 Yi-ming Wang
Yi-ming Wang Ling-cong Kong
Ling-cong Kong Hong-xia Ma
Hong-xia Ma- College of Animal Science and Technology, Jilin Agricultural University, Changchun, China
Eugenol, the major active essential oil component of clove, was reported to possess QS (quorum sensing) inhibitory activity. A previous study found that eugenol could bind to quorum sensing receptors of Pseudomonas aeruginosa and down-regulate the expression of Streptococcus mutans virulence genes at sub-MIC (minimum inhibitory concentration) without affecting the bacterial growth. However, the alterations of QS signal molecules at transcription levels was not well understood. To better understand interactions of Klebsiella pneumoniae in response to eugenol and explore molecular regulations, transcriptome sequencing was performed. A total of 5779 differentially expressed genes (DEGs) enriched in a variety of biological processes and pathways were identified. The transcriptional data was validated by qPCR and the results showed that the expression profiles of 4 major genes involved in autoinducers-2 (AI-2) synthesis, including luxS, pfs, and lsrK were consistent with transcriptome analysis except for lsrR, a transcriptional repressor gene of lsr operon, which may repress the expression of following genes responsible for AI-2 signal transmission in vivo. In vitro AI-2 synthesis assay also revealed that eugenol could inhibit AI-2 generation. The results of our study offer insights into the mechanisms of QS inhibitory activity and K. pneumoniae AI-2 alterations after eugenol treatment.
Introduction
Klebsiella pneumoniae (K. pneumoniae) is a member of Enterobacteriaceae that is widely distributed on the skin, and digestive and respiratory tracts of both humans and animals. In nature, although K. pneumoniae is an opportunistic pathogen, it can cause pneumonia, sepsis, and inflammation of the urinary tract of human beings and has been recognized as one of the most common community-acquired pulmonary pathogen (Podschun and Ullmann, 1998; Fung et al., 2000; Sahly et al., 2000; Ko et al., 2002). In the near decades, with widespread use of antibiotics, the incidence of multi-drug resistant and hyper virulent K. pneumoniae isolates has increased frequently and globally (Shon et al., 2013; Shin and Ko, 2014; Lee et al., 2016; Zhang et al., 2017). In veterinary field, K. pneumoniae has been thought to be less harmful to animal production for a long time. However, the emergence of newly reported multi-drug resistant and hyper virulent K. pneumoniae related animal infections have increased the importance of prevention and control.
Quorum sensing is an intercellular communication mechanism that regulates certain gene expressions of most bacteria by producing extracellular signaling molecules. The regulation of QS was reported to be cell-density dependent which was first found in the marine luminous bacteria Vibrio fischeri and Vibrio harveyi. In Gram-negative bacteria, the autoinducers-1 (AI-1) have been mostly identified as derivates of N-acyl homoserine lactones (AHL) which is responsible for intraspecies communication, while autoinducers-2 (AI-2), found in both Gram-negative and Gram-positive bacteria, were reported to be in charge of interspecies communication with a common chemical component of furanosyl borate diester (Diggle et al., 2007; Antunes et al., 2010; Castillo-Juarez et al., 2015). It is generally accepted that the production of AI-2 depends on LuxS encoded by the luxS gene (Lu et al., 2017; Park et al., 2017). It has been shown that bacterial communication mediated by LuxS is also involved in biofilm formation and lipopolysaccharide synthesis (Schauder et al., 2001; De Keersmaecker et al., 2006). Previous studies have elucidated that the substrate S-ribosylhomocysteine (SAM), a methyl donor in vital cellular processes, can yield the toxic metabolic intermediate S-adenosylhomocysteine (SAH) under catalysis of S-adenosylmethionine synthetase. The S-adenosylhomocysteine nucleosidase (Pfs) rapidly hydrolyzes SAH to adenine and S-ribosylhomocysteine (SRH). Then, SRH is converted to 4,5-dihydroxy-2,3-pentanedione (DPD) and homocysteine by LuxS. However, DPD is unstable and it has been proposed that it spontaneously rearranges to form a furanosyl borate diester, namely AI-2 (Ascenso et al., 2011). Several reports have shown the involvement of AI-2 in the regulation of expression of virulence-related factors, motility, secretion systems, and regulatory proteins (Stroeher et al., 2003).
Eugenol (Figure 1) was reported to inhibit biofilm formation at early stage and reduce preformed biofilm of Streptococcus mutans without affecting bacterial viability (Adil et al., 2014). Another study showed that eugenol exhibited anti-virulence properties by competitively binding to quorum sensing receptors (Rathinam et al., 2017). Moreover, it can down-regulate the expression of S. mutans virulence genes involved in the adhesion. Eugenol was also reported to inhibit formation of biofilm las and pqs QS systems of Escherichia coli biosensors at sub-MIC level (Zhou et al., 2013). K. pneumoniae possesses pfs like (mntn) and luxS orthologs in genome and can synthesize functional AI-2 molecules, so it can be inferred that group behaviors are regulated by the AI-2-mediated quorum sensing system (Balestrino et al., 2005). Despite possessing QS inhibitor activity, there was no report of AI-2 alterations at transcription levels and molecular regulation of quorum sensing in K. pneumoniae after treated with eugenol at sub-MIC.
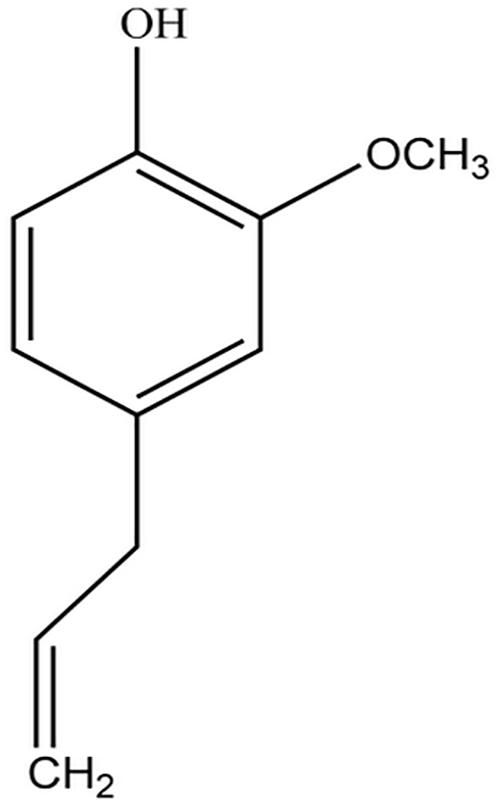
Figure 1. Chemical structure of eugenol.
Therefore, the current study analyzed transcriptional changes and interactions of AI-2 of clinical K. pneumoniae isolate treated with eugenol at exponential phase, and aimed to provide a better understanding of the QS inhibitory mechanism of eugenol.
Materials and Methods
Chemicals, Bacteria, and Culture Medium
The eugenol used in this study was obtained from Macklin Biochemical (Shanghai, China) Co., Ltd. Dimethyl sulfoxide (DMSO, Sigma-Aldrich, United States) was added with a final concentration of 1%. Triphenyltetrazolium chloride (TTC) was purchased from Biotopped Co., LTD (Beijing, China). K. pneumoniae ATCC 46117 and E. coli ATCC 25922 were kept in our laboratory. The clinical K. pneumoniae was isolated from nasal swabs and the feces sample of a porcine with respiratory disease was collected from a small pig farm in Gongzhuling City, Jilin province. Tryptone soybean broth (TSB) and Tryptone soybean agar (TSA) were obtained from Qingdao Hope Bio-Technology Co., LTD (Qingdao, China). Commercial antibiotic paper disks: AMO (Amoxicillin, 20 μg), AZM (Azithromycin, 15 μg), AZL (Azlocillin, 75 μg), AZT (Aztreonam, 30 μg), CFZ (Cefazolin, 30 μg), CHL (Chloramphenicol, 30 μg), CTX (Cefotaxime, 30 μg), DOX (Doxycycline, 30 μg), FRZ (furazolidone, 300 μg), GEN (gentamicin, 10 μg), IPM (imipenem, 10 μg), KAN (kanamycin, 30 μg), LVX (levofloxacin, 5 μg), OXA (oxacillin, 1 μg), NOR (Norfloxacin, 10 μg), POL (PolymyxinB, 300IU), STR (Streptomycin, 10 μg) were purchased from Hangzhou Microbial Reagent Co., Ltd (Hangzhou, China). S-adenosylhomocysteine (SAH) and DTNB were purchased from Sigma-Aldrich, Inc. (United States).
Drug Susceptibility and Antibacterial Test
The drug susceptibility of the clinical isolated K. pneumoniae was determined using agar disk diffusion method. Briefly, bacterial isolate was cultured in TSB at 37°C to reach a turbidity of 0.05 at 600 nm (approximately 108 cfu/mL) and was further diluted to obtain 106 cfu/mL. Sterile plates containing TSA medium were seeded with 100 μl each bacterial strain. Seventeen kinds of commercial antibiotic paper disks were placed on the surface of the inoculated plates. Incubations were carried out for 16 h at 37°C. Antibacterial susceptibility was determined by measuring the diameter of the inhibition zone (in mm) generated around the disc according to the manufacturer’s protocol. Discs of cefazolin were used as positive controls. All tests were performed in triplicate. K. pneumoniae ATCC 46117 and E. coli ATCC 25922 were employed in this study as quality control strains. For MIC determination, the eugenol was serially diluted and inoculated with an equal volume of bacterial suspension to make final concentrations of 8, 4, 2, 1, 0.5 μg/mL. 1% TTC was added as growth indicator. The tubes containing TSB only and TSB with bacteria were set as negative and positive controls, respectively. MIC value was defined as the lowest concentration of the samples that resulted in complete inhibition of visible growth after incubation. Each test was repeated triplicate at separate times.
Samples Treatment and Collection
The K. pneumoniae isolate was cultured to reach a turbidity of 0.05 at 600 nm. Then, eugenol was added into tube to reach a final concentration of sub-MIC (0.5 μg/mL) for a further culture of 1 h. In the control, an equal volume of TSB was added instead of eugenol. 2 mL bacterial culture was centrifuged at 8000g, for 5 min for RNA extraction. In each group, triplicate samples were prepared under the same experimental conditions.
RNA Extraction, Library Construction, and Transcriptome Sequencing
The library construction and sequencing were performed at Biomarker Technology (Beijing, China) using Illumina HiSeq 2500 platform. Total RNA extraction of bacterial samples was performed using a RNAiso Pure RNA Isolation Kit (TaKaRa, JP) followed by DNase I digestion, carried out according to the manufacturer’s instruction. The quality and quantity of RNA was assessed using a Thermo Scientific NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific, United States), a Qubit 2.0 fluorometer (Life Technologies, United States) and an Agilent 2100 Bioanalyzer system (Agilent Technologies, United States).
The prokaryotic rRNA was eliminated by complementary probe sequences. The rRNA-depleted RNA was randomly fragmented into small pieces by adding a fragmentation buffer and served as template for synthesis of the first strand of cDNA by using superScript II reverse transcriptase and random hexamers, then, the second strand of cDNA was synthesized by adding a buffer, dNTPs, dATP, dGTP, dCUP, RNaseH, and DNA polymerase I subsequently. The double-stranded cDNA was purified using AMPure XP beads and was subjected to end repair, adapter ligation, base A addition, adaptor ligation and agarose gel electrophoresis filtration. Suitable fragments was selected and enriched as templates by PCR amplification and used for cDNA library preparation. For library quality control, the cDNA library concentration and insert size was measured by using a Qubit 2.0 fluorometer and Agilent 2100 Bioanalyzer system, and effective concentration was qualified by qPCR.
Transcriptome analysis was done using reference genome-based reads mapping. Briefly, raw reads were pre-processed by removing adaptor, primer sequences and ambiguous reads. Additionally, low-quality reads (including reads with more than 10% unknown nucleotides and reads with the percentage of the bases with quality scores Q20 lower than 50%) were filtered before alignment. Hereafter, high quality clean reads were mapped with reference K. pneumoniae strain Kp52.145 sequence (GenBank: FO834906.1) by Bowtie2 software to obtain position and characteristic information. RNA-seq data were further analyzed and transcript expression levels were determined by calculating FPKM (Fragments Per Kilobase of transcript per Million mapped reads) by Cuffdiff section of Cufinks software.
Differential Expression Gene Analysis
The control (CK1) and eugenol treated groups (EK1) were subjected to differential gene expression analysis using Empirical Analysis of Digital Gene Expression Data (edgeSeq) R package version 3.22.3 to identify DEGs. For comparison, the P-values calibrated for multiple tests were performed according to the Benjamini and Hochberg method. False discovery rate (FDR) threshold ≤0.05 and | log2fold change (FC)| >1 were set as the criteria to determine significant DEGs (Sun et al., 2015).
GO and KEGG Enrichment Analysis
Gene Ontology (GO) enrichment analysis and KEGG (Kyoto Encyclopedia of Genes and Genome) pathway analysis were performed based on selected DEGs (Sun et al., 2017; Wang et al., 2018). Briefly, DEG was mapped to terms in GO database and the gene number of each term was counted to analyze the GO and KEGG pathway enrichment. The GO terms and pathway with corrected P-value by FDR, Q-value ≤ 0.05 are considered significantly enriched. The major biological functions that DEGs perform can be evaluated by analysis of significant GO and KEGG pathway enrichment.
qPCR
Expression of luxS, pfs(mtnn), lsrR, and lsrK genes at mRNA level was evaluated by qPCR. Bacteria was cultured to reach turbidity of 0.05 at 600 nm, eugenol was added to make a final concentration of 1/4 MIC. In the control group, TSB was added instead of eugenol. The mixture was incubated at 37°C and shaken at 150 rpm for 1 h. Total RNA was extracted using RNAiso Plus (TaKaRa) and was reversely transcribed to cDNA by PrimeScript RTase. Then, the cDNA was used as template for Real Time PCR amplification in the same reaction tube using One Step SYBR®PrimeScriptTM RT-PCR Kit (TaKaRa).
Real-time PCR process was performed on Applied Biosystems 7500 Real-Time PCR System (United States). The primers (Table 1) were designed according to a previous study and K. pneumoniae str.52.145 complete genome sequence. The amplifications were performed in 20 μl reaction mixtures containing 10 μl One Step SYBR RT-PCR Buffer Ø, 0.4 μl TaKaRa Ex Taq HS, 0.4 μl PrimeScript RT Enzyme Mix Π, 0.4 μl ROX Reference Dye Π, 6.0 μl RNase Free H2O, 0.4 μl forward and reverse primer (10 μM), respectively, and 2.0 μl template. 16S rRNA was used as reference gene (Table 1). The reacting condition was set as a two steps method as follows: 1 cycle for reverse transcription at 42°C, 5 min and 95°C for 10 s, then, 40 cycles consisting of denaturation at 95°C for 5s and annealing at 60°C for 34 s. All templates were run in triplicate.
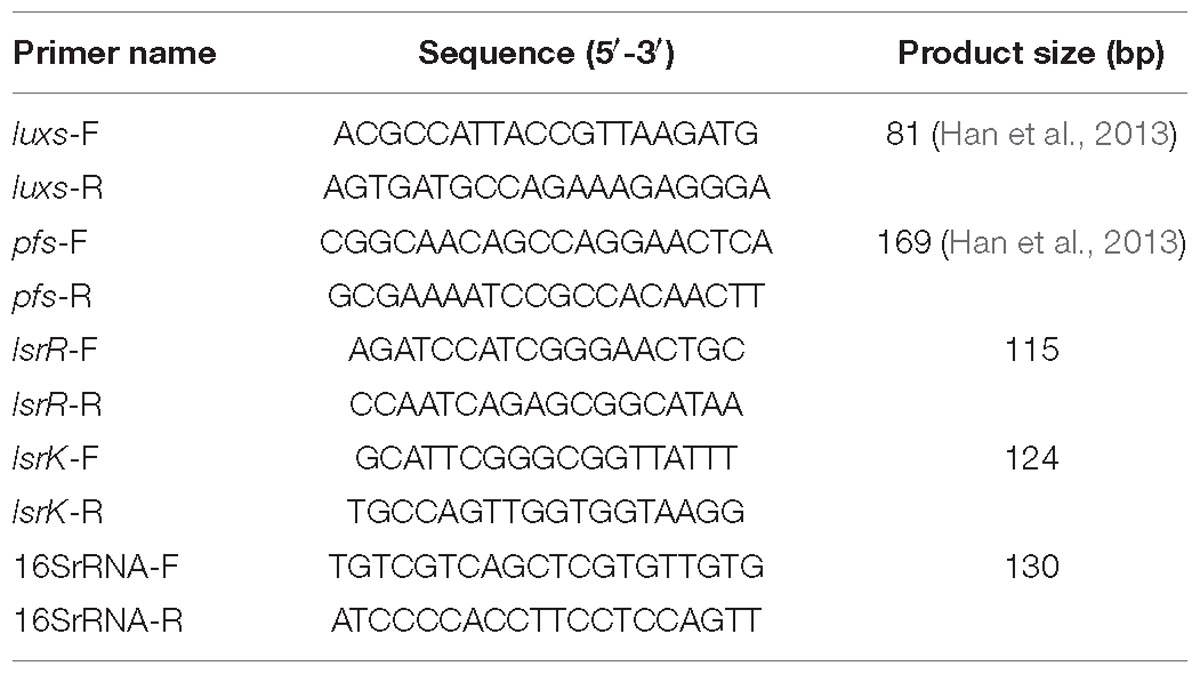
Table 1. Sequences of primers used in this study.
Enzyme Activity Inhibition Assay
In vitro synthesis of AI-2 was performed on a 96-well plate according to the method described before with little modification (Ascenso et al., 2011). Briefly, LuxS and Pfs (MtnN) were pre-prepared by prokaryotic expression previously and both were purified by AKTA pure system (GE, United States). SAH was incubated with 100 μl LuxS and Pfs to obtain an initial concentration of 1 mg/mL each in PBS (pH 7.5). Eugenol was added to the mixture with final concentrations of 0.5, 1, and 2 μg/mL, respectively. For control, equal volume of PBS was inoculated with the same experimental condition instead. The plate was incubated at 37°C for 1 h and then ultrafiltered to removing extra reaction protein. Then, 100 μl filtrate, with the major component of homocysteine, was added to 5 mmol/L equal volume Ellman’s reagent (DTNB, dissolved in 100 mmol/L PBS and 0.1 mmol/L EDTA, pH 7.2) and incubated at 37°C for 15 min. The absorbance of obtained supernatant was measured by Thermo Fisher Scientific NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific, United States) at 412 nm.
Results
Drug Susceptibility Test
As shown in Table 2, the clinical isolated porcine-derived K. pneumoniae was resistant to three out of seventeen kinds of commonly used antibiotics: amoxicillin, azithromycin, and oxacillin. The result suggested that the K. pneumoniae isolate exhibited a multi-drug resistant phenotype and is mainly resistant to beta-lactam antibiotics. The MIC of eugenol against K. pneumoniae isolate was 2 μg/ml.

Table 2. Antimicrobial resistance profile of clinical K. pneumoniae isolate.
Transcriptome Sequencing and Clustering DEGs
Raw reads data were sequenced and filtered for further analysis, yielding approximately 14.7–20.1 million clean reads. The number of high-quality clean reads in both the eugenol treated group and control group is over 98.5%. The rate of low-quality reads is lower than 0.7%. Statistical analysis revealed high total number of reads of sequencing samples and high ratio of high-quality reads. The results suggested good quality sequencing data. After aligning the high-quality clean reads with the rRNA, the reads were mapped to a reference genome. Transcriptome sequencing data were deposited to NCBI SRA database (SRA accession: PRJNA504310). In total, 5779 reference genes were detected, with a known gene number of 4514 (78.11%) in control group and 4561 (78.92%) in the eugenol treated group. Based on FDR threshold ≤0.05 and | log2 fold change (FC)| >1, 890 significant DEGs were identified. Among which, 771 genes were up-regulated while the rest 119 genes were down-regulated in control group vs. eugenol treated group (Figure 2).
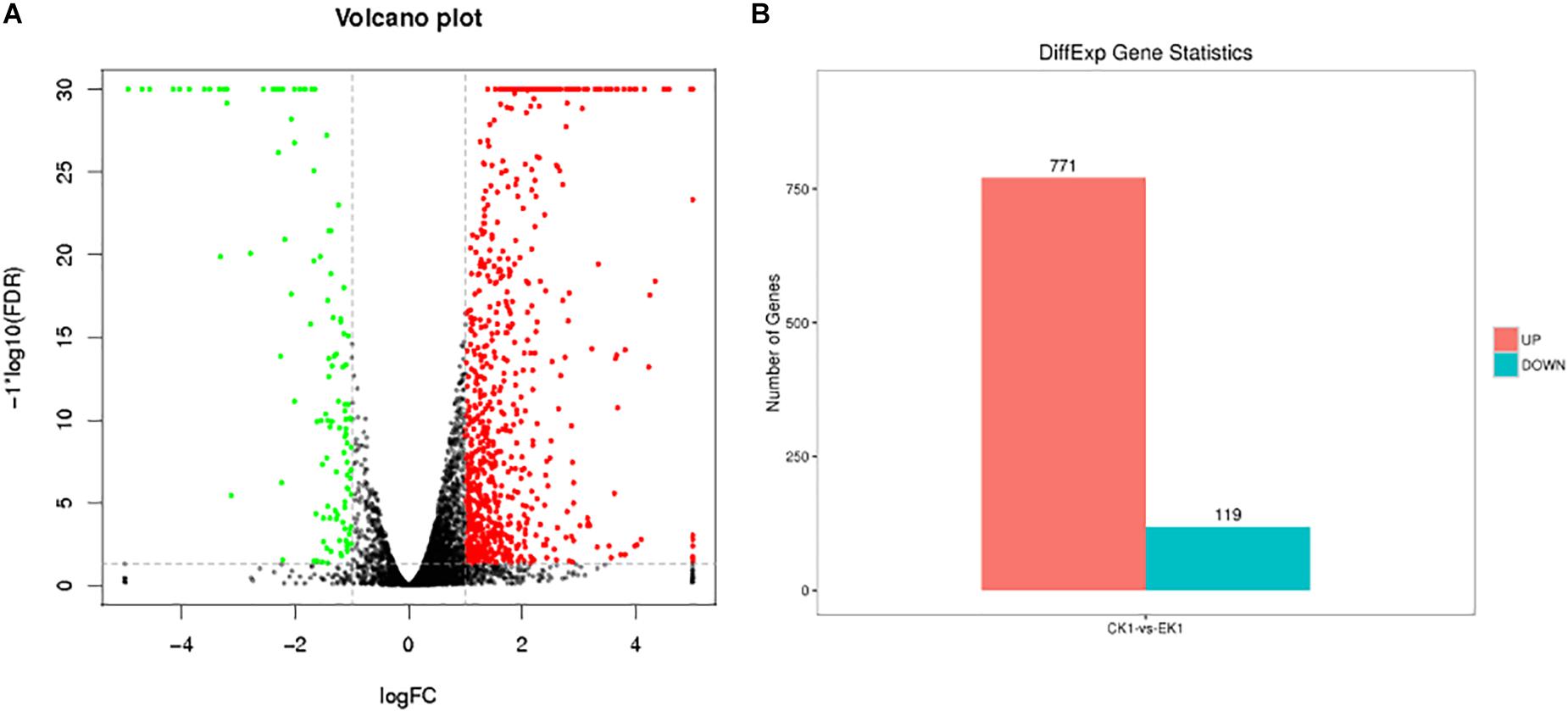
Figure 2. Volcano graph of DEGs (A) and up-regulated and down-regulated genes (B) between eugenol treated group (EK1) and control group (CK1).
Functional Annotation and Pathway Analysis of DEGs
Gene Ontology analysis can not only provide reliable gene product descriptions from various databases but also offers a set of dynamic, controlled, and structured terminologies to describe gene functions and products in organism. According to GO functions, all DEGs were classified into three categories: biological process, cellular component, and molecular function (Figure 3). There were a total of 77 terms enriched in GO terms (eugenol treated group vs. control group) among which 39 for biological process, 6 for cellular component and 32 for molecular function (Supplementary Figures S1 A–C). The most represented categories were “membrane” (GO:0016020), “intrinsic component of membrane” (GO:0031224), “membrane part” (GO:0044425), “oxidoreductase activity” (GO:0016491), “transporter activity” (GO:0005215), “transmembrane transporter activity” (GO:0022857), “tetrapyrrole binding” (GO:0046906), “establishment of localization” (GO:0051234), “localization” (GO:0051179) and “single-organism process” (GO:0044699). Among which, 146 out of 604 genes in “membrane,” 126 out of 529 genes in “intrinsic component of membrane,” 126 out of 536 genes in “membrane part,” 63 out of 228 genes in “oxidoreductase activity,” 77 out of 314 genes in “transporter activity,” 68 out of 270 genes in “transmembrane transporter activity,” 7 out of 9 genes in “tetrapyrrole binding” 157 out of 642 genes in “localization,” and 157 out of 640 genes in “establishment of localization” were up-regulated in control vs. eugenol treated groups. Results showed that gene expression of K. pneumoniae after eugenol treatment was up-regulated or down-regulated compared with untreated cells indicating molecular interactions to different gene ontology.
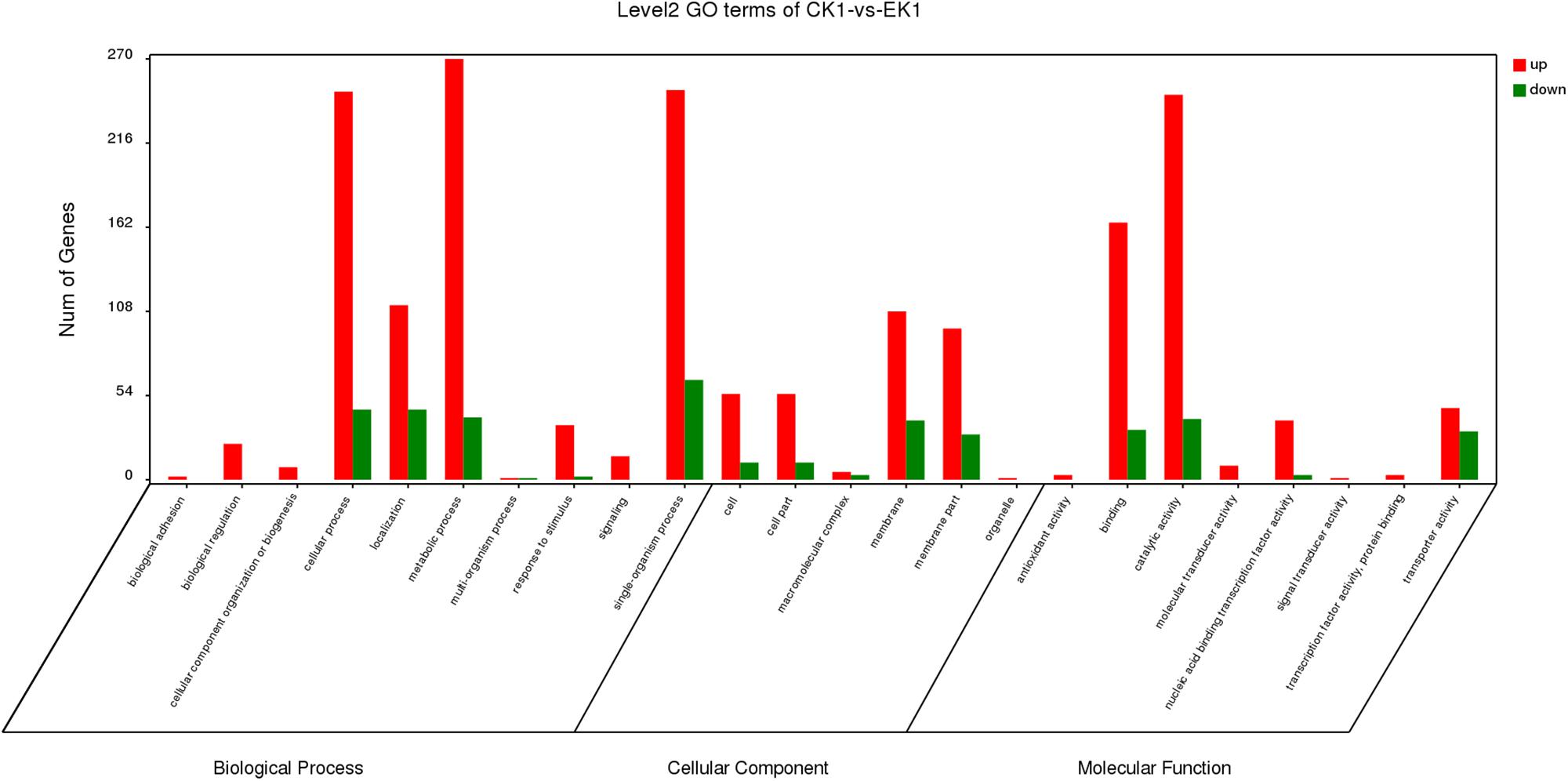
Figure 3. DEGs of biological process, cellular component and molecular function GO terms.
KEGG Pathway Analysis
The DEGs were also enriched in KEGG pathways, which could provide data on biological systems and their relationships at molecular, cellular, and organism levels. The KEGG pathways were annotated from the assembled K. pneumoniae transcriptome, and results were mapped with GO terms. As shown in Figure 4, among the top 20 of pathway enrichment, there were 60 DEGs belonging to ABC transporters and microbial metabolism in diverse environments pathways each. 40 DEGs were secondly enriched in two-component system pathway which in charge of signal transduction. Moreover, citrate cycle (TCA cycle), sulfur metabolism, ABC transporters and two-component system were the most significantly enriched pathways at both P-value and Q-value level (<0.05). The rest of DEGs, mainly enriched in metabolic pathways, were up-regulated (Supplementary Figures S2 A–C). The results indicated that eugenol affected bacterial metabolic pathways which may further inhibit AI-2 molecular production and biofilm formation.
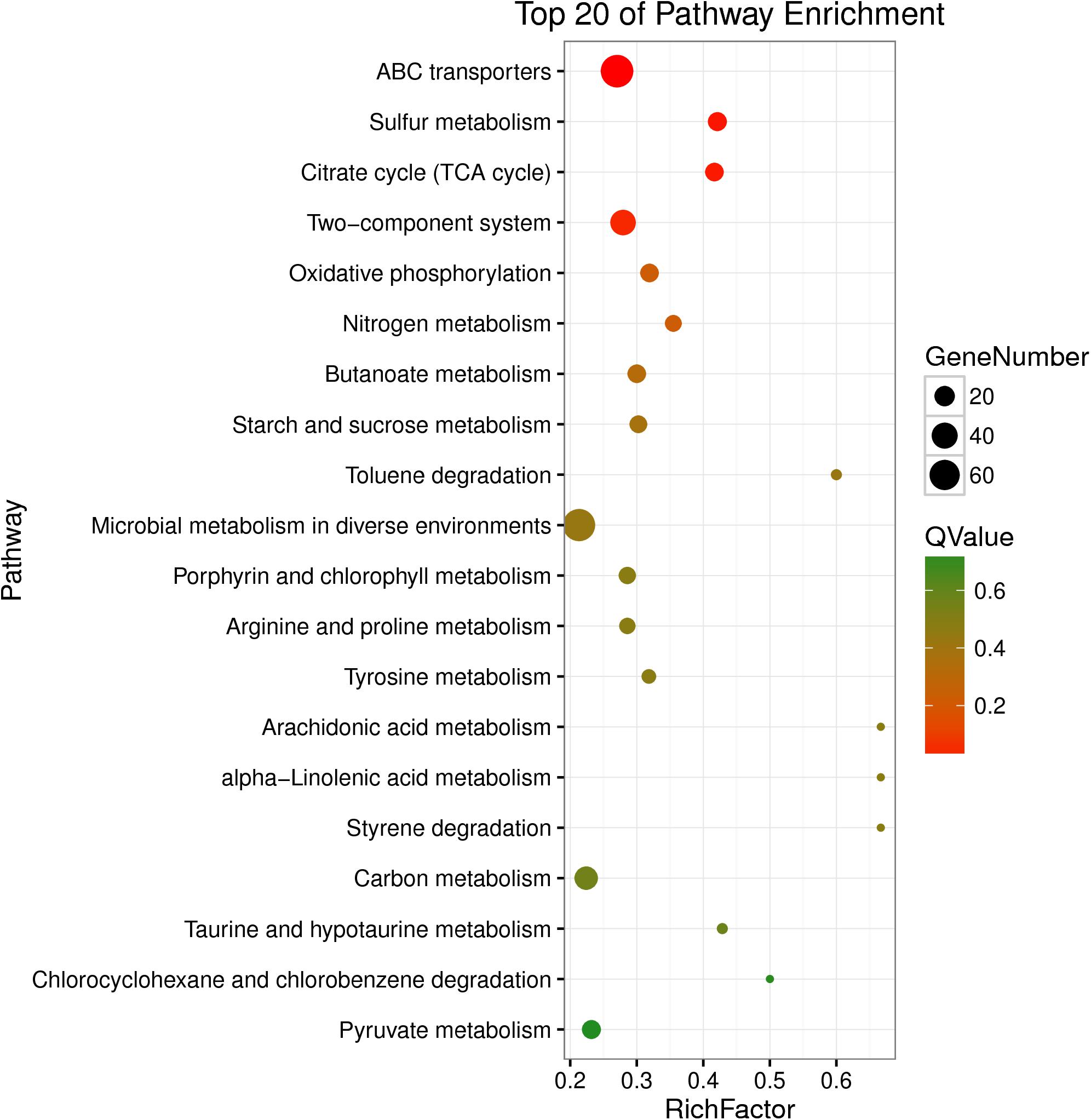
Figure 4. Statistical enrichment of DEGs in KEGG pathways.
Gene Expression Profile Analysis
qPCR analysis was performed to validate the expression profiles of DEGs in bacteria. As shown in Figure 5, genes involved in AI-2 production were selected as targets for qPCR analysis. The qPCR results of luxS, pfs(mtnn), and lsrK agreed with the fpkm dataset. In contrast, the relative expression of lsrR in eugenol added group was significantly higher than non-eugenol group despite very little difference between fpkms. Although the fold changes in K. pneumoniae slightly differed between qPCR and RNA-seq, the general expression trends matched.
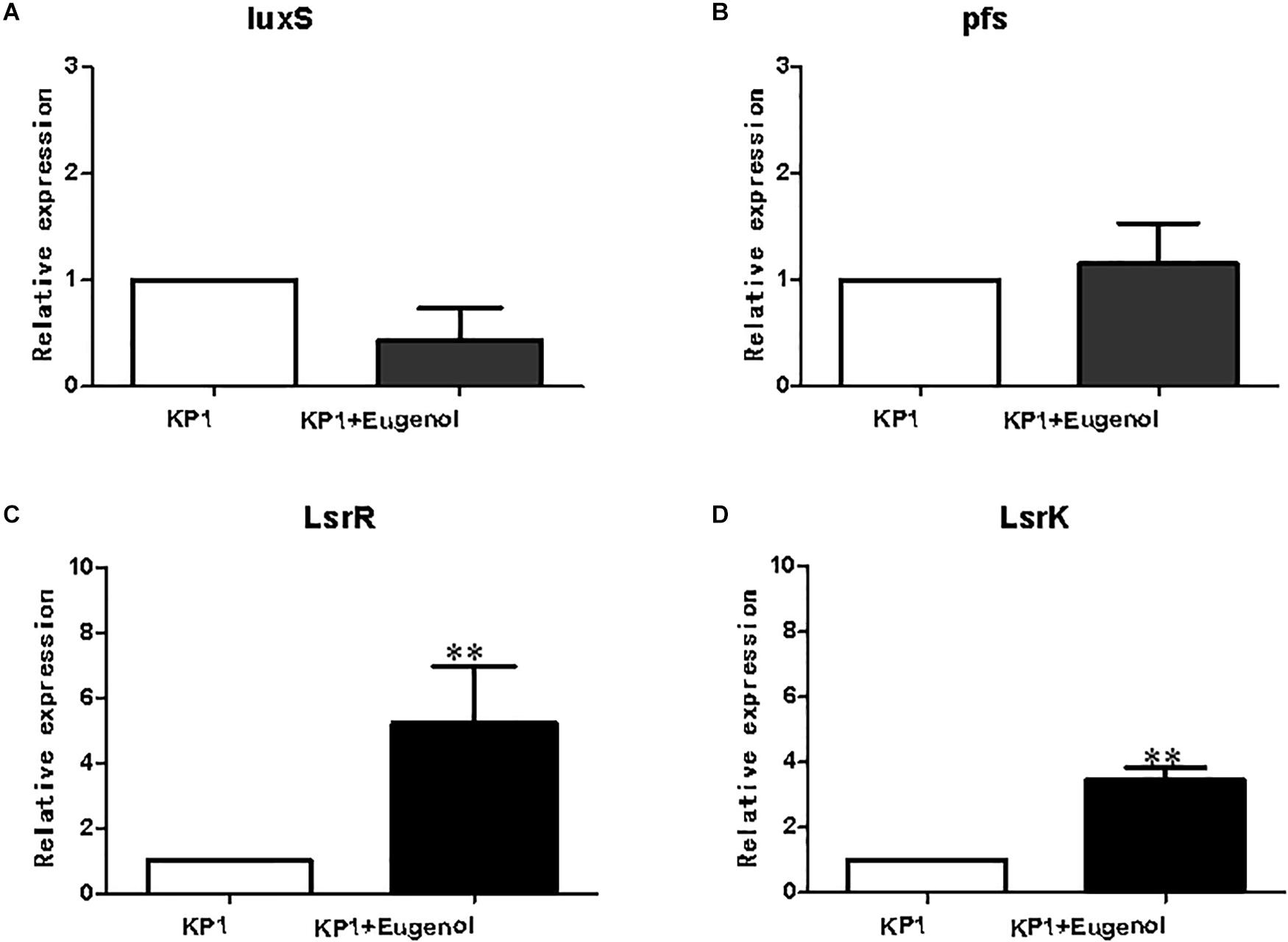
Figure 5. Relative expression of AI-2 relevant genes. luxS (A), pfs (B), lsrR (C), lsrK (D). All data were expressed as mean ± SD., n = 3. ∗∗P < 0.01 vs. control group.
AI-2 Synthesis Inhibition Test
The synthesis of AI-2 in vitro can be evaluated via measurement of homocysteine that is generated with AI-2 simultaneously. As can be seen from Figure 6, addition of eugenol at MIC could significantly (P = 0.0385) inhibit the production of AI-2 molecule compared with the control group. The absorbance of homocysteine decreased after treatment with eugenol at 1/2 MIC and 1/4 MIC, despite the non-significant difference, indicating lower AI-2 levels. The above result clearly demonstrated that eugenol could interact with LuxS and Pfs (MtnN) and thus interfere the generation of AI-2. The effect presented a dose-dependent manner. Furthermore, the result was in accordance with transcriptome sequencing and qPCR analysis, which further indicated the inhibiting effect of eugenol on K. pneumoniae AI-2 molecule and quorum sensing system.
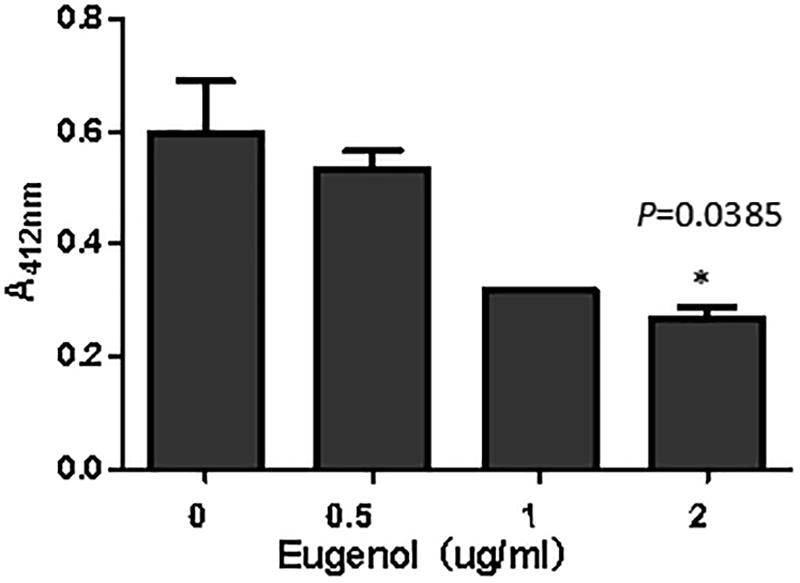
Figure 6. Absorption of homocysteine incubated with different concentrations eugenol. ∗P < 0.05 vs. control group.
Discussion
Animal-derived multi-drug resistant K. pneumoniae has been reported more frequently in recent years. The emergence of animal infection caused by K. pneumoniae not only results in economic loss of animal husbandry, but will pose a great challenge to human public health. However, due to the wide use of antibiotics, fast occurring bacterial drug-resistance has made most conventional antibiotics invalid, which make the situation more severe (Zou et al., 2011; Overdevest et al., 2014; Vuotto et al., 2014, 2017; Anes et al., 2017). According to drug-resistance result, the isolated porcine-derived K. pneumoniae was multi-drug resistant to β-lactamase antibiotics, which was commonly found in both human and animal clinical trials. As an effective herbal monomer, eugenol was proven to possess QS inhibitor activity. However, little is known about how K. pneumoniae AI-2 responses to eugenol at the transcriptional level. The present study revealed that eugenol could decrease AI-2 signal molecules mainly by down-regulating luxS gene expression, up-regulating negative feedback gene lsrR, and inhibit catalytic activity of LuxS.
Quorum sensing consists of an enzyme that catalyzes the synthesis of the signal molecules and receptors that bind to the signal and reprograms the expression of several genes. Previous study has investigated the theory that AI-1 was not produced by K. pneumoniae due to the AI-1 type QS quenching enzymes, AHL lactonases (Park et al., 2003; Sun et al., 2016). In contrast, class II autoinducer, AI-2 could be detected in culture supernatant and was LuxS dependent (Wang et al., 2005; Ascenso et al., 2011; Zhu et al., 2011). Moreover, it is well accepted that QS can affect biofilm formation and virulent gene expression by regulating LPS synthesis and virulent genes expression, both of which are closely related to drug-resistance (De Araujo et al., 2010; Defoirdt, 2018). Therefore, on the basis of the above study, the transcriptional analysis was performed and validated by qPCR. Interactions between eugenol and AI-2 were confirmed by AI-2 inhibition test.
Concerning the results of transcriptome sequencing, the total number of reads and ratio of high quality was high, and the error rate was low, suggesting good quality sequencing data. The correlation between control and eugenol treated groups was assessed using Pearson’s correlation coefficient (Figure 7). The paired Pearson’s correlation co-efficient within groups were all higher than 0.97, demonstrating good sample correlations. The heat maps (Figure 8) of the induced and suppressed transcripts of eugenol-exposed samples matched to controls in the opposite way. This result demonstrated that exposure to eugenol involved a series of gene ontologies mainly in biological processes, cellular component and molecular function, and could consequently result in the up-regulating most genes. Moreover, the result also explained the eugenol mode of action on bacteria at sub-MIC.
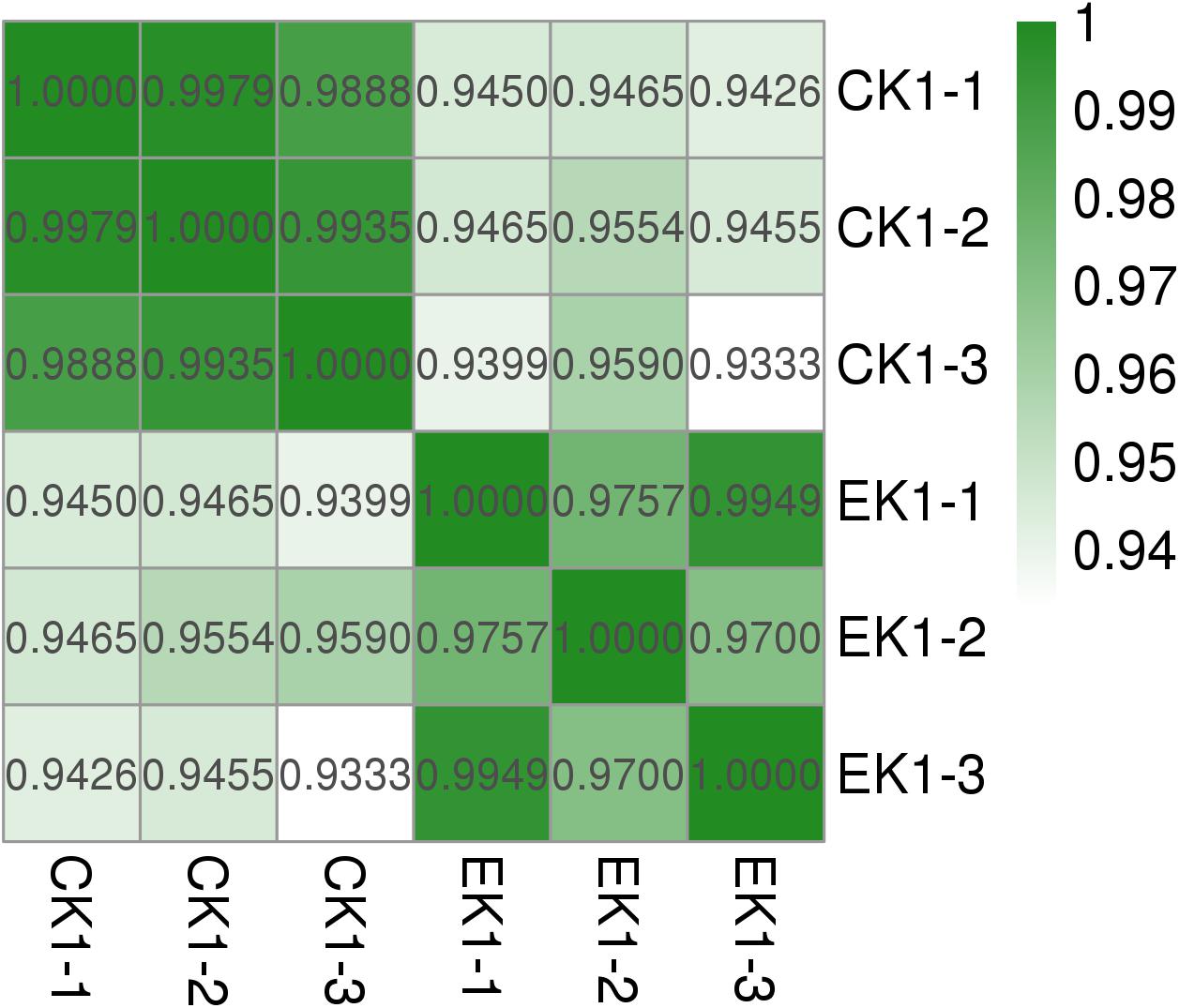
Figure 7. Pearson’s correlation coefficient and volcano graph of eugenol treated group (EK1) and control group (CK1).
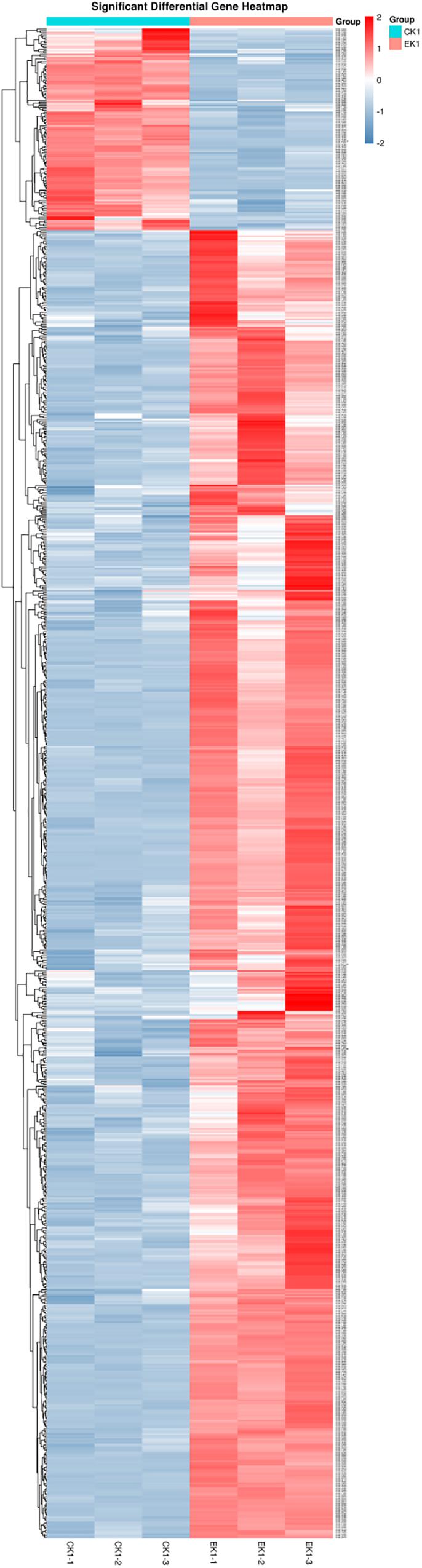
Figure 8. Heatmap expression pattern clustering analysis of eugenol treated group (EK1) and control group (CK1).
lsrK and lsrR are positive and negative regulating genes of luxS, respectively. lsrK can positively regulate phospho-AI-2 production and negatively regulate lsrR. lsrR, inversely, not only negatively regulate subsequent gene cluster from lsrA to lsrG, but lsrK and itself (Taga et al., 2003; Torres-Escobar et al., 2013; Adams et al., 2014). In the study, the fpkms of luxS and lsrR were a little lower in the eugenol treated group vs. control, while mtnN and lsrk were higher. For luxS and pfs, it could be explained that although expression of luxS and pfs was coincidence at the initial culture period, the speed and level really differed when bacterial growth reach logarithmic phase, which has been proven in previous research. The results of qPCR were consistent with luxS, pfs(mtnN), and lsrK except for lsrR, a transcriptional repressor of lsr operon. In the eugenol treated group, the relative expression of lsrR was significantly higher than that of the control. Concerning the non-significant fpkm data between control and eugenol-treated group, we speculate that the high level of lsrR does repress the expression of the following genes responsible for AI-2 signal transmission, which will reduce the level of phospho-AI-2 in vivo. The multidrug resistance related proteins (MRPs) function as efflux transporters of a variety of large organic anions or their conjugates. Eugenol was reported to decrease MRP2 levels more effectively than Piper betel leaves extract with less of a sensitizing effect and therefore has the potential to serve as an additive to reduce drug resistance (Young et al., 2006). ATP-binding cassette (ABC) transporters are membrane-bound transporters involved in various physiological processes. Previous study found that ABC transporter encoding genes MgAtr1, MgAtr2, MgAtr4, and MgAtr5 from the wheat pathogen Mycosphaerella graminicola could be upregulated. Although the mode of action of eugenol is unclear, there is some evidence that it might act as an uncoupler of oxidative phosphorylation and thus can inhibit ABC transporter activity by depleting ATP. Moreover, these findings also support the hypothesis that the encoded ABC transporters provide protection against toxic compounds present in the environment of the fungus (Zwiers and De Waard, 2000; Stergiopoulos et al., 2002).
In vitro synthesis of AI-2 has been applied to detect and qualify the AI-2 of Avian Pathogenic E. coli before (Han et al., 2013). In this study, the addition of eugenol at MIC significantly inhibited the generation of AI-2 molecule, and 1/2 MIC and 1/4 MIC also exhibited inhibiting effects. The above results clearly demonstrated that eugenol could interact with luxS and mtnN and thus interfere with the generation of AI-2, and the effect was dose-dependent. Combined with transcriptome sequencing and qPCR analysis, we think sub-MIC eugenol exerted AI-2 inhibitory activity mainly depends on luxS and AI-2 synthesis regulator gene affiliated to biological process and molecular function ontology.
Conclusion
In conclusion, the study revealed the transcriptional profile of clinical isolated porcine-derived K. pneumoniae treated with sub-MIC eugenol. The DEGs involved in AI-2 molecules were identified. Eugenol could reduce AI-2 molecule generation largely by lowering AI-2 synthesis gene category e.g., luxS and lsrK, increasing the expression of repressive gene lsrR, and decelerating LuxS catalytic activity. The research provides a basis for elucidating the mechanism of interactions between eugenol and K. pneumoniae.
Author Contributions
Y-MW contributed to drafting and revising the manuscript. Y-MW, L-CK, and W-LD participated in the design of the study and data analysis. HX-M participated in experiment design and article revise, and provided constructive advice to the study. Y-MW and KO performed the antimicrobial susceptibility test, qPCR, and enzyme activity inhibition assay. All authors read and approved the final manuscript.
Funding
This work was supported by the National Key Research and Development Plan (Grant No: 2016YFD0501301), National Natural Science Foundation of China (Grant No: 31702293), Science and Technology Development Plan of Jilin Province (Grant No: 20170520074JH), and Natural Science Foundation of Jilin Province (Grant No: 20180101003JC).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the Jilin Provincial Key Laboratory of New Veterinary Drug Innovation Research and Development for support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01159/full#supplementary-material
FIGURE S1 | GO enrichment analysis of P-value heat map (A), Q-value heat map (B), and enrichment factor heat map (C).
FIGURE S2 | KEGG enrichment analysis of P-value heat map (A), Q-value heat map (B), and enrichment factor heat map (C).
References
Adams, B. L., Carter, K. K., Guo, M., Wu, H. C., Tsao, C. Y., Sintim, H. O., et al. (2014). Evolved quorum sensing regulator, LsrR, for altered switching functions. ACS Synth. Biol. 3, 210–219. doi: 10.1021/sb400068z
Adil, M., Singh, K., Verma, P. K., and Khan, A. U. (2014). Eugenol-induced suppression of biofilm-forming genes in Streptococcus mutans: an approach to inhibit biofilms. J. Glob. Antimicrob. Resist. 2, 286–292. doi: 10.1016/j.jgar.2014.05.006
Anes, J., Hurley, D., Martins, M., and Fanning, S. (2017). Exploring the genome and phenotype of multi-drug resistant Klebsiella pneumoniae of clinical origin. Front. Microbiol. 8:1913. doi: 10.3389/fmicb.2017.01913
Antunes, L. C., Ferreira, R. B., Buckner, M. M., and Finlay, B. B. (2010). Quorum sensing in bacterial virulence. Microbiology 156, 2271–2282. doi: 10.1099/mic.0.038794-0
Ascenso, O. S., Marques, J. C., Santos, A. R., Xavier, K. B., Ventura, M. R., and Maycock, C. D. (2011). An efficient synthesis of the precursor of AI-2, the signalling molecule for inter-species quorum sensing. Bioorg. Med. Chem. 19, 1236–1241. doi: 10.1016/j.bmc.2010.12.036
Balestrino, D., Haagensen, J. A., Rich, C., and Forestier, C. (2005). Characterization of type 2 quorum sensing in Klebsiella pneumoniae and relationship with biofilm formation. J. Bacteriol. 187, 2870–2880. doi: 10.1128/jb.187.8.2870-2880.2005
Castillo-Juarez, I., Maeda, T., Mandujano-Tinoco, E. A., Tomas, M., Perez-Eretza, B., Garcia-Contreras, S. J., et al. (2015). Role of quorum sensing in bacterial infections. World J. Clin. Cases 3, 575–598. doi: 10.12998/wjcc.v3.i7.575
De Araujo, C., Balestrino, D., Roth, L., Charbonnel, N., and Forestier, C. (2010). Quorum sensing affects biofilm formation through lipopolysaccharide synthesis in Klebsiella pneumoniae. Res. Microbiol. 161, 595–603. doi: 10.1016/j.resmic.2010.05.014
De Keersmaecker, S. C., Sonck, K., and Vanderleyden, J. (2006). Let LuxS speak up in AI-2 signaling. Trends Microbiol. 14, 114–119. doi: 10.1016/j.tim.2006.01.003
Defoirdt, T. (2018). Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 26, 313–328. doi: 10.1016/j.tim.2017.10.005
Diggle, S. P., Griffin, A. S., Campbell, G. S., and West, S. A. (2007). Cooperation and conflict in quorum-sensing bacterial populations. Nature 450, 411–414. doi: 10.1038/nature06279
Fung, C. P., Hu, B. S., Chang, F. Y., Lee, S. C., Kuo, B. I., Ho, M., et al. (2000). A 5-year study of the seroepidemiology of Klebsiella pneumoniae: high prevalence of capsular serotype K1 in Taiwan and implication for vaccine efficacy. J. Infect. Dis. 181, 2075–2079. doi: 10.1086/315488
Han, X., Bai, H., Liu, L., Dong, H., Liu, R., Song, J., et al. (2013). The luxS gene functions in the pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 55, 21–27. doi: 10.1016/j.micpath.2012.09.008
Ko, W. C., Paterson, D. L., Sagnimeni, A. J., Hansen, D. S., Von Gottberg, A., Mohapatra, S., et al. (2002). Community-acquired Klebsiella pneumoniae bacteremia: global differences in clinical patterns. Emerg. Infect. Dis. 8, 160–166. doi: 10.3201/eid0802.010025
Lee, C. R., Lee, J. H., Park, K. S., Kim, Y. B., Jeong, B. C., and Lee, S. H. (2016). Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front. Microbiol. 7:895. doi: 10.3389/fmicb.2016.00895
Lu, S. Y., Zhao, Z., Avillan, J. J., Liu, J., and Call, D. R. (2017). Autoinducer-2 quorum sensing contributes to regulation of microcin PDI in Escherichia coli. Front. Microbiol. 8:2570. doi: 10.3389/fmicb.2017.02570
Overdevest, I. T., Heck, M., Van Der Zwaluw, K., Huijsdens, X., Van Santen, M., Rijnsburger, M., et al. (2014). Extended-spectrum beta-lactamase producing Klebsiella spp. in chicken meat and humans: a comparison of typing methods. Clin. Microbiol. Infect. 20, 251–255. doi: 10.1111/1469-0691.12277
Park, H., Lee, K., Yeo, S., Shin, H., and Holzapfel, W. H. (2017). Autoinducer-2 quorum sensing influences viability of Escherichia coli O157:H7 under osmotic and in vitro gastrointestinal stress conditions. Front. Microbiol. 8:1077. doi: 10.3389/fmicb.2017.01077
Park, S. Y., Lee, S. J., Oh, T. K., Oh, J. W., Koo, B. T., Yum, D. Y., et al. (2003). AhlD, an N-acylhomoserine lactonase in Arthrobacter sp., and predicted homologues in other bacteria. Microbiology 149, 1541–1550. doi: 10.1099/mic.0.26269-0
Podschun, R., and Ullmann, U. (1998). Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 11, 589–603. doi: 10.1128/cmr.11.4.589
Rathinam, P., Vijay Kumar, H. S., and Viswanathan, P. (2017). Eugenol exhibits anti-virulence properties by competitively binding to quorum sensing receptors. Biofouling 33, 624–639. doi: 10.1080/08927014.2017.1350655
Sahly, H., Podschun, R., and Ullmann, U. (2000). Klebsiella infections in the immunocompromised host. Adv. Exp. Med. Biol. 479, 237–249. doi: 10.1007/0-306-46831-x_21
Schauder, S., Shokat, K., Surette, M. G., and Bassler, B. L. (2001). The LuxS family of bacterial autoinducers: biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 41, 463–476. doi: 10.1046/j.1365-2958.2001.02532.x
Shin, J., and Ko, K. S. (2014). Comparative study of genotype and virulence in CTX-M-producing and non-extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 58, 2463–2467. doi: 10.1128/AAC.02499-13
Shon, A. S., Bajwa, R. P., and Russo, T. A. (2013). Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: a new and dangerous breed. Virulence 4, 107–118. doi: 10.4161/viru.22718
Stergiopoulos, I., Gielkens, M. M., Goodall, S. D., Venema, K., and De Waard, M. A. (2002). Molecular cloning and characterisation of three new ATP-binding cassette transporter genes from the wheat pathogen Mycosphaerella graminicola. Gene 289, 141–149. doi: 10.1016/s0378-1119(02)00505-x
Stroeher, U. H., Paton, A. W., Ogunniyi, A. D., and Paton, J. C. (2003). Mutation of luxS of Streptococcus pneumoniae affects virulence in a mouse model. Infect. Immun. 71, 3206–3212. doi: 10.1128/iai.71.6.3206-3212.2003
Sun, D., Zhang, W., Mou, Z., Chen, Y., Guo, F., Yang, E., et al. (2017). Transcriptome analysis reveals silver nanoparticle-decorated quercetin antibacterial molecular mechanism. ACS Appl. Mater. Interfaces 9, 10047–10060. doi: 10.1021/acsami.7b02380
Sun, Q. L., Zhao, C. P., Wang, T. Y., Hao, X. B., Wang, X. Y., Zhang, X., et al. (2015). Expression profile analysis of long non-coding RNA associated with vincristine resistance in colon cancer cells by next-generation sequencing. Gene 572, 79–86. doi: 10.1016/j.gene.2015.06.087
Sun, S., Zhang, H., Lu, S., Lai, C., Liu, H., and Zhu, H. (2016). The metabolic flux regulation of Klebsiella pneumoniae based on quorum sensing system. Sci. Rep. 6:38725. doi: 10.1038/srep38725
Taga, M. E., Miller, S. T., and Bassler, B. L. (2003). Lsr-mediated transport and processing of AI-2 in Salmonella typhimurium. Mol. Microbiol. 50, 1411–1427. doi: 10.1046/j.1365-2958.2003.03781.x
Torres-Escobar, A., Juarez-Rodriguez, M. D., Lamont, R. J., and Demuth, D. R. (2013). Transcriptional regulation of Aggregatibacter actinomycetemcomitans lsrACDBFG and lsrRK operons and their role in biofilm formation. J. Bacteriol. 195, 56–65. doi: 10.1128/JB.01476-12
Vuotto, C., Longo, F., Balice, M. P., Donelli, G., and Varaldo, P. E. (2014). Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 3, 743–758. doi: 10.3390/pathogens3030743
Vuotto, C., Longo, F., Pascolini, C., Donelli, G., Balice, M. P., Libori, M. F., et al. (2017). Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J. Appl. Microbiol. 123, 1003–1018. doi: 10.1111/jam.13533
Wang, L., Li, J., March, J. C., Valdes, J. J., and Bentley, W. E. (2005). luxS-dependent gene regulation in Escherichia coli K-12 revealed by genomic expression profiling. J. Bacteriol. 187, 8350–8360. doi: 10.1128/jb.187.24.8350-8360.2005
Wang, S., Zheng, Y., Gu, C., He, C., Yang, M., Zhang, X., et al. (2018). Bacillus cereus AR156 activates defense responses to Pseudomonas syringae pv. Tomato in Arabidopsis thaliana Similarly to flg22. Mol. Plant Microbe Interact. 31, 311–322. doi: 10.1094/MPMI-10-17-0240-R
Young, S. C., Wang, C. J., Hsu, J. D., Hsu, J. L., and Chou, F. P. (2006). Increased sensitivity of Hep G2 cells toward the cytotoxicity of cisplatin by the treatment of piper betel leaf extract. Arch. Toxicol. 80, 319–327. doi: 10.1007/s00204-005-0051-3
Zhang, L., Li, Y., Shen, W., Wang, S. M., Wang, G., and Zhou, Y. (2017). Whole-genome sequence of a carbapenem-resistant hypermucoviscous Klebsiella pneumoniae isolate SWU01 with capsular serotype K47 belonging to ST11 from a patient in China. J. Glob. Antimicrob. Resist. 11, 87–89. doi: 10.1016/j.jgar.2017.09.001
Zhou, L., Zheng, H., Tang, Y., Yu, W., and Gong, Q. (2013). Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 35, 631–637. doi: 10.1007/s10529-012-1126-x
Zhu, H., Liu, H. J., Ning, S. J., and Gao, Y. L. (2011). A luxS-dependent transcript profile of cell-to-cell communication in Klebsiella pneumoniae. Mol. Biosyst. 7, 3164–3168. doi: 10.1039/c1mb05314k
Zou, L. K., Wang, H. N., Zeng, B., Zhang, A. Y., Li, J. N., Li, X. T., et al. (2011). Phenotypic and genotypic characterization of beta-lactam resistance in Klebsiella pneumoniae isolated from swine. Vet. Microbiol. 149, 139–146. doi: 10.1016/j.vetmic.2010.09.030
Keywords: eugenol, K. pneumoniae, quorum sensing, transcriptome analysis, AI-2 synthesis, inhibitory effect
Citation: Wang Y-m, Dong W-l, Odah KA, Kong L-c and Ma H-x (2019) Transcriptome Analysis Reveals AI-2 Relevant Genes of Multi-Drug Resistant Klebsiella pneumoniae in Response to Eugenol at Sub-MIC. Front. Microbiol. 10:1159. doi: 10.3389/fmicb.2019.01159
Received: 07 November 2018; Accepted: 07 May 2019;
Published: 28 May 2019.
Edited by:
Hua-Bin Li, Sun Yat-sen University, ChinaReviewed by:
Jianfeng Wang, Jilin University, ChinaTommaso Felicetti, Università Degli Studi Di Perugia, Italy
Copyright © 2019 Wang, Dong, Odah, Kong and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ling-cong Kong, Y29uZ3dic0AxMjYuY29t; Hong-xia Ma, aG9uZ3hpYTA3MzEwMDFAMTYzLmNvbQ==