 Agustina Rodríguez-Sorrento1
Agustina Rodríguez-Sorrento1 Lorena Castillejos1*
Lorena Castillejos1* Paola López-Colom1
Paola López-Colom1 Gloria Cifuentes-Orjuela2
Gloria Cifuentes-Orjuela2 María Rodríguez-Palmero2
María Rodríguez-Palmero2 José Antonio Moreno-Muñoz2
José Antonio Moreno-Muñoz2 Diana Luise3
Diana Luise3 Paolo Trevisi3
Paolo Trevisi3 Susana María Martín-Orúe1
Susana María Martín-Orúe1- 1Servicio de Nutrición y Bienestar Animal, Departament de Ciència Animal i dels Aliments, Universitat Autònoma de Barcelona, Bellaterra, Spain
- 2Laboratorios Ordesa S.L., Parc Científic de Barcelona, Barcelona, Spain
- 3Department of Agricultural and Food Science, University of Bologna, Bologna, Italy
We evaluated the potential of multi-strain probiotic (Bifidobacterium longum subsp. infantis CECT 7210 and Lactobacillus rhamnosus HN001) with or without galacto-oligosaccharides against enterotoxigenic Escherichia coli (ETEC) F4 infection in post-weaning pigs. Ninety-six piglets were distributed into 32 pens assigned to five treatments: one non-challenged (CTR+) and four challenged: control diet (CTR−), with probiotics (>3 × 1010 CFU/kg body weight each, PRO), prebiotic (5%, PRE), or their combination (SYN). After 1 week, animals were orally inoculated with ETEC F4. Feed intake, weight, and clinical signs were recorded. On days 4 and 8 post-inoculation (PI), one animal per pen was euthanized and samples from blood, digesta, and tissues collected. Microbiological counts, ETEC F4 real-time PCR (qPCR) quantification, fermentation products, serum biomarkers, ileal histomorphometry, and genotype for mucin 4 (MUC4) polymorphism were determined. Animals in the PRO group had similar enterobacteria and coliform numbers to the CTR+ group, and the ETEC F4 prevalence, the number of mitotic cells at day 4 PI, and villus height at day 8 PI were between that observed in the CTR+ and CTR− groups. The PRO group exhibited reduced pig major acute-phase protein (Pig-MAP) levels on day 4 PI. The PRE diet group presented similar reductions in ETEC F4 and Pig-MAP, but there was no effect on microbial groups. The SYN group showed reduced fecal enterobacteria and coliform counts after the adaptation week but, after the inoculation, the SYN group showed lower performance and more animals with high ETEC F4 counts at day 8 PI. SYN treatment modified the colonic fermentation differently depending on the MUC4 polymorphism. These results confirm the potential of the probiotic strains and the prebiotic to fight ETEC F4, but do not show any synergy when administered together, at least in this animal model.
Introduction
Diarrhea is the second leading infectious cause of death – after pneumonia – among children younger than 5 years (Liu et al., 2015). The main pathogenic agents responsible for these diseases are viruses – namely rotavirus and calicivirus, which are responsible for 38 and 13% of the cases, respectively – followed by enteropathogenic and enterotoxigenic Escherichia coli (EPEC and ETEC, respectively), which contribute to 12 and 8%, respectively, of the deaths (Lanata et al., 2013). ETEC is also one of most common pathogens in pigs; it is responsible for post-weaning diarrhea syndrome (Luppi, 2017). ETEC carrying fimbriae F4 and F18 is considered as the most common pathogen and, thus, these pathogens are used to perform standardized challenges with in vivo trials (Luise et al., 2019a). Although E. coli is part of the normal intestinal microbiota, some strains, like the above mentioned, have developed pathogenic mechanisms to cause intestinal or extraintestinal disease (Clements et al., 2012). Due to the increasing development of antibiotic resistance in this organism (Poirel et al., 2018), new strategies for prevention and therapy must be implemented. Probiotics are defined by the Food and Agriculture Organization (FAO) as “live microorganisms which, when administered in adequate amounts, confer a health benefit on the host.” These advantageous effects are strain dependent and can include gastrointestinal disorder prophylaxis and treatment, immune system enhancement, cancer prevention, and cholesterol normalization, among others (Kechagia et al., 2013). Regarding the potential of probiotics to ameliorate diarrhea caused by ETEC, different strains of bifidobacteria and lactobacilli have proven antagonistic activity against E. coli in vitro (Vazquez-Gutierrez et al., 2016; Fijan et al., 2018; Song et al., 2019). There have also been positive results when tested in vivo (Romond et al., 1997; Kumar et al., 2016). Our previous work has shown that the particular strain Bifidobacterium longum subsp. infantis CECT 7210 can reduce ileal colonization by ETEC, a phenomenon that improves the local immune response in a piglet model (Barba-Vidal et al., 2017). Regarding lactobacilli strains, Lactobacillus rhamnosus has been reported to have positive effects against pathogenic E. coli, although the benefits could depend on the probiotic strain. In this sense, L. rhamnosus HN001 has been demonstrated to reduce enterohemorrhagic E. coli translocation and to increase immunoglobulin A (IgA) concentration and blood leukocyte phagocytic activity in mice (Shu and Gill, 2001). However, administration of L. rhamnosus ATCC 53103 to post-weaning piglet infected with ETEC F4 does not confer protection (Trevisi et al., 2009). A combination of B. longum and L. rhamnosus HN001 has also been reported as an effective way to modulate the intestinal environment in humans, with the reductions of potential harmful bacteria and an increase of beneficial ones (Toscano et al., 2017).
On the other hand, prebiotics can also be useful against pathogenic bacteria (de Vrese and Marteau, 2007). Galacto-oligosaccharides (GOSs) are obtained from lactose by transgalactosylation reactions catalyzed by β-galactosidases. These reactions result in a chain of galactose units with a terminal glucose unit (Tzortzis, 2009). This prebiotic has been associated with increases in lactobacilli and bifidobacteria in vitro as well as in human clinical trials (Grimaldi et al., 2016; Canfora et al., 2017; Paganini et al., 2017). Moreover, GOS are considered to be highly similar to oligosaccharides from human milk (HMO), a factor that makes it an attractive prebiotic when designing synbiotics with B. longum subsp. infantis or other probiotic strains isolated from the infant intestine. Notably, genomic adaptations for HMO utilization have been described for B. longum subsp. infantis (Sela et al., 2008). Furthermore, GOS have been shown to interfere with E. coli adhesion to tissue culture cells (Shoaf et al., 2006). Moreover, recent studies have demonstrated the ability of the probiotic strain B. longum subsp. infantis CECT 7210 to metabolize GOS as substrate, promoting a higher growth than the other prebiotics. This combination has also shown significant antimicrobial properties against E. coli (Ruiz et al., 2020).
Based on this information, we hypothesized that the efficacy of combined probiotic strains of Bifidobacterium and Lactobacillus will be increased by the addition of fermentable carbohydrates that may promote their growth and activity in the gut. Hence, the objective of this work was to evaluate whether combining multi-strain probiotic, composed of B. longum subsp. infantis CECT 7210 and L. rhamnosus HN001, with GOS could improve activity against ETEC F4 in post-weaning piglets.
Materials and Methods
The trial was performed at the Experimental Unit of the Universitat Autònoma de Barcelona (UAB) and received prior approval (Permit No. CEEAH: 4026 DMAH: 10118) from the Animal and Human Experimental Ethical Committee of this Institution. The treatment, management, housing, husbandry, and slaughtering conditions conformed to European Union Guidelines (Directive 2010/63/EU; European Commission, 2010). All efforts were made to minimize animal suffering.
Animals, Housing, and Experimental Design
This trial was carried out as level 2 high-risk biosecurity procedures and involved personnel who had received appropriate training. A total of 96 male piglets (Landrace × Large White) × Pietrain, 21 (±2) days of age and weighting 5.04 ± 0.32 kg, were used. Only males were selected to reduce possible residual variability associated to sex as sample size is limited in trials involving pathogen challenges. All animals came from a high-sanitary-status farm in which mothers were not vaccinated against E. coli.
Piglets were transferred to the experimental unit located in the UAB, which consisted of four boxes of eight pens each (32 pens, three animals per pen). Each 2 m2 pen comprised a feeder and water nipple to provide feed and water ad libitum. All weaning rooms were equipped with an automatic heater and forced ventilation, and each pen had an individual heating light.
At arrival, animals were distributed among treatment groups according to their initial body weight. The trial was a completely randomized design that included five experimental groups: (i) positive control (CTR+), pigs not challenged with ETEC but orally inoculated with a sterile placebo solution; (ii) negative control (CTR−), pigs orally challenged with ETEC F4; (iii) probiotic (PRO), pigs receiving a combination of B. longum subsp. infantis CECT 7210 and L. rhamnosus HN001 and orally dosed with ETEC F4; (iv) prebiotic (PRE), pigs receiving galacto-oligosaccharides and orally dosed with ETEC F4; and (v) synbiotic (SYN), pigs receiving the combination of the products supplied in the PRO and PRE groups and orally dosed with ETEC F4. These challenged groups were equally distributed in three of the four rooms, while one full room was kept for non-inoculated control pigs; hence, the design was unbalanced. Each experimental group had six replicates, except for the non-challenged groups, which had eight replicates. In the challenged rooms, probiotic, and synbiotic treatments were distributed within four pens on one side of the room, and the control and prebiotic pens were on the other side of the room, separated by a corridor in between to avoid cross-contamination.
Probiotic Strains, Prebiotic, and Diets
The tested probiotics were B. longum subsp. infantis CECT 7210, supplied by Laboratorios Ordesa S.L., and L.s rhamnosus HN001 (Danisco USA Inc.). Both strains were provided lyophilized, containing 5 × 1010 and 3 × 1010 colony forming units (CFU) per gram of product, respectively, in a maltodextrin base. The lyophilized probiotics were mixed daily into the feed for a final dosage of 5.5 × 107 and 3.3 × 107 CFU/g, respectively. The feed was totally replaced daily. Before the trial, we confirmed the viability of the probiotics in the dry feed during the day.
The experimental prebiotic containing galacto-oligosaccharides (GOS) was in a syrup form and was thoroughly mixed with the diet by hand every day to guarantee a homogenous blend (no visual lumps). The final GOS concentration was 5% (w/w) trying to reach the amount of oligosaccharides provided by human milk (5–10 g/L; Kunz et al., 2017) and also based in previous studies supplementing GOS to neonatal and weaned piglets (Alizadeh et al., 2016; Tian et al., 2019; Wang et al., 2019). When mixed in the SYN diet, GOS was mixed prior to the probiotics.
Pre-starter diets were formulated in concordance with the nutrient requirement standards for pigs [National Research Council (NRC), 2012] and given in a mash form. The possible amino acid dilution in the PRE and SYN diets due to the incorporation of the prebiotic was compensated by the addition of synthetic amino acids: 0.5 g L-valine, 0.9 g L-lysine HCl, 1.2 g DL-methionine, 0.5 g L-threonine, and 0.2 g L-tryptophan per kg of feed. Details for their ingredient and chemical composition are given in Table 1.
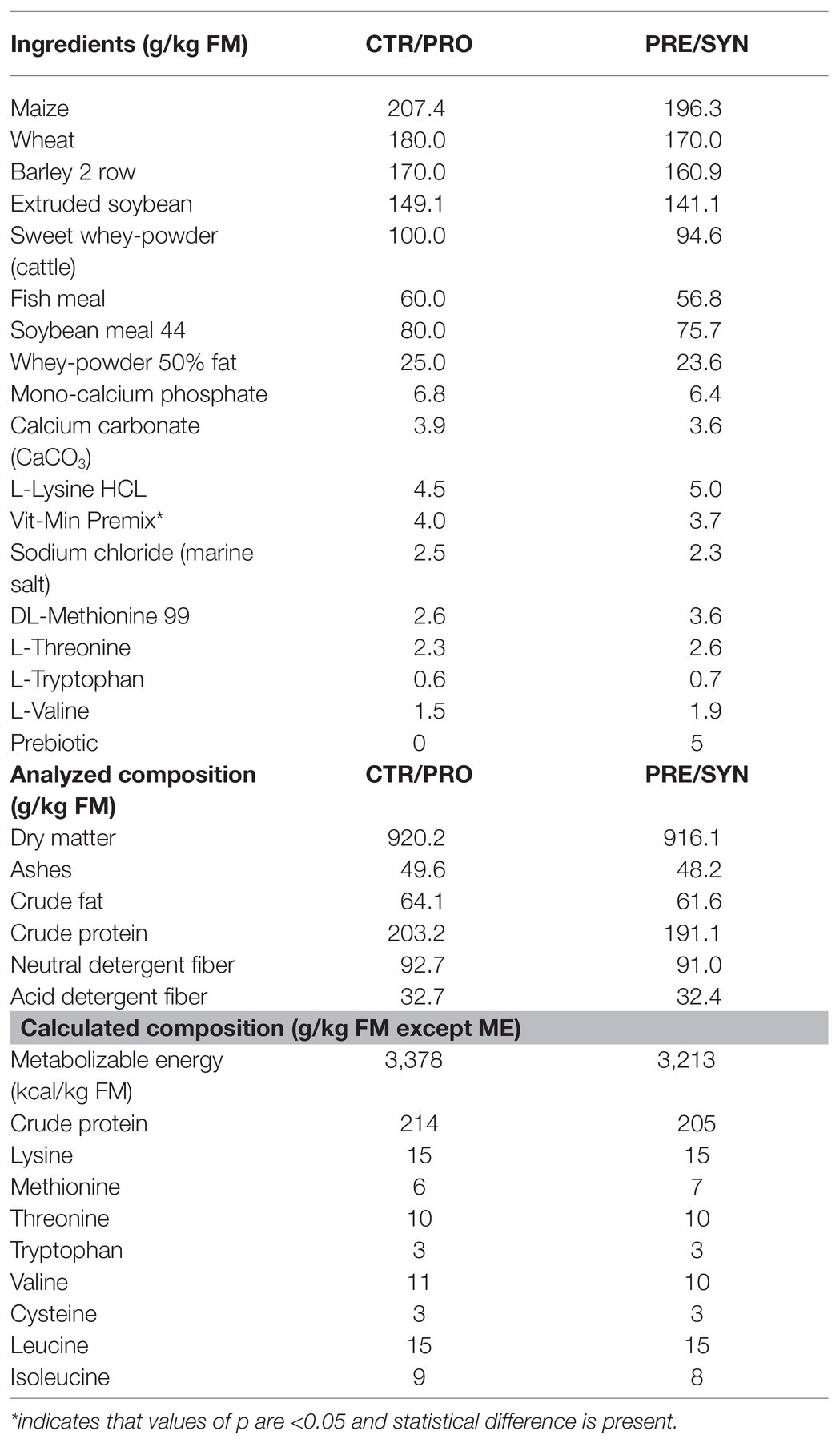
Table 1. Ingredient and nutritional composition of the diets.
Escherichia coli F4 Strain
The bacterial strain of ETEC F4 used was isolated from feces of 14-week old pigs and provided by the Infectious Diseases Laboratory (Ref. 30/14) of the UAB. This strain presented the following virulence factors: F4ab, F4ac, LT, STb, and EAST1, and it was negative for K99, F6, F18, F41, STa, VT1, VT2, and EAE. The oral inoculum was prepared by a 12-h overnight incubation at 37°C in brain heart infusion broth (Oxoid; Hampshire, England) with slow agitation (250 rpm) in an orbital incubator. A total volume of 6 ml from the culture was given directly to the animals; the final concentration was 1 × 109 CFU/ml. Inoculum concentrations were also determined before the inoculation by McFarland standards and were plated in Luria Agar (LA; made in house: tryptase, yeast extract, NaCl, and agar, Oxoid; Hampshire, United Kingdom) for the same day manual plate counting.
Experimental Procedure
The experiment lasted 15 days. Animals were offered the experimental diets from the first moment after arrival. After an adaptation period of 7 days (day 7), animals were orally challenged with the pathogen. The inoculum was given to the challenged groups via the oral route as a single dose of 6 ml of ETEC F4 bacterial culture containing 6 × 109 CFU. The same amount of sterile broth was administered to non-challenged piglets. To ensure that the animals had a full stomach at the moment of the oral challenge, pigs were starved for 12 h (night period from 8:00 PM to 8:00 AM) and feed was reintroduced 30 min before the inoculation.
From the challenge onward, animals were checked daily to determine clinical signs and evaluate their post-inoculation status – always by the same person. The fecal score was measured using a scale: 1 = solid and cloddy; 2 = soft with shape; 3 = very soft or viscous liquid; and 4 = watery or with blood. The rectal temperature was assessed with a digital thermometer (Accuvet, Sanchung City, Taiwan) on days 1 and 2 PI.
For microbiological analysis, fecal samples were collected aseptically after spontaneous defecation or by digital stimulation at arrival and on the day of the inoculation (day 0 PI). Fecal samples were always obtained from the largest animal of each pen (N = 32), except at day 4 PI, when the average piglet was sampled.
Animal performance was monitored. Individual body weight was registered at arrival (day 7) and on days 0, 4, and 8 PI, and feed intake was monitored daily, associated with the regular feed change to maintain probiotic viability. The average daily gain (ADG) and average daily feed intake (ADFI) were calculated by pen for the adaptation period (days 0–6), days 0–4 PI (days 7–11), and days 4–8 PI (days 12–15). The G:F ratio was calculated by pen for the adaptation period (days 0–6), post inoculation period (days 7–15), and total length of the trial (days 0–15). The animals received no antibiotic treatment.
On days 4 and 8 PI, one pig per pen was euthanized. On day 4 PI, the selected animal was the one with an intermediate initial BW, while on day 8 PI, the chosen piglet was the heaviest from each pen. Animals were euthanized and sequentially sampled during the morning of each day (between 8:00 and 13:00 h). An intramuscular injection containing 20 mg/kg ketamine (Ketamidor; Wels, Austria) and 2 mg/kg of xylazine (Xilagesic; Les Franqueses del Vallès, Spain) was given to the animals to induce deep sedation. Prior to injection of the euthanasia drug, 10 ml sample blood was taken from each animal via venipuncture of the cranial cava vein using 10 ml blood collection tubes without anticoagulant (Aquisel; Madrid, Spain). Right after blood sampling, pigs were euthanized with an intravenous injection of sodium pentobarbital (140 mg/kg BW, Euthasol, Le Vet B.V., Oudewater, Netherlands). Once dead, animals were bled, the abdomen was immediately open, and the gastrointestinal tract extracted.
A fecal sample taken from rectum was kept in ice and used for culture-based microbiology; it was analyzed within 4 h of collection. Subsequently, content from the ileum and proximal colon was collected and homogenized prior to pH determination with a pH meter calibrated on each day of use (Crison 52–32 electrode, Net Interlab; Barcelona, Spain). Subsamples of colonic and ileal contents were collected for different analyses. One aliquot of colonic content was kept frozen at −80°C for ETEC F4 quantification by real-time PCR (qPCR). A set of ileal and colonic digesta aliquots were stored at −20°C in H2SO4 solution (3 ml of content plus 3 ml of 0.2 N H2SO4) for ammonia (NH3) determination and an additional sample (~10 g) was also frozen at −20°C until analysis for short-chain fatty acids (SCFA) and lactic acid determination.
From each animal, 5 cm of distal ileum were collected, washed thoroughly with sterile phosphate-buffered saline (PBS), opened longitudinally, and scraped with a glass microscope slide to obtain the mucosa sample to determine the number of enterobacteria and coliforms attached to the intestinal mucosa. A subsample was also stored at −80°C for ETEC F4 quantification by qPCR.
For the histological study, 1 cm of ileum was removed, opened longitudinally, and washed thoroughly with 4% formaldehyde solution (Panreac; Castellar del Vallès, Spain) before fixing them by immersion in the same solution. Blood samples were centrifuged (3,000 g for 15 min) after blood coagulation; the obtained serum was stored at −20°C.
Analytical Procedures
Feed Analysis
Chemical analyses of the diets, including dry matter (DM), ash, crude protein, and diethyl ether extract, were performed according to the Association of Official Agricultural Chemists standard procedures (AOAC International, 1995). Neutral detergent fiber and acid-detergent fiber were determined according to the method of Van Soest et al. (1991).
Microbiological Analysis
For enterobacteria and coliform counts, samples were serially diluted in Ringer’s lactate solution (Sigma-Aldrich, Madrid, Spain) and proper dilutions seeded in MacConkey agar (Oxoid; Madrid, Spain) and eosin methylene blue agar (Scharlab; Barcelona, Spain). Plaques were incubated for 24 h at 37°C and colonies were manually counted. From colon content and ileal mucosal scrapings, the total bacterial DNA was extracted using QIAmp DNA Stool Mini Kit (Qiagen; West Sussex, United Kingdom) following the manufacturer’s instructions. The ETEC F4 concentration was than assessed by qPCR targeting the gene coding the F4 fimbria of E. coli, according to the procedure described by Hermes et al. (2013), using SYBR green dye with the ABI 7900 HT Sequence Detection System (PE Biosystems, Warrington, United Kingdom) with optical grade 96-well plates. The results were scored in five levels according to the number of gene copies per gram of fresh matter (FM). Scores were defined as following: negative = less than 4 logarithmic units of gene copies per gram FM; low = 4–5.5 logarithmic units of gene copies per gram FM; medium = 5.5–7 logarithmic units of gene copies per gram FM; high = 7–8.5 logarithmic units of gene copies per gram FM; and very high = more than 8.5 logarithmic units of gene copies per gram FM.
Short-Chain Fatty Acids, Lactic Acid, and Ammonia Analyses
Short-chain fatty acids and lactic acid analyses were performed on ileal and colonic digesta samples by gas liquid chromatography. Samples were subjected to acid-base treatment prior an ether extraction and derivatization with N-(tertbutyldimethylsilyl)-N-methyl-trifluoroacetamide (MBTSTFA) plus 1% tert-butyldimethylchlorosilane (TBDMCS) agent, using the method of Richardson et al. (1989), modified by Jensen et al. (1995).
Ammonia concentrations were assessed on ileal and colonic digesta samples using a gas-sensitive electrode (Hatch Co.; CO, United States) combined with a digital voltmeter (Crison GLP 22, Crison Instruments, S.A.; Barcelona, Spain) and following a procedure described by Hermes et al. (2009). Samples were diluted (1:2) in 0.16 M NaOH and, after homogenization, were centrifuged at 1500 g for 10 min. Once the ammonia was released, it was measured in the supernatants as a change in voltage (in mV).
Serum Analysis
Serum samples were analyzed for Tumor Necrosis Factor-α (TNF-α) and pig major acute-phase protein (Pig-MAP). The concentrations of TNF-α were determined by enzyme-linked immunosorbent assay (ELISA) using the Quantikine Porcine TNF-α kit (R&D Systems; Minneapolis, United States), using a 4-parameter logistic regression (4PL) fit for calibration curve adjustment. Pig-MAP concentrations were determined by a turbidimetric method using the TURBOVET Pig-MAP (Acuvet Biotech; Zaragoza, Spain). Samples were not diluted in any of the analyses.
Histological Analysis
For histological study, ileal samples were dehydrated and embedded in paraffin wax, sectioned at 4 μm, and stained with hematoxylin and eosin. Measurements of 10 different villus-crypt complexes per sample were considered including counting of intraepithelial lymphocytes (IEL), Goblet cells (GC), and the number of mitotic cells in each complex. Analyses were performed with a light microscope (BHS, Olympus, Barcelona, Spain) following the procedure described by Nofrarías et al. (2006).
Piglet Genotyping for ETEC Susceptibility
The mucin 4 (MUC4) gene has been suggested as one of the candidate gene associated with piglet susceptibility to ETEC F4 (Jørgensen et al., 2003; Luise et al., 2019a). Thus, in the present study, the animals were genotyped for the polymorphism described by Jørgensen et al. (2003). For MUC4 genotype determination, hair follicles were collected from 81 pigs. DNA was extracted following the procedure described by Luise et al. (2019b). A restriction fragment length polymorphism PCR (PCR-RFLP) was performed following the guidelines described by Jørgensen et al. (2003). Pigs were classified into two groups: susceptible pigs having the MUC4GG or MUC4CG genotype (MUC4+) or resistant pigs having MUC4CC homozygotes (MUC4−).
Statistical Analysis
The results are expressed as means with their standard errors unless otherwise stated. Microbiological counts were log transformed for analysis. A two-way analysis of variance (ANOVA) was used to examine the effect of the five experimental treatments and the MUC4 (MUC4+ and MUC4−) with the following model:
where Yijk relates to each observation of the outcome variable, m is the global mean, treati is the main effect of treatment, MUC4j is the main effect of MUC4 polymorphism, and treat*MUC4ij corresponds to the interaction between treatment and MUC4. Finally, eij is the experimental error term. Regarding MUC4 effect and interaction term, they were removed from the model when found not to be significant.
The effects on the post-slaughter measurements were examined using the R v3.4 (R Core Team, 2013) lm function for two-way ANOVA, with the treatment and MUC4 effects as main effects. When the MUC4 effect was not observed (p > 0.05), one-way ANOVA was performed with only the treatment effect. For E. coli F4 prevalence values, data were subjected to frequency analysis using the fisher.test function in the same package.
ADFI and daily fecal scores were also analyzed using the lme4 package (Bates et al., 2015) lmer function for a generalized linear mixed model (GLMM) with a treatment-by-time interaction term and considering the animal as the random effect.
The experimental unit was the pen. For all analyses, significance was set at p < 0.05 and p ≥ 0.05, but ≤ 0.10 were considered to indicate a statistical trend. When treatment effects were established, the mean comparison was adjusted with the Tukey-Kramer test. Data are presented as means and residual standard error (RSE).
Results
Following the oral challenge with ETEC, animals developed moderate clinical signs of diarrhea that began to resolve spontaneously at the end of the study. Eight spontaneous casualties occurred (by group: 3 CTR−, 3 PRE, 1 PRO, and 1 SYN), and no humane euthanasia was required. Table 2 presents the analysis of mucin 4 (MUC4) polymorphism for each treatment group.
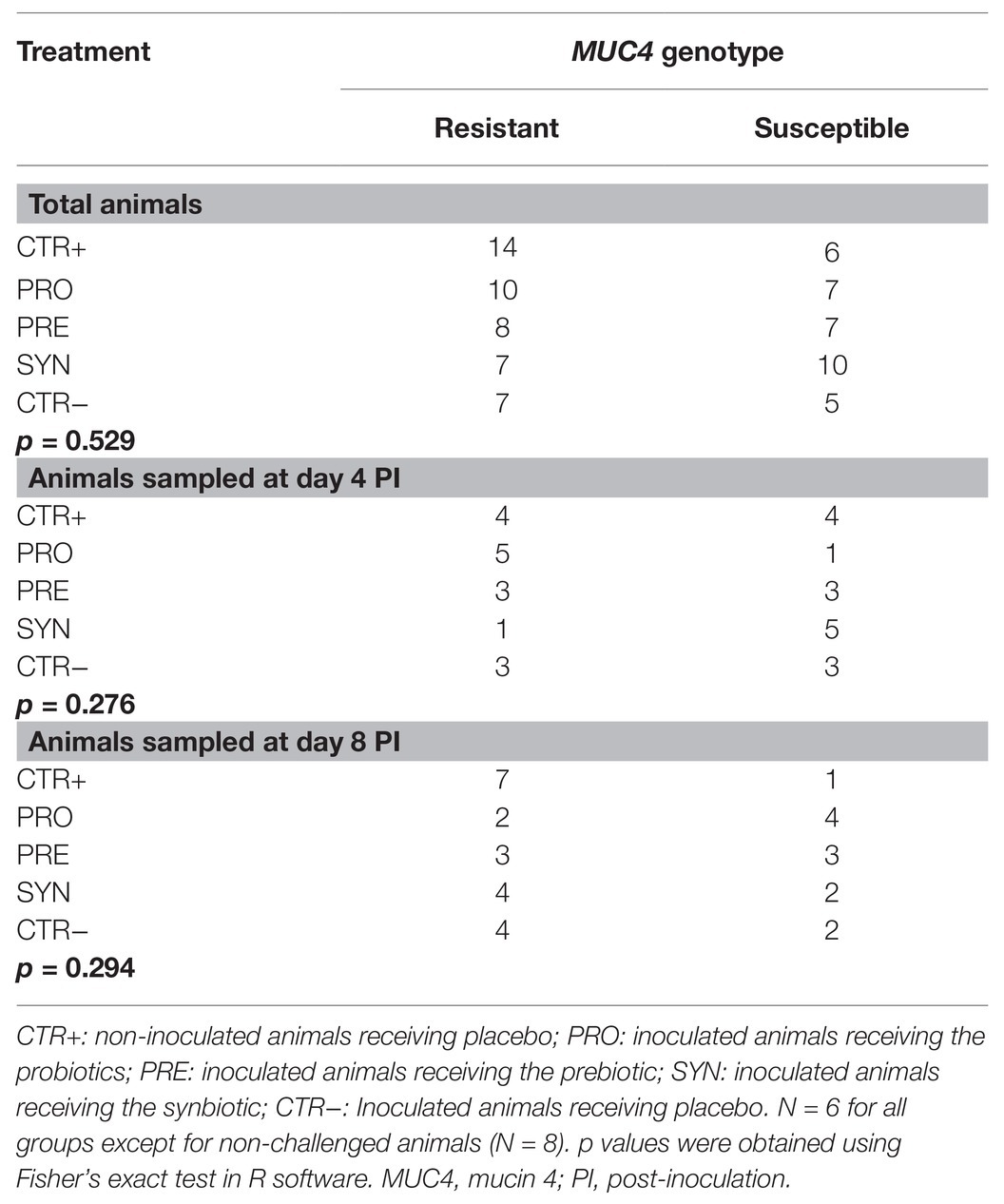
Table 2. Distribution of resistant and susceptible animals for MUC4 gene in each experimental group.
Performance Parameters
Table 3 presents the results obtained for body weight (BW), average daily feed intake (ADFI), average daily weight gain (ADG), and gain to feed (G:F) ratio. During the 1st week, the animals from all experimental groups had a similar feed intake. However, after pathogen inoculation, all challenged groups showed numerical reductions in intake, although the differences only reached statistical significance for the CTR− and SYN groups for the days 0–4 post-inoculation (PI) period (p = 0.002) and for the SYN group for the days 4–8 PI period (p < 0.001).
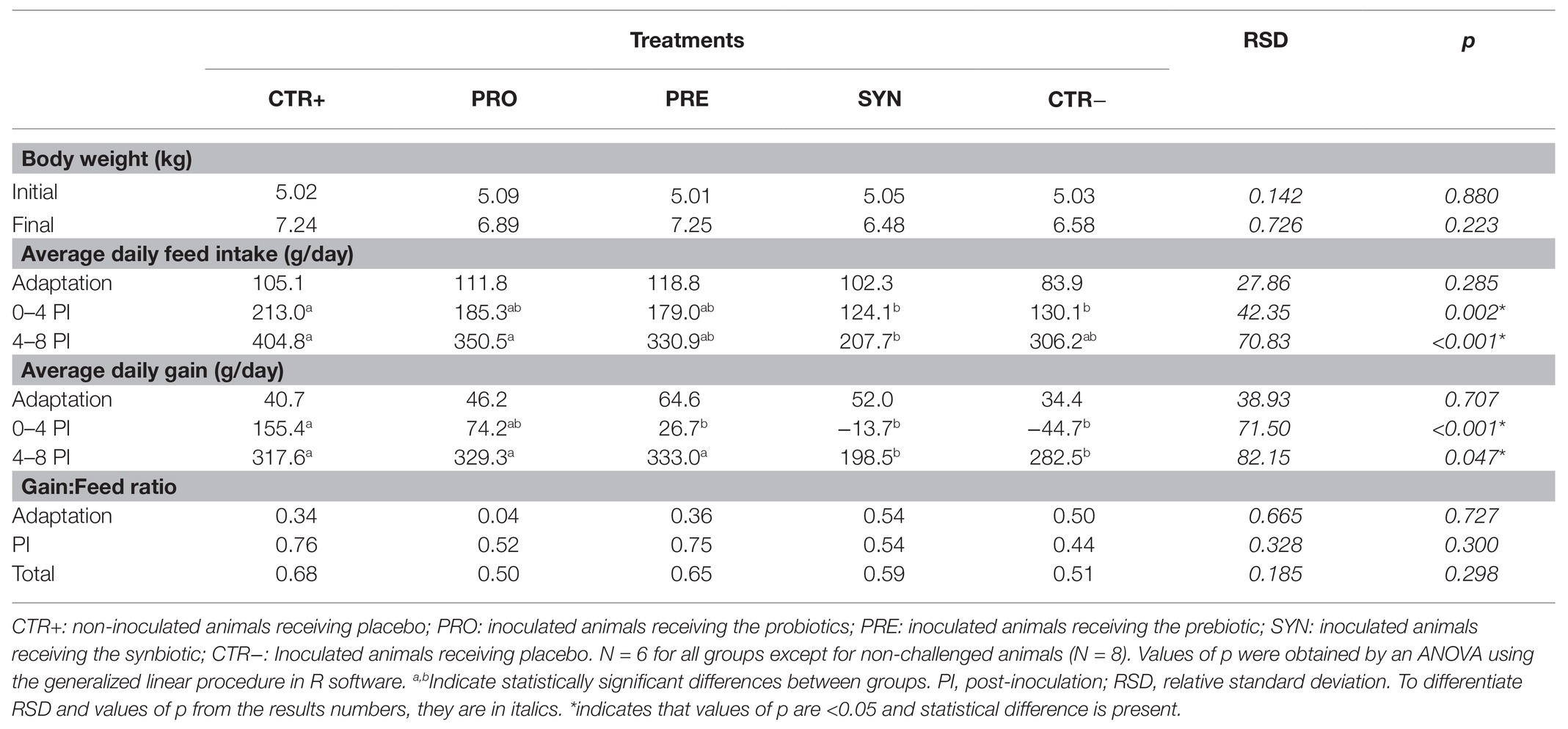
Table 3. Effects of experimental treatments on feed intake and weight gain.
Figure 1 shows the evolution of feed intake per day during the experimental period. There were statistical differences between groups beginning on day 3. The SYN group was the most affected: this group consistently showed the lowest feed intakes; being significantly different from CTR+ on days 3, 4, 5, 7, and 8 PI.
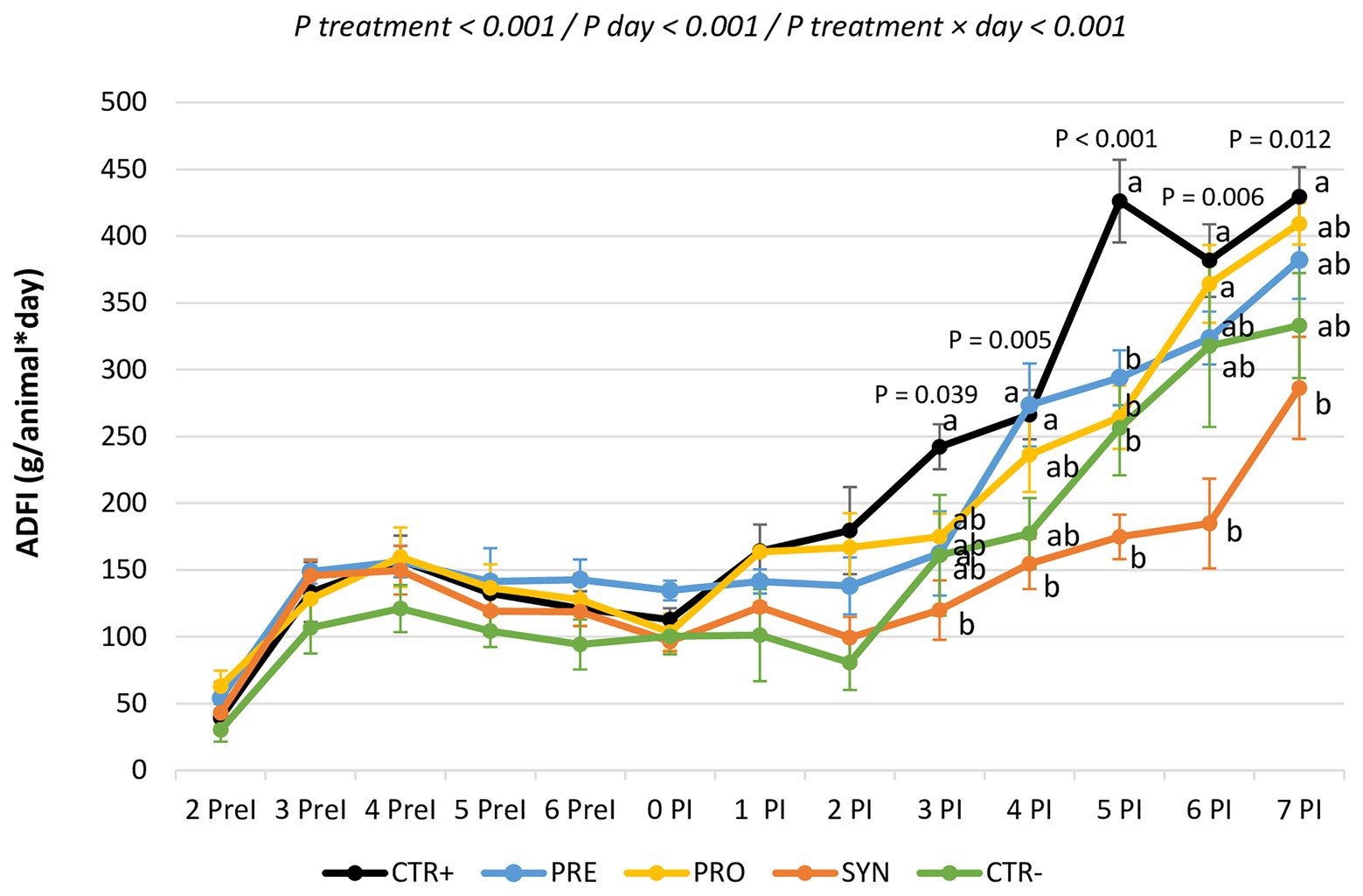
Figure 1. Evolution of feed consumption for the experimental groups during the entire experimental period. CTR+: non-inoculated animals receiving placebo; PRO: inoculated animals receiving the probiotics; PRE: inoculated animals receiving the prebiotic; SYN−: inoculated animals receiving the synbiotic; CTR−: inoculated animals receiving placebo. N = 6 for all groups except for non-challenged animals (N = 8). a and b indicate statistically significant differences between groups. Bars correspond to the standard error. ADFI, average daily feed intake; PreI, pre-inoculation; PI, post-inoculation.
Similarly, ADG did not differ between treatments during the 1st week of the trial, but it decreased after the challenge. During the first period after the inoculation (days 0–4 PI), all the challenged groups showed significant decreases in growth, except for the PRO group, in which numerical reductions in weight gains did not reach statistical significance. In the second phase (days 4–8 PI), the PRO and PRE groups showed a fast recovery of weight gain. Indeed, they reached similar levels (even numerically higher) to non-inoculated piglets. However, the SYN group showed a tendency toward lower weight gain compared to the CTR+ group (p = 0.083). There were no differences among the groups for G:F ratio during the trial.
Clinical Signs
Figure 2 shows the evolution of fecal consistency along with the post-infection period. ETEC F4-challenged animals showed higher fecal scores (lower fecal consistency) immediately after the inoculation; these differences were significant on days 1 and 2 PI and recovered from day 3 PI onward. MUC4+ animals had worse scale numbers compared to MUC4− animals on days 1 and 2 PI (2.03 vs. 2.47; p = 0.009 and 1.95 vs. 2.34; p = 0.013 for not susceptible and susceptible on days 1 and 2 PI, respectively). Among the challenged groups, the PRO group had the best consistency and the SYN group the worst (1.60, 1.87, 1.95, 2.07, and 2.10 for CTR+, PRO, PRE, SYN, and CTR−, respectively). There was no treatment × MUC4 interaction. Rectal temperature was within the normal range and was not affected by the challenge, experimental diets, or MUC4 status.
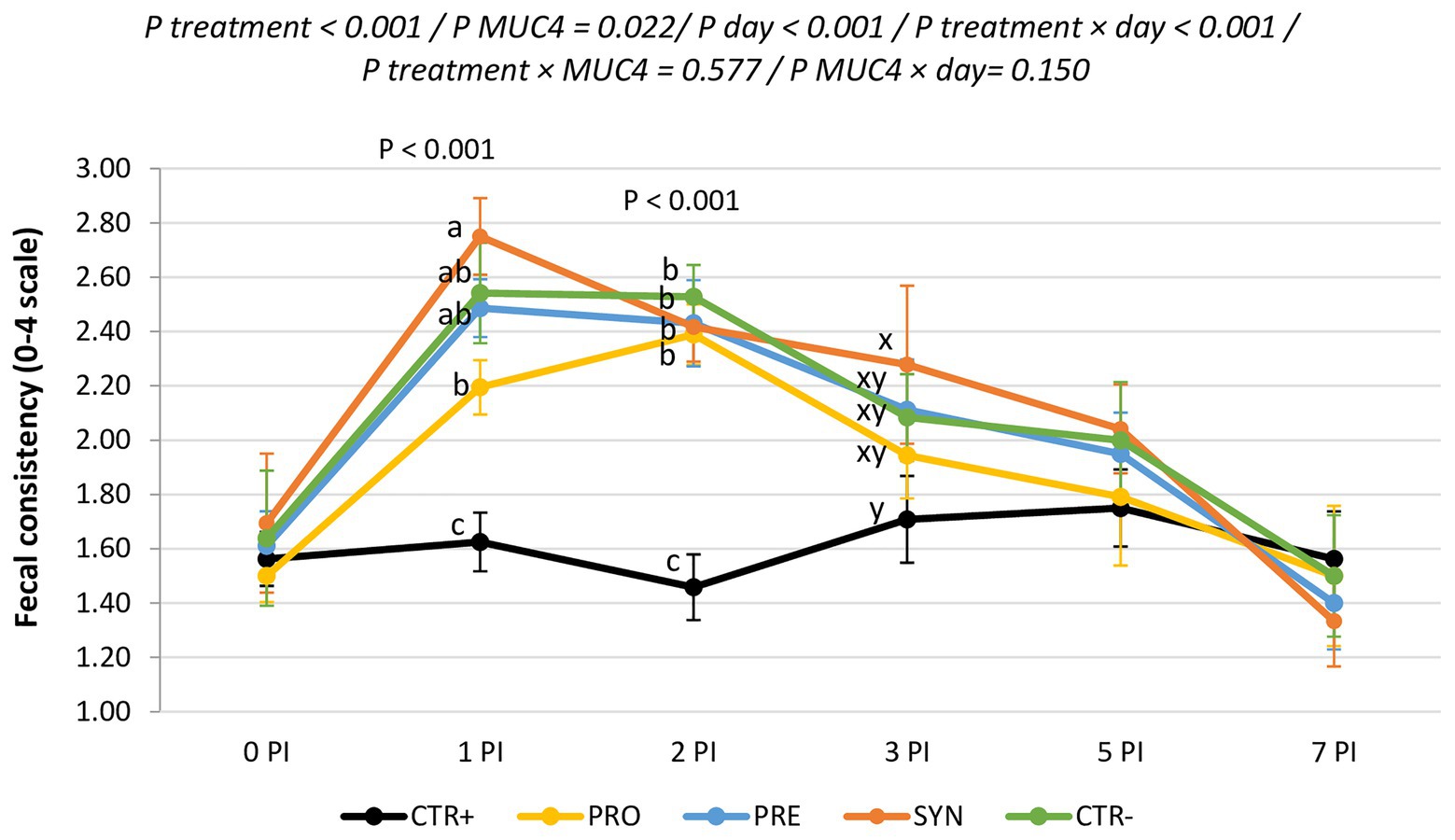
Figure 2. Evolution of average fecal scores for the different experimental groups during the post-inoculation (PI) period. CTR+: non-inoculated animals receiving placebo; PRO−: inoculated animals receiving the probiotics; PRE−: inoculated animals receiving the prebiotic; SYN−: inoculated animals receiving the synbiotic; CTR−: inoculated animals receiving placebo. N = 6 for all groups except for non-challenged animals (N = 8). MUC4 represents the effect of polymorphism of the MUC4 gene. a,b, and c indicate statistically significant differences between groups. x and y indicate statistical trends among groups. Bars correspond to the standard error. PI, post-inoculation.
Microbiological Analysis
Table 4 shows enterobacteria and coliform plate counts from fecal samples taken at the arrival of the animals, before the oral challenge, and at days 4 and 8 PI. The table also includes plate counts from ileal mucosa scrapings at day 4 and 8 PI.
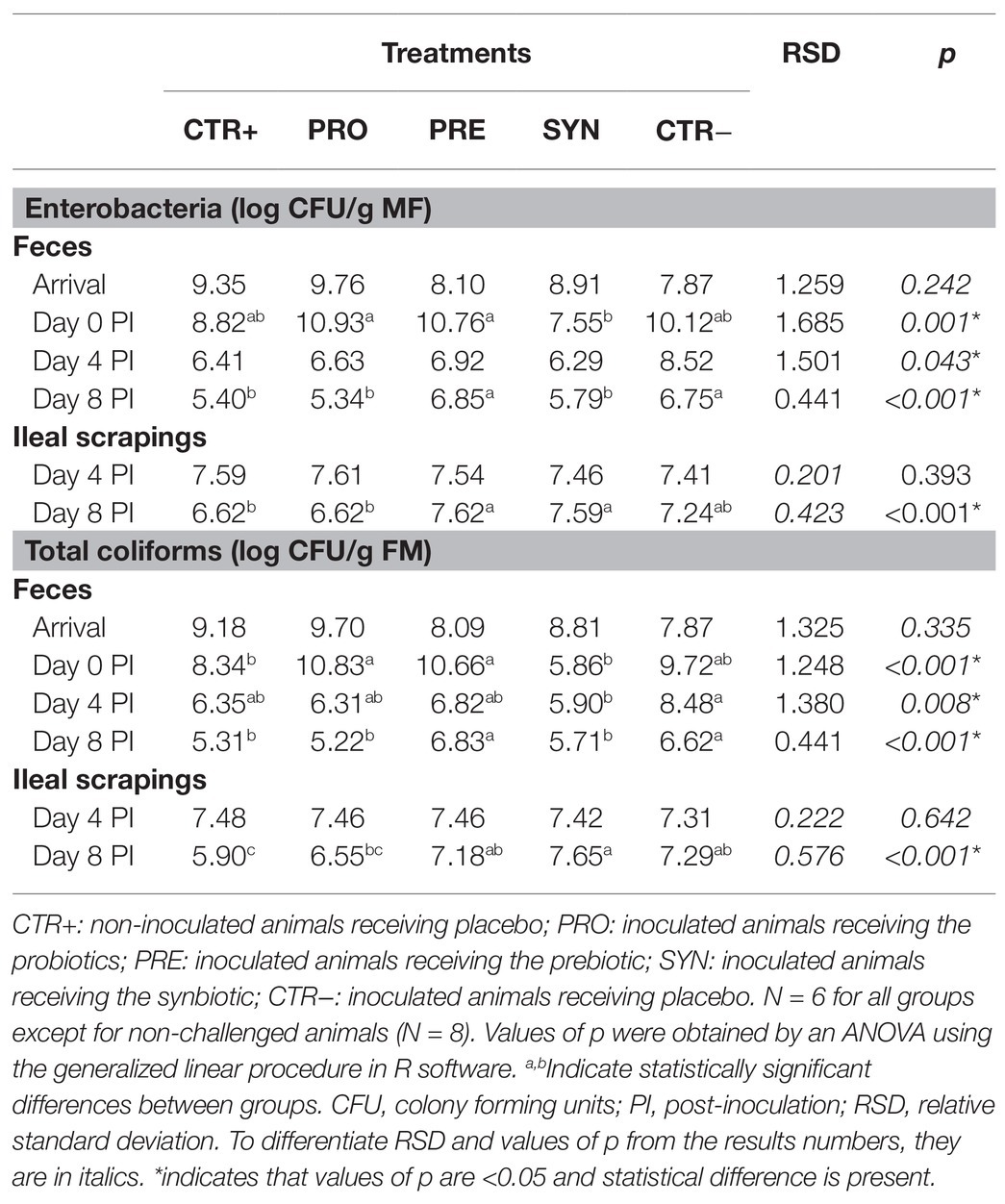
Table 4. Effects of experimental treatments on enterobacteria and coliform counts in fecal samples and ileal scrapings.
There were no differences between experimental groups on the day of arrival. However, at the end of the adaptation phase, the SYN group showed lower values of fecal enterobacteria (p = 0.001) and coliforms (p < 0.001) compared to the PRE and PRO groups. On day 4 PI, the SYN group was the only one with lower fecal coliforms counts compared to the CTR− group (p = 0.023). On day 8 PI, only the PRE and CTR groups maintained higher plate counts compared to non-challenged pigs (p < 0.001 for both groups).
MUC4 also had an impact on fecal counts of enterobacteria and coliforms on day 4 PI: resistant animals had lower counts compared to susceptible carriers (enterobacteria 6.27 vs. 7.44 CFU/g; P MUC4 = 0.015; coliforms 6.23 vs. 7.22 CFU/g; P MUC4 = 0.010). The interaction between MUC4 and treatment was not significant, except for fecal enterobacteria before the challenge, when there were differences in the SYN group compared to the other treatment groups only in non-susceptible pigs who had a lower number of enterobacteria (9.17, 10.86, 10.77, 5.92, and 10.05 CGU/g for CTR+, PRO, PRE, SYN, and CTR−, respectively; P interaction = 0.003).
Regarding ileal scrapings, only on day 8 PI were there differences between treatments (p < 0.001). Specifically, the PRO group was the only one not that different from the CTR+ group.
The ETEC F4 quantification results by qPCR in colonic digesta and ileal scrapings showed that MUC4 status significantly affected the prevalence of ETEC F4 in colonic digesta at day 4 PI. The challenge effect was much clearer in carrier animals (P MUC4 < 0.001), with no significant difference among treatments (Figure 3). At day 8 PI, there was an interaction between MUC4 and treatment, with a higher prevalence of ETEC F4 in the SYN group – but only in the MUC4 non-carrier animals. MUC4 did not significantly impact the ileal scraping counts at days 4 or 8 PI.
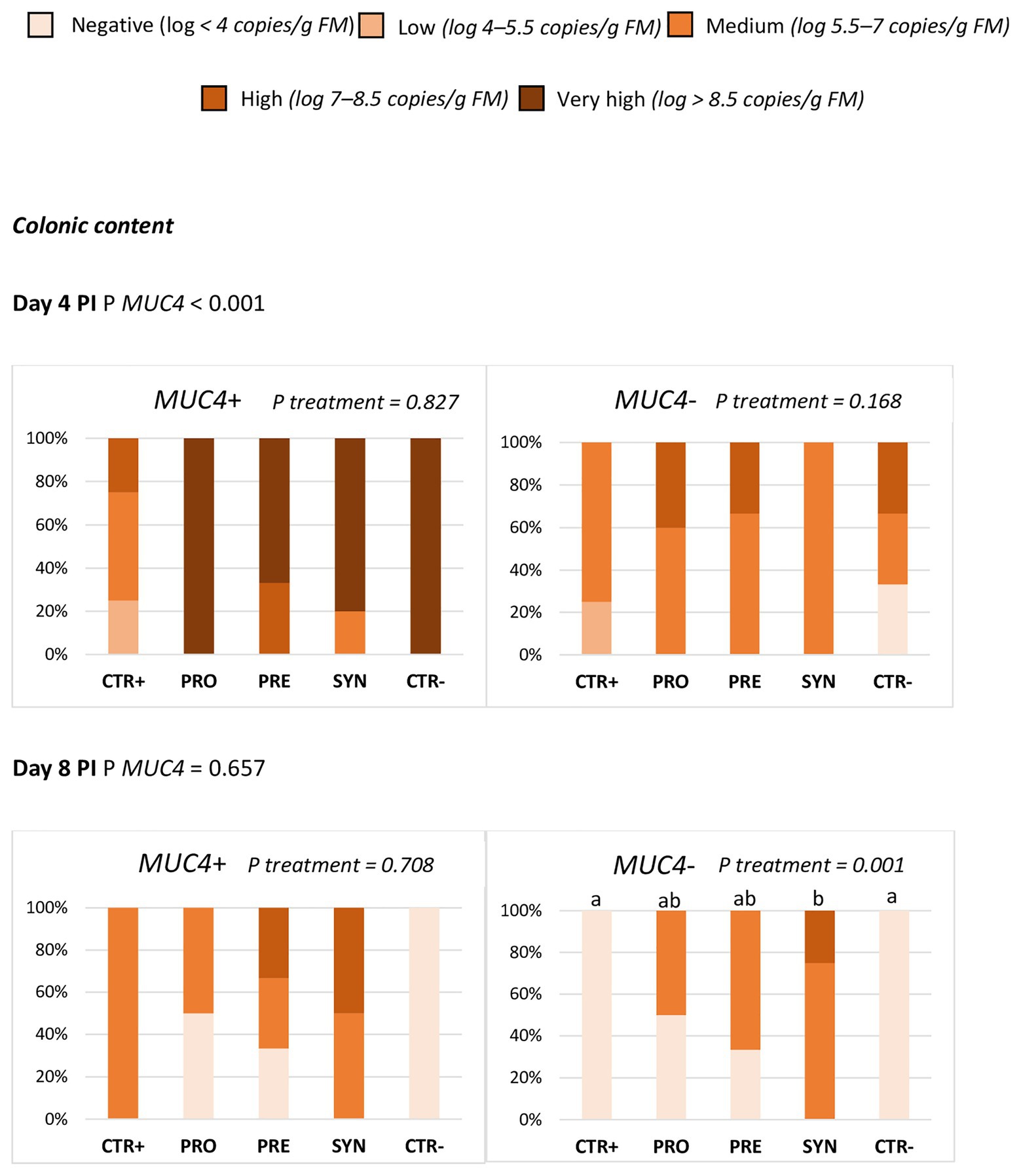
Figure 3. Effect of MUC4 polymorphism on the percentage of animals with different levels of ETEC F4 counts in colon content on days 4 and 8 post-inoculation (PI). Different animals were sampled on days 4 and 8 post-inoculation (PI). CTR+: non-inoculated animals receiving placebo; PRO−: inoculated animals receiving the probiotics; PRE: inoculated animals receiving the prebiotic; SYN: inoculated animals receiving the synbiotic; CTR−: inoculated animals receiving placebo. N = 6 for all groups except for non-challenged animals (N = 8). Values of p were obtained using Fisher’s exact test in R software. a and b indicate statistically significant differences between groups. MUC4+, Mucin 4 susceptible genotype; MUC4−, Mucin 4 resistant genotype; PI, post-inoculation.
When considering all animals, regardless of MUC4 status (Figure 4), there were differences between treatments in the colonic prevalence of ETEC F4. On day 4 PI, the SYN group showed a higher prevalence compared to the CTR+ and similar to the CTR− group; the PRO and PRE groups showed intermediate levels (p = 0.01). On day 8 PI, the SYN group maintained high excretion levels (SYN vs. CTR+; p = 0.002), while animals from the CTR− group recovered from the challenge and the values decreased; the PRO and PRE groups showed intermediate values.
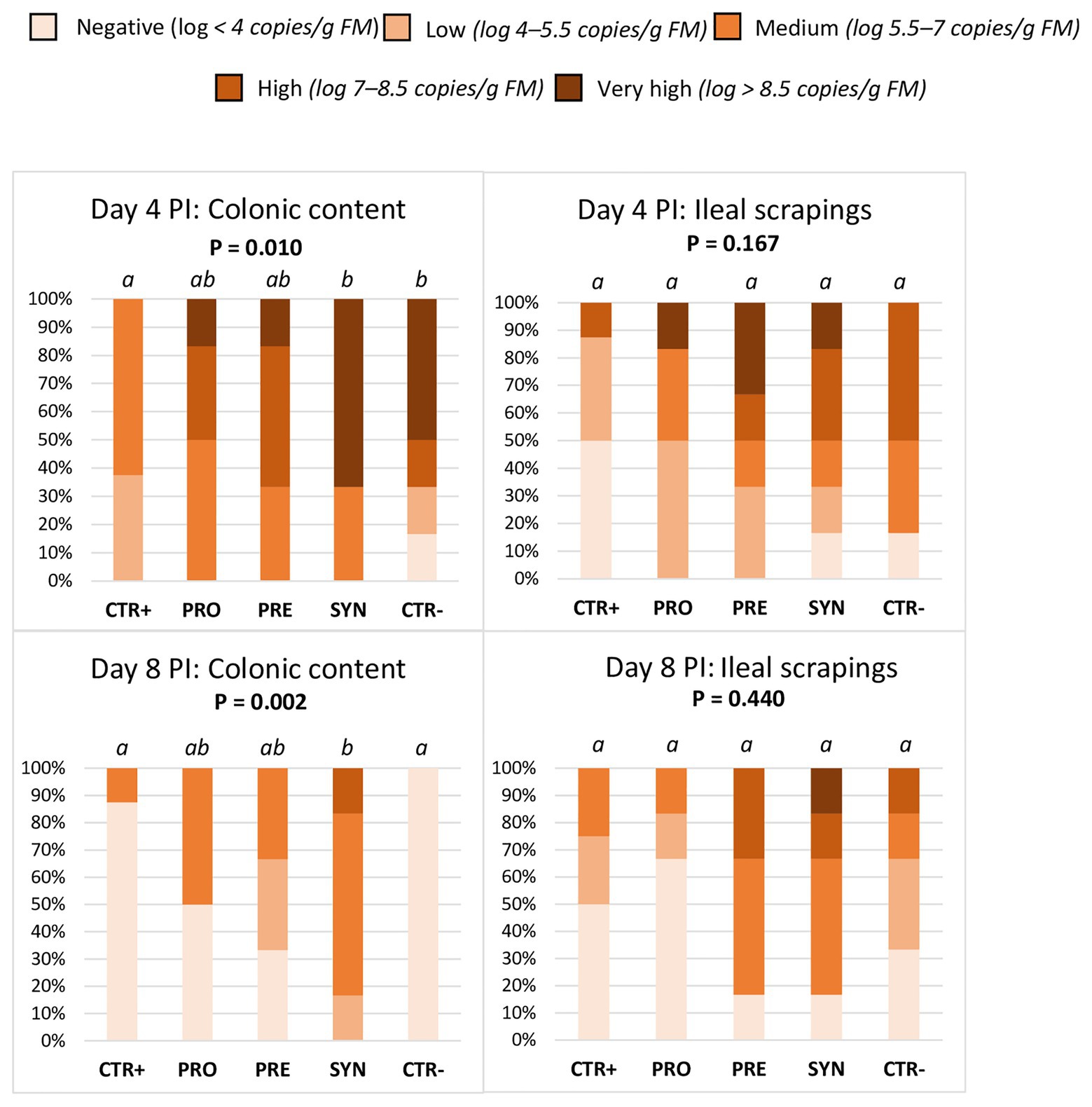
Figure 4. Effect of experimental treatments on the percentage of animals with different levels of ETEC F4 counts in colonic digesta and ileal scrapings on days 4 and 8 post-inoculation (PI). Different animals were sampled on day 4 PI and 8 PI. CTR+: non-inoculated animals receiving placebo; PRO−: inoculated animals receiving the probiotics; PRE: inoculated animals receiving the prebiotic; SYN: inoculated animals receiving the synbiotic; CTR−: inoculated animals receiving placebo. N = 6 for all groups except for non-challenged animals (N = 8). Values of p were obtained using Fisher’s exact test in R software. a and b indicate statistically significant differences between groups. FM, fresh matter; PI, post-inoculation.
Intestinal Fermentation
Table 5 shows values of intestinal pH, ammonia, lactic acid, and short-chain fatty acid (SCFA) concentrations in ileal and colonic content for the different treatments.
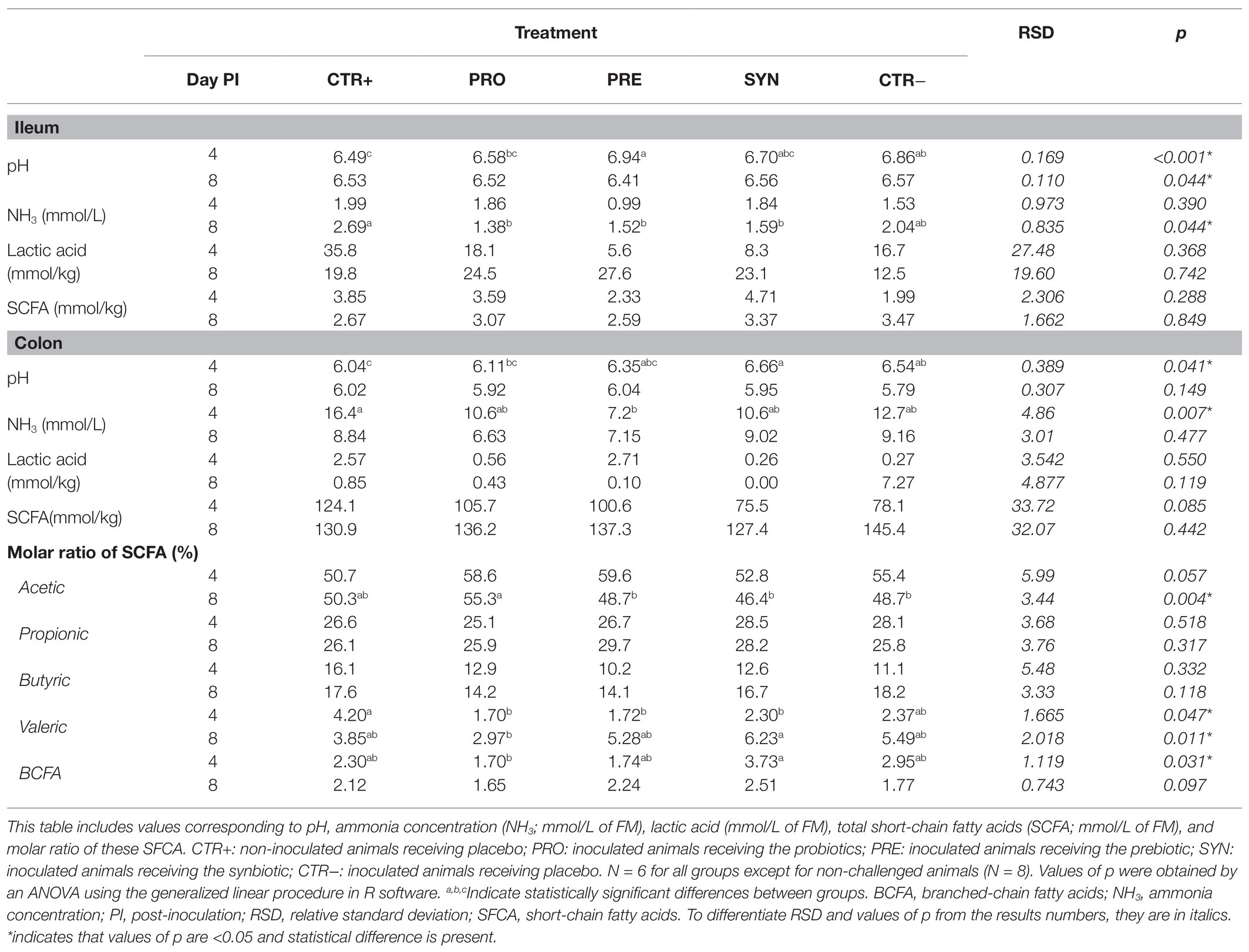
Table 5. Effects of experimental treatments on ileal and colonic fermentation.
At the ileal level, the PRE group had the highest pH values at day 4 PI; they were different from the CTR+ and PRO groups (p < 0.05), but not from the other challenged groups. At day 8 PI, the PRE group had the lowest pH values (P treatment = 0.044). Ammonia concentrations diminished at day 8 PI in the PRO, PRE, and SYN groups compared to the CTR+ and CTR− groups (p = 0.044). There was no treatment effect in lactic acid or total SCFA concentrations.
In the colon, the pH was numerically increased at day 4 PI in all challenged groups, although the differences compared to the CTR+ group were only significant for the SYN and CTR− groups (P treatment = 0.041). The PRE group was the only treatment with reduced ammonia values at day 4 PI compared to the CTR+ group (P treatment = 0.007). There were no differences in lactic acid concentration at any time.
Regarding total SCFA, the SYN and CTR− groups showed a trend for a decreased concentration compared to the other groups (P treatment = 0.085) at day 4 PI but not at day 8 PI. At day 8 PI, there was a marked increase in lactic acid concentration in the CTR− group, although differences with other treatments did not reach statistical significance (p = 0.119). The molar ratios of different SCFA showed some changes related to the experimental treatments. On day 4 PI, there was a numerical trend for an increased acetic acid percentage in the challenged groups (P treatment = 0.057), and at day 8 PI, the PRO group showed a higher acetic acid concentration compared to other inoculated piglets (p = 0.004). The PRO group showed lower percentages of branched chain fatty acids (BCFA) compared to the SYN group (p = 0.049), and valeric acid was reduced in the PRO, PRE, and SYN groups compared to the CTR+ group at day 4 PI (P treatment = 0.047).
Notably, these treatment-mediated differences with regard to the fermentation profile were influenced by MUC4 polymorphism when analyzed at day 8 PI. The identified interactions are shown in Figure 5. The SYN group animals showed a lower colonic pH and a higher amount of total SCFA in the colon, but only in the carrier pigs (P interaction < 0.001). Regarding valerate, the SYN group had a sharp increase in the molar ratio, but only in susceptible pigs (P interaction = 0.019). The BCFA molar ratio was increased in resistant animals in the SYN group and in susceptible animals in the PRE group. Finally, piglets with a MUC4 resistant phenotype showed increased colon ammonia levels on day 4 PI (13.31 vs. 9.79 mmol/L; P MUC4 = 0.006); there was no interaction with treatments.
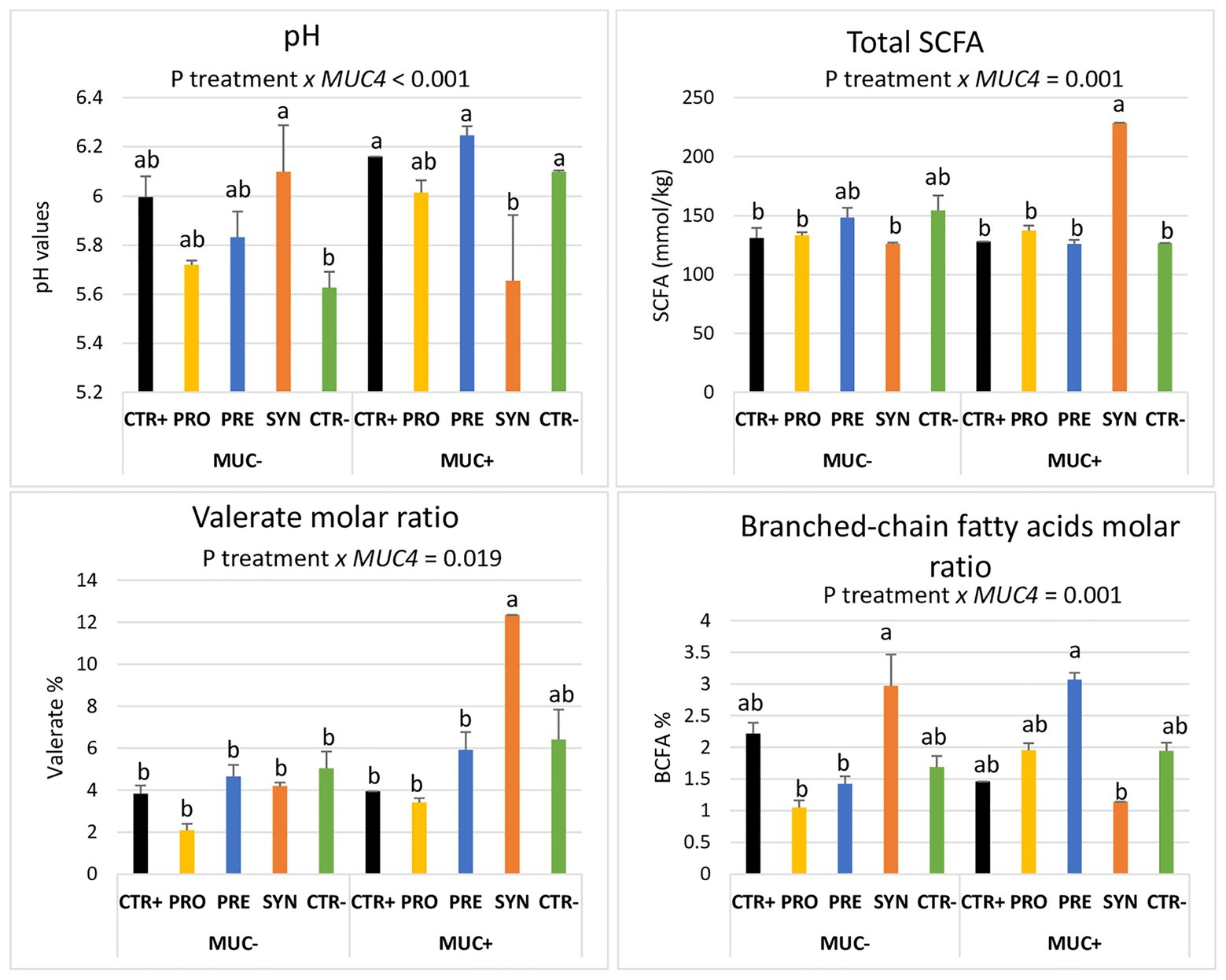
Figure 5. Interactions between MUC4 gene polymorphism and diets on pH values, total short-chain fatty acids (SCFA), valerate molar ratio, and branched-chain fatty acids molar ratio (BCFA) on day 8 PI. CTR+: non-inoculated animals receiving placebo; PRO−: inoculated animals receiving the probiotics; PRE−: inoculated animals receiving the prebiotic; SYN−: inoculated animals receiving the synbiotic; CTR−: inoculated animals receiving placebo. N = 6 for all groups except for non-challenged animals (N = 8). MUC4 represents the effect of polymorphism of the MUC4 gene. a and b indicate statistically significant differences between groups. Bars correspond to standard error. BCFA, branched-chain fatty acids; MUC4+, Mucin 4 susceptible genotype; MUC4−, Mucin 4 resistant genotype; SCFA, short-chain fatty acids.
Immune Response
Table 6 presents values corresponding to serum levels of the pro-inflammatory cytokine tumor necrosis factor alpha (TNF-α) and the acute-phase protein pig major acute-phase protein (Pig-MAP).
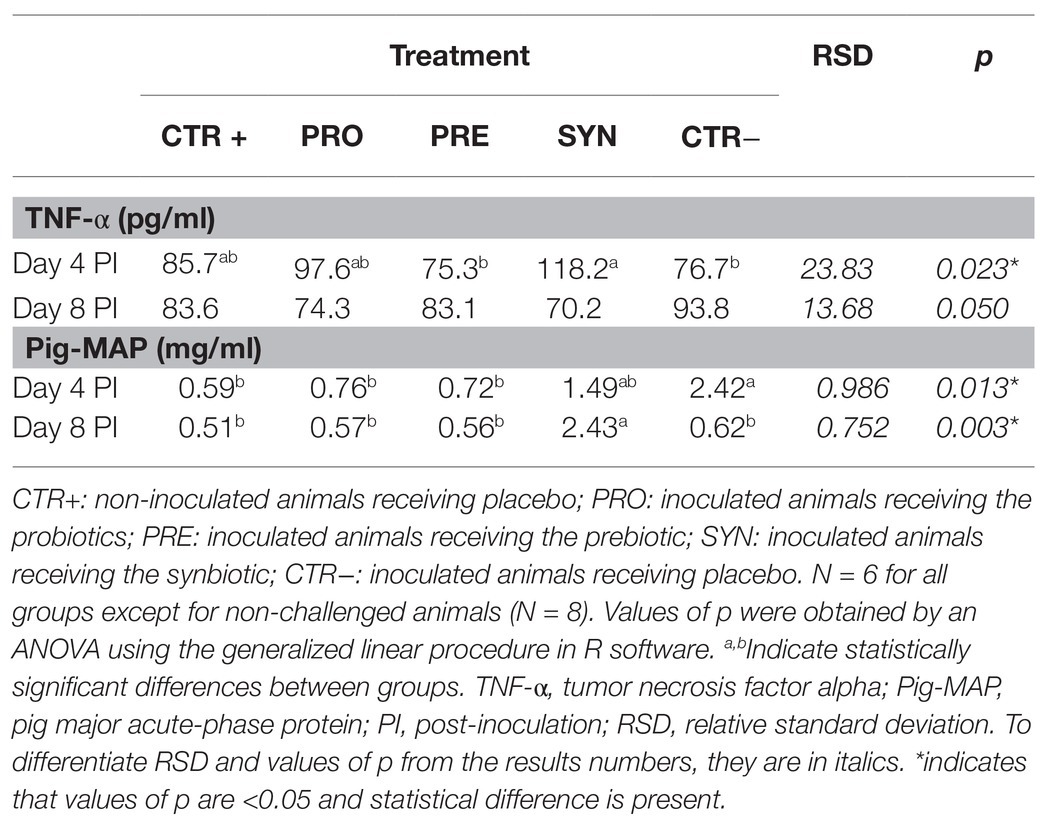
Table 6. Effect of experimental treatments in serum levels of acute-phase protein Pig-MAP and TNF-α.
The TNF-α concentration on day 4 PI was higher in the SYN group compared to the CTR− and PRE groups (P treatment = 0.023); any differences did not reach significance with the rest of the groups. At day 8 PI, the SYN group showed the lowest values (p = 0.050).
On day 4 PI, the Pig-MAP value was higher in the CTR− compared to the CTR+ group. The PRO and PRE groups had levels closer to the CTR+ group, and the SYN group showed intermediate levels (P treatment = 0.013). On day 8 PI, the Pig-MAP concentration was clearly higher in the SYN group, whereas the rest of challenged groups resembled the CTR+ group level (P treatment = 0.003).
There were no effects due to MUC4 gene polymorphism.
Intestinal Histological Structure
The effects of the experimental treatments on villi height, crypt depth, and mitosis number are shown in Table 7.
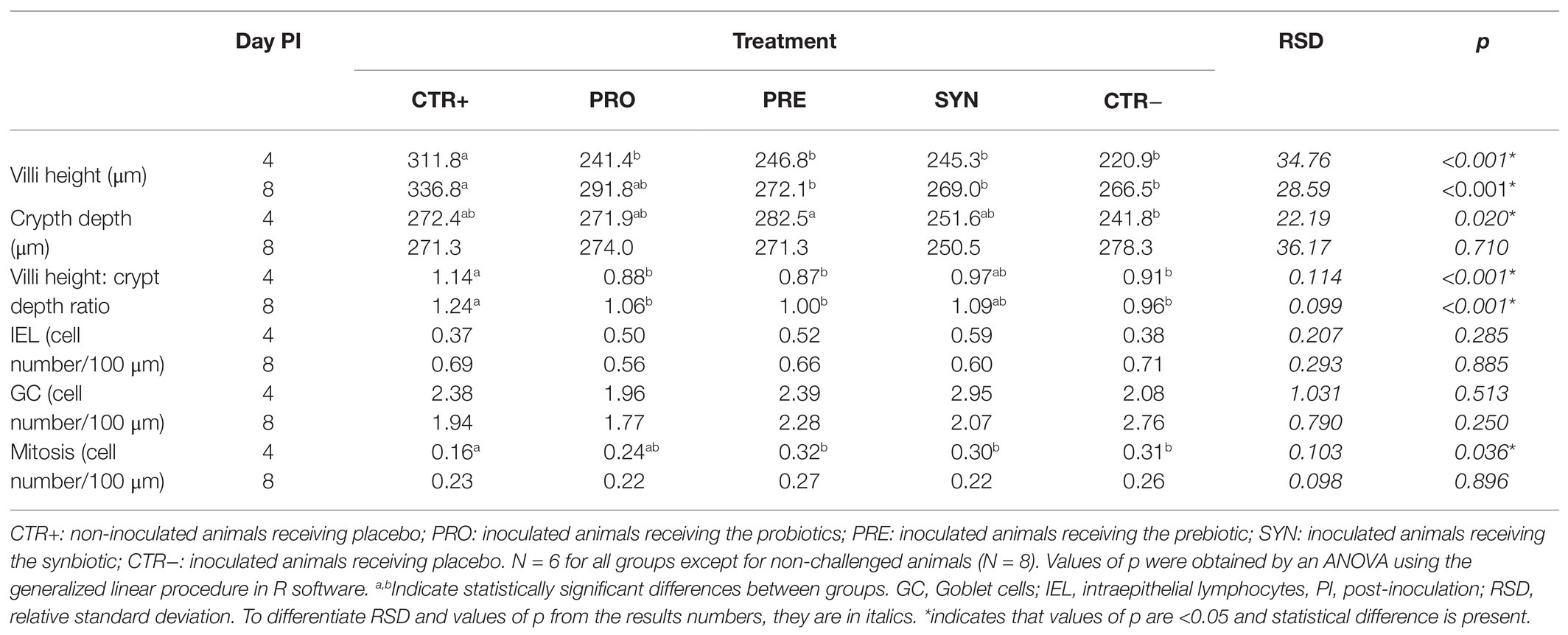
Table 7. Effects of treatments on ileal histomorphological parameters on days 4 and 8 post-inoculation (PI).
The impact of the ETEC F4 oral challenge was evidenced by the ileal epithelium structure. On day 4 PI, all the challenged groups showed a reduction in the villi height (p < 0.001), a phenomenon which was still significant on day 8 PI, except for the PRO group (P treatment < 0.001). When analyzing crypt depth on day 4 PI, the CTR− group had the lowest values, whereas the PRE group had the highest; the difference between the two treatments was statistically significant (p = 0.028). According to this result, the villous height:crypt depth ratio was modified by the oral challenge on both sampling days (p < 0.001), with reductions in the ratio for all the challenged groups except SYN. In that group, the reductions did not reach statistical significance compared to the CTR+ group (p = 0.064).
The challenge was also associated with a higher number of mitotic cells (P treatment = 0.036), although this increase was not significant in the PRO group when compared to the CTR+ group (p = 0.649) at day 4 PI. Regarding goblet cells and interepithelial lymphocytes (IELs), there were no significant differences related to the experimental treatments.
MUC4 significantly affected the number of mitotic cells on day 4 PI: susceptible animals presented reduced mitosis compared to resistant animals (0.23 vs. 0.29 cell number/100 μm, P MUC4 = 0.049). Furthermore, the villous height:crypt depth ratio on day 8 PI showed an interaction effect (p = 0.004). Specifically, the SYN group exhibited a higher villous:crypt ratio compared to the challenged groups, but only in the susceptible animals (1.33, 1.07, 1.01, 1.32, and 0.95 for CTR+, PRO, PRE, SYN and CTR−, respectively).
Discussion
In this study, a piglet model of ETEC F4 colibacillosis was used to evaluate the efficacy of treatment with B. longum subsp. infantis CECT 7210 and L. rhamnosus HN001, galacto-oligosaccharides, and their combination. The current experimental trial clearly demonstrated the effects caused by the pathogen challenge. Specifically, in all the challenged groups, there was a decrease in feed intake, followed by a reduction in weight gain. Piglets also showed impaired fecal consistency immediately after the inoculation, with evident effects on the intestinal epithelium structure and the Pig-MAP response.
The work also analyses the possible role of the MUC4 gene in the development of the disease. Actually the presence of ETEC F4-specific receptors on the intestinal brush border of pigs has been associated with MUC4 polymorphism (Peng et al., 2007; Trevisi et al., 2009) and suggested as criteria for the inclusion of animals in ETEC F4 challenge models (Luise et al., 2019a). As expected, in the present study, animals that were heterozygous or homozygous for the susceptible MUC4 allele showed the elevated levels of enterobacteria, coliforms, and ETEC F4 on some of the sampling days, as well as a worsened fecal consistency. These results support the usefulness of this candidate gene as marker for the genetic selection of farmed pig toward increased resistance to diarrhea (Jørgensen et al., 2003). However, it is also fair to remark that nor MUC4 polymorphism or any other candidate marker has yet been confirmed as the univocal causative gene for ETEC F4 susceptibility. Actually Rasschaert et al. (2007) showed that 30% of the genotypic resistant pigs are positive in the in vitro villous adhesion assay, suggesting that polymorphisms other than MUC4 could also been involved. Between those other candidate markers there has been described single nucleotide polymorphisms (SNPs) located on MUC4, MUC13, MUC20, the transferrin receptor (TFRC), tyrosine kinase non-receptor 2 (ACK1), or the UDP-GlcNAc:betaGalbeta-1,3-N-acetylglucosaminyltransferase 5 (B3GNT5) gene (reviewed by Luise et al., 2019a). Moreover, Goetstouwers et al. (2014) found a refined candidate region for F4ab/ac ETEC susceptibility situated proximal to MUC13 gene, however, it should be noted that this marker maps on a non-coding region. No protocols, in addition to an Illumina chip or a next-generation sequencing (NGS) technique, are available for genotyping the pigs for this marker, making nowadays difficult to routinely use for animal selection. To better dilucidated the role of MUC4 gen in this study, it had been interesting to determine the phenotypical presence of F4 receptors on brush borders, enterocytes, or villi of the small intestine by in vitro test (Van den Broeck et al., 1999). However, as characterization of the F4ab/ac ETEC susceptibility was not initially planned, unfortunately the receptor expression in susceptible pigs could not be confirmed.
Regarding the potential of the probiotic to fight the disease, the probiotic combination of B. infantis CECT 7210 and L. rhamnosus HN001 ameliorated the impairment in weight gain immediately after ETEC F4 inoculation (day 0–4 PI); the only challenged group that was not significantly different from the CTR+ group. Moreover, in the 4–8 PI period, the PRO group also showed improved weight gain, reaching levels similar to the CTR+ group. This enhanced response could be the result of a competitive exclusion of the pathogen by the probiotics. In fact, at day 4 PI this group showed an ETEC F4 colonic prevalence with values intermediate to those in the CTR+ and CTR− groups. In addition, there was a lower number of fecal enterobacteria and coliform plate counts compared to the CTR− group as well as a reduced number of attached bacteria to the ileal mucus – only the experimental diet group was not different from the CTR+ group. Other authors have reported the beneficial effects of these two probiotic strains against pathogenic agents, albeit when used separately. Barba-Vidal et al. (2017) showed, in a similar piglet model, that B. longum subsp. infantis CECT 7210 tends to reduce the percentage of animals with countable ileal coliforms and decreases fecal Salmonella excretion after an ETEC F4 or a Salmonella Typhimurium oral challenge, respectively. Moreover, this strain has antiviral activity when tested against rotavirus in mice (Muñoz et al., 2011) as well as antidiarrhoeal properties in healthy infants (Escribano et al., 2018). Similarly, L. rhamnosus HN001 is successful against pathogens such as ETEC and Staphylococcus aureus (Gopal et al., 2001; Inturri et al., 2016; Eggers et al., 2018). More recently, in previous studies of our group, we could also demonstrate the positive effects of this probiotic combination against a Salmonella challenge, leading to a faster clearance of the pathogen and recovery from the intestinal damage (Rodríguez-Sorrento et al., 2020). This putative reduction in the enteropathogen challenge promoted by the probiotic treatment could have also explained the intermediate level of mitosis observed between the CTR+ and CTR− groups at day 4 PI and the faster recovery of villus height at day 8 PI. These effects could also be associated with the reduced response observed in Pig-MAP at day 4 PI. This treatment showed values between 0.43 and 1.50 mg/ml, which are clearly below the 2 mg/ml considered to be normal in weanlings [Piñeiro et al., 2009; normal (≤1 mg/ml), borderline (1–2 mg/ml) and high levels (>2 mg/ml)]. Pig-MAP is commonly induced by interleukin 6 (IL-6; González-Ramón et al., 2000) that is simultaneously stimulated by nuclear factor kappa B (NF-κB) activation (Brasier, 2010). NF-κB is a protein complex, which controls the expression of genes implicated in inflammation process (Baker et al., 2011). Bifidobacterium infantis and L. rhamnosus can also modify the expression of IL-6 or NF-kB (Khailova et al., 2014; Gamallat et al., 2016; Ishizuka et al., 2016), an eventuality that is consistent with our results. A possible modulation of the immune response by this probiotic should also be considered. In this regard, previous works with B. infantis CECT 7210 have shown consistent increases in ileal IEL (13), and L. rhamnosus HN001 is attributed immune-modulating properties (Gill et al., 2001). However, we could not find such an effect in the present study when combined with L. rhamnosus HN001.
Galacto-oligosaccharides supplementation alone (the PRE group) in the piglets’ diet also promoted favorable outcomes. Like the probiotics alone, the PRE group presented improved weight gain for the 4–8 PI period, reaching similar values to the CTR+ group (and even numerically higher). Supplementing diets with the GOS alone affected neither enterobacteria nor coliform populations nor ileal histomorphometry, although, like the PRO group, the prevalence of colonic ETEC F4 was between the levels of the CTR+ and CTR− groups. At day 4 PI, the PRE group showed the lowest Pig-MAP level compared to the other challenged groups; only the PRE group was significantly different from the CTR− group and similar to the CTR+ group. In fact, GOS may alleviate inflammation, as shown in several studies (Vulevic et al., 2013; Verheijden et al., 2015). Wang et al. (2019) attributed the modulatory effects of inflammatory state to the ability of GOS to increase anti-inflammatory cytokine IL-10 while decreasing IL-8 by modulating, once again, NF-κB protein complex. Regarding fermentation, the prebiotic encouraged a decrease in colonic and ileal ammonia, data that suggest a possible shift toward a less proteolytic and beneficial microbiota due to its inclusion, as, for example, ammonia may buffer SCFA and block their activity (Davila et al., 2013; Shen et al., 2015).
The interest of mixing the tested probiotic strains with GOS is based on the reported ability of Bifidobacteria and Lactobacillus to degrade and use GOS as energy source. β-glycosidic linkages connecting saccharides that comprise GOS are hydrolyzed in the colon by these two genera of bacteria that bear β-galactosidases (Andersen et al., 2012; Garrido et al., 2013) and, specifically, B. infantis CECT 7210 prefers GOS as a growth substrate rather than other prebiotics (Ruiz et al., 2020). Several works have reported a rise in Bifidobacteria and Lactobacillus when tested together with different types of GOS, whether or not it is very pure (Hong et al., 2016; Monteagudo-Mera et al., 2016; Kittibunchakul et al., 2018). However, GOS utilization varies depending on the bacterial strain and the oligosaccharide composition (Thongaram et al., 2017). Hence, considering the previously mentioned characteristics, administering these probiotics with GOS potentially would lead to synergistic activity enhancing the benefits that each component produces on its own, however, in the present study, we were not able to demonstrate any additive o synergic effect. In the literature, works evaluating the synbiotic strategy are limited, and the results have not always been consistent. For example, Tanner et al. (2014) and Abrahamse-Berkeveld et al. (2016) observed positive outcomes, including enhanced growth and pathogen inhibition, when mixing a bifidobacteria strain with GOS in both in vitro and in vivo studies. By contrast, Krumbeck et al. (2018) did not find any synergy in the capacity of improvement intestinal barrier function when combining bifidobacteria and GOS in humans. Previous studies of our group also failed to demonstrate the synergic activities of combination of the strains CECT 7210 and HN001 with FOS in Samonella orally challenged piglets (Rodríguez-Sorrento et al., 2020). In the present study, we observed some synergy before the challenge, particularly in the number of enterobacteria and coliforms, which were lower compared to the rest of supplemented groups. This effect could suggest a positive impact of the synbiotic treatment on the resident microbiota, specifically promoting the growth of specific microorganisms, which together with the probiotics, would have displaced enterobacteria. These results are particularly relevant since the 1st week after weaning is one of the most critical periods in the pig’s life, in which they have to cope with numerous stressors and dysbiosis is commonly present (Gresse et al., 2017; Motta et al., 2019). Shifts of microbiota toward bifidobacteria via the administration of a synbiotic that contains GOS [±oligofructose (OF)] and strains of Bifidobacterium have been reported in trials with healthy newborn babies and infants (Simeoni et al., 2016; Chua et al., 2017). In particular, the latter authors attributed this outcome to the increase of endogenous bifidobacteria and also in piglets by OF (Trevisi et al., 2008; Modesto et al., 2009). However, the outcomes in the SYN group were different after the challenge. During the acute period of the infection (days 0–4 PI), the SYN group showed a larger decrease in feed intake compared to the other treatments, with similar values to challenged pigs that were not supplemented. This feed intake depression was even more pronounced in the 4–8 PI period. According to this reduced feed intake, SYN and CTR− were the only two groups that lost weight in the 0–4 PI period, and the SYN group showed a trend for lower gains compared to the CTR− group in the 4–8 PI period. Together with reduced intake, impaired nutrient utilization associated with diarrhea could also explain weight loss (Clements et al., 2012). Although we were unable to find differences in the fecal score due to the dietary supplementation, the highest fecal scores on day 1 PI occurred in the SYN group, with values that were significantly higher than those found in the PRO group. This trend for a more acute peak of diarrhea in the SYN group is supported by the higher prevalence of ETEC F4 found in the colon on day 8 PI compared to the CTR+ group. This outcome was particularly evident in the groups of animals with the MUC4 susceptible genotype, consistent with previous findings (Luise et al., 2019b). Moreover, the concentration of inflammatory serum markers was higher in the SYN group. On day 4 PI, the TNF-α value was higher in the SYN group compared to the CTR− group, although it was not different from the PRO or CTR+ groups. Regarding Pig-MAP, whereas on day 4 PI the SYN group showed a level between the CTR+ and CTR− groups, at the end of the trial (day 8 PI), the SYN group maintained markedly high values, in contrast to the rest of the challenged groups that were able to normalize Pig-MAP levels. A higher energy expenditure associated with an inflammatory response could have also contributed to explain the lower performance of the SYN animals.
Based on the above findings, it is evident that the synbiotic combination did not show any additive or beneficial effect on the challenged animals but could even have provided a better opportunity for ETEC F4+ to thrive. Whereas before the inoculation the SYN treatment showed a positive outcome with reductions in the number of enterobacteria and coliforms, after the ETEC challenge the combination of the probiotic and prebiotic could have reduced the ability of the animal to fight the pathogen. An explanation for that phenomenon could be the complex interactions between members of the intestinal microbiota that could have been potentially disturbed by the synbiotic. It is known that a well-established and developed microbiota is characterized by an equilibrium between different microbial groups through cross-feeding, competitive exclusion, and quorum sensing mechanisms that provide to the microbial community with a high resilience against external inputs (Walter et al., 2018). The administration of SYN could have determined changes in the sequence of gut colonization of the young pig during the 1st week post-weaning leading to a microbial ecosystem more susceptible to be colonized by opportunistic pathogens like ETEC F4. The fermentation of GOS by the probiotic along the small intestine, could have increased the amount of probiotic reaching the hindgut and also have reduced the amount of GOS arriving to the colon as intact prebiotic. This hypothesis could explain why the ammonia concentration was only decreased in colon at day 4 PI in the PRE but not in the SYN group as less GOS would have arrived to the colon. Moreover, the reduction observed in enterobacteria and coliforms with SYN, although generally regarded as positive, could have been a consequence of a reduced biodiversity and resilience of the ecosystem. Not necessarily a reduction in number of enterobacteria in the microbial community is always positive. Previous authors have described how commensal Enterobacteriaceae can protect neonates against Salmonella colonization through oxygen competition (Litvak and Bäumler, 2019) as well as producing bacteriocins able to eliminate close-related competitors (Stecher, 2015). Seems therefore plausible to hypothesize that the biodiversity of the intestinal community could have been reduced by the synbiotic treatment, making the ecosystem more susceptible to dysbiosis with more niches becoming available for opportunistic/pathogenic bacteria.
Interestingly, the impact of the synbiotic treatment on microbial metabolism seemed to be MUC4 genotype dependent. The SCFA concentration was increased, and also the molar ratio of valeric acid at day 8 PI, but only in pigs with the MUC4 susceptible genotype. A more acute course of diarrhea, determined by the susceptible genotype, could have promoted a differential effect on microbial activity, suggesting that the effect of the SYN was somehow challenge dependent. In support of this hypothesis, it is known that the MUC4 genotype can influence the small intestinal glycomic patterns (Luise et al., 2019b) and the intestinal microbiota (Massacci et al., 2020), facts that could explain the differential response of the susceptible animals to the SYN treatment. The increase observed in the molar proportion of valeric acid with SYN supplementation could be attributed to favored valeric-producing bacteria. In this vain, Van Nevel et al. (2005) observed that the inclusion of galactomannans from locust bean gum increases valeric acid and decreases the mitotic index in crypts, similarly to the present study, where we also found a lower mitosis index and shorter crypts in the susceptible animals of the SYN group.
It is also fair to remark that results from challenge studies like this one are always conditioned by the experimental model of disease and need to be approached with caution. An oral challenge with a single high dose of ETEC F4 is undoubtedly far from what would naturally occur, namely repeated lower doses. Factors, as the inoculated bacterial strains, doses, timing of the inoculum, post-weaning age, and immunity status of the animals, undoubtedly are variables that could determine differential responses (Luise et al., 2019a). One of the factors that could deserve some discussion is timing for inoculation. We inoculated animals 7 days post-weaning (around 28 days of age) aiming to test the potential prophylactic effect of the symbiotic. Inoculating animals in shorter times (i.e., 1–2 days post-weaning) could had better resembled the situation in the farm, tackling the time of the highest risk for intestinal dysbiosis and colonization by ETEC. However, challenging animals only 1–2 days after weaning had limited the possibilities for the symbiotic to exert a prophylactic effect. Considering this, we decided to inoculate animals after 7 days post-weaning, although undoubtedly this could had determined differences with what is found in the field.
Regarding the possible role of the MUC4 gene on results of this trial, it is known that ETEC attaches to the jejunal brush border through the binding of its fimbriae to many putative receptors. These receptors are carbohydrates of glycoproteins in the intestinal epithelial cells and intestinal mucus, which have been shown to differ among pigs (Van den Broeck et al., 2000; Rasschaert et al., 2007). Different antigenic variants of F4 fimbriae have been identified (Bakker et al., 1991), including the F4ab and F4ac variants, in the strain used in this study. MUC4 codes for a membrane-bound O-glycoprotein in the mucus layer. A mutation in this gene has been proposed as a useful genetic marker to identify susceptible genotypes (Jørgensen et al., 2003; Luise et al., 2019a). In our study, as expected, MUC4 susceptible piglets showed a worsened fecal consistency, as well as a lower weight daily gain and higher numbers of enterobacteria, coliforms, and ETEC F4 in colon. Other authors have also described similar associations in challenge and non-challenge conditions (Casini et al., 2003; Trevisi et al., 2015; Luise et al., 2019b; Sterndale et al., 2019); these data support the usefulness of this candidate gene. Nevertheless, not all the evaluated parameters were modified by MUC4 status. Specifically, neither the number of enterobacteria and coliforms attached to the ileal epithelium nor ileal histomorphology were altered. This outcome could be due to other genes and receptors involved in the susceptibility to ETEC F4 infection (Nguyen et al., 2012). In this regard, Rasschaert et al. (2007) confirmed that the in vitro adhesion of F4ac and F4ab ETEC to the villous brush borders is not always associated with the MUC4 gene, suggesting the existence of at least one other receptor for F4ab/ac fimbria. Indeed, other genes like MUC13 or TNRC (transferrin receptor gene) have also been associated with ETEC F4 susceptibility (Zhang et al., 2008; Jacobsen et al., 2010; Ren et al., 2012).
In our experiment, we did not select animals based on MUC4 susceptibility, but we determined this characteristic at the end of the trial. Although many studies with ETEC challenges do not apply any control a priori or a posteriori on the genetic susceptibility of the animals, we found that this could be of interest to better understand the variability of the data and to elucidate the role of this candidate gene in the piglet response in front of an ETEC F4 challenge. The MUC4 analysis showed certain unbalanced distribution of susceptible animals between treatments. This could have had an impact on the results, particularly on those parameters that were influenced by MUC4 polymorphism (performance, fecal consistency, numbers of fecal coliforms, and colonic ETEC F4). When the whole group of animals are considered, the ratio between resistant and susceptible animals varied between 0.7 for SYN to 2.3 for CTR+ (see Table 2). Although it was a reasonable difference, it could partially explain the lower ADFI and ADG observed for the SYN treatment. When contemplating only the sampled animals after euthanasia, the distribution of resistant and susceptible animals was more unbalanced between treatments causing a different scenario between treatments at days 4 and 8 PI. Whereas at day 4 PI, PRO treatment presented a favorable ratio of 5 and SYN of 0.7, at day 8 PI PRO showed a ratio of 0.5 and SYN of 2. Nonetheless, SYN treatment always exhibited higher ETEC F4 counts than PRO, evidencing that, despite the uneven distribution, SYN treatment consistently brought about the same effects both sampling days. Also, it is interesting to remark that in defiance of the association between the MUC4 polymorphism and fecal counts of coliforms, particularly relevant at day 4 PI (7.43 vs. 6.27 log CFU/g for MUC4+ and MUC4−, respectively; p = 0.027), the SYN treatment was always associated with lower plate counts compared to CTR−, regardless the dissimilar distribution of resistant and susceptible animals (5.90 vs. 8.48 log CFU/g, p < 0.01 at day 4 PI and 5.71 vs. 6.62; p < 0.001 at day 8 PI). Altogether, it appears that, although the unbalanced distribution of MUC4 polymorphism could have increased residual variability and have had an impact on the magnitude of the response, the direction and meaning of the observed differences would be consistent.
In conclusion, the multi-strain probiotic B. longum subsp. infantis CECT 7210 and L. rhamnosus HN001 reduced growth impairment after oral ETEC F4 challenge. Furthermore, this supplementation seems to provide competitive exclusion for ETEC F4, because there were fewer colonic enterobacteria and coliforms in the gut and a trend to diminish the pathogen. This outcome could explain the lower Pig-MAP levels and improved villus height found after 1 week of the challenge with this treatment. The supplementation of the diets with galacto-oligosaccharides also diminished the growth impairment induced by the challenge and is associated with lower levels of plasmatic Pig-MAP. This phenomenon suggests a modulation of the inflammatory response by this prebiotic. These beneficial effects were not synergistic when the probiotics and the prebiotic were administered together. The synbiotic treatment had a differential impact on colonic fermentation depending on MUC4 susceptibility. The differential response in animals with more acute diarrhea suggests that the absence of synergistic effect could be related to the limitations of experimental model that rely on a single exposure to high doses of the pathogen rather than the low-dose exposure that would occur naturally. More research should be performed in this field to understand the complex interactions produced in the gastrointestinal tract, with an emphasis on the microbiota establishment at early ages.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by Animal and Human Experimental Ethical Committee of the Autonomous University of Barcelona (Permit No. CEEAH: 4026 DMAH: 10118).
Author Contributions
AR-S participated in the experimental design and was responsible for the animal trial, laboratory analysis, data analysis, and writing. LC participated in the experimental design, animal trials, data analysis, and writing. PL-C participated in animal trials and data analysis. GC-O, JM-M, and MR-P participated in the experimental design and contributed to data analysis and writing. DL and PT contributed to laboratory analysis, data analysis, and writing. SM-O participated in the experimental design, animal trials, laboratory analysis, data analysis, and writing. All authors contributed to the article and approved the submitted version.
Funding
This project has been supported by the CDTI (Ministerio de Ciencia e Innovación de España) and the Fondo Europeo de Desarrollo Regional (FEDER; Acronym: SMARTFOODS, project number IDI-20141206).
Conflict of Interest
GC-O, JM-M, and MR-P were employed by the company Laboratorios Ordesa S.L.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abrahamse-Berkeveld, M., Alles, M., Franke-Beckmann, E., Helm, K., Knecht, R., Köllges, R., et al. (2016). Infant formula containing galacto-and fructo-oligosaccharides and Bifidobacterium breve M-16V supports adequate growth and tolerance in healthy infants in a randomised, controlled, double-blind, prospective, multicentre study. J. Nutr. Sci. 5:e42. doi: 10.1017/jns.2016.35
Alizadeh, A., Akbari, P., Difilippo, E., Schols, H. A., Ulfman, L. H., Schoterman, M. H., et al. (2016). The piglet as a model for studying dietary components in infant diets: effects of galacto-oligosaccharides on intestinal functions. Br. J. Nutr. 115, 605–618. doi: 10.1017/S0007114515004997
Andersen, J. M., Barrangou, R., Hachem, M. A., Lahtinen, S. J., Goh, Y. J., Svensson, B., et al. (2012). Transcriptional analysis of prebiotic uptake and catabolism by Lactobacillus acidophilus NCFM. PLoS One 7:e44409. doi: 10.1371/journal.pone.0044409
AOAC International (1995). Official methods of analysis of AOAC international. Washington, DC: AOAC.
Baker, R. G., Hayden, M. S., and Ghosh, S. (2011). NF-κB, inflammation, and metabolic disease. Cell Metab. 13, 11–22. doi: 10.1016/j.cmet.2010.12.008
Bakker, D., Willemsen, P. T., Willems, R. H., Huisman, T. T., Mooi, F. R., Oudega, B., et al. (1991). Identification of minor fimbrial subunits involved in biosynthesis of K88 fimbriae. J. Bacteriol. 174, 6350–6358. doi: 10.1128/jb.174.20.6350-6358.1992
Barba-Vidal, E., Castillejos, L., López-Colom, P., Rivero Urgell, M., Moreno Muñoz, J. A., and Martín-Orúe, S. M. (2017). Evaluation of the probiotic strain Bifidobacterium longum subsp. infantis CECT 7210 capacities to improve health status and fight digestive pathogens in a piglet model. Front. Microbiol. 8:533. doi: 10.3389/fmicb.2017.00533
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2014). Package Lme4: linear mixed-effects models using Eigen and S4. R package version. 67.
Brasier, A. R. (2010). The nuclear factor-κB–interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 86, 211–218. doi: 10.1093/cvr/cvq076
Canfora, E. E., van der Beek, C. M., Hermes, G. D. A., Goossens, G. H., Jocken, J. W. E., Holst, J. J., et al. (2017). Supplementation of diet with galacto-oligosaccharides increases bifidobacteria, but not insulin sensitivity, in obese prediabetic individuals. Gastroenterology 153, 87–97.e3. doi: 10.1053/j.gastro.2017.03.051
Casini, L., Bosi, P., Gremokolini, C., Trevisi, P., Mazzoni, M., and Bonilauri, P. (2003). Oral challenge with E.coli K88 as a tool to assess growth and health performance in feeding trials of-weaned pigs. Ital. J. Anim. Sci. 2(Suppl. 1), 358–360. doi: 10.4081/ijas.2003.11676010
Chua, M. C., Ben-Amor, K., Lay, C., Neo, A. G. E., Chiang, W. C., Rao, R., et al. (2017). Effect of synbiotic on the gut microbiota of cesarean delivered infants: a randomized, double-blind, multicenter study. J. Pediatr. Gastroenterol. Nutr. 65, 102–106. doi: 10.1097/MPG.0000000000001623
Clements, A., Young, J. C., Constantinou, N., and Frankel, G. (2012). Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes 3, 71–87. doi: 10.4161/gmic.19182
Davila, A. M., Blachier, F., Gotteland, M., Andriamihaja, M., Benetti, P. H., Sanz, Y., et al. (2013). Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol. Res. 68, 95–107. doi: 10.1016/j.phrs.2012.11.005
de Vrese, M., and Marteau, P. R. (2007). Probiotics and prebiotics: effects on diarrhea. J. Nutr. 137(3 Suppl. 2), 803S–811S. doi: 10.1093/jn/137.3.803S
Eggers, S., Barker, A. K., Valentine, S., Hess, T., Duster, M., and Safdar, N. (2018). Effect of Lactobacillus rhamnosus HN001 on carriage of Staphylococcus aureus: results of the impact of probiotics for reducing infections in veterans (IMPROVE) study. BMC Infect. Dis. 18:129. doi: 10.1186/s12879-018-3028-6
Escribano, J., Ferré, N., Gispert-Llaurado, M., Luque, V., Rubio-Torrents, C., Zaragoza-Jordana, M., et al. (2018). Bifidobacterium longum subsp infantis CECT7210-supplemented formula reduces diarrhoea in healthy infants: a randomized controlled trial. Pediatr. Res. 83, 1120–1128. doi: 10.1038/pr.2018.34
Fijan, S., Šulc, D., and Steyer, A. (2018). Study of the in vitro antagonistic activity of various single-strain and multi-strain probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 15:E1539. doi: 10.3390/ijerph15071539
Gamallat, Y., Meyiah, A., Kuugbee, E. D., Hago, A. M., Chiwala, G., Awadasseid, A., et al. (2016). Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 83, 536–541. doi: 10.1016/j.biopha.2016.07.001
Garrido, D., Ruiz-Moyano, S., Jimenez-Espinoza, R., Eom, H. J., Block, D. E., and Mills, D. A. (2013). Utilization of galactooligosaccharides by Bifidobacterium longum subsp. infantis isolates. Food Microbiol. 33, 262–270. doi: 10.1016/j.fm.2012.10.003
Gill, H. S., Shu, Q., Lin, H., Rutherfurd, K. J., and Cross, M. L. (2001). Protection against translocating Salmonella typhimurium infection in mice by feeding the immuno-enhancing probiotic Lactobacillus rhamnosus strain HN001. Med. Microbiol. Immunol. 190, 97–104. doi: 10.1007/s004300100095
Goetstouwers, T., Van Poucke, M., Coppieters, W., Nguyen, V. U., Melkebeek, V., Coddens, A., et al. (2014). Refined candidate region for F4ab/ac enterotoxigenic Escherichia coli susceptibility situated proximal to MUC13 in pigs. PLoS One 9:e105013. doi: 10.1371/journal.pone.0105013
González-Ramón, N., Hoebe, K., Alava, M. A., Van Leengoed, L., Piñeiro, M., Carmona, S., et al. (2000). Pig MAP/ITIH4 and haptoglobin are interleukin-6-dependent acute-phase plasma proteins in porcine primary cultured hepatocytes. Eur. J. Biochem. 267, 1878–1885. doi: 10.1046/j.1432-1327.2000.01195.x
Gopal, P. K., Prasad, J., Smart, J., and Gill, H. S. (2001). In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int. J. Food Microbiol. 67, 207–216. doi: 10.1016/S0168-1605(01)00440-8
Gresse, R., Chaucheyras-Durand, F., Fleury, M. A., Van de Wiele, T., Forano, E., and Blanquet-Diot, S. (2017). Gut microbiota dysbiosis in postweaning piglets: understanding the keys to health. Trends Microbiol. 25, 851–873. doi: 10.1016/j.tim.2017.05.004
Grimaldi, R., Swann, J. R., Vulevic, J., Gibson, G. R., and Costabile, A. (2016). Fermentation properties and potential prebiotic activity of Bimuno® galacto-oligosaccharide (65% galacto-oligosaccharide content) on in vitro gut microbiota parameters. Br. J. Nutr. 116, 480–486. doi: 10.1017/S0007114516002269
Hermes, R. G., Molist, F., Pérez, J. F., Gómez de Segura, A., Ywazaki, M., Davin, R., et al. (2013). Casein glycomacropeptide in the diet may reduce Escherichia coli attachment to the intestinal mucosa and increase the intestinal lactobacilli of early weaned piglets after an enterotoxigenic E. coli K88 challenge. Br. J. Nutr. 109, 1001–1012. doi: 10.1017/S0007114512002978
Hermes, R. G., Molist, F., Ywazaki, M., Nofrarías, M., Gomez de Segura, A., Gasa, J., et al. (2009). Effect of dietary level of protein and fiber on the productive performance and health status of piglets. J. Anim. Sci. 87, 3569–3577. doi: 10.2527/jas.2008-1241
Hong, K. B., Kim, J. H., Kwon, H. K., Han, S. H., Park, Y., and Suh, H. J. (2016). Evaluation of prebiotic effects of high-purity galactooligosaccharides in vitro and in vivo. Food Technol. Biotechnol. 54, 156–163. doi: 10.17113/ftb.54.02.16.4292
Inturri, R., Stivala, A., Furneri, P. M., and Blandino, G. (2016). Growth and adhesion to HT-29 cells inhibition of gram-negatives by Bifidobacterium longum BB536 e Lactobacillus rhamnosus HN001 alone and in combination. Eur. Rev. Med. Pharmacol. Sci. 20, 4943–4949.
Ishizuka, T., Kanmani, P., Kobayashi, H., Miyazaki, A., Soma, J., Suda, Y., et al. (2016). Immunobiotic bifidobacteria strains modulate rotavirus immune response in porcine intestinal epitheliocytes via pattern recognition receptor signaling. PLoS One 11:e0152416. doi: 10.1371/journal.pone.0152416
Jacobsen, M., Kracht, S. S., Esteso, G., Cirera, S., Edfors, I., Archibald, A. L., et al. (2010). Refined candidate region specified by haplotype sharing for Escherichia coli F4ab/F4ac susceptibility alleles in pigs. Anim. Genet. 41, 21–25. doi: 10.1111/j.1365-2052.2009.01952.x
Jensen, M. T., Cox, R. P., and Jensen, B. B. (1995). Microbial production of skatole in the hind gut of pigs given different diets and its relation to skatole deposition in backfat. Anim. Sci. 61, 293–304. doi: 10.1017/S1357729800013837
Jørgensen, C. B., Cirera, S., Anderson, S. I., Archibald, A. L., Raudsepp, T., Chowdhary, B., et al. (2003). Linkage and comparative mapping of the locus controlling susceptibility towards E. coli F4ab/ac diarrhoea in pigs. Cytogenet. Genome Res. 102, 157–162. doi: 10.1159/000075742
Kechagia, M., Basoulis, D., Konstantopoulou, S., Dimitriadi, D., Gyftopoulou, K., Skarmoutsou, N., et al. (2013). Health benefits of probiotics: a review. ISRN Nutr. 2013:481651. doi: 10.5402/2013/481651
Khailova, L., Petrie, B., Baird, C. H., Dominguez Rieg, J. A., and Wischmeyer, P. E. (2014). Lactobacillus rhamnosus GG and Bifidobacterium longum attenuate lung injury and inflammatory response in experimental sepsis. PLoS One 9:e97861. doi: 10.1371/journal.pone.0097861
Kittibunchakul, S., Maischberger, T., Domig, K. J., Kneifel, W., Nguyen, H. M., Haltrich, D., et al. (2018). Fermentability of a novel galacto-oligosaccharide mixture by Lactobacillus spp. and Bifidobacterium spp. Molecules 23:E3352. doi: 10.3390/molecules23123352
Krumbeck, J. A., Rasmussen, H. E., Hutkins, R. W., Clarke, J., Shawron, K., Keshavarzian, A., et al. (2018). Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 6:121. doi: 10.1186/s40168-018-0494-4
Kumar, M., Dhaka, P., Vijay, D., Vergis, J., Mohan, V., Kumar, A., et al. (2016). Antimicrobial effects of Lactobacillus plantarum and Lactobacillus acidophilus against multidrug-resistant enteroaggregative Escherichia coli. Int. J. Antimicrob. Agents 48, 265–270. doi: 10.1016/j.ijantimicag.2016.05.014
Kunz, C., Meyer, C., Collado, M. C., Geiger, L., García-Mantrana, I., Bertua-Ríos, B., et al. (2017). Influence of gestational age, secretor, and Lewis blood group status on the oligosaccharide content of human Milk. J. Pediatr. Gastroenterol. Nutr. 64, 789–798. doi: 10.1097/MPG.0000000000001402
Lanata, C. F., Fischer-Walker, C. L., Olascoaga, A. C., Torres, C. X., Aryee, M. J., Black, R. E., et al. (2013). Global causes of diarrheal disease mortality in children <5 years of age: a systematic review. PLoS One 8:e72788. doi: 10.1371/journal.pone.0072788
Litvak, Y., and Bäumler, A. J. (2019). The founder hypothesis: A basis for microbiota resistance, diversity in taxa carriage, and colonization resistance against pathogens. PLoS Pathog. 15:e1007563. doi: 10.1371/journal.ppat.1007563
Liu, L., Oza, S., Hogan, D., Perin, J., Rudan, I., Lawn, J. E., et al. (2015). Global, regional, and national causes of child mortality in 2000-13, with projections to inform post-2015 priorities: an updated systematic analysis. Lancet 385, 430–440. doi: 10.1016/S0140-6736(14)61698-6
Luise, D., Lauridsen, C., Bosi, B., and Trevisi, P. (2019a). Methodology and application of Escherichia coli F4 and F18 encoding infection models in post-weaning pigs. J. Anim. Sci. Biotechnol. 10:53. doi: 10.1186/s40104-019-0352-7
Luise, D., Motta, V., Bertocchi, M., Salvarani, C., Clavenzani, P., Fanelli, F., et al. (2019b). Effect of Mucine 4 and Fucosyltransferase 1 genetic variants on gut homoeostasis of growing healthy pigs. J. Anim. Physiol. Anim. Nutr. 103, 801–812. doi: 10.1111/jpn.13063
Luppi, A. (2017). Swine enteric colibacillosis: diagnosis, therapy and antimicrobial resistance. Porcine Health Manag. 3:16. doi: 10.1186/s40813-017-0063-4
Massacci, F. R., Tofani, S., Forte, C., Bertocchi, M., Lovito, C., Orsini, S., et al. (2020). Host genotype and amoxicillin administration affect the incidence of diarrhoea and faecal microbiota of weaned piglets during a natural multiresistant ETEC infection. J. Anim. Breed. Genet. 137, 60–72. doi: 10.1111/jbg.12432
Modesto, M., D'Aimmo, M., Stefanini, I., Trevisi, P., Filippi, S., Casini, L., et al. (2009). A novel strategy to select Bifidobacterium strains and prebiotics as natural growth promoters in newly weaned pigs. Livest. Sci. 122, 248–258. doi: 10.1016/j.livsci.2008.08.017
Monteagudo-Mera, A., Arthur, J. C., Jobin, C., Keku, T., Bruno-Barcena, J. M., and Azcarate-Peril, M. (2016). High purity galacto-oligosaccharides enhance specific Bifidobacterium species and their metabolic activity in the mouse gut microbiome. Benefic. Microbes 7, 247–264. doi: 10.3920/BM2015.0114
Motta, V., Luise, D., Bosi, P., and Trevisi, P. (2019). Faecal microbiota shift during weaning transition in piglets and evaluation of AO blood types as shaping factor for the bacterial community profile. PLoS One 14:e0217001. doi: 10.1371/journal.pone.0217001
Muñoz, J. A., Chenoll, E., Casinos, B., Bataller, E., Ramón, D., Genovés, S., et al. (2011). Novel probiotic Bifidobacterium longum subsp. infantis CECT 7210 strain active against rotavirus infections. Appl. Environ. Microbiol. 77, 8775–8783. doi: 10.1128/AEM.05548-11
National Research Council (NRC) (2012). Nutrient requirements of swine. Washington, DC: National Academy Press.
Nguyen, V. U., Goetstouwers, T., Coddens, A., Van Poucke, M., Peelman, L., Deforce, D., et al. (2012). Differentiation of F4 receptor profiles in pigs based on their mucin 4 polymorphism, responsiveness to oral F4 immunization and in vitro binding of F4 to villi. Vet. Immunol. Immunopathol. 152, 93–100. doi: 10.1016/j.vetimm.2012.09.015
Nofrarías, M., Manzanilla, E. G., Pujols, J., Gibert, X., Majó, N., Segalés, J., et al. (2006). Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J. Anim. Sci. 84, 2735–2742. doi: 10.2527/jas.2005-414
Paganini, D., Uyoga, M. A., Kortman, G. A. M., Cercamondi, C. I., Moretti, D., Barth-Jaeggi, T., et al. (2017). Prebiotic galacto-oligosaccharides mitigate the adverse effects of iron fortification on the gut microbiome: a randomised controlled study in Kenyan infants. Gut 66, 1956–1967. doi: 10.1136/gutjnl-2017-314418
Peng, Q. L., Ren, J., Yan, X. M., Huang, X., Tang, H., Wang, Y. Z., et al. (2007). The g.243A>G mutation in intron 17 of MUC4 is significantly associated with susceptibility/resistance to ETEC F4ab/ac infection in pigs. Anim. Genet. 38, 397–400. doi: 10.1111/j.1365-2052.2007.01608.x
Piñeiro, C., Piñeiro, M., Morales, J., Andrés, M., Lorenzo, E., Pozo, M. D., et al. (2009). Pig-MAP and haptoglobin concentration reference values in swine from commercial farms. Vet. J. 179, 78–84. doi: 10.1016/j.tvjl.2007.08.010
Poirel, L., Madec, J. Y., Lupo, A., Schink, A. K., Kieffer, N., Nordmann, P., et al. (2018). Antimicrobial resistance in Escherichia coli. Microbiol. Spectr. 6. doi: 10.1128/microbiolspec.ARBA-0026-2017
Rasschaert, K., Verdonck, F., Goddeeris, B. M., Duchateau, L., and Cox, E. (2007). Screening of pigs resistant to F4 enterotoxigenic Escherichia coli (ETEC) infection. Vet. Microbiol. 123, 249–253. doi: 10.1016/j.vetmic.2007.02.017
Ren, J., Yan, X., Ai, H., Zhang, Z., Huang, X., Ouyang, J., et al. (2012). Susceptibility towards enterotoxigenic Escherichia coli F4ac diarrhea is governed by the MUC13 gene in pigs. PLoS One 7:e44573. doi: 10.1371/journal.pone.0044573
Richardson, A. J., Calder, A. G., Stewart, C. S., and Smith, A. (1989). Simultaneous determination of volatile and non-volatile acidic fermentation products of anaerobes by capillary gas chromatography. Lett. Appl. Microbiol. 9, 5–8. doi: 10.1111/j.1472-65X.1989.tb00278.x
Rodríguez-Sorrento, A., Castillejos, L., López-Colom, P., Cifuentes-Orjuela, G., Rodríguez-Palmero, M., Moreno-Muñoz, J. A., et al. (2020). Effects of Bifidobacterium longum Subsp. infantis CECT 7210 and lactobacillus rhamnosus HN001, combined or not With Oligofructose-enriched inulin, on weaned pigs orally challenged With Salmonella Typhimurium. Front. Microbiol. 11:2012. doi: 10.3389/fmicb.2020.02012
Romond, M. B., Haddou, Z., Mielcareck, C., and Romond, C. (1997). Bifidobacteria and human health: regulatory effect of indigenous bifidobacteria on Escherichia coli intestinal colonization. Anaerobe 3, 131–136. doi: 10.1006/anae.1997.0089
Ruiz, L., Flórez, A. B., Sánchez, B., Moreno-Muñoz, J. A., Rodriguez-Palmero, M., Jiménez, J., et al. (2020). Bifidobacteriumlongum subsp. infantis CECT7210 (B. infantis IM-1®) displays In vitro activity against Some intestinal pathogens. Nutrients 12:3259. doi: 10.3390/nu12113259
Sela, D. A., Chapman, J., Adeuya, A., Kim, J. H., Chen, F., Whitehead, T. R., et al. (2008). The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc. Natl. Acad. Sci. U. S. A. 105, 18964–18969. doi: 10.1073/pnas.0809584105
Shen, T. C., Albenberg, L., Bittinger, K., Chehoud, C., Chen, Y. Y., Judge, C. A., et al. (2015). Engineering the gut microbiota to treat hyperammonemia. J. Clin. Invest. 125, 2841–2850. doi: 10.1172/JCI79214
Shoaf, K., Mulvey, G. L., Armstrong, G. D., and Hutkins, R. W. (2006). Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect. Immun. 74, 6920–6928. doi: 10.1128/IAI.01030-06
Shu, Q., and Gill, H. S. (2001). A dietary probiotic (Bifidobacterium lactis HN019) reduces the severity of Escherichia coli O157:H7 infection in mice. Med. Microbiol. Immunol. 189, 147–152. doi: 10.1007/s430-001-8021-9
Simeoni, U., Berger, B., Junick, J., Blaut, M., Pecquet, S., Rezzonico, E., et al. (2016). Gut microbiota analysis reveals a marked shift to bifidobacteria by a starter infant formula containing a synbiotic of bovine milk-derived oligosaccharides and Bifidobacterium animalis subsp. lactis CNCM I-3446. Environ. Microbiol. 18, 2185–2195. doi: 10.1111/1462-2920.13144
Song, H., Zhang, J., Qu, J., Liu, J., Yin, P., Zhang, G., et al. (2019). Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 41, 1007–1014. doi: 10.1007/s10529-019-02694-2
Stecher, B. (2015). The roles of inflammation, nutrient availability and the commensal microbiota in enteric pathogen infection. Microbiol. Spectr. 3. doi: 10.1128/microbiolspec.MBP-0008-2014
Sterndale, S. O., Evans, D. J., Mansfield, J. P., Clarke, J., Sahibzada, S., Abraham, S., et al. (2019). Effect of mucin 4 allele on susceptibility to experimental infection with enterotoxigenic F4 Escherichia coli in pigs fed experimental diets. J. Anim. Sci. Biotechnol. 10:56. doi: 10.1186/s40104-019-0366-1
Tanner, S. A., Chassard, C., Zihler Berner, A., and Lacroix, C. (2014). Synergistic effects of Bifidobacterium thermophilum RBL67 and selected prebiotics on inhibition of Salmonella colonization in the swine proximal colon PolyFermS model. Gut Pathog. 6:44. doi: 10.1186/s13099-014-0044-y
Thongaram, T., Hoeflinger, J. L., Chow, J., and Miller, M. J. (2017). Prebiotic galactooligosaccharide metabolism by probiotic lactobacilli and bifidobacteria. J. Agric. Food Chem. 65, 4184–4192. doi: 10.1021/acs.jafc.7b00851
Tian, S., Wang, J., Yu, H., Wang, J., and Zhu, W. (2019). Changes in Ileal microbial composition and microbial metabolism by an early-life Galacto-oligosaccharides intervention in a neonatal porcine model. Nutrients 11:1753. doi: 10.3390/nu11081753
Toscano, M., De Grandi, R., Stronati, L., De Vecchi, E., and Drago, L. (2017). Effect of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536 on the healthy gut microbiota composition at phyla and species level: A preliminary study. World J. Gastroenterol. 23, 2696–2704. doi: 10.3748/wjg.v23.i15.2696
Trevisi, P., Corrent, E., Mazzoni, M., Messori, S., Priori, D., Gherpelli, Y., et al. (2015). Effect of added dietary threonine on growth performance, health, immunity and gastrointestinal function of weaning pigs with differing genetic susceptibility to Escherichia coli infection and challenged with E. coli K88ac. J. Anim. Physiol. Anim. Nutr. 99, 511–520. doi: 10.1111/jpn.12216
Trevisi, P., De Filippi, S., Minieri, L., Mazzoni, M., Modesto, M., Biavati, B., et al. (2008). Effect of fructo-oligosaccharides and different doses of Bifidobacterium animalis in a weaning diet on bacterial translocation and toll-like receptor gene expression in pigs. Nutrition 24, 1023–1029. doi: 10.1016/j.nut.2008.04.008
Trevisi, P., Melchior, D., Mazzoni, M., Casini, L., De Filippi, S., Minieri, L., et al. (2009). A tryptophan-enriched diet improves feed intake and growth performance of susceptible weanling pigs orally challenged with E. coli K88. J. Anim. Sci. 87, 148–156. doi: 10.2527/jas.2007-0732
Tzortzis, G. (2009). Development and functional properties of Bimuno: a second-generation prebiotic mixture. Food Sci. Technol. Bull. 6, 81–89. doi: 10.1616/1476-2137.15818
Van den Broeck, W., Cox, E., and Goddeeris, B. M. (1999). Receptor-specific binding of purified F4 to isolated villi. Vet. Microbiol. 68, 255–263. doi: 10.1016/S0378-1135(99)00076-0
Van den Broeck, W., Cox, E., Oudega, B., and Goddeeris, B. (2000). The F4 fimbrial antigen of Escherichia coli and its receptors. Vet. Microbiol. 71, 223–244. doi: 10.1016/S0378-1135(99)00174-1
Van Nevel, C. J., Decuypere, J. A., Dierick, N. A., and Molly, K. (2005). Incorporation of galactomannans in the diet of newly weaned piglets: effect on bacteriological and some morphological characteristics of the small intestine. Arch. Anim. Nutr. 59, 123–138. doi: 10.1080/17450390512331387936
Van Soest, P. J., Robertson, J. B., and Lewis, B. A. (1991). Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597. doi: 10.3168/jds.S0022-0302(91)78551-2
Vazquez-Gutierrez, P., de Wouters, T., Werder, J., Chassard, C., and Lacroix, C. (2016). High iron-sequestrating bifidobacteria inhibit enteropathogen growth and adhesion to intestinal epithelial cells in vitro. Front. Microbiol. 7:1480. doi: 10.3389/fmicb.2016.01480
Verheijden, K. A., Akbari, P., Willemsen, L. E., Kraneveld, A. D., Folkerts, G., Garssen, J., et al. (2015). Inflammation-induced expression of the alarmin interleukin 33 can be suppressed by galacto-oligosaccharides. Int. Arch. Allergy Immunol. 167, 127–136. doi: 10.1159/000437327
Vulevic, J., Juric, A., Tzortzis, G., and Gibson, G. R. (2013). A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 143, 324–331. doi: 10.3945/jn.112.166132
Walter, J., Maldonado-Gómez, M. X., and Martínez, I. (2018). To engraft or not to engraft: an ecological framework for gut microbiome modulation with live microbes. Curr. Opin. Biotechnol. 49, 129–139. doi: 10.1016/j.copbio.2017.08.008
Wang, J., Tian, S., Yu, H., Wang, J., and Zhu, W. (2019). The response of colonic mucosa-associated microbiota composition, mucosal immune homeostasis, and barrier function to early-life galactooligosaccharides intervention in suckling piglets. J. Agric. Food Chem. 67, 578–588. doi: 10.1021/acs.jafc.8b05679
Zhang, B., Ren, J., Yan, X., Huang, X., Ji, H., Peng, Q., et al. (2008). Investigation of the porcine MUC13 gene: isolation, expression, polymorphisms and strong association with susceptibility to enterotoxigenic Escherichia coli F4ab/ac. Anim. Genet. 39, 258–266. doi: 10.1111/j.1365-2052.2008.01721.x
Keywords: synbiotic, enterotoxigenic Escherichia coli, piglet, probiotic, prebiotic, galacto-oligosaccharides, Bifidobacterium, Lactobacillus
Citation: Rodríguez-Sorrento A, Castillejos L, López-Colom P, Cifuentes-Orjuela G, Rodríguez-Palmero M, Moreno-Muñoz JA, Luise D, Trevisi P and Martín-Orúe SM (2021) Effects of the Administration of Bifidobacterium longum subsp. infantis CECT 7210 and Lactobacillus rhamnosus HN001 and Their Synbiotic Combination With Galacto-Oligosaccharides Against Enterotoxigenic Escherichia coli F4 in an Early Weaned Piglet Model. Front. Microbiol. 12:642549. doi: 10.3389/fmicb.2021.642549
Edited by:
Hang Xiao, University of Massachusetts Amherst, United StatesCopyright © 2021 Rodríguez-Sorrento, Castillejos, López-Colom, Cifuentes-Orjuela, Rodríguez-Palmero, Moreno-Muñoz, Luise, Trevisi and Martín-Orúe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lorena Castillejos, bG9yZW5hLmNhc3RpbGxlam9zQHVhYi5jYXQ=