 Thanaporn Eiamsam-ang
Thanaporn Eiamsam-ang Pakpoom Tadee2
Pakpoom Tadee2 Ben Pascoe
Ben Pascoe Prapas Patchanee
Prapas Patchanee- 1Graduate Program in Veterinary Science, Faculty of Veterinary Medicine, Chiang Mai University, Chiang Mai, Thailand
- 2Integrative Research Center for Veterinary Preventive Medicine, Faculty of Veterinary Medicine, Chiang Mai University, Chiang Mai, Thailand
- 3The Milner Center for Evolution, University of Bath, Bath, United Kingdom
Salmonella is a prevalent zoonotic foodborne pathogen. Swine and pork are implicated as important sources of salmonellosis in humans. In Chiang Mai and Lamphun Provinces in northern Thailand, there has been a high prevalence of Salmonella persistence for over a decade. Infection is usually with dominant S. enterica serotypes, including serotypes Rissen and 1,4,[5],12:i:-. However, other serotypes also contribute to disease but are less well characterized. The whole genome sequencing data of 43 S. enterica serotypes isolated from pork production chain through 2011–2014, were used to evaluate genetic diversity and ascertain the possible source of Salmonella contamination based on Core Genome Multilocus Sequence Typing (cgMLST) approach. The Salmonella serotypes recovered from farms and slaughterhouses were re-circulating by swine environmental contamination. Conversely, the Salmonella contamination in the retail market represents cross-contamination from multiple sources, including contaminated foodstuffs. Salmonella contamination in the pork production chain has the competency for host cell adhesion, host cell invasion, and intracellular survival, which is enough for the pathogenicity of salmonellosis. In addition, all of these isolates were multi-drug resistant Salmonella, which contained at least 10 antimicrobial resistance genes. This result indicated that these S. enterica serotypes also pose a significant public health risk. Our findings support the need for appropriate surveillance of food-animal products going to market to reduce public exposure to highly pathogenic, multi-drug resistant Salmonella. Acquiring information would motivate all stakeholders to reinforce sanitation standards throughout the pork production chain in order to eradicate Salmonella contamination and reduce the risk of salmonellosis in humans.
Introduction
Salmonella is recognized as a prevalent bacterial-zoonotic pathogen that causes acute foodborne illness in humans and is a global public health concern (Chen et al., 2013). In the United States, approximately 1.35 million people suffer from salmonellosis with 26,500 hospitalizations and 420 deaths reported annually [Center of Disease Control and Prevention (CDC), 2021]. In Southeast Asia, Salmonella has consistently contaminated the production chain for a decade, suggesting that eradication of this disease will indeed be complicated (Sinwat et al., 2016; Trongjit et al., 2017). According to the Bureau of Epidemiology of Thailand’s annual surveillance report for 2018, Salmonella is the most frequently detected pathogen causing food poisoning in hospitalized patients (Bureau of Epidemiology, 2018). Swine has been recognized as the one of the important Salmonella’s carriers, where the bacteria can multiply in the digestive tract and can be spread to other steps of the production chain via feces (Rostagno and Callaway, 2012). Furthermore, pork has been reported to be an important source of Salmonella contamination especially in the retail market which is the predisposing factor of salmonellosis in human (Patchanee et al., 2016).
Investigations have been undertaken to quantify the prevalence of Salmonella throughout the pork production chain in Chiang Mai, Thailand. These studies have reported a high prevalence of Salmonella isolated from swine farms as 30.56% (Tadee et al., 2014), and up to 41.5% in retail pork circulating in the Chiang Mai municipality area (Patchanee et al., 2016). These finding suggest that the burden of Salmonella has been continuing and substantial for a decade. All aspects of the production chain have been contaminated, from the farm-slaughterhouse-retail market, which is likely related to the levels of sanitation and hygienic at each step of production (Savall et al., 2016; Chen et al., 2019). Salmonella Rissen and Salmonella 1,4,[5],12:i:- have been the most common serotypes isolated in Chiang Mai and Lamphun provinces for more than a decade and their epidemiology investigated using molecular typing methods (Prasertsee et al., 2019; Patchanee et al., 2020). Many other serotypes are found at lower frequencies and have not been as well characterized, such as S. enterica serotypes Anatum, Panama, Stanley and Give have not been studied further.
Whole genome sequencing-based methods are now being used to identify transmission networks and assess the genetic relatedness of the clonal or closely related strains during an outbreak (Oakeson et al., 2017). Core genome MLST provides high-resolution data and reveals the precise relatedness within the species by comparing allelic variation to equivalent loci in other isolates (Pearce et al., 2020). The emergence of antimicrobial resistance (AMR) in foodborne pathogens particularly Salmonella species has posed a significant threat to public health (Ferri et al., 2017). Indiscriminate antimicrobial use in the livestock industry has been identified as a driver for multidrug-resistant (MDR) organisms, which can be spread to humans through the food chain (Van Boeckel et al., 2015; Nhung et al., 2016). In addition, the pathogenic potential of Salmonella has been linked to expression of virulence genes. Adhesion and invasion genes are essential virulence genes, which when expressed allow colonization of infecting Salmonella of the host cell (Wang et al., 2020). Expression of genes required for growth and replication within the host are then required for adequate nutrient uptake (Saha et al., 2019). Furthermore, the virulence genes that are encoded for resistance to host defense and resistance to antimicrobial peptide are the one of the important mechanisms to allowing the chronic infection of Salmonella (Kintz et al., 2015; Jajere, 2019). Whole genome sequence-based techniques can help characterize isolates according to their putative pathogenic potential, by identifying known antimicrobial resistance genes (AMRs) and virulence-related genes (Campioni et al., 2012).
In this study, we sequenced isolates from these infrequent Salmonella serotypes isolated from farms, slaughterhouses, and retail markets in Chiang Mai and Lamphun provinces between 2011 and 2014. We characterized the genetic diversity of these isolates and compared their genetic relatedness throughout the production process to assess feasibility of transmission. In addition, we characterized the antimicrobial resistance genes and virulence genes of each isolate in order to better understand the potential risk of these infrequent lineages. These data will helped assess public health risk and inform public health guidance and prevention strategies for minimize Salmonella contamination in the study area.
Materials and methods
Bacterial isolates
A total of 43 S. enterica isolates from the pork production chain in Chiang Mai and Lamphun municipality area were included in this study. These samples were collected from three steps of the pork production chain, including farms (n = 17), slaughterhouses (n = 16) and retail markets (n = 10). Classification by serotype including Salmonella enterica serotypes Stanley (n = 12), Typhimurium (n = 10), Panama (n = 6), Give (n = 6), Krefeld (n = 2), Kedougou (n = 2), Anatum (n = 1), Agona (n = 1), Lexington (n = 1), Newport (n = 1) and Yoruba (n = 1). Isolates were serotyped according to the WHO National Salmonella and Shigella Center Laboratory (NSSC) in Non-thaburi, Thailand using the slide agglutination method and serotypes were assigned according to the Kauffmann–White scheme (Brenner et al., 2000). Typing details for each isolate is shown in Table 1.
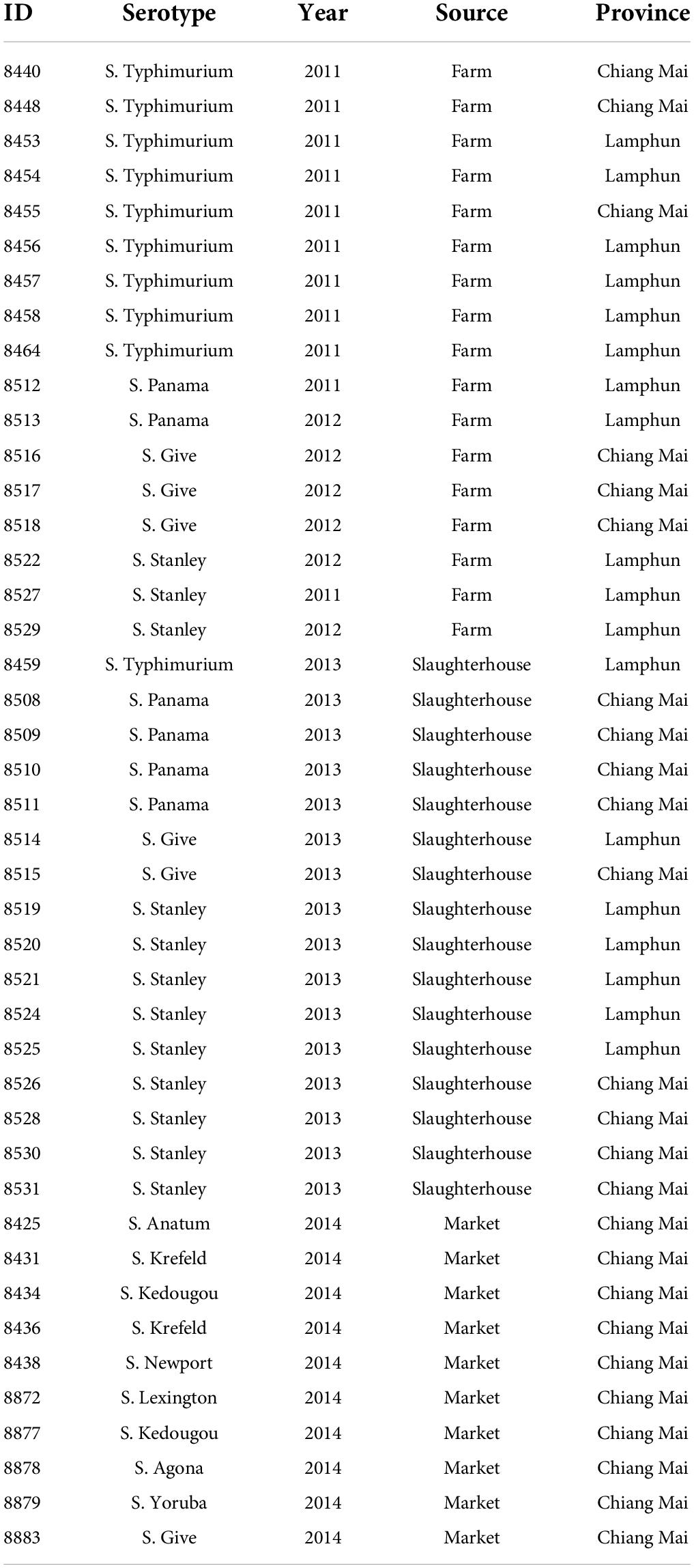
Table 1. Origin and characteristic of S. enterica isolates tested recovered from farms, slaughterhouses and retail markets during the period of 2011–2014.
Whole genome sequencing
DNA of all 43 S. enterica isolates were extracted using QIAamp DNA mini kits (Qiagen, Crawley, United Kingdom). The Nextera XT DNA Library Preparation Kit was used for the library preparation according to the manufacturer’s instructions (Illumina, Cambridge, United Kingdom). Salmonella genomes were sequenced as short reads using an Illumina MiSeq 300 bp paired-end sequencer (Illumina, Cambridge, United Kingdom). Short reads were filtered, trimmed with TRIMMOMATIC (Bolger et al., 2014), and assembled de novo with SPAdes software (version 3.8.0, using the-careful command) (Bankevich et al., 2012). The average number of contigs was 341 (range: 89–2,365) for an average total assembled sequence size of 4,918,241 bp (range: 4,169,306–5,234,359). The average N50 was 38,380 (range: 2,249–128,256) and the average GC content was 52.2% (range: 52.0–52.6). Short read data are available on the NCBI Sequence Read Archive, associated with BioProjects PRJNA573746 and PRJNA4199261.
Core genome multilocus sequence typing
Whole genome sequencing data of all 43 S. enterica isolates were uploaded to a Bacterial Isolate Genome Sequence Database (BIGSdb) and the genome comparator tool (constructed for the and provided by pubMLST)2 used to assess gene presence among the isolates. The genome comparator tool analyzes all selected isolates using the EnteroBase Salmonella database’s core genome multilocus sequence typing (cgMLST) scheme, which considers a total of 3,002 loci (Pearce et al., 2020).
Context dataset
To expand our collection, we included an additional 61 S. enterica isolates from public repositories: Enterobase Salmonella enterica WGS database3 (Achtman et al., 2020; Supplementary Table 1). These additional genomes included isolates from different source reservoirs in Thailand during the period of 2001 through 2016. Isolates were typed to Salmonella enterica serotypes Stanley (n = 8), Typhimurium (n = 21), Panama (n = 2), Give (n = 1), Anatum (n = 8), Kedougou (n = 9), Agona (n = 7) and Lexington (n = 5); from animals (2 wild animals, and 1 poultry), food (38 pork, 6 frozen seafood, 5 spices, 3 foods and 2 vegetables), and human (n = 4). A minimum spanning tree of all isolates was constructed, based on advanced cluster analysis for categorical data of allelic number for cgMLST using Bionumerics version 7.6 (Applied Maths, Ghent, Belgium).
Virulence genes and antimicrobial resistance genes investigation
We used the RASTtk algorithm (Brettin et al., 2015) to annotate whole genome sequencing data of all isolates via the PATRIC v3.6.12 annotation server using default parameters (Wattam et al., 2017)4. The Virulence Factor Database (VFDB; database version 2019) (Liu et al., 2019) was used to define the presence of known virulence genes including adhesion effector, invasion effectors, intracellular survival effectors and toxin-producing genes. Antimicrobial resistance gene presence was explored using the Comprehensive Antibiotic Resistance Database (CARD; database version 2020) (Alcock et al., 2020). The antimicrobial resistance genes (AMR genes) related to the expression of aminoglycoside, beta-lactam, trimethoprim, fluoroquinolone, fosfomycin, macrolide, macrolide-lincosamide-streptogramin B, peptide, phenicol, sulfonamide, and tetracycline were investigated. The dendrogram of the virulence genes and antimicrobial resistance genes investigation were constructed using the unweighted pair group method with arithmetic mean (UPGMA) algorithms according to the cluster analysis of categorical values of genes concluded in this study.
Results
S. enterica serotypes differ in their ability survive through the pork production chain
From the minimum spanning tree analysis, all S. enterica isolates tested were divided into 4 major clusters with 9 additional singleton isolates (Figure 1). According to the source of origin, isolates from farms and slaughterhouses were grouped together into the similar clusters. Conversely, most isolates recovered from retail markets (9/10) did not cluster with farm or slaughterhouse isolates. There was a single cluster that was comprised of isolates from all three sources of S. enterica isolates, supporting persistence of this serotype (S. enterica serotype Give) from the swine farm to slaughter and contamination of retail pork products (Supplementary Figure 1). This cluster was comprised solely of isolates collected in Chiang Mai and Lamphun province during the period of 2012–2014.
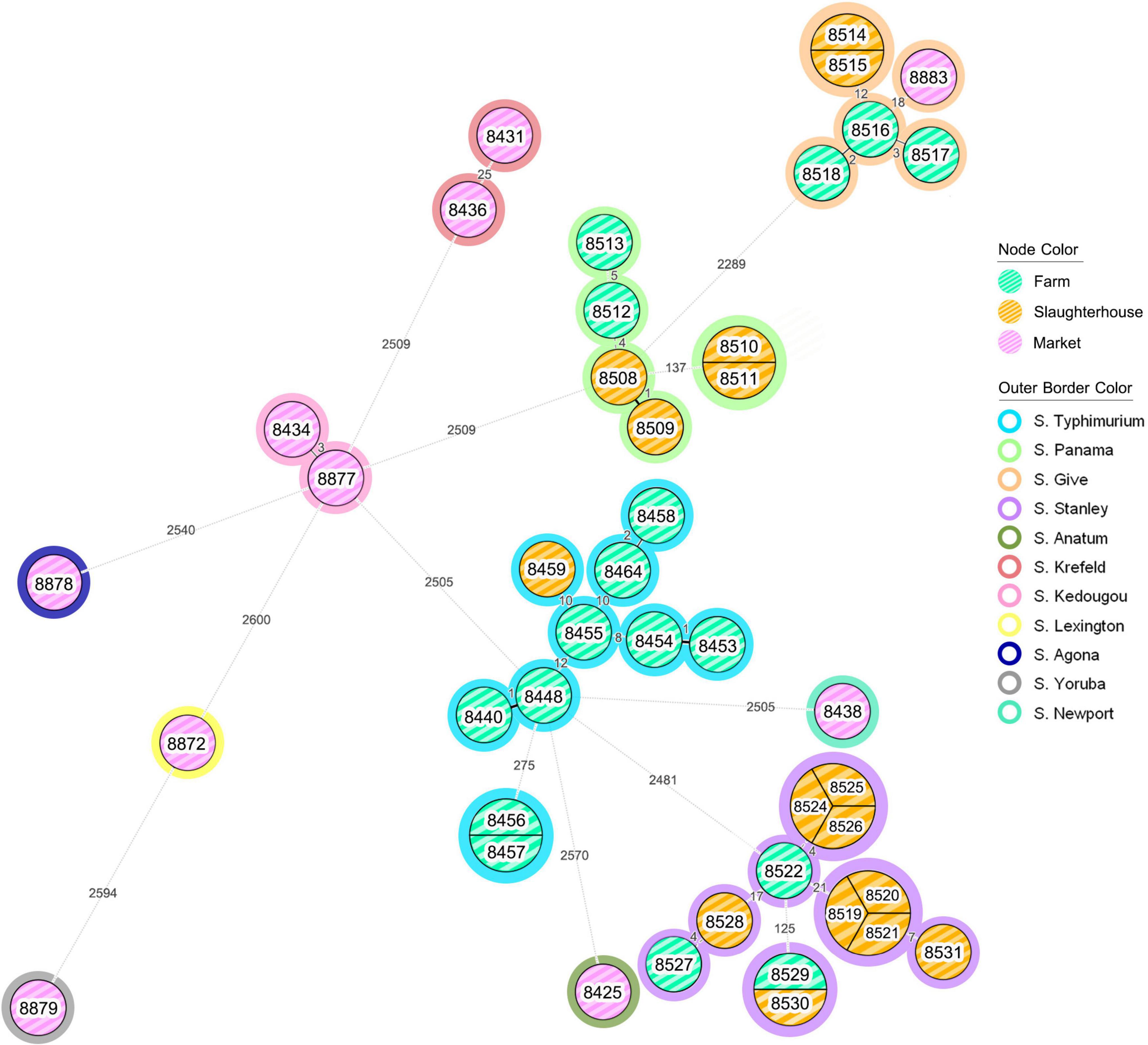
Figure 1. The minimum spanning tree (MST) analysis of Salmonella isolates recovered from the pork production chain. Each isolate was grouped according to the loci different of EnteroBase Salmonella database’s cgMLST scheme, which considers a total of 3,002 loci (Pearce et al., 2020). The number on the branch represent the number of loci different between each isolate. Node color coding: green color, yellow color and pink color represent the Salmonella isolates recovered from farm, slaughterhouse and retail market, respectively. Outer border color represents each serotype of Salmonella isolates.
Some Salmonella isolates shared the same cgMLST profiles, including serotype Typhimurium isolates ID 8456 and 8457—both recovered from farms in Lamphun province in the period of 2011; six serotype Stanley isolates (IDs: 8519, 8520, 8521, 8524, 8525, and 8526) recovered from slaughterhouses in Chiang Mai and Lamphun during the period of 2013; and two serotype Panama isolates (IDs: 8510 and 8511) from Chiang Mai which were recovered from slaughterhouse in 2013 (Figure 1). In addition, some clonal isolates were able to persist between different production steps and time periods, including two serotype Stanley isolates (IDs: 8529 and 8530): collected from a farm in Lamphun in 2012, and a slaughterhouse in Chiang Mai in 2013, respectively. Two others clonal serotype Give isolates were collected from slaughterhouses in different provincial areas during 2013 (IDs: 8514 and 8515; Figure 1).
Notwithstanding S. enterica serotype Rissen and the 1,4,[5],12:i:-, which are the most common serotypes identified in the northern Thai pork production chain, S. enterica serotypes Typhimurium, Stanley and Panama were the predominant serotypes recovered from farms and slaughterhouses. These Salmonella isolates were found in Chiang Mai and Lamphun Province during the period of 2011 through 2013. Other serotypes, including S. enterica serotypes Agona, Anatum, Krefeld, Kedougou, Lexington, Newport, and Yoruba were not grouped into any clusters and were isolated from sources in the retail markets in the Chiang Mai municipality area. S. enterica serotype Give was the only serotype that was found in all 3 different steps during 2012–2014 (Supplementary Figure 1).
Not all high-risk S. enterica isolates collected in the retail markets were from pork
To better understand the genetic relatedness of isolates in our collection, we constructed a minimum spanning tree based on cgMLST profiles of our collected isolates (n = 43), compared with all publicly available genomes from Thailand (n = 61; Supplementary Table 1). In total, 104 S. enterica isolates were clustered according to their cgMLST profiles by their source of origin (Figure 2), and there was a distinction in isolates from the farms and slaughterhouses and those collected in the retail markets. Isolates collected from the retail markets demonstrated greater diversity in cgMLST profiles (6 cgMLST clusters) and serotypes (7 Serotypes). Four cgMLST clusters and 4 serotypes overlapped between the farms and slaughterhouses (Supplementary Figure 2). Many of the isolates collected in Chiang Mai municipality markets were of unique cgMLST profiles and serotypes that were not present in any of our pork production samples, instead grouping with clusters from other Thai food products, including spices, food, frozen seafood and pork. Although pork products are a frequent source of salmonellosis infection, they are not the only high-risk food product available in the retail markets. The limited number of clinical S. enterica isolates that were publicly available (n = 4) clustered (same/similar cgMLST profile) predominantly with isolates identified in the pre-harvest steps of pork production (n = 3), with only a single isolate clustering with isolates identified at retail markets (Figure 2). Given the low number of isolates compared it’s difficult to draw robust conclusions but does suggest that isolates from the pork production industry are able to persist and pose a public health risk.
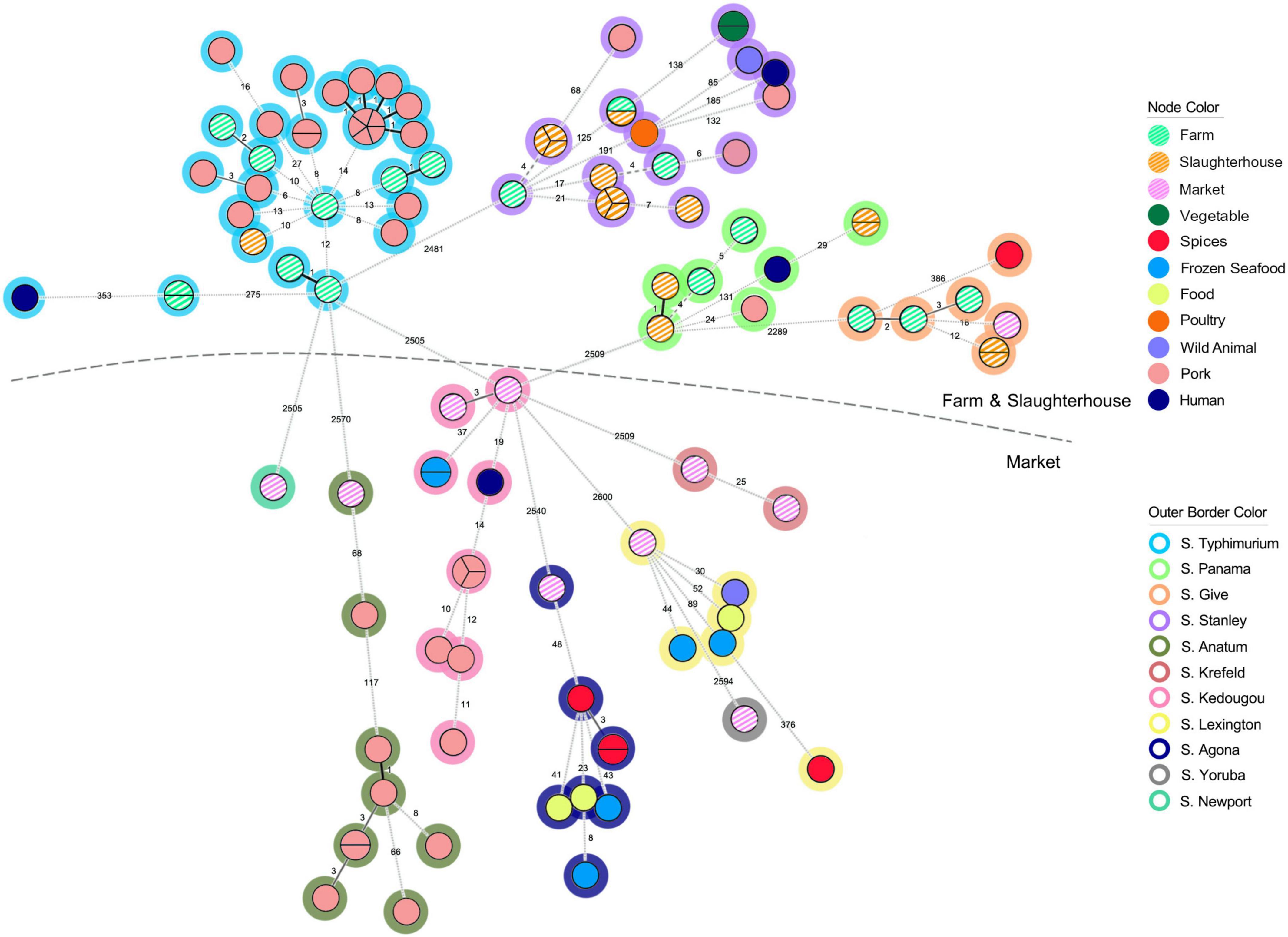
Figure 2. The minimum spanning tree (MST) analysis of 43 Salmonella isolates recovered from pork production chain (striped nodes) and additional 61 Salmonella isolates circulating in Thailand. Each isolate was grouped according to the loci different of EnteroBase Salmonella database’s cgMLST scheme, which considers a total of 3,002 loci (Pearce et al., 2020). The number on the branch represent the number of loci different between each isolate. Node color coding were representing the sources of the Salmonella isolates. Outer border color represents each serotype of Salmonella isolates.
Virulence gene profiling of S. enterica from the pork production chain
Known virulence genes were identified in all isolates through nucleotide comparisons with the VFDB. All isolates carried the virulence genes encoding for host cell adhesion (csg, fim, mis, sin), host cell invasion (che, flg, fli, inv, mot, omp, org, prg, sic, sif, sip, slr, sop, spa, spt, ssa, ssc, sse) and intracellular survival (ent, fep, gmh, iro, mgt, kds, mig). The virulence genes were harbored in the Salmonella isolates regardless of serotype, production steps, year, and geographical area. However, serotype Give isolates seemed to contain alternative host adhesion genes (fae and shd) in place of the more common lpf and rat genes. We also identified cdt genes in these serotypes Give isolates (and serotype Panama), which are typically associated with S. enterica serotype Typhi and encode for toxin production. This is particularly alarming as serotype Give isolates were able to persist through the pork production chain, and therefore these highly virulent genes were identified in isolates from farms, slaughterhouses, and retail markets (Figure 3).
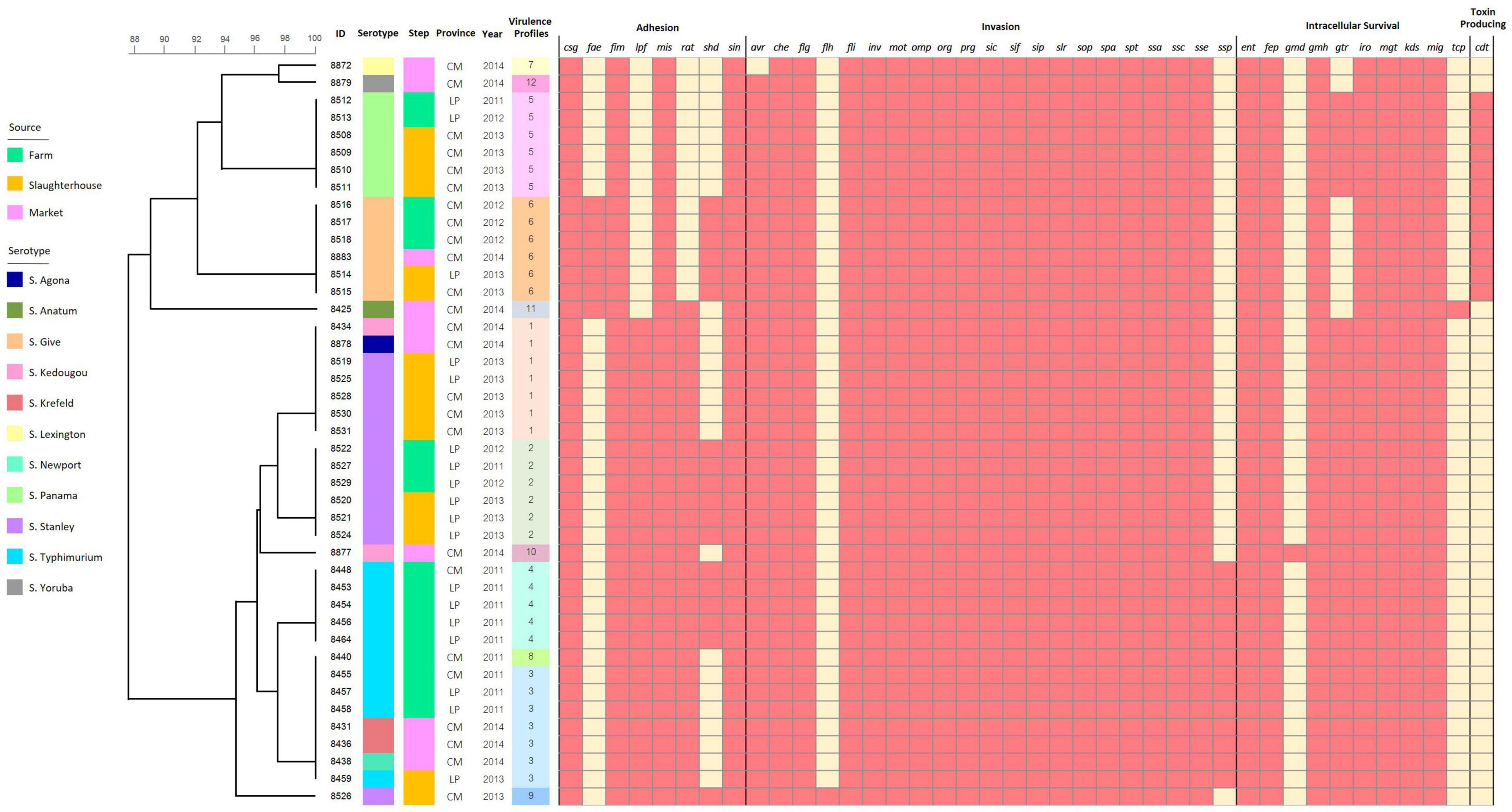
Figure 3. Binary heatmap analysis of virulence genes harbored in the Salmonella isolates recovered from pork production chain circulating in Chiang Mai and Lamphun municipality area during the period 2011–2014.
Isolates were characterized according to presence of each of the 40 identified virulence genes and classified into 12 virulence profiles. Profiles 1 and 3 (16.27%) were the most frequently encoded profile, followed by profiles 2, 5 and 6 with 13.95% of frequency. Several genes were found only in specific sources, resulting in unique virulence profiles, including virulence profiles 9 (with flh genes isolated from the slaughterhouse), profiles 10 and 11 (with gmd and tcp genes isolated from the retail markets, respectively). There was also diversity in virulence profiles among serotypes and the different steps of the production chain. In another words, there are the less concordance between serotyping results and virulence profiles (Figure 3).
An intersection analysis of virulence genes demonstrated that 37 virulence genes were shared among the three steps of pork production. Many Salmonella isolates collected from the retail markets had the most virulence genes (39/40 virulence genes), followed by Salmonella isolates recovered from slaughterhouses and farms (38 and 37 virulence genes were identified, respectively) (Supplementary Figure 3).
Widespread multidrug resistance in isolates from all steps of the pork production chain
According to the heatmap analysis, all the Salmonella isolates in this study carried at least one antimicrobial resistance gene (ARG), and all of them were multi-drug resistant Salmonella—demonstrating putative resistance to three or more antimicrobial classes. Similar to the distribution of virulence genes, ARGs were predominantly lineage dependent—with isolates from the same serotype sharing similar ARG content, even across different isolation sources, time and geographical area. In total, ten antimicrobial resistance genes were harbored in all Salmonella isolates, including acrD, gyrA, mfd, parC, parE, glpT, murA, bacA, pmrC, and pmrE, which contribute toward aminoglycoside, fluoroquinolone, fosfomycin, and peptide resistance, while CTX-M-14 and erm (42), which encoded for beta-lactam and macrolide-lincosamide-streptogramin B resistance, was not found in this study (Figure 4).
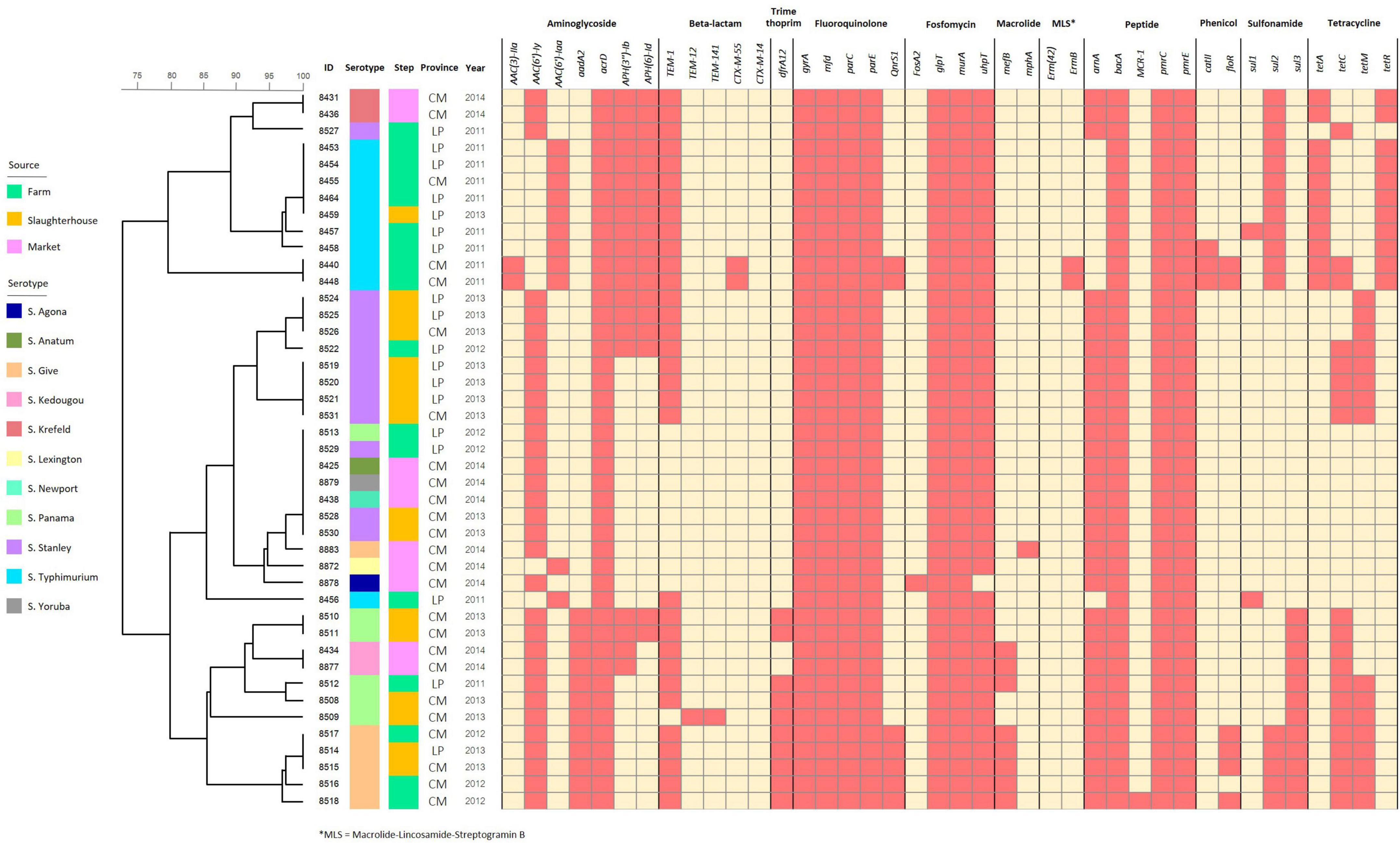
Figure 4. Binary heatmap analysis of antimicrobial resistance genes harbored in the Salmonella isolates recovered from pork production chain circulating in Chiang Mai and Lamphun municipality area during the period 2011–2014.
According to intersection analysis, twenty-four antimicrobial resistance genes were shared throughout all three pork production steps. Additionally, four antimicrobial resistance genes (drfA12, QnrS1, florR and tetM) were identified in isolates sampled from farms and slaughterhouses, which are linked to trimethoprim, fluoroquinolone, phenicol and tetracycline, respectively. The Salmonella isolates recovered from farms harbored the highest number of antimicrobial resistance genes (34/40 ARGs), followed by the Salmonella isolates recovered from slaughterhouses and retail markets (30 and 26 ARGs identified, respectively). Some ARGs were identified in specific sources of Salmonella isolates, including AAC (3)-IIa, CTX-M-55, ErmB, MCR-1, catII and sul1 genes in farm isolates, while TEM-12 and TEM-141 genes were only found in slaughterhouses. Furthermore, FosA2 and mphA were only found in Salmonella isolates that were on the retail markets (Supplementary Figure 4).
Discussion
In Chiang Mai and Lamphun Province, the Salmonella contamination in the pork production chain has been burden for over a decade. From the study of Tadee et al. (2015) and Patchanee et al. (2016) were collected the Salmonella contamination from farm, slaughterhouse and market in this study areas include serotypes Rissen, 1,4,[5],12:i:-, Stanley, Typhimurium, Panama, Give, Krefeld, Kedougou, Anatum, Agona, Lexington, Newport, and Yoruba. The most common Salmonella serotypes implicated in widespread contamination of the pork production chain in northern Thailand are serotype 1,4,[5],12:i:- and Rissen. Previous genomics studies have focused on these common serotypes and represent the relationship of farm-slaughterhouse-retail transmission, with infrequent serotypes overlooked (Prasertsee et al., 2019; Patchanee et al., 2020). In this study we focus on these infrequent serotypes to investigate their genetic diversity and virulence potential. Isolates originating from farm and slaughterhouse were closely related, with isolates from serotypes Typhimurium, Panama, and Stanley collected from both production steps. This is consistent with previous studies suggesting that these serotypes are common at the pre-harvest level (Sanguankiat et al., 2010; Niyomdecha et al., 2016). Together, these findings support the feasibility of Salmonella spreading from the farms to slaughterhouses via live animals (Rostagno and Callaway, 2012; Savall et al., 2016).
S. enterica serotype Give was the only serotype found at all three steps of the pork production chain. This serotype has previously been found in Thailand from diverse sources of food-animal production, including poultry, pork and other food products, such as chili powder (Wang et al., 2015; Phongaran et al., 2019). Another study implicated this serotype as the causative agent of salmonellosis with a splenic abscess in a male patient who had traveled to southern Thailand and consumed raw minced pork (Girardin et al., 2006). Despite this, many isolates found primarily on the farms and in the slaughterhouses apparently pose no direct impact on consumers, i.e., they are not able to survive (or out compete other strains) to contaminate pork products sold at market. However, there is evidence that S. enterica serotypes found predominantly at the pre-harvest level can cause infection in workers at the operational level. Estimates suggest that as many as 43% of workers are colonized by Salmonella, compared to an overall prevalence of Salmonella contamination in farmed pigs of 52% (Sringam et al., 2017). This suggests that transmission between pigs and humans is very common in the farm environment, with S. enterica serotype Typhimurium implicated as the primary infecting agent (Punpanich et al., 2012). S. enterica serotypes Stanley and Panama were also among the most common serotypes isolated from farms and slaughterhouses and have been recovered from the stool samples of salmonellosis patients in other studies (Pulford et al., 2019).
The S. enterica isolates that we collected from retail markets were more diverse and represented several Salmonella serotypes. Of the 10 samples we collected from the retail markets, we identified 8 different serotypes. There was little overlap between serotypes found in retail market isolates and those sampled on the farms or slaughterhouses. It is likely that this diversity, in both phenotypic and genotypic characteristics are due to the wider sources of contamination of retail products than the pork production industry (Chen et al., 2019). We identified a single isolate from the retail market samples that was from S. enterica serotype Anatum, which is typically one of the most common serotypes found in pork products in Thailand (Padungtod and Kaneene, 2006)—sometimes considered the 3rd most common serotype isolated from pork in Asia (Ferrari et al., 2019). Other serotypes, including Agona, Kedougou and Lexington are grouped in clusters with isolates from frozen seafood and spices in Thailand. Contamination with Salmonella from uncooked seafood products sold at market has been estimated at the rate of 21% (Woodring et al., 2012). However, these serotypes have been identified from multiple agricultural products and retail food products, including seafood, meat, spices, and herbs (Zhao et al., 2006). Spices such as clove, oregano, black pepper, red chili, and pepper powder have become important sources of Salmonella contamination, and have been implicated in foodborne outbreaks in several countries, e.g., contaminated fresh basil in Denmark; and herbal tea in Germany (Koch et al., 2005; Pakalniskiene et al., 2009). Cross-contamination between foodstuffs at the retail point cannot be discounted due to the remaining of Salmonella on the food contact surface, which difficult to eradicate from the retail environment and facilities (De Cesare et al., 2003; Campos et al., 2019).
Although some virulence genes were detected at low frequency, many of the virulence genes we identified were found in nearly all isolates, including those associated with host cell adhesion, invasion and intracellular survival (Jajere, 2019). Alarmingly, we also were able to identify cytolethal distending toxin genes (cdt) in a small number of isolates from serotypes Give and Panama. Typically, cdt genes are found in highly virulent Salmonella Typhi isolates, although there is precedence for their presence in non-typhoidal Salmonella (Haghjoo and Galán, 2004). Previous work has identified cdt genes in non-typhoidal Salmonella serotypes Javiana, Montevideo, and Oranienburg, and were implicated in their higher capability to persist and cause infection (Miller and Wiedmann, 2016). Virulence genes were found in all isolates at all steps of the pork production chain, timescale, or geographical area. Isolates from the same serotype shared similar virulence profiles, which transcended sampling source. This suggests that all isolates that we collected had the potential to colonize and infect humans, whether through direct consumption by consumers of retail food products or exposure of workers on the farms and slaughterhouses (Poonchareon et al., 2019).
The global rise in AMR pathogens is a significant public health risk and forecasts on death tolls resulting from AMR infections are shocking—with an estimated death toll of 10 million people by 2050, if no action is taken (Balouiri et al., 2016). Fluoroquinolone resistant Salmonella are among the WHO’s high priority organisms for development of new antibiotics [World Health Organization (WHO), 2017]. In our collection, all 43 isolates were multidrug resistant and resistant to at least three antimicrobial classes. Previous studies have identified widespread dissemination of MDR Salmonella throughout the pork production chain, with high prevalence (98%) in Thailand (Sinwat et al., 2016). Multi-drug resistant Salmonella have also been observed in the neighboring countries, including Laos (98.4%) and Cambodia (52%) (Trongjit et al., 2017). Isolates carried at least different 10 ARGs, associated with aminoglycoside, fluoroquinolone, fosfomycin, and peptide resistance. All antimicrobials where putative resistance was identified are widely used in veterinary practice, especially in swine production—where farmed swine are thought to consume more antimicrobials than any other livestock animal (Van Boeckel et al., 2015; Nhung et al., 2016). The highest number of ARG was found in farm isolates, followed by the slaughterhouses and then the retail markets. Clearly, there is a strong selective pressure imposed by incongruous antimicrobial usage, evidenced here and in other studies by a high prevalence of AMR organisms (not just Salmonella) in industrially farmed swine (Harada and Asai, 2010; Tadee et al., 2015). Our findings support the bleak WHO outlook where a rise in fluoroquinolone resistant organisms continues to erode the efficacy of antimicrobials for clinical cases and there is an urgent need to either curtail this rise; or develop novel antimicrobials (Lee et al., 2009).
The lack of strong regulation and indiscriminate use of antimicrobials has promoted dissemination of ARGs in the Thai food chain (Lekagul et al., 2021). As a result, the Department of Livestock Development, Ministry of Agriculture and cooperatives have implemented a ban on the use of any antimicrobials as a growth promoter in animal feed to combat antimicrobial resistance in livestock animals. Even though the government agency has already issued some policies for reducing the antimicrobial resistance pathogens’ occurrence in the livestock section, some antimicrobial agents, including aminoglycoside and fluoroquinolone, are still available for treatment at the farm level (Nuangmek et al., 2021). Our findings indicate that multidrug resistant Salmonella are still a problem in the pork production chain, and larger scale surveillance studies are required.
Conclusion
In the Chiang Mai and Lamphun Municipality areas, Salmonella contamination in the pork production chain has been burdened and substantial for a decade. Salmonella serotypes Rissen and 1,4,[5],12:i:- are the predominant serotypes found in this area and already investigated by using molecular typing method (Prasertsee et al., 2019; Patchanee et al., 2020). By the way, other infrequent Salmonella serotypes found in the pork production chain also harmful to all stakeholders along the production chain. This study can provide additional information about the evidence of Salmonella’s cross-contamination at the pre-harvest level. Furthermore, the whole genome sequence-based analysis can substantiate the possibility of Salmonella contamination from other agricultural products to the pork at the retail level. All of these isolates contained the necessary virulence genes for the pathogenicity of salmonellosis. In addition, the Salmonella isolates in this study were the multi-drug resistance Salmonella which harmful to the public health worldwide. Additional information from this study would motivate all stakeholders to be aware of and pay attention to the reinforcement of standardized sanitation throughout the pork production chain in order to eradicate and reduce the risk of Salmonella contamination.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA573746; https://www.ncbi.nlm.nih.gov/bioproject/PRJNA419926.
Author contributions
TE-A conceived and designed the experiments, performed the experiment, analyzed the data, prepared figures and/or tables, and approved the final draft. PT conceived and designed the experiments, prepared figures and/or tables, authored or reviewed drafts of the manuscript, and approved the final draft. BP conceived and designed the experiments, performed the experiment, analyzed the data, and approved the final draft. PP conceived and designed the experiments, authored or reviewed drafts of the manuscript, and approved the final draft. All authors contributed to the article and approved the submitted version.
Funding
This research project was supported by National Research Council of Thailand (NRCT): NRCT5-RGJ63004-073. BP was funded by the Medical Research Council (MR/V001213/1) and National Institutes of Health (1R01AI158576-01). All high-performance computing was performed on MRC CLIMB (supported by MRC grants MR/L015080/1 and MR/T030062/1).
Acknowledgments
We would like to express our appreciation to our colleagues at Chiang Mai University for noteworthy contributions and partial funding support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.968695/full#supplementary-material
Supplementary Figure 1 | The minimum spanning tree (MST) analysis of Salmonella isolates recovered from the pork production chain. Each isolate was grouped according to the EnteroBase Salmonella database’s cgMLST scheme, which considers a total of 3,002 loci (Pearce et al., 2020). The number on the branch represent the number of loci different between each isolate. Node color represents each serotype of Salmonella isolates. Outer border color coding: green color, yellow color and pink color represent the Salmonella isolates recovered from farm, slaughterhouse and retail market, respectively.
Supplementary Figure 2 | The minimum spanning tree (MST) analysis of 43 Salmonella isolates recovered from pork production chain (striped nodes) and additional 61 Salmonella isolates circulating in Thailand. Each isolate was grouped according to the loci different of EnteroBase Salmonella database’s cgMLST scheme, which considers a total of 3,002 loci (Pearce et al., 2020). The number on the branch represent the number of loci different between each isolate. Node color coding were representing the serotypes of the Salmonella isolates. Outer border color represents each source of Salmonella isolates.
Supplementary Figure 3 | The Venn diagram of intersection analysis of virulence genes among different steps of pork production chain. The Venn diagram represent the number of unique and shared virulence genes in 43 Salmonella isolates recovered from farms, slaughterhouses and retail markets.
Supplementary Figure 4 | The Venn diagram of intersection analysis of antimicrobial resistance genes among different steps of the pork production chain. The Venn diagram represents the number of unique and shared antimicrobial resistance genes in 43 Salmonella isolates recovered from farms, slaughterhouses, and retail markets.
Supplementary Table 1 | Origin and characteristic of S. enterica isolates originating from various sources in Thailand during the period of 2001 through 2016 acquired from public repositories (Enterobase Salmonella database).
Supplementary Table 2 | Kirby-Bauer disk diffusion susceptibility test of 43 Salmonella isolates recovered from pork production chain.
Footnotes
- ^ https://www.ncbi.nlm.nih.gov/sra
- ^ https://pubmlst.org/bigsdb?db=pubmlst_salmonella_isolates&page=plugin&name=GenomeComparator
- ^ https://enterobase.warwick.ac.uk/species/index/senterica
- ^ https://patricbrc.org/app/Annotation
References
Achtman, M., Zhou, Z., Alikhan, N. F., Tyne, W., Parkhill, J., Cormican, M., et al. (2020). Genomic diversity of Salmonella enterica – the UoWUCC 10K genomes project. Wellcome Open Res. 5:223. doi: 10.12688/wellcomeopenres.16291.1
Alcock, B. P., Raphenya, A. R., Lau, T. T., Tsang, K. K., Bouchard, M., Edalatmand, A., et al. (2020). CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 48, D517–D525. doi: 10.1093/nar/gkz935
Balouiri, M., Sadiki, M., and Ibnsouda, S. K. (2016). Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 6, 71–79.
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Brenner, F. W., Villar, R. G., Angulo, F. J., Tauxe, R., and Swaminathan, B. (2000). Salmonella nomenclature. J. Clin. Microbiol. 38, 2465–2467. doi: 10.1128/JCM.38.7.2465-2467.2000
Brettin, T., Davis, J. J., Disz, T., Edwards, R. A., Gerdes, S., Olsen, G. J., et al. (2015). RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 5:8365. doi: 10.1038/srep08365
Bureau of Epidemiology (2018). Annual epidemiological surveillance report. Available online at: https://apps-doe.moph.go.th/boeeng/download/AW_Annual_Mix
Campioni, F., Bergamini, A. M. M., and Falcão, J. P. (2012). Genetic diversity, virulence genes and antimicrobial resistance of Salmonella Enteritidis isolated from food and humans over a 24-year period in Brazil. Food Microbiol. 32, 254–264. doi: 10.1016/j.fm.2012.06.008
Campos, J., Mourão, J., Peixe, L., and Antunes, P. (2019). Non-typhoidal Salmonella in the pig production chain: A comprehensive analysis of its impact on human health. Pathogens 8:19. doi: 10.3390/pathogens8010019
Center of Disease Control and Prevention (2021). Salmonella. U.S. Department of Health and Human Services, CDC. Available online at: https://www.cdc.gov/salmonella/index.html (accessed April 20, 2021).
Chen, H. M., Wang, Y., Su, L. H., and Chiu, C. H. (2013). Nontyphoid Salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatr. Neonatol. 54, 147–152. doi: 10.1016/j.pedneo.2013.01.010
Chen, T., Jiang, J., Ye, C., Xie, J., Chen, X., Xu, D., et al. (2019). Genotypic characterization and antimicrobial resistance profile of Salmonella isolated from chicken, pork and the environment at abattoirs and supermarkets in Chongqing, China. BMC Vet. Res. 15:456. doi: 10.1186/s12917-019-2202-4
De Cesare, A., Sheldon, B. W., Smith, K. S., and Jaykus, L. A. (2003). Survival and persistence of Campylobacter and Salmonella species under various organic loads on food contact surfaces. J. Food Prot. 66, 1587–1594. doi: 10.4315/0362-028x-66.9.1587
Ferrari, R. G., Rosario, D. K., Cunha-Neto, A., Mano, S. B., Figueiredo, E. E., and Conte-Junior, C. A. (2019). Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 85:e00591-19. doi: 10.1128/AEM.00591-19
Ferri, M., Ranucci, E., Romagnoli, P., and Giaccone, V. (2017). Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 57, 2857–2876. doi: 10.1080/10408398.2015.1077192
Girardin, F., Mezger, N., Hächler, H., and Bovier, P. A. (2006). Salmonella serovar Give: An unusual pathogen causing splenic abscess. Eur. J. Clin. Microbiol. Infect. Dis. 25, 272–274. doi: 10.1007/s10096-006-0122-2
Haghjoo, E., and Galán, J. E. (2004). Salmonella typhi encodes a functional cytolethal distending toxin that is delivered into host cells by a bacterial-internalization pathway. Proc. Natl. Acad. Sci. U.S.A. 101, 4614–4619. doi: 10.1073/pnas.0400932101
Harada, K., and Asai, T. (2010). Role of antimicrobial selective pressure and secondary factors on antimicrobial resistance prevalence in Escherichia coli from food-producing animals in Japan. J. Biomed. Biotechnol. 2010:180682. doi: 10.1155/2010/180682
Jajere, S. M. (2019). A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 12, 504–521. doi: 10.14202/vetworld.2019.504-521
Kintz, E., Davies, M. R., Hammarlöf, D. L., Canals, R., Hinton, J. C., and van der Woude, M. W. (2015). A BTP 1 prophage gene present in invasive non-typhoidal Salmonella determines composition and length of the O-antigen of the lipopolysaccharide. Mol. Microbiol. 96, 263–275. doi: 10.1111/mmi.12933
Koch, J., Schrauder, A., Alpers, K., Werber, D., Frank, C., Prager, R., et al. (2005). Salmonella Agona outbreak from contaminated aniseed, Germany. Emerg. Infect. Dis. 11, 1124–1247. doi: 10.3201/eid1107.041022
Lee, H. Y., Su, L. H., Tsai, M. H., Kim, S. W., Chang, H. H., Jung, S. I., et al. (2009). High rate of reduced susceptibility to ciprofloxacin and ceftriaxone among nontyphoid Salmonella clinical isolates in Asia. Antimicrob. Agents Chemother. 53, 2696–2699. doi: 10.1128/AAC.01297-08
Lekagul, A., Tangcharoensathien, V., Liverani, M., Mills, A., Rushton, J., and Yeung, S. (2021). Understanding antibiotic use for pig farming in Thailand: A qualitative study. Antimicrob. Resist. Infect. Control 10:3. doi: 10.1186/s13756-020-00865-9
Liu, B., Zheng, D., Jin, Q., Chen, L., and Yang, J. (2019). VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 47, D687–D692. doi: 10.1093/nar/gky1080
Miller, R. A., and Wiedmann, M. (2016). The cytolethal distending toxin produced by nontyphoidal Salmonella serotypes Javiana, Montevideo, Oranienburg, and Mississippi induces DNA damage in a manner similar to that of serotype Typhi. mBio 7:e2109-16. doi: 10.1128/mBio.02109-16
Nhung, N. T., Cuong, N. V., Thwaites, G., and Carrique-Mas, J. (2016). Antimicrobial usage and antimicrobial resistance in animal production in Southeast Asia: A review. Antibiotics 5:37. doi: 10.3390/antibiotics5040037
Niyomdecha, N., Mungkornkaew, N., and Samosornsuk, W. (2016). Serotypes and antimicrobial resistance of Salmonella enterica isolated from pork, chicken meat and lettuce, Bangkok and central Thailand. Southeast Asian J. Trop. Med. Public Health 47, 31–39.
Nuangmek, A., Rojanasthien, S., Yamsakul, P., Tadee, P., Eiamsam-ang, T., Thamlikitkul, V., et al. (2021). Perspectives on antimicrobial use in pig and layer farms in Thailand: Legislation, policy, regulations and potential. Vet. Integr. Sci. 19, 1–21.
Oakeson, K. F., Wagner, J. M., Mendenhall, M., Rohrwasser, A., and Atkinson-Dunn, R. (2017). Bioinformatic analyses of whole-genome sequence data in a public health laboratory. Emerg. Infect. Diseases 23, 1441–1445. doi: 10.3201/eid2309.170416
Padungtod, P., and Kaneene, J. B. (2006). Salmonella in food animals and humans in northern Thailand. Int. J. Food Microbiol. 108, 346–354. doi: 10.1016/j.ijfoodmicro.2005.11.020
Pakalniskiene, J., Falkenhorst, G., Lisby, M., Madsen, S. B., Olsen, K. E. P., Nielsen, E. M., et al. (2009). A foodborne outbreak of enterotoxigenic E. coli and Salmonella Anatum infection after a high-school dinner in Denmark, November 2006. Epidemiol. Infect. 137, 396–401. doi: 10.1017/S0950268808000484
Patchanee, P., Tanamai, P., Tadee, P., Hitchings, M. D., Calland, J. K., Sheppard, S. K., et al. (2020). Whole-genome characterization of multidrug resistant monophasic variants of Salmonella Typhimurium from pig production in Thailand. PeerJ 8:e9700. doi: 10.7717/peerj.9700
Patchanee, P., Tansiricharoenkul, K., Buawiratlert, T., Wiratsudakul, A., Angchokchatchawal, K., Yamsakul, P., et al. (2016). Salmonella in pork retail outlets and dissemination of its pulsotypes through pig production chain in Chiang Mai and surrounding areas, Thailand. Prev. Vet. Med. 130, 99–105. doi: 10.1016/j.prevetmed.2016.06.013
Pearce, M. E., Chattaway, M. A., Grant, K., and Maiden, M. C. (2020). A proposed core genome scheme for analyses of the Salmonella genus. Genomics 112, 371–378. doi: 10.1016/j.ygeno.2019.02.016
Phongaran, D., Khang-Air, S., and Angkititrakul, S. (2019). Molecular epidemiology and antimicrobial resistance of Salmonella isolates from broilers and pigs in Thailand. Vet. World 12, 1311–1318. doi: 10.14202/vetworld.2019.1311-1318
Poonchareon, K., Pulsrikarn, C., Nuanmuang, N., and Khamai, P. (2019). Effectiveness of BOX-PCR in differentiating genetic relatedness among Salmonella enterica serotype 4,[5], 12: I: - isolates from hospitalized patients and minced pork samples in northern Thailand. Int. J. Microbiol. 2019:5086240. doi: 10.1155/2019/5086240
Prasertsee, T., Chuammitri, P., Deeudom, M., Chokesajjawatee, N., Santiyanont, P., Tadee, P., et al. (2019). Core genome sequence analysis to characterize Salmonella enterica serovar Rissen ST469 from a swine production chain. Int. J. Food Microbiol. 304, 68–74. doi: 10.1016/j.ijfoodmicro.2019.05.022
Pulford, C. V., Perez-Sepulveda, B. M., Rodwell, E. V., Weill, F. X., Baker, K. S., and Hinton, J. C. (2019). Salmonella enterica serovar Panama, an understudied serovar responsible for extraintestinal salmonellosis worldwide. Infect. Immun. 87:e00273-19. doi: 10.1128/IAI.00273-19
Punpanich, W., Netsawang, S., and Thippated, C. (2012). Invasive salmonellosis in urban Thai children: A ten-year review. Pediatr. Infect. Dis. J. 31:e105-10. doi: 10.1097/INF.0b013e31825894b0
Rostagno, M. H., and Callaway, T. R. (2012). Pre-harvest risk factors for Salmonella enterica in pork production. Food Res. Int. 45, 634–640. doi: 10.1016/j.foodres.2011.04.041
Saha, P., Xiao, X., Yeoh, B. S., Chen, Q., Katkere, B., Kirimanjeswara, G. S., et al. (2019). The bacterial siderophore enterobactin confers survival advantage to Salmonella in macrophages. Gut Microbes 10, 412–423. doi: 10.1080/19490976.2018.1546519
Sanguankiat, A., Pinthong, R., Padungtod, P., Baumann, M. P., Zessin, K. H., Srikitjakarn, L., et al. (2010). A cross-sectional study of Salmonella in pork products in Chiang Mai, Thailand. Foodborne Pathog. Dis. 7, 873–878. doi: 10.1089/fpd.2009.0436
Savall, J. F., Bidot, C., Leblanc-Maridor, M., Belloc, C., and Touzeau, S. (2016). Modelling Salmonella transmission among pigs from farm to slaughterhouse: Interplay between management variability and epidemiological uncertainty. Int. J. Food Microbiol. 229, 33–43. doi: 10.1016/j.ijfoodmicro.2016.03.020
Sinwat, N., Angkittitrakul, S., Coulson, K. F., Pilapil, F. M. I. R., Meunsene, D., and Chuanchuen, R. (2016). High prevalence and molecular characteristics of multidrug-resistant Salmonella in pigs, pork and humans in Thailand and Laos provinces. J. Med. Microbiol. 65, 1182–1193. doi: 10.1099/jmm.0.000339
Sringam, P., Tungtanatanich, P., Suwannachot, P., Suksawat, F., Khang-Air, S., and Angkititrakul, S. (2017). Molecular epidemiology and antibiogram of Salmonella isolates from humans, swine and pork. Southeast Asian J. Trop. Med. Public Health 48, 1017–1028.
Tadee, P., Boonkhot, P., Pornruangwong, S., and Patchanee, P. (2015). Comparative phenotypic and genotypic characterization of Salmonella spp. in pig farms and slaughterhouses in two provinces in northern Thailand. PLoS One 10:e0116581. doi: 10.1371/journal.pone.0116581
Tadee, P., Kumpapong, K., Sinthuya, D., Yamsakul, P., Chokesajjawatee, N., Nuanualsuwan, S., et al. (2014). Distribution, quantitative load and characterization of Salmonella associated with swine farms in upper-northern Thailand. J. Vet. Sci. 15, 327–334. doi: 10.4142/jvs.2014.15.2.327
Trongjit, S., Angkititrakul, S., Tuttle, R. E., Poungseree, J., Padungtod, P., and Chuanchuen, R. (2017). Prevalence and antimicrobial resistance in Salmonella enterica isolated from broiler chickens, pigs and meat products in Thailand–Cambodia border provinces. Microbiol. Immunol. 61, 23–33. doi: 10.1111/1348-0421.12462
Van Boeckel, T. P., Brower, C., Gilbert, M., Grenfell, B. T., Levin, S. A., Robinson, T. P., et al. (2015). Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. U.S.A. 112, 5649–5654. doi: 10.1073/pnas.1503141112
Wang, H., Chen, Y., Ayers, S., Melka, D., Laasri, A., Payne, J. S., et al. (2015). Draft genome sequence of Salmonella enterica subsp. enterica serovar Give, isolated from an imported chili powder product. Genome Announc. 3:e00726-15. doi: 10.1128/genomeA.00726-15
Wang, M., Qazi, I. H., Wang, L., Zhou, G., and Han, H. (2020). Salmonella virulence and immune escape. Microorganisms 8:407. doi: 10.3390/microorganisms8030407
Wattam, A. R., Davis, J. J., Assaf, R., Boisvert, S., Brettin, T., Bun, C., et al. (2017). Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 45, D535–D542. doi: 10.1093/nar/gkw1017
Woodring, J., Srijan, A., Puripunyakom, P., Oransathid, W., Wongstitwilairoong, B., and Mason, C. (2012). Prevalence and antimicrobial susceptibilities of Vibrio, Salmonella, and Aeromonas isolates from various uncooked seafoods in Thailand. J. Food Prot. 75, 41–47. doi: 10.4315/0362-028X.JFP-11-211
World Health Organization (2017). WHO publishes list of bacteria for which new antibiotics are urgently needed. Available online at: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed May 15, 2021).
Keywords: Salmonella, whole genome sequencing, core genome MLST (cgMLST), antimicrobial resistance genes (ARG), virulence genes, pork production chain
Citation: Eiamsam-ang T, Tadee P, Pascoe B and Patchanee P (2022) Genome-based analysis of infrequent Salmonella serotypes through the Thai pork production chain. Front. Microbiol. 13:968695. doi: 10.3389/fmicb.2022.968695
Received: 14 June 2022; Accepted: 05 August 2022;
Published: 25 August 2022.
Edited by:
Magaly Toro, University of Chile, ChileReviewed by:
Jessica L. Jones, United States Food and Drug Administration, United StatesAndrea Isabel Moreno Switt, Pontificia Universidad Católica de Chile, Chile
Copyright © 2022 Eiamsam-ang, Tadee, Pascoe and Patchanee. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ben Pascoe, Yi5wYXNjb2VAYmF0aC5hYy51aw==; Prapas Patchanee, cHJhcGFzLnBhdEBjbXUuYWMudGg=