. doi: 10.1038/nbt0702-702
Kruger, C., Hultberg, A., Marcotte, H., Hermans, P., Bezemer, S., Frenken, L. G. , et al. (2006). Therapeutic effect of llama derived VHH fragments against Streptococcus mutans on the development of dental caries. Appl. Microbiol. Biotechnol. 72, 732–737. doi: 10.1007/s00253-006-0347-0
Kruger, C., Hultberg, A., van Dollenweerd, C., Marcotte, H., and Hammarstrom, L. (2005). Passive immunization by Lactobacilli expressing single-chain antibodies against Streptococcus mutans. Mol. Biotechnol. 31, 221–231. doi: 10.1111/j.1834-7819.2005.tb00370.x
La-Beck, N. M., Jean, G. W., Huynh, C., Alzghari, S. K., and Lowe, D. B. (2015). Immune checkpoint inhibitors: new insights and current place in cancer therapy. Pharmacotherapy 35, 963–976. doi: 10.1002/phar.1643
Landete, J. M. (2017). A review of food-grade vectors in lactic acid bacteria: from the laboratory to their application. Crit. Rev. Biotechnol. 37, 296–308. doi: 10.3109/07388551.2016.1144044
Lauwereys, M., Arbabi Ghahroudi, M., Desmyter, A., Kinne, J., Holzer, W., De Genst, E. , et al. (1998). Potent enzyme inhibitors derived from dromedary heavy-chain antibodies. EMBO J. 17, 3512–3520. doi: 10.1093/emboj/17.13.3512
Le, D. T., Durham, J. N., Smith, K. N., Wang, H., Bartlett, B. R., Aulakh, L. K. , et al. (2017). Mismatch-repair deficiency predicts response of solid tumors to PD-1 blockade. Science (New York, N.Y.) 357, 409–413. doi: 10.1126/science.aan6733
Le, D. T., Uram, J. N., Wang, H., Bartlett, B. R., Kemberling, H., Eyring, A. D. , et al. (2015). PD-1 blockade in tumors with mismatch-repair deficiency. N. Engl. J. Med. 372, 2509–2520. doi: 10.1056/NEJMoa1500596
Lee, P., Abdul-Wahid, A., and Faubert, G. M. (2009). Comparison of the local immune response against Giardia lamblia cyst wall protein 2 induced by recombinant Lactococcus lactis and Streptococcus gordonii. Microbes Infect. 11, 20–28. doi: 10.1016/j.micinf.2008.10.002
Leow, C. H., Fischer, K., Leow, C. Y., Cheng, Q., Chuah, C., and McCarthy, J. (2017). Single domain antibodies as new biomarker detectors. Diagnostics 7:52. doi: 10.3390/diagnostics7040052
Li, X., Xing, Y., Guo, L., Lv, X., Song, H., and Xi, T. (2014). Oral immunization with recombinant Lactococcus lactis delivering a multi-epitope antigen CTB-UE attenuates Helicobacter pylori infection in mice. Pathog. Dis. 72, 78–86. doi: 10.1111/2049-632x.12173
Lievin-Le Moal, V., and Servin, A. L. (2014). Anti-infective activities of Lactobacillus strains in the human intestinal microbiota: from probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 27, 167–199. doi: 10.1128/cmr.00080-13
Lin, C. F., Lo, T. C., Kuo, Y. C., and Lin, T. H. (2013). Stable integration and expression of heterologous genes in several lactobacilli using an integration vector constructed from the integrase and attP sequences of phage PhiAT3 isolated from Lactobacillus casei ATCC 393. Appl. Microbiol. Biotechnol. 97, 3499–3507. doi: 10.1007/s00253-012-4393-5
Lin, T. H., Lin, C. H., and Pan, T. M. (2018). The implication of probiotics in the prevention of dental caries. Appl. Microbiol. Biotechnol. 102, 577–586. doi: 10.1007/s00253-017-8664-z
Linares, D. M., Alvarez-Sieiro, P., del Rio, B., Ladero, V., Redruello, B., Martin, M. C. , et al. (2015). Implementation of the agmatine-controlled expression system for inducible gene expression in Lactococcus lactis. Microb. Cell Fact. 14:208. doi: 10.1186/S12934-015-0399-X
Linares, D. M., Perez, M., Ladero, V., Del Rio, B., Redruello, B., Martin, M. C. , et al. (2014). An agmatine-inducible system for the expression of recombinant proteins in Enterococcus faecalis. Microb. Cell Fact. 13:169. doi: 10.1186/s12934-014-0169-1
Lipson, E. J. (2013). Re-orienting the immune system: durable tumor regression and successful re-induction therapy using anti-PD1 antibodies. Oncoimmunology 2:e23661. doi: 10.4161/onci.23661
Liu, H., Wang, Y., Duan, H., Zhang, A., Liang, C., Gao, J. , et al. (2015). An intracellularly expressed Nsp9-specific nanobody in MARC-145 cells inhibits porcine reproductive and respiratory syndrome virus replication. Vet. Microbiol. 181, 252–260. doi: 10.1016/j.vetmic.2015.10.021
Liu, J. K., Hou, X. L., Wei, C. H., Yu, L. Y., He, X. J., Wang, G. H. , et al. (2009). Induction of immune responses in mice after oral immunization with recombinant Lactobacillus casei strains expressing enterotoxigenic Escherichia coli F41 fimbrial protein. Appl. Environ. Microbiol. 75, 4491–4497. doi: 10.1128/aem.02672-08
Liu, S., Li, Y., Deng, B., and Xu, Z. (2016). Recombinant Lactococcus lactis expressing porcine insulin-like growth factor I ameliorates DSS-induced colitis in mice. BMC Biotechnol. 16:25. doi: 10.1186/s12896-016-0255-z
Liu, Y., and Huang, H. (2018). Expression of single-domain antibody in different systems. Appl. Microbiol. Biotechnol. 102, 539–551. doi: 10.1007/s00253-017-8644-3
Ma, L., Gu, K., Zhang, C. H., Chen, X. T., Jiang, Y., Melcher, K. , et al. (2016). Generation and characterization of a human nanobody against VEGFR-2. Acta Pharmacol. Sin. 37, 857–864. doi: 10.1038/aps.2016.2
Maffey, L., Vega, C. G., Parreno, V., and Garaicoechea, L. (2015). Controlling Rotavirus-associated diarrhea: could single-domain antibody fragments make the difference? Rev. Argent. Microbiol. 47, 368–379. doi: 10.1016/j.ram.2015.09.005
Malanovic, N., and Lohner, K. (2016). Gram-positive bacterial cell envelopes: the impact on the activity of antimicrobial peptides. Biochim. Biophys. Acta 1858, 936–946. doi: 10.1016/j.bbamem.2015.11.004
Mathiesen, G., Sveen, A., Piard, J. C., Axelsson, L., and Eijsink, V. G. (2008). Heterologous protein secretion by Lactobacillus plantarum using homologous signal peptides. J. Appl. Microbiol. 105, 215–226. doi: 10.1111/j.1365-2672.2008.03734.x
Mao, R., Wu, D., and Wang, Y. (2016). Surface display on lactic acid bacteria without genetic modification: strategies and applications. Appl. Microbiol. Biotechnol. 100, 9407–9421. doi: 10.1007/s00253-016-7842-8
Mamat, U., Wilke, K., Bramhill, D., Schromm, A. B., Lindner, B., Kohl, T. A. , et al. (2015). Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Fact. 14:57. doi: 10.1186/s12934-015-0241-5
Marcobal, A., Liu, X., Zhang, W., Dimitrov, A. S., Jia, L., Lee, P. P. , et al. (2016). Expression of human immunodeficiency virus Type 1 neutralizing antibody fragments using human vaginal Lactobacillus. AIDS Res. Hum. Retrovir. 32, 964–971. doi: 10.1089/aid.2015.0378
Marcotte, H., Koll-Klais, P., Hultberg, A., Zhao, Y., Gmur, R., Mandar, R. , et al. (2006). Expression of single-chain antibody against RgpA protease of Porphyromonas gingivalis in Lactobacillus. J. Appl. Microbiol. 100, 256–263. doi: 10.1111/j.1365-2672.2005.02786.x
Marelli, B., Perez, A. R., Banchio, C., de Mendoza, D., and Magni, C. (2011). Oral immunization with live Lactococcus lactis expressing rotavirus VP8 subunit induces specific immune response in mice. J. Virol. Methods 175, 28–37. doi: 10.1016/j.jviromet.2011.04.011
Marshall, B. J., Barrett, L. J., Prakash, C., McCallum, R. W., and Guerrant, R. L. (1990). Urea protects Helicobacter (Campylobacter) pylori from the bactericidal effect of acid. Gastroenterology 99, 697–702. doi: 10.1016/0016-5085(90)90957-3
Martin, M. C., Alonso, J. C., Suarez, J. E., and Alvarez, M. A. (2000). Generation of food-grade recombinant lactic acid bacterium strains by site-specific recombination. Appl. Environ. Microbiol. 66, 2599–2604. doi: 10.1128/AEM.66.6.2599-2604.2000
Martin, M. C., Pant, N., Ladero, V., Gunaydin, G., Andersen, K. K., Alvarez, B. , et al. (2011). Integrative expression system for delivery of antibody fragments by lactobacilli. Appl. Environ. Microbiol. 77, 2174–2179. doi: 10.1128/AEM.02690-10
Mays, Z. J., and Nair, N. U. (2018). Synthetic biology in probiotic lactic acid bacteria: at the frontier of living therapeutics. Curr. Opin. Biotechnol. 53, 224–231. doi: 10.1016/j.copbio.2018.01.028
McFarland, L. V., Ship, N., Auclair, J., and Millette, M. (2018). Primary prevention of Clostridium difficile infections with a specific probiotic combining Lactobacillus acidophilus, L. casei, and L. rhamnosus strains: assessing the evidence. J. Hosp. Infect. 99, 443–452. doi: 10.1016/j.jhin.2018.04.017
Medina, E., and Guzman, C. A. (2001). Use of live bacterial vaccine vectors for antigen delivery: potential and limitations. Vaccine 19, 1573–1580. doi: 10.1016/S0264-410X(00)00354-6
Mejias, M. P., Hiriart, Y., Lauche, C., Fernandez-Brando, R. J., Pardo, R., Bruballa, A. , et al. (2016). Development of camelid single chain antibodies against Shiga toxin type 2 (Stx2) with therapeutic potential against Hemolytic Uremic Syndrome (HUS). Sci. Rep. 6:24913. doi: 10.1038/srep24913
Michon, C., Kuczkowska, K., Langella, P., Eijsink, V. G., Mathiesen, G., and Chatel, J. M. (2015). Surface display of an anti-DEC-205 single chain Fv fragment in Lactobacillus plantarum increases internalization and plasmid transfer to dendritic cells in vitro and in vivo. Microb. Cell Fact. 14:95. doi: 10.1186/s12934-015-0290-9
Michon, C., Langella, P., Eijsink, V. G., Mathiesen, G., and Chatel, J. M. (2016). Display of recombinant proteins at the surface of lactic acid bacteria: strategies and applications. Microb. Cell Fact. 15:70. doi: 10.1186/s12934-016-0468-9
Mohamadzadeh, M., and Klaenhammer, T. R. (2008). Specific Lactobacillus species differentially activate Toll-like receptors and downstream signals in dendritic cells. Expert Rev. Vaccines 7, 1155–1164. doi: 10.1586/14760584.7.8.1155
Monedero, V., Rodriguez-Diaz, J., Viana, R., Buesa, J., and Perez-Martinez, G. (2004). Selection of single-chain antibodies against the VP8* subunit of rotavirus VP4 outer capsid protein and their expression in Lactobacillus casei. Appl. Environ. Microbiol. 70, 6936–6939. doi: 10.1128/aem.70.11.6936-6939.2004
Mukherjee, J., Tremblay, J. M., Leysath, C. E., Ofori, K., Baldwin, K., Feng, X. , et al. (2012). A novel strategy for development of recombinant antitoxin therapeutics tested in a mouse botulism model. PLoS One 7:e29941. doi: 10.1371/journal.pone.0029941
Muyldermans, S. (2013). Nanobodies: natural single-domain antibodies. Annu. Rev. Biochem. 82, 775–797. doi: 10.1146/annurev-biochem-063011-092449
Muyldermans, S., Cambillau, C., and Wyns, L. (2001). Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains. Trends Biochem. Sci. 26, 230–235. doi: 10.1016/S0968-0004(01)01790-X
Nakano, K., Nomura, R., Matsumoto, M., and Ooshima, T. (2010). Roles of oral bacteria in cardiovascular diseases—from molecular mechanisms to clinical cases: cell-surface structures of novel serotype k Streptococcus mutans strains and their correlation to virulence. J. Pharmacol. Sci. 113, 120–125. doi: 10.1254/jphs.09R24FM
Narita, J., Okano, K., Kitao, T., Ishida, S., Sewaki, T., Sung, M. H. , et al. (2006). Display of alpha-amylase on the surface of Lactobacillus casei cells by use of the PgsA anchor protein, and production of lactic acid from starch. Appl. Environ. Microbiol. 72, 269–275. doi: 10.1128/aem.72.1.269-275.2006
Norton, P. M., Wells, J. M., Brown, H. W., Macpherson, A. M., and Le Page, R. W. (1997). Protection against tetanus toxin in mice nasally immunized with recombinant Lactococcus lactis expressing tetanus toxin fragment C. Vaccine 15, 616–619.
Oh, Y., Varmanen, P., Han, X. Y., Bennett, G., Xu, Z., Lu, T. , et al. (2007). Lactobacillus plantarum for oral peptide delivery. Oral Microbiol. Immunol. 22, 140–144. doi: 10.1111/j.1399-302X.2007.00338.x
O’Neil, B. H., Wallmark, J. M., Lorente, D., Elez, E., Raimbourg, J., Gomez-Roca, C. , et al. (2017). Safety and antitumor activity of the anti-PD-1 antibody pembrolizumab in patients with advanced colorectal carcinoma. PLoS One 12:e0189848. doi: 10.1371/journal.pone.0189848
Oggioni, M. R., Beninati, C., Boccanera, M., Medaglini, D., Spinosa, M. R., Maggi, T. , et al. (2001). Recombinant Streptococcus gordonii for mucosal delivery of a scFv microbicidal antibody. Int. Rev. Immunol. 20, 275–287. doi: 10.3109/08830180109043039
Oliveira, S., Heukers, R., Sornkom, J., Kok, R. J., and van Bergen en Henegouwen, P. M. P. (2013). Targeting tumors with nanobodies for cancer imaging and therapy. J. Control. Release 172, 607–617. doi: 10.1016/j.jconrel.2013.08.298
Olivera-Severo, D., Uberti, A. F., Marques, M. S., Pinto, M. T., Gomez-Lazaro, M., Figueiredo, C. , et al. (2017). A new role for Helicobacter pylori urease: contributions to angiogenesis. Front. Microbiol. 8:1883. doi: 10.3389/fmicb.2017.01883
Ouwehand, A. C., Salminen, S., and Isolauri, E. (2002). Probiotics: an overview of beneficial effects. Antonie Van Leeuwenhoek 82, 279–289. doi: 10.1023/a:1020620607611
Pant, N., Hultberg, A., Zhao, Y., Svensson, L., Pan-Hammarstrom, Q., Johansen, K. , et al. (2006). Lactobacilli expressing variable domain of llama heavy-chain antibody fragments (lactobodies) confer protection against rotavirus-induced diarrhea. J. Infect. Dis. 194, 1580–1588. doi: 10.1086/508747
Pant, N., Marcotte, H., Hermans, P., Bezemer, S., Frenken, L., Johansen, K. , et al. (2011). Lactobacilli producing bispecific llama-derived anti-rotavirus proteins in vivo for rotavirus-induced diarrhea. Future Microbiol. 6, 583–593. doi: 10.2217/fmb.11.32
Pardoll, D. M. (2012). The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 12, 252–264. doi: 10.1038/nrc3239
Piñero-Lambea, C., Ruano-Gallego, D., and Fernandez, L. A. (2015). Engineered bacteria as therapeutic agents. Curr. Opin. Biotechnol. 35, 94–102. doi: 10.1016/j.copbio.2015.05.004
Ren, C., Zhang, Q., Wang, G., Ai, C., Hu, M., Liu, X. , et al. (2014). Modulation of peanut-induced allergic immune responses by oral lactic acid bacteria-based vaccines in mice. Appl. Microbiol. Biotechnol. 98, 6353–6364. doi: 10.1007/s00253-014-5678-7
Riazi, A., Strong, P. C., Coleman, R., Chen, W., Hirama, T., van Faassen, H. , et al. (2013). Pentavalent single-domain antibodies reduce Campylobacter jejuni motility and colonization in chickens. PLoS One 8:e83928. doi: 10.1371/journal.pone.0083928
Ricci, A., Allende, A., Bolton, D., Chemaly, M., Davies, R., Girones, R. , et al. (2018). EFSA panel on biological hazards (EFSA BIOHAZ Panel). Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 7: suitability of taxonomic units notified to EFSA until September 2017. EFSA J. 16:e05131. doi: 10.2903/j.efsa.2018.5131
Riera-Montes, M., O’Ryan, M., and Verstraeten, T. (2017). Norovirus and rotavirus disease severity in children: systematic review and meta-analysis. Pediatr. Infect. Dis. J. 37, 501–505. doi: 10.1097/inf.0000000000001824
Robinson, K., Chamberlain, L. M., Schofield, K. M., Wells, J. M., and Le Page, R. W. (1997). Oral vaccination of mice against tetanus with recombinant Lactococcus lactis. Nat. Biotechnol. 15, 653–657. doi: 10.1038/nbt0797-653
Rozan, C., Cornillon, A., Petiard, C., Chartier, M., Behar, G., Boix, C. , et al. (2013). Single-domain antibody-based and linker-free bispecific antibodies targeting FcgammaRIII induce potent antitumor activity without recruiting regulatory T cells. Mol. Cancer Ther. 12, 1481–1491. doi: 10.1158/1535-7163.mct-12-1012
Saerens, D., Stijlemans, B., Baral, T. N., Nguyen Thi, G. T., Wernery, U., Magez, S. , et al. (2008). Parallel selection of multiple anti-infectome nanobodies without access to purified antigens. J. Immunol. Methods 329, 138–150. doi: 10.1016/j.jim.2007.10.005
Saez-Lara, M. J., Gomez-Llorente, C., Plaza-Diaz, J., and Gil, A. (2015). The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: a systematic review of randomized human clinical trials. Biomed. Res. Int. 2015:505878. doi: 10.1155/2015/505878
Salminen, S., Nybom, S., Meriluoto, J., Collado, M. C., Vesterlund, S., and El-Nezami, H. (2010). Interaction of probiotics and pathogens—benefits to human health? Curr. Opin. Biotechnol. 21, 157–167. doi: 10.1016/j.copbio.2010.03.016
Sarker, S. A., Jakel, M., Sultana, S., Alam, N. H., Bardhan, P. K., Chisti, M. J. , et al. (2013). Anti-rotavirus protein reduces stool output in infants with diarrhea: a randomized placebo-controlled trial. Gastroenterology 145, 740–748.e8. doi: 10.1053/j.gastro.2013.06.053
Saxelin, M., Tynkkynen, S., Mattila-Sandholm, T., and de Vos, W. M. (2005). Probiotic and other functional microbes: from markets to mechanisms. Curr. Opin. Biotechnol. 16, 204–211. doi: 10.1016/j.copbio.2005.02.003
Schneewind, O., Mihaylova-Petkov, D., and Model, P. (1993). Cell wall sorting signals in surface proteins of gram-positive bacteria. EMBO J. 12, 4803–4811. doi: 10.1002/j.1460-2075.1993.tb06169.x
Schneewind, O., and Missiakas, D. (2014). Sec-secretion and sortase-mediated anchoring of proteins in Gram-positive bacteria. Biochim. Biophys. Acta 1843, 1687–1697. doi: 10.1016/j.bbamcr.2013.11.009
Shi, T., Ma, Y., Yu, L., Jiang, J., Shen, S., Hou, Y. , et al. (2018). Cancer immunotherapy: a focus on the regulation of immune checkpoints. Int. J. Mol. Sci. 19, 1389–1403. doi: 10.3390/ijms19051389
Shibasaki, S., Karasaki, M., Tafuku, S., Aoki, W., Sewaki, T., and Ueda, M. (2014). Oral immunization against candidiasis using Lactobacillus casei displaying enolase 1 from Candida albicans. Sci. Pharm. 82, 697–708. doi: 10.3797/scipharm.1404-07
Shiroza, T., Shinozaki-Kuwahara, N., Hayakawa, M., Shibata, Y., Hashizume, T., Fukushima, K. , et al. (2003). Production of a single-chain variable fraction capable of inhibiting the Streptococcus mutans glucosyltransferase in Bacillus brevis: construction of a chimeric shuttle plasmid secreting its gene product. Biochim. Biophys. Acta 1626, 57–64. doi: 10.1016/s0167-4781(03)00038-1
Siegel, S. D., Reardon, M. E., and Ton-That, H. (2017). Anchoring of LPXTG-like proteins to the gram-positive cell wall envelope. Curr. Top. Microbiol. Immunol. 404, 159–175. doi: 10.1007/82_2016_8
Sindhu, K. N. C., Sowmyanarayanan, T. V., Paul, A., Babji, S., Ajjampur, S. S. R., Priyadarshini, S. , et al. (2014). Immune response and intestinal permeability in children with acute gastroenteritis treated with Lactobacillus rhamnosus GG: a randomized, double-blind, placebo-controlled trial. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 58, 1107–1115. doi: 10.1093/cid/ciu065,
Singh, P. P., Sharma, P. K., Krishnan, G., and Lockhart, A. C. (2015). Immune checkpoints and immunotherapy for colorectal cancer. Gastroenterol. Rep. (Oxf) 3, 289–297. doi: 10.1093/gastro/gov053
Siontorou, C. G. (2013). Nanobodies as novel agents for disease diagnosis and therapy. Int. J. Nanomedicine 8, 4215–4227. doi: 10.2147/IJN.S39428
Slordahl, T. S., Denayer, T., Moen, S. H., Standal, T., Borset, M., Ververken, C. , et al. (2013). Anti-c-MET nanobody—A new potential drug in multiple myeloma treatment. Eur. J. Haematol. 91, 399–410. doi: 10.1111/ejh.12185
Song, A. A.-L., In, L. L. A., Lim, S. H. E., and Rahim, R. A. (2017). A review on Lactococcus lactis: from food to factory. Microb. Cell Fact. 16:55. doi: 10.1186/s12934-017-0669-x
Stadtmann, A., and Zarbock, A. (2012). CXCR2: from bench to bedside. Front. Immunol. 3:263. doi: 10.3389/fimmu.2012.00263
Steeland, S., Vandenbroucke, R. E., and Libert, C. (2016). Nanobodies as therapeutics: big opportunities for small antibodies. Drug Discov. Today 21, 1076–1113. doi: 10.1016/j.drudis.2016.04.003
Steidler, L., Hans, W., Schotte, L., Neirynck, S., Obermeier, F., Falk, W. , et al. (2000). Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 289, 1352–1355. doi: 10.1126/science.289.5483.1352
Steidler, L., Neirynck, S., Huyghebaert, N., Snoeck, V., Vermeire, A., Goddeeris, B. , et al. (2003). Biological containment of genetically modified Lactococcus lactis for intestinal delivery of human interleukin 10. Nat. Biotechnol. 21, 785–789. doi: 10.1038/nbt840nbt840
Su, C., Nguyen, V. K., and Nei, M. (2002). Adaptive evolution of variable region genes encoding an unusual type of immunoglobulin in camelids. Mol. Biol. Evol. 19, 205–215. doi: 10.1093/oxfordjournals.molbev.a004073
Szatraj, K., Szczepankowska, A. K., and Chmielewska-Jeznach, M. (2017). Lactic acid bacteria – promising vaccine vectors: possibilities, limitations, doubts. J. Appl. Microbiol. 123, 325–339. doi: 10.1111/jam.13446
Thueng-in, K., Thanongsaksrikul, J., Jittavisutthikul, S., Seesuay, W., Chulanetra, M., Sakolvaree, Y. , et al. (2014). Interference of HCV replication by cell penetrable human monoclonal scFv specific to NS5B polymerase. MAbs 6, 1327–1339. doi: 10.4161/mabs.29978
Tokuhara, D., Alvarez, B., Mejima, M., Hiroiwa, T., Takahashi, Y., Kurokawa, S. , et al. (2013). Rice-based oral antibody fragment prophylaxis and therapy against rotavirus infection. J. Clin. Invest. 123, 3829–3838. doi: 10.1172/jci70266
Tremblay, J. M., Mukherjee, J., Leysath, C. E., Debatis, M., Ofori, K., Baldwin, K. , et al. (2013). A single VHH-based toxin-neutralizing agent and an effector antibody protect mice against challenge with Shiga toxins 1 and 2. Infect. Immun. 81, 4592–4603. doi: 10.1128/iai.01033-13
Tsai, H.-F., and Hsu, P.-N. (2017). Cancer immunotherapy by targeting immune checkpoints: mechanism of T cell dysfunction in cancer immunity and new therapeutic targets. J. Biomed. Sci. 24:35. doi: 10.1186/s12929-017-0341-0
Unger, M., Eichhoff, A. M., Schumacher, L., Strysio, M., Menzel, S., Schwan, C. , et al. (2015). Selection of nanobodies that block the enzymatic and cytotoxic activities of the binary Clostridium difficile toxin CDT. Sci. Rep. 5:7850. doi: 10.1038/srep07850
van der Linden, R. H., Frenken, L. G., de Geus, B., Harmsen, M. M., Ruuls, R. C., Stok, W. , et al. (1999). Comparison of physical chemical properties of llama VHH antibody fragments and mouse monoclonal antibodies. Biochim. Biophys. Acta 1431, 37–46.
van der Vaart, J. M., Pant, N., Wolvers, D., Bezemer, S., Hermans, P. W., Bellamy, K. , et al. (2006). Reduction in morbidity of rotavirus induced diarrhoea in mice by yeast produced monovalent llama-derived antibody fragments. Vaccine 24, 4130–4137. doi: 10.1016/j.vaccine.2006.02.045
van Pijkeren, J. P., and Barrangou, R. (2017). Genome editing of food-grade Lactobacilli to develop therapeutic probiotics. Microbiol. Spectr. 5, 1–25. doi: 10.1128/microbiolspec.BAD-0013-2016
Vandenbroucke, K., de Haard, H., Beirnaert, E., Dreier, T., Lauwereys, M., Huyck, L. , et al. (2010). Orally administered L. lactis secreting an anti-TNF Nanobody demonstrate efficacy in chronic colitis. Mucosal Immunol. 3, 49–56. doi: 10.1038/mi.2009.116
Vandenbroucke, K., Hans, W., Van Huysse, J., Neirynck, S., Demetter, P., Remaut, E. , et al. (2004). Active delivery of trefoil factors by genetically modified Lactococcus lactis prevents and heals acute colitis in mice. Gastroenterology 127, 502–513. doi: 10.1053/j.gastro.2004.05.020
Vanlandschoot, P., Stortelers, C., Beirnaert, E., Ibañez, L. I., Schepens, B., Depla, E. , et al. (2011). Nanobodies®: new ammunition to battle viruses. Antivir. Res. 92, 389–407. doi: 10.1016/j.antiviral.2011.09.002
Vega, C. G., Bok, M., Vlasova, A. N., Chattha, K. S., Gomez-Sebastian, S., Nunez, C. , et al. (2013). Recombinant monovalent llama-derived antibody fragments (VHH) to rotavirus VP6 protect neonatal gnotobiotic piglets against human rotavirus-induced diarrhea. PLoS Pathog. 9:e1003334. doi: 10.1371/journal.ppat.1003334
Venerito, M., Vasapolli, R., Rokkas, T., and Malfertheiner, P. (2015). Helicobacter pylori and gastrointestinal malignancies. Helicobacter 20 (Suppl 1), 36–39. doi: 10.1111/hel.12255
Vesa, T., Pochart, P., and Marteau, P. (2000). Pharmacokinetics of Lactobacillus plantarum NCIMB 8826, Lactobacillus fermentum KLD, and Lactococcus lactis MG 1363 in the human gastrointestinal tract. Aliment. Pharmacol. Ther. 14, 823–828. doi: 10.1046/j.1365-2036.2000.00763.x
Vosjan, M. J., Vercammen, J., Kolkman, J. A., Stigter-van Walsum, M., Revets, H., and van Dongen, G. A. (2012). Nanobodies targeting the hepatocyte growth factor: potential new drugs for molecular cancer therapy. Mol. Cancer Ther. 11, 1017–1025. doi: 10.1158/1535-7163.mct-11-0891
Wang, J., Mukhtar, H., Ma, L., Pang, Q., and Wang, X. (2018). VHH antibodies: reagents for mycotoxin detection in food products. Sensors (Basel) 18, 1–13. doi: 10.3390/s18020485
Wang, M., Gao, Z., Zhang, Y., and Pan, L. (2016). Lactic acid bacteria as mucosal delivery vehicles: a realistic therapeutic option. Appl. Microbiol. Biotechnol. 100, 5691–5701. doi: 10.1007/s00253-016-7557-x
Watkins, R. R., and Bonomo, R. A. (2016). Overview: global and local impact of antibiotic resistance. Infect. Dis. Clin. N. Am. 30, 313–322. doi: 10.1016/j.idc.2016.02.001
Watterlot, L., Rochat, T., Sokol, H., Cherbuy, C., Bouloufa, I., Lefevre, F. , et al. (2010). Intragastric administration of a superoxide dismutase-producing recombinant Lactobacillus casei BL23 strain attenuates DSS colitis in mice. Int. J. Food Microbiol. 144, 35–41. doi: 10.1016/j.ijfoodmicro.2010.03.037
Wei, C. H., Liu, J. K., Hou, X. L., Yu, L. Y., Lee, J. S., and Kim, C. J. (2010). Immunogenicity and protective efficacy of orally or intranasally administered recombinant Lactobacillus casei expressing ETEC K99. Vaccine 28, 4113–4118. doi: 10.1016/j.vaccine.2009.05.088
Weiner, L. M., Surana, R., and Wang, S. (2010). Monoclonal antibodies: versatile platforms for cancer immunotherapy. Nat. Rev. Immunol. 10, 317–327. doi: 10.1038/nri2744
Wells, J. (2011). Mucosal vaccination and therapy with genetically modified lactic acid bacteria. Annu. Rev. Food Sci. Technol. 2, 423–445. doi: 10.1146/annurev-food-022510-133640
Wells, J. M., and Mercenier, A. (2008). Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat. Rev. Microbiol. 6, 349–362. doi: 10.1038/nrmicro1840
Wells, J. M., Wilson, P. W. , Norton, P. M. , Gasson, M. J., and Le Page, R. W. (1993). Lactococcus lactis: high-level expression of tetanus toxin fragment C and protection against lethal challenge. Mol. Microbiol. 8, 1155–1162.
Wen, L. J., Hou, X. L., Wang, G. H., Yu, L. Y., Wei, X. M., Liu, J. K. , et al. (2012). Immunization with recombinant Lactobacillus casei strains producing K99, K88 fimbrial protein protects mice against enterotoxigenic Escherichia coli. Vaccine 30, 3339–3349. doi: 10.1016/j.vaccine.2011.08.036
Wu, Y., Jiang, S., and Ying, T. (2017). Single-domain antibodies as therapeutics against human viral diseases. Front. Immunol. 8:1802. doi: 10.3389/fimmu.2017.01802
Wyszynska, A., Kobierecka, P., Bardowski, J., and Jagusztyn-Krynicka, E. K. (2015). Lactic acid bacteria—20 years exploring their potential as live vectors for mucosal vaccination. Appl. Microbiol. Biotechnol. 99, 2967–2977. doi: 10.1007/s00253-015-6498-0
Xu, Y., Xiong, L., Li, Y., Xiong, Y., Tu, Z., Fu, J. , et al. (2015). Citrinin detection using phage-displayed anti-idiotypic single-domain antibody for antigen mimicry. Food Chem. 177, 97–101. doi: 10.1016/j.foodchem.2015.01.007
Yang, X. Q., Zhao, Y. G., Chen, X. Q., Jiang, B., and Sun, D. Y. (2013). The protective effect of recombinant Lactococcus lactis oral vaccine on a Clostridium difficile-infected animal model. BMC Gastroenterol. 13:117. doi: 10.1186/1471-230x-13-117
Yang, Z., Schmidt, D., Liu, W., Li, S., Shi, L., Sheng, J. , et al. (2014). A novel multivalent, single-domain antibody targeting TcdA and TcdB prevents fulminant Clostridium difficile infection in mice. J. Infect. Dis. 210, 964–972. doi: 10.1093/infdis/jiu196
Yao, G., Lam, K. H., Weisemann, J., Peng, L., Krez, N., Perry, K. , et al. (2017). A camelid single-domain antibody neutralizes botulinum neurotoxin A by blocking host receptor binding. Sci. Rep. 7:7438. doi: 10.1038/s41598-017-07457-5
Yuvaraj, S., Al-Lahham, S., Marreddy, R. K., Dijkstra, G., Wolken, W. A., Lolkema, J. S. , et al. (2008). Human scFv SIgA expressed on Lactococcus lactis as a vector for the treatment of mucosal disease. Mol. Nutr. Food Res. 52, 913–920. doi: 10.1002/mnfr.200700132
Zabana, Y., Domenech, E., Manosa, M., Garcia-Planella, E., Bernal, I., Cabre, E. , et al. (2010). Infliximab safety profile and long-term applicability in inflammatory bowel disease: 9-year experience in clinical practice. Aliment. Pharmacol. Ther. 31, 553–560. doi: 10.1111/j.1365-2036.2009.04206.x
Zadravec, P., Strukelj, B., and Berlec, A. (2015). Heterologous surface display on lactic acid bacteria: non-GMO alternative? Bioengineered 6, 179–183. doi: 10.1080/21655979.2015.1040956
Zadravec, P., Mavric, A., Bogovic Matijasic, B., Strukelj, B., and Berlec, A. (2014). Engineering BmpA as a carrier for surface display of IgG-binding domain on Lactococcus lactis. Protein Eng. Des. Sel. 27, 21–27. doi: 10.1093/protein/gzt059
Zhang, F., Wei, H., Wang, X., Bai, Y., Wang, P., Wu, J. , et al. (2017). Structural basis of a novel PD-L1 nanobody for immune checkpoint blockade. Cell Discov. 3:17004. doi: 10.1038/celldisc.2017.4
Zhang, Z. H., Jiang, P. H., Li, N. J., Shi, M., and Huang, W. (2005). Oral vaccination of mice against rodent malaria with recombinant Lactococcus lactis expressing MSP-1(19). World J. Gastroenterol. 11, 6975–6980. doi: 10.3748/wjg.v11.i44.6975
Keywords: VHH, nanobodies, lactic acid bacteria, in situ delivery, gastrointestinal tract, prophylaxis, therapy
Citation: del Rio, Redruello, Fernandez, Martin, Ladero and Alvarez (2019) Lactic Acid Bacteria as a Live Delivery System for the in situ Production of Nanobodies in the Human Gastrointestinal Tract. Front. Microbiol. 9:3179. doi: 10.3389/fmicb.2018.03179
Edited by:
Paloma López, Centro de Investigaciones Biológicas (CIB), SpainCopyright © 2019 del Rio, Redruello, Fernandez, Martin, Ladero and Alvarez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beatriz del Rio, YmVhZGVscmlvQGlwbGEuY3NpYy5lcw==