 Friedrich Kirsch1
Friedrich Kirsch1 Stephan Klähn
Stephan Klähn Martin Hagemann
Martin Hagemann- 1Department of Plant Physiology, Institute for Biosciences, University of Rostock, Rostock, Germany
- 2Department of Solar Materials, Helmholtz-Centre for Environmental Research–UFZ, Leipzig, Germany
Cyanobacteria are prokaryotes that can assimilate inorganic carbon via oxygenic photosynthesis, which results in the formation of organic compounds essentially from CO2, water, and light. Increasing concerns regarding the increase in atmospheric CO2 due to fossil energy usage fueled the idea of a photosynthesis-driven and CO2-neutral, i.e., cyanobacteria-based biotechnology. The ability of various cyanobacteria to tolerate high and/or fluctuating salinities attenuates the requirement of freshwater for their cultivation, which makes these organisms even more interesting regarding a sustainable utilization of natural resources. However, those applications require a detailed knowledge of the processes involved in salt acclimation. Here, we review the current state of our knowledge on the regulation of compatible solute accumulation in cyanobacteria. The model organism Synechocystis sp. PCC 6803 responds to increasing salinities mainly by the accumulation of glucosylglycerol (GG) and sucrose. After exposure toward increased salt concentrations, the accumulation of the main compatible solute GG is achieved by de novo synthesis. The key target of regulation is the enzyme GG-phosphate synthase (GgpS) and involves transcriptional, posttranscriptional, and biochemical mechanisms. Recently, the GG-degrading enzyme GG hydrolase A (GghA) was identified, which is particularly important for GG degradation during exposure to decreasing salinities. The inversely ion-regulated activities of GgpS and GghA could represent the main model for effectively tuning GG steady state levels according to external salinities. Similar to GG, the intracellular amount of sucrose is also salt-regulated and seems to be determined by the balance of sucrose synthesis via sucrose-phosphate synthase (Sps) and its degradation via invertase (Inv). In addition to their role as stress protectants, both compatible solutes also represent promising targets for biotechnology. Hence, the increasing knowledge on the regulation of compatible solute accumulation not only improves our understanding of the stress physiology of cyanobacteria but will also support their future biotechnological applications.
Introduction
Cyanobacteria are the only prokaryotes performing oxygenic photosynthesis, a process which uses light energy to assimilate CO2 into carbohydrates and biomass, thereby releasing oxygen as byproduct from water splitting. It is commonly accepted that oxygenic photosynthesis has evolved within the group of cyanobacteria. The evolution of cyanobacteria initiated the rise of free oxygen in the ancient atmosphere, which is known as the great oxygenation event (Hohmann-Marriott and Blankenship, 2011; Lyons et al., 2014; Soo et al., 2017). Accordingly, ancient cyanobacteria had a huge impact on Earth’s biogeochemical history not only by producing the oxygen-containing atmosphere and provoking the evolution of aerobic life, but also by providing organic matter in the global food web. Moreover, an early representative of cyanobacteria served as an endosymbiont within a eukaryotic cell, subsequently establishing the chloroplast as photosynthetic organelle in eukaryotic algae and plants (Martin et al., 2015). During their long evolution, cyanobacteria developed a large variability in metabolism, morphology, and tolerance to abiotic stresses (Tandeau de Marsac and Houmard, 1993). These abilities enable cyanobacteria to colonize almost all aquatic and terrestrial habitats, as long as minimal amounts of water, light, and some trace minerals are present. To date, cyanobacteria still play an important role in global biogeochemical cycles. For example, marine picocyanobacteria of the genera Prochlorococcus and Synechococcus account for approximately 25% of primary production in the oceans (Flombaum et al., 2013). In addition to inorganic carbon, the fixation of atmospheric nitrogen by several cyanobacterial strains is a major source of reduced inorganic nitrogen compounds in marine habitats (Foster et al., 2011; Zehr and Kudela, 2011).
Due to their photosynthetic lifestyle, there is a rising interest for cyanobacteria in green biotechnology (for reviews, see Ducat et al., 2011; Savakis and Hellingwerf, 2015; Hagemann and Hess, 2018). For instance, cyanobacterial cells can be used as sustainable producers of various chemical compounds or fuel components. The interest in biofuels is mainly driven by the increasing global energy demand, which in turn forces mankind to find alternatives for the limited fossil energy reserves and the high CO2 emission during their utilization.
The advantage of using cyanobacteria is essentially based on three basic ideas: first, phototrophic microorganisms do not require energy to build differentiated organs, such as roots, leaves, and flowers, which promise greater efficiency in the conversion of light energy into the desired product (Mata et al., 2010). Secondly, their cultivation does not depend on agricultural land–there is no competition with the cultivation of crops for food and feed production (Ahmad et al., 2011). And thirdly, many of these organisms tolerate enhanced salinities; hence, their cultivation does not necessarily rely on limited freshwater availability (Chisti, 2013; Pade and Hagemann, 2015). In recent years, great efforts have been made to produce a wide variety of products using cyanobacteria as hosts. These include ethanol and biodiesel, fatty acids, isobutanol, isoprene, and many more (reviewed in Case and Atsumi, 2016; Singh et al., 2017). Relatively high production titers have been reported, for example, for cyanobacterial ethanol production with rates of 230 up to 270 mg L−1 d−1 (Gao et al., 2012; Dienst et al., 2014). However, albeit a variety of different approaches have been attempted; no economically competitive biotechnological production of bioenergy and chemical feedstock has yet been established with cyanobacteria. Therefore, further research is required which tackles, for instance, the design of cost-effective bioreactors. In contrast to large-scale industrial fermenters for heterotrophic bacteria, appropriate light delivery represents one of the major challenges for cyanobacterial mass cultivation. This is due to the fact that shading effects occur proportional with cell growth, which in turn limit biomass and often product formation (for a review on photobiorectors, see Posten, 2009). Nevertheless, significant progress has been made during the last two decades, for instance by using flat panel reactors in a pilot scale (e.g., Subitec GmbH, https://subitec.com/en). Furthermore, the application of new methods in the field of synthetic biology might enable more efficient delivery of carbon into the respective end product in the future (Hagemann and Hess, 2018). In addition, the stress tolerance of the used cyanobacterial strains comes to the fore considering the indispensable effects of fluctuating temperatures and changing light conditions when exposed to natural sunlight (Singh et al., 2017). This also counts for the anticipated cultivation in salty media, i.e., naturally available brackish or seawater. In this context, the tolerance of the producer strains to salt is one of the most important challenges.
Salt Stress and the Accumulation of Compatible Solutes
Salinity is one of the major abiotic environmental factors in aquatic but also terrestrial habitats. Fluctuating salt concentrations essentially confront organisms with two problems. First, high amounts of dissolved ions decrease the osmotic potential in the surrounding medium, which has a direct influence on the water content within the cell. An increasing salinity is subsequently accompanied by an osmotic loss of water, which is conflicting the maintenance of turgor pressure as a driving force for cellular growth (Ladas and Papageorgiou, 2000). Second, especially during sudden increases of the external salinity (salt shock), which, e.g., naturally occur in river estuaries, large amounts of inorganic ions penetrate into the cell, thereby disturbing intracellular ion homeostasis. This is problematic because many cellular processes are directly or indirectly dependent on various inorganic ions such as potassium (K+), calcium (Ca2+), and magnesium (Mg2+). However, sodium (Na+) and chloride (Cl−) have the largest percentage of dissolved salt ions in natural saline habitats (Pawlowicz, 2013). Thus, salt stress can lead to a drastic change in the intracellular ion composition. In addition, high intracellular Na+ concentrations are directly toxic due to negative effects on the hydration shell and the surface charge of macromolecules (Klähn and Hagemann, 2011).
To accommodate increasing salinity, most microorganisms including cyanobacteria use the salt-out strategy, which involves the active export of Na+ and Cl− as well as the intracellular accumulation of osmoprotective organic compounds (Hagemann, 2011). The former counteracts the influx of toxic ions in the cell and serves to maintain ion homeostasis. The accumulation of osmoprotective compounds, the so-called compatible solutes, serves to balance the osmotic potential between the cell interior and exterior. Compatible solutes are low-molecular mass substances that are readily soluble in water and can be accumulated in high concentrations within the cell without interfering with metabolism and other cellular processes (Brown, 1976). By accumulating these substances, it is possible to compensate for changes in the osmotic potential and thus ensuring the uptake of water to maintain turgor and cellular growth. Structurally, compatible solutes can be assigned to different substance classes (Figure 1); these are often sugars (trehalose, sucrose), polyols (glycerol, sorbitol), heterosides (glucosylglycerol, floridoside; Hagemann and Pade, 2015) or amino acids (glutamate, proline), and amino acid derivatives (glycine betaine, ectoine). The intracellular accumulation can be achieved either by the uptake of externally available compatible solutes or by de novo synthesis.
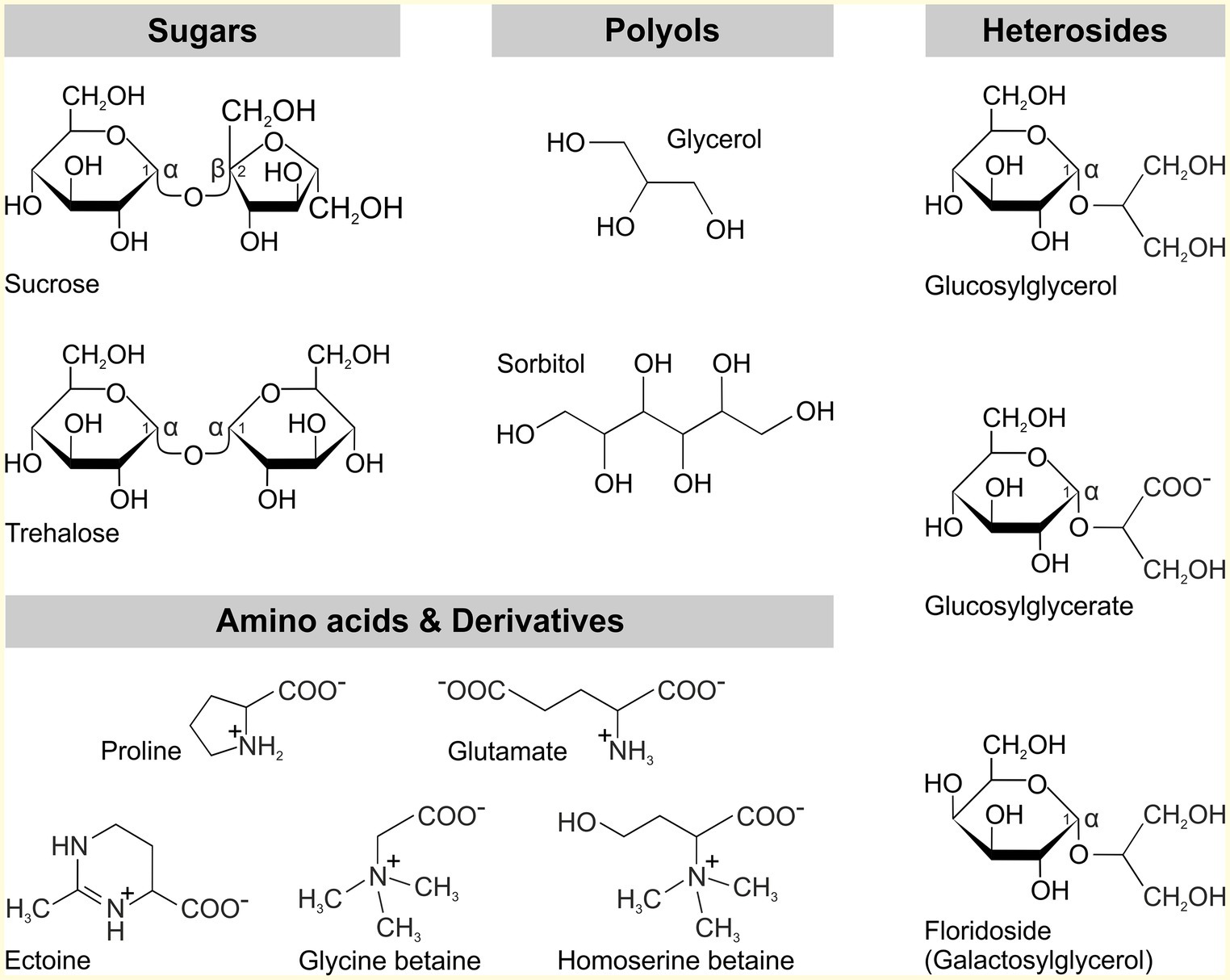
Figure 1. Classes of compatible solutes found in microorganisms.
Within the group of cyanobacteria, there is a rough correlation between the compatible solutes used to compensate osmotic potential and the natural habitat of the respective strain. For instance, freshwater strains with low salt tolerance usually use the disaccharides sucrose and trehalose, while in moderately salt-tolerant, e.g., marine species the heteroside glucosylglycerol (GG) is predominantly accumulated. Glycine betaine and glutamate betaine were identified as compatible solutes in halophilic species at extremely saline sites (Mackay et al., 1984; Hagemann, 2011). The preference of specific compatible solutes at different salinity levels might be correlated with the specific degree of protection for the stabilization of macromolecules such as enzymes or membranes (Borges et al., 2002; Hincha and Hagemann, 2004). Nevertheless, it should be noted that the classification into these three groups is not always reflected among cyanobacteria. Within the marine representatives, there are exceptions, for example, Crocosphaera watsonii exclusively accumulates trehalose (Pade et al., 2012), while most oceanic Prochlorococcus spp. mainly accumulate sucrose (Klähn et al., 2010a). Moreover, some Prochlorococcus and Synechococcus strains also synthesize glycine betaine as well as unusual solutes such as the negatively charged glucosylglycerate (Klähn et al., 2010a). In the important nitrogen-fixing strain Trichodesmium erythraeum, in turn, none of the compounds mentioned are found as compatible solutes, but this organism uses the recently identified homoserine betaine (Pade et al., 2016a).
Cyanobacterial Salt Acclimation and Biotechnology
To increase sustainability, large scale cultivation of cyanobacteria for the biotechnological production of biofuels or bulk chemicals shall avoid the use of large quantities of limited freshwater resources and should be preferentially done in brackish or seawater (Chisti, 2013). The influence of increased salt content on the production of various substances has been scarcely investigated. A few reports are available for ethanol (Pade et al., 2017), isoprene (Chaves et al., 2015; Pade et al., 2016b), and glycogen (Aikawa et al., 2014) production in salt-stressed cyanobacteria. With the model strains Synechocystis sp. PCC 6803 (hereafter Synechocystis) and Synechococcus sp. PCC 7002, two moderately salt-tolerant cyanobacteria accumulating GG as main compatible solute were used in these studies. At least with respect to the mentioned compounds, an externally supplied NaCl concentration of 2% (w/v) did not significantly affect the productivity of these strains. At a NaCl concentration of 4%, which is slightly above the average salinity of seawater, the respective production rates decreased. In this regard, the high rate of GG synthesis, thereby reducing the flow of carbon into the respective product, might be crucial for the decreased product yield in these strains (Pade et al., 2017). However, the low fluxes of carbon from the basal metabolism into the desired products represent a general problem of cyanobacterial biotechnology (Angermayr et al., 2009).
In addition to the influence of salt on the production of various substances, the compatible solutes themselves are increasingly coming into the focus of a biotechnological utilization. For example, GG can be used as stabilizing agent for enzymes or antibodies, and hence enables their long-term storage in freezers (Luley-Goedl et al., 2010; Tan et al., 2015). Moreover, due to its biochemical properties, it is also commercially used in cosmetics, i.e., as a component of several skin revitalizing creams (GG is produced by the bitop AG, Germany, and designated as Glycoin®). Sucrose is of interest as a sweetener in food industry, as a high-energy sugar for fuel production as well as a feedstock for the cultivation of heterotrophic microorganisms (Ducat et al., 2012; Du et al., 2013; Löwe et al., 2017; Kirsch et al., 2018).
Accumulation of Sucrose and Glucosylglycerol Among Salt-Stressed Cyanobacteria
The salt-induced accumulation of sucrose was first reported in Synechococcus elongatus PCC 6301 (Blumwald et al., 1983) and Anabaena variabilis (Erdmann, 1983). In the meantime, the widespread distribution of sucrose as a compatible solute has become evident on the basis of the numerous sequenced cyanobacterial genomes (Hagemann, 2013; Kolman et al., 2015). In fact, phylogenetic studies suggest that the synthesis of this sugar occurred very early in cyanobacterial evolution, and that sucrose was the first osmoprotective substance in ancient cyanobacteria (Blank, 2013). By acquiring the genes for the synthesis of other compatible solutes and concomitant conquest of marine and hypersaline habitats, the genes for sucrose metabolism were lost in some groups. Among cyanobacteria, sucrose remains the dominant compatible solute in freshwater habitats in addition to trehalose and also occurs in some moderately salt-tolerant strains such as Synechocystis as a secondary compatible solute besides GG. Presumably, the genes of sucrose metabolism were transferred to algae and plants in the endosymbiotic formation of chloroplasts (Lunn, 2002; Salerno and Curatti, 2003).
The first detection of GG in cyanobacteria was reported for extracts of Agmenellum quadruplicatum (former name of Synechococcus sp. PCC 7002; Kollman et al., 1979). Subsequently, the salt-dependent accumulation of GG and thus its function as a compatible solute could be demonstrated (Borowitzka et al., 1980). In the meantime, GG has been identified as an osmoprotective substance in more than 60 cyanobacteria (Hagemann, 2011). In many marine cyanobacteria, GG represents the primary compatible solute, but there are also numerous strains in which GG does not occur. For example, key genes of GG metabolism are absent in most strains of the ecologically important picocyanobacteria Prochlorococcus spp. (Scanlan et al., 2009). Only recently, the first member of this genus could be identified with an intact GG synthesis in the Red Sea (Shibl et al., 2018). Conversely, the synthesis of GG has also been demonstrated in some cyanobacteria originally isolated from freshwater habitats, including the model strain Synechocystis (Reed and Stewart, 1985). GG synthesis, however, is not restricted to cyanobacteria. Several heterotrophic bacteria possess a full GG synthesis pathway and accumulate GG in response to osmotic stress (Pocard et al., 1994; Roder et al., 2005).
Enzymes of Glucosylglycerol and Sucrose Synthesis
The synthesis of GG has mainly been investigated in Synechocystis. It is formed within two enzymatic steps by GG-phosphate synthase (GgpS, Marin et al., 1998) and GG-phosphate phosphatase (GgpP, Hagemann et al., 1997a). GgpS is a glucosyltransferase catalyzing the synthesis of GG 3-phosphate from ADP-glucose and glycerol 3-phosphate. This intermediate is subsequently dephosphorylated by GgpP (Figure 2; Hagemann and Erdmann, 1994). Albeit these two enzymes are synthesized at a basal level under low salt conditions, they are largely inactive and are activated after a salt shock. All GG-accumulating cyanobacteria investigated so far possess two separate genes for GgpS and GgpP, whereas some heterotrophic bacteria also harbor fusion proteins with both catalytic activities (Hagemann et al., 2008).
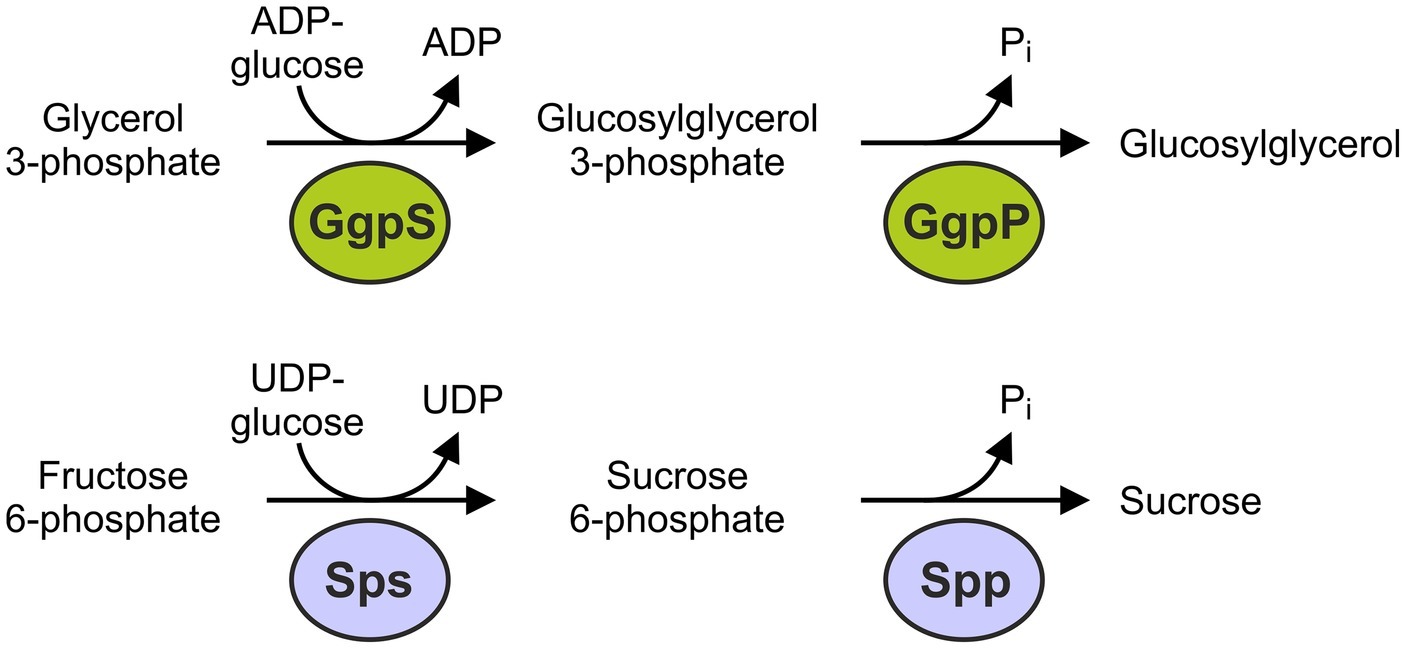
Figure 2. Reactions and enzymes involved in the de novo synthesis of glucosylglycerol (GG) and sucrose in cyanobacteria.
As in plants, the biosynthesis of sucrose in cyanobacteria is executed in a two step reaction by the enzymes sucrose-phosphate synthase (Sps, Curatti et al., 1998; Hagemann and Marin, 1999) and sucrose-phosphate phosphatase (Spp, Lunn, 2002). The Sps catalyzes the reaction of UDP-glucose and fructose 6-phosphate to sucrose 6-phosphate. Spp causes dephosphorylation and sucrose is released (Figure 2). Thus, the biosynthetic pathways of sucrose and GG are basically similar because they are formed in two enzymatic steps starting from an activated nucleotide sugar via a phosphorylated intermediate, which is synthesized by a glucosyltransferase. This also applies to the compatible solutes trehalose and glucosylglycerate (Klähn and Hagemann, 2011). In cyanobacteria, there are two structurally different forms of Sps. The proteins of Nostoc (Anabaena) sp. PCC 7119 and some other strains only require the glucosyltransferase domain for catalytic activity (Porchia and Salerno, 1996; Salerno and Curatti, 2003). More common are Sps proteins in which the glucosyltransferase domain is fused to an inactive phosphohydrolase domain, while the second reaction is catalyzed by a separate Spp protein. The first Sps variant appears to be exclusive in a few filamentous cyanobacteria, while the latter is present in most cyanobacteria, including Synechocystis, as well as algae and plants (Blank, 2013). The fusion of the glucosyltransferase and phosphohydrolase domains raises the question of whether bifunctional Sps proteins, which can catalyze the two steps in the synthesis of sucrose, can also be found in some organisms. In fact, the Sps of Synechococcus elongatus PCC 7942 also appears to have phosphohydrolase activity in vitro (Martinez-Noel et al., 2013). It is unclear to what extent the synthesis of sucrose in this strain is performed via this single protein, or is completed by the occurring separate Spp in vivo. An alternative route for the synthesis of sucrose can be catalyzed by the enzyme sucrose synthase (SuSy) through the reversible linkage of NDP-glucose and fructose with NDP cleavage (Porchia et al., 1999). In cyanobacteria, SuSy is important in heterocystous, nitrogen-fixing strains, in which it catalyzes the reverse reaction – i.e., the cleavage of sucrose to produce carbon skeletons (Curatti et al., 2002; Kolman et al., 2015).
Salt-Induced Accumulation of Glucosylglycerol and Sucrose in Synechocystis
In the model strain Synechocystis, the accumulation of GG and sucrose is induced immediately after a salt shock. In the case of GG, it reaches a final concentration after about 24 h, which then remains stable in salt-acclimated cells (Figure 3A) and is adjusted to the external salt level (Marin et al., 2002). Sucrose also accumulates rapidly in the cell within few hours (Figure 3B). However, this is a transient accumulation with a maximum of about 6–12 h after the salt shock (Desplats et al., 2005; Kirsch et al., 2018). As a result, the sucrose content drops again almost to the initial level within 24 h, likely due to the replacement by GG. In cyanobacteria of low salt tolerance, in which sucrose is the primary compatible solute (e.g., Synechococcus elongatus PCC 7942), this accumulation occurs to a final concentration and remains stable at that level thereafter (Blumwald et al., 1983).
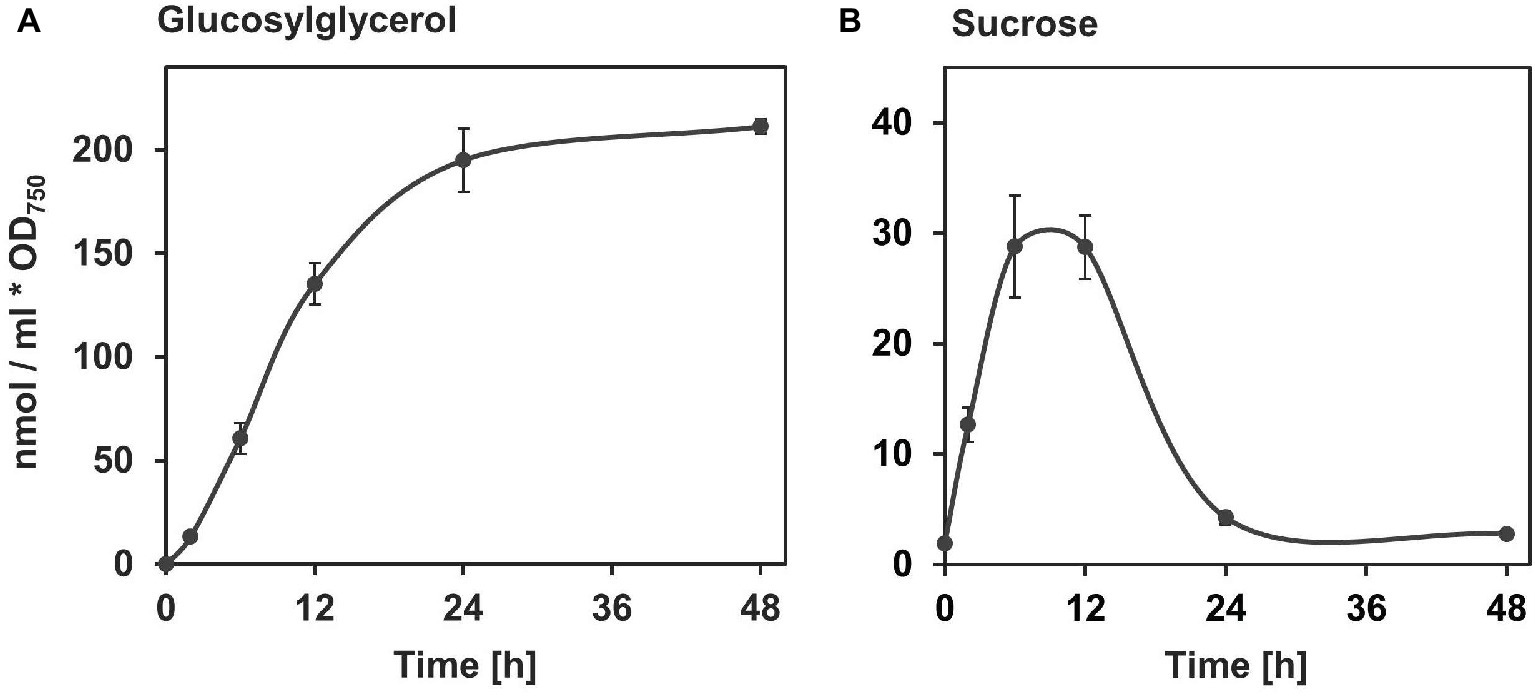
Figure 3. Salt-induced accumulation of the compatible solutes GG (A) and sucrose (B) in Synechocystis sp. PCC 6803. Synechocystis cells pre-cultivated in NaCl-free media were exposed to 3.5% NaCl (~ 600 mM, equivalent to full seawater conditions) at time point 0 and the compatible solutes were quantified via gas chromatography (Kirsch et al., 2017, 2018). The milliliter in the unit at the ordinate refers to culture volume.
Activation of Glucosylglycerol-Synthesis After Salt Shock – Gene Expression and Biochemical Regulation
Increases in salt content require a rapid and coordinated cell response. In many heterotrophic bacteria, the synthesis of compatible solutes is mainly activated at the transcriptional level, e.g., the salt-induced synthesis of proline is initiated by the transcriptional activation of the proHJ operon in Bacillus subtilis (Brill et al., 2011). This operon codes for the enzymes responsive for osmoadaptive proline synthesis, which is independent from proteins involved in the anabolic proline synthesis (Hoffmann and Bremer, 2016). In the case of GG, Synechocystis activates the pathway at both the biochemical and transcriptional level. Cells cultivated under NaCl-free conditions already contain a basal amount of inactive GgpS protein, which is activated very rapidly by a sudden increase in salinity. As it turned out, this biochemical activation takes place directly through NaCl or other salts (Hagemann and Erdmann, 1994). Since recombinant GgpS protein purified from E. coli has NaCl-independent activity, it was concluded that an inhibitor bound to GgpS is removed by NaCl in vivo (Hagemann et al., 2001). In fact, at low salinities, GgpS can be bound to nucleic acids due to electrostatic binding to the negatively charged phosphate groups, which results in an inactive enzymatical state (Novak et al., 2011). After salt shock, inorganic ions are flowing into the cell and can disrupt this electrostatic bond, which results in GgpS release and its activation. Thus, the initial biochemical activation of GG synthesis is mediated by the ions entering a cell upon salt addition, as has been shown for salt-shocked cyanobacteria (Figure 4; Reed et al., 1985; Hagemann et al., 1994). Interestingly, the salt-dependent activity of the Synechocystis GgpS was also observed when the gene was expressed in the heterologous host Corynebacterium glutamicum (Roenneke et al., 2018). In Corynebacterium as in Synechocystis, GgpS was active when cells were exposed to salt stress, however, in contrast to Synechocystis GG continuously leaked from the engineered Corynebacterium cells. The increases in salinity also induce ggpS gene expression in Synechocystis (Marin et al., 2004), thereby adjusting the amount of enzyme in addition to its activity regulation by changes in the salt content. Nevertheless, some strain-specific aspects should be noted here. For instance, in the marine strain Synechococcus sp. PCC 7002, the biochemical regulation of GgpS seems to play a minor role whereas the transcriptional induction of ggpS in salt-stressed cells is dominating (Engelbrecht et al., 1999).
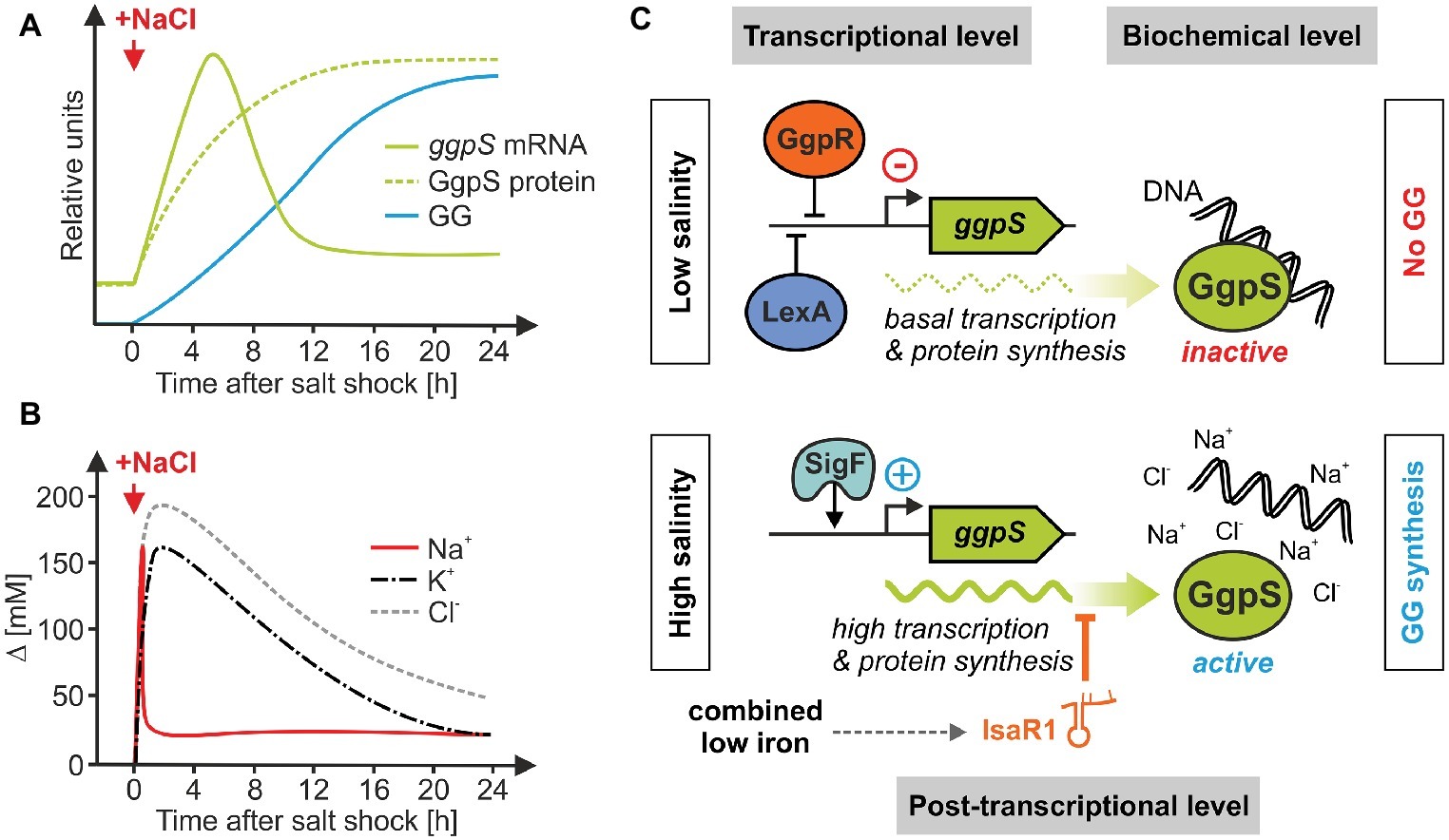
Figure 4. Regulation of salt-induced GG-synthesis via the key enzyme GgpS involves different regulatory layers in Synechocystis. (A,B) Kinetics of ggpS mRNA, GgpS protein, GG accumulation, and changes of salt ion concentrations after sudden salt shocks in Synechocystis [data have been obtained from Marin et al., 2002 (A) and Reed et al., 1985 (B)]. (C) Three proteins act on the transcriptional level, GgpR (Klähn et al., 2010b), LexA (Kizawa et al., 2016), and SigF (Huckauf et al., 2000; Marin et al., 2002). The sRNA IsaR1 interacts with ggpS mRNA at the postranscriptional level (Rübsam et al., 2018). The rapid activation of GG synthesis and its adjustment to the final salt content seem to be mainly regulated at the biochemical level via the cellular amount of inorganic ions (Novak et al., 2011).
Remarkably, the ggpS mRNA shows a salt-dependent accumulation that roughly follows the kinetics of the intracellular salt ion concentrations in response to salt shocks (Figures 4A,B). The induction of ggpS expression involves several regulatory components in Synechocystis (Figure 4C). First, on a superior level, the adjustment of ggpS transcription involves the use of alternative sigma factors. In Synechocystis for instance, the absence of the SigB and SigD factors lead to a decreased salt tolerance, which, however, is probably due to a generally reduced stress tolerance (Pollari et al., 2008) as found before by the knock out of the stress-regulatory sigma factors in B. subtilis (Hecker et al., 2007). A more specific role in salt acclimation appears to play SigF, because a sigF mutant strain shows markedly reduced ggpS expression accompanied by an impaired acclimation capacity to salt stress (Huckauf et al., 2000; Marin et al., 2002). Second, the genes of GG metabolism in Synechocystis have been reported to be under negative control of the transcription factor LexA, because the lexA mutant strain showed increased ggpS transcript levels already under low salt conditions (Kizawa et al., 2016). However, given the large number of LexA-regulated genes, this is likely more a global rather than a salt-specific regulation that interferes with GG synthesis in addition to various other cellular processes. Third, likely more GG-specific, regulatory functions are mediated at the transcriptional level by a small protein that is encoded by a gene located directly upstream of ggpS. It was shown that a deletion of this gene resulted in de-regulated ggpS expression. Hence, the respective protein appears to repress the expression of ggpS under low-salt conditions and was named ggpS repressor (GgpR) (Klähn et al., 2010b). It should be noted that clear homologs of GgpR are missing in the databases. However, a similar gene organization, namely a short open reading frame upstream of ggpS, was also found in a few other GG-accumulating cyanobacterial strains suggesting that such an organization rather than a conserved, DNA-binding repressor protein could be responsible for the salt-dependent transcriptional induction.
Moreover, the ggpS gene of Synechocystis was also shown to be target of post-transcriptional regulation by a small regulatory RNA (Figure 4C). The iron-stress activated RNA 1 (IsaR1) was previously identified as key regulator for adjusting the photosynthetic apparatus of various cyanobacteria to iron deficiency (Georg et al., 2017). This regulation is mainly conducted by complementary base-pairing between IsaR1 and its target mRNAs, which in turn interferes with protein biosynthesis, i.e., under iron-depletion when IsaR1 levels are strongly increased. Interestingly, the ggpS mRNA is also targeted by IsaR1. Thereby, IsaR1 clearly diminishes the de novo synthesis of GgpS protein as well as GG (Rübsam et al., 2018). However, the biological meaning of post-transcriptional ggpS regulation by a small RNA that is predominantly expressed under iron deficiency is not yet understood. Most likely, this is of importance when cells are exposed to parallel fluctuations in both salinity (triggering ggpS expression) and iron availability (triggering IsaR1 expression). In this regard, it should be noted that in natural environments, both situations frequently occur in parallel, e.g., in estuary brackish water habitats, where an increase in salinity is accompanied by a rapid removal of iron from river water (Boyle et al., 1977). Obviously, IsaR1 coordinates the response of Synechocystis to iron starvation with osmotic acclimation and hence is involved in the integration of responses to different environmental perturbations.
Altogether, GG accumulation in salt-shocked Synechocystis cells is regulated at different levels; however, the interplay and communication between these systems is largely unknown. The rapid start of GG accumulation immediately after salt shock seems to be mainly achieved via the direct activation of the basically synthesized GgpS protein. To ensure an efficient GG synthesis in salt-acclimated cells, an enhanced GgpS amount is necessary, which is apparently ensured by the release of the repressor GgpR and positive action of SigF. The GG synthesis must be imbedded into the overall carbon utilization network, which is probably regulated via LexA, since this transcriptional activator also affects the expression of many other genes involved in primary carbon metabolism (Kizawa et al., 2016). Finally, salt stress is often accompanied by multiple other stressors and the responses need to be integrated. This might be exemplified by IsaR1 that is likely part of a coordinated acclimation to simultaneous iron deficiency and salt stress frequently occurring in natural habitats.
Activation of Sucrose-Synthesis After Salt Shock – Gene Expression and Biochemical Regulation
Compared to the regulatory mechanisms involved in the synthesis of GG, less is known for sucrose. However, in view of the very rapid accumulation of this sugar in salt-stressed cells of Synechocystis, a rapid activation of enzymes is to be expected here as well (Figure 5). Similar to GgpS in GG synthesis, Sps appears to be the rate-limiting enzyme in sucrose synthesis, as modifications of Sps expression have the greatest effect on the rate of sucrose accumulation (Du et al., 2013). Likewise, as with GgpS, stimulation of Sps activity by NaCl has been shown in S. elongatus PCC 6301 (Hagemann and Marin, 1999). However, this is not true for all Sps proteins, such in vitro stimulation could not be observed with the enzyme from Nostoc (Anabaena) sp. PCC 7120. These results suggest that within cyanobacteria, different regulatory aspects exist instead of a conserved mechanism for biochemical activation of sucrose synthesis. To which extent the biochemical activation of Sps is based on a protein-DNA interaction, similar to GgpS, is still unclear.
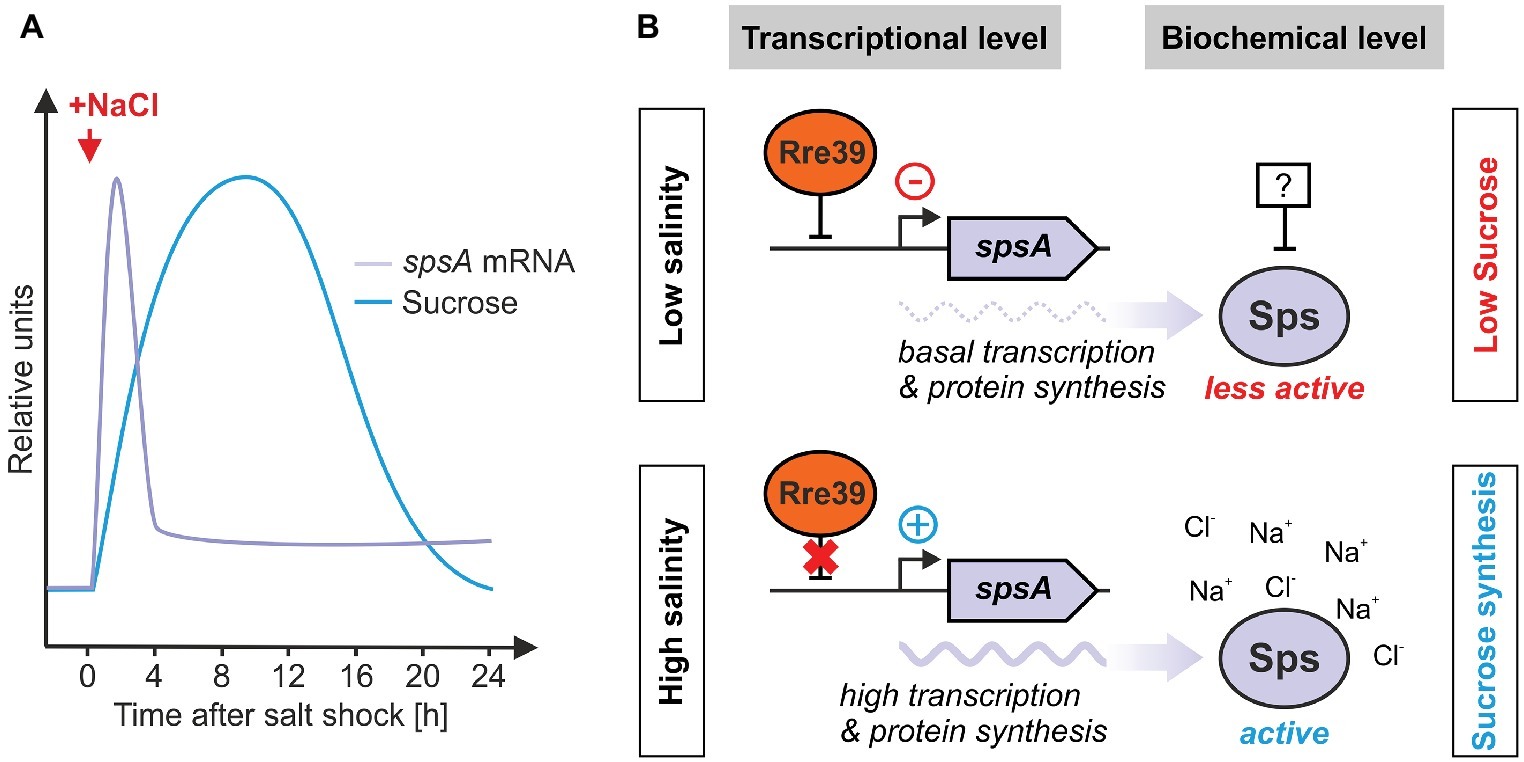
Figure 5. Regulation of salt-induced sucrose accumulation in Synechocystis. (A) The relative spsA mRNA and sucrose accumulation kinetics in response to a salt shock have been approximated from Marin et al. (2004) and Kirsch et al. (2018). (B) Regulation of expression and activity of sucrose-phosphate synthase by the response regulator Rre39 and the intracellular ion levels.
At the transcriptional level, the induction of spsA gene expression occurs within minutes after a salt shock (Marin et al., 2004; Desplats et al., 2005). However, this induction is significantly lower than for ggpS (induction factor of ~ 85 for ggpS versus a factor of ~ 3 for spsA, Marin et al., 2004). The factors involved in the transcriptional regulation of Sps are still largely unknown. Recently, the influence of response regulator 39 (Rre39; Slr1588) on Sps expression has been identified (Song et al., 2017). In mutant Δslr1588, increased expression of spsA was observed, indicating a possible negative regulation by Rre39. Sucrose synthesis in salt-stressed cells of Nostoc (Anabaena) sp. PCC 7120 is regulated by the Rre named OrrA (Ehira et al., 2014). Rre’s are usually components of bacterial two-component signal transduction systems, which consist of a sensory histidine kinase (Hik) and a Rre and are also important sensor systems for various types of stress in cyanobacteria (Los et al., 2010). In E. coli, such two-component systems are also involved in the sensing of salt stress. The Hik/Rre pairs KdpD/KdpE (salt-dependent regulation of K+ uptake, Jung and Altendorf, 2002) and EnvZ/OmpR (salt-dependent regulation of permeability of the outer membrane, Mizuno and Mizushima, 1990) play a role here. Synechocystis Rre39, however, is an “orphan” response regulator, i.e., so far no associated Hik could be identified. Overall, five Hik/Rre systems have been reported that play a role in signal transduction under salt stress in Synechocystis (Marin et al., 2003; Shoumskaya et al., 2005). However, among the genes regulated, there are none that play a role in the synthesis of GG or sucrose. Hence, the signal transduction cascades triggering the transcriptional resonse of ggpS and spsA remain enigmatic.
Response to Decreasing Salinities Involves the Degradation of Compatible Solutes
Unlike the accumulation of compatible solutes upon increasing salt levels, little is known about the cellular response of cyanobacteria to a decreasing salt concentration. Despite the opposite direction, salinity reduction may be just as challenging for a microorganism as it is an increase in salinity. In particular, due to the highly accumulated compatible solutes, the osmotic potential of the cytosol becomes lower compared to the extracellular space when salinity decreases. Accordingly, this leads to a flow of water into the cell that can represent a serious problem. For example, a hypoosmotic shock equivalent to 0.3 M has been shown to increase the turgor pressure from 4 (~400 kPa) to 11 atm (~1.1 MPa) in E. coli (Booth and Louis, 1999). This poses a dramatic burden on the cytoplasmic membrane and the cell wall. To prevent the cell from bursting, the particular organism must rapidly establish the osmotic balance between the cell’s interior and exterior. Two scenarios are conceivable for intracellularly enriched compatible solutes–either these are enzymatically degraded and metabolized or the intracellular substance concentration is adjusted by release from the cell. The latter represents a widely used strategy for adapting to rapidly and strongly decreasing salinity and is mediated by mechanosensitive channels (Levina et al., 1999). These channels are opened following a hypoosmotic shock, when the water influx induces a mechanical tension on the cell membrane exceeding a certain threshold. The enzymatic degradation of compatible solutes represents a slower response to less strong hypoosmotic conditions. Nevertheless, this strategy also has advantages, e.g., the large amounts of organic carbon (and sometimes nitrogen) incorporated into compatible solutes are retained for cellular metabolism. It is therefore not surprising that microorganisms have evolved precisely regulated ways of recycling compatible solutes internally.
The Release of Compatible Solutes
When salt-acclimated cells of Synechocystis are exposed to a sudden hypoosmotic shock, i.e., when the salt concentration of the medium is lowered by at least 250 mM, large amounts of GG are immediately released into the medium (Fulda et al., 1990). This rapid reaction is likely relying on the opening of mechanosensitive channels (Msc) through which dissolved substances escape from the cell as shown for E. coli (Booth and Louis, 1999). The genome of Synechocystis contains eight candidate genes for homologs of MscS and one gene for MscL (Pivetti et al., 2003). So far, however, only the MscL (encoded by gene slr0875) has been demonstrated to function in adapting to hypoosmotic shocks (Nanatani et al., 2013). The mechanosensitive channels show a low specificity because, in addition to compatible solutes, a whole series of other dissolved substances such as inorganic ions and amino acids also escape into the medium (Fulda et al., 1990). Presumably, this release of solutes after a hypoosmotic shock is a rapid emergency reaction that prevents the cell from bursting due to osmotic water influx.
Intracellular Degradation of Glucosylglycerol
In natural habitats such rapid changes in salinity are rather rare, but it could appear in mixing zones at sea shores and dry soils during heavy rain. Since the release of compatible solutes also results in a significant loss of previously fixed carbon, the existence of degradation pathways for the intracellular metabolisation of these substances can be assumed. In fact, compatible solutes in many bacteria also serve as storage substances for carbon and nitrogen (Welsh, 2000). This was demonstrated, for example, for the substances glycine betaine (Wargo, 2013), proline (Moses et al., 2012), and ectoine (Schulz et al., 2017). In cyanobacteria, specific evidence emerged from experiments, in which salt-adapted Synechocystis cultures were supplemented with the alternative compatible solute trehalose (Mikkat et al., 1997). It was found that the externally available trehalose was taken up and accumulated intracellularly. At the same time, the GG content in the cells decreased without GG being detected in the medium. These results suggested that GG was degraded intracellularly and replaced by the ingested trehalose. Another indication of a GG-degrading pathway was found in Microcoleus chthonoplastes (Moezelaar et al., 1996). Cultures that were grown in darkness and under anaerobic conditions performed fermentation processes in which GG was degraded in addition to glycogen. The glucose residue was fermented and the glycerol residue was released into the medium. Moreover, 13C-labeling experiments in Synechococcus sp. PCC 7002 revealed a strong exchange of labeled carbon between the pools of glycogen and GG (Baran et al., 2017). This indicates a continuous conversion of GG by synthesis and degradation at least in this strain.
The above mentioned studies suggested that GG-synthesizing cyanobacteria also have a degradation pathway for GG and hence, its intracellular recycling. Recently, the responsible enzyme glucosylglycerol hydrolase A (GghA) was identified in Synechocystis. This enzyme hydrolyzes GG into glycerol and glucose (Savakis et al., 2016; Kirsch et al., 2017). It could be shown that GghA is responsible for the depletion of intracellular GG in response to decreasing salinities and thus represents a significant element of the cyanobacterial salt acclimation process (Kirsch et al., 2017). Interestingly, in the genome of Synechocystis, the respective gene gghA (slr1670) is located upstream of the ggpS gene, however on the complementary DNA strand (Figure 6). The promoters of both genes overlap in the region encoding the small regulatory protein GgpR (Klähn et al., 2010b). Transcriptome analyses verified a salt-dependent expression of gghA (Marin et al., 2004). The gghA gene is likely co-transcribed with the genes glpK (slr1672) encoding glycerol kinase and spoU (slr1673) that encodes a putative tRNA/rRNA methyltransferase. GlpK plays a role in the formation of glycerol 3-phosphate and thus enables reassimilation of the released glycerol back into primary metabolism. The co-localization of gghA with ggpS and glpK appears to be conserved among GG-synthesizing strains (Figure 6).
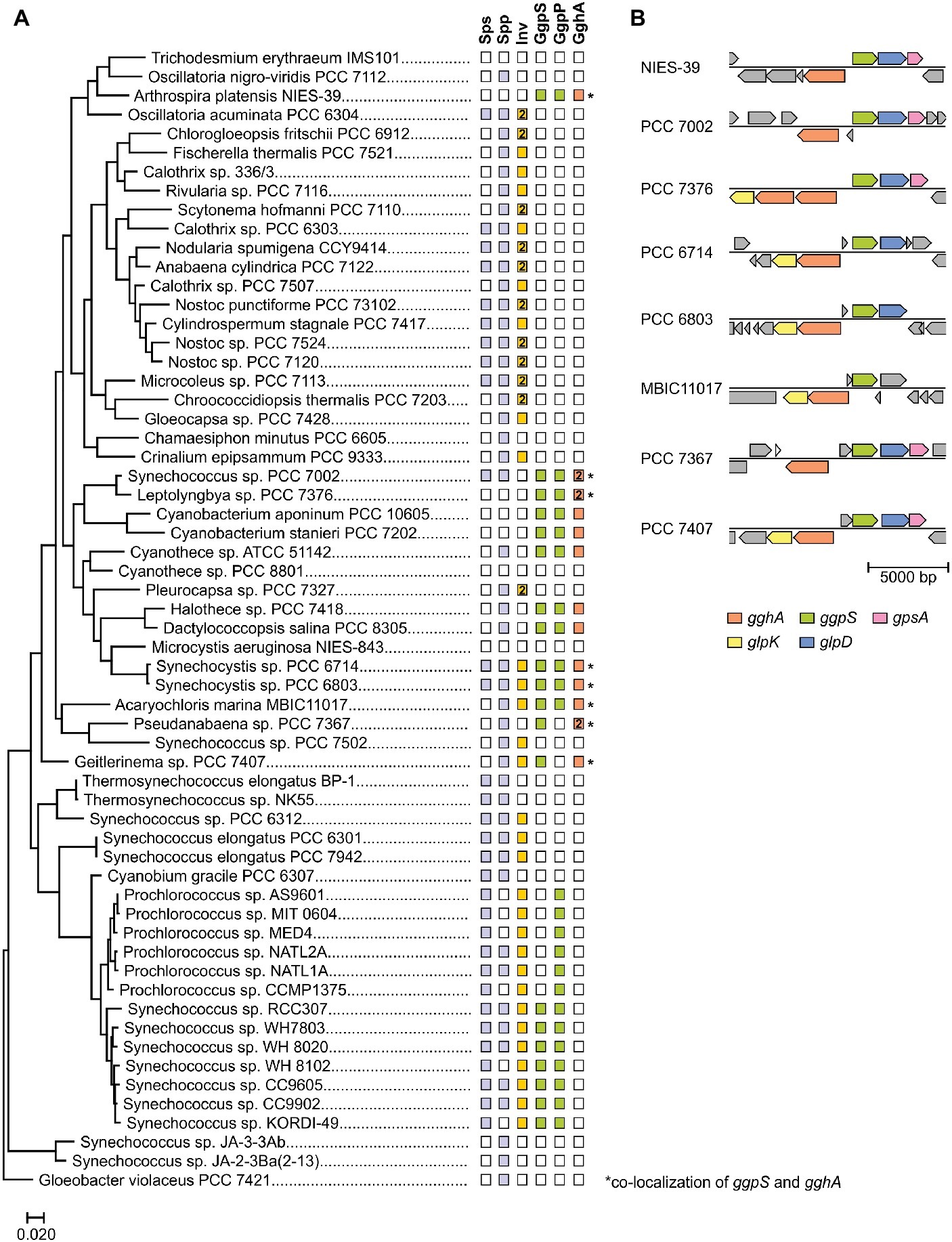
Figure 6. Distribution of GG-synthesis (ggpS) and GG-degradation (gghA) genes among cyanobacteria. (A) Phylogenetic tree of selected cyanobacteria based on 16S rDNA sequences. The tree was generated with the MEGA X (Kumar et al., 2018) software package and the Maximum Likelihood method. The presence of genes (illustrated by filled rectangle) was investigated using the BLASTP algorithm. As reference the amino acid sequences of functionally verified GgpS, GgpP, GghA, etc. from Synechocystis sp. PCC 6803 were used. (B) In various cyanobacteria ggpS and gghA are co-localized (glpD, glycerolphosphate dehydrogenase; ggpS, GG-phosphate synthase; gghA, GG hydrolase A; glpK, glycerol kinase; gpsA, glycerol 3-phosphate dehydrogenase).
With the hydrolytic cleavage of GG into the products glucose and glycerol, GghA differs from the previously identified GG-splitting enzyme GG-phosphorylase (GGP) from Bacillus selenitireducens, which catalyzes the reversible cleavage of GG into glucose-1-phosphate and glycerol (Nihira et al., 2014). Interestingly, the activity of GghA becomes increasingly inhibited by adding inorganic salts to in vitro enzyme assays (Kirsch et al., 2017). Moreover, a ΔgghA mutant of Synechocystis showed a clear phenotype, as this strain lost its ability to adjust the GG content to decreasing salt concentrations. This was demonstrated by two experiments in which salt-adapted cultures of the wild type and the ΔgghA mutant were exposed to either a gradual reduction in salinity or an abrupt hypoosmotic shock. Continuous dilution of the salinity over several hours did not result in the release of GG into the medium, but the wild type simultaneously adjusted the intracellular GG concentration to the altered salinity. This was not the case in the ΔgghA mutant, in which the amount of GG remained unchanged (Kirsch et al., 2017).
Moreover, GghA also plays an important role in the acclimation to sudden hypoosmotic shock treatments. As mentioned above, after such a treatment, both the wild type and the ΔgghA mutant strain release GG into the culture medium. However, GG quickly disappears from the medium and cells of wild type, whereas it remains in supernatants of the mutant ΔgghA (Kirsch et al., 2017). It has been shown that an ABC-type transporter named Ggt is involved in the uptake of GG but also trehalose and sucrose when externally provided to the cells. Mutation of the transporter resulted in the continuous release of GG into the medium, which gave raise to the hypothesis that the main function of the Ggt system is related to the re-uptake of compatible solutes leaked out the cells to avoid losses of organic carbon (Hagemann et al., 1997b; Mikkat and Hagemann, 2000). Correspondingly, the released GG after hypoosmotic shocks is likely re-imported into cells of the wild type by the Ggt system, where it is hydrolyzed by GghA and the glucose is then used to provide carbon and energy supporting the quick acclimation to the new growth conditions. In contrast to wild type, the ΔgghA mutant was unable to use the released GG and showed a decreased ability to restart growth after hypoosmotic treaments. These experiments illustrated that the GG degradation is crucial for the acclimation to fluctuating salinities.
The importance of GghA is underlined by the fact that several GG synthesizing cyanobacteria even harbor two copies of the gghA gene. However, a gghA gene is absent from marine picocyanobacterial strains of the genus Synechococcus all of which harbor ggpS and ggpP genes (Figure 6). These genes are functional and lead to intracellular GG accumulation, which has been verified, e.g., for Synechococcus sp. WH7803 and WH8102 (Klähn et al., 2010a). The lack of a GG-degrading pathway in these strains, however, could be explained by a rather constant saline environment in the open oceans and the needlessness of osmotic adjustments in response to decreasing salinities. Nevertheless, an alternative, so far unknown GG-degradation pathway cannot be excluded.
In addition to the described defect in GG degradation, another interesting phenotypical change was observed for the strain lacking GghA. The cultures of the ΔgghA mutant became very viscous when grown under NaCl-free conditions, suggesting the release of extracellular polysaccharides (EPS) from the cells (Kirsch et al., 2017). It is therefore of interest to investigate a possible role of GG or its intermediate GGP in the synthesis of these substances. Research on the complex functions of EPS in biofilm formation and adaptation to different types of stress have become increasingly interesting in recent years (Pereira et al., 2009).
Intracellular Degradation of Sucrose
Within cyanobacteria, three different degradation pathways for sucrose have been identified so far (Kolman et al., 2015). The responsible enzymes are sucrose synthase, amylosucrase and invertase (Inv). Sucrose synthase can catalyze both synthesis and cleavage of sucrose, but appears to be active primarily in the direction of the cleavage reaction to (A/U)DP-glucose and fructose in vivo (Porchia et al., 1999; Curatti et al., 2002). This enzyme is mainly found in heterocyst-forming cyanobacteria and plays an essential role for N2-fixation (Kolman et al., 2015). Amylosucrase has recently been described in Synechococcus sp. PCC 7002 (Perez-Cenci and Salerno, 2014), where it splits sucrose into free fructose and glucose, the latter is then linked via glycosidic bond to oligosaccharides or glycogen. In most cases, however, the degradation of sucrose among cyanobacteria occurs by the enzyme Inv, which catalyzes the irreversible cleavage of sucrose into the monosaccharides glucose and fructose. Phylogenetic studies suggested that invertases were part of the original sucrose metabolism and were transferred from cyanobacteria into plants (Vargas and Salerno, 2010). The invertases form a large and diverse group of sucrose-cleaving enzymes. A biochemical classification can be made on the basis of the pH optimum for enzyme activity, distinguishing invertases with a pH optimum in the acid range from pH 4.5 to 5 (acid invertases, Ac-Inv) and invertases having an optimum in the neutral to slightly alkaline range from pH 6.5 to 8 (alkaline/neutral invertases; A/N-Inv). Phylogenetically, Ac-Inv and A/N-Inv are unrelated. The Ac-Inv are β-fructofuranosidases (EC 3.2.1.26) that are not only specific for sucrose but can also cleave other β-fructose-containing oligosaccharides such as raffinose and stachyose (Sturm, 1999). They occur mainly in heterotrophic bacteria, yeasts, and plants; e.g., in plants, they are extracellularly bound to the cell wall or localized in the vacuole. Ac-Inv are of great importance for the food industry and hence, have been extensively studied (Nadeem et al., 2015). The occurrence of A/N-Inv is, however, limited to cyanobacteria and plants (Vargas et al., 2003). These enzymes differ from the Ac-Inv not only in terms of their pH optimum, but also catalytically. Thus, they do not belong to the family of β-fructofuranosidases because they are specific for sucrose and do not cleave other β-fructose-containing substrates (Vargas and Salerno, 2010). Recently, the crystal structure of the alkaline invertase from Anabaena (InvA) has provided clues for the catalytic mechanism of these enzymes. The A/N Inv of cyanobacteria and plants specifically cleave the α, β-1,2-glycosidic linkage of sucrose and represent a distinct family of glucosidases (Xie et al., 2016).
The role of sucrose degradation by the enzyme Inv in salt acclimation of Synechocystis has recently been investigated (Kirsch et al., 2018). Similar to most unicellular cyanobacteria (Kolman et al., 2015), also the genome of Synechocystis harbors one gene that is annotated as A/N-Inv (CyanoBase–http://genome.annotation.jp/cyanobase). To gain evidence, the respective gene sll0626 was expressed in E. coli. The recombinant protein was able to cleave sucrose in vitro, with an optimum at pH 7. In the slightly alkaline region at pH 8, the activity of Sll0626 was reduced by 23%. These data show that Sll0626 is a neutral invertase, named Synechocystis invertase (SyInv) (Kirsch et al., 2018). The measured Km for sucrose of 14.7 mM is in good agreement with the invertases from Anabaena (36 mM for A-Inv and 15 mM for N-Inv, Vargas et al., 2003) and plants (10 mM for A/N-Inv, Sturm, 1999). Interestingly, the activity of SyInv becomes increasingly inhibited by adding inorganic salts to in vitro enzyme assays. The activity was almost halved at 75 mM NaCl and reached only about 15% of the initial level at 145 mM NaCl (Kirsch et al., 2018). These inhibitory ion concentrations are markedly exceeded in salt-stressed cells of Synechocystis (Hagemann et al., 1994).
The mutation of SyInv provided strong evidence that this protein is responsible for in vivo sucrose degradation in Synechocystis (Kirsch et al., 2018). Unlike the wild type, the Δinv mutant already showed significant amounts of sucrose when grown under NaCl-free conditions, which indicates active sucrose turnover and explains the low steady state content of sucrose in wild-type cells. The addition of 3.5% NaCl induced the synthesis of sucrose in the wild type as well as in the mutant. However, sucrose enrichment was not transient in the Δinv mutant. In contrast to the sucrose accumulation kinetics in wild type (see Figure 3), the Δinv mutant showed constantly enhanced sucrose content also after long-term salt acclimation (Kirsch et al., 2018). Hence, the quick decrease of sucrose accumulation in salt-shocked wild-type cells is mainly based on the action of sucrose degradation via SyInv.
Biochemical Model of the Salt-Dependent Activation of Glucosylglycerol and Sucrose Accumulation in Synechocystis
The biochemical data obtained for compatible solute synthesizing as well as degrading enzymes suggest opposite controlling in cyanobacteria. On the one hand, increased inorganic ion amounts (represented by NaCl) stimulate the activity of enzymes for the synthesis of GG and sucrose in vitro. On the other hand, the activity of degrading enzymes is inhibited by higher concentrations of inorganic ions. Based on these findings we suggest a biochemical model for the control of compatible solute accumulation in Synechocystis (Figure 7). Exposure to enhanced salinities immediately activates the accumulation of sucrose and GG due to ion-mediated enzyme activation. It has been shown that Na+ ions move inside the cells within seconds, and are later exchanged by K+, while compatible solute biosynthesis starts only thereafter within minutes (Reed et al., 1985, see Figure 4B). While GgpS and probably also Sps activities are activated, the degradation enzymes GghA and SyInv are switched off at the same time. This inverse enzyme behavior may result in the observed rapid initial GG and sucrose accumulation (Figure 7).
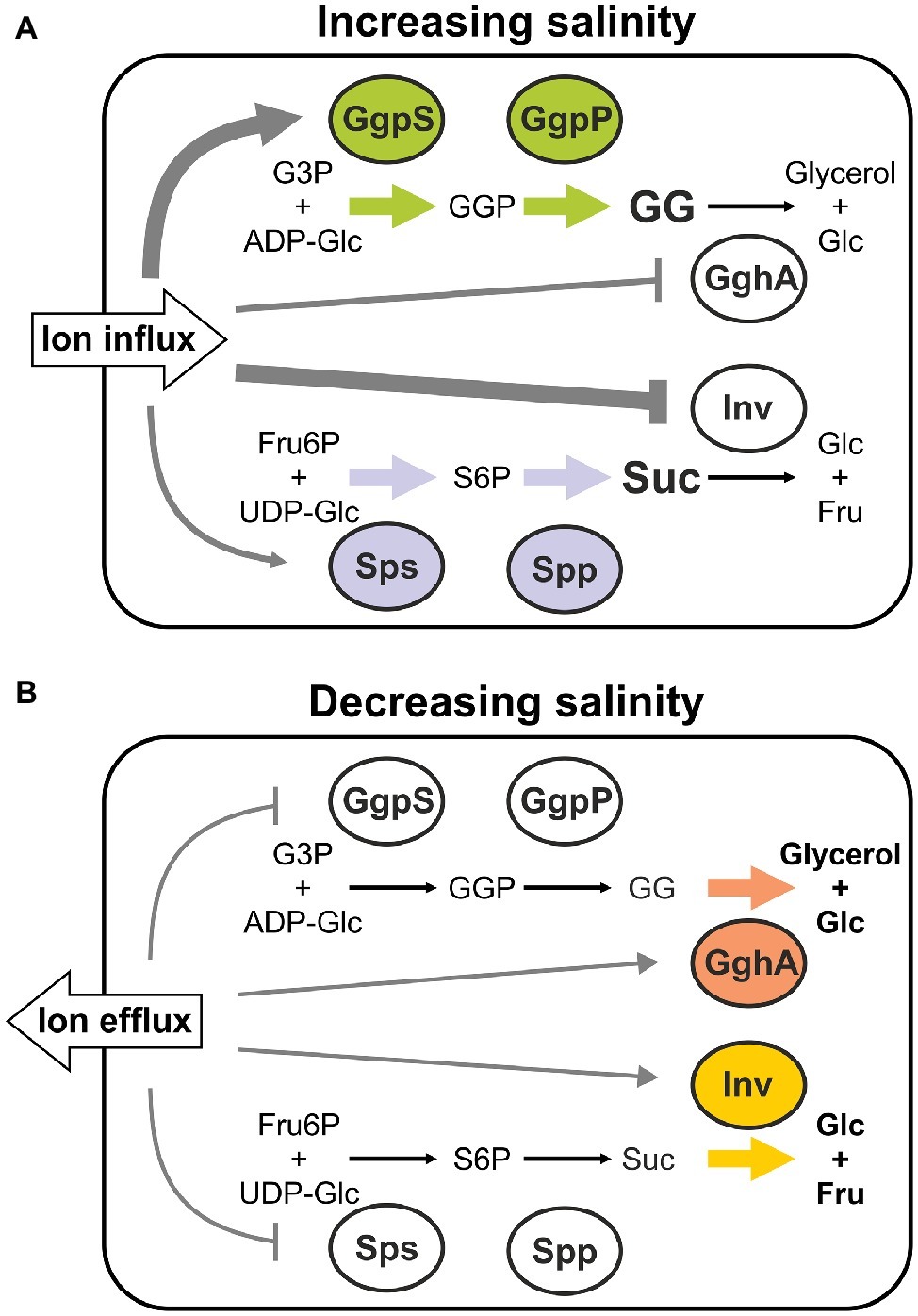
Figure 7. Biochemical regulation of GG and sucrose accumulation in salt-treated cells of Synechocystis sp. PCC 6803 via inorganic ions. The hypothetical model is based on the different response of corresponding synthesis and degradation enzymes toward changing inorganic ion contents. The increase (A) or decrease (B) of external salinity results in the influx or efflux of inorganic ions, which in turn modulate the activities of enzymes involved in GG and sucose biosynthesis and degradation, respectively. (GgpS, GG-phosphate synthase; GgpP, GG-phosphate phosphatase; GghA, GG hydrolase A; Sps, sucrose-phosphate synthase; Spp, sucrose-phosphate phosphatase; Inv, invertase; G3P, glycerol 3-phosphate; ADP-Glc, ADP-glucose; GG, glucosylglycerol; GGP, GG-phosphate; Glc, glucose; Fru, fructose; Fru6P, fructose 6-phosphate; UDP-Glc, UDP-glucose; S6P, sucrose 6-phosphate; Suc, sucrose).
At a later stage, the osmotic balance between the cell interior and the exterior is increasingly achieved by the accumulated compatible solutes, which permits to reduce the intracellular inorganic ions. According to our model, the decrease of the intracellular ion pool is responsible for the decreased sucrose accumulation after 12 h, because lower ion contents result in lowered Sps activity but particularly increased SyInv activity. The predominant effect of increased Inv activity is supported by the finding that the decrease of the sucrose pool is almost absent in mutant Δinv (Kirsch et al., 2018). The synthesis of GG continues with maximum speed for longer times until the steady state level is reached. Probabably the decreased GgpS activity in the presence of lowered intracellular ion contents is compensated by the massive expression stimulation of ggpS after salt shocks (Marin et al., 2004). In this context, the sucrose content is preferably regulated by the ion-mediated regulation of its degradation by the invertase, whereas for the high and continuous GG accumulation the ion-mediated activation and synthesis of GgpS is of greater importance in salt-shocked cells of Synechocystis (Figure 7).
Despite the active export, long-term salt-acclimated cells still have a higher steady state content of inorganic ions. For example, the intracellular Na+ concentration in Synechocystis increases from 32 mM in cells cultivated under NaCl-free conditions to 70 mM or even 215 mM when grown in media supplemented with 2 or 4% (w/v) additional NaCl (Hagemann et al., 1994). It can therefore be assumed that GghA is only slightly active due to these inhibitory ion concentrations in salt-adapted cells, whereas a sufficient part of the large GgpS pool is released from the inhibitory binding of DNA to synthesize GG (Figure 7).
Biotechnological Utilization of Salt-Induced Glucosylglycerol Accumulation
The interest in GG as a biotechnological product is based on its applicability in the pharmaceutical and cosmetic sector. It can serve as a stabilizer for the storage of enzymes and antibodies (Borges et al., 2002; Luley-Goedl et al., 2010). In skin care products, it is used as an additive due to its hydrating and revitalizing effect and marketed under the product name Glycoin® by the bitop AG. Presently, GG is commercially synthesized by the enzyme sucrose phosphorylase (from Leuconostoc mesenteroides), which converts the cheap precursors sucrose and glycerol via a transglycosylation reaction into GG and fructose (Luley-Goedl et al., 2010). However, it is conceivable to establish a GG production directly from cyanobacterial cultures, e.g., as byproduct from biocatalysts used for biofuel production in saline media. For example, optimized production strains of Synechocystis can produce approximately 981 mg GGL-1 (Tan et al., 2015). To achieve this high productivity, the genes ggtC and ggtD (uptake of GG) as well as ggpR (repressor of GG synthesis) were mutated. It has been previously shown that high GG yields can be obtained from wild type by a cyclic cultivation, during which the cultures are alternately exposed to salt stress for GG synthesis and hypoosmotic shocks that release the previously enriched GG (Reed et al., 1986; Erdmann et al., 1992). Using such a strategy and the optimized producer strain, it was possible to effectively harvest the produced GG in up to four consecutive production cycles (24 days in total) (Tan et al., 2015). Continuous release of GG without hypoosmotic shocks would require an effective exporter. A candidate gene for such a protein is ycaD from Stenotrophomonas rizophila (Roder et al., 2005; Alavi et al., 2013). Unfortunately, an exact characterization of the corresponding protein is still pending, but the expression of ycaD in GG-producing cyanobacteria may potentially lead to continuous export and high GG titers (Pade and Hagemann, 2015). Recently, a GG-producing Corynebacterium glutamicum strain has been engineered using the GG synthesis genes from Synechocystis, which continuously released GG into the culture medium and produced under optimized conditions more than 10 mM GG (approximately, 2 g L−1) (Roenneke et al., 2018). It has been further reported for Synechocystis that mutation of the slr1670 gene (later annotated as gghA) resulted in increased cellular GG accumulation (Savakis et al., 2016). In our own work, however, no significantly increased steady state concentration of GG could be confirmed in salt-adapted cultures of the ΔgghA mutant (Kirsch et al., 2017). Moreover, GG production has been also achieved with transgenic potato plants, which accumulated 10 μmol GGg-1 FM in leaves and up to 2.6 μmol GGg-1 FM in tubers (Sievers et al., 2013).
Biotechnological Use of Sucrose
With respect to biotechnology, salt-induced sucrose production has also moved into focus, because it is an attractive nutritive sugar and feedstock (Hagemann and Hess, 2018). The potential of cyanobacterial sucrose production has been studied to date with the organisms Synechococcus elongatus PCC 7942 (Ducat et al., 2012), Synechocystis (Du et al., 2013; Kirsch et al., 2018), and Synechococcus elongatus UTEX 2973 (Song et al., 2016). The expression of sucrose permease (sucrose/proton symporter, CscB) from E. coli proved to be very effective for sucrose production in both S. elongatus strains, PCC 7942 and UTEX 2973. In these freshwater strains, sucrose is the only compatible solute, and continuous loss via CscB resulted in a dramatic increase in sucrose synthesis when cultured in saline media (Ducat et al., 2012). Approximately, 35 mg sucrose L−1 h−1 were produced by strain Synechococcus CscB during cultivation at 200 mM NaCl. The authors performed a scaling up calculation of potential annual cyanobacterial sucrose production rates, which showed that cyanobacterial sucrose yields could be similar to those obtained with sugar cane (Ducat et al., 2012).
In Synechocystis, the sucrose content could be increased by the mutation of ggpS blocking GG synthesis and thereby enabling increased carbon flux in sucrose (Du et al., 2013; Kirsch et al., 2018). In addition, it has been shown that overexpression of the Sps also contributes to increasing sucrose synthesis (Du et al., 2013). As discussed above, the Δinv mutant of Synechocystis also has an increased sucrose content (Kirsch et al., 2018). It should be emphasized as a unique feature of this mutant that there are significant amounts of sucrose in the cells even under salt-free culture conditions. Based on this, the inactivation of the invertase was carried out in the strain WD25, which represented the most potent sucrose-producing Synechocystis strain (Du et al., 2013). Even with this genetic background, the inv mutation proved to be very effective, as in the resulting strain QL240, a 40% increase in sucrose production was observed compared to the parental WD25 (Kirsch et al., 2018). Thus, using the example of sucrose, the potential of genetic modifications to metabolically direct carbon into a desired product becomes apparent. However, it remains to be verified if sucrose from cyanobacterial production will be able to compete with traditionally obtained sucrose from sugar cane and sugar beet in future. Compared to the established method for obtaining sucrose from the plant material, a purification from cyanobacterial cultures appears to be labor-intensive and therefore cost-intensive, because sucrose must be separated from NaCl of the used saline growth medium. Hence, the future use of sucrose-producing cyanobacteria in co-cultivation with heterotrophic microorganisms seems more promising as has been also suggested by Ducat et al. (2012) and Löwe et al. (2017). Here, the sugar obtained from photosynthesis can serve as carbon and energy source for the synthesis of high value substances. The advantage of those systems is the elimination of downstream processing, i.e., a purification of sucrose from cells or the medium is not required. The work of Ducat et al. (2012) showed the positive effect of the sucrose-exporting S. elongatus PCC 7942 on the growth of yeast. Such a strain was also proven to be an effective co-cultivation partner for a polyhydroxyalkanoate-producing Pseudomonas putida strain (Löwe et al., 2017).
Outlook
Our knowledge on cyanobacterial GG and sucrose biosynthesis has been strongly improved during recent years, particularly the aspect of compatible solute degradation has come into focus of current research. The biochemical characterization of GG and sucrose synthesizing as well as degrading enzymes allowed us to propose a simple biochemical model to describe the ion-mediated accumulation and consumption of these compatible solutes in the model strain Synechocystis under hyperosmotic and hypoosmotic salt conditions, respectively (see Figure 7). Contrary to older views, labeling studies even revealed that compatible solutes are not accumulated as a dead end of metabolism but rather show a quick turnover–at least observed for the compatible solute GG in Synechococcus sp. PCC 7002 (Baran et al., 2017). Other studies indicated a close connection between sucrose and glycogen pools in salt-stressed cells of S. elongatus (Qiao et al., 2018). Nevertheless, one important question remains unanswered: to which extent can the knowledge obtained from Synechocystis or other well-investigated models transferred toward different cyanobacterial strains on the one hand and even toward other compatible solutes on the other hand? For example, differences in the relative importance of biochemical or transcriptional regulation of GG synthesis seem to exist between euryhaline strains or stenohaline such as marine strains adapted to constantly high salinities. Moreover, there are hints that similar to GgpS other enzyme activities are also stimulated in salt-stressed cells (e.g., Sps in Synechocystis). However, it is completely uncertain whether or not the model of inhibitory electrostatic interaction of GgpS with inhibitory DNA, which can be influenced by changed intracellular ion concentration (Novak et al., 2011) may play a more general role. Hence, it would be interesting to study a larger spectrum of cyanobacterial strains regarding GG as well as sucrose synthesis, for example to identifiy enzymes depending or not depending on salt for their activity, which would permit to analyze the structural basis for the salt-mediated enzyme regulation and also provide compatible solute synthesis enzymes that could be active in the absence of high NaCl amounts improving downstream processing.
In addition to the direct regulatory effects of ions on enzyme activities, the regulatory circuits involved in the sensing of salt stress and the signal transduction triggering gene expression programs are still not well understood. It has been proposed that instead of specific two-component systems or sigma factors, the changed ion and metabolic signature, e.g., accumulated K+ glutamate, might influence the specificity of promoter recognition by RNA polymerase (Gralla and Vargas, 2005). However, recently, the second messenger nucleotide cyclic-diAMP has been identified as potential signal molecule involved in the sensing of salt stress signals and crucial for K+ homeostasis (e.g., Nelson et al., 2013).
The closure of these knowledge gaps is not only of academic interest. As mentioned above, the increasing utilization of cyanobacteria for biotechnological applications will depend on mass cultivation of suitable strains in saline media to save limited freshwater resources. For sure, strain-specific salt acclimation modes and their regulatory circuits need to be revealed to minimize negative effects of stress acclimation on product formation. Moreover, there might be a broader spectrum of compatible solues that can be used for different pharmaceutical or other commercial purposes as high value byproducts of cyanobacterial mass cultivation. Hence, the selection of new cyanobacterial chassis organisms and the introduction of new biosynthetic routes for valuable bioproducts should always include an integrated study of salt acclimation and product synthesis under fluctuating salinities.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
The work on cyanobacterial salt tolerance of MH at the University of Rostock has been supported by grants of the DFG, BMBF, and Sino-German Research Center for many years.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ahmad, A. L., Mat Yasin, N. H., Derek, C. J. C., and Lim, J. K. (2011). Microalgae as a sustainable energy source for biodiesel production: a review. Renew. Sust. Energ. Rev. 15, 584–593. doi: 10.1016/j.rser.2010.09.018
Aikawa, S., Nishida, A., Ho, S. H., Chang, J. S., Hasunuma, T., and Kondo, A. (2014). Glycogen production for biofuels by the euryhaline cyanobacteria Synechococcus sp. strain PCC 7002 from an oceanic environment. Biotechnol. Biofuels 7:88. doi: 10.1186/1754-6834-7-88
Alavi, P., Starcher, M. R., Zachow, C., Müller, H., and Berg, G. (2013). Root-microbe systems: the effect and mode of interaction of stress protecting agent (SPA) Stenotrophomonas rhizophila DSM14405T. Front. Plant Sci. 4:141. doi: 10.3389/fpls.2013.00141
Angermayr, S. A., Hellingwerf, K. J., Lindblad, P., and de Mattos, M. J. (2009). Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 20, 257–263. doi: 10.1016/j.copbio.2009.05.011
Baran, R., Lau, R., Bowen, B. P., Diamond, S., Jose, N., Garcia-Pichel, F., et al.(2017). Extensive turnover of compatible solutes in cyanobacteria revealed by deuterium oxide (D2O) stable isotope probing. ACS Chem. Biol. 12, 674–681. doi: 10.1021/acschembio.6b00890
Blank, C. E. (2013). Phylogenetic distribution of compatible solute synthesis genes support a freshwater origin for cyanobacteria. J. Phycol. 49, 880–895. doi: 10.1111/jpy.12098
Blumwald, E., Mehlhorn, R. J., and Packer, L. (1983). Studies of osmoregulation in salt adaptation of cyanobacteria with ESR spin-probe techniques. Proc. Natl. Acad. Sci. USA 80, 2599–2602. doi: 10.1073/pnas.80.9.2599
Booth, I. R., and Louis, P. (1999). Managing hypoosmotic stress: aquaporins and mechanosensitive channels in Escherichia coli. Curr. Opin. Microbiol. 2, 166–169. doi: 10.1016/S1369-5274(99)80029-0
Borges, N., Ramos, A., Raven, N. D., Sharp, R. J., and Santos, H. (2002). Comparative study of the thermostabilizing properties of mannosylglycerate and other compatible solutes on model enzymes. Extremophiles 6, 209–216. doi: 10.1007/s007920100236
Borowitzka, L. J., Demmerle, S., Mackay, M. A., and Norton, R. S. (1980). Carbon-13 nuclear magnetic resonance study of osmoregulation in a blue-green alga. Science 210, 650–651. doi: 10.1126/science.210.4470.650
Boyle, E. A., Edmond, J. M., and Sholkovitz, E. R. (1977). The mechanism of iron removal in estuaries. Geochim. Cosmochim. Acta 41, 1313–1324. doi: 10.1016/0016-7037(77)90075-8
Brill, J., Hoffmann, T., Bleisteiner, M., and Bremer, E. (2011). Osmotically controlled synthesis of the compatible solute proline is critical for cellular defense of Bacillus subtilis against high osmolarity. J. Bacteriol. 193, 5335–5346. doi: 10.1128/JB.05490-11
Case, A. E., and Atsumi, S. (2016). Cyanobacterial chemical production. J. Biotechnol. 231, 106–114. doi: 10.1016/j.jbiotec.2016.05.023
Chaves, J. E., Kirst, H., and Melis, A. (2015). Isoprene production in Synechocystis under alkaline and saline growth conditions. J. Appl. Phycol. 27, 1089–1097. doi: 10.1007/s10811-014-0395-2
Chisti, Y. (2013). Constraints to commercialization of algal fuels. J. Biotechnol. 167, 201–214. doi: 10.1016/j.jbiotec.2013.07.020
Curatti, L., Folco, E., Desplats, P., Abratti, G., Limones, V., Herrera-Estrella, L., et al. (1998). Sucrose-phosphate synthase from Synechocystis sp. strain PCC 6803: identification of the spsA gene and characterization of the enzyme expressed in Escherichia coli. J. Bacteriol. 180, 6776–6779.
Curatti, L., Flores, E., and Salerno, G. (2002). Sucrose is involved in the diazotrophic metabolism of the heterocyst-forming cyanobacterium Anabaena sp. FEBS Lett. 513, 175–178. doi: 10.1016/S0014-5793(02)02283-4
Desplats, P., Folco, E., and Salerno, G. L. (2005). Sucrose may play an additional role to that of an osmolyte in Synechocystis sp. PCC 6803 salt-shocked cells. Plant Physiol. Biochem. 43, 133–138. doi: 10.1016/j.plaphy.2005.01.008
Dienst, D., Georg, J., Abts, T., Jakorew, L., Kuchmina, E., Börner, T., et al. (2014). Transcriptomic response to prolonged ethanol production in the cyanobacterium Synechocystis sp. PCC6803. Biotechnol. Biofuels 7:21. doi: 10.1186/1754-6834-7-21
Du, W., Liang, F., Duan, Y., Tan, X., and Lu, X. (2013). Exploring the photosynthetic production capacity of sucrose by cyanobacteria. Metab. Eng. 19, 17–25. doi: 10.1016/j.ymben.2013.05.001
Ducat, D. C., Way, J. C., and Silver, P. A. (2011). Engineering cyanobacteria to generate high-value products. Trends Biotechnol. 29, 95–103. doi: 10.1016/j.tibtech.2010.12.003
Ducat, D. C., Avelar-Rivas, J. A., Way, J. C., and Silver, P. A. (2012). Rerouting carbon flux to enhance photosynthetic productivity. Appl. Environ. Microbiol. 78, 2660–2668. doi: 10.1128/AEM.07901-11
Ehira, S., Kimura, S., Miyazaki, S., and Ohmori, M. (2014). Sucrose synthesis in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120 is controlled by the two-component response regulator OrrA. Appl. Environ. Microbiol. 80, 5672–5679. doi: 10.1128/AEM.01501-14
Engelbrecht, F., Marin, K., and Hagemann, M. (1999). Expression of the ggpS gene, involved in osmolyte synthesis in the marine cyanobacterium Synechococcus sp. strain PCC 7002, revealed regulatory differences to the freshwater strain Synechocystis sp. PCC 6803. Appl. Environ. Microbiol. 65, 4822–4829.
Erdmann, N. (1983). Organic osmoregulatory solutes in blue-green algae. Z. Pflanzenphysiol. 110, 147–155. doi: 10.1016/S0044-328X(83)80161-5
Erdmann, N., Zuther, E., and Abarzua, S. (1992). Comparative studies on the photoproduction of nonhydrogenous resources by cyanobacteria. Curr. Microbiol. 25, 83–87. doi: 10.1007/BF01570964
Flombaum, P., Gallegos, J. L., Gordillo, R. A., Rincón, J., Zabala, L. L., Jiao, N., et al. (2013). Present and future global distributions of the marine cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 110, 9824–9829. doi: 10.1073/pnas.1307701110
Foster, R. A., Kuypers, M. M. M., Vagner, T., Paerl, R. W., Musat, N., and Zehr, J. P. (2011). Nitrogen fixation and transfer in open ocean diatom–cyanobacterial symbioses. ISME J. 5, 1484–1493. doi: 10.1038/ismej.2011.26
Fulda, S., Hagemann, M., and Libbert, E. (1990). Release of glucosylglycerol from the cyanobacterium Synechocystis spec. SAG 92.79 by hypoosmotic shock. Arch. Microbiol. 153, 405–408. doi: 10.1007/BF00249013
Gao, Z., Zhao, H., Li, Z., Tan, T., and Lu, X. (2012). Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ. Sci. 5, 9857–9865. doi: 10.1039/C2EE22675H
Georg, J., Kostova, G., Vuorijoki, L., Schön, V., Kadowaki, T., Huokko, T., et al. (2017). Acclimation of oxygenic photosynthesis to iron starvation is controlled by the sRNA IsaR1. Curr. Biol. 27, 1425–1436. doi: 10.1016/j.cub.2017.04.010
Gralla, J. D., and Vargas, D. R. (2005). Potassium glutamate as transcriptional inhibitor during bacterial osmoregulation. EMBO J. 25, 1515–1521. doi: 10.1038/sj.emboj.7601041
Hagemann, M. (2011). Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 35, 87–123. doi: 10.1111/j.1574-6976.2010.00234.x
Hagemann, M. (2013). “Genomics of salt acclimation: synthesis of compatible solutes among cyanobacteria” in Genomics of cyanobacteria. Advances in botanical research. Vol. 65, eds. F. Chauvat and C. Cassier-Chauvat (Amsterdam, the Netherlands: Elsevier Ltd.), 27–55.
Hagemann, M., and Erdmann, N. (1994). Activation and pathway of GG synthesis in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 140, 1427–1431. doi: 10.1099/00221287-140-6-1427
Hagemann, M., and Hess, W. R. (2018). Systems and synthetic biology for the biotechnological application of cyanobacteria. Curr. Opin. Biotechnol. 49, 94–99. doi: 10.1016/j.copbio.2017.07.008
Hagemann, M., and Marin, K. (1999). Salt-induced sucrose accumulation is mediated by sucrose-phosphate-synthase in cyanobacteria. J. Plant Physiol. 155, 424–430. doi: 10.1016/S0176-1617(99)80126-6
Hagemann, M., and Pade, N. (2015). Heterosides – compatible solutes occurring in prokaryotic and eukaryotic phototrophs. Plant Biol. 17, 927–934. doi: 10.1111/plb.12350
Hagemann, M., Fulda, S., and Schubert, H. (1994). DNA, RNA, and protein synthesis in the cyanobacterium Synechocystis sp. PCC 6803 adapted to different salt concentrations. Curr. Microbiol. 28, 201–207. doi: 10.1007/BF01575962
Hagemann, M., Schoor, A., Jeanjean, R., Zuther, E., and Joset, F. (1997a). The stpA gene from Synechocystis sp. strain PCC 6803 encodes the glucosylglycerol-phosphate phosphatase involved in cyanobacterial osmotic response to salt shock. J. Bacteriol. 179, 1727–1733.
Hagemann, M., Richter, S., and Mikkat, S. (1997b). The ggtA gene encodes a subunit of the transport system for the osmoprotective compound glucosylglycerol in Synechocystis sp. strain PCC 6803. J. Bacteriol. 179, 714–720.
Hagemann, M., Effmert, U., Kerstan, T., Schoor, A., and Erdmann, N. (2001). Biochemical characterization of glucosylglycerolphosphate synthase of Synechocystis sp. strain PCC 6803: purification and overexpression change its salt dependence. Curr. Microbiol. 43, 278–283. doi: 10.1007/s002840010301
Hagemann, M., Ribbeck-Busch, K., Klähn, S., Hasse, D., Steinbruch, R., and Berg, G. (2008). The plant-associated bacterium Stenotrophomonas rhizophila expresses a new enzyme for the synthesis of the compatible solute glucosylglycerol. J. Bacteriol. 190, 5898–5906. doi: 10.1128/JB.00643-08
Hecker, M., Pane-Farre, J., and Völker, U. (2007). SigB-dependent general stress response in Bacillus subtilis and related gram-positive bacteria. Annu. Rev. Microbiol. 61, 215–236. doi: 10.1146/annurev.micro.61.080706.093445
Hincha, D. K., and Hagemann, M. (2004). Stabilization of model membranes during drying by compatible solutes involved in the stress tolerance of plants and microorganisms. Biochem. J. 383, 277–283. doi: 10.1042/BJ20040746
Hoffmann, T., and Bremer, E. (2016). “Management of osmotic stress by Bacillus subtilis: genetics and physiology” in Stress and environmental regulation of gene expression and adaptation in bacteria. ed. F. J. de Bruijn (London, UK: John Wiley & Sons), 657–676.
Hohmann-Marriott, M. F., and Blankenship, R. E. (2011). Evolution of photosynthesis. Annu. Rev. Plant Biol. 62, 515–548. doi: 10.1146/annurev-arplant-042110-103811
Huckauf, J., Nomura, C., Forchhammer, K., and Hagemann, M. (2000). Stress responses of Synechocystis sp. strain PCC 6803 mutants impaired in genes encoding putative alternative sigma factors. Microbiology 146, 2877–2889. doi: 10.1099/00221287-146-11-2877
Jung, K., and Altendorf, K. (2002). Towards an understanding of the molecular mechanisms of stimulus perception and signal transduction by the KdpD/KdpE system of Escherichia coli. J. Mol. Microbiol. Biotechnol. 4, 223–228.
Kirsch, F., Pade, N., Klähn, S., Hess, W. R., and Hagemann, M. (2017). The glucosylglycerol-degrading enzyme GghA is involved in acclimation to fluctuating salinities by the cyanobacterium Synechocystis sp. strain PCC 6803. Microbiology 163, 1319–1328. doi: 10.1099/mic.0.000518
Kirsch, F., Luo, Q., Lu, X., and Hagemann, M. (2018). Inactivation of invertase enhances sucrose production in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 164, 1220–1228. doi: 10.1099/mic.0.000708
Kizawa, A., Kawahara, A., Takimura, Y., Nishiyama, Y., and Hihara, Y. (2016). RNA-seq profiling reveals novel target genes of LexA in the cyanobacterium Synechocystis sp. PCC 6803. Front. Microbiol. 7, 193.1–193.14. doi: 10.3389/fmicb.2016.00193
Klähn, S., and Hagemann, M. (2011). Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 13, 551–562. doi: 10.1111/j.1462-2920.2010.02366.x
Klähn, S., Steglich, C., Hess, W. R., and Hagemann, M. (2010a). Glucosylglycerate: a secondary compatible solute common to marine cyanobacteria from nitrogen-poor environments. Environ. Microbiol. 12, 83–94. doi: 10.1111/j.1462-2920.2009.02045.x
Klähn, S., Höhne, A., Simon, E., and Hagemann, M. (2010b). The gene ssl3076 encodes a protein mediating the salt-induced expression of ggpS for the biosynthesis of the compatible solute glucosylglycerol in Synechocystis sp. strain PCC 6803. J. Bacteriol. 192, 4403–4012. doi: 10.1128/JB.00481-10
Kollman, V. H., Hanners, L. J., London, R. E., Adame, E. G., and Walker, T. E. (1979). Photosynthetic preparation and characterization of 13C-labeled carbohydrates in Agmenellum quadruplicatum. Carbohydr. Res. 73, 193–202.
Kolman, M. A., Nishi, C. N., Perez-Cenci, M., and Salerno, G. L. (2015). Sucrose in cyanobacteria: from a salt-response molecule to play a key role in nitrogen fixation. Life 5, 102–126. doi: 10.3390/life5010102
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Ladas, N. P., and Papageorgiou, G. C. (2000). Cell turgor: a critical factor for the proliferation of cyanobacteria at unfavorable salinity. Photosynth. Res. 65, 155–164. doi: 10.1023/A:1006423221150
Levina, N., Totemeyer, S., Stokes, N. R., Louis, P., Jones, M. A., and Booth, I. R. (1999). Protection of Escherichia coli cells against extreme turgor by activation of MscS and MscL mechanosensitive channels: identification of genes required for MscS activity. EMBO J. 18, 1730–1737. doi: 10.1093/emboj/18.7.1730
Los, D. A., Zorina, A., Sinetova, M., Kryazhov, S., Mironov, K., and Zinchenko, V. V. (2010). Stress sensors and signal transducers in cyanobacteria. Sensors 10, 2386–2415. doi: 10.3390/s100302386
Löwe, H., Hobmeier, K., Moos, M., Kremling, A., and Pflüger-Grau, K. (2017). Photoautotrophic production of polyhydroxyalkanoates in a synthetic mixed culture of Synechococcus elongatus cscB and Pseudomonas putida cscAB. Biotechnol. Biofuels 10:190. doi: 10.1186/s13068-017-0875-0
Luley-Goedl, C., Sawangwan, T., Mueller, M., Schwarz, A., and Nidetzky, B. (2010). Biocatalytic process for production of α-glucosylglycerol using sucrose phosphorylase. Food Technol. Biotechnol. 48, 276–283. ISSN 1330-9862.
Lunn, J. E. (2002). Evolution of sucrose synthesis. Plant Physiol. 128, 1490–1500. doi: 10.1104/pp.010898
Lyons, T. W., Reinhard, C. T., and Planavsky, N. J. (2014). The rise of oxygen in Earth’s early ocean and atmosphere. Nature 506, 307–315. doi: 10.1038/nature13068
Mackay, M. A., Norton, R. S., and Borowitzka, L. J. (1984). Organic osmoregulatory solutes in cyanobacteria. J. Gen. Microbiol. 130, 2177–2191.
Marin, K., Zuther, E., Kerstan, T., Kunert, A., and Hagemann, M. (1998). The ggpS gene from Synechocystis sp. strain PCC 6803 encoding glucosyl-glycerol-phosphate synthase is involved in osmolyte synthesis. J. Bacteriol. 180, 4843–4849.
Marin, K., Huckauf, J., Fulda, S., and Hagemann, M. (2002). Salt-dependent expression of glucosylglycerol-phosphate synthase, involved in osmolyte synthesis in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 184, 2870–2877. doi: 10.1128/JB.184.11.2870-2877.2002
Marin, K., Suzuki, I., Yamaguchi, K., Ribbeck, K., Yamamoto, H., Kanesaki, Y., et al. (2003). Identification of histidine kinases that act as sensors in the perception of salt stress in Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 100, 9061–9066. doi: 10.1073/pnas.1532302100
Marin, K., Kanesaki, Y., Los, D. A., Murata, N., Suzuki, I., and Hagemann, M. (2004). Gene expression profiling reflects physiological processes in salt acclimation of Synechocystis sp. strain PCC 6803. Plant Physiol. 136, 3290–3300. doi: 10.1104/pp.104.045047
Martin, W. F., Garg, S., and Zimorski, V. (2015). Endosymbiotic theories for eukaryote origin. Philos. Trans. R Soc. Lond. B Biol. Sci. 370:20140330. doi: 10.1098/rstb.2014.0330
Martinez-Noel, G., Cumino, A. C., Kolman, M. A., and Salerno, G. L. (2013). First evidence of sucrose biosynthesis by single cyanobacterial bimodular proteins. FEBS Lett. 587, 1669–1674. doi: 10.1016/j.febslet.2013.04.012
Mata, T. M., Martins, A. A., and Caetano, N. S. (2010). Microalgae for biodiesel production and other applications: a review. Renew. Sust. Energ. Rev. 14, 217–232. doi: 10.1016/j.rser.2009.07.020
Mikkat, S., and Hagemann, M. (2000). Molecular analysis of the ggtBCD gene cluster of Synechocystis sp. strain PCC6803 encoding subunits of an ABC transporter for osmoprotective compounds. Arch. Microbiol. 174, 273–282. doi: 10.1007/s002030000201
Mikkat, S., Effmert, U., and Hagemann, M. (1997). Uptake and use of the osmoprotective compounds trehalose, glucosylglycerol, and sucrose by the cyanobacterium Synechocystis sp. PCC 6803. Arch. Microbiol. 167, 112–118. doi: 10.1007/s002030050423
Mizuno, T., and Mizushima, S. (1990). Signal transduction and gene regulation through the phosphorylation of two regulatory components: the molecular basis for the osmotic regulation of the porin genes. Mol. Microbiol. 4, 1077–1082. doi: 10.1111/j.1365-2958.1990.tb00681.x
Moezelaar, R., Bijvank, S. M., and Stal, L. J. (1996). Fermentation and sulfur reduction in the mat-building cyanobacterium Microcoleus chthonoplastes. Appl. Environ. Microbiol. 62, 1752–1758.
Moses, S., Sinner, T., Zaprasis, A., Stöveken, N., Hoffmann, T., Belitsky, B. R., et al. (2012). Proline utilization by Bacillus subtilis: uptake and catabolism. J. Bacteriol. 194, 745–758. doi: 10.1128/JB.06380-11
Nadeem, H., Rashid, M. H., Siddique, M. H., Azeem, F., Muzammil, S., Javed, M. R., et al. (2015). Microbial invertases: a review on kinetics, thermodynamics, physiochemical properties. Process Biochem. 50, 1202–1210. doi: 10.1016/j.procbio.2015.04.015
Nanatani, K., Shijuku, T., Akai, M., Yukutake, Y., Yasui, M., Hamamoto, S., et al. (2013). Characterization of the role of a mechanosensitive channel in osmotic down shock adaptation in Synechocystis sp. PCC 6803. Channels 7, 238–242. doi: 10.4161/chan.25350
Nelson, J. W., Sudarsan, N., Furukawa, K., Weinberg, Z., Wang, J. X., and Breaker, R. R. (2013). Riboswitches in eubacteria sense the second messenger c-di-AMP. Nat. Chem. Biol. 9, 834–839. doi: 10.1038/nchembio.1363
Nihira, T., Saito, Y., Ohtsubo, K., Nakai, H., and Kitaoka, M. (2014). 2-O-α-D-glucosylglycerol phosphorylase from Bacillus selenitireducens MLS10 possessing hydrolytic activity on β-D-glucose 1-phosphate. PLoS One 9:86548. doi: 10.1371/journal.pone.0086548
Novak, J. F., Stirnberg, M., Roenneke, B., and Marin, K. (2011). A novel mechanism of osmosensing, a salt-dependent protein-nucleic acid interaction in the cyanobacterium Synechocystis species PCC 6803. J. Biol. Chem. 286, 3235–3241. doi: 10.1074/jbc.M110.157032
Pade, N., and Hagemann, M. (2015). Salt acclimation of cyanobacteria and their application in biotechnology. Life 5, 25–49. doi: 10.3390/life5010025
Pade, N., Compaoré, J., Klähn, S., Stal, L. J., and Hagemann, M. (2012). The marine cyanobacterium Crocosphaera watsonii WH8501 synthesizes the compatible solute trehalose by a laterally acquired OtsAB fusion protein. Environ. Microbiol. 14, 1261–1271. doi: 10.1111/j.1462-2920.2012.02709.x
Pade, N., Michalik, D., Ruth, W., Belkin, N., Hess, W. R., Berman-Frank, I., et al. (2016a). Trimethylated homoserine functions as the major compatible solute in the globally significant oceanic cyanobacterium Trichodesmium. Proc. Natl. Acad. Sci. USA 113, 13191–13196. doi: 10.1073/pnas.1611666113
Pade, N., Erdmann, S., Enke, H., et al. (2016b). Insights into isoprene production using the cyanobacterium Synechocystis sp. PCC 6803. Biotechnol. Biofuels 9:89. doi: 10.1186/s13068-016-0503-4
Pade, N., Mikkat, S., and Hagemann, M. (2017). Ethanol, glycogen and glucosylglycerol represent competing carbon pools in ethanol-producing cells of Synechocystis sp. PCC 6803 under high-salt conditions. Microbiology 163, 300–307. doi: 10.1099/mic.0.000433
Pawlowicz, R. (2013). Key physical variables in the ocean: temperature, salinity, and density. Nat. Educ. Knowl. 4:13
Pereira, S., Zille, A., Micheletti, E., Moradas-Ferreira, P., De Philippis, R., and Tamagnini, P. (2009). Complexity of cyanobacterial exopolysaccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 33, 917–941. doi: 10.1111/j.1574-6976.2009.00183.x
Perez-Cenci, M., and Salerno, G. L. (2014). Functional characterization of Synechococcus amylosucrase and fructokinase encoding genes discovers two novel actors on the stage of cyanobacterial sucrose metabolism. Plant Sci. 224, 95–102. doi: 10.1016/j.plantsci.2014.04.003
Pivetti, C. D., Yen, M. R., Miller, S., Busch, W., Tseng, Y. H., Booth, I. R., et al. (2003). Two families of mechanosensitive channel proteins. Microbiol. Mol. Biol. Rev. 67, 66–85. doi: 10.1128/MMBR.67.1.66-85.2003
Pocard, J. A., Smith, L. T., Smith, G. M., and Le Rudulier, D. (1994). A prominent role for glucosylglycerol in the adaptation of Pseudomonas mendocina SKB70 to osmotic stress. J. Bacteriol. 176, 6877–6884. doi: 10.1128/jb.176.22.6877-6884.1994
Pollari, M., Gunnelius, L., Tuominen, I., Ruotsalainen, V., Tyystjärvi, E., Salminen, T., et al. (2008). Characterization of single and double inactivation strains reveals new physiological roles for group 2 sigma factors in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 147, 1994–2005. doi: 10.1104/pp.108.122713
Porchia, A. C., and Salerno, G. L. (1996). Sucrose biosynthesis in a prokaryotic organism: presence of two sucrose-phosphate synthases in Anabaena with remarkable differences compared with the plant enzymes. Proc. Natl. Acad. Sci. USA 93, 13600–13604. doi: 10.1073/pnas.93.24.13600
Porchia, A. C., Curatti, L., and Salerno, G. L. (1999). Sucrose metabolism in cyanobacteria: sucrose synthase from Anabaena sp. strain PCC 7119 is remarkably different from the plant enzymes with respect to substrate affinity and amino-terminal sequence. Planta 210, 34–40. doi: 10.1007/s004250050651
Posten, C. (2009). Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 9, 165–177. doi: 10.1002/elsc.200900003
Qiao, C., Duan, Y., Zhang, M., Hagemann, M., Luo, Q., and Lu, X. (2018). Effects of lowered and enhanced glycogen pools on salt-induced sucrose production in a sucrose-secreting strain of Synechococcus elongatus PCC 7942. Appl. Environ. Microbiol. 84, e02023–e02017. doi: 10.1128/AEM.02023-17
Reed, R. H., and Stewart, W. D. P. (1985). Osmotic adjustment and organic solute accumulation in unicellular cyanobacteria from freshwater and marine habitats. Mar. Biol. 88, 1–9. doi: 10.1007/BF00393037
Reed, R. H., Warr, S. R. C., Richardson, D. L., Moore, D. J., and Stewart, W. D. P. (1985). Multiphasic osmotic adjustment in a euryhaline cyanobacterium. FEMS Microbiol. Lett. 28, 225–229. doi: 10.1111/j.1574-6968.1985.tb00796.x
Reed, R. H., Warr, S. R. C., Kerby, N. W., and Stewart, W. D. P. (1986). Osmotic shock-induced release of low molecular weight metabolites from free-living and immobilized cyanobacteria. Enzym. Microb. Technol. 8, 101–104. doi: 10.1016/0141-0229(86)90080-3
Roder, A., Hoffmann, E., Hagemann, M., and Berg, G. (2005). Synthesis of the compatible solutes glucosylglycerol and trehalose by salt-stressed cells of Stenotrophomonas strains. FEMS Microbiol. Lett. 243, 219–226. doi: 10.1016/j.femsle.2004.12.005
Roenneke, B., Rosenfeldt, N., Derya, S. M., Novak, J. F., Marin, K., Krämer, R., et al. (2018). Production of the compatible solute α-D-glucosylglycerol by metabolically engineered Corynebacterium glutamicum. Microb. Cell Factories 17:94. doi: 10.1186/s12934-018-0939-2
Rübsam, H., Kirsch, F., Reimann, V., Erban, A., Kopka, J., Hagemann, M., et al. (2018). The iron-stress activated RNA 1 (IsaR1) coordinates osmotic acclimation and iron starvation responses in the cyanobacterium Synechocystis sp. PCC 6803. Environ. Microbiol. 20, 2757–2768. doi: 10.1111/1462-2920.14079
Salerno, G. L., and Curatti, L. (2003). Origin of sucrose metabolism in higher plants: when, how and why? Trends Plant Sci. 8, 63–69. doi: 10.1016/S1360-1385(02)00029-8
Savakis, P., and Hellingwerf, K. J. (2015). Engineering cyanobacteria for direct biofuel production from CO2. Curr. Opin. Biotechnol. 33, 8–14. doi: 10.1016/j.copbio.2014.09.007
Savakis, P., Tan, X., Qiao, C., Song, K., Lu, X., Hellingwerf, K. J., et al. (2016). Slr1670 from Synechocystis sp. PCC 6803 is required for the re-assimilation of the osmolyte glucosylglycerol. Front. Microbiol. 7:1350. doi: 10.3389/fmicb.2016.01350
Scanlan, D. J., Ostrowski, M., Mazard, S., Dufresne, A., Garczarek, L., Hess, W. R., et al. (2009). Ecological genomics of marine picocyanobacteria. Microbiol. Mol. Biol. Rev. 73, 249–299. doi: 10.1128/MMBR.00035-08
Schulz, A., Stöveken, N., Binzen, I. M., Hoffmann, T., Heider, J., and Bremer, E. (2017). Feeding on compatible solutes: a substrate-induced pathway for uptake and catabolism of ectoines and ist genetic control by EnuR. Environ. Microbiol. 19, 926–946. doi: 10.1111/1462-2920.13414
Shibl, A. A., Ngugi, D. K., Talarmin, A., Thompson, L. R., Blom, J., and Stingl, U. (2018). The genome of a novel isolate of Prochlorococcus from the Red Sea contains transcribed genes for compatible solute biosynthesis. FEMS Microbiol. Ecol. 94:fiy182. doi: 10.1093/femsec/fiy182
Shoumskaya, M. A., Paithoonrangsarid, K., Kanesaki, Y., Los, D. A., Zinchenko, V. V., Tanticharoen, M., et al. (2005). Identical Hik-Rre systems are involved in perception and transduction of salt signals and hyperosmotic signals but regulate the expression of individual genes to different extents in Synechocystis. J. Biol. Chem. 280, 21531–21538. doi: 10.1074/jbc.M412174200
Sievers, N., Muders, K., Henneberg, M., Klähn, S., Effmert, M., Junghans, H., et al. (2013). Establishing glucosylglycerol synthesis in potato (Solanum tuberosum L. cv. Albatros) by expression of the ggpPS gene from Azotobacter vinelandii. J. Plant Sci. Mol. Breed. doi: 10.7243/2050-2389-2-1
Singh, R., Parihar, P., Singh, M., Bajguz, A., Kumar, J., Singh, S., et al. (2017). Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: current status and future prospects. Front. Microbiol. 8:515. doi: 10.3389/fmicb.2017.00515
Song, K., Tan, X., Liang, Y., and Lu, X. (2016). The potential of Synechococcus elongatus UTEX 2973 for sugar feedstock production. Appl. Microbiol. Biotechnol. 100, 7865–7875. doi: 10.1007/s00253-016-7510-z
Song, K., Hagemann, M., Tan, X., and Lu, X. (2017). The response regulator Slr1588 regulates spsA but is not crucial for salt acclimation of Synechocystis sp. PCC 6803. Front. Microbiol. 8:1176. doi: 10.3389/fmicb.2017.01176
Soo, R. M., Hemp, J., Parks, D. H., Fischer, W. W., and Hugenholtz, P. (2017). On the origins of oxygenic photosynthesis and aerobic respiration in cyanobacteria. Science 355, 1436–1440. doi: 10.1126/science.aal3794
Sturm, A. (1999). Invertases. Primary structures, functions and roles in plant development and sucrose partitioning. Plant Physiol. 121, 1–7. doi: 10.1104/pp.121.1.1
Tan, X., Du, W., and Lu, X. (2015). Photosynthetic and extracellular production of glucosylglycerol by genetically engineered and gel-encapsulated cyanobacteria. Appl. Microbiol. Biotechnol. 99, 2147–2154. doi: 10.1007/s00253-014-6273-7
Tandeau de Marsac, N. T., and Houmard, J. (1993). Adaptation of cyanobacteria to environmental stimuli: new steps toward molecular mechanisms. FEMS Microbiol. Rev. 104, 119–190.
Vargas, W., Cumino, A., and Salerno, G. L. (2003). Cyanobacterial alkaline/neutral invertases. Origin of sucrose hydrolysis in the plant cytosol? Planta 216, 951–960. doi: 10.1007/s00425-002-0943-x
Vargas, W. A., and Salerno, G. L. (2010). The Cinderella story of sucrose hydrolysis: alkaline/neutral Invertases, from cyanobacteria to unforeseen roles in plant cytosol and organelles. Plant Sci. 178, 1–8. doi: 10.1016/j.plantsci.2009.09.015
Wargo, M. J. (2013). Homeostasis and catabolism of choline and glycine betaine: lessons from Pseudomonas aeruginosa. Appl. Environ. Microbiol. 79, 2112–2120. doi: 10.1128/AEM.03565-12
Welsh, D. T. (2000). Ecological significance of compatible solute accumulation by microorganisms: from single cells to global climate. FEMS Microbiol. Rev. 24, 263–290. doi: 10.1111/j.1574-6976.2000.tb00542.x
Xie, J., Cai, K., Hu, H. X., Jiang, Y. L., Yang, F., Hu, P. F., et al. (2016). Structural analysis of the catalytic mechanism and substrate specificity of Anabaena alkaline invertase InvA reveals a novel glucosidase. J. Biol. Chem. 291, 25667–25677. doi: 10.1074/jbc.M116.759290
Keywords: cyanobacteria, compatible solute, salt stress, sucrose, glucosylglycerol, glucosylglycerol-phosphate synthase/phosphatase, sucrose phosphate synthase
Citation: Kirsch F, Klähn S and Hagemann M (2019) Salt-Regulated Accumulation of the Compatible Solutes Sucrose and Glucosylglycerol in Cyanobacteria and Its Biotechnological Potential. Front. Microbiol. 10:2139. doi: 10.3389/fmicb.2019.02139
Edited by:
Wendy Schluchter, University of New Orleans, United StatesReviewed by:
Klaas J. Jan Hellingwerf, University of Amsterdam, NetherlandsErhard Bremer, University of Marburg, Germany
Copyright © 2019 Kirsch, Klähn and Hagemann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Hagemann, bWFydGluLmhhZ2VtYW5uQHVuaS1yb3N0b2NrLmRl