 Francisco Arenas-Huertero1*
Francisco Arenas-Huertero1* Montserrat Zaragoza-Ojeda1
Montserrat Zaragoza-Ojeda1 Juana Sánchez-Alarcón2
Juana Sánchez-Alarcón2 Mirta Milić3
Mirta Milić3 Maja Šegvić Klarić4
Maja Šegvić Klarić4 José M. Montiel-González2
José M. Montiel-González2 Rafael Valencia-Quintana2*
Rafael Valencia-Quintana2*- 1Experimental Pathology Research Laboratory, Children’s Hospital of Mexico Federico Gómez, Mexico, Mexico
- 2Rafael Villalobos-Pietrini Laboratory of Genomic Toxicology and Environmental Chemistry, Faculty of Agrobiology, Autonomous University of Tlaxcala, Tlaxcala, Mexico
- 3Mutagenesis Unit, Institute for Medical Research and Occupational Health, Zagreb, Croatia
- 4Department of Microbiology, Faculty of Pharmacy and Biochemistry, University of Zagreb, Zagreb, Croatia
The purpose of this review is to present information about the role of activation of aflatoxins and other mycotoxins, of the aryl hydrocarbon receptor (AhR) pathway. Aflatoxins and other mycotoxins are a diverse group of secondary metabolites that can be contaminants in a broad range of agricultural products and feeds. Some species of Aspergillus, Alternaria, Penicilium, and Fusarium are major producers of mycotoxins, some of which are toxic and carcinogenic. Several aflatoxins are planar molecules that can activate the AhR. AhR participates in the detoxification of several xenobiotic substances and activates phase I and phase II detoxification pathways. But it is important to recognize that AhR activation also affects differentiation, cell adhesion, proliferation, and immune response among others. Any examination of the effects of aflatoxins and other toxins that act as activators to AhR must consider the potential of the disruption of several cellular functions in order to extend the perception thus far about the toxic and carcinogenic effects of these toxins. There have been no Reviews of existing data between the relation of AhR and aflatoxins and this one attempts to give information precisely about this dichotomy.
Introduction
Aflatoxins and other mycotoxin are activators of the aryl hydrocarbon receptor (AhR), are a structurally diverse group of fungal secondary metabolites and a toxigenically and chemically heterogeneous assemblage. They are ubiquitous contaminants in a broad range of agricultural commodities and feed (Table 1) and can contaminate the food supply at any time during production, processing, transport or storage (Bräse et al., 2009). Currently, far more than 400 mycotoxins are produced by some 350 fungi species (Kuhn and Ghannoum, 2003), but scientific attention is given mainly to those that have proven to be carcinogenic and/or toxic (Zain, 2011). The aflatoxins and other toxins that can act as activators to AhR can be acutely or chronically toxic, depending on the kind of toxin, the dose, health, age and nutritional status of the exposed individual or animal, and the possible synergistic effects between mycotoxins (Bennett and Klich, 2003). Their ingestion, inhalation, or skin absorption can cause diseases in humans and animals and their greater impact on human health in industrial countries is due to the chronic exposure (Gil-Serna et al., 2014). In this case, aflatoxins are associated with different biological effects like carcinogenic, mutagenic, teratogenic, estrogenic, hemorrhagic, immunotoxic, nephrotoxic, hepatotoxic, dermotoxic and neurotoxic ones, including displays of both tumor and antitumor effect, cytotoxic, and antimicrobial properties (see Table 1). Their impact in human diseases has been the subject of intensive study since the early 1960s, since aflatoxicosis and other mycotoxicosis can provoke tumor formation or even rapid death (Bräse et al., 2009).
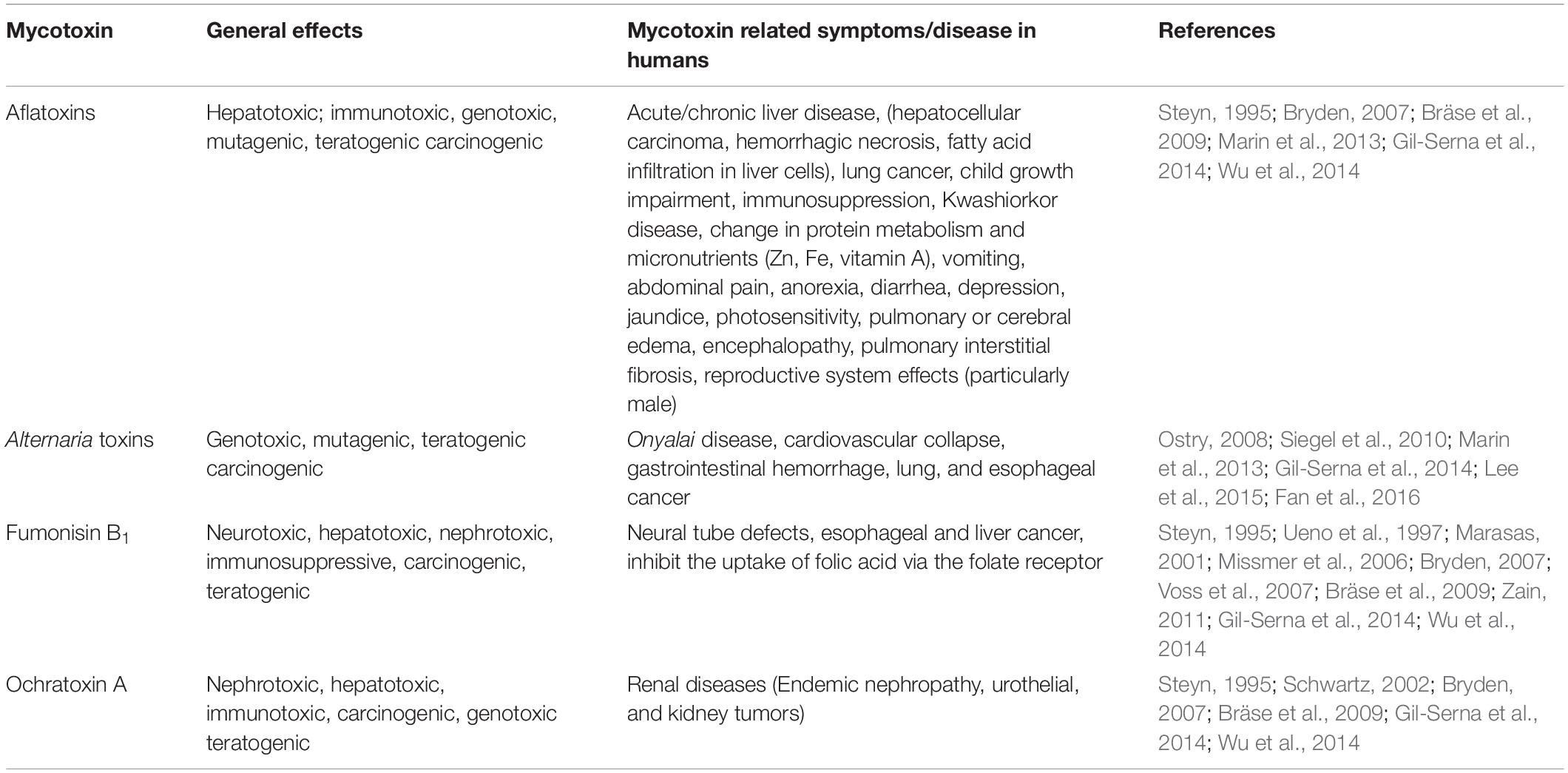
Table 1. Toxic effects of several mycotoxins in human.
Some molds are capable of producing more than one toxin and some toxins are produced by more than one fungal species (Bräse et al., 2009; Zain, 2011). Aflatoxins are commonly found in many food products, such as cereals, fruits, nuts, and meat, among others.
Interactions of Aflatoxins and Other Foodborne Mycotoxins With AhR
The major foodborne mycotoxins of public health interest are aflatoxins. However, there are several foodborne mycotoxins (Alternaria toxins, fumonisin B1, ochratoxin A, and patulin) that should not be excluded due to their broad toxicity, as well as ability to activate AhR-pathway, the focus of this review. These mycotoxins account annually for millions of dollars losses worldwide in human health, animal health and discarded agricultural products (Zain, 2011).
Aflatoxins
The name “aflatoxin” (A. flavus toxin) was assigned to a number of polyketide-derived furanocoumarins carcinogenic metabolites produced mainly by fungi Aspergillus flavus and Aspergillus parasiticus (Kensler et al., 2011). The aflatoxins were structurally identified in the early 1960s and over the last 50 years have been extensively studied with respect to their mechanism of action, including their mutagenic and carcinogenic activity (Valencia-Quintana et al., 2014). Collectively, aflatoxins are a group of approximately 20 chemically related metabolites, although the most important and naturally occurring are aflatoxins B1 (AFB1), B2, G1, and G2 (Figure 1). The AFB1 is in this group the most potent hepatotoxin with highly toxic and carcinogenic characteristics for many animal species (Hendricks, 1994).
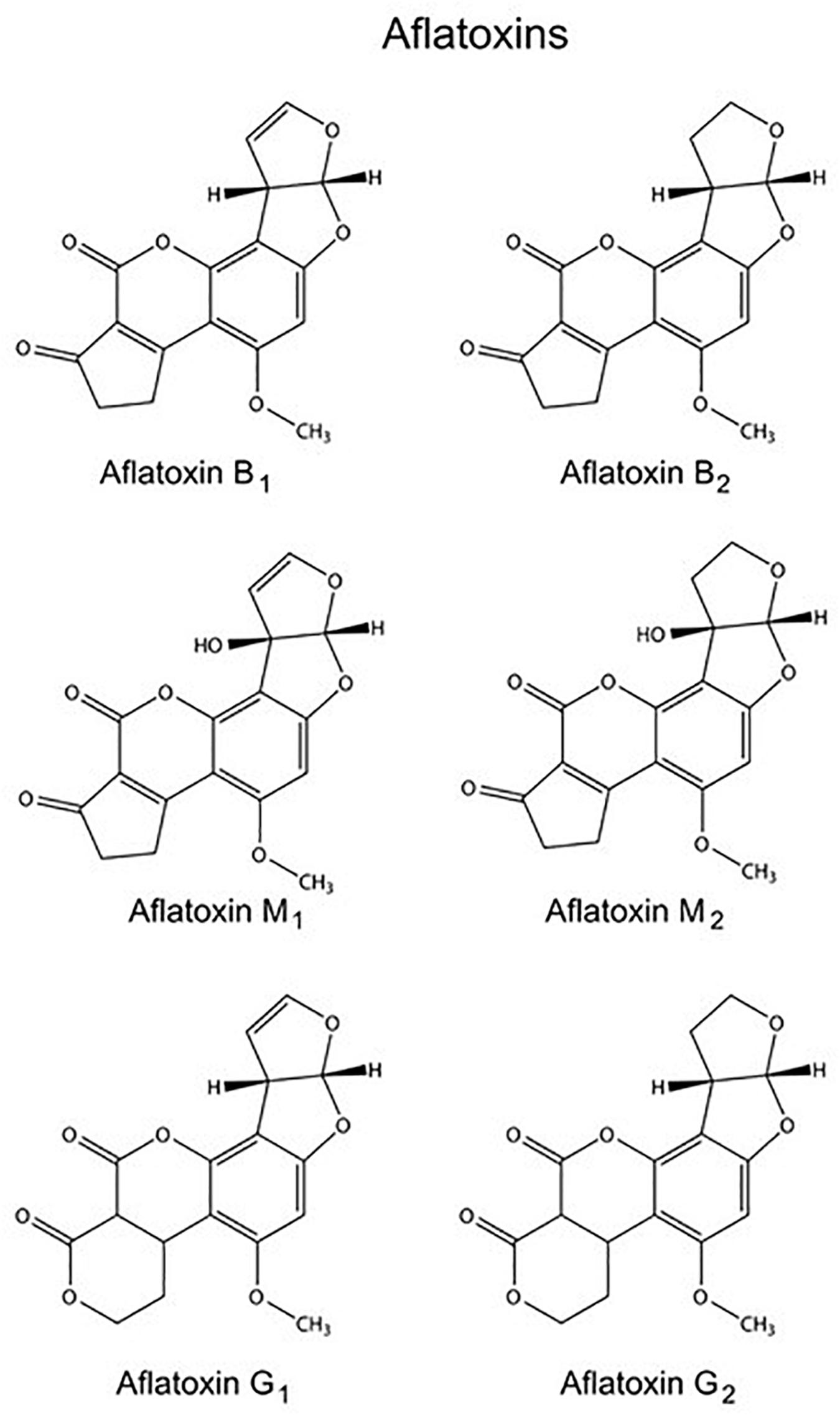
Figure 1. Molecular structures of several aflatoxins.
Aflatoxins are metabolically activated in the liver by major CYP isoenzymes as CYP1A1, CYP1A2, and CYP3A4, their biotransformation produces a highly reactive AFB1-8,9-exo-epoxide and 8,9-endo-epoxide that can form adducts with DNA and RNA. In addition, CYP1A1 which also participates in AFB1 transformation into epoxide, represents the highest fraction of extrahepatic CYP. After binding to DNA, the predominant 8,9-dihydro-8-(N7-guanyl)-9-hydroxy AFB1 (AFB1-N7-Gua) adduct is formed. AFB1-N7-Gua may be converted to two secondary lesions, the one with apurinic site and the other with a more stable ring opened AFB1-formamidopyrimidine (AFB1-FAPY) adduct; the latter far more persistent in vivo than AFB1-N7-Gua. Sufficient evidence from animal and epidemiological studies, such as the demonstration of a specific mutation in the TP53 gene, led to the AFB1 classification as a human carcinogen by the International Agency for Research on Cancer (IARC) (Hendricks, 1994; IARC, 2002; Krska et al., 2008; Valencia-Quintana et al., 2014). Among mycotoxicosis described in humans, aflatoxicosis is of the greatest concern; repetitive aflatoxin outbreaks were reported in recent years in Kenya, India, and Malaysia, often accompanied by fumonisins (Shephard, 2004; Lewis et al., 2005). In acute mycotoxication, it can cause impaired growth in children, vomiting, abdominal pain, pulmonary edema and liver necrosis (Valencia-Quintana et al., 2012).
Because of its planar structure, it could act as activators for the AhR, at the moment there is not evidence to prove that any mycotoxin is an agonist of AhR. That was first seen in H4IIE cells; AFB1 induced an increase in CYP1A activity and CYP1A transcription, which was associated with an enhanced AhR activity, suggesting AhR pathway activation as a toxicity mechanism of AFB1 (Mary et al., 2015). The major human cytochrome P450 (CYP) enzymes involved in aflatoxin metabolism are CYP1A1, CYP3A4, 3A5, 3A7, and 1A2 (Tan et al., 2004; Marin et al., 2013). The CYP enzymes activation is an indirect evidence of activating AhR pathway. It is known that AFB1 causes carcinogenic, mutagenic and teratogenicity effects (Krska et al., 2008; Zhang et al., 2015), as well as immunotoxicity, hepatotoxicity and even death in farm animals and humans (Table 1). The AFB1 causes micronucleus (MN), sister chromatid exchanges (SCE), unscheduled DNA synthesis, and chromosomal strand breaks as well as adducts in rodent and human cells (Turkez and Geyikoglu, 2010). The mechanism of AFB1 cellular damage has not been completely elucidated. Reactive oxygen species (ROS) and lipid peroxidation (LPO) have been considered to be an important mechanisms in the toxicity. The ROS and LPO are created via AhR activation and its target genes as several CYPs, already mentioned above. In the following sections, this review will describe how the AhR pathway participates via cytochrome activation and the ROS production.
Alternaria Toxins
The Alternaria genus is able to produce different mycotoxins, A. alternata is considered to be the main mycotoxin producer. Alternaria mycotoxins include a small percentage of more than 70 phytotoxins, and only few have been physicochemically characterized (Dall’Asta et al., 2014). They belong to three different structural groups: the dibenzopyrone derivatives, alternariol (AOH), alternariol monomethyl ether (AME), and altenuene (ALT); the perylene derivatives altertoxins (ATX-I, -II, -III); and the tetramic acid derivatives, tenuazonic acid (TeA) and iso-tenuazonic acid (iso-TeA) (Schrader et al., 2001; Ostry, 2008). The chemical structures of these mycotoxins are presented in Figure 2 (Zain, 2011).
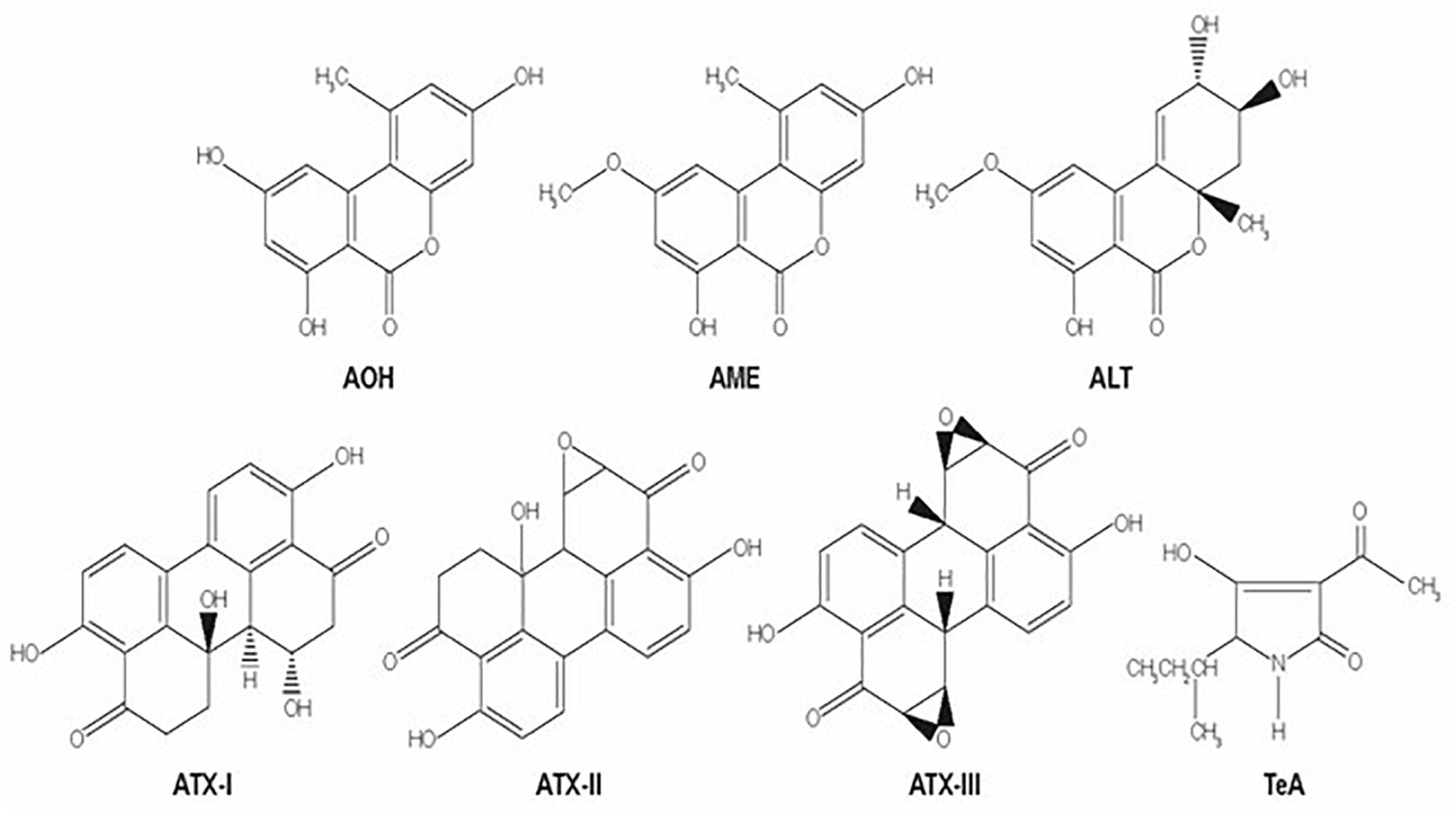
Figure 2. Molecular structures of several Alternaria mycotoxins: alternariol (AOH), alternariol monomethyl ether (AME), altenuene (ALT); the perylene derivatives altertoxins (ATX-I, -II, -III); and the tetramic acid derivatives, tenuazonic acid (TeA).
Alternariol monomethyl ether (AME) and alternariol (AOH) are not very toxic, but exert mutagenic, genotoxic and estrogenic activity (Schrader et al., 2001; Fehr et al., 2009; Dall’Asta et al., 2014), and also inhibit topoisomerase I and II (Fehr et al., 2009; Schreck et al., 2012; Dall’Asta et al., 2014). These toxins are planar benzopyrones that are quite similar to benzo(a)pyrene [B[a]P], a polycyclic aromatic hydrocarbon, and are metabolized by CYP1A1 and 1A2 cytochromes, which generate epoxides with marked activity on DNA and protein (Schreck et al., 2012). AOH and AME induce DNA strand breaks in cell lines (Pfeiffer et al., 2007), unscheduled DNA synthesis in cultured human amnion FL cells, chromosomal aberrations and sisters chromatid exchange in human peripheral blood lymphocytes, mutation in V79 cells and transformation of NIH 3T3 cells (Brugger et al., 2006; Schobert and Schlenk, 2008; Pahlke et al., 2016). These two toxins are AhR activators in mouse hepatoma cells (Schobert and Schlenk, 2008; Schreck et al., 2012; Pahlke et al., 2016). At very low concentrations altertoxin-II induces CYP1A1, which results in the production of ROS (Pahlke et al., 2016) that can cause DNA damage. However, DNA damage does occur, likely due to the generation of direct DNA adducts or the ROS induction, independently of cytochrome activity (Schreck et al., 2012; Pahlke et al., 2016). TeA is considered to be the most toxic among the Alternaria mycotoxins. It inhibits protein synthesis and is biologically active, exerting cytotoxic, phytotoxic, antitumor, antiviral and antibiotic effects (Schobert and Schlenk, 2008; Siegel et al., 2010), and has been deemed responsible for the outbreak of “onyalai” disease (Siegel et al., 2010). Also Alternaria toxins in grains might be responsible for esophageal cancer and other diseases (Dall’Asta et al., 2014; Table 1). Still, little is known about the mechanisms of action of Alternaria toxins.
Fumonisin B1
Since their discovery in 1988 (Gelderblom et al., 1988), fumonisins have been the subject of numerous toxicological investigations (Voss et al., 2007). Fumonisins are a group of mycotoxins with a strong structural similarity to sphinganine, the backbone precursor of sphingolipids, and are produced primarily by Fusarium species commonly associated mainly with cereal grains Fusarium verticillioides, F. proliferatum strains (García and Heredia, 2006; Marin et al., 2013). These toxins have been epidemiologically and experimentally associated with human esophageal cancer (Cortez-Rocha et al., 2003) and birth defects due to interference with cellular folate uptake (Stevens and Tang, 1997; Voss et al., 2007). FB1 is a cancer promoter, but a poor cancer initiator; based on toxicological evidence, the IARC has classified FB1 as a possible human carcinogen (group 2B) (IARC, 2002). FB1 co-occurred with AFB1 in a high-incidence area in human primary hepatocellular carcinoma in China, suggesting that the mixture may be involved in the development of the disease (Li et al., 2001). Toxic properties in general and the symptoms related to human exposure to FB1 are listed in Table 1.
From a toxicological perspective, fumonisin B1 (FB1) is the most important fumonisin (Figure 3). The chemical name of this mycotoxin is 1,2,3-propanetricarboxylic acid, 1,10-(1-(12- amino-4,9,11-trihydroxy-2-methyltridecyl)-2-(1-methylpentyl)-1,2-ethanediyl)ester (European Food Safety Authority [EFSA], 2005), and it elicits a spectrum of toxicities likely mediated through mechanisms involving disruption of sphingolipid metabolism and sphingolipid-mediated processes (Voss et al., 2007). This mycotoxin inhibits ceramide (CER) synthase that catalyzes the acylation of sphinganine (Sa) and recycling of sphingosine (So), increases intracellular Sa and other cytotoxic sphingoid compounds. This imbalance is mainly responsible for the toxicity (Voss et al., 2007). Thus, alterations of Sa to So ratio in tissues, urine and blood have been proposed as potential biomarkers of FB exposure, but studies have not allowed an accurate validation (Solfrizzo et al., 2011). Also, the AhR activation was demonstrated when FB1 was evaluated in H4IIE hepatoma cells. As a result, this activation initiated the CYP1A1 and CYP1A2 activity (Mary et al., 2015).
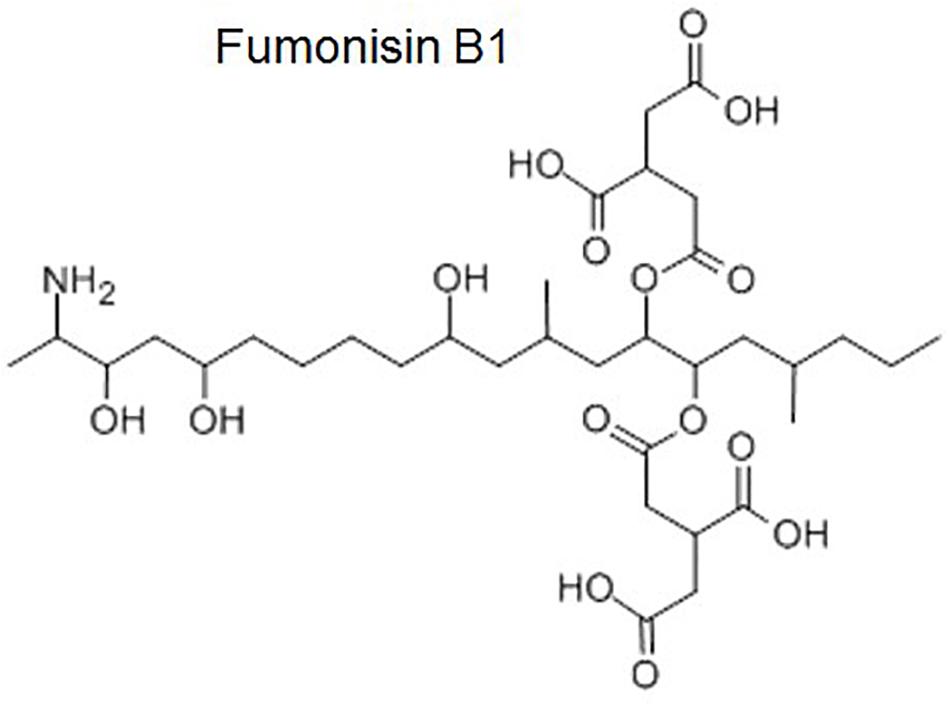
Figure 3. Molecular structure of fumonisin B1.
Ochratoxin A
Ochratoxin A (OTA) is a secondary metabolite produced by Penicillium verrucosum, many species of the Aspergillus section Circumdati (A. ochraceus, A. steynii, and A. westerdijkiae are most important regarding OTA contamination in agricultural products) and a few Aspergilli section Nigri (Samson et al., 2007; Visagie et al., 2014). OTA (Figure 4) is recognized as a potential human health hazard (Table 1) associated with kidney disease in humans (García and Heredia, 2006) and has been classified as a possible human carcinogen (Tangni et al., 2002). Structurally, it is a phenylalanyl derivative of a substituted isocoumarin(R)-N-(5-chloro-3,4-dihydro-8-hydroxy-3-methyl-1-oxo-1H-2-benzopyran-7-y1)-carbonyl)-L-phenylalanine (Marin et al., 2013). Several mechanisms of OTA toxicity have been proposed including DNA damage coupled with forming direct DNA adducts via quinone formation and a network of interacting epigenetic mechanisms including inhibition of protein synthesis, oxidative stress, mitosis disruption and activation of specific signaling pathways (Mally, 2012; Pfohl-Leszkowicz and Manderville, 2012; Vettorazzi et al., 2013). In primary human hepatocytes cultures, it induced an increase in the transcription of the AhR gene and, thus, also CYP1A1 and 1A2 cytochromes gene transcription (Ayed-Boussema et al., 2012b).
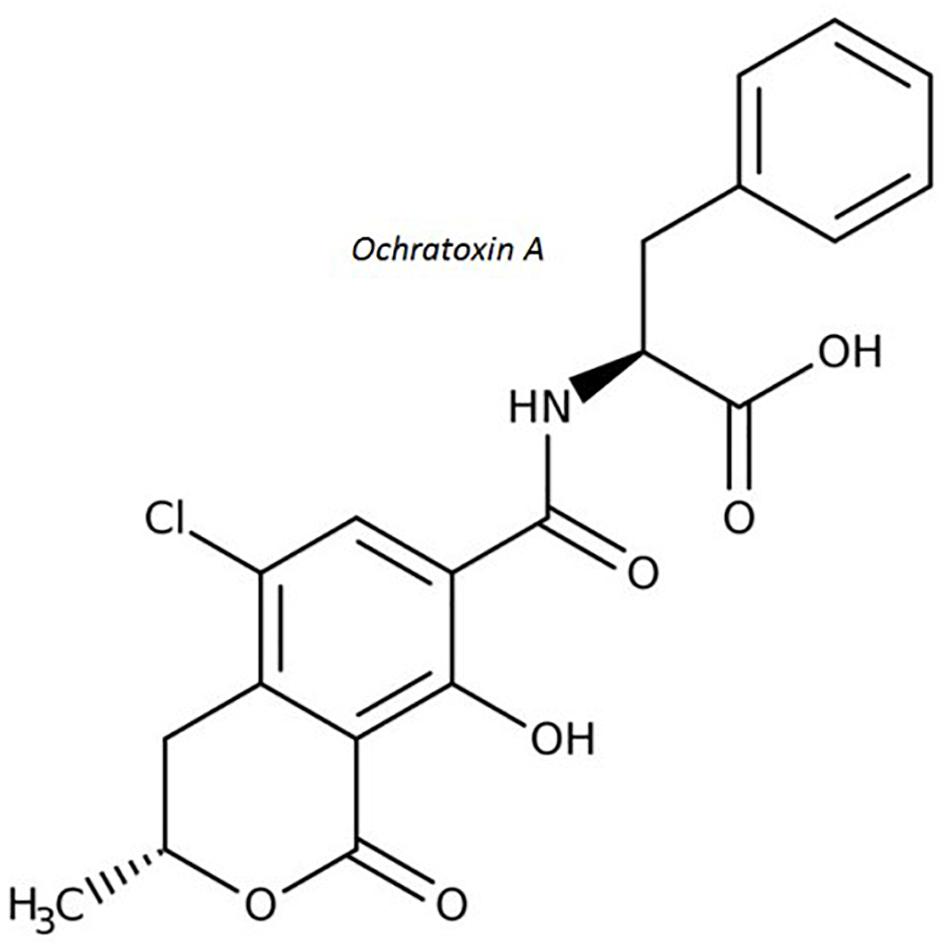
Figure 4. Molecular structure of ochratoxin A.
Patulin
Patulin (PAT) (4-38 hidroxi-4H-furo(3,2-c)piran-2(6H)-ona) is mainly produced by Penicillium expansum, but other Penicillia can produce it including P. crustosum, P. patulum as well as Aspergillus clavatus (Samson et al., 2009; Figure 5). Apples, apple-based products and other fruits are major commodities contaminated with PAT and are responsible for PAT human exposure (Piqué et al., 2013). Animal model studies obtained convulsions, agitation, ulceration, edema, intestinal inflammation, vomiting and DNA damage in brain, liver and kidneys upon PAT exposure (McKinley and Carlton, 1980; de Melo et al., 2012). Chronic toxicity is related to neurotoxic, immunotoxic, genotoxic, and teratogenic effects in rodents (Boon et al., 2009). However, based on a lack of toxicological human data, the IARC has classified PAT in Group 3 as a non-human carcinogen (IARC, 2002).
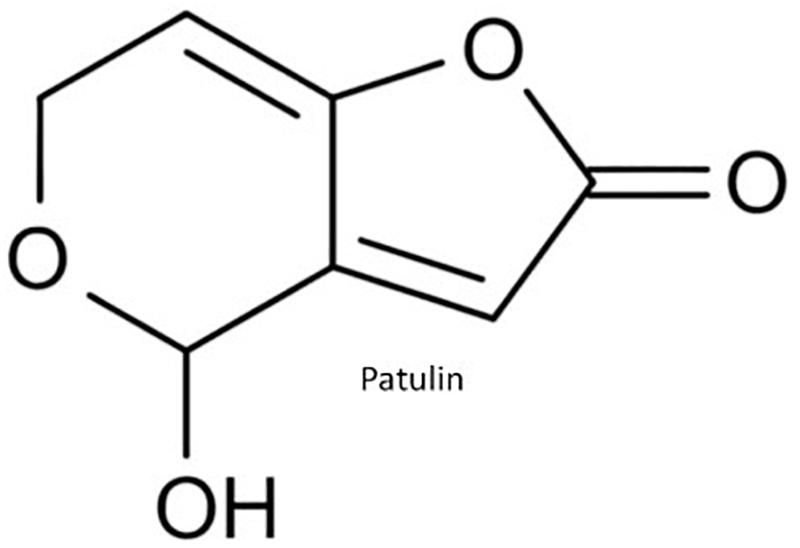
Figure 5. Molecular structure of patulin.
PAT biotransformation has been poorly investigated; increase in CYP P450 content was reported in male mice and no increase was seen in the metabolic activity in human cells previously immortalized and transfected with human CYP 450 genes (Siraj et al., 1980; Lewis et al., 1999). The study that investigated involvement of AhR pathway in PAT toxicity is discussed later on in this review (section Activation of AhR Pathway by Other Mycotoxins).
The Role of the Aryl Hydrocarbon Receptor: Historical Aspects
The study of the AhR reveals a two-dimensional history, each with its own characteristics. One is a reality of our time; namely, exposure to synthetic organic compounds and its consequences. Back at the beginning, in the 1970s, many toxicologists and, later, biochemists and molecular biologists, centered their attention on the toxicological implications of exposure to the halogenated polycyclic hydrocarbon 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) which was identified as a contaminant in the process of producing 2,4,5-trichlorophenol, a herbicide to which the workers who participated in its synthesis were exposed to (Schultz, 1957). After exposure, those workers suffered chloracne and porphyria cutanea tarda (Kimmig and Schulz, 1957). Later, studies proved that TCDD can elevate the expression of the δ- aminolevulinate synthase enzyme that participates in the heme biosynthesis (Poland and Glover, 1973). The second aspect is a rather serendipitous finding that appeared for reasons unknown at that time. In the early 1950s, studies observed that treating rats with the carcinogen 3-methylcholanthrene inhibited the hepatocarcinogenicity of 3′-methyl-4-dimethylamino azobenzene (Richardson et al., 1952). Afterward, it was proven that this carcinogenicity-inhibiting activity can be induced as well by other polycyclic aromatic hydrocarbons (PAH), such as B[a]P. Thus, it was demonstrated that these PAH can block the activity of an enzyme that modifies carcinogens, and it was suggested that, due to the constant use of B[a]P, it should be called B[a]P hydroxylase (Conney et al., 1957), which today corresponds to CYP1A1 hepatic cytochrome. Back then, in the 1960s, it was determined that the enzyme B[a]P hydroxylase, renamed at that moment as aryl hydrocarbon hydroxylase, or AHH, was inducible in some, but not all, syngeneic strains of mice (Nebert and Bausserman, 1970; Nebert et al., 1982). This response suggested the existence of a gene and AHH locus, denominated Ah, which is expressed, together with the enzyme, in the C57BL/6 strain, and corresponded to the Ahb allele. Mice of the DBA/2 strain were not responsive, and that corresponded to the Ahd allele (Nebert et al., 1972, 1982). Later, the role of the Ah locus was demonstrated in regulating the carcinogenicity, mutagenicity, teratogenicity and toxic responses to the PAH, based on inducing AHH activity (Nebert, 1986).
This provided the opportunity to test the inductive strength of AHH for TCDD and 3-methylcholanthrene, and subsequent research demonstrated that TCDD was 30,000 times more potent in AHH enzyme induction (Poland and Glover, 1974). As a result, this became the ideal test molecule for measuring all metabolisms of xenobiotic compounds in that period. Thanks to this, strains with phenotype Ahd to 3-methylcholanthrene could be made responsive to TCDD (Poland and Glover, 1974). At that time, the study of steroid receptors was also in its apogee and researchers pondered the existence of a “receptor” with smaller affinity for 3-methylcholanthrene and a greater TCDD affinity, from the Ah locus that induces AHH (Yueh et al., 2003). Once the first radioactively-marked TCDD was synthetized, in 1976, the existence of a fraction bonded to the cytoplasm and another to the nucleus was successfully demonstrated by the differential fractioning method. Similarly to what occurred with the steroid receptors, once the TCDD bonded to its receptor, it was translocated to the nucleus with its Ah receptor (Poland et al., 1976). The weight of the receptor varied if it was isolated from the cytoplasm and, in fact, was heavier than that found in the nucleus (Gasiewicz and Bauman, 1987). This raised suspicions of the possible presence of other protein compounds. It was then demonstrated that induction of AHH activity requires the formation of the TCDD-AHR compound (Jones et al., 1984). In the 1980s, the nucleotide sequence to which the TCDD-AHR compound bonded on the AHH gene was identified, and it was confirmed that it corresponded to a gene of the group of enzymes denominated cytochromes, in this case Cyp1a1. This sequence was named the dioxin response element (DRE) (Jones et al., 1985), which was shown to have the ability to activate expression of heterologous genes. At first, its existence was demonstrated in mice, but later in rats and humans as well. In another part of the world, in Japan, studies were being conducted, not regarding the response to TCDD, but to other xenobiotic compounds, and those reports found the same sequence where the TCDD-AHR compound should bond, though they called it the xenobiotic response element (XRE) (Fujisawa-Sehara et al., 1988), this sequence is: 5′-TNGCGTG-3′ (Pohjanvirta, 2011). Biochemical evidences continue to reveal differences between the molecular form of AhR found in the cytoplasm (i.e., heavier) and the one existing in the nucleus. This latter finding suggested the presence of another, very similar, weight unit. This led to the clonation and determination of the AhR nuclear receptor translocator (ARNT), a protein that forms the complex with the receptor and generates the bond with TCDD (Hoffman et al., 1991). First, the cloning of the AhR gene in humans in 1994 made it possible to prove that the initial enzyme, AHH, corresponded to the cytochrome CYP1A1. After that, researchers undertook explorations of its expression in a large number of tissues as well as in other organisms (Pohjanvirta, 2011). This information then made it possible to look for its expression in different pathologies, such as those characterized by high Cyp1a1 activity, which suggested a “cross-talk” in different pathways that run from the one in charge of metabolizing and eliminating toxic xenobiotic compounds to the alteration of such processes as the response to steroid hormones in organs like the ovaries and mammary glands (Kociba et al., 1978). From that moment forward, expression of Cyp1a1 and Cyp1b1 in response to the application of natural compounds, drugs, other PAHs, and other compounds in general such as mycotoxins, could indirectly reflect the activation and/or participation of the AhR pathway, as shown below, in a wide variety of processes, including proliferation, death/apoptosis, differentiation, cellular adhesion, and drug-expulsing proteins (Pohjanvirta, 2011).
Finally, we now know that this molecule that emerged initially from the response to toxic compounds, providing on this way a linear explanation of toxicology, specific “only” to this response, is actually a protein that is a master regulator situated above all general cellular processes, including proliferation, differentiation, cellular adhesion, death and others that will surely be described in the years to come.
General Characteristics of the AhR Receptor
Organization of the Gene
In order to know and understand better the gene function, it is important to focus the analysis on the sequence organization, from the promoter zone to the codifying region. Before beginning to describe its important molecular and sequential characteristics, it is vital to mention that the AhR gene provides a clear example of the evolution of proteins in organisms. Genes homologous to this receptor exist, from Caenorhabditis elegans, called AHR-1, which also possesses a protein that forms a pair with AHR-1 and AHA-1, which is the homolog of ARNT. Also, other organisms, like Drosophila melanogaster, have a Spineless that is homologous to human AhR; and Tango which is homologous to human ARNT. Gene conservation during evolution in pluricellular organisms has an element in common related to the differentiation processes in the early stages of development (Pohjanvirta, 2011). Detailed information related to what occurs in humans is presented below.
Thanks to the cloning of the close promoter region (Eguchi et al., 1994), it is important to mention that one of the principle characteristics is that this promoter is without a TATA box, which characterizes the promoters of specific tissue genes, and has three cis GC sites that act as bonding sites to the factor trans Sp1; a feature of gene promoters that have a constitutive expression. It also possesses a cis bonding site to CRE factors, and another E-box type site for binding to Myc. This analysis is a product of the study of the sequence -468 to -911 close to the promoter. The human gene promoter is longer than the mouse one and therefore generates a longer complete gene (promoter plus codifying region) of 6.6 pb vs. 5.5 pb, respectively. Harper et al. (2006) analyzed the presence of these cis binding sites to other transcription factors, at -5000 pb over the promoter sequence. Their results showed multiple cis sites to the factor trans HNF (hepatic nuclear factor) that numbered approximately 23 (Kaestner et al., 1994). There were also 2 cis DLX3 regions (Distal-less 3), to which this home box factor binds and functions as a placenta-specific transcriptional regulator (Morasso et al., 1999). In addition, it contains 11 BRN3-like sites that serve to bind to transcription factors of the POU family specifically expressed in the nervous system of mammals in development and in adult state (Korkalainen et al., 2005). Moreover, this complex participates in the differentiation and survival of motor and sensory neurons (McEvilly et al., 1996). There are also four cis sites to STAT6 (a signal transducer and transcription 6 activator) which belong to the family of the transcription factors that respond to cytokines such as IL-4, and to growth factors like TGF-β1 (Tanaka et al., 2005). In addition, there are 7 cis regions that bind to factors such as Tcf/Lef, other transcription factors that realize their nuclear response to Wnt signaling by interacting with β-catenin (Eastman and Grosschedl, 1999). Finally, there are 11 cis binding sites to potential nuclear receptors of the progesterone-, androgen-, glucocorticoid-, proliferation-activated peroxisome-, the farsenoid X-, and vitamin D- receptors (Harper et al., 2006). This full information on the cis sites inside the complete promoter makes it possible to see and understand the constitutive expression and specific tissue of the receptor, while also allowing us to comprehend one additional characteristic: the fact that it is a constitutive gene with ubiquitous expression; due to the important biological functions it fulfills (Hahn, 2002). It is especially critical during embryogenesis and fetal development in the first post-natal stages, after which its levels decrease significantly. This suggests that its role is required less in the final phases of an organism’s growth (Harper et al., 2006) and also allows for the explanation of the broad responses observed in several tissues and organs where xenobiotic compounds such as the aflatoxins, exert their hepato-, nefro- and neurotoxic activity as well as mutagenic and carcinogenic properties.
The open reading frame has 12 exons, which are organized and formed in the mature messenger and the important domains of the protein, as it will be described below.
Important Domains of the Protein
The detailed information regarding the domains present in the protein was obtained principally by cloning mouse genes. The mouse’s peptic sequence revealed the presence of 805 residues of amino acids organized in domains present in those bHLH-PAS transcription factors (Burbach et al., 1992). These factors contain a binding region to DNA basic Helix-Loop-Helix (bHLH) and a pair of domains that have been found in the Per proteins (a factor that participates in regulating the circadian cycle), in ARNT (i.e., the protein with which AhR realizes heterodimerization) and, finally, with a protein (that participates in neuronal development in Drosophila) called “single-minded,” or Sim, which gives this domain its name: PAS (Pohjanvirta, 2011). Analyses based on the terminal amino domain show a region of basic amino acids residues, followed by the hHLH motif. The basic residues region is important for the AhR interaction with the response element cis sequence to the aryl hydrocarbons, ARE; while the hHLH motif is important for the realization of heterodimerization between AhR and Arnt. Ahead of this hHLH motif there are 2 repeated inversions regions of 51 amino acids residues that are also found in the PAS factors described above. These motifs perform multiple functions, such as directing the AhR union with 2 Hsp90 molecules and with the protein AIP (AhR interacting protein), or XAP2 (X-associated protein 2) in the cytoplasm (Ma and Whitlock, 1997), the bond to the ligand and the interaction of AhR and Arnt in the nucleus (Figure 2). The carboxy terminal contains three domains for the transcription activation (AT) that contain residues of amino acids that are acidic and rich in serine/threonine and glutamate. Between the PAS B and AT domains there is a small sequence of approximately 81 residues of amino acids that exerts an inhibiting activity on AT and so has been named the inhibiting domain (ID) (Ma et al., 1995).
Associated Proteins and Formation of the AhR Complex
One must recall that, historically, the study of the AhR was always conducted based on the results observed during exposure to, and interaction with, TCDD. The AhR in the cytosolic fraction was shown by ultracentrifugation to have a 9S sedimentation value. Upon adding the ligand, i.e., TCDD– this value decreases to 6S and, moreover, is found in the nuclear fraction (Pohjanvirta, 2011). These differences revealed the existence of two different forms in the two cellular compartments. In addition, electrophoretic studies demonstrated that this weight difference was due to the fact that the receptor protein in the cytoplasm is found in complex tetrameric compounds by that same receptor and three proteins with molecular weights of 96, 88, and 37 kDa, the first two corresponding to two isoforms of mouse HSP90 (Chen and Perdew, 1994). The protein XAP2 (X-associated protein 2) (Meyer et al., 1998), as well as AIP (AhR-interacting protein) (Ma and Whitlock, 1997) or ARA9 (AhR-associated protein 9) (Carver et al., 1998) is the one that corresponds to molecular weight 43–37 kDa (Figure 6). It has been shown that the interaction between Hsp90 and the AhR takes place in the PAS-B motif, and that this interaction is required to create the conformational change that allows binding with the ligand. The protein AIP has three repeated sequences of tetratricopeptides that allow protein-protein interaction (Schreiber, 1991). This explains why the receptor protein in the cytoplasm is more stable and has a half-life of approximately 28 h. This data corresponds to the earlier description which indicated that the onset of the induction of CYP1A1 does not require protein synthesis. After treatment with a ligand (TCDD or B[a]P), the receptor’s half-life is reduced to just 3 h (Ma and Baldwin, 2000). Afterward, the AhR protein suffers degradation by the 26S proteasome (Pohjanvirta, 2011), which takes place in the nucleus (Figure 6), an important site for the degradation of other transcription factors in addition to AhR (Roberts and Whitelaw, 1999), such as TGF-β1 (Nebert, 1986; Lo and Massagué, 1999) and MyoD (Floyd et al., 2001).
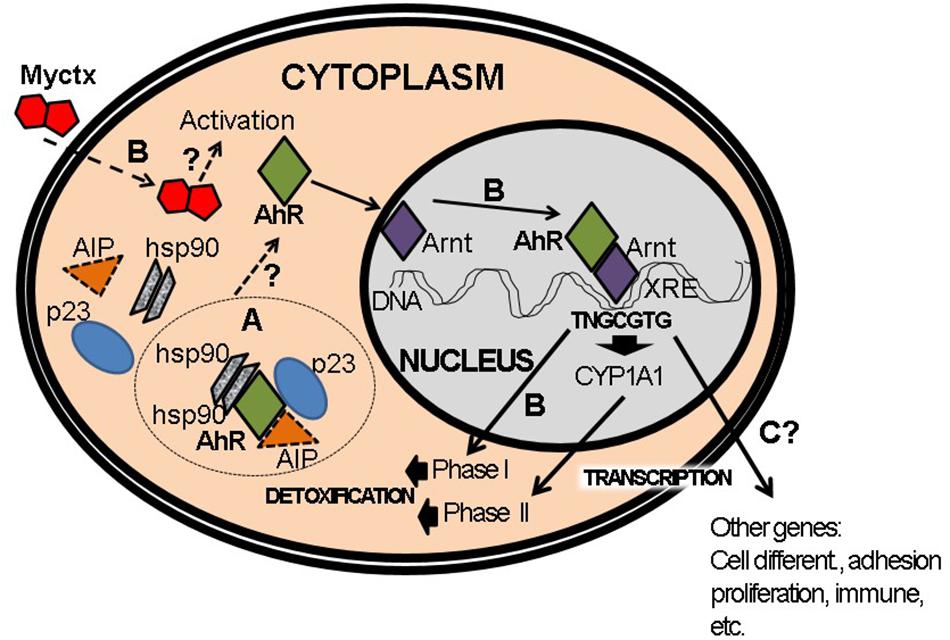
Figure 6. Activation of AhR pathway by mycotoxins (Myctx). (A) The AhR is a protein that interacts with hsp90, AIP, and p23 proteins in cytoplasm. (B) Under activation by Myctx, the transcription of phase I and phase II genes is started in order to metabolize and detoxify. It is not proved if the Myctx are ligands/agonists of AhR, which is indicated by a question mark (?). (C) This activation can also start transcription of several genes such as those involved in cell differentiation, adhesion, proliferation and immune response, but in the case of Myctx these have to be proved if it is performed as ligands/agonists (?).
The Canonical Pathway of AhR Activation Through Interaction With Arnt
In order to explain the canonical pathway of AhR activation (Figure 2), the description must be based strongly on the activation of the genes studied that participate in the detoxification mechanism, or in phase I and phase II of the xenobiotic compounds metabolism such as the cytochromes Cyp1a1 and Cyp1a2. This history starts with the binding of the ligand to the AhR, that binds and causes a conformational change in the PAS A domain which enables demonstration from the residues of amino acids 55–75 to a nuclear localization signal, allowing its translocation to the nucleus (Figure 2). As mentioned previously, it dissociates in the cytoplasm of the complex of 2 molecules of Hsp90, and translocates to the nucleus in a process fostered by importins (Ikuta et al., 1998). Once inside the nucleus, AhR is dimerized with another protein from the same family as factors bHLH-PAS, denominated Arnt. Arnt is only localized in the nucleus and this protein is also organized in domains similar to those that contain AhR, i.e., toward the amino terminal end we find the bHLH domain and the nuclear localization signal, followed by the PAS A and PAS B domains (Figure 2). Finally, the activation domain is contained toward the carboxyl terminal end (Pohjanvirta, 2011). The dimerization between AhR and Arnt is realized through the HLH regions (Reisz-Porszasz et al., 1994; Fukunaga et al., 1995) and is stabilized by a conformational change in the PAS A region (Soshilov and Denison, 2008). Observations have shown that modifications by phosphorylation occur for the binding function to the DNA via the protein kinase C (Mahon and Gasiewicz, 1995). Binding to the ARE cis site in the promoter sequences takes place with the consensus sequence 5′-TNGCGTG-3′, which is seen in many genes, such as cytochrome CYP1A1, which contains 8; CYP1A2, which contains 1, and CYP1B1, which contains 3 sequences (Pohjanvirta, 2011). For this reason, it is said that these genes are regulated by the canonical pathway and that CYP1A1 is the model gene for the study of the response to AhR. In the context of the binding, AhR binds to the middle of the 5′-T/NGC site, while Arnt binds by occupying the second half of 5′GTG (Bacsi et al., 1995). We also know that a series of nucleotides located near the ARE are important because its cis element contains the optimal sequence: GGGTGNAT(C/T)GCGTGACNNCC. This sequence establishes contact with the bHLH domain of the AHR/ARNT heterodimer (Chapman-Smith and Whitelaw, 2006). We also know that the promoter has a second sequence or response element to AHR II, whose sequence is CATG(N)C(T/A)TG, and which is conserved in humans, rats and mice, but does not seem to influence the canonical response to ARE’s action on the cis site (Pohjanvirta, 2011). Thanks to the exhaustive studies that have been conducted on the regulation of the expression of the cytochromes during AhR activation, it was possible to identify various genes that have AREs (Gasiewicz et al., 2008), which can be grouped as follows:
1. Phase I genes of the metabolism of xenobiotics: CYP1A1, CYP1A2, CYP1B1, CYP2A5, CYP2S1, and CYP4B1.
2. Phase II genes of the metabolism of xenobiotics: aldehyde dehydrogenase 3A1, glutation-S-transferase, NAD(P)H- quinone oxidoreductase-1, UDP glucuronosyltransferase 1A1, and 1A6.
3. Cell cycle suppressors: p21, p27.
4. Cell cycle activators: c-jun, s-myc, junD, Insulin-like growth factor binding protein-1.
5. Others: Bax, cathepsin D, cyclooxygenase -2, epiregulin, filaggrin, Slug, etc.
There is also one exceptional case; that of poly/ADP-ribose polymerase, which contains 16 XRE cis sequences (Ma, 2002). One aspect of the consequences of cytochrome activation is the generation of modifications of the xenobiotic compounds that induce their activation and achieve control of the receptor activation while reducing ligand concentrations in the cell (Gu et al., 2000). All currently available evidence demonstrate that the AhR protein can function as an exogenous ligand sensor, and that these compounds appeared only recently in the human ecosystem such as polycyclic aromatic hydrocarbons (the technosphere). Thus, it can be said that the canonical pathway of AhR-Arnt may well be activated as a response to the presence of toxic compounds produced by humans, and that thanks to a process of conserved evolution, the AhR protein – belonging to a group of proteins known to be environmental sensors – is a response mechanism with the function to detoxify the organism from those “new” compounds that are foreign to nature. In this context, activation of the canonical pathway by AhR-Arnt is separated from its physiological role, in which surely many endogenous compounds perform functions less related to toxicity, and much more focused on the processes of development and differentiation of mammals, since this receptor and this group of PAS proteins also play important roles in development, a function in which AhR perhaps operates initially in the early stages of mammals, as mentioned above.
The Non-canonical AhR Pathway
In the past 10 years, the development of research analyzing the microarrays of genes in samples of the AhR from the organs of knockout mice, and cell lines principally from the liver, and their comparison with WT mice to AhR and in normal lines of gene expression, have revealed that in the absence of ligands exogenous to the receptor exists an expression of genes for which, upon analyzing the cis regions of some promoters using chromatin immunoprecipitation (or CHIP), it was possible to show that they present sequences distinct from those of ARE described in the canonical pathway, which have been denominated as non-consensus response elements to xenobiotics (NC-XRE) (Peters et al., 1999; Dere et al., 2011). As a result, the non-canonical AhR pathway refers to the expression of genes which promoters even in the presence of some ligand, such as TCDD, do not contain the characteristic cis element of the ARE. One of these genes is the plasminogen-1 activator inhibitor (PAI-1) (Tijet et al., 2006). Later studies corroborated that treatment with TCDD suppresses hepatic regeneration, and we now understand that this is due to PAI-1 inhibition of the urokinase-type plasminogen activator that is necessary for the activation of the hepatic growth factor (Mitchell et al., 2006). The characteristic of these promoters is that they contain a repeated tetranucleotide: 5′-GGGA-3′. In the case of the PAI-1 promoter, it is the second NC-XRE that is important for the binding of AhR. One important feature is the absence of interaction with Arnt in this gene activation (Huang and Elferink, 2012). It has also been proven that suppression of hepatic regeneration is realized by stopping cell proliferation by inducing a delay in phase G1 by inhibiting the activity of kinase 2-dependent cyclin (CDK2) (Mitchell et al., 2006). This is dependent on the increase in the expression of kinase-dependent cyclin inhibitors such as p21 and p27. These inhibitors negatively regulate the progression of the cell cycle by controlling CDK2, CDK4 and CDK6 (Harper et al., 1995). Later, through analyses of the p21 promoter sequence, it was discovered that they contain NC-XRE cis regions (Jackson et al., 2015). Also, an endogenous ligand must exist that allows, in the absence of a bond to some ligand, expression of AhR-dependent p21, as was proven in the hepatic regeneration model through partial hepatectomy (Jackson et al., 2014).
Recent evidence demonstrates that the Kruppel-like factor (KLF) forms a heterodimer with the AhR receptor (Wilson et al., 2013). Structurally, the carboxy terminal of AhR is where the bHLH and PAS A domains are found that must interact with the amino terminal end of KLF6 (Wilson et al., 2013). The KLF6 factor regulates numerous cellular processes that participate in differentiation, proliferation and apoptosis (Philipsen and Suske, 1999). Alterations to the KLF6 gene are associated with various types of cancer, including astrocytomas and gliomas (Jeng and Hsu, 2003). The KLF6 factor also activates expression of p21, forming the heterodimer, which is part of the activation of the non-canonical pathway of AhR activation (Andreoli et al., 2010). The KLF6 factor induces an increase in the expression of the E-cadherin genes, transforming the growth factor and the insulin-like growth factor 1 receptor (Rubinstein et al., 2004).
Implication of the AhR Pathway in Toxicity of Aflatoxins and Some Other Mycotoxins
It is important to understand, firstly, that the proposal for this review arose because so little has been written on this topic up to date. Secondly, we must be aware that the reports that do exist are mainly related to AFB1 (Ayed-Boussema et al., 2011, 2012a; Merrick et al., 2013; Mary et al., 2015), and few to Alternaria toxins (Pahlke et al., 2016), ochratoxin A (Ayed-Boussema et al., 2012b), fumonisin and patulin (Ayed-Boussema et al., 2011, 2012a; Mary et al., 2015). All of these studies have demonstrated the activation of members of different families of CYP during transcription (Ayed-Boussema et al., 2012b; Mary et al., 2015; Pahlke et al., 2016), enzymes with UDP-glucuronidase activity (Fleck et al., 2012; Hanioka et al., 2012), and the role played by such receptors as pregnane X (PXP), and the constitutive androstane (CAR), retinoid X (RXR), glucocorticoids (RG), and Ah receptors (Fleck et al., 2012; Hanioka et al., 2012; Mary et al., 2015; Pahlke et al., 2016). Some of these studies also evaluated the cytotoxic and genotoxic potential of these toxins (Koliopanos et al., 2002; Ayed-Boussema et al., 2012a; Fleck et al., 2012). The following describe the key results that different working groups have obtained regarding aflatoxins and other mycotoxins and their relation to the AhR.
Activation of AhR Pathway by Aflatoxins
AFB1 is a powerful hepatocarcinogen (Merrick et al., 2013). Due to its planar structure and activation of transcriptional expression of the CYP1A1 in H4IIE hepatoma cells, it was suggested to be agonist for the AhR, but at the moment the direct evidence is lacking (Mary et al., 2012). Increased expression has been observed in carcinomas of the pancreas and lungs (Koliopanos et al., 2002; Lin et al., 2003). This increase in its expression is also related to an increase in the invasion of gastric cancer cells. Consequences of the binding and activation of the AhR include the induction of free radicals and ROS (Mary et al., 2012). This seems to play an important role in carcinogenesis and the progression of carcinomas. In fact, their operation functions by reducing immunovigilance of the carcinogenic process because activation of the AhR affects the regulatory function of T CD4 + lymphocytes (Pot, 2012). The activation of AFB1 to the AhR allows the activation of the rapid response of AhR and the activation of tyrosine kinases and the ERK and PKC1 signaling pathways (Tan et al., 2004; Schreck et al., 2012). In cultures of human hepatocytes, AFB1 also increased the transcriptional expression of RXR due to the increased expression of the genes of CYP2B6 and CYP3A5 cytochromes, whose expression is regulated by this receptor (Ayed-Boussema et al., 2012b).
An analysis of genes differentially-expressed by RNAseq (transcriptome) from rat livers revealed an at least twofold increase, to a total of 1026 genes. These genes participate in the AhR, NF-E2-related factor 2 (Nrf2) and glutathione (GSH) pathways, the cell cycle of the metabolism of xenobiotic compounds, the extracellular matrix and cell differentiation (Merrick et al., 2013). Observations of the H4IIE hepatocellular carcinoma lines showed activation of the AhR and of the transcription of the CYP1A1 gene (Mary et al., 2015). Finally, in HepG2 hepatocellular carcinoma cells, a significant expression of UGTs of the families 1 and 2 was observed (Hanioka et al., 2012).
There are transcription factors (TF) associated to DNA damage response. After a combination of transcriptomic analyses and prediction of binding of TF, in HepG2 cells after exposure to 5μM of AFB1, it was revealed that the canonical pathway is important in the response to modulate the DNA repair response by assessing the phosphorylation of H2AX histone (γH2AX), via ARNT. In the absence of ARNT, reduction in CYP1A1 expression and in γH2AX response demonstrated that canonical pathway of AhR is necessary for the DNA repair response after exposure to AFB1 (Smit et al., 2017).
Finally, the efflux transporter ABCG2 plays an important role in the mammary excretion of toxins in humans. AFB1 can interact with the ABCG2 transporter even at higher aflatoxin concentrations. This interaction produces an efflux effect of dioxin-like compounds such as PCB 126, but this function depends of AhR activation since it is inhibited by CH233191, an inhibitor of the AhR pathway. Therefore, this evidence opens new issues to research the role produced by the interaction between AFB1 and the close action of the AhR pathway (Manzini et al., 2017).
Activation of AhR Pathway by Alternaria Toxins
Alternaria toxins, alternariol and its monomethyl ether are planar benzopyrones that are quite similar to B[a]P, compounds metabolized by CYP1A1 and 1A2 cytochromes, which generate epoxides with marked activity on DNA and protein (Schreck et al., 2012). They have both mutagenic and genotoxic effects, induce micronuclei, and show estrogenic activity (Table 1; Fleck et al., 2012; Schreck et al., 2012; Pahlke et al., 2016). They are also related to the development of esophageal cancer and are inhibitors of DNA topoisomerases (Fehr et al., 2009). Only monomethyl ether stops the cells in G2/M. Apoptosis is induced at 48 h of exposure to both compounds (Schreck et al., 2012). These two toxins are AhR activators in mouse hepatoma cells (Burkhardt et al., 2012; Schreck et al., 2012; Pahlke et al., 2016). Also altertoxin-II, at very low concentrations, induces CYP1A1, resulting in the ROS production (Pahlke et al., 2016) that can cause DNA damage, independently of the AhR, since the siRNA of the receptor does not induce CYP1A1. However, DNA damage does occur, likely due to the generation of direct DNA adducts, or the ROS induction independently of cytochrome activity (Burkhardt et al., 2012). For this reason, it is important to evaluate, as well, whether they have the capacity to induce phase II enzymes of detoxification, such as glucuronidation, because the metabolites of these mycotoxins do not cause DNA damage. This could represent a strategy for reducing the toxic properties of these mycotoxins through conjugation with glucuronic acid.
Activation of AhR Pathway by Other Mycotoxins
So far, there have been only a few studies related to other mycotoxins, including OTA, FB1 and patulin as activators of AhR. In primary cultured human hepatocytes treated with subcytotoxic OTA concentrations, significant upregulation of CYP3A4, CYP2B6, and, to a lesser extent, CYP3A5 and CYP2C9 was obtained. PXR mRNA expression increased in only 1 treated liver, whereas CAR mRNA expression was not affected. OTA induced an overexpression of CYP1A1 and CYP1A2 genes accompanied by an increase in AhR mRNA expression suggesting that OTA could activate both PXR and AhR in human hepatocytes (Ayed-Boussema et al., 2012b). Also, in human kidney cells HK-2 OTA activated both AhR and PXR by induction of the CYP1A1, CYP1A2, and CYP3A4 genes. The mRNA expression of phase II enzymes such as heme oxygenase-1, nicotinamide adenine dinucleotide phosphate-quinone oxidoreductase 1, and glutamate cysteine ligase catalytic subunit were upregulated by the activation of Nrf2. Nrf2 activation is directly induced by OTA-activated AhR and PXR, or indirectly by the adaptive response to ROS generated through metabolic processes between AhR, PXR-induced CYP enzymes, and OTA. Taken together, these studies indicate that OTA induces phase I and II enzymes through the activation of AhR, PXR, and Nrf2 signaling pathways, which may lead to cell injury (Lee et al., 2018).
Induction of CYP enzymes by FB1 has been poorly investigated so far; but an increase of CYP1A activity in the liver of Wistar rats exposed to FB1 has been confirmed (Martínez-Larrañaga et al., 1996). Recent study demonstrated that FB1 applied alone caused a small increase in CYP1A activity and cyp1A gene transcription in H4IIE and rat spleen mononuclear cells, but to a lesser extent and duration than single AFB1. However, cyp1A gene expression by FB1 applied alone did not involve Ahr activation. On the other hand, a two-toxin mixture induced an overexpression of cyp1A and ahr genes in spleen cells indicating the interaction between both toxins in AhR pathway activation and subsequently in carcinogenesis (Mary et al., 2015).
In primary human hepatocytes, PAT induced an upregulation of PXR and CYP2B6, 3A5, 2C9, and 3A4 genes accompanied by AhR and CYP1A1 and 1A2 mRNA expression. Since this is the only study available in literature related to AhR activation by PAT, further studies should determine whether this mechanism could modulate PAT toxicity (Ayed-Boussema et al., 2012a).
Conclusion and Perspectives
‐ Due to their toxic properties, aflatoxins and other mycotoxins are AhR activators and are also mutagenic and carcinogenic.
‐ As lipophilic compounds, they can easily cross cells biological membranes that are a part of the entrance pathway; for example, the intestinal wall. For this reason, they constitute as pro-carcinogenic molecules.
‐ Their activation as carcinogenic metabolites has centered on modification by mono-oxygenases, represented primarily by CYP1A1, that are expressed abundantly in the liver. Furthermore, the CYP1A1 promoter has 8 XRE-elements which are used by the AhR activation.
‐ The gene expression increase and the activity of such cytochromes such as CYP1A1 are important factors due to their activity, triggered by exposure to aflatoxins can go on for weeks or months, meaning that the carcinogenic process can be sustained due to the consequences of this activity.
‐ Aflatoxin metabolites are mutagenic and some produce adducts on the nitrogenous bases that can cause base changes and modifications in the methylation states of gene promoters and thereby establish a modified expression pattern. This area has not been yet fully explored.
‐ These metabolites also induce an increase in ROS that can cause damage to DNA, lipids or other proteins, and thereby initiate the process of carcinogenesis.
‐ Mutations can be produced in genes with important functions but they are also targets for mutagen modification such as p53 and K-Ras, among others.
‐ The increase in mRNA expression of the AhR is significant due to significant expression of this gene in liver and adipose tissue, with the latter potentially serving as a reservoir for lipophilic compounds like aflatoxins. It is therefore important to mention that in the AhR promoter there are several important cis elements which have an important function in interactions with trans factors. Some cis elements are: HNF (binding of the hepatic nuclear factor); DLX3 (regulatory placenta-specific); BRN2-like (binding of factors participating in neural development and adult stage in mammals, also performing differentiation of sensitive and motor neurons); STAT6 (responding to cytokines such as IL-4 and TGF-β1); Tcf/Lef (Wnt via/neurodevelopment). Binding sites to potential nuclear receptors to the progesterone receptor, androgen receptors, glucocorticoid receptors, proliferation-activated peroxisome receptors, the farsenoid X receptor and the vitamin D receptor could explain the biological effects of these toxins. These cis elements partly enable the comprehension of the toxic effects caused by the activation of AhR, but this is a less studied area.
‐ Up to date, the AhR activation has been limited to groups of genes such as the cytochromes and UDP-glucuronidases. However, we know that AhR activation can participate in inflammatory responses, cell proliferation and differentiation, and the loss of cell adhesion. Thus, the implications of AhR activation by aflatoxins may be more serious than thought thus far.
‐ Certain polymorphisms can predispose tissues to cancer development, when they are exposed to aflatoxins. This field needs to be studied broadly since we know that at least some polymorphisms of enzymes like GCTP1, AKR, EPHX1, and EERC2 may participate in cancer development.
‐ The pathways that depend on the AhR activation have been explored, but there must be others, such as the non-canonical receptor pathway that participates in the toxic and carcinogenic effects by aflatoxins. At this point it is important to emphasize that AhR protein heterodimerizes with KLF6 and this complex can activate CYP1A1. Finally, KLF6 can regulate differentiation, proliferation and apoptosis.
‐ The study of the aflatoxins effects thus represents an area of opportunity in which diverse specialists, including researchers in omics, must work together in efforts to integrate the data that make their field of study so complex.
Author Contributions
FA-H, RV-Q, and MS contributed mostly in final shaping and improving the manuscript, while the other authors also contributed in writing but also in designing the tables, figures, and correcting the manuscript with English and reference editing. All authors have contributed to the idea and form of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank to Prof. Maxo Herman for the help with English editing of the manuscript. This work was also a part of collaboration on a CA Environment and Genetics UATLX-CA-223 project under the name Evaluación del riesgo genotóxico por exposición a contaminantes ambientales, leader Rafael Valencia, Ph.D. (Universidad Autonoma de Tlaxcala) and collaborators Prof Stefano Bonassi (Italy) and Dr. Mirta Milic (Croatia). We thank to Dirección de Investigación of Hospital Infantil de Mexico Federico Gomez for its financial support.
References
Andreoli, V., Gehrau, R. C., and Bocco, J. L. (2010). Biology of Krüppel-like factor 6 transcriptional regulator in cell life and death. IUBMB Life 62, 896–905. doi: 10.1002/iub.396
Ayed-Boussema, I., Pascussi, J. M., Maurel, P., Bacha, H., and Hassen, W. (2011). Activates pregnane X receptor, constitutive androstane receptor and aryl hydrocarbon receptor and corresponding phase I target genes mRNA in primary cultures of human hepatocytes. Environ. Toxicol. Pharmacol. 31, 79–87. doi: 10.1016/j.etap.2010.09.008
Ayed-Boussema, I., Pascussi, J. M., Rjiba, K., Maurel, P., Bacha, H., and Hassen, W. (2012a). The mycotoxin, patulin, increases the expression of PXR and AhR and their target cytochrome P450s in primary cultured human hepatocytes. Drug Chem. Toxicol. 35, 241–250. doi: 10.3109/01480545.2011.592194
Ayed-Boussema, I., Pascussi, J. M., Zaied, C., Maurel, P., Bacha, H., and Hassen, W. (2012b). Ochratoxin A induces CYP3A4, 2B6, 3A5, 2C9, 1A1, and CYP1A2 gene expression in primary cultured human hepatocytes: a possible activation of nuclear receptors. Drug Chem. Toxicol. 35, 71–80. doi: 10.3109/01480545.2011.589438
Bacsi, S. G., Reisz-Porszasz, S., and Hankinson, O. (1995). Orientation of the heterodimeric aryl hydrocarbon (dioxin) receptor complex on its asymmetric DNA recognition sequence. Mol. Pharmacol. 47, 432–438.
Bennett, J. W., and Klich, M. (2003). Mycotoxins. Clin. Microbiol. Rev. 16, 497–516. doi: 10.1128/CMR.16.3.497-516.2003
Boon, P. E., Ruprich, J., Petersen, A., Moussavian, S., Debegnach, F., and van Klaveren, J. D. (2009). Harmonisation of food consumption data format for dietary exposure assessments of chemicals analysed in raw agricultural commodities. Food Chem. Toxicol. 47, 2883–2889. doi: 10.1016/j.fct.2009.08.003
Bräse, S., Encinas, A., Keck, J., and Nising, C. F. (2009). Chemistry and biology of mycotoxins and related fungal metabolites. Chem. Rev. 109, 3903–3990. doi: 10.1021/cr050001f
Brugger, E.-M., Wagner, J., Schumacher, D. M., Koch, K., Podlech, J., Metzler, M., et al. (2006). Mutagenicity of the mycotoxin alternariol in cultured mammalian cells. Toxicol. Lett. 164, 221–230. doi: 10.1016/j.toxlet.2006.01.001
Bryden, W. L. (2007). Mycotoxins in the food chain: human health implications. Asia Pac. J. Clin. Nutr. 16(Suppl. 1), 95–101. doi: 10.6133/apjcn.2007.16.s1.18
Burbach, K. M., Poland, A., and Bradfield, C. A. (1992). Cloning of the Ah-receptor cDNA reveals a distinctive ligand-activated transcription factor. Proc. Natl. Acad. Sci. U.S.A. 89, 8185–8189. doi: 10.1073/pnas.89.17.8185
Burkhardt, B., Jung, S. A., Pfeiffer, E., Weiss, C., and Metzler, M. (2012). Mouse hepatoma cell lines differing in aryl hydrocarbon receptor-mediated signaling have different activities for glucuronidation. Arch. Toxicol. 86, 643–649. doi: 10.1007/s00204-011-0789-8
Carver, L. A., LaPres, J. J., Jain, S., Dunham, E. E., and Bradfield, C. A. (1998). Characterization of the Ah receptor-associated protein, ARA9. J. Biol. Chem. 273, 33580–33587. doi: 10.1074/jbc.273.50.33580
Chapman-Smith, A., and Whitelaw, M. L. (2006). Novel DNA binding by a basic helix-loop-helix protein. The role of the dioxin receptor PAS domain. J. Biol. Chem. 281, 12535–12545. doi: 10.1074/jbc.M512145200
Chen, H. S., and Perdew, G. H. (1994). Subunit composition of the heteromeric cytosolic aryl hydrocarbon receptor complex. J. Biol. Chem. 269, 27554–27558.
Conney, A. H., Miller, E. C., and Miller, J. A. (1957). Substrate-induced synthesis and other properties of benzpyrene hydroxylase in rat liver. J. Biol. Chem. 228, 753–766.
Cortez-Rocha, M. O., Ramírez-Astudillo, W. R., Sánchez-Mariñez, R. I., Rosas-Burgos, E. C., Wong-Corral, F. J., Borboa-Flores, J., et al. (2003). Fumonisins and fungal species in corn from Sonora, Mexico. Bull. Environ. Contam. Toxicol. 70, 668–673. doi: 10.1007/s00128-003-0036-y
Dall’Asta, C., Cirlini, M., and Falavigna, C. (2014). Mycotoxins from Alternaria: toxicological implications. Adv. Mol. Biol. 8, 107–121. doi: 10.1016/B978-0-444-63406-1.00003-9
de Melo, F. T., de Oliveira, I. M., Greggio, S., Dacosta, J. C., Guecheva, T. N., Saffi, J., et al. (2012). DNA damage in organs of mice treated acutely with patulin, a known mycotoxin. Food Chem. Toxicol. 50, 3548–3555. doi: 10.1016/j.fct.2013.07.052
Dere, E., Lo, R., Celius, T., Matthews, J., and Zacharewski, T. R. (2011). Integration of genome-wide computation DRE search, AhR ChIP-chip and gene expression analyses of TCDD-elicited responses in the mouse liver. BMC Genomics 12:365. doi: 10.1186/1471-2164-12-365
Eastman, Q., and Grosschedl, R. (1999). Regulation of LEF-1/TCF transcription factors by Wnt and other signals. Curr. Opin. Cell Biol. 11, 233–240. doi: 10.1016/S0955-0674(99)80031-3
Eguchi, H., Hayashi, S., Watanabe, J., Gotoh, O., and Kawajiri, K. (1994). Molecular cloning of the human AH receptor gene promoter. Biochem. Biophys. Res. Commun. 203, 615–622. doi: 10.1006/bbrc.1994.2227
European Food Safety Authority [EFSA] (2005). Opinion of the Scientific Panel on contaminants in the food chain (CONTAM) related to fumonisins as undesirable substances in animal feed. EFSA J. 235, 1–32. doi: 10.2903/j.efsa.2005.235
Fan, C., Cao, X., Liu, M., and Wang, W. (2016). Determination of Alternaria mycotoxins in wine and juice using ionic liquid modified countercurrent chromatography as a pretreatment method followed by high-performance liquid chromatography. J. Chromatogr. A 1436, 133–140. doi: 10.1016/j.chroma.2016.01.069
Fehr, M., Pahlke, G., Fritz, J., Christensen, M. O., Boege, F., Altemöller, M., et al. (2009). Alternariol acts as a topoisomerase poison, preferentially affecting the II alpha isoform. Mol. Nutr. Food Res. 53, 441–451. doi: 10.1002/mnfr.200700379
Fleck, S. C., Burkhardt, B., Pfeiffer, E., and Metzler, M. (2012). Alternaria toxins: altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol. Lett. 214, 27–32. doi: 10.1016/j.toxlet.2012.08.003
Floyd, Z. E., Trausch-Azar, J. S., Reinstein, E., Ciechanover, A., and Schwartz, A. L. (2001). The nuclear ubiquitin-proteasome system degrades MyoD. J. Biol. Chem. 276, 22468–22475. doi: 10.1074/jbc.M009388200
Fujisawa-Sehara, A., Yamane, M., and Fujii-Kuriyama, Y. (1988). A DNA-binding factor specific for xenobiotic responsive elements of P-450c gene exists as a cryptic form in cytoplasm: its possible translocation to nucleus. Proc. Natl. Acad. Sci. U.S.A. 85, 5859–5863. doi: 10.1073/pnas.85.16.5859
Fukunaga, B. N., Probst, M. R., Reisz-Porszasz, S., and Hankinson, O. (1995). Identification of functional domains of the aryl hydrocarbon receptor. J. Biol. Chem. 270, 29270–29278. doi: 10.1074/jbc.270.49.29270
García, S., and Heredia, N. (2006). Mycotoxins in Mexico: epidemiology, management, and control strategies. Mycopathologia 162, 255–264. doi: 10.1007/s11046-006-0058-1
Gasiewicz, T. A., and Bauman, P. A. (1987). Heterogeneity of the rat hepatic Ah receptor and evidence for transformation in vitro and in vivo. J. Biol. Chem. 262, 2116–2120.
Gasiewicz, T. A., Henry, E. C., and Collins, L. L. (2008). Expression and activity of aryl hydrocarbon receptors in development and cancer. Crit. Rev. Eukaryot. Gene Expr. 18, 279–321. doi: 10.1615/CritRevEukarGeneExpr.v18.i4.10
Gelderblom, W. C., Jaskiewicz, K., Marasas, W. F., Thiel, P. G., Horak, R. M., Vleggaar, R., et al. (1988). Fumonisins-novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 54, 1806–1811.
Gil-Serna, Y., Vázquez, C., González-Jaén, M. T., and Patiño, B. (2014). “Discrimination of the main ochratoxin A-producing species,” in Encyclopedia of Food Microbiology, 2nd Edn, Vol. 1, eds C. A. Batt, and M. A. Tortorello (London: Elsevier), 887–892.
Gu, Y.-Z., Hogenesch, J. B., and Bradfield, C. A. (2000). The PAS superfamily: sensors of environmental and developmental signals. Annu. Rev. Pharmacol. Toxicol. 40, 519–561. doi: 10.1146/annurev.pharmtox.40.1.519
Hahn, M. E. (2002). Aryl hydrocarbon receptors: diversity and evolution. Chem. Biol. Interact. 141, 131–160. doi: 10.1016/S0009-2797(02)00070-4
Hanioka, N., Nonaka, Y., Saito, K., Negishi, T., Okamoto, K., Kataoka, H., et al. (2012). Effect of aflatoxin B1 on UDP-glucuronosyltransferase mRNA expression in HepG2 cells. Chemosphere 89, 526–529. doi: 10.1016/j.chemosphere.2012.05.039
Harper, J. W., Elledge, S. J., Keyomarsi, K., Dynlacht, B., Tsai, L. H., Zhang, P., et al. (1995). Inhibition of cyclin-dependent kinases by p21. Mol. Biol. Cell 6, 387–400. doi: 10.1091/mbc.6.4.387
Harper, P. A., Riddick, D. S., and Okey, A. B. (2006). Regulating the regulator: factors that control levels and activity of the aryl hydrocarbon receptor. Biochem. Pharmacol. 72, 267–279. doi: 10.1016/j.bcp.2006.01.007
Hendricks, J. D. (1994). “Carcinogenicity of aflatoxins in non-mammalian organisms,” in The Toxicology of Aflatoxins: Human Health, Veterinary, and Agricultural Significance, eds J. D. Eaton, and D. L. Groopman (New York, NY: Academic Press), 103–136. doi: 10.1016/b978-0-12-228255-3.50011-8
Hoffman, E. C., Reyes, H., Chu, F. F., Sander, F., Conley, L. H., Brooks, B. A., et al. (1991). Cloning of a factor required for activity of the Ah (dioxin) receptor. Science 252, 954–958. doi: 10.1126/science.1852076
Huang, G., and Elferink, C. J. (2012). A novel nonconsensus xenobiotic response element capable of mediating aryl hydrocarbon receptor-dependent gene expression. Mol. Pharmacol. 81, 338–347. doi: 10.1124/mol.111.075952
IARC (2002). Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. IARC Monogr. Eval. Carcinog. Risks Hum. 82, 1–556.
Ikuta, T., Eguchi, H., Tachibana, T., Yoneda, Y., and Kawajiri, K. (1998). Nuclear localization and export signals of the human aryl hydrocarbon receptor. J. Biol. Chem. 273, 2895–2904. doi: 10.1074/jbc.273.5.2895
Jackson, D. P., Joshi, A. D., and Elferink, C. J. (2015). Ah receptor pathway intricacies, signaling through diverse protein partners and DNA-motifs. Toxicol. Res. 4, 1143–1158. doi: 10.1039/C4TX00236A
Jackson, D. P., Li, H., Mitchell, K. A., Joshi, A. D., and Elferink, C. J. (2014). Ah receptor-mediated suppression of liver regeneration through NC-XRE-driven p21Cip1 expression. Mol. Pharmacol. 85, 533–541. doi: 10.1124/mol.113.089730
Jeng, Y.-M., and Hsu, H.-C. (2003). KLF6, a putative tumor suppressor gene, is mutated in astrocytic gliomas. Int. J. Cancer 105, 625–629. doi: 10.1002/ijc.11123
Jones, P. B., Galeazzi, D. R., Fisher, J. M., and Whitlock, J. P. (1985). Control of cytochrome P1-450 gene expression by dioxin. Science 227, 1499–1502. doi: 10.1126/science.3856321
Jones, P. B., Miller, A. G., Israel, D. I., Galeazzi, D. R., and Whitlock, J. P. (1984). Biochemical and genetic analysis of variant mouse hepatoma cells which overtranscribe the cytochrome P1-450 gene in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 259, 12357–12363.
Kaestner, K. H., Hiemisch, H., Luckow, B., and Schütz, G. (1994). The HNF-3 gene family of transcription factors in mice: gene structure, cDNA sequence, and mRNA distribution. Genomics 20, 377–385. doi: 10.1006/geno.1994.1191
Kensler, J. D., Roebuck, T. W., Wogan, B. D., and Groopman, G. N. (2011). Aflatoxin: a 50-year odyssey of mechanistic and translational toxicology. Tox. Sci. 120, S28–S48. doi: 10.1093/toxsci/kfq283
Kimmig, J., and Schulz, K. H. (1957). Occupational acne (so-called chloracne) due to chlorinated aromatic cyclic ethers. Dermatologica 115, 540–546.
Kociba, R. J., Keyes, D. G., Beyer, J. E., Carreon, R. M., Wade, C. E., Dittenber, D. A., et al. (1978). Results of a two-year chronic toxicity and oncogenicity study of 2,3,7,8-tetrachlorodibenzo-p-dioxin in rats. Toxicol. Appl. Pharmacol. 46, 279–303. doi: 10.1016/0041-008X(78)90075-3
Koliopanos, A., Kleeff, J., Xiao, Y., Safe, S., Zimmermann, A., Büchler, M. W., et al. (2002). Increased arylhydrocarbon receptor expression offers a potential therapeutic target for pancreatic cancer. Oncogene 21, 6059–6070. doi: 10.1038/sj.onc.1205633
Korkalainen, M., Lindén, J., Tuomisto, J., and Pohjanvirta, R. (2005). Effect of TCDD on mRNA expression of genes encoding bHLH/PAS proteins in rat hypothalamus. Toxicology 208, 1–11. doi: 10.1016/j.tox.2004.11.003
Krska, R., Schubert-Ullrich, P., Molinelli, A., Sulyok, M., MacDonald, S., and Crews, C. (2008). Mycotoxin analysis: an update. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 25, 152–163. doi: 10.1080/02652030701765723
Kuhn, D. M., and Ghannoum, M. A. (2003). Indoor mold, toxigenic fungi, and Stachybotrys chartarum: infectious disease perspective. Clin. Microbiol. Rev. 16, 144–172. doi: 10.1128/CMR.16.1.144-172.2003
Lee, H. B., Patriarca, A., and Magan, N. (2015). Alternaria in food: ecophysiology, mycotoxin production and toxicology. Mycobiology 43, 93–106. doi: 10.5941/MYCO.2015.43.2.93
Lee, H. J., Pyo, M. C., Shin, H. S., Ryu, D., and Lee, K. W. (2018). Renal toxicity through AhR, PXR, and Nrf2 signaling pathway activation of ochratoxin A-induced oxidative stress in kidney cells. Food Chem. Toxicol. 122, 59–68. doi: 10.1016/j.fct.2018.10.00
Lewis, D. F., Dickins, M., Eddershaw, P. J., Tarbit, M. H., and Goldfarb, P. S. (1999). Cytochrome P450 substrate specificities, substrate structural templates and enzyme active site geometries. Drug Metabol. Drug Interact. 15, 1–49.
Lewis, L., Onsongo, M., Njapau, H., Schurz-Rogers, H., Luber, G., Kieszak, S., et al. (2005). Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 113, 1763–1767. doi: 10.1289/ehp.7998
Li, F. Q., Yoshizawa, T., Kawamura, O., Luo, X. Y., and Li, Y. W. (2001). Aflatoxins and fumonisins in corn from the high-incidence area for human hepatocellular carcinoma in Guangxi, China. J. Agric. Food Chem. 49, 4122–4126. doi: 10.1021/jf010143k
Lin, P., Chang, H., Tsai, W. T., Wu, M. H., Liao, Y. S., Chen, J. T., et al. (2003). Overexpression of aryl hydrocarbon receptor in human lung carcinomas. Toxicol. Pathol. 31, 22–30. doi: 10.1080/01926230390173824
Lo, R. S., and Massagué, J. (1999). Ubiquitin-dependent degradation of TGF-beta-activated smad2. Nat. Cell Biol. 1, 472–478. doi: 10.1038/70258
Ma, Q. (2002). Induction and superinduction of 2,3,7,8-tetrachlorodibenzo-rho-dioxin-inducible poly(ADP-ribose) polymerase: role of the aryl hydrocarbon receptor/aryl hydrocarbon receptor nuclear translocator transcription activation domains and a labile transcription r. Arch. Biochem. Biophys. 404, 309–316. doi: 10.1016/S0003-9861(02)00339-9
Ma, Q., and Baldwin, K. T. (2000). 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced degradation of aryl hydrocarbon receptor (AhR) by the ubiquitin-proteasome pathway. Role of the transcription activaton and DNA binding of AhR. J. Biol. Chem. 275, 8432–8438. doi: 10.1074/jbc.275.12.8432
Ma, Q., Dong, L., and Whitlock, J. P. (1995). Transcriptional activation by the mouse Ah receptor. Interplay between multiple stimulatory and inhibitory functions. J. Biol. Chem. 270, 12697–12703. doi: 10.1074/jbc.270.21.12697
Ma, Q., and Whitlock, J. P. (1997). A novel cytoplasmic protein that interacts with the Ah receptor, contains tetratricopeptide repeat motifs, and augments the transcriptional response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 272, 8878–8884. doi: 10.1074/jbc.272.14.8878
Mahon, M. J., and Gasiewicz, T. A. (1995). Ah receptor phosphorylation: localization of phosphorylation sites to the C-terminal half of the protein. Arch. Biochem. Biophys. 318, 166–174. doi: 10.1006/abbi.1995.1217
Mally, A. (2012). Ochratoxin A and mitotic disruption: mode of action analysis of renal tumor formation by ochratoxin A. Toxicol. Sci. 127, 315–330. doi: 10.1093/toxsci/kfs105
Manzini, L., Halwachs, S., Girolami, F., Badino, P., Honscha, W., and Nebbia, C. (2017). Interaction of mammary bovine ABCG2 with AFB1 and its metabolites and regulation by PCB in a MDCKII in vitro model. J. Vet. Pharmacol. Ther. 40, 591–598. doi: 10.1111/jvp.12397
Marasas, W. F. (2001). Discovery and occurrence of the fumonisins: a historical perspective. Environ. Health Perspect. 109(Suppl. 2), 239–243. doi: 10.1289/ehp.01109s2239
Marin, V., Ramos, S., Cano-Sancho, A. J., and Sanchis, G. (2013). Mycotoxins: occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 60, 218–237. doi: 10.1016/j.fct.2013.07.047
Martínez-Larrañaga, M. R., Anadon, A., Díaz, M. J., Fernández, R., Sevil, B., Fernández-Cruz, M. L., et al. (1996). Induction of cytochrome P4501A1 and P4504A1 activities and peroxisomal proliferation by fumonisin B1. Toxicol. Appl. Pharmacol. 141, 185–194. doi: 10.1006/taap.1996.0275
Mary, V. S., Theumer, M. G., Arias, S. L., and Rubinstein, H. R. (2012). Reactive oxygen species sources and biomolecular oxidative damage induced by aflatoxin B1 and fumonisin B1 in rat spleen mononuclear cells. Toxicology 302, 299–307. doi: 10.1016/j.tox.2012.08.012
Mary, V. S., Valdehita, A., Navas, J. M., Rubinstein, H. R., and Fernández-Cruz, M. L. (2015). Effects of aflatoxin B1, fumonisin B1 and their mixture on the aryl hydrocarbon receptor and cytochrome P450 1A induction. Food Chem. Toxicol. 75, 104–111. doi: 10.1016/j.fct.2014.10.030
McEvilly, R., Erkman, J. L., Luo, L., Sawchenko, P. E., Ryan, A. F., and Rosenfeld, M. G. (1996). Requirement for Brn-3.0 in differentiation and survival of sensory and motor neurons. Nature 384, 574–577. doi: 10.1038/384574a0
McKinley, E. R., and Carlton, W. W. (1980). Patulin mycotoxicosis in Swiss ICR mice. Food Cosmet. Toxicol. 18, 181–187. doi: 10.1016/0015-6264(80)90072-3
Merrick, B. A., Phadke, D. P., Auerbach, S. S., Mav, D., Stiegelmeyer, S. M., Shah, R. R., et al. (2013). RNA-Seq profiling reveals novel hepatic gene expression pattern in aflatoxin B1 treated rats. PLoS One 8:61768. doi: 10.1371/journal.pone.0061768
Meyer, B. K., Pray-Grant, M. G., Vanden Heuvel, J. P., and Perdew, G. H. (1998). Hepatitis B virus X-associated protein 2 is a subunit of the unliganded aryl hydrocarbon receptor core complex and exhibits transcriptional enhancer activity. Mol. Cell Biol. 18, 978–988. doi: 10.1128/mcb.18.2.978
Missmer, S. A., Suarez, L., Felkner, M., Wang, E., Merrill, A. H. Jr., Rothman, K. J., et al. (2006). Exposure to fumonisins and the occurrence of neural tube defects along the Texas-Mexico border. Environ. Health Perspect. 114, 237–241. doi: 10.1289/ehp.8221
Mitchell, K. A., Lockhart, C. A., Huang, G., and Elferink, C. J. (2006). Sustained aryl hydrocarbon receptor activity attenuates liver regeneration. Mol. Pharmacol. 2006, 163–170. doi: 10.1124/mol.106.023465
Morasso, M. I., Grinberg, A., Robinson, G. T., Sargent, D., and Mahon, K. A. (1999). Placental failure in mice lacking the homeobox gene Dlx3. Proc. Natl. Acad. Sci. U.S.A. 96, 162–167. doi: 10.1073/pnas.96.1.162
Nebert, D. W. (1986). The 1986 bernard B. brodie award lecture. The genetic regulation of drug-metabolizing enzymes. Drug Metab. Dispos. 16, 1–8.
Nebert, D. W., and Bausserman, L. L. (1970). Genetic differences in the extent of aryl hydrocarbon hydroxylase induction in mouse fetal cell cultures. J. Biol. Chem. 245, 6373–6382.
Nebert, D. W., Goujon, F. M., and Gielen, J. E. (1972). Aryl hydrocarbon hydroxylase induction by polycyclic hydrocarbons: simple autosomal dominant trait in the mouse. Nat. New Biol. 236, 107–110. doi: 10.1038/newbio236107a0
Nebert, D. W., Negishi, M., Lang, M. A., Hjelmeland, L. M., and Eisen, H. J. (1982). The Ah locus, a multigene family necessary for survival in a chemically adverse environment: comparison with the immune system. Adv. Genet. 21, 1–52. doi: 10.1016/S0065-2660(08)60296-5
Ostry, V. (2008). Alternaria mycotoxins: an overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 1, 175–188. doi: 10.3920/WMJ2008.x013
Pahlke, G., Tiessen, C., Domnanich, K., Kahle, N., Groh, I. A., Schreck, I., et al. (2016). Impact of Alternaria toxins on CYP1A1 expression in different human tumor cells and relevance for genotoxicity. Toxicol. Lett. 240, 93–104. doi: 10.1016/j.toxlet.2015.10.003
Peters, J. M., Narotsky, M. G., Elizondo, G., Fernandez-Salguero, P. M., Gonzalez, F. J., and Abbott, B. D. (1999). Amelioration of TCDD-induced teratogenesis in aryl hydrocarbon receptor (AhR)-null mice. Toxicol. Sci. 47, 86–92. doi: 10.1093/toxsci/47.1.86.E
Pfeiffer, E., Eschbach, S., and Metzler, M. (2007). Alternaria toxins: DNA strand-breaking activity in mammalian cells in vitro. Mycotoxin Res. 23, 152–157. doi: 10.1007/BF02951512
Pfohl-Leszkowicz, A., and Manderville, R. A. (2012). An update on direct genotoxicity as molecular mechanism of ochratoxin A carcinogenicity. Chem. Res. Toxicol. 25, 252–262. doi: 10.1021/tx200430f
Philipsen, S., and Suske, G. (1999). A tale of three fingers: the family of mammalian Sp/XKLF transcription factors. Nucleic Acids Res. 27, 2991–3000. doi: 10.1093/nar/27.15.2991
Piqué, E., Vargas-Murga, L., Gómez-Catalán, J., Lapuente, J. D., and Llobet, J. M. (2013). Occurrence of patulin in organic and conventional apple-based food marketed in Catalonia and exposure assessment. Food Chem. Toxicol. 60, 199–204. doi: 10.1016/j.fct.2013.07.052
Pohjanvirta, R. (2011). The Ah Receptor in Biology and Toxicology. New Jersey, NJ: Wiley, 3–32. doi: 10.1002/9781118140574
Poland, A., and Glover, E. (1973). 2,3,7,8-Tetrachlorodibenzo-p-dioxin: a potent inducer of -aminolevulinic acid synthetase. Science 179, 476–477. doi: 10.1126/science.179.4072.476
Poland, A., and Glover, E. (1974). Comparison of 2,3,7,8-tetrachlorodibenzo-p-dioxin, a potent inducer of aryl hydrocarbon hydroxylase, with 3-methylcholanthrene. Mol. Pharmacol. 10, 349–359.
Poland, A., Glover, E., and Kende, A. S. (1976). Stereospecific, high affinity binding of 2,3,7,8-tetrachlorodibenzo-p-dioxin by hepatic cytosol. Evidence that the binding species is receptor for induction of aryl hydrocarbon hydroxylase. J. Biol. Chem. 251, 4936–4946.
Pot, C. (2012). Aryl hydrocarbon receptor controls regulatory CD4+ T cell function. Swiss Med. Wkly. 142:w13592. doi: 10.4414/smw.2012.13592
Reisz-Porszasz, S. M., Probst, R., Fukunaga, B. N., and Hankinson, O. (1994). Identification of functional domains of the aryl hydrocarbon receptor nuclear translocator protein (ARNT). Mol. Cell. Biol. 14, 6075–6086. doi: 10.1128/mcb.14.9.6075
Richardson, H. L., Stier, A. R., and Borsos-Nachtnebel, E. (1952). Liver tumor inhibition and adrenal histologic responses in rats to which 3’-methyl-4-dimethylaminoazobenzene and 20-methylcholanthrene were simultaneously administered. Cancer Res. 12, 356–361.
Roberts, B. J., and Whitelaw, M. L. (1999). Degradation of the basic helix-loop-helix/Per-ARNT-Sim homology domain dioxin receptor via the ubiquitin/proteasome pathway. J. Biol. Chem. 274, 36351–36356. doi: 10.1074/jbc.274.51.36351
Rubinstein, M., Idelman, G., Plymate, S. R., Narla, G., Friedman, S. L., and Werner, H. (2004). Transcriptional activation of the insulin-like growth factor I receptor gene by the Kruppel-like factor 6 (KLF6) tumor suppressor protein: potential interactions between KLF6 and p53. Endocrinology 145, 3769–3777. doi: 10.1210/en.2004-0173
Samson, R. A., Houbraken, J., Varga, J., and Frisvad, J. C. (2009). Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 22, 14–27. doi: 10.3767/003158509X418925
Samson, R. A., Noonim, P., Meijer, M., Houbraken, J., Frisvad, J. C., and Varga, J. (2007). Diagnostic tools to identify black Aspergilli. Stud. Mycol. 59, 129–145. doi: 10.3114/sim.2007.59.13
Schobert, R., and Schlenk, A. (2008). Tetramic and tetronic acids: an update on new derivatives and biological aspects. Bioorg. Med. Chem. 16, 4203–4221. doi: 10.1016/j.bmc.2008.02.069
Schrader, T. J., Cherry, W., Soper, K., Langlois, I., and Vijay, H. M. (2001). Examination of Alternaria alternata mutagenicity and effects of nitrosylation using the Ames salmonella test. Teratog. Carcinog. Mutagen. 21, 261–274. doi: 10.1002/tcm.1014
Schreck, I., Deigendesch, U., Burkhardt, B., Marko, D., and Weiss, C. (2012). The Alternaria mycotoxins alternariol and alternariol methyl ether induce cytochrome P450 1A1 and apoptosis in murine hepatoma cells dependent on the aryl hydrocarbon receptor. Arch. Toxicol. 86, 625–632. doi: 10.1007/s00204-011-0781-3
Schreiber, S. L. (1991). Chemistry and biology of the immunophilins and their immunosuppressive ligands. Science 251, 283–287. doi: 10.1126/science.1702904
Schultz, K. H. (1957). Clinical and experimental studies on the etiology of chloracne. Arch. Klin. Exp. Dermatol. 206, 589–596.
Schwartz, G. G. (2002). Hypothesis: does ochratoxin A cause testicular cancer? Cancer Causes Control 13, 91–100. doi: 10.1023/A:1013973715289
Shephard, G. S. (2004). “Mycotoxins worldwide: current issues in Africa,” in Meeting the Mycotoxin Menace, eds D. Barug, H. P. van Egmond, R. Lopez-Garcia, W. A. van Osenbruggen, and A. Visconti (Wageningen: Wageningen Academic Publishers), 81–88.
Siegel, I. N., Merkel, S., Koch, M., and Nehls, I. (2010). Quantification of the Alternaria mycotoxin tenuazonic acid in beer. Food Chem. 120, 902–906. doi: 10.1016/j.foodchem.2009.10.070
Siraj, M. Y., Hayes, A. W., Takanaka, A., and Ho, I. K. (1980). Effect of patulin on hepatic monooxygenase in male mice. J Environ Pathol Toxicol. 4, 545–553.
Smit, E., Souza, T., Jennen, D. G. J., Kleinjans, J. C. S., and van der Beucken, T. (2017). Identificaion of essential transcription factors for adequate DNA damage response after benzo(a)pyrene and AFB1 exposure by combining transcriptomics with functional genomics. Toxicology 390, 74–82. doi: 10.1016/j.tox.2017.09.002
Solfrizzo, M., Gambacorta, L., Lattanzio, V. M. T., Powers, S., and Visconti, A. (2011). Simultaneous LC-MS/MS determination of aflatoxin M1, ochratoxin A, deoxynivalenol, de-epoxydeoxynivalenol, α and β-zearalenols and fumonisin B1 in urine as a multi-biomarker method to assess exposure to mycotoxins. Anal. Bioanal. Chem. 401, 2831–2841. doi: 10.1007/s00216-011-5354-z
Soshilov, A., and Denison, M. S. (2008). Role of the Per/Arnt/Sim domains in ligand-dependent transformation of the aryl hydrocarbon receptor. J. Biol. Chem. 283, 32995–33005. doi: 10.1074/jbc.M802414200
Stevens, V. L., and Tang, J. (1997). Fumonisin B1-induced sphingolipid depletion inhibits vitamin uptake via the glycosylphosphatidylinositol-anchored folate receptor. J. Biol. Chem. 272, 18020–18025. doi: 10.1074/jbc.272.29.18020
Steyn, P. S. (1995). Mycotoxins, general view, chemistry and structure. Toxicol. Lett. 8, 843–851. doi: 10.1016/0378-4274(95)03525-7
Tan, Z., Huang, M., Puga, A., and Xia, Y. (2004). A critical role for MAP kinases in the control of Ah receptor complex activity. Toxicol. Sci. 82, 80–87. doi: 10.1093/toxsci/kfh228
Tanaka, G., Kanaji, S., Hirano, A., Arima, K., Shinagawa, A., Goda, C., et al. (2005). Induction and activation of the aryl hydrocarbon receptor by IL-4 in B cells. Int. Immunol. 17, 797–805. doi: 10.1093/intimm/dxh260
Tangni, E. K., Ponchaut, S., Maudoux, M., Rozenberg, R., and Larondelle, Y. (2002). Ochratoxin A in domestic and imported beers in Belgium: occurrence and exposure assessment. Food Addit. Contam. 19, 1169–1179. doi: 10.1080/02652030210007859
Tijet, N., Boutros, P. C., Moffat, I. D., Okey, A. B., Tuomisto, J., and Pohjanvirta, R. (2006). Aryl hydrocarbon receptor regulates distinct dioxin-dependent and dioxin-independent gene batteries. Mol. Pharmacol. 69, 140–153. doi: 10.1124/mol.105.018705
Turkez, H., and Geyikoglu, F. (2010). Boric acid: a potential chemoprotective agent against aflatoxin b(1) toxicity in human blood. Cytotechnology 62, 157–165. doi: 10.1007/s10616-010-9272-2
Ueno, Y., Iijima, K., Wang, S. D., Sugiura, Y., Sekijima, M., Tanaka, T., et al. (1997). Fumonisins as a possible contributory risk factor for primary liver cancer: a 3-year study of corn harvested in Haimen, China, by HPLC and ELISA. Food Chem. Toxicol. 35, 1143–1150. doi: 10.1016/S0278-6915(97)00113-0
Valencia-Quintana, R., Sánchez-Alarcón, J., Tenorio, M. G., Deng, Y., Waliszewski, S. M., and Valera, M. Á. (2012). Preventive strategies aimed at reducing the health risks of Aflatoxin B1. Toxicol. Environ. Health Sci. 4, 71–79. doi: 10.1007/s13530-012-0138-1
Valencia-Quintana, R., Sánchez-Alarcón, J., Tenorio-Arvide, M. G., Deng, Y., Montiel-González, J. M., Gómez-Arroyo, S., et al. (2014). The microRNAs as potential biomarkers for predicting the onset of aflatoxin exposure in human beings: a review. Front. Microbiol. 5:102. doi: 10.3389/fmicb.2014.00102
Vettorazzi, A., van Delft, J., and López de Carain, A. (2013). A review on ochratoxin A transcriptomic studies. Food Chem. Toxicol. 59, 766–783. doi: 10.1016/j.fct.2013.05.043
Visagie, C. M., Varga, J., Houbraken, J., Meijer, M., Kocsubé, S., Yilmaz, N., et al. (2014). Ochratoxin production and taxonomy of the yellow Aspergilli (Aspergillus section Circumdati). Stud. Mycol. 78, 1–61. doi: 10.1016/j.simyco.2014.07.001
Voss, K. A., Smith, G. W., and Haschek, W. M. (2007). Fumonisins: toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 137, 299–325. doi: 10.1016/j.anifeedsci.2007.06.007
Wilson, S. R., Joshi, A. D., and Elferink, C. J. (2013). The tumor suppressor Kruppel-like factor 6 is a novel aryl hydrocarbon receptor DNA binding partner. J. Pharmacol. Exp. Ther. 345, 419–429. doi: 10.1124/jpet.113.203786
Wu, F., Groopman, J. D., and Pestka, J. J. (2014). Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 5, 351–372. doi: 10.1146/annurev-food-030713-092431
Yueh, M.-F., Huang, Y.-H., Hiller, A., Chen, S., Nguyen, N., and Tukey, R. H. (2003). Involvement of the xenobiotic response element (XRE) in Ah receptor-mediated induction of human UDP-glucuronosyltransferase 1A1. J. Biol. Chem. 278, 15001–15006. doi: 10.1074/jbc.M300645200
Zain, M. E. (2011). Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 15, 129–144. doi: 10.1016/j.jscs.2010.06.006
Keywords: Aspergillus, Alternaria, AFB1, alternariol (AOH), CYP1A1
Citation: Arenas-Huertero F, Zaragoza-Ojeda M, Sánchez-Alarcón J, Milić M, Šegvić Klarić M, Montiel-González JM and Valencia-Quintana R (2019) Involvement of Ahr Pathway in Toxicity of Aflatoxins and Other Mycotoxins. Front. Microbiol. 10:2347. doi: 10.3389/fmicb.2019.02347
Received: 07 August 2019; Accepted: 26 September 2019;
Published: 10 October 2019.
Edited by:
Mehdi Razzaghi-Abyaneh, Pasteur Institute of Iran (PII), IranReviewed by:
Thomas Haarmann-Stemmann, Leibniz-Institut für Umweltmedizinische Forschung (IUF), GermanyRichard Manderville, University of Guelph, Canada
Copyright © 2019 Arenas-Huertero, Zaragoza-Ojeda, Sánchez-Alarcón, Milić, Šegvić Klarić, Montiel-González and Valencia-Quintana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco Arenas-Huertero, ZmFyZW5hc2h1ZXJ0ZXJvQHlhaG9vLmNvbS5teA==; Rafael Valencia-Quintana, cHJ2cTIwMDRAeWFob28uY29tLm14