Abstract
Methanogens are anaerobic archaea which conserve energy by producing methane. Found in nearly every anaerobic environment on earth, methanogens serve important roles in ecology as key organisms of the global carbon cycle, and in industry as a source of renewable biofuels. Environmentally, methanogenic archaea play an essential role in the reintroducing unavailable carbon to the carbon cycle by anaerobically converting low-energy, terminal metabolic degradation products such as one and two-carbon molecules into methane which then returns to the aerobic portion of the carbon cycle. In industry, methanogens are commonly used as an inexpensive source of renewable biofuels as well as serving as a vital component in the treatment of wastewater though this is only the tip of the iceberg with respect to their metabolic potential. In this review we will discuss how the efficient central metabolism of methanoarchaea could be harnessed for future biotechnology applications.
Methanogen ecology and diversity
Methanogens are single-celled organisms that conserve energy via the conversion of substrate carbon compounds into methane gas (Ferry, 2012). The majority of methanogens subsist in anaerobic environments by the reduction of one carbon (C1) compounds including carbon dioxide and carbon monoxide, methanol, methylamines, and methyl sulfides as well as the fermentation of acetate (Daniels et al., 1977; Rother and Metcalf, 2004; Buan, 2018). The gaseous methane they produce then bubbles back into the aerobic world where it is consumed by methanotrophic organisms and is returned to the carbon cycle. The methane produced by methanogens is of interest due to methane’s ecological impact resulting from agricultural production by livestock (Johnson and Johnson, 1995) and rice cultivation (Schütz et al., 1989) as well as methane’s benefits as a renewable source of natural gas (Luo and Angelidaki, 2012; Huang et al., 2017) which is a high energy fuel used for heat, electricity generation, and for transportation including as a propellant for rocket engines (Neill et al., 2009; Sheehan, 2021). In nature, methanogenic archaea have been identified in environments spanning the boundaries of life sustaining conditions, from acidic to alkaline (pH 3.0–10.2), thermophilic to psychrophilic (−2°C to 110°C), and including both fresh and saline aquatic environments (Martin and Sousa, 2016). In addition to these environments, methanogens are found symbiotically communing in a wide range of single-and multi-cellular hosts ranging from amoebae (Holmes et al., 2014) and protozoa (Stumm and Zwart, 1986) to termites, (Brune, 1998, 2018) bovines (Whitford et al., 2001), and humans (Fricke et al., 2006; Rajilić-Stojanović et al., 2007).
As more methanogen species are discovered, it is becoming evident that methanogens may be able to use a wider variety of substrates than previously known. Biomethane generation has been observed from subsurface coal beds (Ulrich and Bower, 2008; Mayumi et al., 2016) as well as oceanic oil sinks (Laso-Pérez et al., 2019; Zhou et al., 2022). Methanogens may form syntrophic partnerships with other microorganisms such as hydrocarbon-degrading bacteria, thereby indirectly facilitating the reintroduction of crude oil carbon into a bioavailable state (Zengler et al., 1999; Dolfing et al., 2008; Jones et al., 2008). However, methanogens are suspected of being capable of alkane oxidation independent of any other archaeal or bacterial partner (Borrel et al., 2019; Laso-Pérez et al., 2019). Ecological methane accumulation has been observed in correlation with colonized oil droplets at deep-sea oil seeps. These proposed alkane utilizing methanogens are not limited to short-chain alkanes; Candidatus Methanoliparum has been shown to degrade long-chain hydrocarbons with methanogenesis (Zhou et al., 2022). The mechanism by which these so far uncultured alkanotrophic methanogens are capable of utilizing hydrocarbons is still being investigated, though the phenomenon does not appear to be a rare occurrence. Alkane-degrading methanogens are widely distributed, (Zengler et al., 1999; Laso-Pérez et al., 2019; Zhou et al., 2022) indicating that methanogens are directly or indirectly involved in the bioconversion of crude oil to methane on a large scale and may serve a benefit to bioremediation efforts in anaerobic environments such as deep-sea sediments.
The ability for methanogens to thrive in these diverse environments is testament to their metabolic robustness. Regardless of the environment they inhabit, methanogens share a similar metabolic niche, the bioconversion of low-energy substrates into biomass and high-energy molecules with a high degree of efficiency. All cultured methanogens to date are strict obligate anaerobes and produce methane as an essential byproduct of metabolism (Daniels et al., 1977; Rother and Metcalf, 2004; Ferry, 2012; Buan, 2018). To grow on these energy poor substrates methanogens have adopted a highly efficient pathway for conserving energy called methanogenesis (Figure 1; Thauer, 2012; Gonnerman et al., 2013). In this review we discuss how methanogen metabolism allows these organisms to thrive under strict energetic conditions and how their special metabolic features could be utilized in biotechnology.
Figure 1
Expanding the methanogenesis pathway
Despite their ability to live in a wide diversity of habitats, methanogens are united by their unique central metabolism. In the five characterized versions of the methanogenesis pathway, substrates are reduced to methane while formate, primary alcohols/amines/thiols, or H2 are oxidized to CO2 or H2O (Ermler et al., 1997; Buan, 2018). Redox cofactors associated with the methanogenesis pathway are regenerated through formation of a transmembrane ion gradient which is coupled to ATP synthesis via ATP synthase (Costa and Leigh, 2014; Diender et al., 2015). These reactions yield a small amount of energy for the methanogen amounting to between 0.5 and 2 moles of ATP per mole of substrate (Buan, 2018). A result of this low energy yield is a high relative flux through energy conservation pathways, with over 99% of the chemistry within the cell being directly tied to methanogenesis (Feist et al., 2006). The remaining 1–2% of carbon substrate is used to generate biomass for replication. The average macromolecular composition of a methanogen includes 63% protein, 0.1% fatty acid lipids, 5% isoprenoid lipids, 0.5% carbohydrates, 28% nucleic acids, and 4% metabolites and metabolic precursors (Gonnerman et al., 2013). The relatively high abundance of isoprenoid lipids and high protein concentration make them an appealing source of difficult-to-synthesize lipids and molecules from inexpensive C1 compounds or acetate, yields and titers of which could be further enhanced through genetic engineering. The unique properties of methanogenesis and highly efficient energy conservation mechanisms make methanoarchaea ideal organisms for the production of renewable biofuels as the vast majority of feed substrate is converted efficiently to methane.
It should be noted, however, that while methanogenesis is highly conserved and exceedingly efficient, it can also be modified to better serve biotechnological goals without necessarily undermining methanogenic growth. Methanosarcina in particular may be well-suited to metabolic engineering, as they can use multiple methanogenesis pathways and are genetically tractable (Metcalf et al., 1997; Ehlers et al., 2005).
Methanogenesis is inherently limited by substrate availability though this limitation can be overcome by expanding the carbon and energy sources available to methanoarchaea. Methanosarcina acetivorans has been successfully engineered to expand its substrate use and to enhance metabolic efficiency. M. acetivorans is a marine methanogen that can use methylotrophic and acetotrophic methanogenesis pathways, but unusually cannot use H2 for methanogenesis (Sowers et al., 1984; Guss et al., 2009). As a result, M. acetivorans appears to use very efficient intracellular redox balancing mechanisms, thus avoiding loss of H2 reducing equivalents by gas diffusion, which is a possibility for methanogens that use H2 cycling to generate transmembrane proton gradients (Kulkarni et al., 2009). Methylotrophic methanogenesis relies on substrate specific methyltransferases to convert substrates to CH3-CoM for entry into the pathway. It has been demonstrated that heterologous expression of the bacterial broad-specificity esterase from Pseudomonas veronii in M. acetivorans increased esterase activity 80-fold and greatly enhanced growth on methyl acetate and methyl propionate substrates (Lessner et al., 2010). Once substrates have entered the methanogenesis pathway, energy conservation occurs by the regeneration of methanogenic cofactors (Thauer, 2012). Cofactor regeneration is catalyzed by membrane bound, redox-driven enzyme complexes such as Rnf (Schlegel et al., 2012) and HdrED (Duszenko and Buan, 2017), which combine cofactor regeneration with ion transport or by cytoplasmic enzymes such as Fpo (Welte and Deppenmeier, 2011), or the terminal oxidase HdrABC (Catlett et al., 2015; Buckel and Thauer, 2018). By enhancing cofactor regeneration it is possible to stimulate increased methanogenesis. For example, when the cytoplasmic enzyme heterodisulfide reductase (HdrABC) is overexpressed, methane production on methanol is 30% faster without a detectable change in growth rate compared to the parent strain (Catlett et al., 2015). The exogenous addition of methanophenzine (MPh), an electron carrier found in methanogens which fulfils a similar role as quinones in other electron transport chains, was found to significantly increase growth in Methanosarcina spp (Duszenko and Buan, 2017). Additionally, it is possible that the regeneration of methanogenic cofactors could be achieved through pathway engineering (Aldridge et al., 2021). The reduction of the disulfide complex between coenzyme M and coenzyme B is the final step in all methanogenic pathways and is restricted to the heterodisulfide reductases HdrABC and HdrED (Buan and Metcalf, 2010; Yan and Ferry, 2018). Providing an alternative means of cofactor reduction would eliminate this metabolic bottleneck, freeing up cofactors at a greater rate (Aldridge et al., 2021). If a methanogen were engineered to produce a non-native metabolite which allows for the reduction of ferredoxin, F420, coenzyme M, or coenzyme B then production of that metabolite has the potential to increase the rate of methanogenesis while also synthesizing the desired product (Aldridge et al., 2021).
Due to the tight energetic restrictions methanogenesis is proposed to rely heavily on substrate channeling to minimize entropic effects (Costa et al., 2010; Matschiavelli et al., 2012; Catlett et al., 2015; Yan and Ferry, 2018; Watanabe et al., 2021). Substrate channeling allows methanogenesis to function efficiently but presents challenges for metabolic engineers as the metabolite pools for methanogenesis have limited availability outside of the channeled enzyme complexes. To overcome this metabolic obstacle metabolic engineers must choose products which draw from metabolites which are not directly channeled or incorporate the production of their products within methanogenesis. Table 1 lists potential strategies to increase substrate variety, optimize growth rates and culture conditions, or generate new metabolic products by engineering methanogenesis.
Table 1
| Desired trait | Potential mechanism |
|---|---|
| Increased methanogenesis and methane production |
|
| Increased substrate uptake rates |
|
| Increased substrate diversity and mixotrophy |
|
| Controlled energy conservation |
|
| Increased stress resistance |
|
| Multiple |
|
Strategies for expanding the metabolic potential of the methanogenesis pathway.
Genetic methods in methanoarchaea are available in several species. Current tools have been recently reviewed or published (Tumbula et al., 1994; Pritchett et al., 2004; Ehlers et al., 2005; Guss et al., 2008; Atomi et al., 2012; Kohler and Metcalf, 2012; Nayak and Metcalf, 2017; Innard, 2018).
Anaerobic oxidation of methane and reverse methanogenesis
Given the efficiency of methanogenesis and the abundance of anaerobic environments around the world, methanogens are distributed across every continent. Yet of the approximately 1 Gt of methane produced by methanogens in the wild each year in anaerobic and microanaerobic environments, roughly half escapes into the aerobic carbon cycle (Conrad, 2009). It is estimated between 43–90% of biogenic methane is oxidized my aerobic methanotrophs at the anaerobic/aerobic interface (Hao et al., 1988; Roslev and King, 1995; Le Mer and Roger, 2001; Conrad, 2009). The remainder of this methane is either trapped within anaerobic environments (as gas or methane gas hydrates) or consumed by methanotrophic archaea and bacteria (Knittel and Boetius, 2009; Thauer, 2011). Given the estimated 70Gt of CO2 fixed by photosynthesis into biomass, methane as a product of methanogenesis accounts for approximately 2% of the annual total carbon utilization (Thauer et al., 2008). Previously it was believed that the anaerobic oxidation of methane (AOM) was possible through the symbiotic exchange of metabolites and electrons between the methanotrophic archaea and sulfate reducers (Alperin and Hoehler, 2009; Summers et al., 2010). Within this process anaerobic methane-oxidizing archaea (ANME) consisting of Methanomicrobiales (ANME-1) and Methanosarcinales (ANME-2 and ANME-3) form granular aggregates with delta-proteobacteria in which electrons are transferred between organisms via multi-heme cytochromes (McGlynn et al., 2015). Metabolic modeling has suggested that iron and sulfate can be co-substrates in AOM (Riedinger et al., 2014) and 16S rRNA gene-sequences for Candidadus Methanoperedens correlated with increased AOM in sulfate-rich anoxic sediments suggesting the occurrence of AOM independent of a bacterial partner (Su et al., 2020). In laboratory conditions it was found that trace amounts of AOM was observed in Methanothermobacter marburgenis (Scheller et al., 2010) and Methanosarcina acetivorans (Moran et al., 2005) though it was not observed that these strains were able to use methane as the major source of carbon and energy for growth. However, by scouring the metagenomes of unculturable ANME-1 samples from aquatic regions with high amounts of AOM, a novel variant of methyl-coenzyme M reductase (Mcr) was discovered which correlated to AOM without the need for a syntrophic sulfate-reducing partner (Meyerdierks et al., 2010; Shima et al., 2012). When the uncultured ANME-1 Mcr was introduced into M. acetivorans it was found that isotope labeled methane was converted into acetate while also facilitating growth, (Soo et al., 2016). Furthermore, methanogen strains containing this ANME-1 Mcr gene can be utilized along with a consortia of microbes including Geobacter sulfurreducens to produce electricity in a microbial fuel cell utilizing only methane as a substrate (McAnulty et al., 2017). As every step of methanogenesis is reversible, reverse methanogenesis is theoretically possible for any methanogen though under most conditions these reactions are non-energy yielding (Thauer, 2012). These observations indicate that the bidirectionality of methanogenesis enables methane to be utilized as growth substrate for methanogens, particularly by Methanosarcina spp. For example: a Methanosarcina culture which has been engineered to produce a high-value terpenoid product is grown using methyl compounds until stationary phase is achieved and biomass accumulation is no longer necessary; this culture could then be induced to produce the terpenoids utilizing potentially any C1 compound or mixtures of compounds including CO, CO2, or CH4 based on substrate availability. This potential extends beyond the production of secreted products, as the biomass of methanogens itself can be utilized as a source of valuable lipids.
Potential for engineering the lipid membrane biosynthesis pathway as a valorization strategy
Methanogen membranes, like those found in all archaea, are distinct from those found in bacteria and eukarya. In bacterial and eukaryotic organisms lipid membrane structures are composed of fatty acid chains ester liked to glycerol-3-phosphate (G3P) (Koga and Morii, 2007). Archaeal lipids membranes instead utilize isoprenoid alkyl chains ether linked to glycerol-1-phosphate (G1P; Figure 2; Koga and Morii, 2007; Koga, 2012). This fundamental differentiation in membrane composition is the basis of the so called ‘lipid divide’ separating archaea from the other two domains of life (Villanueva et al., 2021). Given the high quantity and the molecular uniformity of lipid membranes, comprising on average 5% of total methanogen dry weight (Gonnerman et al., 2013), and the relatively high metabolic flux through the archaeal mevalonate lipid biosynthesis pathway, high-value isoprenoid lipids are attractive metabolic engineering targets. The isoprenoid lipids used by archaea allow them to tolerate a wide range of environmental stressors. The most abundant archaeal lipid structures are archeol, consisting of a pair of phytanyl chains ether linked to G1P and caldarcheol, a cyclic dimer of archeol. Caldarcheol is of particular biotechnological interest as the cyclized tetraether lipids maintain cellular homeostasis in the presence of extreme pH and thermal stress (Boyd et al., 2013; Siliakus et al., 2017). Archaeal ether linked lipids are more stable than ester linked membranes when exposed to extremes of pH and thermal conditions, and the unique monolayer structure of tetraether linked lipids imparts resistance to degradation to phospholipases (Jacquemet et al., 2009). These stable properties and the intrinsic monolayer formed by caldarcheol represents an enticing alternative to traditional phospholipids in liposome-based commercial applications. One such application is in the delivery of chemotherapeutic compounds via archaeal derived liposomes. It has been found that tetraether linked artificial liposomes reduce leakage of chemotherapeutic compounds by 9-fold compared to conventional eukaryotic derived liposomes, which results in a lower dose required for therapeutic effects (Leriche et al., 2017). The archaeal liposomes themselves also contribute therapeutic effects as archaeal liposomes utilized to transport vaccine components induce robust antigen specific humoral and cellular immune responses exceeding those found from traditional delivery mechanisms (Conlan et al., 2001; Patel and Chen, 2010; Haq et al., 2016; Landi et al., 2017).
Figure 2
In addition to the direct application of archaeal lipids, the high metabolic flux through the archaeal mevalonate pathway presents an opportunity for low-cost production of terpene compounds. Terpenes are the largest class of natural compounds and have a wide range of commercial applications. Odorant terpenes such as limonene, eucalyptol, and linalool are cornerstones of the $29B flavor and fragrance industry (Markets, 2021). In addition to odorants, terpenes are often the active compound in pharmaceuticals including the anti-cancer drug paclitaxel and the antimalarial artemisinin. Hundreds of natural terpenes have shown promising bioactivity (Gould, 1997; Sgadari et al., 2000; Friedman et al., 2002; Paduch et al., 2007; Ajikumar et al., 2008) yet are limited in application due to their availability. Many of these terpenes are currently harvested from their native plant, fungal, and marine producers which are limited by the endogenous expression levels which are prohibitively low (Long et al., 1998; Sills et al., 1998; Newman and Cragg, 2004) or non-renewably synthesized from petroleum precursors. Organically produced terpenes are primarily produced via compounds derived from one of two isoprenoid synthesis pathways, the mevalonate (MVA) pathway and the deoxyxylose 5-phosphate (DXP) pathway (Lange et al., 2000). These pathways in non-archaeal organisms suffer low carbon flux and depletion of precursors towards non-target compounds (McGarvey and Croteau, 1995; Rodriguez-Concepcion and Boronat, 2002; Vranova et al., 2012). Archaea, however, synthesize the majority of membrane lipids through the mevalonate pathway, accounting for a higher flux as compared to eukaryal or bacterial organisms (Boucher et al., 2004; Jain et al., 2014; Villanueva et al., 2014). As such, there is a naturally higher abundance of metabolic precursors available for the synthesis of isoprenoid and terpene products using methanoarchaea. Concerns over the depletion of these membrane precursors have been alleviated by the synthesis of mono-isoprene from engineered strains of M. acetivorans and Methanosarcina barkeri (Aldridge et al., 2021; Carr et al., 2021). These strains demonstrate that methanogens are able to withstand the metabolic burden of membrane substrate depletion without a significant decrease in growth rate or final carrying capacity, opening the door for further isoprenoid products that could be produced by addition of relatively few genes (Table 2). Inducible promoters such as Ptet could also be used to drive expression of genes for terpenoid biosynthesis in two-stage fermentation processes to increase bioreactor carrying capacity and maximize terpenoid titer and yield (Urlinger et al., 2000; Loew et al., 2010). One challenge is that some terpenes require molecular oxygen for complete biosynthesis and this might be difficult for anaerobic organisms to achieve. However, Methanosarcina acetivorans is remarkably oxygen-tolerant and it is possible to further enhance resistance to oxidative stress through engineering or adaptation (Horne and Lessner, 2013; Jasso-Chávez et al., 2015). Therefore, it is theoretically feasible to use O2 availability as a biosynthetic inducer during terpene fermentation with oxygen-tolerant methanogens.
Table 2
| Name | Terpene class | Structure | Synthesis Enzyme | Substrate | Initial citation |
|---|---|---|---|---|---|
| Isoprene | Hemiterpene | 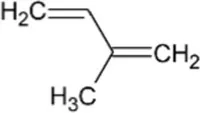 | Isoprene synthase (4.2.3.27) | Dimethylallyl pyrophosphate | Silver and Fall (1991) |
| Geranyl pyrophosphate (GPP) | Monoterpene | 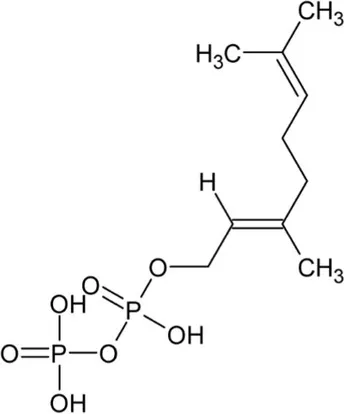 | (2E,6E)-farnesyl diphosphate synthase (2.5.1.10) | Dimethylallyl pyrophosphate | Cornforth et al. (1966) |
| Geraniol | Monoterpene | 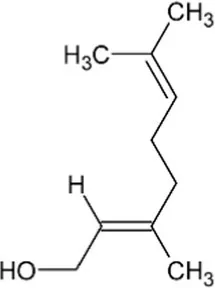 | Geraniol synthase (3.1.7.11) | Geranyl diphosphate | Iijima et al. (2004) |
| Linalool | Monoterpene | 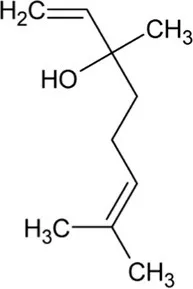 | S-linalool synthase (4.2.3.25) | Geranyl diphosphate | Pichersky et al. (1994) |
| Ocimene | Monoterpene | 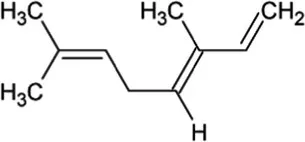 | (E)-beta-ocimene synthase (4.2.3.106) | Geranyl diphosphate | Bohlmann et al. (2000) |
| Myrcene | Monoterpene | 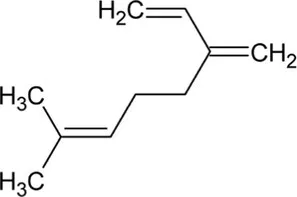 | Myrcene synthase (4.2.3.15) | Geranyl diphosphate | Bohlmann et al. (1997) |
| Sabinene | Bicyclic Monoterpenoid | 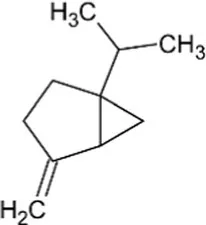 | (+)-sabinene synthase (4.2.3.110) | Geranyl diphosphate | Wise et al. (1998) |
| Pinene | Bicyclic Monoterpenoid | 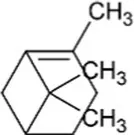 | Pinene synthase (4.2.3.14) | Geranyl diphosphate | Gambliel and Croteau (1984) |
| Farnesyl diphosphate | Acyclic Sesquiterpenoid | 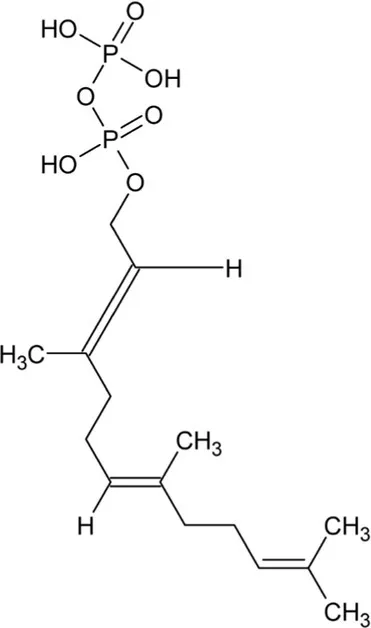 | Farnesyl diphosphate synthase (2.5.1.1) | Dimethylallyl diphosphate and isopentenyl diphosphate | Vandermoten et al. (2009) |
| Farnesol | Acyclic Sesquiterpenoid | 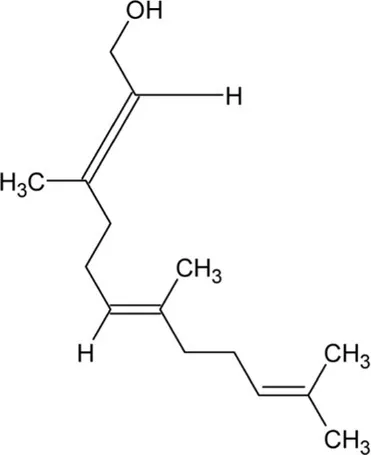 | Farnesyl diphosphatase (3.1.7.6) | Farnesyl diphosphate | Meigs and Simoni (1997) |
| Nerolidol | Acyclic Sesquiterpenoid | 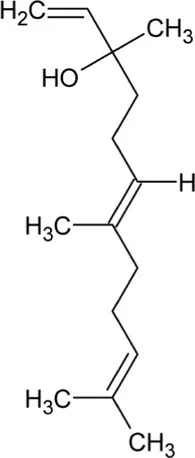 | (3S,6E)-nerolidol synthase (4.2.3.48) | Farnesyl diphosphate | Donath and Boland (1995) |
| Farnesene | Acyclic Sesquiterpenoid | 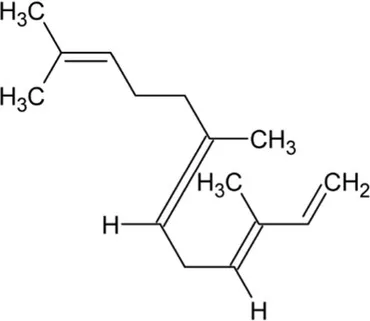 | Alpha-farnesene synthase (4.2.3.46) and beta-farnesene synthase (4.2.3.47) | Farnesyl diphosphate | Pechous and Whitaker (2004) |
| Humulene | Monocyclic Sesquiterpenoid | 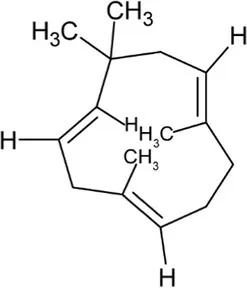 | Alpha-humulene synthase (4.2.3.104) | Farnesyl diphosphate | van Der Hoeven et al. (2000) |
| Bisabolene | Monocyclic Sesquiterpenoid | 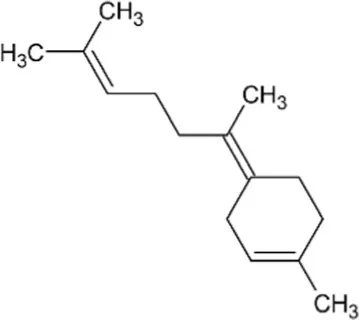 | Alpha-bisbolene synthase (4.2.3.38) | Farnesyl diphosphate | Bohlmann et al. (1998) |
| Zingiberene | Monocyclic Sesquiterpenoid | 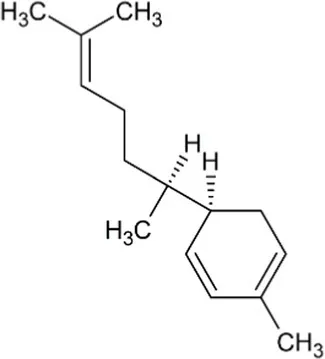 | Zingiberene synthase (4.2.3.65) | Farnesyl diphosphate | Zhuang et al. (2012) |
| Curcumene | Monocyclic Sesquiterpenoid | 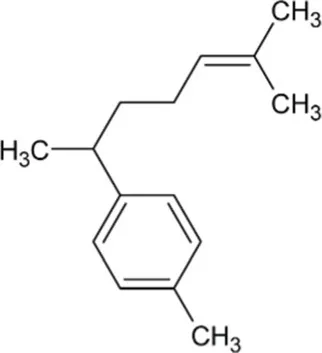 | Gamma-curcumene synthase (4.2.3.94) | Farnesyl diphosphate | Deguerry et al. (2006) |
| Amorphadiene | Bicyclic Sesquiterpenoid | 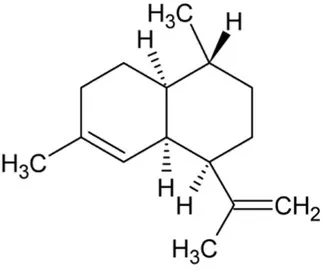 | Amorpha-4,11-diene synthase (4.2.3.24) | Farnesyl diphosphate | Bouwmeester et al. (1999) |
| Valencene | Bicyclic Sesquiterpenoid | 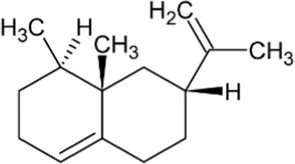 | Valencene synthase (4.2.3.73) | Farnesyl diphosphate | Sharon-Asa et al. (2003) |
Potential terpenoids to be produced by methanogens based on category.
Terpenoid clasess requiring molecular oxygen for biosynthesis have been omitted. However, oxygen-tolerant Methanosarcina have potential to be used in this context.
Benefits and challenges of methanogen biotechnologies
The use of methanogens in bioproduction is beneficial in a myriad of ways including ease of selection, low cost of media, and flexibility of products (Table 3). Methanogens have been shown to be an excellent source of metabolically active compounds such as coenzyme M (CoM) which acts as a potent chemotherapy adjuvant as the drug mesna (Shaw and Graham, 1987) as well as immune stimulating lipids for vaccine delivery (Patel and Chen, 2010; Haq et al., 2016). Due to their anaerobic metabolism which requires a lack of O2, they are able to produce novel chiral precursors which could later be tailored by chemists through custom oxidation steps and subsequent functionalization.
In large scale industrial fermentations pure aseptic environments are difficult to maintain, and often media and growth conditions are utilized to ensure continuous selection during the fermentation (Mosier and Ladisch, 2011; Doran, 2013). Methanogens circumvent this issue by growing in selective environments free of oxygen using substrates that cannot be used by the majority of common contaminating factors such as lactic acid bacteria and fungi (Skinner and Leathers, 2004; Beckner et al., 2011). Methanogens are prototrophic organisms, able to synthesize all vitamins and cofactors required for growth from inorganic material, allowing for additional selection by limiting available vitamins and nutrients required for contaminating growth by exclusion (Patil and Muskan, 2009; Thauer, 2012; Buan, 2018). While viral predation on methanogens has been observed (Park et al., 2007) there is little evidence that these methanophage/methanovirus particles have a substantial effect on methanogenic digestor performance as viral titers did not correlate with a significant decrease in methane output and methanogen carrying capacity.
Another major challenge in industrial fermentations is the large amounts of fresh water required for E. coli or yeast (Chen, 2012). Methanogens, however, thrive in environments with high salt concentrations, allowing for the utilization of seawater in fermentations. Non-sterile hypersaline environments such ocean water and hydraulic fracking fluids have been demonstrated to select for methylotrophic methanogens such as Methanohalophilus, Methanohalobium, and Methanosarcina spp. while also presenting a high concentration of non-competitive substrates such as methylamines (McGenity, 2010; Guan et al., 2015). Methanogens are utilized worldwide for the production of renewable biogas in non-selective environments with high degrees of contamination such as municipal and agricultural wastewater treatment. In these environments methanogens are exposed to a wide variety of stressors including dramatic shifts in ammonia, osmotic shifts, and exposure to heavy metals (Yan et al., 2020). Many methanogens are natively capable of withstanding these stressors (Macario and Macario, 2003) though as stated above, using genetic tools it is possible to combine or “stack” desirable traits onto a single methanogen strain to gain the maximum benefit from a single organism.
Table 3
| Benefits | Challenges |
|---|---|
| Methanogens are some of the fastest-replicating organisms, particularly members of Methanococcus (Jones et al., 1983; Goyal et al., 2016; Long et al., 2017) and Methanopyrus (Takai et al., 2008) genus. (Jones et al., 1983; Takai et al., 2008; Goyal et al., 2016; Long et al., 2017). | Strain differences in growth rate and carrying capacity. Growth is flux-controlled depending on substrate feed rates. Gas-phase fermentation presents similar problems as oxygenation in traditional fermentations (Mosier and Ladisch, 2011; Chen, 2012; Luo and Angelidaki, 2012). |
| Methanogens can grow on inexpensive substrates including negative value substrates such as wastewater (Daniels et al., 1977; Schiraldi et al., 2002; McGenity, 2010; Ferry, 2012; Costa and Leigh, 2014; Borrel et al., 2016; Buan, 2018; Chadwick et al., 2022) Methanogens already scaled up worldwide for water treatment and biogas production. | Process disfavors growth of aerobic pathogens. Co-product can be water ready for discharge to aquifers and waterways. |
| Can be coupled directly or indirectly to electrodes for carbon capture by electrosynthesis or for electricity generation from biomass (Ragab et al., 2020). | Surface-to-area, substrate solubility, and other challenges commensurate with microbial fuel cell technologies. |
| Oxygenation not required. Can grow on non-gas substrates. No contamination by aerobic organisms. | Methanogens require specialized culture environments to maintain anaerobicity (Balch et al., 1979; Rouviere and Wolfe, 1988; Buan, 2018). |
| Mesophilic and thermophilic strains available to tailor to the desired product and process needs. | Methanogen chassis organisms may need different optimization strategies. |
| Novel metabolic pathways are constantly being discovered (Costa and Leigh, 2014; Guan et al., 2015; Borrel et al., 2016; Mayumi et al., 2016; Buan, 2018; Yan and Ferry, 2018; Chadwick et al., 2022; Zhou et al., 2022). | Methanogen genetics and biochemistry are less characterized than other model organisms. |
| Synthetic biology pathways often use archaeal or methanogen genes to improve yields and reduce feedback inhibition. | |
| Bacterial synthetic biology and genetic strategies have been successfully translated to methanogens. | |
| Methanogens have a high substrate to volume ratio with low accumulation of biomass relative to products (Thauer et al., 2008; Ferry, 2012; Buan, 2018). | High titers of intracellular products may be difficult to obtain unless accumulated into vacuoles or secreted extracellularly. |
| Multiple validated genetic tools available including tools for Methanosarcina spp., (Metcalf et al., 1997; Buan et al., 2011; Nayak and Metcalf, 2017) Methanococcus maripaludis,(Blank et al., 1995; Bao and Scheller, 2021) and Methanothermobacter thermautotrophicus (Buan et al., 2011; Sarmiento et al., 2011; Nayak and Metcalf, 2017; Bao and Scheller, 2021; Fink et al., 2021). | Variability in genome copy number can present challenges when performing chromosomal modifications (Hildenbrand et al., 2011; Aldridge et al., 2021). |
| The lack of cell wall and envelope in most methanogens ensures that products generated through methanogen fermentations are not contaminated with peptidoglycan or endotoxin (Jones et al., 1987; Claus and König, 2010). | Some methanogen species produce pseudomurein cell walls or extracellular polysaccharide capsules, although these are generally non-or weakly immunogenic (Sirohi et al., 2010; Subedi et al., 2021). |
| Methanoarchaea are non-pathogenic, though there have been studies suggesting a link between methanogens and other microbes in dysbiotic anaerobic abscesses (Drancourt et al., 2017; Sogodogo et al., 2019). | Not currently recognized as a GRAS (Generally Regarded as Safe) organism. |
Benefits and challenges of methanogen biotechnology.
Conclusion
Methanogens are biologically important organisms with a wide-reaching impact both in ecological and biotechnological applications. Their extremely efficient central metabolism makes them an ideal source of renewable biofuels that can be captured through anaerobic digestion or fermentation processes. They are able to grow prototrophically with inexpensive feedstocks and can produce endotoxin-free protein, carbohydrates, and valuable isoprenoid lipids. Their unique membrane composition can be used to expand the biotechnological toolbox for the delivery of chemotherapeutics as well as source for novel terpene compounds previously not available via conventional extraction means. By continuing to investigate the molecular, genetic, and synthetic biology potential of these unique organisms, researchers may unlock a wide range of applications from environmental and ecological management, renewable energy, agriculture, chemical manufacturing, and pharmaceutic industries.
Funding
This work was supported by grants from the National Science Foundation (IOS-1938948) and the Nebraska Center for Energy Sciences Research (Cycle 15). Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the funding agencies.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Statements
Author contributions
SC conceived and wrote the manuscript. NRB conceived, wrote, and edited the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of interest
NRB has disclosed a significant financial interest in RollingCircle Biotech, LLC and Molecular Trait Evolution, LLC.
SC declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
AjikumarP. K.TyoK.CarlsenS.MuchaO.PhonT. H.StephanopoulosG. (2008). Terpenoids: opportunities for biosynthesis of natural product drugs using engineered microorganisms. Mol. Pharm.5, 167–190. doi: 10.1021/mp700151b
2
AldridgeJ.CarrS.WeberK. A.BuanN. R. (2021). Anaerobic production of isoprene by engineered Methanosarcina species archaea. Appl. Environ. Microbiol.87, e02417–e02420. doi: 10.1128/AEM.02417-20
3
AlperinM. J.HoehlerT. M. (2009). Anaerobic methane oxidation by archaea/sulfate-reducing bacteria aggregates: 1. Thermodynamic and physical constraints. Am. J. Sci.309, 869–957. doi: 10.2475/10.2009.02
4
AndersonK. L.ApolinarioE. E.SowersK. R. (2012). Desiccation as a long-term survival mechanism for the archaeon Methanosarcina barkeri. Appl. Environ. Microbiol.78, 1473–1479. doi: 10.1128/AEM.06964-11
5
AtomiH.ImanakaT.FukuiT. (2012). Overview of the genetic tools in the archaea. Front. Microbiol.3:337. doi: 10.3389/fmicb.2012.00337
6
BalchW.FoxG.MagrumL.WoeseC.WolfeR. (1979). Methanogens: reevaluation of a unique biological group. Microbiol. Rev.43:260. PMCID:
7
BaoJ.SchellerS. (2021). Efficient CRISPR/Cas12a-based genome editing toolbox for metabolic engineering in Methanococcus maripaludis. bioRxiv. doi: 10.1101/2021.12.29.474413
8
BecknerM.IveyM. L.PhisterT. G. (2011). Microbial contamination of fuel ethanol fermentations. Lett. Appl. Microbiol.53, 387–394. doi: 10.1111/j.1472-765X.2011.03124.x
9
BlankC. E.KesslerP. S.LeighJ. A. (1995). Genetics in methanogens: transposon insertion mutagenesis of a Methanococcus maripaludis nifH gene. J. Bacteriol.177, 5773–5777.
10
BohlmannJ.CrockJ.JetterR.CroteauR. (1998). Terpenoid-based defenses in conifers: cDNA cloning, characterization, and functional expression of wound-inducible (E)-alpha-bisabolene synthase from grand fir (Abies grandis). Proc. Natl. Acad. Sci. U. S. A.95, 6756–6761. doi: 10.1073/pnas.95.12.6756
11
BohlmannJ.MartinD.OldhamN. J.GershenzonJ. (2000). Terpenoid secondary metabolism in Arabidopsis thaliana: cDNA cloning, characterization, and functional expression of a myrcene/(E)-β-ocimene synthase. Arch. Biochem. Biophys.375, 261–269. doi: 10.1006/abbi.1999.1669
12
BohlmannJ.SteeleC. L.CroteauR. (1997). Monoterpene synthases from grand fir (Abies grandis). cDNA isolation, characterization, and functional expression of myrcene synthase, (−)-(4S)-limonene synthase, and (−)-(1S,5S)-pinene synthase. J. Biol. Chem.272, 21784–21792. doi: 10.1074/jbc.272.35.21784
13
BorrelG.AdamP. S.GribaldoS. (2016). Methanogenesis and the Wood–Ljungdahl pathway: an ancient, versatile, and fragile association. Genome Biol. Evol.8, 1706–1711. doi: 10.1093/gbe/evw114
14
BorrelG.AdamP. S.McKayL. J.ChenL.-X.Sierra-GarcíaI. N.SieberC. M.et al. (2019). Wide diversity of methane and short-chain alkane metabolisms in uncultured archaea. Nat. Microbiol.4, 603–613. doi: 10.1038/s41564-019-0363-3
15
BoucherY.KamekuraM.DoolittleW. F. (2004). Origins and evolution of isoprenoid lipid biosynthesis in archaea. Mol. Microbiol.52, 515–527. doi: 10.1111/j.1365-2958.2004.03992.x
16
BouwmeesterH. J.WallaartT. E.JanssenM. H.van LooB.JansenB. J.PosthumusM. A.et al. (1999). Amorpha-4,11-diene synthase catalyses the first probable step in artemisinin biosynthesis. Phytochemistry52, 843–854. doi: 10.1016/s0031-9422(99)00206-x
17
BoydE.HamiltonT.WangJ.HeL.ZhangC. (2013). The role of tetraether lipid composition in the adaptation of thermophilic archaea to acidity. Front. Microbiol.4:62. doi: 10.3389/fmicb.2013.00062
18
BruneA. (1998). Termite guts: the world's smallest bioreactors. Trends Biotechnol.16, 16–21. doi: 10.1016/S0167-7799(97)01151-7
19
BruneA. (2018). In (Endo) symbiotic Methanogenic Archaea 81–101Berlin: Springer.
20
BuanN. R. (2018). Methanogens: pushing the boundaries of biology. Emerg. Topics Life Sci.2, 629–646. doi: 10.1042/ETLS20180031
21
BuanN.KulkarniG.MetcalfW. (2011). Genetic methods for Methanosarcina species. in Methods in Enzymology (Vol. 494, pp. 23–42). Academic Press.
22
BuanN. R.MetcalfW. W. (2010). Methanogenesis by Methanosarcina acetivorans involves two structurally and functionally distinct classes of heterodisulfide reductase. Mol. Microbiol.75, 843–853. PMID: . doi: 10.1111/j.1365-2958.2009.06990.x
23
BuckelW.ThauerR. K. (2018). Flavin-based electron bifurcation, ferredoxin, Flavodoxin, and anaerobic respiration with protons (Ech) or NAD(+) (Rnf) as electron acceptors: a historical review. Front. Microbiol.9. doi: 10.3389/fmicb.2018.00401
24
CarrS.AldridgeJ.BuanN. R. (2021). Isoprene production from municipal wastewater biosolids by engineered archaeon Methanosarcina acetivorans. Appl. Sci.11:3342. doi: 10.3390/app11083342
25
CatlettJ. L.CarrS.CashmanM.SmithM. D.WalterM.SakkaffZ.et al. (2022). R. Metabolic synergy between human symbionts Bacteroides and Methanobrevibacter. Microbiol Spectr.10:e0106722. doi: 10.1128/spectrum.01067-22 Epub 2022 May 10. PMID: ; PMID: PMC9241691.
26
CatlettJ. L.OrtizA. M.BuanN. R. (2015). Rerouting cellular electron flux to increase the rate of biological methane production. Appl. Environ. Microbiol.81, 6528–6537. doi: 10.1128/AEM.01162-15
27
ChadwickG. L.SkennertonC. T.Laso-PérezR.LeuA. O.SpethD. R.YuH.et al. (2022). Comparative genomics reveals electron transfer and syntrophic mechanisms differentiating methanotrophic and methanogenic archaea. PLoS Biol.20:e3001508. PMID: ; PMID: PMC9012536. doi: 10.1371/journal.pbio.3001508
28
ChenG.-Q. (2012). New challenges and opportunities for industrial biotechnology. Microb. Cell Factories11, 1–3. PMID: ; PMID: PMC3520750. doi: 10.1186/1475-2859-11-111
29
ClausH.KönigH. (2010). In Prokaryotic Cell Wall CompoundsBerlin: Springer.
30
ConlanJ. W.KrishnanL.WillickG. E.PatelG. B.SprottG. D. (2001). Immunization of mice with lipopeptide antigens encapsulated in novel liposomes prepared from the polar lipids of various Archaeobacteria elicits rapid and prolonged specific protective immunity against infection with the facultative intracellular pathogen Listeria monocytogenes. Vaccine19, 3509–3517. PMID: . doi: 10.1016/s0264-410x(01)00041-x
31
ConradR. (2009). The global methane cycle: recent advances in understanding the microbial processes involved. Environ. Microbiol. Rep.1, 285–292. doi: 10.1111/j.1758-2229.2009.00038.x
32
CornforthJ.CornforthR. H.PopjakG.YengoyanL. (1966). Studies on the biosynthesis of cholesterol: XX. Steric course of decarboxylation of 5-pyrophosphomevalonate and of the carbon to carbon bond formation in the biosynthesis of farnesyl pyrophosphate. J. Biol. Chem.241, 3970–3987. doi: 10.1016/S0021-9258(18)99800-5
33
CostaK. C.LeighJ. A. (2014). Metabolic versatility in methanogens. Curr. Opin. Biotechnol.29, 70–75. PMID: . doi: 10.1016/j.copbio.2014.02.012
34
CostaK. C.WongP. M.WangT.LieT. J.DodsworthJ. A.SwansonI.et al. (2010). Protein complexing in a methanogen suggests electron bifurcation and electron delivery from formate to heterodisulfide reductase. Proc. Natl. Acad. Sci. U. S. A.107, 11050–11055. PMID: ; PMID: PMC2890747. doi: 10.1073/pnas.1003653107
35
DanielsL.FuchsG.ThauerR.ZeikusJ. (1977). Carbon monoxide oxidation by methanogenic bacteria. J. Bacteriol.132, 118–126. PMID: ; PMID: PMC221834. doi: 10.1128/jb.132.1.118-126.1977
36
DeguerryF.PastoreL.WuS.ClarkA.ChappellJ.SchalkM. (2006). The diverse sesquiterpene profile of patchouli, Pogostemon cablin, is correlated with a limited number of sesquiterpene synthases. Arch. Biochem. Biophys.454, 123–136. doi: 10.1016/j.abb.2006.08.006
37
DienderM.StamsA. J.SousaD. Z. (2015). Pathways and bioenergetics of anaerobic carbon monoxide fermentation. Front. Microbiol.6:1275. doi: 10.3389/fmicb.2015.01275
38
DolfingJ.LarterS. R.HeadI. M. (2008). Thermodynamic constraints on methanogenic crude oil biodegradation. ISME J.2, 442–452. doi: 10.1038/ismej.2007.111
39
DonathJ.BolandW. (1995). Biosynthesis of acyclic homoterpenes: enzyme selectivity and absolute configuration of the nerolidol precursor. Phytochemistry39, 785–790. doi: 10.1016/0031-9422(95)00082-I
40
DoranP. M. ed. (2013). Biotechnology of hairy root systems. Springer.
41
DrancourtM.NkamgaV. D.LakheN. A.RégisJ.-M.DufourH.FournierP.-E.et al. (2017). Evidence of archaeal methanogens in brain abscess. Clin. Infect. Dis.65, 1–5. PMID: . doi: 10.1093/cid/cix286
42
DuszenkoN.BuanN. R. (2017). Physiological evidence for isopotential tunneling in the electron transport chain of methane-producing archaea. Appl. Environ. Microbiol.83, e00950–e00917. PMID: ; PMID: PMC5583484. doi: 10.1128/AEM.00950-17
43
EhlersC.WeidenbachK.VeitK.DeppenmeierU.MetcalfW. W.SchmitzR. A. (2005). Development of genetic methods and construction of a chromosomal glnK1 mutant in Methanosarcina mazei strain Gö1. Mol. Gen. Genomics.273, 290–298. doi: 10.1007/s00438-005-1128-7
44
ErmlerU.GrabarseW.ShimaS.GoubeaudM.ThauerR. K. (1997). Crystal structure of methyl-coenzyme M reductase: the key enzyme of biological methane formation. Science278, 1457–1462. PMID: . doi: 10.1126/science.278.5342.1457
45
FeistA. M.ScholtenJ. C.PalssonB. Ø.BrockmanF. J.IdekerT. (2006). Modeling methanogenesis with a genome-scale metabolic reconstruction of Methanosarcina barkeri. Mol. Syst. Biol.2:2006.0004. PMID: ; PMID: PMC1681478. doi: 10.1038/msb4100046
46
FerryJ. G. (2012). Methanogenesis: Ecology, Physiology, Biochemistry & Genetics. BerlinSpringer Science & Business Media. ISBN: 9781461523918.
47
FinkC.BeblawyS.EnkerlinA. M.MühlingL.AngenentL. T.MolitorB. (2021). A shuttle-vector system allows heterologous gene expression in the thermophilic methanogen Methanothermobacter thermautotrophicus ΔH. MBio12, e02766–e02721. PMID: ; PMCID: PMC8609365. doi: 10.1128/mBio.02766-21
48
FrickeW. F.SeedorfH.HenneA.KrüerM.LiesegangH.HedderichR.et al. (2006). The genome sequence of Methanosphaera stadtmanae reveals why this human intestinal archaeon is restricted to methanol and H2 for methane formation and ATP synthesis. J. Bacteriol.188, 642–658. PMID: ; PMCID: PMC1347301. doi: 10.1128/JB.188.2.642-658.2006
49
FriedmanM.HenikaP. R.MandrellR. E. (2002). Bactericidal activities of plant essential oils and some of their isolated constituents against campylobacter jejuni, Escherichia coli, listeria monocytogenes, and salmonella enterica. J. Food Protect.65, 1545–1560. PMID: . doi: 10.4315/0362-028x-65.10.1545
50
FuL.ZhouT.WangJ.YouL.LuY.YuL.et al. (2019). NanoFe3O4 as solid electron shuttles to accelerate acetotrophic methanogenesis by Methanosarcina barkeri. Front. Microbiol.10:388. doi: 10.3389/fmicb.2019.00388
51
GamblielH.CroteauR. (1984). Pinene cyclases I and II. Two enzymes from sage (Salvia officinalis) which catalyze stereospecific cyclizations of geranyl pyrophosphate to monoterpene olefins of opposite configuration. J. Biol. Chem.259, 740–748. PMID: .
52
GonnermanM. C.BenedictM. N.FeistA. M.MetcalfW. W.PriceN. D. (2013). Genomically and biochemically accurate metabolic reconstruction of Methanosarcina barkeri Fusaro, iMG746. Biotechnol. J.8, 1070–1079. doi: 10.1002/biot.201200266
53
GouldM. N. (1997). Cancer chemoprevention and therapy by monoterpenes. Environ. Health Persp.105, 977–979. doi: 10.2307/3433313
54
GoyalN.ZhouZ.KarimiI. A. (2016). Metabolic processes of Methanococcus maripaludis and potential applications. Microb. Cell Factories15, 1–19. doi: 10.1186/s12934-016-0500-0
55
GuanY.HikmawanT.AntunesA.NgugiD.StinglU. (2015). Diversity of methanogens and sulfate-reducing bacteria in the interfaces of five deep-sea anoxic brines of the Red Sea. Res. Microbiol.166, 688–699. doi: 10.1016/j.resmic.2015.07.002
56
GussA. M.KulkarniG.MetcalfW. W. (2009). Differences in hydrogenase gene expression between Methanosarcina acetivorans and Methanosarcina barkeri. J. Bacteriol.191, 2826–2833. doi: 10.1128/JB.00563-08
57
GussA. M.RotherM.ZhangJ. K.KulkkarniG.MetcalfW. W. (2008). New methods for tightly regulated gene expression and highly efficient chromosomal integration of cloned genes for Methanosarcina species. Archaea2, 193–203. PMID: ; PMID: PMC2685592. doi: 10.1155/2008/534081
58
HaoW.ScharffeD.CrutzenP.SanhuezaE. (1988). Production of N2O, CH4, and CO2 from soils in the tropical savanna during the dry season. J. Atmos. Chem.7, 93–105. doi: 10.1007/BF00048256
59
HaqK.JiaY.KrishnanL. (2016). Archaeal lipid vaccine adjuvants for induction of cell-mediated immunity. Expert Rev. Vaccines15, 1557–1566. doi: 10.1080/14760584.2016.1195265
60
HildenbrandC.StockT.LangeC.RotherM.SoppaJ. (2011). Genome copy numbers and gene conversion in methanogenic archaea. J. Bacteriol.193, 734–743. doi: 10.1128/JB.01016-10
61
HolmesD. E.GiloteauxL.OrellanaR.WilliamsK. H.RobbinsM. J.LovleyD. R. (2014). Methane production from protozoan endosymbionts following stimulation of microbial metabolism within subsurface sediments. Front. Microbiol.5:366. doi: 10.3389/fmicb.2014.00366
62
HorneA. J.LessnerD. J. (2013). Assessment of the oxidant tolerance of Methanosarcina acetivorans. FEMS Microbiol. Lett.343, 13–19. doi: 10.1111/1574-6968.12115
63
HuangZ.SednekC.UrynowiczM. A.GuoH.WangQ.FallgrenP.et al. (2017). Low carbon renewable natural gas production from coalbeds and implications for carbon capture and storage. Nat. Commun.8, 1–11. doi: 10.1038/s41467-017-00611-7
64
IijimaY.GangD. R.FridmanE.LewinsohnE.PicherskyE. (2004). Characterization of geraniol synthase from the peltate glands of sweet basil. Plant Physiol.134, 370–379. PMID: ; PMID: PMC316316. doi: 10.1104/pp.103.032946
65
InnardN. (2018). Developing Genetic Tools in Methanococcus maripaludis, Heslington: University of York. Available at: https://etheses.whiterose.ac.uk/20486/
66
JacquemetA.BarbeauJ.LemiègreL.BenvegnuT. (2009). Archaeal tetraether bipolar lipids: structures, functions and applications. Biochimie91, 711–717. doi: 10.1016/j.biochi.2009.01.006
67
JainS.CaforioA.DriessenA. J. (2014). Biosynthesis of archaeal membrane ether lipids. Front. Microbiol.5:641. doi: 10.3389/fmicb.2014.00641
68
Jasso-ChávezR.Santiago-MartínezM. G.Lira-SilvaE.PinedaE.Zepeda-RodríguezA.et al. (2015). Air-adapted Methanosarcina acetivorans shows high methane production and develops resistance against oxygen stress. PLoS One10:e0117331. PMID: ; PMCID: PMC4338226. doi: 10.1371/journal.pone.0117331
69
JenningsM. E.SchaffC. W.HorneA. J.LessnerF. H.LessnerD. J. (2014). Expression of a bacterial catalase in a strictly anaerobic methanogen significantly increases tolerance to hydrogen peroxide but not oxygen. Microbiology160, 270–278. doi: 10.1099/mic.0.070763-0
70
JinD.KangK.WangH.WangZ.XueB.WangL.et al. (2017). Effects of dietary supplementation of active dried yeast on fecal methanogenic archaea diversity in dairy cows. Anaerobe44, 78–86. PMID: . doi: 10.1016/j.anaerobe.2017.02.007
71
JohnsonK. A.JohnsonD. E. (1995). Methane emissions from cattle. J. Anim. Sci.73, 2483–2492. doi: 10.2527/1995.7382483x
72
JonesD.HeadI.GrayN.AdamsJ.RowanA.AitkenC.et al. (2008). Crude-oil biodegradation via methanogenesis in subsurface petroleum reservoirs. Nature451, 176–180. doi: 10.1038/nature06484
73
JonesW. J.NagleD.WhitmanW. B. (1987). Methanogens and the diversity of archaebacteria. Microbiol. Rev.51, 135–177. PMID: ; PMID: PMC373095. doi: 10.1128/mr.51.1.135-177.1987
74
JonesW. J.PaynterM.GuptaR. (1983). Characterization of Methanococcus maripaludis sp. nov., a new methanogen isolated from salt marsh sediment. Arch. Microbiol.135, 91–97. doi: 10.1007/BF00425213
75
KnittelK.BoetiusA. (2009). Anaerobic oxidation of methane: progress with an unknown process. Annu. Rev. Microbiol.63, 311–334. PMID: . doi: 10.1146/annurev.micro.61.080706.093130
76
KogaY. (2012). Thermal adaptation of the archaeal and bacterial lipid membranes. Archaea2012, 1–6. doi: 10.1155/2012/789652
77
KogaY.MoriiH. (2007). Biosynthesis of ether-type polar lipids in archaea and evolutionary considerations. Microbiol. Mol. Biol. Rev.71, 97–120. PMID: ; PMCID: PMC1847378. doi: 10.1128/MMBR.00033-06
78
KohlerP. R.MetcalfW. W. (2012). Genetic manipulation of Methanosarcina spp. Front. Microbiol.3:259. PMID: ; PMCID: PMC3403347. doi: 10.3389/fmicb.2012.00259
79
KulkarniG.KridelbaughD. M.GussA. M.MetcalfW. W. (2009). Hydrogen is a preferred intermediate in the energy-conserving electron transport chain of Methanosarcina barkeri. Proc. Natl. Acad. Sci. U. S. A.106, 15915–15920. doi: 10.1073/pnas.0905914106
80
KurthJ. M.Op den CampH. J.WelteC. U. (2020). Several ways one goal—methanogenesis from unconventional substrates. Appl. Microbiol. Biotechnol.104, 6839–6854. PMID: ; PMCID: PMC7374477. doi: 10.1007/s00253-020-10724-7
81
LandiA.LawJ.HockmanD.LoganM.CrawfordK.ChenC.et al. (2017). Superior immunogenicity of HCV envelope glycoproteins when adjuvanted with cyclic-di-AMP, a STING activator or archaeosomes. Vaccine35, 6949–6956. doi: 10.1016/j.vaccine.2017.10.072
82
LangeB. M.RujanT.MartinW.CroteauR. (2000). Isoprenoid biosynthesis: the evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. U. S. A.97, 13172–13177. PMID: ; PMCID: PMC27197. doi: 10.1073/pnas.240454797
83
Laso-PérezR.HahnC.van VlietD. M.TegetmeyerH. E.SchubotzF.SmitN. T.et al. (2019). Anaerobic degradation of non-methane alkanes by “Candidatus Methanoliparia” in hydrocarbon seeps of the Gulf of Mexico. MBio10, e01814–e01819. PMID: ; PMCID: PMC6703427. doi: 10.1128/mBio.01814-19
84
Le MerJ.RogerP. (2001). Production, oxidation, emission and consumption of methane by soils: a review. Eur. J. Soil Biol.37, 25–50. doi: 10.1016/S1164-5563(01)01067-6
85
LemaireO. N.WagnerT. (2022). A structural view of alkyl-coenzyme M reductases, the first step of alkane anaerobic oxidation catalyzed by archaea. Biochemistry61, 805–821. doi: 10.1021/acs.biochem.2c00135
86
LericheG.CifelliJ. L.SibucaoK. C.PattersonJ. P.KoyanagiT.GianneschiN. C.et al. (2017). Characterization of drug encapsulation and retention in archaea-inspired tetraether liposomes. Org. Biomol. Chem.15, 2157–2162. doi: 10.1039/C6OB02832B
87
LessnerD. J.LhuL.WahalC. S.FerryJ. G. (2010). An engineered methanogenic pathway derived from the domains bacteria and archaea. MBio1, e00243–e00210. PMID: ; PMCID: PMC2975365. doi: 10.1128/mBio.00243-10
88
LoewR.HeinzN.HampfM.BujardH.GossenM. (2010). Improved Tet-responsive promoters with minimized background expression. BMC Biotechnol.10, 1–13. PMID: ; PMCID: PMC3002914. doi: 10.1186/1472-6750-10-81
89
LongB. H.CarboniJ. M.WassermanA. J.CornellL. A.CasazzaA. M.JensenP. R.et al. (1998). Eleutherobin, a novel cytotoxic agent that induces tubulin polymerization, is similar to paclitaxel (Taxol (R)). Cancer Res.58, 1111–1115. PMID: .
90
LongF.WangL.LupaB.WhitmanW. B. (2017). A flexible system for cultivation of Methanococcus and other formate-utilizing methanogens. Archaea2017, 1–12. doi: 10.1155/2017/7046026
91
López MuñozM. M.SchönheitP.MetcalfW. W. (2015). Genetic, genomic, and transcriptomic studies of pyruvate metabolism in Methanosarcina barkeri fusaro. J. Bacteriol.197, 3592–3600. doi: 10.1128/JB.00551-15
92
LuoG.AngelidakiI. (2012). Integrated biogas upgrading and hydrogen utilization in an anaerobic reactor containing enriched hydrogenotrophic methanogenic culture. Biotechnol. Bioeng.109, 2729–2736. doi: 10.1002/bit.24557
93
MacarioE. C. D.MacarioA. J. (2003). Molecular biology of stress genes in methanogens: potential for bioreactor technology. BiomethanationI, 95–150. PMID: . doi: 10.1007/3-540-45839-5_4
94
MarketsM. A. (2021). Flavors & Fragrances Market by ingredients (natural, synthetic), end use (beverage, Savory & Snacks, bakery, dairy products, confectionery, consumer products, fine fragrances), and region (Asia Pacific, North America, Europe) – global forecast to 2026. Available at: https://www.marketsandmarkets.com/Market-Reports/flavors-fragrance-market-175163912.html
95
MartinW. F.SousaF. L. (2016). Early microbial evolution: the age of anaerobes. Cold Spring Harb. Perspect. Biol.8:a018127. doi: 10.1101/cshperspect.a018127
96
MatschiavelliN.OelgeschlägerE.CocchiararoB.FinkeJ.RotherM. (2012). Function and regulation of isoforms of carbon monoxide dehydrogenase/acetyl coenzyme a synthase in Methanosarcina acetivorans. J. Bacteriol.194, 5377–5387. doi: 10.1128/JB.00881-12
97
MayumiD.MochimaruH.TamakiH.YamamotoK.YoshiokaH.SuzukiY.et al. (2016). Methane production from coal by a single methanogen. Science354, 222–225. doi: 10.1126/science.aaf8821
98
McAnultyM. J.PoosarlaV.KimK.-Y.Jasso-ChávezR.LoganB. E.WoodT. K. (2017). Electricity from methane by reversing methanogenesis. Nat. Commun.8, 1–8. doi: 10.1038/ncomms15419
99
McGarveyD. J.CroteauR. (1995). Terpenoid metabolism. Plant Cell7, 1015–1026. doi: 10.1105/tpc.7.7.1015
100
McGenityT. J. (2010). “Methanogens and methanogenesis in hypersaline environments” in Handbook of Hydrocarbon and Lipid Microbiology. ed. TimmisK. N. Berlin, Heidelberg: Springer, 665–680.
101
McGlynnS. E.ChadwickG. L.KempesC. P.OrphanV. J. (2015). Single cell activity reveals direct electron transfer in methanotrophic consortia. Nature526, 531–535. doi: 10.1038/nature15512
102
MeigsT. E.SimoniR. D. (1997). Farnesol as a regulator of HMG-CoA reductase degradation: characterization and role of farnesyl pyrophosphatase. Arch. Biochem. Biophys.345, 1–9. doi: 10.1006/abbi.1997.0200
103
MetcalfW. W.ZhangJ. K.ApolinarioE.SowersK. R.WolfeR. S. (1997). A genetic system for archaea of the genus Methanosarcina: liposome-mediated transformation and construction of shuttle vectors. Proc. Natl. Acad. Sci. U. S. A.94, 2626–2631. PMID: ; PMCID: PMC20139. doi: 10.1073/pnas.94.6.2626
104
MeyerdierksA.KubeM.KostadinovI.TeelingH.GlöcknerF. O.ReinhardtR.et al. (2010). Metagenome and mRNA expression analyses of anaerobic methanotrophic archaea of the ANME-1 group. Environ. Microbiol.12, 422–439. doi: 10.1111/j.1462-2920.2009.02083.x
105
MoranJ. J.HouseC. H.FreemanK. H.FerryJ. G. (2005). Trace methane oxidation studied in several Euryarchaeota under diverse conditions. Archaea1, 303–309. PMID: ; PMCID: PMC2685550. doi: 10.1155/2005/650670
106
MosierN. S.LadischM. R. (2011). Modern Biotechnology: Connecting Innovations in Microbiology and Biochemistry to Engineering Fundamentals. Hoboken: John Wiley & Sons, ISBN: 1118210204.
107
NayakD. D.MetcalfW. W. (2017). Cas9-mediated genome editing in the methanogenic archaeon Methanosarcina acetivorans. Proc. Natl. Acad. Sci.114, 2976–2981. doi: 10.1073/pnas.1618596114
108
NeillT.JuddD.VeithE.RousarD. (2009). Practical uses of liquid methane in rocket engine applications. Acta Astronaut.65, 696–705. doi: 10.1016/j.actaastro.2009.01.052
109
NewmanD. J.CraggG. M. (2004). Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod.67, 1216–1238. doi: 10.1021/np040031y
110
PaduchR.Kandefer-SzerszenM.TrytekM.FiedurekJ. (2007). Terpenes: substances useful in human healthcare. Arch. Immunol. Ther. Ex.55, 315–327. doi: 10.1007/s00005-007-0039-1
111
ParkM.-O.IkenagaH.WatanabeK. (2007). Phage diversity in a methanogenic digester. Microb. Ecol.53, 98–103. PMID: . doi: 10.1007/s00248-006-9053-9
112
PatelG. B.ChenW. (2010). Archaeal lipid mucosal vaccine adjuvant and delivery system. Expert Rev. Vaccines9, 431–440. PMID: . doi: 10.1586/erv.10.34
113
PatilU. K.MuskanK. (2009). Essentials of Biotechnology. Bengaluru: IK International Pvt Ltd. ISBN: 9380026528.
114
PauloL. M.StamsA. J.SousaD. Z. (2015). Methanogens, sulphate and heavy metals: a complex system. Rev. Environ. Sci. Biotechnol.14, 537–553. doi: 10.1007/s11157-015-9387-1
115
PechousS. W.WhitakerB. D. (2004). Cloning and functional expression of an (E, E)-alpha-farnesene synthase cDNA from peel tissue of apple fruit. Planta219, 84–94. doi: 10.1007/s00425-003-1191-4
116
PicherskyE.RagusoR. A.LewinsohnE.CroteauR. (1994). Floral Scent Production in Clarkia (Onagraceae) (I. Localization and Developmental Modulation of Monoterpene Emission and Linalool Synthase Activity). Plant Physiol.106, 1533–1540. doi: 10.1104/pp.106.4.1533
117
PritchettM. A.ZhangJ. K.MetcalfW. W. (2004). Development of a markerless genetic exchange method for Methanosarcina acetivorans C2A and its use in construction of new genetic tools for methanogenic archaea. Appl. Environ. Microbiol.70, 1425–1433. doi: 10.1128/AEM.70.3.1425-1433.2004
118
RagabA.ShawD. R.KaturiK. P.SaikalyP. E. (2020). Effects of set cathode potentials on microbial electrosynthesis system performance and biocathode methanogen function at a metatranscriptional level. Sci. Rep.10, 1–15. doi: 10.1038/s41598-020-76229-5
119
Rajilić-StojanovićM.SmidtH.De VosW. M. (2007). Diversity of the human gastrointestinal tract microbiota revisited. Environ. Microbiol.9, 2125–2136. doi: 10.1111/j.1462-2920.2007.01369.x
120
RiedingerN.FormoloM. J.LyonsT. W.HenkelS.BeckA.KastenS. (2014). An inorganic geochemical argument for coupled anaerobic oxidation of methane and iron reduction in marine sediments. Geobiology12, 172–181. doi: 10.1111/gbi.12077
121
Rodriguez-ConcepcionM.BoronatA. (2002). Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol.130, 1079–1089. doi: 10.1104/pp.007138
122
RoslevP.KingG. M. (1995). Aerobic and anaerobic starvation metabolism in methanotrophic bacteria. Appl. Environ. Microbiol.61, 1563–1570. PMID: ; PMCID: PMC1388422. doi: 10.1128/aem.61.4.1563-1570.1995
123
RotherM.MetcalfW. W. (2004). Anaerobic growth of Methanosarcina acetivorans C2A on carbon monoxide: an unusual way of life for a methanogenic archaeon. Proc. Natl. Acad. Sci. U. S. A.101, 16929–16934. doi: 10.1073/pnas.0407486101
124
RouviereP. E.WolfeR. (1988). Novel biochemistry of methanogenesis. J. Biol. Chem.263, 7913–7916. PMID: .
125
SarmientoF. B.LeighJ. A.WhitmanW. B. (2011). Methods in EnzymologyAmsterdam: Elsevier. PMID: . doi: 10.1016/B978-0-12-385112-3.00003-2
126
SchellerS.GoenrichM.BoecherR.ThauerR. K.JaunB. (2010). The key nickel enzyme of methanogenesis catalyses the anaerobic oxidation of methane. Nature465, 606–608. doi: 10.1038/nature09015
127
SchiraldiC.GiulianoM.De RosaM. (2002). Perspectives on biotechnological applications of archaea. Archaea1, 75–86. doi: 10.1155/2002/436561
128
SchlegelK.WelteC.DeppenmeierU.MullerV. (2012). Electron transport during aceticlastic methanogenesis by Methanosarcina acetivorans involves a sodium-translocating Rnf complex. FEBS J.279, 4444–4452. doi: 10.1111/febs.12031
129
SchützH.SeilerW.ConradR. (1989). Processes involved in formation and emission of methane in rice paddies. Biogeochemistry7, 33–53. doi: 10.1007/BF00000896
130
SgadariC.ToschiE.PalladinoC.BarillariG.CarleiD.CeresetoA.et al. (2000). Mechanism of paclitaxel activity in Kaposi's sarcoma. J. Immunol.165, 509–517. PMID: . doi: 10.4049/jimmunol.165.1.509
131
Sharon-AsaL.ShalitM.FrydmanA.BarE.HollandD.OrE.et al. (2003). Citrus fruit flavor and aroma biosynthesis: isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J.36, 664–674. doi: 10.1046/j.1365-313x.2003.01910.x
132
ShawI.GrahamM. (1987). Mesna—a short review. Cancer Treat. Rev.14, 67–86. PMID: . doi: 10.1016/0305-7372(87)90041-7
133
SheehanS. W. (2021). Electrochemical methane production from CO2 for orbital and interplanetary refueling. Iscience24:102230. doi: 10.1016/j.isci.2021.102230
134
ShimaS.KruegerM.WeinertT.DemmerU.KahntJ.ThauerR. K.et al. (2012). Structure of a methyl-coenzyme M reductase from Black Sea mats that oxidize methane anaerobically. Nature481, 98–101. PMID: . doi: 10.1038/nature10663
135
SiegertM.CichockaD.HerrmannS.GründgerF.FeisthauerS.RichnowH.-H.et al. (2011). Accelerated methanogenesis from aliphatic and aromatic hydrocarbons under iron-and sulfate-reducing conditions. FEMS Microbiol. Lett.315, 6–16. doi: 10.1111/j.1574-6968.2010.02165.x
136
SiliakusM. F.van der OostJ.KengenS. W. (2017). Adaptations of archaeal and bacterial membranes to variations in temperature, pH and pressure. Extremophiles21, 651–670. doi: 10.1007/s00792-017-0939-x
137
SillsA. K.WilliamsJ. I.TylerB. M.EpsteinD. S.SiposE. P.DavisJ. D.et al. (1998). Squalamine inhibits angiogenesis and solid tumor growth in vivo and perturbs embryonic vasculature. Cancer Res.58, 2784–2792. PMID: .
138
SilverG. M.FallR. (1991). Enzymatic synthesis of isoprene from dimethylallyl diphosphate in aspen leaf extracts. Plant Physiol.97, 1588–1591. PMID: ; PMCID: PMC1081206. doi: 10.1104/pp.97.4.1588
139
SirohiS.PandeyN.SinghB.PuniyaA. (2010). Rumen methanogens: a review. Indian J. Microbiol.50, 253–262. doi: 10.1007/s12088-010-0061-6
140
SkinnerK. A.LeathersT. D. (2004). Bacterial contaminants of fuel ethanol production. J. Ind. Microbiol. Biotechnol.31, 401–408. doi: 10.1007/s10295-004-0159-0
141
SogodogoE.DrancourtM.GrineG. (2019). Methanogens as emerging pathogens in anaerobic abscesses. Eur. J. Clin. Microbiol. Infect. Dis.38, 811–818. doi: 10.1007/s10096-019-03510-5
142
SooV. W.McAnultyM. J.TripathiA.ZhuF.ZhangL.HatzakisE.et al. (2016). Reversing methanogenesis to capture methane for liquid biofuel precursors. Microb. Cell Factories15, 1–14. doi: 10.1186/s12934-015-0397-z
143
SowersK. R.BaronS. F.FerryJ. G. (1984). Methanosarcina acetivorans sp. nov., an Acetotrophic methane-producing bacterium isolated from marine sediments. Appl. Environ. Microbiol.47, 971–978. doi: 10.1128/aem.47.5.971-978.1984
144
StummC.ZwartK. (1986). Symbiosis of protozoa with hydrogen-utilizing methanogens. Microbiol. Sci.3, 100–105. PMID: .
145
SuG.ZopfiJ.YaoH.SteinleL.NiemannH.LehmannM. F. (2020). Manganese/iron-supported sulfate-dependent anaerobic oxidation of methane by archaea in lake sediments. Limnol. Oceanogr.65, 863–875. doi: 10.1002/lno.11354
146
SubediB. P.MartinW. F.CarboneV.DuinE. C.CroninB.SauterJ.et al. (2021). Archaeal pseudomurein and bacterial murein cell wall biosynthesis share a common evolutionary ancestry. FEMS Microb.2:xtab012. doi: 10.1093/femsmc/xtab012
147
SummersZ. M.FogartyH. E.LeangC.FranksA. E.MalvankarN. S.LovleyD. R. (2010). Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science330, 1413–1415. doi: 10.1126/science.1196526
148
TakaiK.NakamuraK.TokiT.TsunogaiU.MiyazakiM.MiyazakiJ.et al. (2008). Cell proliferation at 122°C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. U. S. A.105, 10949–10954. doi: 10.1073/pnas.0712334105
149
TannerR. S.WolfeR. S. (1988). Nutritional requirements of Methanomicrobium mobile. Appl. Environ. Microbiol.54, 625–628. PMID: ; PMCID: PMC202515. doi: 10.1128/aem.54.3.625-628.1988
150
ThauerR. K. (2011). Anaerobic oxidation of methane with sulfate: on the reversibility of the reactions that are catalyzed by enzymes also involved in methanogenesis from CO2. Curr. Opin. Microbiol.14, 292–299. doi: 10.1016/j.mib.2011.03.003
151
ThauerR. K. (2012). The Wolfe cycle comes full circle. Proc. Natl. Acad. Sci. U. S. A.109, 15084–15085. doi: 10.1073/pnas.1213193109
152
ThauerR. K.KasterA.-K.SeedorfH.BuckelW.HedderichR. (2008). Methanogenic archaea: ecologically relevant differences in energy conservation. Nat. Rev. Microbiol.6, 579–591. PMID: . doi: 10.1038/nrmicro193
153
TumbulaD. L.MakulaR. A.WhitmanW. B. (1994). Transformation of Methanococcus maripaludis and identification of a Pst I-like restriction system. FEMS Microbiol. Lett.121, 309–314. doi: 10.1111/j.1574-6968.1994.tb07118.x
154
UlrichG.BowerS. (2008). Active methanogenesis and acetate utilization in Powder River Basin coals, United States. Int. J. Coal Geol.76, 25–33. doi: 10.1016/j.coal.2008.03.006
155
UrlingerS.BaronU.ThellmannM.HasanM. T.BujardH.HillenW. (2000). Exploring the sequence space for tetracycline-dependent transcriptional activators: novel mutations yield expanded range and sensitivity. Proc. Natl. Acad. Sci.97, 7963–7968. doi: 10.1073/pnas.130192197
156
van Der HoevenR. S.MonforteA. J.BreedenD.TanksleyS. D.SteffensJ. C. (2000). Genetic control and evolution of sesquiterpene biosynthesis in Lycopersicon esculentum and L. hirsutum. Plant Cell12, 2283–2294. doi: 10.1105/tpc.12.11.2283
157
VandermotenS.SantiniS.HaubrugeE.HeuzeF.FrancisF.BrasseurR.et al. (2009). Structural features conferring dual geranyl/farnesyl diphosphate synthase activity to an aphid prenyltransferase. Insect Biochem. Mol. Biol.39, 707–716. doi: 10.1016/j.ibmb.2009.08.007
158
VillanuevaL.DamsteJ. S.SchoutenS. (2014). A re-evaluation of the archaeal membrane lipid biosynthetic pathway. Nat. Rev. Microbiol.12, 438–448. doi: 10.1038/nrmicro3260
159
VillanuevaL.von MeijenfeldtF. B.WestbyeA. B.YadavS.HopmansE. C.DutilhB. E.et al. (2021). Bridging the membrane lipid divide: bacteria of the FCB group superphylum have the potential to synthesize archaeal ether lipids. ISME J.15, 168–182. PMID: ; PMCID: PMC7852524. doi: 10.1038/s41396-020-00772-2
160
VranovaE.ComanD.GruissemW. (2012). Structure and dynamics of the isoprenoid pathway network. Mol. Plant5, 318–333. doi: 10.1093/mp/sss015
161
WangC.WangC.LiuJ.XuQ.HanZ.XuX.et al. (2020). Tolerance of aceticlastic methanogenesis enhanced by magnetite under the condition of ammonia stress. ACS Sustain. Chem. Eng.8, 1417–1426. doi: 10.1021/acssuschemeng.9b05585
162
WangY.WegenerG.HouJ.WangF.XiaoX. (2019). Expanding anaerobic alkane metabolism in the domain of archaea. Nat. Microbiol.4, 595–602. doi: 10.1038/s41564-019-0364-2
163
WatanabeT.Pfeil-GardinerO.KahntJ.KochJ.ShimaS.MurphyB. J. (2021). Three-megadalton complex of methanogenic electron-bifurcating and CO2-fixing enzymes. Science373, 1151–1156. PMID: . doi: 10.1126/science.abg5550
164
WelteC.DeppenmeierU. (2011). Re-evaluation of the function of the F420 dehydrogenase in electron transport of Methanosarcina mazei. FEBS J.278, 1277–1287. doi: 10.1111/j.1742-4658.2011.08048.x
165
WhitfordM. F.TeatherR. M.ForsterR. J. (2001). Phylogenetic analysis of methanogens from the bovine rumen. BMC Microbiol.1, 1–5. PMID: ; PMCID: PMC32158. doi: 10.1186/1471-2180-1-5
166
WiseM. L.SavageT. J.KatahiraE.CroteauR. (1998). Monoterpene synthases from common sage (Salvia officinalis). cDNA isolation, characterization, and functional expression of (+)-sabinene synthase, 1,8-cineole synthase, and (+)-bornyl diphosphate synthase. J. Biol. Chem.273, 14891–14899. doi: 10.1074/jbc.273.24.14891
167
YanZ.FerryJ. G. (2018). Electron bifurcation and confurcation in methanogenesis and reverse methanogenesis. Front. Microbiol.9:1322. doi: 10.3389/fmicb.2018.01322
168
YanM.TreuL.ZhuX.TianH.BasileA.FotidisI. A.et al. (2020). Insights into ammonia adaptation and methanogenic precursor oxidation by genome-centric analysis. Environ. Sci. Technol.54, 12568–12582. doi: 10.1021/acs.est.0c01945
169
ZenglerK.RichnowH. H.Rosselló-MoraR.MichaelisW.WiddelF. (1999). Methane formation from long-chain alkanes by anaerobic microorganisms. Nature401, 266–269. doi: 10.1038/45777
170
ZhouZ.ZhangC.-J.LiuP.-F.FuL.Laso-PérezR.YangL.et al. (2022). Non-syntrophic methanogenic hydrocarbon degradation by an archaeal species. Nature601, 257–262. doi: 10.1038/s41586-021-04235-2
171
ZhuangX.KöllnerT. G.ZhaoN.LiG.JiangY.ZhuL.et al. (2012). Dynamic evolution of herbivore-induced sesquiterpene biosynthesis in sorghum and related grass crops. Plant J.69, 70–80. doi: 10.1111/j.1365-313X.2011.04771.x
Summary
Keywords
methanogens, archaea, biotechnology, methane, biofuel, terpenoid chemicals
Citation
Carr S and Buan NR (2022) Insights into the biotechnology potential of Methanosarcina. Front. Microbiol. 13:1034674. doi: 10.3389/fmicb.2022.1034674
Received
01 September 2022
Accepted
28 October 2022
Published
15 December 2022
Volume
13 - 2022
Edited by
Sabine Kleinsteuber, Helmholtz Association of German Research Centres (HZ), Germany
Reviewed by
James Ferry, The Pennsylvania State University, United States; Barny Whitman, University of Georgia, United States
Updates
Copyright
© 2022 Carr and Buan.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicole R. Buan, nbuan@unl.edu
This article was submitted to Microbial Physiology and Metabolism, a section of the journal Frontiers in Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.