Abstract
Aedes mosquitoes are important vectors for emerging diseases caused by arboviruses, such as chikungunya (CHIKV). These viruses’ main transmitting species are Aedes aegypti and Ae. albopictus, which are present in tropical and temperate climatic areas all over the globe. Knowledge of vector characteristics is fundamentally important to the understanding of virus transmission. Only female mosquitoes are able to transmit CHIKV to the vertebrate host since they are hematophagous. In addition, mosquito microbiota is fundamentally important to virus infection in the mosquito. Microorganisms are able to modulate viral transmission in the mosquito, such as bacteria of the Wolbachia genus, which are capable of preventing viral infection, or protozoans of the Ascogregarina species, which are capable of facilitating virus transmission between mosquitoes and larvae. The competence of the mosquito is also important in the transmission of the virus to the vertebrate host, since their saliva has several substances with biological effects, such as immunomodulators and anticoagulants, which are able to modulate the host’s response to the virus, interfering in its pathogenicity and virulence. Understanding the Aedes vector-chikungunya interaction is fundamentally important since it can enable the search for new methods of combating the virus’ transmission.
Introduction
The sylvan transmission cycle involving invertebrate vector(s) and vertebrate reservoirs maintains, amplifies and contributes to arboviruses that actively provoke outbreaks of yellow fever, West Nile, Zika, chikungunya, and dengue (Rodríguez-Morales, 2015; Vega-Rúa et al., 2015; Chouin-Carneiro et al., 2016). CHIKV is a mosquito-borne arthritogenic pathogen, classified as an alphavirus of the Togaviridae family, which has an envelope and single strand RNA as nucleic acids. In the urban cycle, this virus is particularly transmitted by the bites of mosquitoes from the genus Aedes, mainly Aedes aegypti and Aedes albopictus, causing arthritis or arthralgia, which is accompanied by fever and rash (Mourya and Yadav, 2006). In this regard, Ae. aegypti is the most important vector and has extensive worldwide distribution, existing mainly in tropical and subtropical areas, whereas Ae. albopictus can thrive in temperate regions. Thus, Aedes mosquitoes are known to be the primary vectors of emerging and re-emerging arbovirus diseases. Arbovirus disease transmission cycles can be initiated by female mosquito during the blood meal (Honório et al., 2015; Kraemer et al., 2015).
These vectors are adapted for indoor and daytime biting in urban areas. They feed primarily in the indoors and the outdoors near dwellings, and choose standing water, such as puddles and water containers, to procreate (Rodríguez-Morales, 2015). Indeed, their larvae can be found in artificial containers where other microorganisms can be found. Consequently, these mosquito species can harbor many microorganisms, which can affect several physiological functions of the mosquito, such as aiding in digestion, nutrition, and reproduction (Gaio et al., 2011). In fact, the vector mosquito microbiota can modulate its immune response, as well as its ability to eliminate pathogens that cause diseases in humans (Dennison et al., 2014; Hegde et al., 2015).
Several studies identify the digestive tract as the first structure to be influenced by microorganisms, since it is the organ that receives and processes the ingested material (Franz et al., 2015). Once in the mosquito midgut, this microbiota is able to interfere with the vector-host-microorganism relationship (Weaver and Reisen, 2010; Dennison et al., 2014). Findings also evidenced by Hegde et al. (2015), who reported that the microbiota may vary according to the site, where an interaction can take place with the same tissue or target protein of arbovirus, with the virus itself, or even both. This microbiota is colocalized with the ingested arboviruses, mainly in the gut of the mosquito, but also in other tissues, such as those of the germline, salivary glands and Malpighian tubules (Dubrulle et al., 2009; Moreira et al., 2009a), as shown in Figure 1.
FIGURE 1
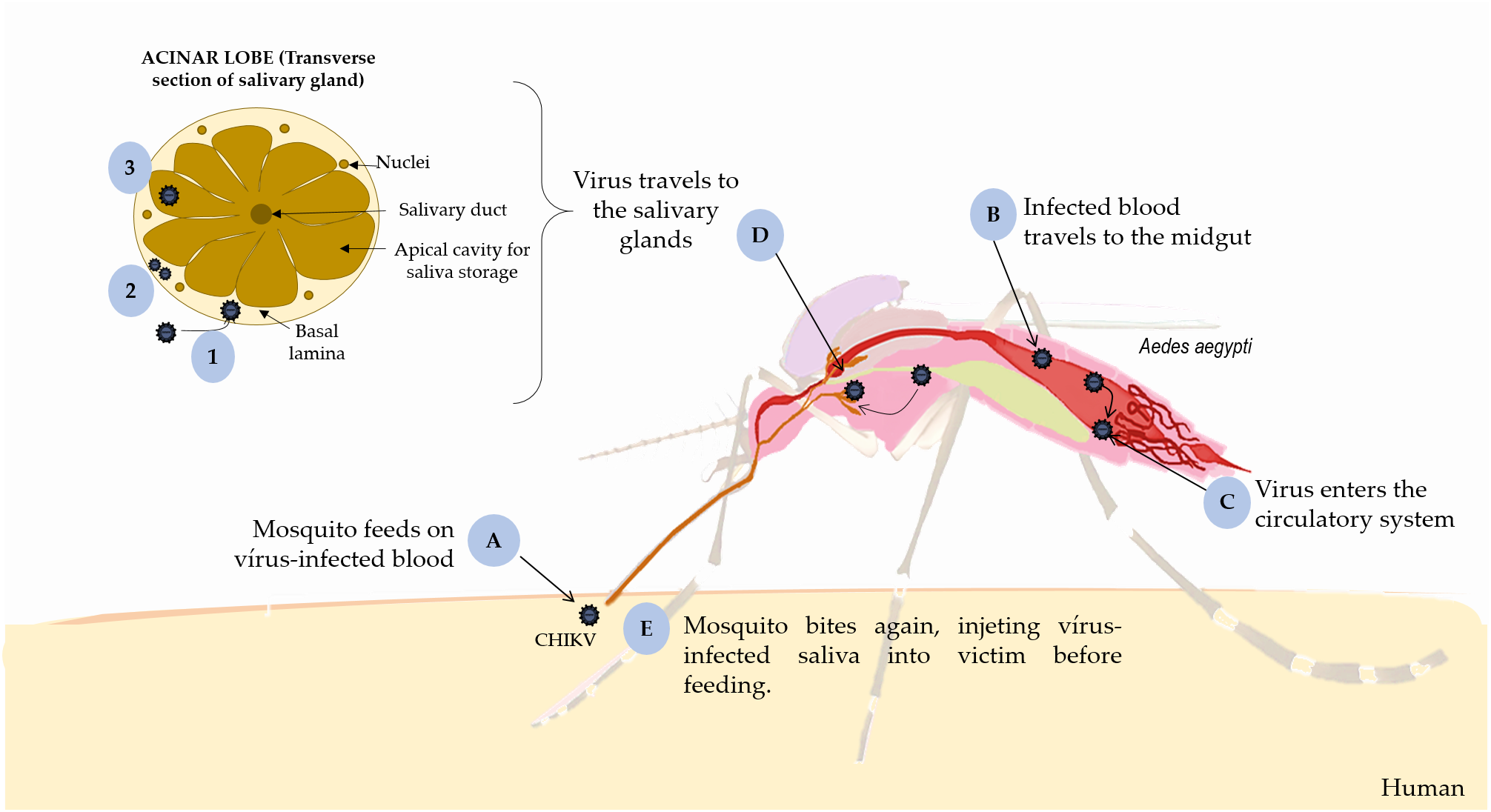
Replication of CHIKV in mosquito. (A) the mosquito feeds on virus-infected blood and occurs intrinsic incubation period in the vector, (B) CHIKV infects the midgut cells, (C) replicates and disseminates to the hemocoel and other organs, such (D) salivary glands. (1) It penetrates the basal lamina of the salivary glands surrounding acinar cells; (2) replicates inside these cells; and (3) is deposited into the apical cavities, (E) where mosquito saliva is stored prior to its release during feeding. This figure used elements from Servier Medical Art (www.servier.com).
In terms of pathogens infection, viral transmission via the Aedes vector is a complex process dependent on intrinsic factors to the mosquito, such as salivary composition, mosquito survival and virus replication index, and extrinsic factors that includes the climate, accessibility of vertebrate hosts and population and competition of vectors (Coffey et al., 2014). There is evidence that mosquito saliva can lead to vascular homeostasis and attenuate the host’s immune response (Melo et al., 2015), thereby enhancing viremia (McCracken et al., 2014) and viral pathogenicity (Schneider et al., 2004, 2010; Wasserman et al., 2004; Schneider and Higgs, 2008; Le Coupanec et al., 2013).
To develop better prophylactic and therapeutic interventions is essential to know how the immune system works against invading pathogens, especially intracellular microorganisms. Thus, some factors may influence CHIKV infectivity both in the vector and in humans. As such, we will describe some bacteria, protozoa and helminthes that may facilitate the infection and maintenance of CHIKV in Aedes mosquitoes, as summarized in Figure 2. In addition, we will also provide a comprehensive overview of the interactions between mosquito saliva and mammalian host immune cells that influence viremia and pathogenicity.
FIGURE 2
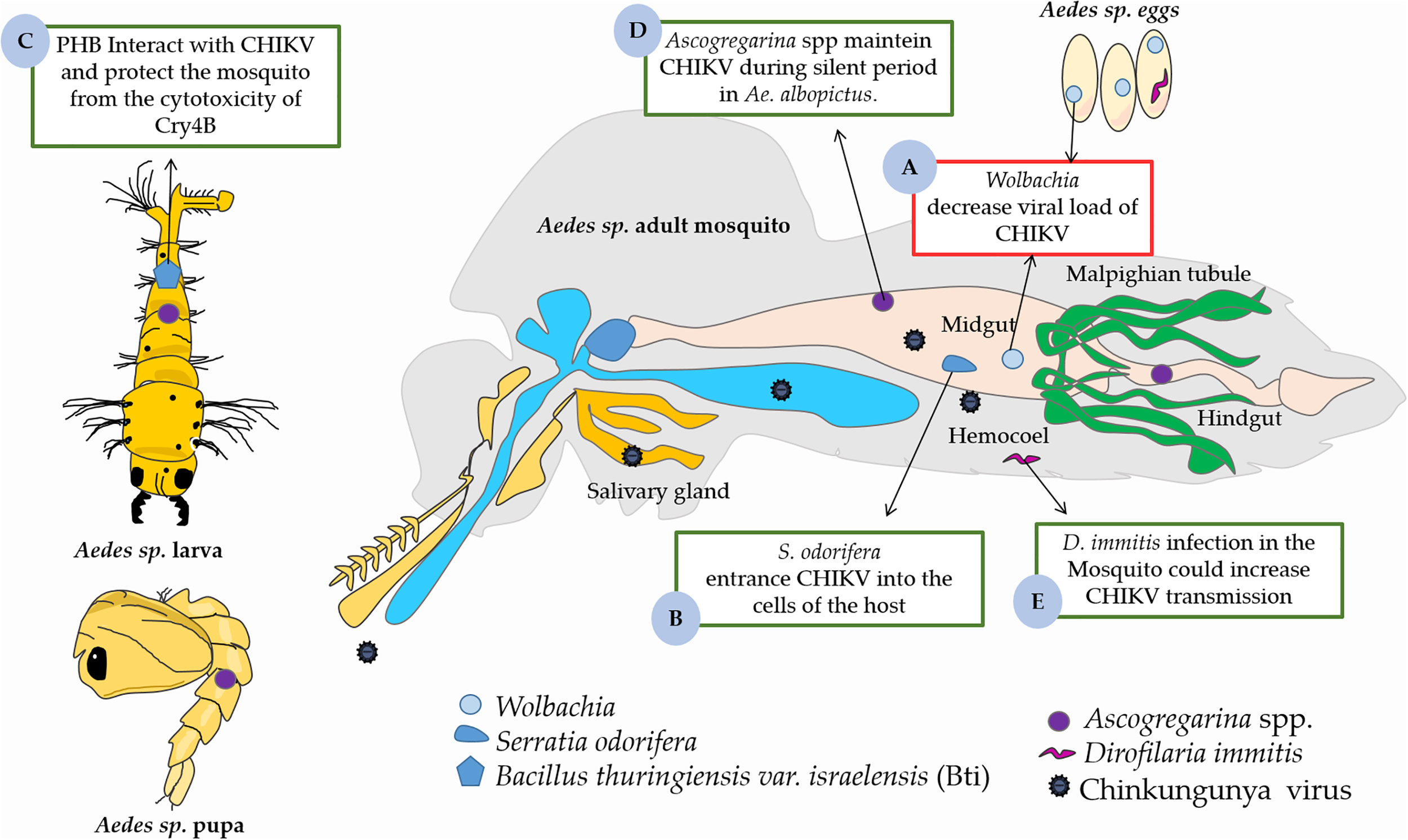
The role midguts microbiota on the CHIKV infection in Aedes vector. (A) The presence of Wolbachia in midgut decreased viral load in Aedes sp. (B)Serratia odorifera facilitates the entrance of CHIKV into the cells of the host. (C) Cry4B of Bacillus thuringiensis var. israelensis (Bti) has a direct interaction with PHB (a receptor of CHIKV) of Aedes sp., which can protect the mosquito from the cytotoxicity of Cry4B. (D)Ascogregarina spp. acts in maintenance of the CHIKV at silent period in Ae. albopictus. (E)Dirofilaria immitis could increasing CHIKV transmission in Aedes sp. This figure used elements from Servier Medical Art (www.servier.com).
Overview of the Aedes sp. Vector and Virus Interaction
There are over than 3,500 species of mosquitoes widely spread worldwide. They can be further classified into 112 genera, with four genera repeatedly linked with disease transmission to humans in the tropics and in cooler climates: Aedes, Anopheles, Culex, and Ochlerotatus (Elbers et al., 2015). Therefore, the Aedes genus has over 950 species, including Ae. aegypti as anthropophilic mosquitoes and Ae. albopictus as endophilic mosquitoes, which are more exophilic under natural field conditions; both are the most relevant species in tropical areas regarding disease transmission due to their adaptability to urban life and a high susceptibility to emerging and re-emerging arboviruses (Carrington and Simmons, 2014).
Aedes aegypti was first described in Linnaeus (1762), originating in African forests. This species is divided into two subspecies: (i) Ae. aegypti formosus, the darker and sylvatic mosquito, which reportedly inhibits forested habitats of Africa and is predominantly zoophilic; (ii) Ae. aegypti aegypti is a predominantly anthropophilic and domestic mosquito having as artificial habitats, mainly in urban environments reproducing in standing water containers and is widely diffused in tropical and subtropical areas (Brown et al., 2011; Coffey et al., 2014). Ae. aegypti has a bright, silvery, lyre-shaped dorsal pattern, white banded legs and can be found in some Asian countries, Australia, equatorial parts of Africa and in the Americas. More specifically, it can be found in the southern United States, Middle America, and South American countries, and has been highly distributed in Brazil (Kraemer et al., 2015).
Aedes albopictus was first described by Skuse (1894) in Calcutta, India. This species has a single, longitudinal, silvery dorsal stripe and white banded legs. It was arose from the forests of southeast Asia, and has been reported in several European countries and some Asian countries; it is poorly distributed in the African continent (Coffey et al., 2014). This species is highly distributed in the southeastern United States and southeast Brazil. In this regard, Brazil is one of the countries where the outbreaks of dengue and CHIKV occurred more frequently (Honório et al., 2015). The possible explanations for these outbreaks were the higher dissemination of both species of Aedes in this region and a single E1-A226V mutation (change of an alanine to a valine at codon 226 of the E1 protein) in different CHIKV strains that led specific genetic adaptations in the Ae. albopictus vector. Thus, this mutation increased the ability of the virus to replicate in Ae. albopictus, favored an increase in virus infectivity in the midgut and its spread to the salivary glands, thereby greatly facilitating the transmission of the disease (Tsetsarkin et al., 2007). Having an overview of the distribution of these species of mosquito vectors, Weaver and Lecuit (2015) also discussed why large epidemic cases of CHIKV have also occurred in Africa and the Indian Ocean basin and are now spreading rapidly throughout Europe and the Americas (Weaver and Lecuit, 2015).
Aedes aegypti can share the same larval habitats with Aedes albopictus, using natural and artificial water to lay their eggs individually on standing water surfaces because they are not susceptible to desiccation. After the eggs hatch, larvae can develop into four larval stages that metamorphose into pupae, and both are aquatic. After 2 days of the pupa cycle, pupa rupture occurs and the adult mosquito will be fully developed (Bidlingmayer, 1974).
For egg development, female mosquitoes should blood feed to acquire amino acids and other components that are important for the synthesis of vitellogenin, a glycophos apolipoprotein that is released into the insect hemolymph. Then, this protein is carried to the ovaries to be absorbed by the oocytes in the follicular epithelia. The female mosquito can oviposit approximately 250 eggs when the oocytes are full. In general, the place where the oviposition will occur is determinate by temperature, rainfall, relative humidity and wind, and hot and humid climates favor the flight and oviposition of the female mosquito (Bidlingmayer, 1974; Day, 2016). In addition, the main Ae. aegypti female behaviors, such as oviposition, blood feeding and sugar feeding, appear to be programmed by an overall circadian activity cycle (Day, 2016). The environment may influence vector-pathogen interactions during larval development, which leads to changes in mosquito competence, distribution and transmission of virus infections including chikungunya, dengue, Zika, and yellow fever to vertebrate hosts. In recent years, several studies from different continents have shown that vector competence can vary between individuals and among mosquito populations, and may be affected by the genetic and the environment components in which the vector is inserted (Diagne et al., 2014; Richard et al., 2016; Agha et al., 2017; Göertz et al., 2017; Chin et al., 2018).
Chikv–Vector Interactions
Vertical transmission in vectors occurs through infected eggs and horizontal transmission occurs during blood meals on a host, during which female mosquitos ingest CHIKV particles. Initially, CHIKV infects and replicates into midgut cells before disseminate to the hemocele and salivary glands during the extrinsic incubation in the vector (Coffey et al., 2014). Indeed, CHIKV needs reach the basal lamina of acinar cells, replicate inside them and get to the apical side, where mosquito saliva is maintained until releasing during blood meal. In mosquito, the salivary glands are sexually dimorphic, paired set in the thorax, and formed by a distinctive tri-lobed structure (two lateral and one median). These lobes consist of a basal lamina bounded by a singular epithelial layer or acinar cells that produces saliva, around a central salivary duct were the saliva is retained and later injected into the host’s skin during feeding (Juhn et al., 2011; Melo et al., 2015; Vega-Rúa et al., 2015) (Figure 1).
Moreover, during egg oviposition in artificial containers, the mosquitoes share larval habitats with some microorganisms. Consequently, they harbor many microorganisms that can develop and colonize their tissues, especially in the digestive system, where they can affect physiological functions, such as nutrition, digestion, and reproduction (Gaio et al., 2011). In addition, the mosquito microbiota can impact the vector’s competence for human pathogens (Hegde et al., 2015). Several studies identify the digestive tract as the first structure to be influenced by microorganisms, since it is the organ that receives and begins to process ingested material (Franz et al., 2015). Once in the mosquito midgut, this microbiota is able to interfere in the vector-host-microorganism relationship (Weaver and Reisen, 2010; Dennison et al., 2014).
In this regards, Hegde et al. (2015) reported an interaction can take place with the same tissue or target protein of the arbovirus, with the virus itself or even both, depending on the site. In addition, the variability of the microbial community in mosquito seems to be modulated by diet, sex, species and life stage. In the midgut of the mosquito, bacteria, protozoa and also helminthes are often located close to the habitats of ingested arboviruses, however, they can also infect other tissues, such as germline, salivary glands and Malpighian tubules (Dubrulle et al., 2009; Moreira et al., 2009a). In addition, Apte-Deshpande et al. (2014) showed that some of these mosquito gut inhabitants can also influence infections of CHIKV, DENV and yellow fever virus (YFV) in Aedes mosquitoes.
Role of the Midguts Microbiota on the Chikungunya Virus Infection in Aedes Vector
Key Role of Endosymbiotic Bacteria
Several bacteria can often be found in the mosquito gut, as well as in the tissues of the germline, Malpighian tubules and salivary glands (Favia et al., 2007; Hughes and Rasgon, 2014; Sharma et al., 2014; Chavshin et al., 2015). For instance, in Anopheles culicifacies, Sharma et al. (2014) found that the salivary gland harbors a broader and more diversified microbiota when compared to the gut of this insect. Moreover, these authors observed a similarity of 11% between the symbiotic bacterial communities of the midgut and the salivary gland of the Anopheles culicifacies, which are involved in the digestion of food (Sharma et al., 2014). Wolbachia is an obligate intracellular gram-negative endosymbiotic bacterium, which naturally infects more than 60% of insect species, including some Aedes species, such as Ae. albopictus, but not Ae. aegypti (Moreira et al., 2009a). This bacterium can be transmitted between mosquitoes by means of vertical transmission (maternally inherited) or by horizontal (e.g., parasitoid-host transfer), providing to its host nutritional benefits and greater resistance to pathogens (Hedges et al., 2008; Hegde et al., 2015; Wiwatanaratanabutr and Zhang, 2016).
The presence of Wolbachia-interfering pathogens, such as virus, was initially observed by Hedges et al. (2008) who showed that Drosophila melanogaster that contained Wolbachia bacteria in their microbiota had a mortality delay induced by Drosophila C virus (DCV), whereas flies without Wolbachia were more susceptible to the virus infection (Hedges et al., 2008). Thus, these authors have suggested that the delay in mortality caused by Wolbachia is associated with the anti-viral action of this bacterium. Similarly, Teixeira et al. (2008) also observed that the presence of Wolbachia decreased viral load in Drosophila, as well as Nora virus, but only slightly Flock House virus (FHV). In addition, these authors reported that genes involved in resistance to DCV (Dcr-2, ago-2, hop and wMel) in Drosophila can be stimulated by the presence of Wolbachia (Teixeira et al., 2008).
Although, Wolbachia does not naturally infect the Ae. aegypti, this mosquito can be transfected with the bacterium by embryo microinjection and adult microinjection (Hughes and Rasgon, 2014). In this regards, Moreira et al. (2009a) also showed that Wolbachia influences the dispersion of the DCV along the body of the mosquito. First, they used three distinct groups of mosquitoes: one infected with the wMelPop-CLA strain of Wolbachia, another treated with tetracycline, and a wild control group, free of all bacteria. The insects were fed blood containing a viral particle and the dispersion of this content along the head and body, as well as along the legs and wings, was observed. Moreira et al. (2009a) highlights the difference in the amount of viral RNA copies after the 14th day of infection, where the group infected with Wolbachia only 17% of the mosquitoes were infected with CHIKV, yet none of the animals had the virus disseminated through their bodies. The authors concluded that the increasing difficulty of Ae. aegypti in completing the process of blood feeding had been related to these strains of Wolbachia (Moreira et al., 2009a).
In addition, when associated with the same host, CHIKV and Wolbachia seem to interact with each other once values such as density, dispersal and movement of the virus within the mosquito change with the presence of the bacterium, and vice versa (Moreira et al., 2009a). Mosquitoes naturally infected by Wolbachia showed some resistance to viruses; however, the resistance seemed less effective (60% for Wolbachia++ and 70% for Wolbachia -) in comparison with transinfected Wolbachia strains such as wMelPop-CLA (Moreira et al., 2009b). Therefore, Wolbachia-infected (with wMelPop strain) Ae. aegypti mosquitoes showed reduced vector capacity and blood feeding behavior through physiological changes, such as reduction of females’ fecundity and egg viability and hatch rate, antiviral protection, stimulation of the immune system and decrease in the amount of saliva produced by the vector (Moreira et al., 2009b; Johnson, 2015; Fraser et al., 2017). In this regard, Moreira et al. (2009b) reported that older Ae. aegypti females infected with wMelPop had an increase in pre-probing and probing time, as well as exhibiting shaking behavior and the bendy proboscis, produced less volume of saliva and less ability to feed on blood. However, Wolbachia infection did not alter the activity of salivary apyrase (Moreira et al., 2009b).
Thus, the presence of the wMelPop strain blocks the accumulation of CHIKV virus (Moreira et al., 2009a). However, other studies have shown that the antiviral action of Wolbachia strains is associated with the induction of antimicrobial peptides and pre-activation of the innate immune response in the insect (Kambris et al., 2009, 2010; Moreira et al., 2009a; Fraser et al., 2017). For instance, Ae. aegypti induced the expression of molecules responsible for the immune effect (cecropin, defensin, thio-ester containing proteins, and C-type lectins) in the presence of wMelPop-CLA Wolbachia (Moreira et al., 2009a).
In Ae. albopictus vector, Zouache et al. (2012) analyzed the impact of CHIKV infection on bacterial community of this insect and found a significant decrease in endosymbiontic bacteria Wolbachia and Blattabacterium. In contrast, the authors showed an augment in bacteria from the Enterobacteriaceae family, as well as CHIKV infection influenced the composition of the bacterial community, and the aging of these infected mosquitoes changed slightly the density of bacteria, probably due to modified nutritional conditions (Zouache et al., 2012). Ae. albopictus is naturally infected by two strains of Wolbachia, named wAlbA and wAlbB (Zouache et al., 2009, 2012), but using C6/36 cell line (from Ae. albopictus cells) infected in vitro with wAlbB strain, it was demonstrated its ability to inhibit the replication and assembly/secretion steps of CHIKV infection (Raquin et al., 2015). Nevertheless, in some cells CHIKV RNA and Wolbachia were co-localized (Raquin et al., 2015).
Mousson et al. (2010) also studied the modulation of CHIKV replication in Ae. albopictus (Mousson et al., 2010). Using RT-PCR techniques to determine the viral load through RNA and PCR, these authors determined the density of Wolbachia by the presence of the actin gene in the DNA. However, mosquitoes with Wolbachia showed a homogeneous pattern in the amount of virus, associated with a decrease in the bacterial load. The authors stated that this had probably occurred due to competition for resources within the host.
van den Hurk et al. (2012) study the influence of wMel strain of Wolbachia on CHIKV infection in Ae. aegypti and showed that CHIKV was detected significantly less often in mosquito bodies infected with the wMel strain (23%), compared to uninfected mosquitoes (83%), and decreased dissemination to the salivary glands (van den Hurk et al., 2012). Regardless of all these studies, differently from Ae. albopictus that is naturally infected by wAlbA and wAlbB strains of Wolbachia, wild Ae. aegypti did not carry this bacterium (Zouache et al., 2009, 2012; Hughes and Rasgon, 2014). Although, recently, some studies have detected Wolbachia in a natural population of Ae. aegypti (Coon et al., 2016; Goindin et al., 2018).
Some species of Serratia (Enterobacteriaceae) have been isolated from the gut microbiota of mosquitoes, where they were considered to be dominant in all isolation assays. They represented more than half of the total microorganisms (fungi and bacteria) in Ae. aegypti (Demaio et al., 1996; Gusmão et al., 2007). Serratia spp. may possess some competitive advantages over other bacteria (Gusmão et al., 2007), thereby Serratia marcescens and Serratia nematodiphila are investigated for the control of some mosquito species (Patil et al., 2011, 2012; Suryawanshi et al., 2015).
Initially, Arnoult et al. (2009) suggested the mitochondrial proteins are important to the immune response of many organisms, since they act on the induction of cellular death after an infection. In this way, the authors suggest that it is possible that Gram-negative Serratia odorifera is a facilitator of the entrance of CHIKV into the cells of the host, which would increase the success of viral infection in the vector (Arnoult et al., 2009).
Apte-Deshpande et al. (2014) fed Ae. aegypti females, free of midgut microbiota, with virus alone, S. odorifera or Microbacterium oxydans along with virus via blood meal. These three groups were monitored for dissemination of CHIKV (Apte-Deshpande et al., 2014). The mosquitoes that were fed with S. odorifera and virus presented a higher susceptibility to CHIKV compared to the groups that received only virus or M. oxydans. On day 10 post feeding (DPF), the group receiving CHIKV+S. odorifera in the blood meal showed slightly higher virus titers (7.563 × 107 pfu/mosquito) compared to groups receiving only virus (4.93 × 107 pfu/mosquito) or virus plus M. oxydans (4.15 × 107 pfu/mosquito), indicating enhanced replication of the virus in Ae. aegypti tissues. In addition, these authors also showed that increased susceptibility of Ae. aegypti to CHIKV mediated by S. odorifera, is due to interaction of the P40 protein of the bacterium with proteins prohibitin and porin present on the midgut of Ae. aegypti that immunosuppress the mosquito, thus accentuating the infection to CHIKV (Apte-Deshpande et al., 2014). Thus, as the virus replication is not even two-fold elevated might be no effect on virus particles due to co-infection (Apte-Deshpande et al., 2014).
Another very important specie of bacteria is Bacillus thuringiensis var. israelensis (Bti), a gram-positive entomopathogenic bacterium that has been widely used in the control of mosquitoes (Boyce et al., 2013). This bacteria’s efficacy can be due to the action of six proteins (Cry4Aa, Cry4Ba, Cry10Aa, Cry11Aa, Cyt1Aa, and Cyt2Ba) (Berry et al., 2002; Park et al., 2005), which are deposited in inclusions that become part of the parasporal crystal of Bti (Bayyareddy et al., 2009). The main mechanism of toxicity of Cry proteins is the lysis of midgut epithelial cells mediated by the formation of pores (Bravo et al., 2007). Cry4B is one of the most effective toxin against Aedes mosquitoes. In this sense, it was reported that target proteins in the midgut of Ae. aegypti larvae, including the prohibitin (PHB), which may also be a receptor for the entry of DENV into mosquito cells (Bayyareddy et al., 2009; Kuadkitkan et al., 2012), as well as it may also be target of the mosquitocidal Cry4B protein. Kuadkitkan et al. (2012) have shown that Aedes cell lines bound to PHB are more susceptible to DENV infection, however, they do not undergo cytolysis in the presence of the Cry4B toxin (Kuadkitkan et al., 2012). These authors also showed a considerable colocalization between Cry4B and PHB in whole cells, which can protect the mosquito from cell death (Kuadkitkan et al., 2012). Therefore, Wintachai et al. (2012) were the first to identify that PHB is also a CHIKV receptor protein in the mosquito gut. Thus, we can suggest that PHB has a direct interaction with CHIKV and Cry4B during infection, which can protect the mosquito from the cytotoxicity of Cry4B (Wintachai et al., 2012).
Key Role of Protozoans
In Ae. aegypti and Ae. albopictus has been reported high prevalence of some species of Ascogregarina (Apicomplexa: Lecudinidae), such as Ascogregarina culicis and Ascogregarina taiwanensis, respectively (Blackmore et al., 1995). Larvae become infected after ingestion of oocysts containing sporozoites from its habitat and are vulnerable to gregarine infection at all larval instars (Kobayashi, 2006; Erthal et al., 2012) The sporozoites of Ascogregarina infects the epithelial cells of mosquitoes, develop the intracellular form (trophozoite) in the midgut, and subsequently rupture the epithelial cells and are released into the intestinal lumen (Chen, 1999; Lantova and Volf, 2014). Muniaraj et al. (2010) related the role of the cyst of Ascogregarina in maintenance of the CHIKV during silent period in Ae. albopictus. As the midgut is the primary region for virus replication in the mosquitoes, Mourya et al. (2003) stated that CHIKV may exploit these parasites for its own survival and transmission, being maintained in the oocysts of the parasites for a some period of time in nature. After filling breeding places with water, a new mosquito cycle starts and emerging larvae pick up the infection of the parasite by ingesting the oocysts that harbor the virus. So, the emerging adults start the mosquito-human-mosquito virus transmission cycle (Mourya et al., 2003).
Mourya et al. (2003) suggest that the gregarine A. culicis could help to maintain the CHIKV inside the mosquitoes. They demonstrated that uninfected first instar Ae. aegypti larvae fed on the dried adult mosquito homogenates obtained from the larvae that were exposed to oocysts, and CHIKV extracts were positive for viral antigens (Mourya et al., 2003). However, the uninfected larvae that fed only on the dried virus stock were negative, indicating that the virus does not survive in dried conditions. Therefore, the authors suggested that Ascogregarina spp. might play a crucial role in the maintenance of CHIKV during the inter-epidemic phases. Later, Muniaraj et al. (2010) described severe outbreaks of chikungunya fever during 2006–2007 in Kerala and Tamil Nadu, India (Muniaraj et al., 2010). Based on the hypotheses of Mourya the authors investigate the prevalence of Ascogregarina spp. in Ae. albopictus in 35 samples from different containers. They found a prevalence of 71.62% of Ascogregarina infection (including larvae, pupae and adults). If Ascogregarina could really be responsible for Aedes mosquito’s infection, as suggested by both authors, a more precise study must be conducted (Muniaraj et al., 2010).
Key Role of Nemathelminthes
To date, just few studies relating Nemathelminthes and Chikungunya virus transmissions have been conducted (Zytoon et al., 1993a,b). Zytoon et al. (1993b) described that Ae. albopictus infected with microfilariae of Dirofilaria immitis were more susceptible to transovarial transmission of CHIKV whereas non-infected mosquitoes did not transmit the CHIKV to the F1 progenitor. They feed the mosquiotes with defibrinated sheep blood containing 5 × 107 PFU of CHIKV and 20,000 microfilariae of D. immitis per milliliter observing that eggs of the first ovarian cycle were infected with CHIKV, which indicates that transovarial transmission occurred following coinfection by dissemination of the virus from female mosquitoes’ ovarian cells to eggs during fertilization (Zytoon et al., 1993b). In addition, the group also described that the microfilariae movement through the gut wall of Ae. albopictus increased CHIKV dissemination in the mosquito allowing some virions attached to the sheath of microfilariae to be carried to the hemocoel or that virus particles could leak through the holes produced when the parasite penetrate the gut wall and move into the hemocoel (Zytoon et al., 1993a).
In addition, Montarsi et al. (2015) described that Aedes koreicus, a new invasive species of mosquito in Europe, is a competent vector of D. immitis. The microfilariae was found in many parts of this mosquito (Montarsi et al., 2015). Later, Ciocchetta et al. (2018) explored the potential of Ae. koreicus to transmit CHIKV (Ciocchetta et al., 2018). They observed the dissemination of the virus to the wings and legs of the mosquito and infection of mosquito saliva, with live virus, occurred in two mosquitoes. These data demonstrate that Ae. koreicus could be a possible vector of CHIKV transmission in Europe (Ciocchetta et al., 2018). Those data corroborating with the fact that the mosquito can infected with D. immitis, could increasing the possibility of virus transmission. However, more studies have to be conducted in order to confirm the competence of this vector for CHIKV.
Chikungunya Virus Infection in Vertebrate Hosts
CHIKV was first discovered in 1952 in Tanzania. Its name is came from the Makonde word, and describes the body movement stiffness associated with persistent arthralgic symptoms presented by the host, after virus infection. Currently, chikungunya fever is a global epidemic, affecting millions of people due to several factors, including an increased number of travelers and the geographic distribution of mosquito vectors around the world (Mourya et al., 2003; Mourya and Yadav, 2006; Tsetsarkin et al., 2007). Symptoms of CHIKV include high fever, headache, rigors, photophobia and a petechial or maculopapular rash which is sometimes associated with arthralgia and myalgia (Schwartz and Albert, 2010; van Duijl-Richter et al., 2015). Some CHIKV-infected individuals present more severe symptoms, including hemorrhage, hepatitis, cranial nerve palsies and/or Guillain-Barre syndrome (Petitdemange et al., 2015; Mehta et al., 2018).
CHIKV belongs to the Togaviridae family, along with 29 different species of the Alphavirus genus; they cause diseases in humans and other mammals. The virus is small, with a 60–70 nm diameter, enveloped, spherical, positive-strand RNA virus with genomes of approximately 12 kb, encoding two polyproteins (Solignat et al., 2009; Hollidge et al., 2011). The first polyprotein consists of four non-structural units (nsP1, nsP2, nsP3, and nsP4) and the second is a structural polyprotein composed of five expression units (Capsid, E3, E2, 6K and E1). A sub genomic positive-strand RNA denominate 26SRNA is replicated from a negative-stranded RNA intermediate, which serves as template for the synthesis of viral structural proteins. A subgenomic positive-strand RNA denominate 26SRNA is transcribed from a negative-stranded RNA intermediate, which serves as template for the synthesis of viral structural proteins. Most of the alphaviruses have conserved domains involved in regulation of viral RNA synthesis. The E1 and E2 glycoproteins can form heterodimers that cover the viral surface uniformly and modulates the attachment of the virus to the host cells (van Duijl-Richter et al., 2015). The control of viral replication is regulated exogenous RNAi pathway in mosquitoes; this could limit potential pathologic effects and facilitate the arthropod survival (Coffey et al., 2014).
The infection starts when an infected mosquito bites the susceptible human host. The CHIKV spreads through the dermis infecting fibroblasts, macrophages and monocytes following an acute viremia that reaches 109–1012 viral particles per milliliter (Her et al., 2010; Wikan et al., 2012). The receptors used by CHIKV are ubiquitously expressed in many cells, specifically prohibitin, phosphatidylserine, glycosaminoglycans and ATP-synthase β subunit receptors that have been previously characterized (van Duijl-Richter et al., 2015). Interestingly, PBMCs are not the main source of the virus upon infection, suggesting that dermal fibroblasts, endothelial cells and monocytes or macrophages keep reproducing the virus during the infection progress (Sourisseau et al., 2007; Kumar et al., 2012). Since most of these target cells reside in the peripheral tissues, this could be the reason why the CHIKV viremia decreases significantly about 7 days after infection, even with the virus remaining in the body. Chronic disease is associated with the viral persistence in muscle and synovial cells and is manifested by myalgia and arthralgia, which can persist for years (Burt et al., 2017).
Several studies have already reported that CHIKV replicates along the skin before reaches the bones and liver (Schwartz and Albert, 2010; Lo Presti et al., 2014). In vitro experiments demonstrated that CHIKV can replicates in human epithelial and endothelial cell lines as well as macrophages and fibroblasts (Sourisseau et al., 2007). Patients with acute CHIKV (5–10 days after the onset of symptoms), when compared to a control group, presented higher systemic levels of IL-2R, IL-6, IL-1RA, IL-8, IL-10, MCP-1, IP-10, and MIG, and low levels of RANTES, whereas those in the chronic phase presented higher levels of MCP-1, IL-1RA, IL-6, IL-8, MIP-1α and MCP-1, returning to levels of uninfected controls in the recovered phase (Chaaithanya et al., 2011). IL-6, IL-8, MCP-1 and MIP-1α, MIP-1β have a determinant role in evolution for chronic disease.
Although very rare, neurological manifestations of CHIKV infections have been fully reviewed elsewhere (Brizzi, 2017; Mehta et al., 2018); these are mainly encephalopathy and encephalitis, encephalomyelopathy, myeloneuropathy, Guillain-Barré syndrome and neuro-ocular manifestations in adults and children infected directly via mosquito bites. Neonatal neurological features from vertical transmission also seem to play an important role in vertebrate infections, as has been previous described and reviewed elsewhere (Contopoulos-Ioannidis et al., 2018).
The molecular targets for CHIKV have already been exhaustively reviewed (Bautista-Reyes et al., 2017; da Silva-Júnior et al., 2017). nSP1, nSP2, and nSP3 are important potent inhibitors of the virus, as well as inhibitors to the known E1-E2. This has led to the compilation of a list of all-important molecular targets, as well as new lead molecules, including synthetic and natural products and designed compounds.
The Immunomodulatory Role of Aedes sp. Saliva on the CHIKV Infection in the Human
Many studies show the effects of Ae. aegypti insect salivary components and their role in pathogen infections (Styer et al., 2011; Surasombatpattana et al., 2012; Ockenfels et al., 2014). Recently, our study shows that SGE of Ae. aegypti was can able improve the survival of murine polymicrobial sepsis modulating neutrophil influx and increasing antioxidant defenses (de Souza Gomes et al., 2018). However, the most studies reported that components of mosquito or tick saliva facilitate the pathogenesis of the virus, improving its replication (Schneider and Higgs, 2008). The mosquito salivary gland produces and secretes molecules (e.g., NADH ubiquinone glutathione s-transferase, animal heme peroxidase, thioredoxin, cytochrome c oxidase) that aid in the digestion of sugars and nectars for their food (Ribeiro and Arca, 2009; Ribeiro et al., 2010); this may affect vascular constriction, blood coagulation, platelet aggregation (Schneider and Higgs, 2008; Ribeiro and Arca, 2009; Ribeiro et al., 2010), D7 protein (Calvo et al., 2009; Ribeiro et al., 2016) and Sialokinin, which modulates host immunity (Schneider et al., 2004; Wichit et al., 2016).
The Ae. aegypti salivary gland components modify the local microenvironment to favor the arbovirus infection (Dhawan et al., 2017). Several studies report that mosquitoes infected with CHIKV have an altered composition of salivary gland proteins, such as: putative inosine-uridine preferring nucleoside hydrolase, 30 kDa allergen-like protein, serpin, and angiopoietin-like protein variants (Tchankouo-Nguetcheu et al., 2012; Shrinet, 2018). SRPN26, protein disulfide-isomerase, tubulin beta chain, malic enzyme and RAN are down-regulated in the presence of CHIKV (Shrinet, 2018).
Some of the proteins that are up regulated with immunomodulatory effects have role in the transmission of CHIKV. For example, some proteins found in Aedes saliva have similarity to nucleoside hydrolases of other insects, such as the putative inosine-uridine preferring nucleoside hydrolase enzyme; which catalyzes preferentially the hydrolysis of inosine and uridine, and is increased in salivary gland of female mosquitoes, playing a role in blood feeding the insect (Ribeiro and Valenzuela, 2003). Mosquito salivary nucleosidases together with adenosine deaminases (ADA) found in Ae. aegypti saliva prevent mast cell degranulation by adenosine converting into hypoxanthine, which inhibits also production of inflammatory cytokines, prostanoids and leukotriene C4. Besides that, ADA produces inosine, which inhibits the production of inflammatory cytokines, decreasing the host response during blood feeding (Ribeiro and Valenzuela, 2003; Ribeiro et al., 2007; Tchankouo-Nguetcheu et al., 2012).
The 30 kDa allergen-like protein (Aed a 3), also called Aegyptin, that inhibits adhesion and aggregation platelet (Chagas et al., 2014). This protein recognizes the specific binding sites integrin a2β1, glycoprotein IV, and Von Willebrand factor (Armiyanti et al., 2015; Peng et al., 2015). The low molecular form (41 kDa) of serpin (serine proteinase inhibitors) is another protein that is up-regulated in Ae. aegypti salivary glands infected by CHIKV (Tchankouo-Nguetcheu et al., 2012). Serpin inhibit endogenous proteases, such as the serine proteases involved in regulating coagulation like factor Xa and thrombin (Wasinpiyamongkol et al., 2010). In this regards Serpin facilitates the blood feeding of the vector and as consequence enhance pathogen transmission from vector to host (Oktarianti et al., 2015).
The immunomodulatory effect of Ae. aegypti saliva in viral transmission involves local induction of chemotaxis and Th2 polarization bias Th1 (Boppana et al., 2009). Beside these proteins being up regulated in CHIKV, the Ae. aegypti salivary gland demonstrates capacity to modulate Th1/Th2 cytokine (Schneider et al., 2004). A study conducted by Thangamani et al. (2010) observed differences in immunological responses of CHIKV infection by needle inoculation compared to by mosquito bite. The infection with the presence of a mosquito’s saliva modulates a Th2 response with IL-4and IL-10 up-regulation (Venugopalan et al., 2014). In addition, the Th1 profile was also down regulated after saliva exposure, reducing IL-2 and IFN-γ levels, and the expression of TLR-3. Moreover, infected and non-infected mosquito bites reduced the expression of Toll-like receptors. The T-cell class shifts to a Th2 phenotype in the first 6 h after inoculation and a high number of eosinophils influx in the area of injection (Thangamani et al., 2010).
On the other hand, the proliferation of naïve lymphocytes is strongly impaired by the presence of SGE. Curiously, antigen-experienced lymphocytes were less sensitive to SGE inhibition, a mechanism dependent on antiapoptotic molecules. Indeed, the suppressive effect of Ae. aegypti on lymphocyte proliferation and biology was previously shown, and specific to T CD4 and T CD8 cells and not to DCs (Niedzwiecki et al., 2013).
Aedes aegypti saliva acts on the modulation of other immune system cells in favor of CHIKV infection. However, it has not yet been well established which components could be involved in this infection. Recently, Wichit et al. (2017) showed that Ae. aegypti saliva facilitates the replication of CHIKV leading to an increasing number of virus in human skin fibroblasts, besides that it also reduces the IFN-inducible gene in CHIKV-infected cells via the JAK-STAT. Among the reservoirs for viral propagation, the saliva of Ae. aegypti induces neutrophils the bite site by a mechanism dependent on the production of CXCL2, with the subsequent secretion of IL-1β and CCL-2 that leads to accumulation of neutrophils by directly mobilizing myeloid cells (Pingen et al., 2016).
In addition, components of the immunomodulatory process of the miRNAs present in Aedes saliva can modulate CHIKV infection. Maharaj et al. (2015) tested different types of miRNAs present in saliva of CHIKV infected mosquitoes, including miR-184, miR-12, miR-375 miR-125 and miR-2490. In this study, these miRNAs were silenced to evaluate their importance on CHIKV replication. Silencing these miRNAs promoted a decrease in viral replication rate, suggesting that the presence of these miRNAs up-regulates CHIKV replication or the activation of immune system cells (Maharaj et al., 2015), as shown in Figure 3.
FIGURE 3
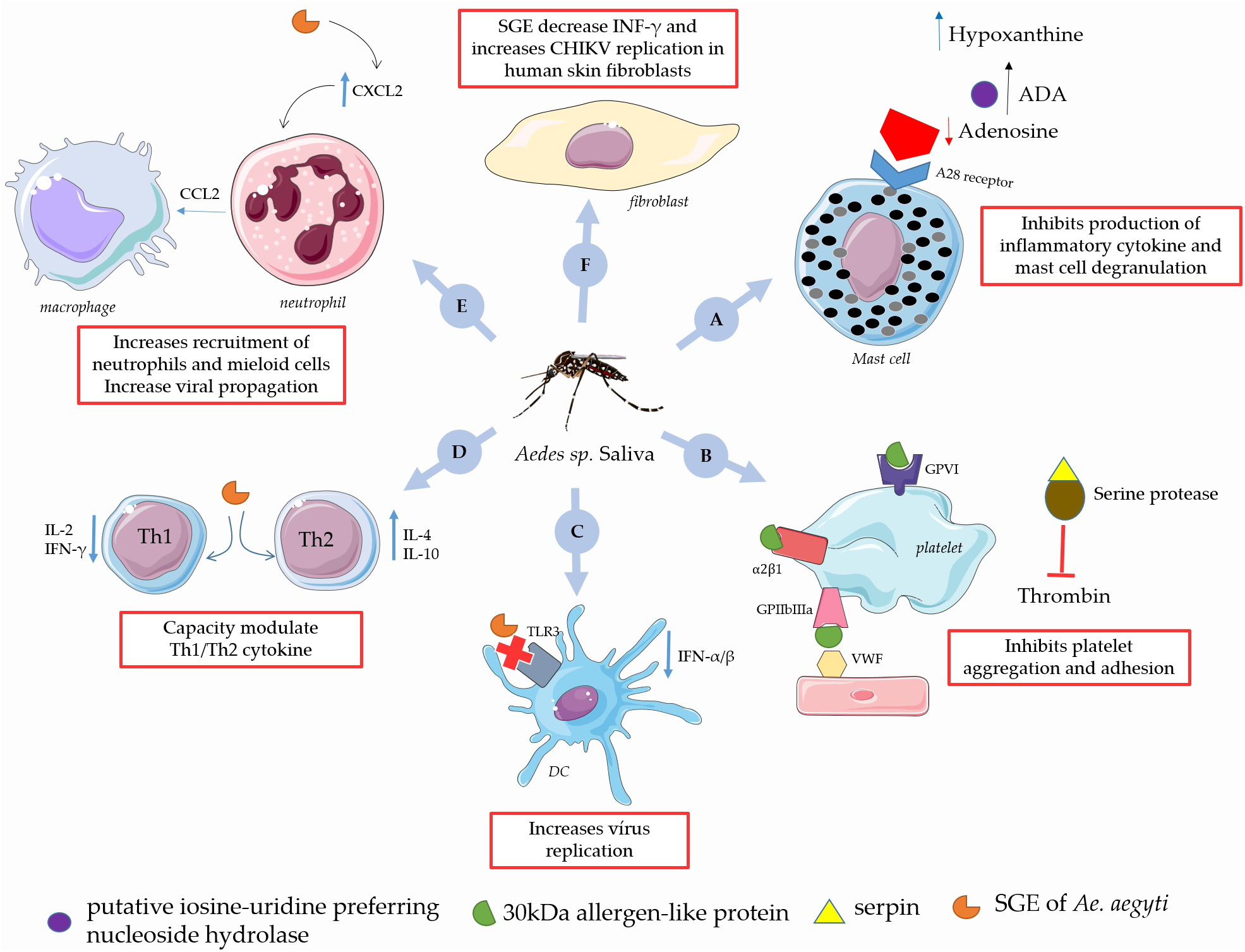
Immunomodulatory role of Aedes sp. saliva on the CHIKV infection in the human. (A)Ae. aegypti salivary putative inosine-uridine preferring nucleoside hydrolase together with adenosine deaminases (ADA) converts adenosine into hypoxanthine, thus preventing production of inflammatory cytokine and mast cell degranulation. (B) The 30 kDa allergen-like protein, is a collagen-binding protein that inhibits platelet aggregation, adhesion and serpin inhibit endogenous proteases which is important in the regulating of coagulation. (C) SGE of Aedes sp. reduce expression of TLR3 and decrease levels of IFN-α/β increasing virus replication. (D) SGE of Ae. aegypti modulates Th1/Th2 response in favor of CHIKV infection. (E)Ae. aegypti saliva increases influx of neutrophils to the local bite by a CXCL2 production, with the subsequent accumulated neutrophils and mobilize myeloid cells, which serve as reservoirs for viral propagation. (F) SGE of Ae. Aegypti increases CHIKV replication in human skin fibroblasts, and down-regulates the interferon IFN-inducible gene in CHIKV-infected cells via the JAK-STAT. This figure used elements from Servier Medical Art (www.servier.com).
Conclusion and Perspectives
Arbovirus infections, such as CHIKV, have been growing worldwide and are becoming a major public health problem, especially in immuno-compromised classes such as children and the elderly. CHIKV is dangerous since it can cause arthritis, fever and other impairments, making it necessary to seek new ways to counter the proliferation of this virus. Knowing the vector’s competence is fundamentally important, including an understanding of the vector virus interaction and the modulation of this interaction. Despite advances in the knowledge of mosquito competence in the transmission of CHIKV, little is known with respect to viral infection in the mosquito. However, it is important to highlight that the competence of the mosquito vector can be influenced by several characteristics, including the age of the mosquito, its nutritional status, stage of the gonotrophic cycle, environmental temperature conditions, salivary components and composition of its microbiota (Carrington et al., 2013; Bartholomay and Michel, 2018; Knecht et al., 2018). Thus, investigating the role of local mosquito populations, as well as kinetic and phenotypic characteristics of the different vectors for the transmission of pathogens, is very important in reducing the risk of possible disease outbreaks caused by many viruses, such as zika virus, dengue fever and yellow fever. In this regard, the mosquito microbiota is of fundamental importance to the viral infection of the mosquito. Bacteria of the Wolbachia genus are an important advance in the understanding of this interaction, since mosquitoes infected by this bacterium will not present viral infections, thus serving as an important tool in combating these viruses and reducing the transmission of DENV in endemic areas (O’Neill et al., 2018). However, we emphasize that there is a need for more robust work with natural populations, since most studies show the antiviral effects of Wolbachia against arboviruses, including DENV and CHIKV, in a model of stable transfection of this bacterium into heterologous mosquito hosts. In addition, mosquito saliva is an important factor in the transmission of CHIKV due to the existence of a series of substances with immunomodulatory and anticoagulants actions. These components may help the mosquito in blood ingestion, but will also allow CHIKV to escape from the host, and are thus fundamentally important to the virulence and pathogenicity of this virus. However, as the microbiota and salivary components of arthropods are highly diverse, together, these factors seem to contribute to the transmission of pathogens; when tested alone, they sometimes oppose the infection and in other situations may facilitate it. Therefore, it is necessary to actively search for new ways to modulate the transmission of CHIKV in the mosquito, which would create a tool to combat the virus.
Statements
Author contributions
All authors participated in the design of the study and drafted the manuscript. MM participated in the study coordination and helped to draft the manuscript. KN-L have designed and prepared the manuscript figures. All authors read and approved the final manuscript.
Funding
The authors were supported by the following Brazilian agencies: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação Amazônia Paraense de Amparo à Pesquisa (FAPESPA), Federal University of Parà and MM thanks for the fellowship from CNPq.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
- BsB
binary protein
- CHIKV
chikungunya virus
- CXCL2
chemokine (C-X-C motif) ligand 2
- DC
dendritic cells
- DENV
dengue virus
- IFN
interferon
- Ig
immunoglobulin
- IL
interleukin
- MCP
monocyte chemotactic protein
- MIG
monokine induce by interferon-γ
- MIP
macrophage inflammatory protein
- NK
natural killer
- PBMCs
peripheral blood mononuclear cells
- SGE
salivary gland extract
- Th
T helper
- TNF
tumor necrosis factor
References
1
Agha S. B. Chepkorir E. Mulwa F. Tigoi C. Arum S. Guarido M. M. et al (2017). Vector competence of populations of Aedes aegypti from three distinct cities in Kenya for chikungunya virus.PLoS Negl. Trop. Dis.11:e0005860. 10.1371/journal.pntd.0005860
2
Apte-Deshpande A. D. Paingankar M. S. Gokhale M. D. Deobagkar D. N. (2014). Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for chikungunya virus.Indian J. Med. Res.139762–768.
3
Armiyanti Y. Nuryady M. M. Arifianto R. P. Nurmariana E. Senjarini K. Fitri L. E. et al (2015). Detection of immunogenic proteins from Anopheles sundaicus salivary glands.Rev. Soc. Bras. Med. Trop.48410–416. 10.1590/0037-8682-0185-2015
4
Arnoult D. Carneiro L. Tattoli I. Girardin S. E. (2009). The role of mitochondria in cellular defense against microbial infection.Semin. Immunol.21223–232. 10.1016/j.smim.2009.05.009
5
Bartholomay L. C. Michel K. (2018). Mosquito immunobiology: the intersection of vector health and vector competence.Annu. Rev. Entomol.63145–167. 10.1146/annurev-ento-010715-023530
6
Bautista-Reyes E. Núñez-Avellaneda D. Alonso-Palomares L. A. Salazar M. I. (2017). Chikungunya: molecular aspects, clinical outcomes and pathogenesis.Rev. Investig. Clin.69299–307. 10.24875/RIC.17002029
7
Bayyareddy K. Andacht T. M. Abdullah M. A. Adang M. J. (2009). Proteomic identification of Bacillus thuringiensis subsp. israelensis toxin Cry4Ba binding proteins in midgut membranes from Aedes (Stegomyia) aegypti Linnaeus (Diptera, Culicidae) larvae.Insect Biochem. Mol. Biol.39279–286. 10.1016/j.ibmb.2009.01.002
8
Berry C. O’Neil S. Ben-Dov E. Jones A. F. Murphy L. Quail M. A. et al (2002). Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis.Appl. Environ. Microbiol.685082–5095. 10.1128/AEM.68.10.5082-5095.2002
9
Bidlingmayer W. L. (1974). The influence of environmental factors and physiological stage on flight patterns of mosquitoes taken in the vehicle aspirator and truck, suction, bait and New Jersey light traps.J. Med. Entomol.11119–146. 10.1093/jmedent/11.2.119
10
Blackmore M. S. Scoles G. A. Craig G. B. (1995). Parasitism of Aedes aegypti and Ae. albopictus (Diptera: Culicidae) by Ascogregarina spp. (Apicomplexa: Lecudinidae) in Florida.J. Med. Entomol.32847–852. 10.1093/jmedent/32.6.847
11
Boppana V. D. Thangamani S. Adler A. J. Wikel S. K. (2009). SAAG-4 is a novel mosquito salivary protein that programmes host CD4 T cells to express IL-4.Parasite Immunol.31287–295. 10.1111/j.1365-3024.2009.01096.x
12
Boyce R. Lenhart A. Kroeger A. Velayudhan R. Roberts B. Horstick O. (2013). Bacillus thuringiensis israelensis (Bti) for the control of dengue vectors: systematic literature review.Trop. Med. Int. Health18564–577. 10.1111/tmi.12087
13
Bravo A. Gill S. S. Soberón M. (2007). Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control.Toxicon49423–435. 10.1016/j.toxicon.2006.11.022
14
Brizzi K. (2017). Neurologic manifestation of chikungunya virus.Curr. Infect. Dis. Rep.19:6. 10.1007/s11908-017-0561-1
15
Brown J. E. Mcbride C. S. Johnson P. Ritchie S. Paupy C. Bossin H. et al (2011). Worldwide patterns of genetic differentiation imply multiple “domestications” of Aedes aegypti, a major vector of human diseases.Proc. R. Soc. B Biol. Sci.2782446–2454. 10.1098/rspb.2010.2469
16
Burt F. J. Chen W. Miner J. J. Lenschow D. J. Merits A. Schnettler E. et al (2017). Chikungunya virus: an update on the biology and pathogenesis of this emerging pathogen.Lancet Infect. Dis.17e107–e117. 10.1016/S1473-3099(16)30385-1
17
Calvo E. Mans B. J. Ribeiro J. M. C. Andersen J. F. (2009). Multifunctionality and mechanism of ligand binding in a mosquito antiinflammatory protein.Proc. Natl. Acad. Sci. U.S.A.1063728–3733. 10.1073/pnas.0813190106
18
Carrington L. B. Seifert S. N. Armijos M. V. Lambrechts L. Scott T. W. (2013). Reduction of Aedes aegypti vector competence for dengue virus under large temperature fluctuations.Am. J. Trop. Med. Hyg.88689–697. 10.4269/ajtmh.12-0488
19
Carrington L. B. Simmons C. P. (2014). Human to mosquito transmission of dengue viruses.Front. Immunol.5:290. 10.3389/fimmu.2014.00290
20
Chaaithanya I. K. Muruganandam N. Sundaram S. G. Kawalekar O. Sugunan A. P. Manimunda S. P. et al (2011). Role of proinflammatory cytokines and chemokines in chronic arthropathy in CHIKV infection.Viral Immunol.24265–271. 10.1089/vim.2010.0123
21
Chagas A. C. Ramirez J. L. Jasinskiene N. James A. A. Ribeiro J. M. C. Marinotti O. et al (2014). Collagen-binding protein, Aegyptin, regulates probing time and blood feeding success in the dengue vector mosquito, Aedes aegypti.Proc. Natl. Acad. Sci. U.S.A.1116946–6951. 10.1073/pnas.1404179111
22
Chavshin A. R. Oshaghi M. A. Vatandoost H. Yakhchali B. Zarenejad F. Terenius O. (2015). Malpighian tubules are important determinants of Pseudomonas transstadial transmission and longtime persistence in Anopheles stephensi.Parasit. Vectors8:36. 10.1186/s13071-015-0635-6
23
Chen W. J. (1999). The life cycle of Ascogregarina taiwanensis (Apicomplexa: Lecudinidae).Parasitol. Today15153–156. 10.1016/S0169-4758(99)01418-0
24
Chin P. A. Ehrbar D. J. Micieli M. V. Fonseca D. M. Ciota A. T. Kramer L. D. (2018). Differential effects of temperature and mosquito genetics determine transmissibility of Arboviruses by Aedes aegypti in Argentina.Am. J. Trop. Med. Hyg.991–19. 10.4269/ajtmh.18-0097
25
Chouin-Carneiro T. Vega-Rua A. Vazeille M. Yebakima A. Girod R. Goindin D. et al (2016). Differential Susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus.PLoS Negl. Trop. Dis.10:e0004543. 10.1371/journal.pntd.0004543
26
Ciocchetta S. Prow N. A. Darbro J. M. Frentiu F. D. Savino S. Montarsi F. et al (2018). The new European invader Aedes (Finlaya) koreicus: a potential vector of chikungunya virus.Pathog. Glob. Health112107–114. 10.1080/20477724.2018.1464780
27
Coffey L. L. Failloux A. B. Weaver S. C. (2014). Chikungunya virus–vector interactions.Viruses64628–4663. 10.3390/v6114628
28
Contopoulos-Ioannidis D. Newman-Lindsay S. Chow C. LaBeaud A. D. (2018). Mother-to-child transmission of Chikungunya virus: a systematic review and meta-analysis.PLoS Negl. Trop. Dis.12:e0006510. 10.1371/journal.pntd.0006510
29
Coon K. L. Brown M. R. Strand M. R. (2016). Mosquitoes host communities of bacteria that are essential for development but vary greatly between local habitats.Mol. Ecol.255806–5826. 10.1111/mec.13877
30
da Silva-Júnior E. F. Leoncini G. O. Rodrigues ÉE. S. Aquino T. M. Araújo-Júnior J. X. (2017). The medicinal chemistry of Chikungunya virus.Bioorg. Med. Chem.254219–4244. 10.1016/j.bmc.2017.06.049
31
Day J. F. (2016). Mosquito oviposition behavior and vector control.Insects7:E65. 10.3390/insects7040065
32
de Souza Gomes R. Navegantes-Lima K. C. Monteiro V. V. S. de Brito Oliveira A. L. Rodrigues D. V. S. Reis J. F. et al (2018). Salivary gland extract from Aedes aegypti improves survival in murine polymicrobial sepsis through oxidative mechanisms.Cells7:182. 10.3390/cells7110182
33
Demaio J. Pumpuni C. B. Kent M. Beier J. C. (1996). The midgut bacterial flora of wild Aedes triseriatus, Culex pipiens, and Psorophora columbiae mosquitoes.Am. J. Trop. Med. Hyg.54219–223. 10.4269/ajtmh.1996.54.219
34
Dennison N. J. Jupatanakul N. Dimopoulos G. (2014). The mosquito microbiota influences vector competence for human pathogens.Curr. Opin. Insect Sci.36–13. 10.1016/j.cois.2014.07.004
35
Dhawan R. Kumar M. Mohanty A. K. Dey G. Advani J. Prasad T. S. K. et al (2017). Mosquito-borne diseases and omics: salivary gland proteome of the female Aedes aegypti Mosquito.Omi. A J. Integr. Biol.2145–54. 10.1089/omi.2016.0160
36
Diagne C. T. Faye O. Guerbois M. Knight R. Diallo D. Faye O. et al (2014). Vector competence of aedes aegypti and aedes vittatus (Diptera: Culicidae) from Senegal and Cape Verde archipelago for West African lineages of chikungunya virus.Am. J. Trop. Med. Hyg.91635–641. 10.4269/ajtmh.13-0627
37
Dubrulle M. Mousson L. Moutailier S. Vazeille M. Failloux A. B. (2009). Chikungunya virus and Aedes mosquitoes: saliva is infectious as soon as two days after oral infection.PLoS One4:e5895. 10.1371/journal.pone.0005895
38
Elbers A. R. W. Koenraadt C. J. M. Meiswinkel R. (2015). Mosquitoes and Culicoides biting midges: vector range and the influence of climate change.Rev. Sci. Tech.34123–137. 10.20506/rst.34.1.2349
39
Erthal J. A. Soghigian J. S. Livdahl T. (2012). Life Cycle Completion of Parasite Ascogregarina taiwanensis (Apicomplexa: Lecudinidae) in Non-Native Host Ochlerotatus japonicus (Diptera: Culicidae).J. Med. Entomol.491109–1117. 10.1603/ME12018
40
Favia G. Ricci I. Damiani C. Raddadi N. Crotti E. Marzorati M. et al (2007). Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector.Proc. Natl. Acad. Sci. U.S.A.1049047–9051. 10.1073/pnas.0610451104
41
Franz A. W. E. Kantor A. M. Passarelli A. L. Clem R. J. (2015). Tissue barriers to arbovirus infection in mosquitoes.Viruses73741–3767. 10.3390/v7072795
42
Fraser J. E. De Bruyne J. T. Iturbe-Ormaetxe I. Stepnell J. Burns R. L. Flores H. A. et al (2017). Novel Wolbachia-transinfected Aedes aegypti mosquitoes possess diverse fitness and vector competence phenotypes.PLoS Pathog.13:e1006751. 10.1371/journal.ppat.1006751
43
Gaio A. D. O. Gusmão D. S. Santos A. V. Berbert-Molina M. A. Pimenta P. F. P. Lemos F. J. A. (2011). Contribution of midgut bacteria to blood digestion and egg production in Aedes aegypti (diptera: Culicidae) (L.).Parasit. Vectors4:105. 10.1186/1756-3305-4-105
44
Göertz G. P. Vogels C. B. F. Geertsema C. Koenraadt C. J. M. Pijlman G. P. (2017). Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti.PLoS Negl. Trop. Dis.11:e0005654. 10.1371/journal.pntd.0005654
45
Goindin D. Cannet A. Delannay C. Ramdini C. Gustave J. Atyame C. et al (2018). Screening of natural Wolbachia infection in Aedes aegypti, Aedes taeniorhynchus and Culex quinquefasciatus from Guadeloupe (French West Indies).Acta Trop.185314–317. 10.1016/j.actatropica.2018.06.011
46
Gusmão D. S. Santos A. V. Marini D. C. Russo ÉD. S. Peixoto A. M. D. Bacci M. et al (2007). First isolation of microorganisms from the gut diverticulum of Aedes aegypti (Diptera: Culicidae): new perspectives for an insect-bacteria association.Mem. Inst. Oswaldo Cruz.102919–924. 10.1590/S0074-02762007000800005
47
Hedges L. M. Brownlie J. C. O’Neill S. L. Johnson K. N. (2008). Wolbachia and virus protection in insects.Science322:702. 10.1126/science.1162418
48
Hegde S. Rasgon J. L. Hughes G. L. (2015). The microbiome modulates arbovirus transmission in mosquitoes.Curr. Opin. Virol.1597–102. 10.1016/j.coviro.2015.08.011
49
Her Z. Malleret B. Chan M. Ong E. K. S. Wong S. C. Kwek D. J. C. et al (2010). Active infection of human blood monocytes by chikungunya virus triggers an innate immune response.J. Immunol.1845903–5913. 10.4049/jimmunol.0904181
50
Hollidge B. S. Weiss S. R. Soldan S. S. (2011). The role of interferon antagonist, non-structural proteins in the pathogenesis and emergence of arboviruses.Viruses3629–658. 10.3390/v3060629
51
Honório N. A. Câmara D. C. P. Calvet G. A. Brasil P. (2015). Chikungunya: an arbovirus infection in the process of establishment and expansion in Brazil.Cad. Saude Publica31906–908. 10.1590/0102-311xpe020515
52
Hughes G. L. Rasgon J. L. (2014). Transinfection: a method to investigate Wolbachia-host interactions and control arthropod-borne disease.Insect Mol. Biol.23141–151. 10.1111/imb.12066
53
Johnson K. N. (2015). The impact of Wolbachia on virus infection in mosquitoes.Viruses75705–5717. 10.3390/v7112903
54
Juhn J. Naeem-Ullah U. MacIel Guedes B. A. Majid A. Coleman J. Paolucci Pimenta P. F. et al (2011). Spatial mapping of gene expression in the salivary glands of the dengue vector mosquito, aedes aegypti.Parasit. Vectors4:1. 10.1186/1756-3305-4-1
55
Kambris Z. Blagborough A. M. Pinto S. B. Blagrove M. S. C. Godfray H. C. J. Sinden R. E. et al (2010). Wolbachia stimulates immune gene expression and inhibits plasmodium development in anopheles gambiae.PLoS Pathog.6:e1001143. 10.1371/journal.ppat.1001143
56
Kambris Z. Cook P. E. Phuc H. K. Sinkins S. P. (2009). Immune activation by life-shortening wolbachia and reduced filarial competence in mosquitoes.Science326134–136. 10.1126/science.1177531
57
Knecht H. Richards S. Balanay J. White A. (2018). Impact of mosquito age and insecticide exposure on susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus.Pathogens7:67. 10.3390/pathogens7030067
58
Kobayashi M. (2006). New findings on the developmental process of ascogregarina taiwanensis and ascogregarina culicis in aedes albopictus and aedes aegypti.Am. Mosq. Control Assoc.2229–36. 10.2987/8756-971X(2006)22[29:NFOTDP]2.0.CO;2
59
Kraemer M. U. G. Sinka M. E. Duda K. A. Mylne A. Q. N. Shearer F. M. Barker C. M. et al (2015). The global distribution of the arbovirus vectors Aedes aegypti and Ae. Albopictus.Elife4:e08347. 10.7554/eLife.08347
60
Kuadkitkan A. Smith D. R. Berry C. (2012). Investigation of the cry4B-prohibitin interaction in Aedes aegypti cells.Curr. Microbiol.65446–454. 10.1007/s00284-012-0178-4
61
Kumar S. Jaffar-Bandjee M.-C. Giry C. Connen de Kerillis L. Merits A. Gasque P. et al (2012). Mouse macrophage innate immune response to chikungunya virus infection.Virol. J.9:313. 10.1186/1743-422X-9-313
62
Lantova L. Volf P. (2014). Infection, Genetics and Evolution Mosquito and sand fly gregarines of the genus Ascogregarina and Psychodiella (Apicomplexa: Eugregarinorida, Aseptatorina) – Overview of their taxonomy, life cycle, host specificity and pathogenicity.Infect. Genet. Evol.28616–627. 10.1016/j.meegid.2014.04.021
63
Le Coupanec A. Babin D. Fiette L. Jouvion G. Ave P. Misse D. et al (2013). Aedes mosquito saliva modulates rift valley fever virus pathogenicity.PLoS Negl. Trop. Dis.7:e2237. 10.1371/journal.pntd.0002237
64
Lo Presti A. Lai A. Cella E. Zehender G. Ciccozzi M. (2014). Chikungunya virus, epidemiology, clinics and phylogenesis: a review.Asian Pac. J. Trop. Med.7925–932. 10.1016/S1995-7645(14)60164-4
65
Maharaj P. D. Widen S. G. Huang J. Wood T. G. Thangamani S. (2015). Discovery of Mosquito Saliva MicroRNAs during CHIKV Infection.PLoS Negl. Trop. Dis.9:e0003386. 10.1371/journal.pntd.0003386
66
McCracken M. K. Christofferson R. C. Grasperge B. J. Calvo E. Chisenhall D. M. Mores C. N. (2014). Aedes aegypti salivary protein “aegyptin” co-inoculation modulates dengue virus infection in the vertebrate host.Virology46133–139. 10.1016/j.virol.2014.07.019
67
Mehta R. Gerardin P. de Brito C. A. A. Soares C. N. Ferreira M. L. B. Solomon T. (2018). The neurological complications of chikungunya virus: a systematic review.Rev. Med. Virol.28:e1978. 10.1002/rmv.1978
68
Melo B. Silva N. Gomes R. Navegantes K. Oliveira A. Almeida L. et al (2015). Bioactive compounds of the salivary glands from aedes aegypti with anti-hemostatic action.Annu. Res. Rev. Biol.81–17. 10.9734/ARRB/2015/20322
69
Montarsi F. Ciocchetta S. Devine G. Ravagnan S. Mutinelli F. Frangipane Di Regalbono A. et al (2015). Development of Dirofilaria immitis within the mosquito Aedes (Finlaya) koreicus, a new invasive species for Europe.Parasit. Vectors8:177. 10.1186/s13071-015-0800-y
70
Moreira L. A. Iturbe-Ormaetxe I. Jeffery J. A. Lu G. Pyke A. T. Hedges L. M. et al (2009a). A Wolbachia Symbiont in Aedes aegypti limits infection with dengue, chikungunya, and plasmodium.Cell1391268–1278. 10.1016/j.cell.2009.11.042
71
Moreira L. A. Saig E. Turley A. P. Ribeiro J. M. C. O’Neill S. L. McGraw E. A. (2009b). Human probing behavior of Aedes aegypti when infected with a life-shortening strain of Wolbachia.PLoS Negl. Trop. Dis.3:e568. 10.1371/journal.pntd.0000568
72
Mourya D. T. Singh D. K. Yadav P. Gokhale M. D. Barde P. V. Narayan N. B. et al (2003). Role of gregarine parasite Ascogregarina culicis (Apicomplexa: Lecudinidae) in the maintenance of Chikungunya virus in vector mosquito.J. Eukaryot. Microbiol.50379–382. 10.1111/j.1550-7408.2003.tb00153.x
73
Mourya D. T. Yadav P. (2006). Vector biology of dengue & chikungunya viruses.Indian J. Med. Res.124475–480.
74
Mousson L. Martin E. Zouache K. Madec Y. Mavingui P. Failloux A. B. (2010). Wolbachia modulates Chikungunya replication in Aedes albopictus.Mol. Ecol.191953–1964. 10.1111/j.1365-294X.2010.04606.x
75
Muniaraj M. Rajendran R. Arunachalam N. Paramasivan R. Philip S. P. (2010). Prevalence of ascogregarina spp. in the container breeding aedes albopictus from chikungunya fever affected areas of Kerala State.India J. Commun. Dis.42157–159.
76
Niedzwiecki M. M. Hall M. N. Liu X. Oka J. Harper K. N. Slavkovich V. et al (2013). A dose-response study of arsenic exposure and global methylation of peripheral blood mononuclear cell DNA in Bangladeshi adults.Environ. Health Perspect.1211306–1312. 10.1289/ehp.6964
77
Ockenfels B. Michael E. McDowell M. A. (2014). Meta-analysis of the effects of insect vector saliva on host immune responses and infection of vector-transmitted pathogens: a focus on leishmaniasis.PLoS Negl. Trop. Dis.8:e3197. 10.1371/journal.pntd.0003197
78
Oktarianti R. Senjarini K. Hayano T. Fatchiyah F. Aulanni’am (2015). Proteomic analysis of immunogenic proteins from salivary glands of Aedes aegypti.J. Infect. Public Health8575–582. 10.1016/j.jiph.2015.04.022
79
O’Neill S. L. Ryan P. A. Turley A. P. Wilson G. Retzki K. Iturbe-Ormaetxe I. et al (2018). Scaled deployment of Wolbachia to protect the community from Aedes transmitted arboviruses [version 1; referees: 1 approved].Gates Open Res.2:36. 10.12688/gatesopenres.12844.1
80
Park H. W. Bideshi D. K. Wirth M. C. Johnson J. J. Walton W. E. Federici B. A. et al (2005). Recombinant larvicidal bacteria with markedly improved efficacy against Culex vectors of West Nile virus.Am. J. Trop. Med. Hyg.72732–738. 10.4269/ajtmh.2005.72.732
81
Patil C. D. Patil S. V. Salunke B. K. Salunkhe R. B. (2011). Prodigiosin produced by Serratia marcescens NMCC46 as a mosquito larvicidal agent against Aedes aegypti and Anopheles stephensi.Parasitol. Res.1091179–1187. 10.1007/s00436-011-2365-9
82
Patil C. D. Patil S. V. Salunke B. K. Salunkhe R. B. (2012). Insecticidal potency of bacterial species Bacillus thuringiensis SV2 and Serratia nematodiphila SV6 against larvae of mosquito species Aedes aegypti, Anopheles stephensi, and Culex quinquefasciatus.Parasitol. Res.1101841–1847. 10.1007/s00436-011-2708-6
83
Peng Z. Xu W. W. Sham Y. Lam H. Sun D. Li C. et al (2015). Mosquito salivary allergen Aed a 3: cloning, comprehensive molecular analysis, and clinical evaluation.Allergy71621–628. 10.1111/all.12812
84
Petitdemange C. Wauquier N. Vieillard V. (2015). Control of immunopathology during chikungunya virus infection.J. Allergy Clin. Immunol.135846–855. 10.1016/j.jaci.2015.01.039
85
Pingen M. Bryden S. R. Pondeville E. Schnettler E. Kohl A. Merits A. et al (2016). Host inflammatory response to mosquito bites enhances the severity of arbovirus infection.Immunity441455–1469. 10.1016/j.immuni.2016.06.002
86
Raquin V. Moro C. V. Saucereau Y. Tran F. H. Potier P. Mavingui P. (2015). Native Wolbachia from Aedes albopictus blocks chikungunya virus infection in cellulo.PLoS One10:e0125066. 10.1371/journal.pone.0125066
87
Ribeiro J. M. Arca B. (2009). From Sialomes to the Sialoverse: an insight into the salivary potion of blood feeding insects.Adv. Insect Physiol.3759–118. 10.1016/S0065-2806(09)37002-2
88
Ribeiro J. M. C. Arcà B. Lombardo F. Calvo E. Phan V. M. Chandra P. K. et al (2007). An annotated catalogue of salivary gland transcripts in the adult female mosquito, Ædes ægypti.BMC Genomics8:6. 10.1186/1471-2164-8-6
89
Ribeiro J. M. C. Mans B. J. Arcà B. (2010). An insight into the sialome of blood-feeding Nematocera.Insect Biochem. Mol. Biol.40767–784. 10.1016/j.ibmb.2010.08.002
90
Ribeiro J. M. C. Martin-Martin I. Arcá B. Calvo E. (2016). A deep insight into the sialome of male and female aedes aegypti mosquitoes.PLoS One11:e0151400. 10.1371/journal.pone.0151400
91
Ribeiro J. M. C. Valenzuela J. G. (2003). The salivary purine nucleosidase of the mosquito, Aedes aegypti.Insect Biochem. Mol. Biol.3313–22. 10.1016/S0965-1748(02)00078-4
92
Richard V. Paoaafaite T. Cao-Lormeau V. M. (2016). Vector Competence of Aedes aegypti and Aedes polynesiensis populations from french polynesia for chikungunya virus.PLoS Negl. Trop. Dis.10:e0004694. 10.1371/journal.pntd.0004694
93
Rodríguez-Morales A. J. (2015). Zika: the new arbovirus threat for latin america.J. Infect. Dev. Ctries.9684–685. 10.3855/jidc.7230
94
Schneider B. S. Higgs S. (2008). The enhancement of arbovirus transmission and disease by mosquito saliva is associated with modulation of the host immune response.Trans. R. Soc. Trop. Med. Hyg.102400–408. 10.1016/j.trstmh.2008.01.024
95
Schneider B. S. Soong L. Coffey L. L. Stevenson H. L. McGee C. E. Higgs S. (2010). Aedes aegypti saliva alters leukocyte recruitment and cytokine signaling by antigen-presenting cells during west nile virus infection.PLoS One5:e11704. 10.1371/journal.pone.0011704
96
Schneider B. S. Soong L. Zeidner N. S. Higgs S. (2004). Aedes aegypti Salivary gland extracts modulate anti-viral and TH1/TH2 cytokine responses to sindbis virus infection.Viral Immunol.17565–573. 10.1089/vim.2004.17.565
97
Schwartz O. Albert M. L. (2010). Biology and pathogenesis of chikungunya virus.Nat. Rev. Microbiol.8491–500. 10.1038/nrmicro2368
98
Sharma P. Sharma S. Maurya R. K. Das De T. Thomas T. Lata S. et al (2014). Salivary glands harbor more diverse microbial communities than gut in Anopheles culicifacies.Parasit. Vectors7:235. 10.1186/1756-3305-7-235
99
Shrinet J. (2018). Understanding oxidative stress in aedes during chikungunya and dengue virus infections using.Viruses10:314. 10.3390/v10060314
100
Solignat M. Gay B. Higgs S. Briant L. Devaux C. (2009). Replication cycle of chikungunya: a re-emerging arbovirus.Virology393183–197. 10.1016/j.virol.2009.07.024
101
Sourisseau M. Schilte C. Casartelli N. Trouillet C. Guivel-Benhassine F. Rudnicka D. et al (2007). Characterization of reemerging chikungunya virus.PLoS Pathog.3:e89. 10.1371/journal.ppat.0030089
102
Styer L. M. Lim P.-Y. Louie K. L. Albright R. G. Kramer L. D. Bernard K. A. (2011). Mosquito saliva causes enhancement of west nile virus infection in mice.J. Virol.851517–1527. 10.1128/JVI.01112-10
103
Surasombatpattana P. Patramool S. Luplertlop N. Yssel H. Missé D. (2012). Aedes aegypti saliva enhances dengue virus infection of human keratinocytes by suppressing innate immune responses.J. Invest. Dermatol.1322103–2105. 10.1038/jid.2012.76
104
Suryawanshi R. K. Patil C. D. Borase H. P. Narkhede C. P. Salunke B. K. Patil S. V. (2015). Mosquito larvicidal and pupaecidal potential of prodigiosin from Serratia marcescens and understanding its mechanism of action.Pestic. Biochem. Physiol.12349–55. 10.1016/j.pestbp.2015.01.018
105
Tchankouo-Nguetcheu S. Bourguet E. Lenormand P. Rousselle J.-C. Namane A. Choumet V. (2012). Infection by chikungunya virus modulates the expression of several proteins in Aedes aegypti salivary glands.Parasit. Vectors5:264. 10.1186/1756-3305-5-264
106
Teixeira L. Ferreira Á Ashburner M. (2008). The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster.PLoS Biol.6:e2. 10.1371/journal.pbio.1000002
107
Thangamani S. Higgs S. Ziegler S. Vanlandingham D. Tesh R. Wikel S. (2010). Host immune response to mosquito-transmitted chikungunya virus differs from that elicited by needle inoculated virus.PLoS One5:e12137. 10.1371/journal.pone.0012137
108
Tsetsarkin K. A. Vanlandingham D. L. McGee C. E. Higgs S. (2007). A single mutation in Chikungunya virus affects vector specificity and epidemic potential.PLoS Pathog.3:e201. 10.1371/journal.ppat.0030201
109
van den Hurk A. F. Hall-Mendelin S. Pyke A. T. Frentiu F. D. McElroy K. Day A. et al (2012). Impact of wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti.PLoS Negl. Trop. Dis.6:e1892. 10.1371/journal.pntd.0001892
110
van Duijl-Richter M. Hoornweg T. Rodenhuis-Zybert I. Smit J. (2015). Early events in chikungunya virus infection—from virus cellbinding to membrane fusion.Viruses73647–3674. 10.3390/v7072792
111
Vega-Rúa A. Lourenço-de-Oliveira R. Mousson L. Vazeille M. Fuchs S. Yébakima A. et al (2015). Chikungunya virus transmission potential by local aedes mosquitoes in the Americas and Europe.PLoS Negl. Trop. Dis.9:e0003780. 10.1371/journal.pntd.0003780
112
Venugopalan A. Ghorpade R. P. Chopra A. (2014). Cytokines in acute chikungunya.PLoS One9:e111305. 10.1371/journal.pone.0111305
113
Wasinpiyamongkol L. Patramool S. Luplertlop N. Surasombatpattana P. Doucoure S. Mouchet F. et al (2010). Blood-feeding and immunogenic Aedes aegypti saliva proteins.Proteomics101906–1916. 10.1002/pmic.200900626
114
Wasserman H. A. Singh S. Champagne D. E. (2004). Saliva of the Yellow Fever mosquito, Aedes aegypti, modulates murine lymphocyte function.Parasite Immunol.26295–306. 10.1111/j.0141-9838.2004.00712.x
115
Weaver S. C. Lecuit M. (2015). Chikungunya virus and the global spread of a mosquito-borne disease.N. Engl. J. Med.3721231–1239. 10.1056/NEJMra1406035
116
Weaver S. C. Reisen W. K. (2010). Present and future arboviral threats.Antiviral Res.85328–345. 10.1016/j.antiviral.2009.10.008
117
Wichit S. Diop F. Hamel R. Talignani L. Ferraris P. Cornelie S. et al (2017). Aedes aegypti saliva enhances chikungunya virus replication in human skin fibroblasts via inhibition of the type I interferon signaling pathway.Infect. Genet. Evol.5568–70. 10.1016/j.meegid.2017.08.032
118
Wichit S. Ferraris P. Choumet V. Missé D. (2016). The effects of mosquito saliva on dengue virus infectivity in humans.Curr. Opin. Virol.21139–145. 10.1016/j.coviro.2016.10.001
119
Wikan N. Sakoonwatanyoo P. Ubol S. Yoksan S. Smith D. R. (2012). Chikungunya virus infection of cell lines: analysis of the east, central and south African lineage.PLoS One7:e31102. 10.1371/journal.pone.0031102
120
Wintachai P. Wikan N. Kuadkitkan A. Jaimipuk T. Ubol S. Pulmanausahakul R. et al (2012). Identification of prohibitin as a Chikungunya virus receptor protein.J. Med. Virol.841757–1770. 10.1002/jmv.23403
121
Wiwatanaratanabutr I. Zhang C. (2016). Wolbachia infections in mosquitoes and their predators inhabiting rice field communities in Thailand and China.Acta Trop.159153–160. 10.1016/j.actatropica.2016.03.026
122
Zouache K. Michelland R. J. Failloux A. B. Grundmann G. L. Mavingui P. (2012). Chikungunya virus impacts the diversity of symbiotic bacteria in mosquito vector.Mol. Ecol.212297–2309. 10.1111/j.1365-294X.2012.05526.x
123
Zouache K. Voronin D. Tran-Van V. Mousson L. Failloux A. B. Mavingui P. (2009). Persistent Wolbachia and cultivable bacteria infection in the reproductive and somatic tissues of the mosquito vector Aedes albopictus.PLoS One4:e6388. 10.1371/journal.pone.0006388
124
Zytoon E. M. El-Belbasi H. I. Matsumura T. (1993a). Mechanism of increased dissemination of chikungunya virus in Aedes albopictus mosquitoes concurrently ingesting microfilariae of Dirofilaria immitis.Am. J. Trop. Med. Hyg.49201–207. 10.4269/ajtmh.1993.49.201
125
Zytoon E. M. El-Belbasi H. I. Matsumura T. (1993b). Transovarial transmission of Chikungunya virus by Aedes albopictus mosquitoes ingesting microfilariae of Dirofilaria immitis under laboratory conditions.Microbiol. Immunol.37419–421. 10.1111/j.1348-0421.1993.tb03232.x
Summary
Keywords
Aedes aegypti , Aedes albopictus , arthropod-borne virus, CHIKV, virus maintenance, microorganisms
Citation
Monteiro VVS, Navegantes-Lima KC, de Lemos AB, da Silva GL, de Souza Gomes R, Reis JF, Rodrigues Junior LC, da Silva OS, Romão PRT and Monteiro MC (2019) Aedes–Chikungunya Virus Interaction: Key Role of Vector Midguts Microbiota and Its Saliva in the Host Infection. Front. Microbiol. 10:492. doi: 10.3389/fmicb.2019.00492
Received
14 September 2018
Accepted
26 February 2019
Published
09 April 2019
Volume
10 - 2019
Edited by
Tonya Michelle Colpitts, Boston University, United States
Reviewed by
Stefanie Christine Becker, University of Veterinary Medicine Hannover, Germany; Grant Leslie Hughes, Liverpool School of Tropical Medicine, United Kingdom; Ian Patterson, Liverpool School of Tropical Medicine, United Kingdom, in collaboration with reviewer GH
Updates
Copyright
© 2019 Monteiro, Navegantes-Lima, de Lemos, da Silva, de Souza Gomes, Reis, Rodrigues Junior, da Silva, Romão and Monteiro.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Chagas Monteiro, martachagas2@yahoo.com.br
This article was submitted to Virology, a section of the journal Frontiers in Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.