 Vera Manageiro1,2
Vera Manageiro1,2 Lurdes Clemente3
Lurdes Clemente3 Raquel Romão1Catarina Silva4Luís Vieira4Eugénia Ferreira1,2
Raquel Romão1Catarina Silva4Luís Vieira4Eugénia Ferreira1,2 Manuela Caniça1,2*
Manuela Caniça1,2*- 1National Reference Laboratory of Antibiotic Resistances and Healthcare Associated Infections, Department of Infectious Diseases, National Institute of Health Dr. Ricardo Jorge, Lisbon, Portugal
- 2Centre for the Studies of Animal Science, Institute of Agrarian and Agri-Food Sciences and Technologies, University of Porto, Porto, Portugal
- 3Bacteriology and Mycology Laboratory, INIAV – National Institute of Agrarian and Veterinary Research, Oeiras, Portugal
- 4Innovation and Technology Unit, Department of Human Genetics, National Institute of Health Dr. Ricardo Jorge, Lisbon, Portugal
We studied a commensal colistin-resistant Escherichia coli isolated from a swine cecum sample collected at a slaughter, in Portugal. Antimicrobial susceptibility phenotype of E. coli LV23529 showed resistance to colistin at a minimum inhibitory concentration of 4 mg/L. Whole genome of E. coli LV23529 was sequenced using a MiSeq system and the assembled contigs were analyzed for the presence of antibiotic resistance and plasmid replicon types using bioinformatics tools. We report a novel mcr-1 gene variant (mcr-1.9), carried by an IncX4 plasmid, where one-point mutation at nucleotide T1238C leads to Val413Ala substitution. The mcr-1.9 genetic context was characterized by an IS26 element upstream of the mcr-pap2 element and by the absence of ISApl1. Bioinformatic analysis also revealed genes conferring resistance to β-lactams, sulphamethoxazole, trimethoprim, chloramphenicol and colistin, corresponding to the phenotype noticed. Moreover, we highlight the presence of mcr-1.9 plus blaCTX-M-8, a blaESBL gene rarely detected in Europe in isolates of animal origin; these two genes were located on different plasmids with 33,303 and 89,458 bp, respectively. MCR-1.9-harboring plasmid showed high identity to other X4-type mcr-1-harboring plasmids characterized worldwide, which strongly suggests that the presence of PMCR-encoding genes in food-producing animals, such as MCR-1.9, represent a potential threat to humans, as it is located in mobile genetic elements that have the potential to spread horizontally.
Introduction
Since the report of a plasmid-mediated colistin resistance (PMCR) mechanism, designated MCR-1, in Escherichia coli and Klebsiella pneumoniae isolated from animals, food and humans in China, further reports exposed the global dissemination of mcr-type gene in various bacterial species isolated from a wide range of different sources (Caniaux et al., 2017). In Portugal, PMCR has also been detected in a wide range of different sources and species, including humans, food-producing animals and meat, and in the environment (Campos et al., 2016; Figueiredo et al., 2016; Jones-Dias et al., 2016; Kieffer et al., 2017; Manageiro et al., 2017; Tacão et al., 2017; Mendes et al., 2018). Noteworthy, are the recent report of two cases presumably associated with the travel of patients from Portugal, one involving animals: a patient repatriated to France after hospitalization for 2 months in Portugal, in 2015 (Beyrouthy et al., 2017), and a New York state patient returning from Portugal in 2016 after staying on a farm with chickens and pigs (Gilrane et al., 2017).
More worrisome is the presence of mcr genes in Enterobacteriaceae carrying other resistance determinants namely, extended-spectrum β-lactamases (ESBL)- and/or carbapenemase-encoding genes. Since the first report of co-localization of mcr-1 and ESBL- in 2016 in bovines in France, an increase encoding genes in the proportion of mcr-1 genes among ESBL-producing E. coli in animals has been noticed, suggesting that the use of extended-spectrum cephalosporins may have simultaneously favored the spread of mcr-1 (Haenni et al., 2016). Here we describe the first detection of a novel mcr variant, hereafter-named mcr-1.9, identified in a commensal E. coli LV23529 isolated from a swine cecum sample collected at a slaughter, in Portugal.
Materials and Methods
Bacterial Isolate
Escherichia coli LV23529 was isolated in 2015 from a swine cecum sample collected at a Portuguese slaughter, during an evaluation study of commensal E. coli recovered from swine samples for antimicrobial susceptibility testing.
Antimicrobial Susceptibility Testing
Minimum inhibitory concentrations (MICs) were determined by microdilution method as previously described (Manageiro et al., 2017). In order to assess decreased susceptibility of the strain, interpretation of the results was done according to the epidemiological cut-off values recommended by the European Committee on Antimicrobial Susceptibility Testing (EUCAST1).
Screening and Characterization of PMCR- and ESBL-Resistance Mechanisms
Molecular Detection of mcr-1 and blaESBL-Encoding Genes
Following phenotypic characteristics, PMCR- and ESBL-resistance mechanisms were searched and identified by molecular methods, as previously described (Manageiro et al., 2017).
Transfer Experiments
Conjugation experiments were performed using sodium azide-resistant E. coli J53 as a recipient strain. Transconjugants were selected on McConkey agar supplemented with sodium azide (150 mg/L) and either cefotaxime (2 mg/L) or colistin (2 mg/L). Plasmid DNA was extracted from E. coli LV23529 using a NucleoBond Xtra Plus kit (Macherey-Nagel), and transformed into E. coli TOP10 OneShot chemically competent cells (Invitrogen), accordingly to manufacture’s protocol. E. coli transformants were selected on MacConkey agar supplemented with 2 mg/L of colistin. PCR for blaCTX-M-8 or mcr-1-type and MICs of recipients and transformants were determined as mentioned above.
Genetic Context of mcr-1.9 Gene
Colistin-resistant E. coli LV23529 was genotypically characterized by whole-genome sequencing (WGS), as previously described (Manageiro et al., 2017). Sequence reads were trimmed and filtered according to quality criteria, and de novo assembled into contigs by means of CLC Genomics Workbench 10.0 (Qiagen). The assembled contigs were analyzed and studied for the presence of antibiotic resistance, virulence genes and plasmid replicon types, serotype, multi-locus sequence type (ST) and fim-type, using bioinformatics tools2. The NCBI prokaryotic genome automatic annotation pipeline (PGAAP) was used for annotation.
Plasmid sequencing was also performed on a MiSeq Illumina platform using 150 bp paired-end reads, after plasmid DNA extraction from TLV23529 (mcr-1.9) using a NucleoBond Xtra Plus kit (Macherey-Nagel), and quantification using Qubit 1.0 Fluorometer (Invitrogen), as previously described (Manageiro et al., 2017). Sequence reads were trimmed and filtered according to quality criteria, and mapped against E. coli ATCC 25922 genome (NZ_CP009073). Unmapped reads (80.2%/total reads) were then used for plasmids structure construction by mapping assembly based on the genetic organization of the closest plasmid sequences obtained by BLASTn; this was followed by contig neighbor’s prediction from assembly information using CLC Genomics Workbench 10.0 (Qiagen). NCBI Microbial genomes BLAST analysis tool3 was used to search for plasmid sequences. Plasmid alignments and ORF representations were also done using EasyFig v. 2.2.3 (Sullivan et al., 2011).
Genomic Epidemiological Analysis
BacWGSTdb database was used for genotyping and source tracking bacterial pathogen (Ruan and Feng, 2016).
Nucleotide Sequence Accession Number
The pLV23529-MCR-1.9 and pLV23529-CTX-M-8 nucleotide sequences from this study were submitted to the NCBI GenBank Database with accession numbers KY964067 and KY964068, respectively. The new mcr-1.9 nucleotide sequence was submitted with accession number KY780959.
This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession SBIH00000000. The version described in this paper is version SBIH01000000.
Results and Discussion
MIC results showed that LV23529 was non-wild-type to third- and fourth-generation cephalosporins (ceftazidime 2 mg/L, cefotaxime 32 mg/L, cefepime 8 mg/L) with synergy with clavulanic acid; this isolate was also non-wild-type to chloramphenicol (>128 mg/L), sulphamethoxazole (>1024 mg/L), trimethoprim (>32 mg/L), tetracycline (>64 mg/L), and colistin (4 mg/L). LV23529 remained wild-type to carbapenems, fluoroquinolones, aminoglycosides and tigecycline (Table 1).
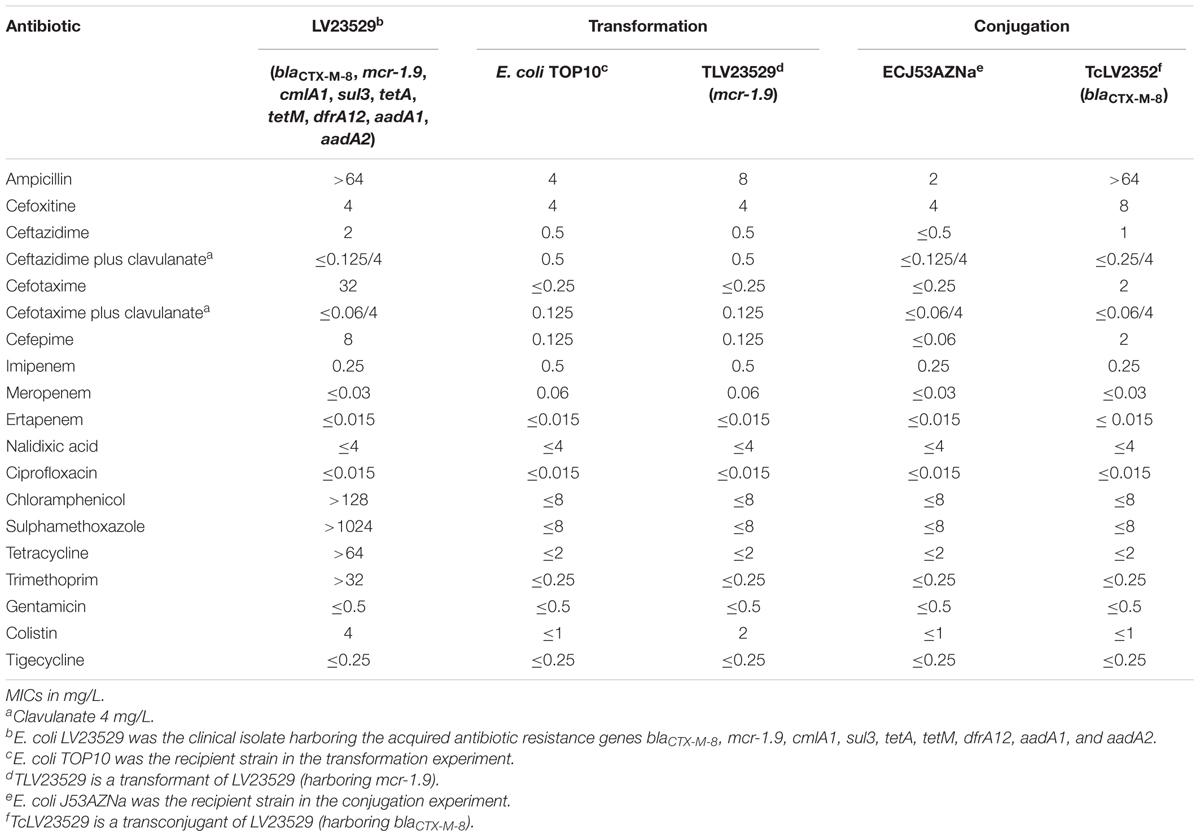
Table 1. Phenotypic and genotypic context of CTX-M-8 and MCR-1.9 producing E. coli clinical isolate, transformant, transconjugant, and the respective recipient strains.
Molecular characterization of the E. coli LV23529 isolate allowed the detection of blaCTX-M-8 and mcr-1-type genes.
Only the transferability of the blaCTX-M-8 gene was achieved by conjugation, with TcLV23529 (blaCTX-M-8) exhibiting the ESBL phenotype from LV23529 isolate (cefotaxime 2 mg/L, cefepime 2 mg/L) with synergy with clavulanic acid, and wild-type to colistin (≤1 mg/L) (Table 1). Although conjugation assays for mcr-1-type were negative, the colistin resistance determinant could be transferred to E. coli TOP10 competent cells; transformant TLV23529 (mcr-1-type) showed the respective resistance to colistin (4 mg/L) (Table 1).
The WGS assembly of E. coli LV23529 yielded 193 contigs (average 143.7-fold coverage), which together comprised 5,122,415bp, showing a GC content of 50.7%. The largest contig was 320,931 bp long; the N50 statistic, which stands for the minimum contig length of at least 50% of the contigs, was 113,197 bp. The average length of the obtained contigs was 26,541 bp. Overall, the genome sequence comprised 5,124 putative genes, among which 5,037 consisted of protein encoding sequences.
The WGS analysis showed that E. coli LV23529 belongs to serotype O8:H19, usually associated with porcine stx2e-producing E. coli (Zweifel et al., 2006; Bai et al., 2015), and to MLST (Achtman scheme) ST201 [clonal complex 469 (CC469)] and to the FimH-type determinant fimH32. This ST201 was encountered worldwide mainly in isolates collected from livestock samples (Escherichia/Shigella Enterobase database, Alikhan et al., 2018). Three virulence factors were detected: astA (heat-stable enterotoxin 1), lpfA (long polar fimbriae), and gad-type (glutamate decarboxylase).
Further bioinformatics analysis of E. coli LV23529 isolate revealed acquired-genes conferring resistance to β-lactams (blaCTX-M-8 and blaTEM-1), aminoglycosides (aadA1 and aadA2), phenicol (cmlA1-type and floR-type), sulphamethoxazole (sul3), tetracycline [tet(A)-type and tet(M)-type], trimethoprim (dfrA12), and colistin (mcr-1-type), justifying the phenotype noticed. Additionally, several unknown mutations in the ampC (promoter region), parC, 16S rrsB, 16S rrsC, 23S and pmrB chromosomal genes were detected, the last gene being described as the primary mechanism for the development of chromosomally encoded resistance to polymyxins (Phan et al., 2017).
The named mcr-1.9, differed from mcr-1 by one-point mutation (T1238C), leading to Val413Ala substitution. The MCR-1 protein contains a transmembrane domain and a phosphoethanolamine (PEA) transferase domain with 8α, 12β, and 12η units (Gao et al., 2016). The amino acid substitution of MCR-1.9 occurred in the region between η7 e η8 of the PEA transferase domain, which have been found not to influence the function of MCR-1 (Gao et al., 2016).
The mcr-1.9 genetic context was characterized by an IS26 element upstream of the mcr-pap2 element and by the absence of ISApl1 (Figure 1), which is in accordance with other studies about mcr-1 gene (Veldman et al., 2016; Sun et al., 2017). The mcr-1.9 gene can be mobilized within an ISApl1-flanked composite transposon (Tn6330), although many sequences have been identified without ISApl1 or with just a single copy (Snesrud et al., 2018). Indeed, it has been described that initially ISApl1 was presumably involved in the transposition of the mcr-1 cassette and then was lost, contributing for the stability of mcr gene on IncX4 plasmids (Sun et al., 2017; Snesrud et al., 2018).
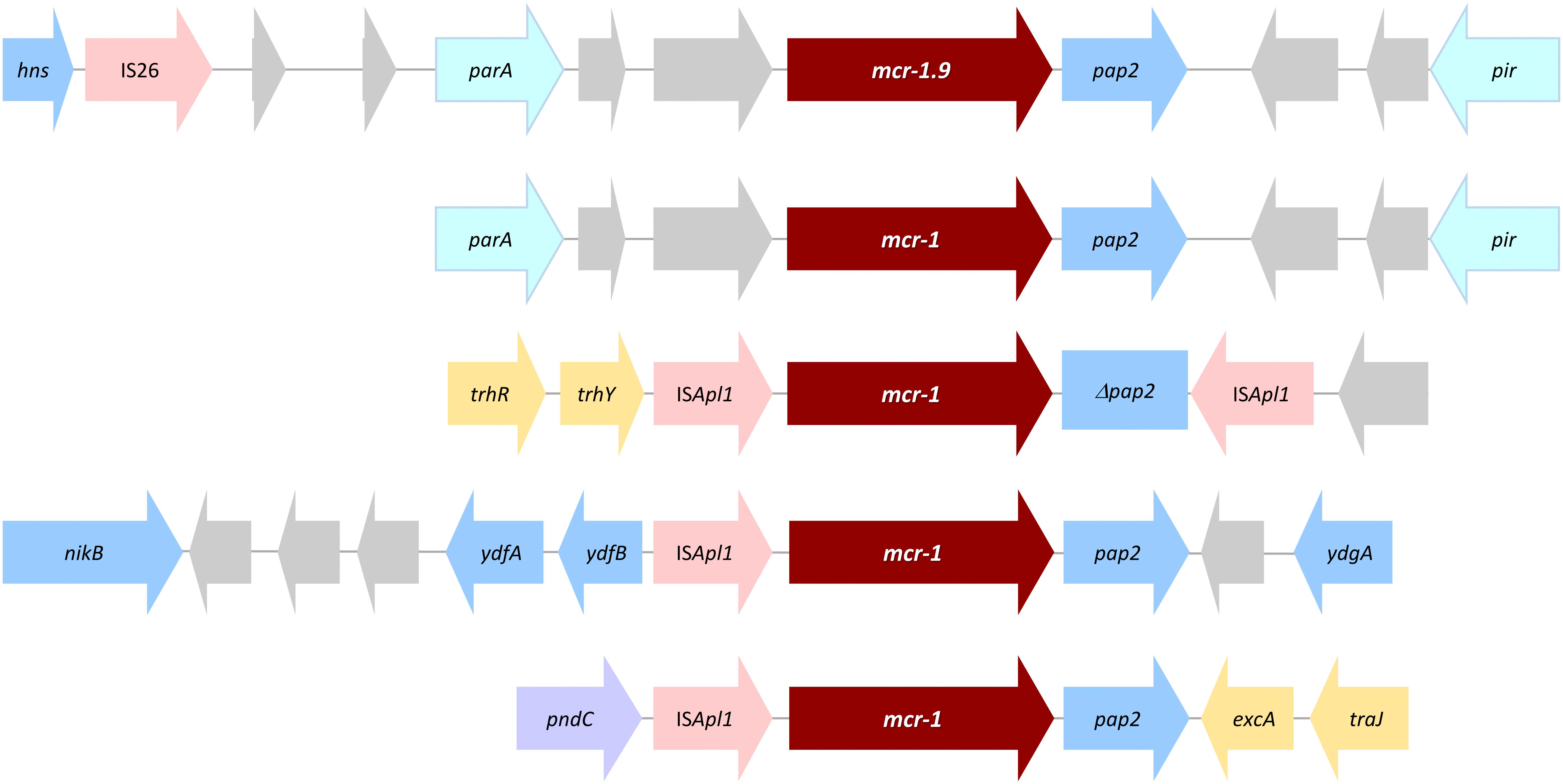
Figure 1. Linear comparison of IncX4-pLV23529-MCR-1.9 with the top six mcr-1-harboring plasmids showing the highest identities (>99.9%, E-value 0.0), in different E. coli isolates. Boxed arrows represent the position and transcriptional direction of ORFs. Gray vertical blocks indicate the shared similarity regions according to TBLASTX identity. Genes associated with pilus and plasmid transfer are colored yellow, antibiotic resistance genes are colored red, mobile genetic elements are colored pink, and other genes are colored gray (hypothetical proteins) or blue (other).
The PMCR-encoding gene was found in an IncX4 plasmid (pLV23529-MCR-1.9), showing highest identities (>99.9%) with six IncX4-type mcr-1-harboring plasmids identified worldwide, in unrelated E. coli isolates, mainly collected from human patients (Figure 2 and Table 2). Indeed, all belonged to different MLST, which might suggest a resistance plasmid dissemination across strains (plasmid outbreak) rather than clonal transmission of MCR-1-type-producing strains. Furthermore, no E. coli LV23529 closely related isolates were detected among those currently deposited in the public database BacWGSTdb (Ruan and Feng, 2016), which reinforce the importance of the horizontal gene transfer in this study.

Figure 2. Schematic representation of the genetic environment of mcr-1.9 in comparison with other mcr-1-type representative environments. Boxed arrows represent the position and transcriptional direction of ORFs. Genes are not drawn to scale. Genes associated with pilus and plasmid transfer are colored yellow, antibiotic resistance genes are in red, mobile genetic elements in pink, plasmid maintenance and stability genes in violet, plasmid replication associated genes are in light blue, and other genes are colored gray (hypothetical proteins) or blue (other).
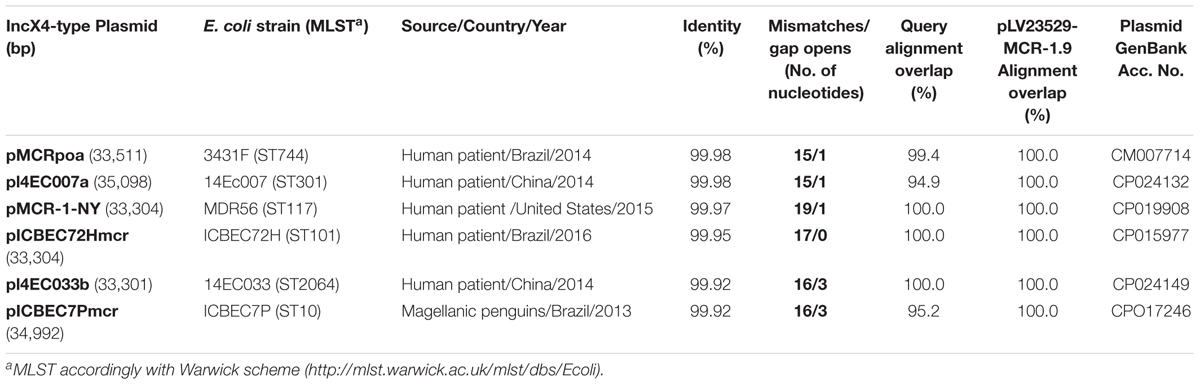
Table 2. Comparison of IncX4-pLV23529-MCR-1.9 with the top six mcr-1-harboring plasmids showing the highest identities (>99.9%, E-value 0.0), in different E. coli isolates.
Like pLV23529-MCR-1.9, the six plasmids (Table 2) doesn’t have the ISApl1 element. Hence, similarities may suggest that the one-point mutation (T1238C) in mcr-1.9 occurred on the X4 plasmid, since mobilization of mcr-1 occurs as part of a composite transposon (Tn6330) and that structures lacking the downstream ISApl1 are not capable of mobilization (Snesrud et al., 2018). The IS26 upstream of the mcr-pap2 element is flanked by an 8bp direct repeat (Figure 3A), indicating that its insertion wouldn’t seems to be related to the mcr-1.9 context, justifying the differences found with other IncX4 mcr-1-harboring plasmids. IncX4 plasmid has been widely implicated in the spread of MCR-1 gene in Europe (Caniaux et al., 2017). In Portugal, this plasmid type is circulating among diverse hosts (humans, pigs, poultry), being responsible for hospital-based outbreak caused by MCR-1 plus KPC-3-producing K. pneumoniae (Mendes et al., 2018), as well as for the diffusion of this PMCR at the farm level (Kieffer et al., 2017). Indeed, IncX4 plasmids seem to be efficiently transferred at different temperatures and different lack-of-fitness burdens among bacterial hosts, which may facilitate the transfer of mcr-type among Enterobacteriaceae (Lo et al., 2014; Wu R. et al., 2018). The pLV23529-MCR-1.9 plasmid backbone contains all the core genes common to IncX plasmids involved in segregation, stability, replication, and conjugative transfer of the plasmid (Figure 3A), namely the IncX-type pilus synthesis operon (pilX1-pilX11). However, pLV23529-MCR-1.9 was mobilizable, but not self-transmissible. Of note, we found a one-point mutation (G64T), leading to Asp22Tyr substitution, in the PilX1, a peptidoglycan hydrolase involved in T-DNA plasmid transfer. This mutation might explain why the attempts to conjugate mcr-1.9 from E. coli LV23529 were unsuccessful (Chen et al., 2009).
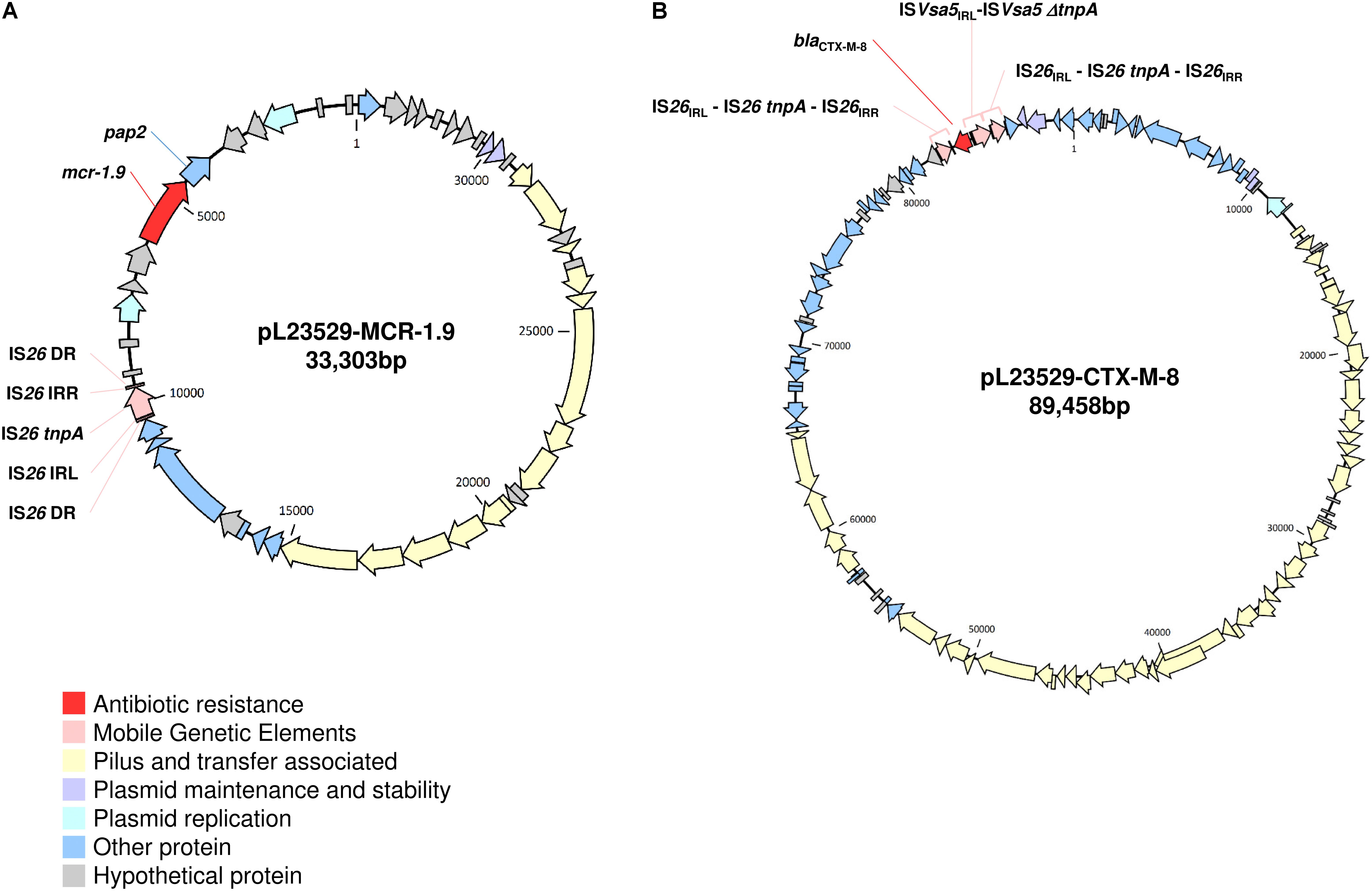
Figure 3. Schematic maps of pLV23529-MCR-1.9 (A) and pLV23529-CTX-M-8 (B). Genes are denoted by arrows and colored based on gene function classification.
Further plasmid analysis revealed the presence of two other plasmids: IncF [F2:A-:B-], IncR and the colicinogenic IncI1-ST113-carrying the blaCTX-M-8 (pLV23529-CTX-M-8, Figure 3B). Of note, the mcr-1.9-positive isolate, co-harboring blaCTX-M-8 and blaTEM-1 genes, is here reported for the first time in an E. coli isolate of animal origin. In fact, blaCTX-M-8 gene is rarely detected in Europe in isolates of animal origin (Börjesson et al., 2016), but in humans seems to be emerging (Eller et al., 2014). Indeed, a recent phylogenetic study suggested an increasing trend of co-existence and transmission of blaCTX-M and mcr-1 in both clinical medicine and veterinary medicine (Wu C. et al., 2018).
In conclusion, the presence of PMCR-encoding genes, such as MCR-1.9, in food-producing animals represents a potential threat to humans, as it is located in mobile genetic elements that have the potential to spread horizontally. As mentioned, in Portugal, PMCR is an emerging problem and its international spread is a worrying reality (Beyrouthy et al., 2017; Gilrane et al., 2017).
Author Contributions
VM designed the study, performed the molecular experiments and bioinformatics analysis, interpreted the data, and wrote the manuscript. LC, RR, and EF performed the microbiological and molecular experiments. CS and LV performed the Illumina genome sequencing experiments. MC designed the study, wrote, reviewed and edited the manuscript. All authors read and approved the final manuscript.
Funding
VM was supported by Fundação para a Ciência e a Tecnologia (FCT) fellowship (Grant No. SFRH/BPD/77486/2011), financed by the European Social Funds (COMPETE-FEDER), and National Funds of the Portuguese Ministry of Education and Science (POPH-QREN). The authors thank to FCT for the project grant UID/MULTI/00211/2013.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
- ^http://mic.eucast.org/Eucast2/
- ^https://cge.cbs.dtu.dk/services/
- ^https://www.ncbi.nlm.nih.gov/genome/microbes/
References
Alikhan, N. F., Zhou, Z., Sergeant, M. J., and Achtman, M. (2018). A genomic overview of the population structure of Salmonella. PLoS Genet. 14:e1007261. doi: 10.1371/journal.pgen.1007261
Bai, X., Wang, H., Xin, Y., Wei, R., Tang, X., Zhao, A., et al. (2015). Prevalence and characteristics of Shiga toxin-producing Escherichia coli isolated from retail raw meats in China. Int. J. Food Microbiol. 200, 31–38. doi: 10.1016/j.ijfoodmicro.2015.01.018
Beyrouthy, R., Robin, F., Lessene, A., Lacombat, I., Dortet, L., Naas, T., et al. (2017). MCR-1 and OXA-48 in vivo acquisition in KPC-producing Escherichia coli after colistin treatment. Antimicrob. Agents Chemother. 61:e02540-16. doi: 10.1128/AAC.02540-16
Börjesson, S., Ny, S., Egervärn, M., Bergström, J., Rosengren,Å, Englund, S., et al. (2016). Limited dissemination of extended-spectrum β-lactamase- and plasmid-encoded Ampc-producing Escherichia coli from food and farm animals, Sweden. Emerg. Infect. Dis. 22, 634–640. doi: 10.3201/eid2204.151142
Campos, J., Cristino, L., Peixe, L., and Antunes, P. (2016). MCR-1 in multidrug-resistant and copper-tolerant clinically relevant Salmonella 1,4,[5],12:i:- and S. Rissen clones in Portugal, 2011 to 2015. Euro Surveill. 21:30270. doi: 10.2807/1560-7917.ES.2016.21.26.30270
Caniaux, I., van Belkum, A., Zambardi, G., Poirel, L., and Gros, M. F. (2017). MCR: modern colistin resistance. Eur. J. Clin. Microbiol. Infect. 36, 415–420. doi: 10.1007/s10096-016-2846-y
Chen, C.-L., Wang, C.-Y., Chu, C., Su, L.-H., and Chiu, C.-H. (2009). Functional and molecular characterization of pSE34 encoding a type IV secretion system in Salmonella enterica serotype Enteritidis phage type 34. FEMS Immunol. Med. Microbiol. 5, 274–283. doi: 10.1111/j.1574-695X.2009.00612.x
Eller, C., Leistner, R., Guerra, B., Fischer, J., Wendt, C., Rabsch, W., et al. (2014). Emergence of extended-spectrum β-lactamase (ESBL) CTX-M-8 in Germany. J. Antimicrob. Chemother. 69, 562–564. doi: 10.1093/jac/dkt387
Figueiredo, R., Card, R. M., Nunez, J., Pomba, C., Mendonça, N., Anjum, M. F., et al. (2016). Detection of an mcr-1-encoding plasmid mediating colistin resistance in Salmonella enterica from retail meat in Portugal. J. Antimicrob. Chemother. 71, 2338–2340. doi: 10.1093/jac/dkw240
Gao, R., Hu, Y., Li, Z., Sun, J., Wang, Q., Lin, J., et al. (2016). Dissemination and mechanism for the MCR-1 colistin resistance. PLoS Pathog. 12:e1005957. doi: 10.1371/journal.ppat.1005957
Gilrane, V. L., Lobo, S., Huang, W., Zhuge, J., Yin, C., Chen, D., et al. (2017). Complete genome sequence of a colistin-resistant Escherichia coli strain harboring mcr-1 on an IncHI2 plasmid in the United States. Genome Announc. 5:e01095-17. doi: 10.1128/genomeA.01095-17
Haenni, M., Métayer, V., Gay, E., and Madec, J. Y. (2016). Increasing trends in mcr-1 prevalence among extended-spectrum-β-lactamase-producing Escherichia coli isolates from french calves despite decreasing exposure to colistin. Antimicrob. Agents Chemother. 60, 6433–6434. doi: 10.1128/AAC.01147-16
Jones-Dias, D., Manageiro, V., Ferreira, E., Barreiro, P., Vieira, L., Moura, I. B., et al. (2016). Architecture of class 1, 2, and 3 integrons from Gram negative bacteria recovered among fruits and vegetables. Front. Microbiol. 7:1400. doi: 10.3389/fmicb.2016.01400
Kieffer, N., Aires-de-Sousa, M., Nordmann, P., and Poirel, L. (2017). High rate of MCR-1-producing Escherichia coli and Klebsiella pneumoniae among pigs. Portugal. Emerg. Infect. Dis. 23, 2023–2029. doi: 10.3201/eid2312.170883
Lo, W. U., Chow, K. H., Law, P. Y., Ng, K. Y., Cheung, Y. Y., Lai, E. L., et al. (2014). Highly conjugative IncX4 plasmids carrying blaCTX-M in Escherichia coli from humans and food animals. J. Med. Microbiol. 63, 835–840. doi: 10.1099/jmm.0.074021-0
Manageiro, V., Clemente, L., Graça, R., Correia, I., Albuquerque, T., Ferreira, E., et al. (2017). New insights into resistance to colistin and third-generation cephalosporins of Escherichia coli in poultry, Portugal: novel blaCTX-M-166 and blaESAC genes. Int. J. Food Microbiol. 263, 67–73. doi: 10.1016/j.ijfoodmicro.2017.10.007
Mendes, A. C., Novais,Â, Campos, J., Rodrigues, C., Santos, C., Antunes, P., et al. (2018). mcr-1 in carbapenemase-producing Klebsiella pneumoniae with hospitalized patients, Portugal, 2016-2017. Emerg. Infect. Dis. 24, 762–766. doi: 10.3201/eid2404.171787
Phan, M. D., Nhu, N. T. K., Achard, M. E. S., Forde, B. M., Hong, K. W., Chong, T. M., et al. (2017). Modifications in the pmrB gene are the primary mechanism for the development of chromosomally encoded resistance to polymyxins in uropathogenic Escherichia coli. J. Antimicrob. Chemother. 72, 2729–2736. doi: 10.1093/jac/dkx204
Ruan, Z., and Feng, Y. (2016). BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acids Res. 44, D682–D687. doi: 10.1093/nar/gkv1004
Snesrud, E., McGann, P., and Chandle, M. (2018). The birth and demise of the ISApl1-mcr-1-ISApl1 composite transposon: the vehicle for transferable colistin resistance. mBio 9:e02381-17. doi: 10.1128/mBio.02381-17
Sullivan, M. J., Petty, N. K., and Beatson, S. A. (2011). Easyfig: a genome comparison visualizer. Bioinformatics 27, 1009–1010. doi: 10.1093/bioinformatics/btr039
Sun, J., Fang, L. X., Wu, Z., Deng, H., Yang, R. S., Li, X. P., et al. (2017). Genetic analysis of the IncX4 plasmids: implications for a unique pattern in the mcr-1 acquisition. Sci. Rep. 7:424. doi: 10.1038/s41598-017-00095-x
Tacão, M., Tavares, R. D. S., Teixeira, P., Roxo, I., Ramalheira, E., Ferreira, S., et al. (2017). mcr-1 and blaKPC-3 in Escherichia coli sequence type 744 after meropenem and colistin therapy. Portugal. Emerg. Infect. Dis. 23, 1419–1421. doi: 10.3201/eid2308.170162
Veldman, K., van Essen-Zandbergen, A., Rapallini, M., Wit, B., Heymans, R., van Pelt, W., et al. (2016). Location of colistin resistance gene mcr-1 in Enterobacteriaceae from livestock and meat. J. Antimicrob. Chemother. 71, 2340–2342. doi: 10.1093/jac/dkw181
Wu, C., Wang, Y., Shi, X., Wang, S., Ren, H., Shen, Z., et al. (2018). Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes. Infect. 7:30. doi: 10.1038/s41426-018-0033-1
Wu, R., Yi, L.-X., Yu, L.-F., Wang, J., Liu, Y., Chen, X., et al. (2018). Fitness Advantage of mcr-1–bearing IncI2 and IncX4 plasmids in vitro. Front. Microbiol. 9:331. doi: 10.3389/fmicb.2018.00331
Keywords: MCR-1.9, plasmid-mediated colistin resistance, IncX4, CTX-M-8, Portugal
Citation: Manageiro V, Clemente L, Romão R, Silva C, Vieira L, Ferreira E and Caniça M (2019) IncX4 Plasmid Carrying the New mcr-1.9 Gene Variant in a CTX-M-8-Producing Escherichia coli Isolate Recovered From Swine. Front. Microbiol. 10:367. doi: 10.3389/fmicb.2019.00367
Received: 21 September 2018; Accepted: 12 February 2019;
Published: 14 March 2019.
Edited by:
Carlos Lodeiro, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, PortugalReviewed by:
María de Toro, Centro de Investigación Biomédica de La Rioja, SpainZhi Ruan, Zhejiang University, China
Copyright © 2019 Manageiro, Clemente, Romão, Silva, Vieira, Ferreira and Caniça. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuela Caniça, bWFudWVsYS5jYW5pY2FAaW5zYS5taW4tc2F1ZGUucHQ=