 Fernando Chimal-Cázares1,2†
Fernando Chimal-Cázares1,2† Gabriela Hernández-Martínez1,2†
Gabriela Hernández-Martínez1,2† Sabino Pacheco3
Sabino Pacheco3 Miguel A. Ares1
Miguel A. Ares1 Jorge Soria-Bustos1,2Manuel Sánchez-Gutiérrez4Jeannett A. Izquierdo-Vega4
Jorge Soria-Bustos1,2Manuel Sánchez-Gutiérrez4Jeannett A. Izquierdo-Vega4 Jose Antonio Ibarra2
Jose Antonio Ibarra2 Jorge A. González-y-Merchand2
Jorge A. González-y-Merchand2 Jean-Pierre Gorvel5
Jean-Pierre Gorvel5 Stéphane Méresse5
Stéphane Méresse5 Miguel A. De la Cruz1*
Miguel A. De la Cruz1*- 1Unidad de Investigación Médica en Enfermedades Infecciosas y Parasitarias, Hospital de Pediatría, Centro Médico Nacional Siglo XXI, Instituto Mexicano del Seguro Social, Mexico City, Mexico
- 2Departamento de Microbiología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Mexico City, Mexico
- 3Departamento de Microbiología Molecular, Instituto de Biotecnología, Universidad Nacional Autónoma de México, Cuernavaca, Mexico
- 4Instituto de Ciencias de la Salud, Universidad Autónoma del Estado de Hidalgo, Pachuca, Mexico
- 5Aix Marseille Univ, CNRS, INSERM, CIML, Marseille, France
Salmonella enterica serotype Typhimurium is a bacterium that causes gastroenteritis and diarrhea in humans. The genome of S. Typhimurium codes for diverse virulence factors, among which are the toxin-antitoxin (TA) systems. SehAB is a type II TA, where SehA is the toxin and SehB is the antitoxin. It was previously reported that the absence of the SehB antitoxin affects the growth of S. Typhimurium. In addition, the SehB antitoxin can interact directly with the SehA toxin neutralizing its toxic effect as well as repressing its own expression. We identified conserved residues on SehB homologous proteins. Point mutations were introduced at both N- and C-terminal of SehB antitoxin to analyze the effect of these changes on its transcription repressor function, on its ability to form homodimers and on the virulence of S. Typhimurium. All changes in amino acid residues at both the N- and C-terminal affected the repressor function of SehB antitoxin and they were required for DNA-binding activity. Mutations in the amino acid residues at the N-terminal showed a lower capacity for homodimer formation of the SehB protein. However, none of the SehB point mutants were affected in the interaction with the SehA toxin. In terms of virulence, the eight single-amino acid mutations were attenuated for virulence in the mouse model. In agreement with our results, the eight amino acid residues of SehB antitoxin were required for its repressor activity, affecting both homodimerization and DNA-binding activity, supporting the notion that both activities of SehB antitoxin are required to confer virulence to Salmonella enterica.
Introduction
Salmonella enterica comprises Gram-negative bacteria with around 2500 serotypes responsible for severe gastroenteritis and systemic infections in warm-blooded animals, including humans (Haraga et al., 2008; Jajere, 2019). By horizontal gene transfer, S. enterica has evolved to adapt to both extracellular and intracellular conditions, expressing a myriad of virulence factors, such as secretion systems, adhesins, flagella, and toxins, among others (Fabrega and Vila, 2013). In this way, toxin-antitoxin systems have emerged as important elements that affect the pathogenicity of many bacteria, regulating their physiology, persistence and virulence (Lobato-Marquez et al., 2016; Harms et al., 2018; Ronneau and Helaine, 2019). Six types of TA systems are currently described, classified mainly according to the ribonucleic or proteinaceous structure of the antitoxin and how it antagonizes the cognate toxin (Page and Peti, 2016). In type I and III, the antitoxin is an RNA molecule, which directly binds to RNA and protein, respectively, while in the others (II, IV, V, and VI), the antitoxin is a protein that directly or indirectly neutralizes the expression or activity of the toxin (Lobato-Marquez et al., 2016). Especially in the type II TA systems, both toxin and antitoxin are proteins and are transcriptionally organized in a bicistronic operon, where mostly the antitoxin gene is upstream of the toxin gene (Yamaguchi and Inouye, 2009).
While type II toxins possess a variety of enzymatic functions (ribonuclease, kinase, adenylyl-, and acetyl-transferase) (Mutschler et al., 2011; Yamaguchi et al., 2011; Harms et al., 2015; Cheverton et al., 2016; Rocker and Meinhart, 2016), type II antitoxins interact with the cognate toxins and also act as transcription repressors of their own expression (Yamaguchi and Inouye, 2011). Most type II antitoxins possess a DNA-binding domain located at the N-terminal and a region involved in homodimerization at the C-terminal (Chan et al., 2016). The higBA (host inhibition of growth) TA genes code for type II systems, which are widely prevalent in bacteria (Pandey and Gerdes, 2005), where HigB is a toxin that cleaves mRNAs in the presence of ribosomes, and HigA is the antitoxin that neutralizes the toxic effect of HigB (Hurley and Woychik, 2009). Contrary to other type II TA genes, in the higBA bicistronic operon the higB toxin gene is upstream of the higA antitoxin gene. Moreover, HigA-type antitoxins in Escherichia coli and Shigella flexneri, unlike Proteus vulgaris and Pseudomonas aeruginosa, present an unusual organization possessing the homo-heterodimerization region and DNA-binding domain at the N- and C-terminal, respectively (Schureck et al., 2014; Yang et al., 2016; Liu et al., 2019; Xu et al., 2019; Yoon et al., 2019).
In Salmonella enterica serotype Typhimurium (S. Typhimurium), SehAB is a type II TA system homologous to HigBA, where SehA is the toxin and SehB is the antitoxin. SehB protein contains 142 amino acids and has a molecular weight and isoelectric point of 15.92 kDa and 5.56, respectively. SehB antitoxin represses its own expression by direct binding to its promoter region, and a SehB homodimer was observed in solution (De la Cruz et al., 2013). In terms of virulence, the absence of SehB dramatically affects the virulence of S. Typhimurium in mice (De la Cruz et al., 2013).
In this work we identified conserved residues of the SehB antitoxin by alignment of amino acid sequence of homologous proteins. Eight plasmid constructs expressing point mutations of the SehB protein located at the N- and C-terminal were generated. All of these mutant proteins were unable to bind to their promoter region, thus affecting the ability of the SehB antitoxin to repress its own expression. This DNA-binding activity was impaired both due to the lack of SehB homodimerization caused by mutations at the N-terminal and by alterations of the helix-turn-helix (HTH)-type domain located at the C-terminal. All SehB amino acid tested were dispensable for the interaction with the SehA toxin. Interestingly, these amino acid residues of the SehB antitoxin were required for the virulence of S. Typhimurium. The functional characterization of the SehB protein showed that Y32, L42, L52, I60, S107, L121, L129, and F140 amino acids are involved in different molecular mechanisms of this protein and they are important for S. Typhimurium virulence.
Materials and Methods
Bacterial Strains and Growth Conditions
Bacterial strains used in this study are listed in Table 1. Strains were cultured in LB broth. Ampicillin (200 μg/ml), kanamycin (50 μg/ml), tetracycline (10 μg/ml), and chloramphenicol (34 μg/ml) were added when required. Bacterial suspensions were prepared from overnight LB broth cultures. Then, 250 ml-flasks containing 50 ml of LB broth were inoculated with bacterial suspensions that presented an initial OD600 nm of 0.05. These cultures were incubated at 37°C in a shaking incubator at 200 rpm.

Table 1. Bacterial strains and plasmids used in this study.
Protein Structure Determination
Amino acid sequence of SehB (STM4030.S) and SehA (STM4031) of S. Typhimurium strain LT2 was used to find templates on SWISS-MODEL server (Waterhouse et al., 2018). The toxin-antitoxin HigBA from S. flexneri (PDB code: 5YCL), which had the highest score of Sequence Coverage, Identity, GMQE (Global Model Quality Estimation) and Quaternary Structure Quality Estimate (QSQE), was chosen to build the 3D-structure of SehB monomeric by homology modeling. The models of SehAB toxin-antitoxin complex and SehB homodimer were built by superimposing of structures on HigBA complex and HigA homodimer. Figures were prepared using PyMol software.
Construction of Plasmids
Plasmids and primers used in this study are listed in Tables 1, 2, respectively. To construct the plasmid pK6-SehB-WT, which carries a His6-tag at N-terminal of SehB protein, a fragment of the sehB gene was amplified by PCR from the pK6-SehB plasmid, using the primers pairs His6-SehB-NcoI-5′ and SehB-HindIII-3′. Site-specific mutations in sehB gene were introduced by overlapping PCR as previously described (Ho et al., 1989). Briefly, pairs of complementary oligonucleotides were designed (Table 2), and both mutagenic primers were combined in parallel PCRs with primers His6-SehB-NcoI-5′ and SehB-HindIII-3′, respectively, using pK6-SehB-WT plasmid as a template. The resulting PCR products were purified and mixed for a second PCR round with primers His6-SehB-NcoI-5′ and SehB-HindIII-3′, which allow the amplification of the entire sehB gene. The final PCR products were purified and digested with NcoI and HindIII and cloned into pMPM-K6 previously digested with the same enzymes, rendering plasmids pK6-SehBY-32A, pK6-SehB-Y42A, pK6-SehB-L52A, pK6-SehB-I60A, pK6-SehB-S107A, pK6-SehB-L121A, pK6-SehB-L129A, and pK6-SehB-F140A. To generate plasmids pSR658-SehB-WT, pSR658-SehB-Y32A, pSR658-SehB-Y42A, pSR658-SehB-L52A, pSR658-SehB-I60A, pSR658-SehB-S107A, pSR658-SehB-L121A, pSR658-SehB-L129A, and pSR658-SehB-F140A, fragments of the sehB gene were amplified by PCR with the primers pair SehB-XhoI-F/SehB-KpnI-R, using as template each construction that carries the sehB point mutation including the wild-type sehB gene. The PCR products were digested with XhoI and KpnI and cloned into the vector pSR658 previously digested with the same restriction enzymes. To generate the pSR659-SehA plasmid, the sehA gene was amplified by PCR using the primers SehA-XhoI-F/SehA-KpnI-R. The resulting PCR product was digested with XhoI and KpnI and cloned into the vector pSR659 digested with the same restriction enzymes. All constructs were verified by DNA sequencing.
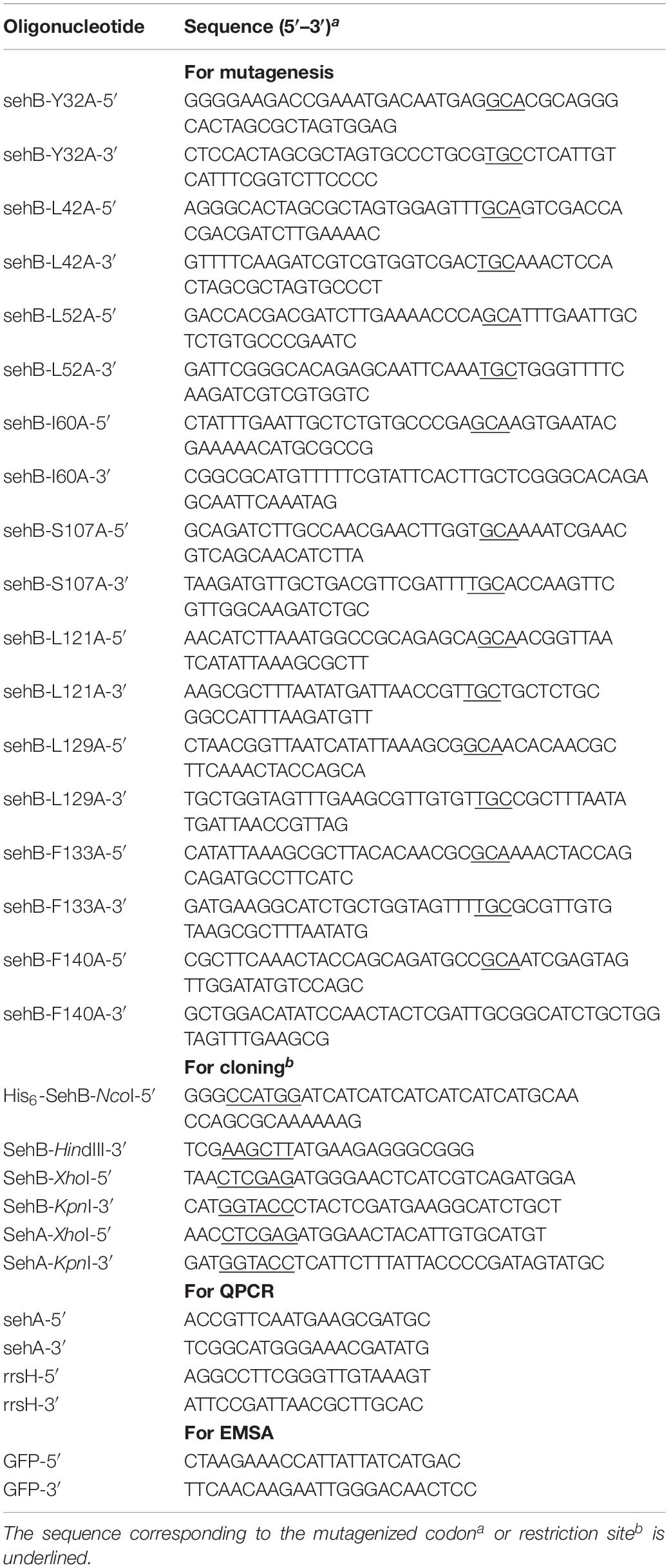
Table 2. Oligonucleotides used in this study.
Flow Cytometry Analysis of Bacteria
Bacteria grown in LB broth were treated and analyzed by flow cytometry as previously described (Aussel et al., 2011).
Quantitative RT-PCR
Total RNA was extracted from bacteria grown under different culture conditions using the hot phenol method (Jahn et al., 2008). DNA was removed with TURBO DNA-free (Ambion, Inc.) and the quality of RNA was assessed using a NanoDrop (ND-1000; Thermo Scientific) and an Agilent 2100 bioanalyzer with a Picochip (Agilent Technologies). The absence of contaminating DNA was controlled by the lack of amplification products after 35 qPCR cycles using RNA as template. Control reactions with no template (water) and with no reverse transcriptase were run in all experiments. cDNA synthesis and qPCR were performed as described (Matter et al., 2018; Ares et al., 2019). Specific primers were designed with the Primer3Plus software and they are listed in Table 2. 16S rRNA (rrsH) was used as a reference gene for normalization and the relative gene expression was calculated using the 2–ΔΔCt method (Livak and Schmittgen, 2001).
Protein Purification
His6-SehB proteins were purified as previously reported (De la Cruz et al., 2013). Briefly, E. coli BL21(DE3) carrying the pK6-SehB plasmids (Table 1) was grown to mid-logarithmic phase. L(+)-arabinose (Sigma-Aldrich) was added to a final concentration of 0.1%, and the bacteria were grown at 6 h at 30°C. Subsequently, cells were pelleted by centrifugation, resuspended in urea buffer (8 M urea, 100 mM NaH2PO4 and 10 mM Tris-HCl, pH 8.0) and disrupted by sonication. This suspension was centrifuged, and the supernatant was filtered through a Ni-nitrilotriacetic acid agarose column (Qiagen) pre-equilibrated with urea buffer. After extensive washing with binding buffer containing 50 mM imidazole (200 ml), proteins were eluted with 500 mM imidazole (10 ml). Fractions were analyzed by SDS-PAGE. Protein concentration was determined by the Bradford procedure. Aliquots of each purified protein were stored at −70°C.
Electrophoretic Mobility Shift Assay (EMSA)
EMSA experiments were performed as described previously (De la Cruz et al., 2007). PCR product corresponding to sehAB promoter region was amplified from sehAB-gfp plasmid using primers GFP-5′ and GFP-3′, which hybridize to the pFPV25 vector. This fragment (100 ng), was mixed with increasing concentrations of wild-type and mutants His6-SehB proteins in PBS/50% glycerol. They were incubated 30 min at room temperature and then separated by electrophoresis in 6% polyacrylamide gels in Tris-borate-EDTA buffer. DNA bands were visualized by staining with ethidium bromide.
Western Blotting
Equal numbers of bacteria were used to prepare whole cell extracts and subjected to SDS-PAGE in 12% polyacrylamide gels and electroblotted onto PVDF membranes. After blocking with 5% non-fat milk, membranes were incubated with anti-His6 (abcam) and anti-LexA (Merck) polyclonal antibodies at 1:3000 and 1:10,000 dilutions, respectively. Goat anti-rabbit IgG (Sigma-Aldrich) conjugated to horseradish peroxidase (1:10,000) was used as secondary antibody, and the reactions were visualized with chemiluminescence reagents (Perkin Elmer).
SehB Dimerization Assays
To test homodimerization of SehB, the pSR658, pSR658-H-NS, pSR658-SehB-WT, pSR658-SehB-Y32A, pSR658-SehB-Y42A, pSR658-SehB-L52A, pSR658-SehB-I60A, pSR658-SehB-S107A, pSR658-SehB-L121A, pSR658-SehB-L129A, and pSR658-SehB-F140A plasmids, were transformed into the E. coli SU101 reporter strain for homodimerization assays, which carries the chromosomal sulA–lacZ transcriptional fusion (Dmitrova et al., 1998). Transformants were grown in LB with tetracycline and 1 mM IPTG to induce expression of LexADBDwt-SehB fusion proteins. Samples were collected at an OD600 nm of 1.0 and used for the determination of β-galactosidase activity.
SehB-SehA Heterodimerization Assays
To test the heterodimerization between SehA and SehB, the E. coli SU202 reporter strain (used for heterodimerization tests), which carries the sulA–lacZ transcriptional fusion with a hybrid LexA operator (Dmitrova et al., 1998) was first transformed with the plasmid pSR659-SehA, and then transformed with the pSR658-SehB-WT and mutagenic variants. Transformants were grown in LB with tetracycline and ampicillin and 1 mM IPTG was used to induce expression of LexADBDwt and LexADBDmut fusion proteins. Samples were collected at an OD600 nm of 1.0 and used for the determination of β-Galactosidase activity.
β-Galactosidase Assays
The β-Galactosidase assay and protein quantification to calculate specific activities were performed as previously described (Oropeza et al., 1999).
Intracellular Replication Assays
RAW264.7 (ATCC TIB-71) mouse macrophages were seeded at a density of 106 cells per well in 24-well tissue culture plates 24 h before use. Bacteria were cultured overnight at 37°C with shaking and were opsonized in DMEM containing FBS and 10% normal mouse serum for 30 min on ice. When pK6-SehB plasmids were used, we did not induce sehB expression from the arabinose promoter since the leaky expression that occurs is enough to complement the absence of sehB in the chromosome. Bacteria were added to cells at a multiplicity of infection (MOI) of 100. Plates were centrifuged at 500 g for 5 min at 4°C and incubated for 30 min at 37°C under an humidified 5% CO2 atmosphere. Cells were washed thrice with DMEM containing 100 μg/ml gentamicin and then incubated in this medium for 1 h to eliminate extracellular bacteria; after, the gentamicin concentration was decreased to 10 μg/ml for the remainder of the experiment. For enumeration of intracellular bacteria, macrophages were washed thrice with PBS and lysed with 0.1% Triton X-100 for 15 min, and 10-fold serial dilutions were plated onto LB agar plates to determine total CFUs. Plates were incubated overnight at 37°C, and colonies were counted. Each time point was performed in triplicate, and each experiment was performed three times.
Survival Curves of Mice Infected With S. Typhimurium
Six- to eight-week-old Balb/c female mice were perorally inoculated with 105 CFUs of WT S. Typhimurium or ΔsehB mutant transformed with different plasmids expressing both wild-type and mutants SehB proteins. When pK6-SehB plasmids were used, we did not induce sehB expression from the arabinose promoter since the leaky expression that occurs is enough to complement the absence of sehB in the chromosome. S. Typhimurium was prepared by growing 1 ml of each culture in 100 ml of LB broth for 3 h (OD600 nm = 1.0) at 37°C, followed by plating 103–106 serial dilutions in triplicate on LB agar, and placed in a 37°C incubator overnight. CFUs were enumerated the following day, and the dose was appropriately prepared to the desired concentration. Infected mice were monitored at least every 12 h during 14 days, and moribund animals were sacrificed by CO2 asphyxiation at the end of the experiment. Survival was recorded as percentage of survival every day after infection. Survival analysis was conducted using the Kaplan-Meier survival test.
Competitive Index
Competitive index experiments were performed as previously described (De la Cruz et al., 2013). Six Balb/c mice (6–8 weeks old) were inoculated perorally with equal amounts of two bacterial strains for a total of 105 bacteria per mouse. When pK6-SehB plasmids were used, we did not induce sehB expression from the arabinose promoter since the leaky expression that occurs is enough to complement the absence of sehB in the chromosome. The spleens were harvested 2 days after inoculation, and homogenized. Bacteria were recovered and enumerated after plating a dilution series onto LB agar with the appropriate antibiotics. Competitive indexes (CI) were determined for each mouse (Beuzon and Holden, 2001).
Statistical Analysis
All data were the averages of three independent experiments performed by triplicate. Data were analyzed with the GraphPad Prism 5.0 software (GraphPad Inc., San Diego, CA, United States) using two-tailed Student’s t-test. P-values of < 0.05 were considered significant.
Results
Identification of Conserved Amino Acid Residues in SehB Homologous Proteins
We began this study by searching homolog proteins of SehB antitoxin in different bacteria. Uncharacterized SehB homologs are present in serotypes of Salmonella enterica (Heidelberg, Paratyphi, Typhi, and Gallinarum), as well as in in other bacteria species such as Serratia marcescens, Xenorhabdus nematophila, Klebsiella pneumoniae, Pectobacterium carotovora, E. coli, S. flexneri, and Enterobacter cloacae (Figure 1A). By alignment of the amino acid sequence of this protein with other homologous SehB antitoxins, conserved amino acids were identified throughout the entire protein (Figure 1A). In addition, an in silico characterization of the SehB antitoxin identified both N- and C-terminal domains, which correspond to the 1-80 and 81-142 amino acid residues, respectively. We used as reference the HigA protein from E. coli K-12 and S. flexneri, which both show 46% identity and 67% similarity to SehB (Figure 1B). Each N- and C-terminal domain of SehB contain four α-helices, which correspond to a putative dimerization and a DNA binding domain, respectively, connected by the helix α5 (Figures 1B,C). Specifically, a helix-turn-helix (HTH) DNA-binding motif was found at the C-terminal from S107 to K134 amino acids (Figure 1C). Moreover, a 3D-model showed the SehB homodimer formation, displaying intermolecular interactions between both SehB monomers. To investigate further the functional role of conserved amino acids residues on the SehB antitoxin, alanine substitutions were performed at four residues of N- and C-terminal: Y32, L42, L52, I60, S107, L121, L129, and F140.
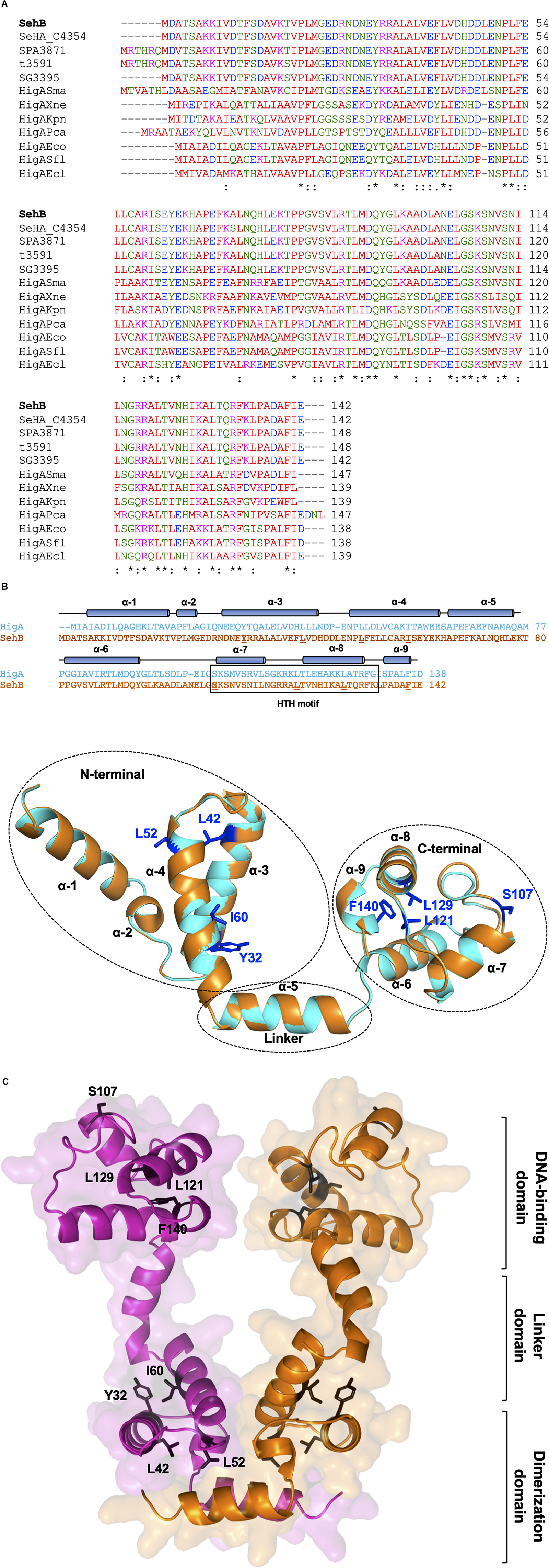
Figure 1. SehB homologs and structure prediction. (A) The amino acid sequence multialignment of SehB (STM4030.S) from S. Typhimurium strain LT2 (AAL22869.1) and homologs proteins SeHA_C4354 from S. Heidelberg strain SL476 (ACF69262.1), SPA3871 from S. Paratyphi A strain ATCC 915 (AAV79638.1), t3591 from S. Typhi strain Ty2 (AAO71094.1), SG3395 from S. Gallinarum strain 287/91 (CAR39186.1), HigASma from S. marcescens strain ATCC 13880 (KFD15043.1), HigAXne from X. nematophila strain ATCC 19061 (CBJ92634.1), HigAKpn from K. pneumoniae strain NTUH-K2044 (BAH66031.1), HigAPca from P. carotovorum strain ICMP 5702 (KML71864.1), HigAEco from E. coli strain MG1655 (NP_417553.1), HigASfl from S. flexneri strain 2457T (AAP18407.1) and HigAEcl from E. cloacae strain ATCC 13047 (YP_003613619.1), was performed using the Clustal Omega software (https://www.ebi.ac.uk/Tools/msa/clustalo/). Identical and similar residues are marked by asterisk and two points, respectively. Hydrophobic, polar, acidic and basic amino acids are colored in red, green, blue and magenta, respectively. (B) Schematic representation of the SehB predicted tertiary structure using the SWISS-MODEL server (Waterhouse et al., 2018). The cylinders represent alpha helices. Amino acids replaced by alanine are underlined. Putative HTH DNA-binding domain correspond to helices α-7 and α-8. Amino acids selected to be mutagenized are shown in the tertiary structure of SehB. (C) Model of SehB homodimer by superposition of monomeric structures on dimeric HigA using PyMol software. Both N- and C-terminal domains are depicted in the figure. Y32, L42, L52, I60, S107, L121, L129, and F140 residues are indicated in one SehB monomer. SehB monomers are colored in magenta and orange.
Point Mutations at Both N- and C-Terminal Affect the Repressor Function of SehB
We previously reported that SehB antitoxin acts as a transcriptional repressor of its own expression (De la Cruz et al., 2013). In order to investigate further the role of amino acids on SehB transcription activity, we generated eight plasmid constructions, each of them containing the sehB gene with single point mutations. Hence, these plasmids were introduced into the ΔsehB mutant and the repressor function of the SehB antitoxin on its own expression was evaluated using sehAB-gfp plasmid fusion. Of note, we did not induce sehB expression from the arabinose promoter since the leaky expression that occurs is enough to complement the absence of sehB in the chromosome (Figure 2). All of these strains displayed growth similar to that of the SehB-WT strain, indicating that mutations in the N- and C-terminal domains of SehB did not affect the function of the SehB protein related to bacterial growth (Figure 2A). Unlike the complemented ΔsehB mutant expressing the SehB-WT protein, none of the point mutants repressed the transcription of the sehAB-gfp fusion (Figure 2B). To corroborate the role of SehB mutant proteins on the repression of the sehAB bicistronic operon, chromosomal expression of the sehA toxin gene was quantified by RT-qPCR. Similar to plasmid reporter fusion, none of SehB mutant proteins were able to complement the levels of sehA mRNA compared to the wild-type strain (Figure 2C). Western blotting experiments showed that all His6-SehB proteins were produced, indicating that the lack of repressor activity of SehB mutagenic variants was not due to the amount of the proteins (Figure 2D). Our observations show that Y32, L42, L52, I60, S107, L121, L129, and F140 amino acids are relevant for the transcription repression activity of the SehB antitoxin on its own promoter region.
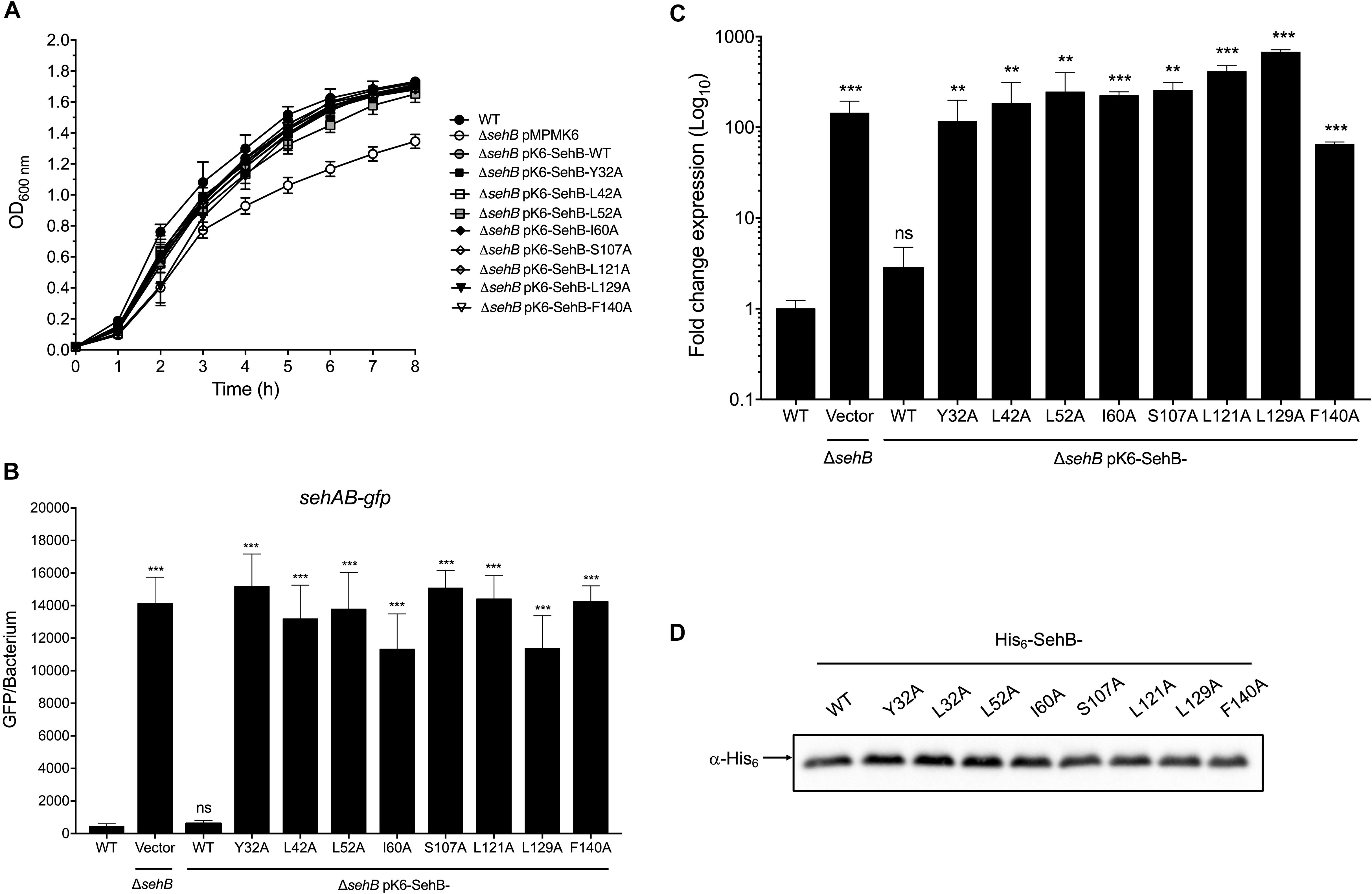
Figure 2. Effect of SehB point mutations on the transcription repression of sehAB. (A) Growth kinetics of S. Typhimurium wild-type (WT), ΔsehB pMPM-K6 and ΔsehB expressing SehB-WT and SehB point mutants. Bacteria were grown at 37°C in LB broth and the optical density was monitored at 600 nm. (B) S. Typhimurium wild-type (WT), ΔsehB pMPM-K6 and ΔsehB expressing the SehB-WT and SehB point mutants carrying a transcriptional fusion sehAB-gfp were grown in LB broth and samples were taken at a final OD600nm of 1.5. The relative fluorescence intensity of bacteria was determined by flow cytometry. 10,000 bacteria were analyzed to calculate the mean GFP/bacterium. (C) Transcription of sehA gene in LB broth at early stationary phase (OD600 nm = 1.5) at 37°C was quantified by RT-qPCR. These graphics represent the mean of three separate experiments performed with triplicate samples with standard deviations. (D) His6-SehB-WT and His6-SehB-point mutations fusion proteins were analyzed by Western Blotting using a polyclonal anti-His6 antibody. Statistically significant with respect to WT S. Typhimurium; **p < 0.01; ***p < 0.001; ns, not significant.
Amino Acids in Both N- and C-Terminal Are Essential for SehB DNA-Binding Activity
SehB represses its own expression by direct binding to its promoter region (De la Cruz et al., 2013). We purified the eight point mutation proteins and wild-type SehB to evaluate their DNA-binding activities. EMSAs were performed using increasing concentrations of the His6-SehB proteins and a DNA fragment encompassing the promoter region of sehAB (−361 to +20 with respect to the putative transcriptional start site). His6-SehB-WT antitoxin was able to bind to its own promoter region at 125 nM and evident DNA-protein complex was detected at 125 and 250 nM (Figure 3) as previously described (De la Cruz et al., 2013). Based on the HTH DNA-binding domain found at the C-terminal, we predicted that mutations in this region may be affected regarding their ability to interact with DNA. Indeed, the DNA-protein complex was not observed with any of the SehB mutants indicating that mutations S107, L121, L129, and F140 affect the binding activity of SehB to the sehAB promoter region (Figure 3). Interestingly, the four mutations at the SehB N-terminal were also affected regarding DNA-binding activity (Figure 3), although SehB-Y32A protein still bound to the sehAB promoter at 250 nM, leading to the formation of a weak DNA-protein complex. These in vitro results show that changes in amino acids located in both N- and C-terminal alter the ability of SehB antitoxin to bind to its own promoter region, which could affect its repressor activity.
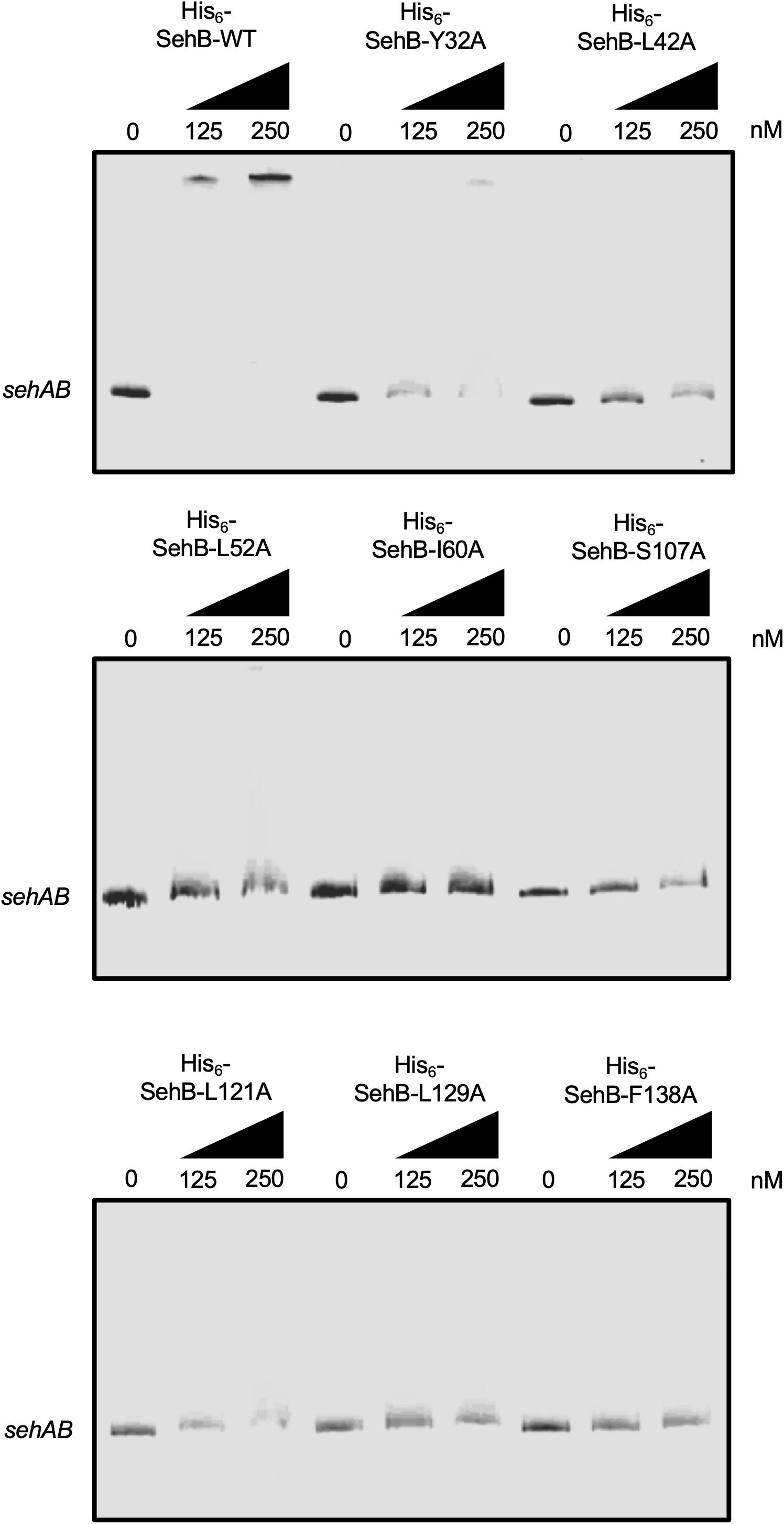
Figure 3. Binding of SehB-WT protein and SehB mutated proteins to the promoter region of sehAB. Electrophoretic mobility shift assays were carried out by incubating 100 ng PCR products covering sehAB (100 ng) promoter region and increasing amounts (0, 125 and 250 nM) of His6-SehB-WT protein and His6-SehB mutated as indicated. The complexes were separated on 6% polyacrylamide gels and DNA was stained with ethidium bromide.
Residues at the N-Terminal of the SehB Antitoxin Are Required for Homodimerization
We have previously reported that the SehB antitoxin forms a stable homodimer in solution or bound to DNA (De la Cruz et al., 2013). In order to corroborate this description, the LexADBDwt-based genetic system (Dmitrova et al., 1998; Daines and Silver, 2000) was used to analyze SehB homodimerization. LexADBDwt-SehB-WT repressed the expression of sulA-lacZ, confirming that the SehB antitoxin forms an active homodimer (Figure 4A). We used the LexADBDwt-alone and LexADBDwt-H-NS fusion proteins as negative and positive controls, respectively (Paredes-Amaya et al., 2018). While changes at the C-terminal were not severely affected in homodimer formation, mutations in the N-terminal domain were defective in their ability to repress the expression of sulA–lacZ compared to LexADBDwt-SehB-WT, suggesting that these amino acids of the N-terminal part are involved in SehB antitoxin dimerization (Figure 4A). All LexADBDwt-SehB fusion proteins were detected by Western Blotting assays, supporting the notion that differences found in homodimerization of LexADBDwt-SehB mutant proteins was not due to the lack of protein production (Figure 4B). Our data indicate that amino acid residues Y32, L42, L52, I60 of SehB are involved in homodimerization of the antitoxin and subsequently in its DNA binding activity.
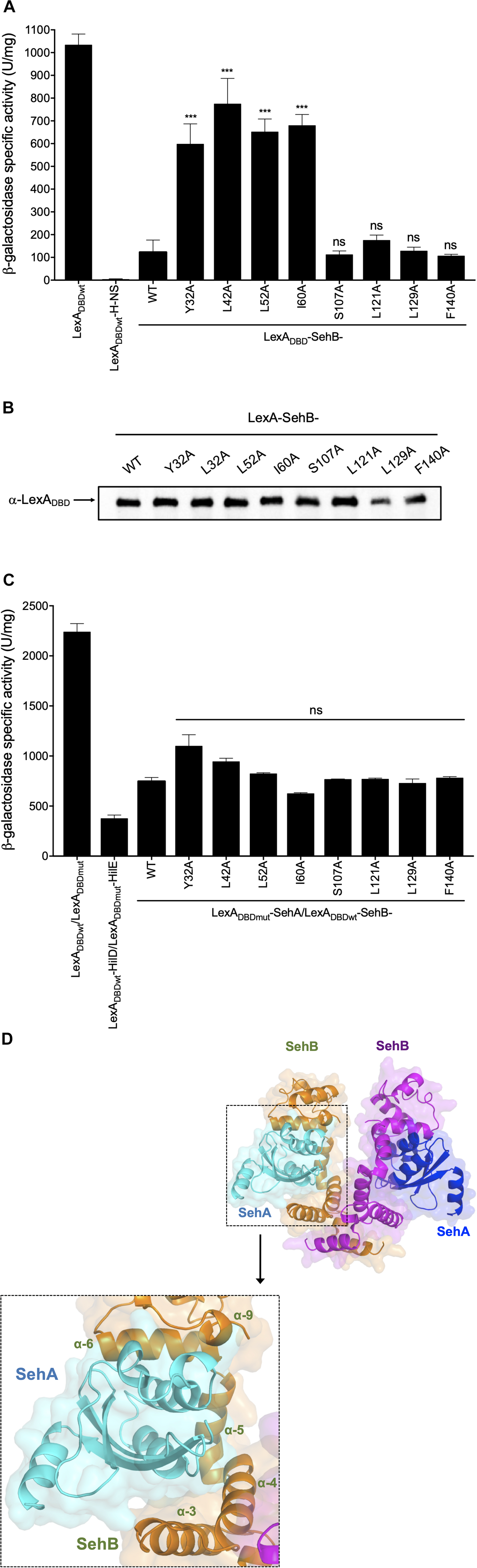
Figure 4. Relevance of SehB amino acids residues in the formation of SehB homodimers and SehB-SehA heterodimers. (A) Expression of the sulA–lacZ fusion was determined in the E. coli SU101 reporter strain containing the plasmids pSR658 (LexADBDwt), pSR658-H-NS (LexADBDwt-H-NS), pSR658-SehB-WT (LexADBDwt-SehB-WT), and pSR658-SehB-point mutations (LexADBDwt-SehB-point mutations). (B) LexADBDwt-SehB-WT and LexADBDwt-SehB-point mutations fusion proteins were analyzed by Western Blotting using a polyclonal anti-LexA antibody. (C) Expression of the sulA–lacZ fusion was determined in the E. coli SU202 reporter strain containing the plasmids pSR658 (LexADBDwt), pSR659 (LexADBDwt) pSR658-HilD (LexADBDwt-HilD), pSR659-HilE (LexADBDmt-HilE), pSR658-SehB (LexADBDwt-SehB), pSR659-SehA (LexADBDmt-SehA), and pSR658-SehB-point mutations (LexADBDwt-SehB-point mutations). The β-Galactosidase activity was determined from samples collected of bacterial cultures grown in LB broth at 37°C to an OD600 nm of 1.0. Expression of LexADBDwt and the LexADBDmt fusion proteins was induced by adding 1 mM IPTG to the medium. (D) Model of SehA-(SehB)2-SehA heterotetrameric complex is shown by superposition of HigBA complex structure using PyMol software. SehB antitoxins are colored in orange and magenta, and SehA toxins are in cyan and blue. Zoomed view of toxin-antitoxin interaction domain is showed. SehB α3-α4-α5-α6 are depicted in the 3D-model. Data are the averages of three independent experiments performed by triplicate. The bars represent the standard deviations. ***p < 0.001; ns, not significant.
SehB Amino Acid Changes Are Dispensable for the Interaction With the SehA Toxin
In addition to repress its own expression, SehB neutralizes the SehA toxin activity by protein-protein interaction (De la Cruz et al., 2013). To investigate if SehB point mutations were affected in the interaction with SehA, we used the LexA-based genetic system for heterodimerization (Dmitrova et al., 1998; Daines and Silver, 2000). LexADBDwt/LexADBDmut and LexADBDwt-HilD/LexADBDmut-HilE fusion protein pairs were used as negative and positive controls, respectively (Paredes-Amaya et al., 2018). LexADBDwt-SehB-WT and LexADBDmut-SehA fusion proteins repressed the expression of sulA-lacZ, corroborating the direct interaction between SehA and SehB, which is a hallmark of type II TA systems (Figure 4C). None of the SehB point mutants were affected in the interaction with SehA toxin, as they still repressed the transcription of the sulA-lacZ fusion (Figure 4C). In fact, the 3D-structure prediction of SehA-(SehB)2-SehA heterotetrameric complex revealed that α3, α4, α5, and α6 of each SehB monomer would be interacting with one SehA toxin, suggesting that the SehB-SehA interaction is more complex and it involves more than just one amino acid (Figure 4D). These observations further indicate that single mutations in Y32, L42, L52, I60, S107, L121, L129, and F140 amino acids were not enough to alter the ability of SehB antitoxin to bind SehA toxin.
SehB Amino Acid Substitutions Affect Salmonella Intracellular Replication
As the absence of the SehB antitoxin affects intracellular replication of S. Typhimurium within macrophages (De la Cruz et al., 2013), the role of SehB point mutations in intracellular Salmonella replication was tested in RAW264.7 mouse macrophages. Macrophages were infected with either WT S. Typhimurium or mutant bacteria expressing the SehB mutant variants, and the fold increase of intracellular Salmonella was calculated between 2 and 16 h, compared to wild-type strain. All SehB mutants showed a reduced level of intra-macrophage replication compared wild-type and complemented ΔsehB strains (Figure 5). We concluded that Y32, L42, L52, I60, S107, L121, L129, and F140 amino acid residues of SehB antitoxin are required for the S. Typhimurium intracellular growth.
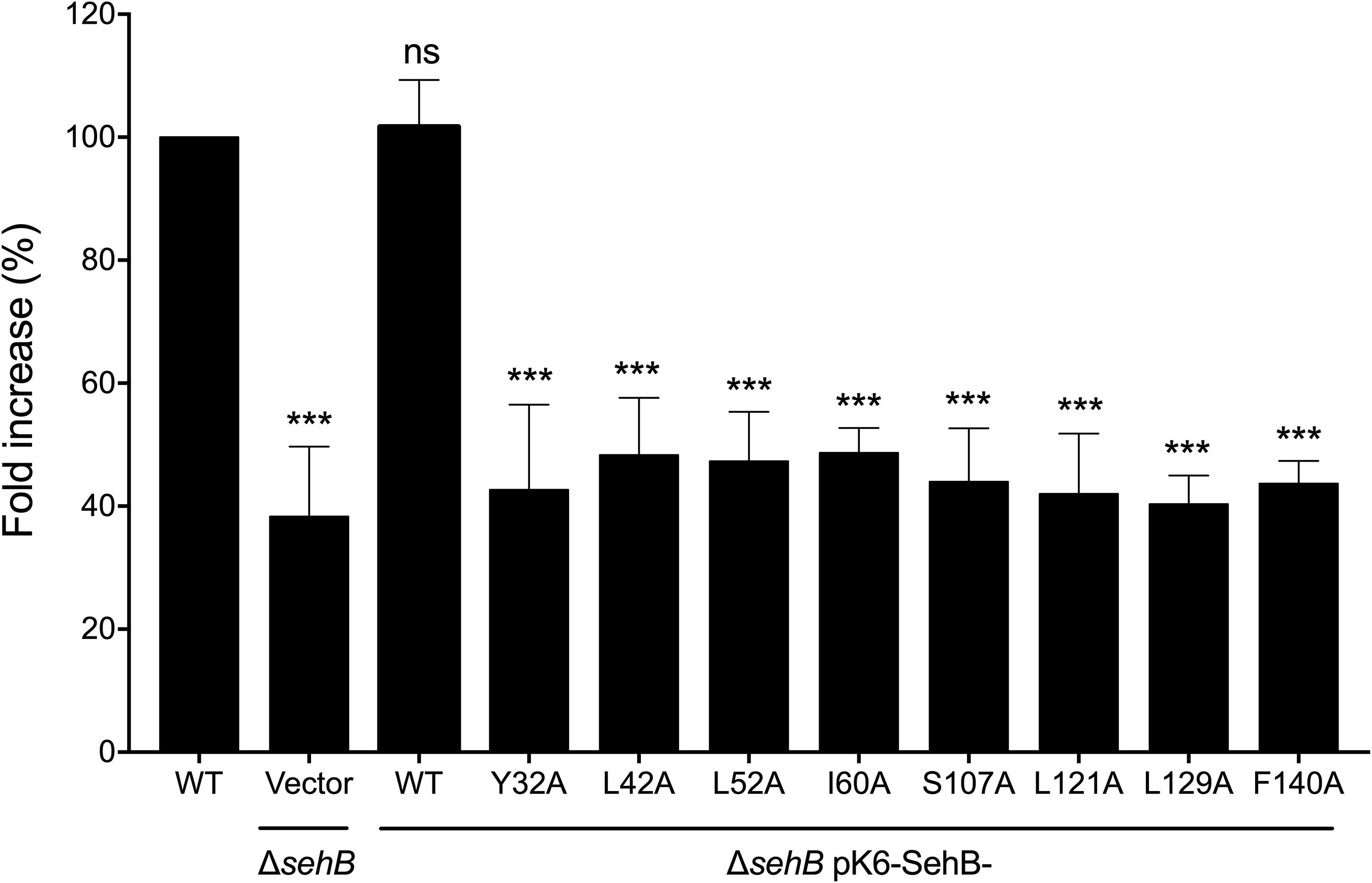
Figure 5. SehB amino acids requirement for the intra-macrophagic replication. RAW264.7 macrophages were infected with S. Typhimurium wild-type (WT), ΔsehB pMPM-K6 and ΔsehB expressing the SehB-WT and SehB point mutations and lysed at 2 h and 16 h p.i. for quantification of intracellular bacteria. The values shown represent the fold increase calculated as a ratio of the intracellular bacteria between 16 and 2 h and normalized to that of the wild-type strain. Values are means ± SD (n = 6). The data are averages of three independent experiments performed by triplicate. The bars represent the standard deviations. ***p < 0.001; ns, not significant.
Amino Acids of the SehB Antitoxin Are Involved in Salmonella Virulence in Mice
We previously showed that the absence of SehB antitoxin dramatically affected the S. Typhimurium virulence in the mouse model (De la Cruz et al., 2013). To evaluate the contribution of the eight amino acid residues of SehB on virulence, the ΔsehB mutant expressing wild-type or mutagenic variants of SehB were tested in survival experiments in which groups of Balb/c mice were infected by gavage with 105 bacteria. These strains were compared to the wild-type (virulent) and to the ΔsehB pMPM-K6 strains (non-virulent) strains (Figure 6A). ΔsehB strains expressing SehB point mutations appeared partially attenuated in the mouse model. While WT S. Typhimurium killed all mice at day 7 p.i., the SehB point mutations located at N-terminal only killed 50% of mice at 14 days (Figure 6B). Changes of amino acids at C-terminal of SehB also attenuated the S. Typhimurium virulence (Figure 6C). In addition, the SehB mutagenic variants were tested for virulence by competitive index (CI) experiments. In this assay, each SehB point mutant was co-administered with WT strain to Balb/c mice and the number of each bacterium from the spleen was determined 2 days after inoculation by gavage. In contrast to the ΔsehB strain complemented with wild-type SehB, both N-, and C-terminal SehB mutations were poorly recovered from the spleen (Figure 7), showing a CI between 0.1 and 0.01. In conclusion, analysis of the different SehB mutants showed that all SehB amino acids tested are required for Salmonella virulence in mice.

Figure 6. Attenuation of S. Typhimurium ΔsehB expressing SehB point mutations. Six Balb/c mice were intragastrically infected with 105 CFUs of S. Typhimurium wild-type (WT), ΔsehB pMPM-K6 and complemented ΔsehB mutant (expressing SehB-WT) (A) and SehB point mutations at N-and C-terminal (B,C) and they were observed during 14 days. The percentages of surviving mice are shown for each strain.
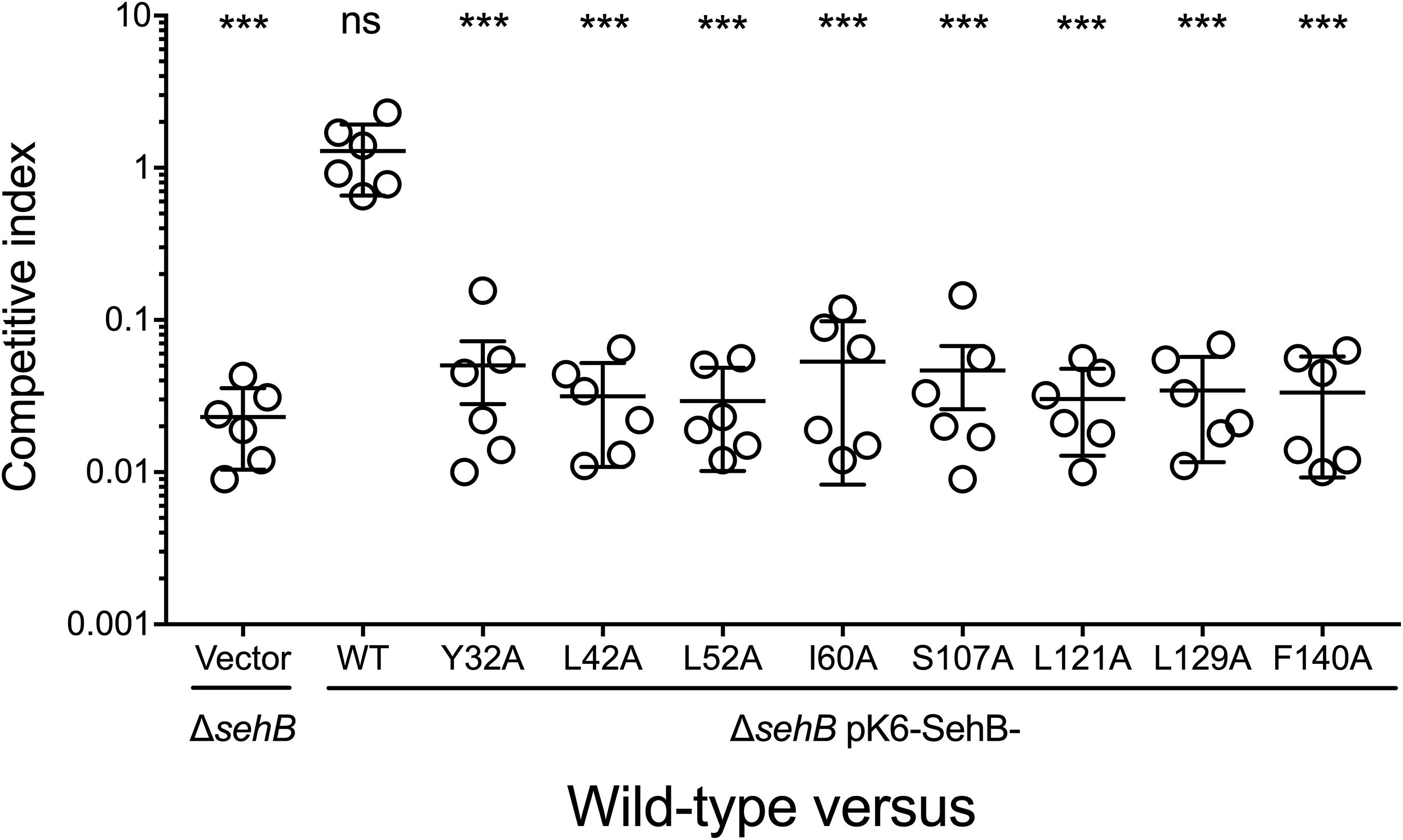
Figure 7. Role of SehB point mutations on the colonization in mice. Six Balb/c mice were inoculated intraperitoneally with a 1:1 mixture of two Salmonella strains as indicated. Spleens were harvested 2 days post-inoculation and bacteria were enumerated. Each symbol represents a mouse and horizontal bars correspond to the means ± SD. An unpaired t-test was used to determine whether two values were significantly different. ***p < 0.001; ns, not significant.
Discussion
During evolution, Salmonella enterica acquired and integrated various genetic elements into its genome, which conferred fast adaptive advantages in specific ecological niches. In this context, the type II TA systems form part of the bacterial mobilome and they can be found in both plasmids and chromosomes. S. Typhimurium chromosome encodes one of the type II TA systems named SehAB, which is homologous to HigBA family, and is required for the virulence of this bacterium (De la Cruz et al., 2013). HigBA-type TA systems are highly prevalent in bacteria, followed only by the VapBC and RelBE-like TA systems (Pandey and Gerdes, 2005). Of these, HigA-type antitoxins differ mainly from two other type II antitoxins: (i) genetically, the antitoxin gene is located after the toxin gene, and (ii) the DNA-binding domains are mostly located at the C-terminal of the protein (Chan et al., 2016). Interestingly, in contrast to most of the type II antitoxins, the absence of the HigA-type antitoxins seems to be lethal in different bacteria or at least to affect their growth (Budde et al., 2007; Fivian-Hughes and Davis, 2010; De la Cruz et al., 2013; Wood and Wood, 2016). In terms of the regulatory mechanisms, antitoxin components seem to be more complex than those of toxin because the first one controls expression of the second at two levels, transcriptional and post-translational, by directly repressing its own expression and neutralizing via protein-protein interaction, respectively (Lobato-Marquez et al., 2016). This functional versatility highlights the importance of characterization of this protein, mainly as a target for molecules that affects its antagonistic activity against the toxin, which results in inhibition of bacterial growth.
Our in silico analysis showed that the SehB antitoxin contained two domains, N- and C-terminals, having four α-helices each and these are connected by one α-helix, which functions as a linker. In solution, SehB was detected forming a dimer, bound or not to DNA containing two palindromic sequences found on the −35 and −10 putative boxes (De la Cruz et al., 2013). A dimer of SehB is required for the transcriptional auto-repression, such as was reported for HigA antitoxin in E. coli (HigAEco), S. flexneri, (HigASfl), V. cholerae (HigAVch), P. aeruginosa (HigAPae), and P. vulgaris (HigAPvu) (Yang et al., 2016; Hadzi et al., 2017; Schureck et al., 2019; Xu et al., 2019; Yoon et al., 2019). Interestingly, the four HigA reported structures show some differences, mainly in structure, organization and location of both dimerization and DNA-binding domains. HigAPae and HigAPvu present a DNA-binding domain similar to HigAEco, although the DNA-binding and dimerization domains are located at the N- and C-terminal, respectively (Schureck et al., 2014). HigAVch protein is more related to E. coli RelB antitoxin, possessing both dimerization and DNA-binding domains at the C-terminal (Hadzi et al., 2017). HigAEco and HigASfl antitoxins were reported to have dimerization and DNA-binding domains located at the N- and C-terminals, respectively (Yang et al., 2016; Xu et al., 2019; Yoon et al., 2019). Similar to both HigAEco and HigASfl, which show 46% identity and 67% similarity with SehB, intact N-and C-terminal domains of SehB were required for self-interaction and for DNA-binding, respectively. Although the crystallographic structure of the SehB antitoxin has not been yet resolved, a 3D-model generated using the HigA structures from E. coli and S. flexneri allowed to observe an homodimer formation (Yang et al., 2016; Xu et al., 2019; Yoon et al., 2019). We propose that L42 and L52 amino acids are participating in the intermolecular contact between both SehB monomers. In contrast, the proximity of Y32 and I60 residues, located at helices α-3 and α-4, respectively, could confer a conformational stability to each SehB monomer, which is required for the homodimer formation. The C-terminal, S107, L121, and L129 amino acid residues form part of the HTH motif necessary for the interaction between SehB antitoxin and its own promoter region. The F140 residue that is conserved in homologous proteins, seems to be interacting with helix α-6 and it probably results in a structure organization required for the conformation of the C-terminal domain, which contains the DNA-binding domain of the antitoxin. In this way, to avoid dramatic changes on the SehB structure, proline or charged amino acids were not selected to be mutagenized in this initial research.
Interestingly, none of the SehB point mutations tested affected heterodimerization between SehB and SehA, keeping the antitoxin neutralizing activity on the toxin. Our 3D-model showed that both homodimerization and toxin interaction domains of SehB are located at the N-terminal, which is similar to what has been reported for HigAEco and HigASfl antitoxins (Yang et al., 2016; Xu et al., 2019; Yoon et al., 2019). Two-hybrid assays and 3D-structure prediction suggest that SehB-SehA interaction is more complex that SehB-SehB, because single mutations on both N- and C-terminal of SehB antitoxin mainly affected the SehB interaction with itself and not the heterodimerization with SehA toxin. Those mutagenic variants still kept the SehA neutralizing activity at the post-translational level despite the lack of repressor activity, explaining why all SehB point mutations were able to complement the growth defect of the sehB chromosomal mutant in LB broth. In E. coli and S. flexneri, HigBA toxin-antitoxin complex has higher affinity to DNA than HigA antitoxin alone (Yang et al., 2016; Yoon et al., 2019). However, HigA alone presents higher affinity to DNA than HigBA complex in P. vulgaris (Schureck et al., 2019). We were unable to analyze if SehAB presented higher or lesser DNA-binding affinity than SehB alone because the SehA-(SehB)2-SehA complex was not possible to reconstitute using both purified proteins, as previously described (De la Cruz et al., 2013).
In the absence of SehB antitoxin, high levels of SehA toxin are expressed, partially affecting the bacterial growth in LB broth resulting in severe attenuation of virulence in the mouse model (De la Cruz et al., 2013). We proposed that in addition to SehA toxin, other elements present in the S. Typhimurium genome, could be affected in the absence of SehB antitoxin, mainly to transcriptional level. This hypothesis is supported by reports where the absence of HigA antitoxin in P. aeruginosa transcriptionally affects other genes that encode for virulence factors, such as pyochelin, pyocyanin, type III secretion system, MvfR transcriptional regulator, and those related with the c-di-GMP metabolism (Li et al., 2016; Wood and Wood, 2016; Zhang et al., 2018; Guo et al., 2019).
The contribution of SehB to S. Typhimurium virulence allowed the identification of amino acid residues involved in the intracellular replication and colonization in mice. According to what is observed in the ability of homodimerization and DNA-binding, all amino acids tested in this study were relevant for the functioning of SehB and needed to confer virulence to S. Typhimurium. Survival curves and competitive indices experiments in mice showed that Y32, L42, L52, I60, S107, L121, L129, and F140 amino acid residues are required for S. Typhimurium virulence. Although ΔsehB mutant expressing pK6-SehB-WT fully complemented the virulence of Salmonella in macrophages and competitive indices assays, this complementation was partial in the mice survival curves. Since pK6-SehB-WT plasmid was stable during the 14 days of mice monitoring (data not shown), we hypothesized that when the bacterium senses some signals at late times during the systemic infection (i.e., after 6 days), the expression of arabinose promoter from plasmid (which drives the sehB transcription) could be down-regulated, or may mechanisms related to copy number or segregation be affected.
The importance of each amino acid residue could be explained by the small size of the SehB antitoxin and the complexity of its regulatory function. Although SehB antitoxin neutralizes the SehA toxin activity by protein-protein interaction, the primary control on the sehAB expression is based in the transcription activity of SehB on its own promoter region. Future work will be aimed to design molecules that function as drugs in order to affect either homodimerization capacity or SehB DNA-binding.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
The animal study was reviewed and approved by the Internal Ethics Committee of the Animal Resource Facility of the Universidad Autónoma del Estado de Hidalgo (Approval number: CIECUAL/002/2018 to Manuel Sánchez-Gutiérrez).
Author Contributions
MD conceived and designed the experiments. FC-C, GH-M, SP, MA, and JS-B performed the experiments. MD, MA, MS-G, JI-V, JI, JG, J-PG, SM, and MD analyzed the data. MD wrote the manuscript.
Funding
This work was supported by grants FIS/IMSS/PROT/G15/1398 (to MD) from the Fondo de Investigación en Salud (FIS)-Instituto Mexicano del Seguro Social (IMSS), México and from the Consejo Nacional de Ciencia y Tecnología (CONACYT), México (256263) to MD. FC-C was supported by a pre-doctoral fellowship from CONACYT (331108) and IMSS (99097541). GH-M was supported by a master fellowship from CONACYT (932252) and IMSS (99097630).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Victor H. Bustamante for the support in β-Galactosidase experiments. We thank to Programa de Cooperación Internacional-IMSS (2017) for supporting a research stay in Marseille France.
References
Ares, M. A., Sansabas, A., Rodriguez-Valverde, D., Siqueiros-Cendon, T., Rascon-Cruz, Q., Rosales-Reyes, R., et al. (2019). The interaction of Klebsiella pneumoniae with lipid rafts-associated cholesterol increases macrophage-mediated phagocytosis due to down regulation of the capsule polysaccharide. Front. Cell. Infect. Microbiol. 9:255. doi: 10.3389/fcimb.2019.00255
Aussel, L., Zhao, W., Hebrard, M., Guilhon, A. A., Viala, J. P., Henri, S., et al. (2011). Salmonella detoxifying enzymes are sufficient to cope with the host oxidative burst. Mol. Microbiol. 80, 628–640. doi: 10.1111/j.1365-2958.2011.07611.x
Beuzon, C. R., and Holden, D. W. (2001). Use of mixed infections with Salmonella strains to study virulence genes and their interactions in vivo. Microbes Infect. 3, 1345–1352. doi: 10.1016/s1286-4579(01)01496-4
Budde, P. P., Davis, B. M., Yuan, J., and Waldor, M. K. (2007). Characterization of a higBA toxin-antitoxin locus in Vibrio cholerae. J. Bacteriol. 189, 491–500. doi: 10.1128/jb.00909-06
Casadaban, M. J. (1976). Transposition and fusion of the lac genes to selected promoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol. Biol. 104, 541–555. doi: 10.1016/0022-2836(76)90119-4
Chan, W. T., Espinosa, M., and Yeo, C. C. (2016). Keeping the wolves at bay: antitoxins of prokaryotic type II toxin-antitoxin systems. Front. Mol. Biosci. 3:9. doi: 10.3389/fmolb.2016.00009
Cheverton, A. M., Gollan, B., Przydacz, M., Wong, C. T., Mylona, A., Hare, S. A., et al. (2016). A Salmonella toxin promotes persister formation through acetylation of tRNA. Mol. Cell 63, 86–96. doi: 10.1016/j.molcel.2016.05.002
Daines, D. A., and Silver, R. P. (2000). Evidence for multimerization of neu proteins involved in polysialic acid synthesis in Escherichia coli K1 using improved LexA-based vectors. J. Bacteriol. 182, 5267–5270. doi: 10.1128/jb.182.18.5267-5270.2000
De la Cruz, M. A., Fernandez-Mora, M., Guadarrama, C., Flores-Valdez, M. A., Bustamante, V. H., Vazquez, A., et al. (2007). LeuO antagonizes H-NS and StpA-dependent repression in Salmonella enterica ompS1. Mol. Microbiol. 66, 727–743. doi: 10.1111/j.1365-2958.2007.05958.x
De la Cruz, M. A., Zhao, W., Farenc, C., Gimenez, G., Raoult, D., Cambillau, C., et al. (2013). A toxin-antitoxin module of Salmonella promotes virulence in mice. PLoS Pathog. 9:e1003827. doi: 10.1371/journal.ppat.1003827
Dmitrova, M., Younes-Cauet, G., Oertel-Buchheit, P., Porte, D., Schnarr, M., and Granger-Schnarr, M. (1998). A new LexA-based genetic system for monitoring and analyzing protein heterodimerization in Escherichia coli. Mol. Gen. Genet. 257, 205–212. doi: 10.1007/s004380050640
Fabrega, A., and Vila, J. (2013). Salmonella enterica serovar typhimurium skills to succeed in the host: virulence and regulation. Clin. Microbiol. Rev. 26, 308–341. doi: 10.1128/CMR.00066-12
Fivian-Hughes, A. S., and Davis, E. O. (2010). Analyzing the regulatory role of the HigA antitoxin within Mycobacterium tuberculosis. J. Bacteriol. 192, 4348–4356. doi: 10.1128/JB.00454-10
Guo, Y., Sun, C., Li, Y., Tang, K., Ni, S., and Wang, X. (2019). Antitoxin HigA inhibits virulence gene mvfR expression in Pseudomonas aeruginosa. Environ. Microbiol. 21, 2707–2723. doi: 10.1111/1462-2920.14595
Hadzi, S., Garcia-Pino, A., Haesaerts, S., Jurenas, D., Gerdes, K., Lah, J., et al. (2017). Ribosome-dependent Vibrio cholerae mRNAse HigB2 is regulated by a beta-strand sliding mechanism. Nucleic Acids Res. 45, 4972–4983. doi: 10.1093/nar/gkx138
Haraga, A., Ohlson, M. B., and Miller, S. I. (2008). Salmonellae interplay with host cells. Nat. Rev. Microbiol. 6, 53–66. doi: 10.1038/nrmicro1788
Harms, A., Brodersen, D. E., Mitarai, N., and Gerdes, K. (2018). Toxins, targets, and triggers: an overview of toxin-antitoxin biology. Mol. Cell 70, 768–784. doi: 10.1016/j.molcel.2018.01.003
Harms, A., Stanger, F. V., Scheu, P. D., De Jong, I. G., Goepfert, A., Glatter, T., et al. (2015). Adenylylation of gyrase and topo IV by FicT toxins disrupts bacterial DNA topology. Cell Rep. 12, 1497–1507. doi: 10.1016/j.celrep.2015.07.056
Ho, S. N., Hunt, H. D., Horton, R. M., Pullen, J. K., and Pease, L. R. (1989). Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene 77, 51–59. doi: 10.1016/0378-1119(89)90358-2
Hurley, J. M., and Woychik, N. A. (2009). Bacterial toxin HigB associates with ribosomes and mediates translation-dependent mRNA cleavage at A-rich sites. J. Biol. Chem. 284, 18605–18613. doi: 10.1074/jbc.M109.008763
Jahn, C. E., Charkowski, A. O., and Willis, D. K. (2008). Evaluation of isolation methods and RNA integrity for bacterial RNA quantitation. J. Microbiol. Methods 75, 318–324. doi: 10.1016/j.mimet.2008.07.004
Jajere, S. M. (2019). A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 12, 504–521. doi: 10.14202/vetworld.2019.504-521
Li, M., Long, Y., Liu, Y., Liu, Y., Chen, R., Shi, J., et al. (2016). HigB of Pseudomonas aeruginosa enhances killing of phagocytes by up-regulating the type III secretion system in ciprofloxacin induced persister cells. Front. Cell. Infect. Microbiol. 6:125. doi: 10.3389/fcimb.2016.00125
Liu, Y., Gao, Z., Liu, G., Geng, Z., Dong, Y., and Zhang, H. (2019). Structural insights into the transcriptional regulation of HigBA toxin-antitoxin system by antitoxin HigA in Pseudomonas aeruginosa. Front. Microbiol. 10:3158. doi: 10.3389/fmicb.2019.03158
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lobato-Marquez, D., Diaz-Orejas, R., and Garcia-Del Portillo, F. (2016). Toxin-antitoxins and bacterial virulence. FEMS Microbiol. Rev. 40, 592–609. doi: 10.1093/femsre/fuw022
Martinez, L. C., Yakhnin, H., Camacho, M. I., Georgellis, D., Babitzke, P., Puente, J. L., et al. (2011). Integration of a complex regulatory cascade involving the SirA/BarA and Csr global regulatory systems that controls expression of the Salmonella SPI-1 and SPI-2 virulence regulons through HilD. Mol. Microbiol. 80, 1637–1656. doi: 10.1111/j.1365-2958.2011.07674.x
Matter, L. B., Ares, M. A., Abundes-Gallegos, J., Cedillo, M. L., Yanez, J. A., Martinez-Laguna, Y., et al. (2018). The CpxRA stress response system regulates virulence features of avian pathogenic Escherichia coli. Environ. Microbiol. 20, 3363–3377. doi: 10.1111/1462-2920.14368
Mayer, M. P. (1995). A new set of useful cloning and expression vectors derived from pBlueScript. Gene 163, 41–46. doi: 10.1016/0378-1119(95)00389-n
Mutschler, H., Gebhardt, M., Shoeman, R. L., and Meinhart, A. (2011). A novel mechanism of programmed cell death in bacteria by toxin-antitoxin systems corrupts peptidoglycan synthesis. PLoS Biol. 9:e1001033. doi: 10.1371/journal.pbio.1001033
Oropeza, R., Sampieri, C. L., Puente, J. L., and Calva, E. (1999). Negative and positive regulation of the non-osmoregulated ompS1 porin gene in Salmonella typhi: a novel regulatory mechanism that involves OmpR. Mol. Microbiol. 32, 243–252. doi: 10.1046/j.1365-2958.1999.01329.x
Page, R., and Peti, W. (2016). Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 12, 208–214. doi: 10.1038/nchembio.2044
Pandey, D. P., and Gerdes, K. (2005). Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 33, 966–976. doi: 10.1093/nar/gki201
Paredes-Amaya, C. C., Valdes-Garcia, G., Juarez-Gonzalez, V. R., Rudino-Pinera, E., and Bustamante, V. H. (2018). The Hcp-like protein HilE inhibits homodimerization and DNA binding of the virulence-associated transcriptional regulator HilD in Salmonella. J. Biol. Chem. 293, 6578–6592. doi: 10.1074/jbc.RA117.001421
Rocker, A., and Meinhart, A. (2016). Type II toxin: antitoxin systems. More than small selfish entities? Curr. Genet. 62, 287–290. doi: 10.1007/s00294-015-0541-7
Ronneau, S., and Helaine, S. (2019). Clarifying the link between toxin-antitoxin modules and bacterial persistence. J. Mol. Biol. 431, 3462–3471. doi: 10.1016/j.jmb.2019.03.019
Schureck, M. A., Maehigashi, T., Miles, S. J., Marquez, J., Cho, S. E., Erdman, R., et al. (2014). Structure of the Proteus vulgaris HigB-(HigA)2-HigB toxin- antitoxin complex. J. Biol. Chem. 289, 1060–1070. doi: 10.1074/jbc.M113.512095
Schureck, M. A., Meisner, J., Hoffer, E. D., Wang, D., Onuoha, N., Ei Cho, S., et al. (2019). Structural basis of transcriptional regulation by the HigA antitoxin. Mol. Microbiol. 111, 1449–1462. doi: 10.1111/mmi.14229
Waterhouse, A., Bertoni, M., Bienert, S., Studer, G., Tauriello, G., Gumienny, R., et al. (2018). SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303. doi: 10.1093/nar/gky427
Wood, T. L., and Wood, T. K. (2016). The HigB/HigA toxin/antitoxin system of Pseudomonas aeruginosa influences the virulence factors pyochelin, pyocyanin, and biofilm formation. Microbiologyopen 5, 499–511. doi: 10.1002/mbo3.346
Xu, B. S., Liu, M., Zhou, K., Geng, Z., Gao, Z. Q., Dong, Y. H., et al. (2019). Conformational changes of antitoxin HigA from Escherichia coli str. K-12 upon binding of its cognate toxin HigB reveal a new regulation mechanism in toxin-antitoxin systems. Biochem. Biophys. Res. Commun. 514, 37–43. doi: 10.1016/j.bbrc.2019.04.061
Yamaguchi, Y., and Inouye, M. (2009). mRNA interferases, sequence-specific endoribonucleases from the toxin-antitoxin systems. Prog. Mol. Biol. Transl. Sci. 85, 467–500. doi: 10.1016/S0079-6603(08)00812-X
Yamaguchi, Y., and Inouye, M. (2011). Regulation of growth and death in Escherichia coli by toxin-antitoxin systems. Nat. Rev. Microbiol. 9, 779–790. doi: 10.1038/nrmicro2651
Yamaguchi, Y., Park, J. H., and Inouye, M. (2011). Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 45, 61–79. doi: 10.1146/annurev-genet-110410-132412
Yang, J., Zhou, K., Liu, P., Dong, Y., Gao, Z., Zhang, J., et al. (2016). Structural insight into the E. coli HigBA complex. Biochem. Biophys. Res. Commun. 478, 1521–1527. doi: 10.1016/j.bbrc.2016.08.131
Yoon, W. S., Seok, S. H., Won, H. S., Cho, T., Lee, S. J., and Seo, M. D. (2019). Structural changes of antitoxin HigA from Shigella flexneri by binding of its cognate toxin HigB. Int. J. Biol. Macromol. 130, 99–108. doi: 10.1016/j.ijbiomac.2019.02.111
Keywords: SehB, SehAB, toxin-antitoxin, Salmonella, virulence
Citation: Chimal-Cázares F, Hernández-Martínez G, Pacheco S, Ares MA, Soria-Bustos J, Sánchez-Gutiérrez M, Izquierdo-Vega JA, Ibarra JA, González-y-Merchand JA, Gorvel J-P, Méresse S and De la Cruz MA (2020) Molecular Characterization of SehB, a Type II Antitoxin of Salmonella enterica Serotype Typhimurium: Amino Acid Residues Involved in DNA-Binding, Homodimerization, Toxin Interaction, and Virulence. Front. Microbiol. 11:614. doi: 10.3389/fmicb.2020.00614
Received: 06 December 2019; Accepted: 19 March 2020;
Published: 09 April 2020.
Edited by:
Daniel Pletzer, University of Otago, New ZealandReviewed by:
Eun-Jin Lee, Korea University, South KoreaYunxue Guo, South China Sea Institute of Oceanology (CAS), China
Copyright © 2020 Chimal-Cázares, Hernández-Martínez, Pacheco, Ares, Soria-Bustos, Sánchez-Gutiérrez, Izquierdo-Vega, Ibarra, González-y-Merchand, Gorvel, Méresse and De la Cruz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel A. De la Cruz, bWlndWVsX2FuZ2VsXzgxQGxpdmUuY29t
†These authors have contributed equally to this work