 Paula Morales
Paula Morales Chanté Muller
Chanté Muller Nadine Jagerovic
Nadine Jagerovic Patricia H. Reggio
Patricia H. Reggio- 1Medicinal Chemistry Institute, Spanish National Research Council, Madrid, Spain
- 2Department of Chemistry and Biochemistry, University of North Carolina at Greensboro, Greensboro, NC, United States
Both metabotropic (CBRs) and ionotropic cannabinoid receptors (ICRs) have implications in a range of neurological disorders. The metabotropic canonical CBRs CB1 and CB2 are highly implicated in these pathological events. However, selective targeting at CB2 versus CB1 offers optimized pharmacology due to the absence of psychoactive outcomes. The ICR transient receptor potential vanilloid type 1 (TRPV1) has also been reported to play a role in CNS disorders. Thus, activation of both targets, CB2 and TRPV1, offers a promising polypharmacological strategy for the treatment of neurological events including analgesia and neuroprotection. This brief research report aims to identify chemotypes with a potential dual CB2/TRPV1 profile. For this purpose, we have rationalized key structural features for activation and performed virtual screening at both targets using curated chemical libraries.
1 Introduction
Well documented pharmacological evidence supports functional crosstalk between the endocannabinoid system (ECS) and the endovanilloid system (EVS) (Di Marzo et al., 2002; Lastres-Becker et al., 2003; Morgese et al., 2007; Avraham et al., 2010; Chávez et al., 2010; Adamczyk et al., 2012; Arnold et al., 2012; Lowin and Straub, 2015; Rossi et al., 2015; Malek and Starowicz, 2016; Assimakopoulou et al., 2017; Bellini et al., 2017; Zhang et al., 2017; Punzo et al., 2018; Bhatta et al., 2019; Wi et al., 2020). Thus, these latest advances provide opportunities to develop innovative strategies for fighting disorders where biological targets of both systems are involved. Here, we emphasize the cannabinoid receptor type 2 (CB2) and the transient receptor potential vanilloid type 1 (TRPV1) channel, both implicated in neurodegenerative diseases and pain.
CB2R is a G-protein-coupled receptor (GPCR) mainly present in the immune cells where they are expressed in lymphocytes, natural killer cells, macrophages, and neutrophils (Cécyre et al., 2020). Thus, they are an attractive target for the treatment of inflammatory processes. The expression of CB2 is also detected in the central nervous system (CNS) under stressful conditions such as cytotoxic and neuroinflammatory injuries within the brainstem, microglia, and astrocytes, suggesting CB2 an interesting target for neuroprotection (Navarro et al., 2016). CB2 is also expressed in the blood brain barrier (BBB), and therefore could be beneficial in the brain and peripheral tissues at different stages of neurodegenerative processes (Aso and Ferrer, 2016; Javed et al., 2016; Cassano et al., 2017; Behl et al., 2020; Berry et al., 2020; Uddin et al., 2020). CB2 selective agonists also represent an attractive approach for pain management among other therapeutic applications (Fowler, 2020). In animal models of chronic inflammation, CB2 agonists lead to beneficial outcomes for diverse pain managements such as neuropathic, osteoarthritic, postoperative, and human immunodeficiency virus (HIV) associated pain relief (Guindon and Hohmann, 2008; Anthony et al., 2020; Aly and Masocha, 2021; Bryk and Starowicz, 2021; Mlost et al., 2021; Ramírez-López et al., 2021).
TRPV1 is a nonselective cation channel mainly expressed in the sensory neurons of the peripheral nervous system (Caterina et al., 1997), acting as a detector of painful stimuli such as heat and pungent chemicals like capsaicin. TRPV1 modulators have attracted much attention as analgesics due to its implication in pathological pain such as inflammatory, visceral, neuropathic, and cancer-related pain (Peppin and Pappagallo, 2014; Malek and Starowicz, 2016; Shuba, 2021). TRPV1 has also been described in the CNS (Gibson et al., 2008; Ho et al., 2012; Shuba, 2021) with expression in neurons, microglia, and astrocytes (Sawamura et al., 2017), and its level of expression can be up- or down-regulated according to age and pathophysiological conditions (Martins et al., 2014). TRPV1 participates to the regulation of neuronal function and synaptic plasticity (Marinelli et al., 2003; Maione et al., 2009; Chávez et al., 2010), the control of motor behavior (Morgese et al., 2007; González-Aparicio and Moratalla, 2014; Martins et al., 2014), and the regulation of neuroinflammation (Kong et al., 2017). Therefore, TRPV1 has been suggested to be implicated in diseases associated with motor dysfunctions, such as Huntington’s, Parkinson’s, and multiple sclerosis, or with cognitive functions like Alzheimer’s disease (González-Aparicio and Moratalla, 2014; Nam et al., 2015; Li et al., 2019; Du et al., 2020).
Co-expression and crosstalk between TRPV1 and CB1 (Cristino et al., 2006; Assimakopoulou et al., 2017) has been established primarily in the modulation of arthritic pain and inflammation (Lowin and Straub, 2015). In addition to CB1, CB2 is also co-expressed with TRPV1 in certain cells including osteoblasts (Rossi et al., 2015), osteoclasts (Bellini et al., 2017), and sensory neurons (Wi et al., 2020). Moreover, CB2 and TRPV1 crosstalk has shown to be engaged diverse pathophysiological processes including pain (Wi et al., 2020; Wilkerson et al., 2022), bone disorders (Rossi et al., 2015; Bellini et al., 2017), inflammatory processes (Lowin et al., 2016; Arnold et al., 2021), cocaine-seeking behavior (Adamczyk et al., 2012), proliferation and apoptosis of T-lymphoblastic leukemia cells (Punzo et al., 2018), and multidrug resistance (Arnold et al., 2012). Benefits of the CBR/TRPV1 axis for neurodegenerative diseases has been suggested by some studies due to CBRs and TRPV1 inhibition of glial activation and expression of proinflammatory cytokines in a mouse model of Parkinson’s disease (Wi et al., 2020). Pharmacologically, strategies targeting CB1/TRPV1 have shown promising therapeutic results in models of pain, spasticity, arthritis, and dyskinesia (Di Marzo et al., 2001, 2002; Brooks et al., 2002; Morgese et al., 2007; Lowin and Straub, 2015) For instance, arvanil, a CB1 agonist, TRPV1 activator, and potent inhibitor of anandamide (AEA) accumulation, alleviates hyperkinesia typical of Huntington’s disease (De Lago et al., 2005). However, few reports have identified dual CB2/TRPV1 modulators thus far.
Current treatments for complex disorders based on selective-target drugs fail in their efficacy. As a consequence, a number of research studies have highlighted the importance of multiple-target strategies for the treatment of multifactorial disorders such as pain and neurodegenerative diseases (Cheong et al., 2019; Gontijo et al., 2019; Maramai et al., 2020). Combinatorial therapies are generally associated with side effects derived from drug-drug interactions. Therefore, single dual-acting drugs should reduce side effects with unique pharmacokinetic or pharmacodynamic profiles. Cannabinoids have been reported to directly modulate TRPV1 (Muller et al., 2019), and among them, few have shown selective CB2 vs CB1 activity. In this brief research report, we will primarily focus on the in silico identification of potential CB2/TRPV1 chemotypes, as well as rationalize reported dual modulators.
2 Methods and Materials
2.1 Receptor Structures
Structures of hCB2 and hTRPV1 were selected based on the reliability and stability of the structures. In a recent publication, an activated structure of hCB2 was resolved via cryo-EM at a resolution of 2.90 Å (PDB: 6KPF) (Hua et al., 2020). This structure was used for our docking screening upon treatment using the protein structure preparation wizard integrated in the Schrödinger software. A model of hTRPV1 was constructed using the cryo-EM structure PDB: 5IRZ congruent to the methods described in Muller et al. and was used for this work (Muller et al., 2020).
2.2 Grid Generation
Prior to using the Glide module high-throughput virtual screening (HTVS) and extra precise (XP) docking within the Schrödinger package (Schrödinger, LLC, New York, NY, 2019), docking grids were generated using the receptor grid generation tool within Glide to ensure ligand screening was performed in the appropriate sites within each receptor. Dimensions for the CB2 receptor grid were set at 20 Å in length along the x, y, and z axes and was centered on the ligand co-crystallized with the CB2 structure (the THC synthetic derivative AM12033).
Similarly, all three TRPV1 grids were generated to adhere to the same dimensions of 20 Å in length in the x, y, and z directions and were centered on residues that are believed and/or reported to be involved with ligand binding at each location. This resulted in three distinctly different grids for TRPV1 that will herein be referred to as “VBP” for the location that capsaicin binds, “tunnel” for the location where anandamide has been reported to interact with TRPV1 via MD simulations, and “CBD-site” for the putative CBD interaction site reported in the TRPV2/CBD cryo-EM structure. Visual representations and further explanation of these TRPV1 sites can be found in Supplementary Figure S1. These grid specifications allow any ligand that is less than or equal to 20 Å in length to be docked within the specified region.
2.3 Curation of Chemical Libraries
2.3.1 CB2
From the CB2 indexed molecules, ligands showing EC50, Emax, and activity data were selected (total of 6356) and retrieved from the ChEMBL webserver as a .csv file. The “activity” category includes compounds with not only agonist activity, but antagonist, inverse agonists, and allosteric modulators as well. DataWarrior, an open-source data visualization software, was used to further analyze the ligand output which included discarding ligands without an agonist profile (−568 ligands), removing duplicates (−2159 ligands), and eliminating ligands with low activity (−773 ligands). This resulted in a final CB2 library of 2856 unique molecules that included a variety of chemotypes.
2.3.2 TRPV1
Ligands that have been indexed for TRPV1 activity within the ChEMBL database were selected and filtered in search of agonists in accordance with the reported EC50 and Emax values and activity. The resulting 7,436 compounds were exported from the ChEMBL webserver as a .csv file and uploaded to DataWarrior. The selection of ligands with TRPV1 activity from the ChEMBL database included antagonists, inverse agonists, possible allosteric modulators, ligands with low activity, and duplicates which were all removed using DataWarrior. The final curated TRPV1 library contained 3,830 unique molecules with a variety of chemotypes.
2.3.3 Internal Standard Ligands
The CB2 agonist resolved with the active hCB2 structure (AM12033) was used as an internal standard for CB2 docking. Three internal standards were used for hTRPV1: capsaicin in the VBP, AEA in the tunnel as observed from MD simulations, and CBD at the putative CBD site.
2.3.4 JWH133 Similarity Library
JWH133, which acts as an agonist at both CB2 and TRPV1, was used as a molecular basis for this additional screen to explore more unique scaffold options that may not be present in the CB2 or TRPV1 curated libraries. A JWH133 similarity library was curated using PubChem Biosays, 2021 which included compounds that shared >0.85 Tanimoto similarity index with JWH133, while also following Lipinsky’s rules of drug likeness (apart from xLogP values, which were set to −1 to 6 due to the lipophilicity of cannabinoid ligands). The JWH133 similarity library consisted of 5081 that were screened at all sites (CB2 and the three TRPV1 sites), and the output was analyzed to identify dual potential chemotypes.
2.4 High-Throughput Virtual Screening Workflow
A general overview of the screening workflows is provided in Supplementary Figure S2.
2.4.1 Ligand Preparation
Each of the curated libraries were exported as .sdf files and their conformations were optimized using the LigPrep module of the Maestro suite (Schrödinger, LLC, New York, NY, 2019). The Epik software was employed to predict pKa values in the pH range of 7.0 ± 0.5 and to return all chemically sensible structures in accordance with the Hammett and Taft methodology. All compounds were minimized using the OPLS3e force field as implemented in Maestro.
2.4.2 HTVS
Molecular docking was performed using the HTVS Glide-dock module integrated in the Schrödinger package. The HTVS was conducted under the default setting, ensuring that high-energy ionization and tautomer states were removed, and the planarity of conjugated pi systems were enhanced. Ligands were docked flexibly, allowing for exploration of an arbitrary number of torsional degrees of freedom, in addition to the six spatial degrees of freedom spanned by the translational and rotational parameters. Up to 10 poses per compound state were generated and ligand poses that were generated in this way were run through a series of hierarchical filters to evaluate ligand interactions with the receptor. Docking score, glide gscore, glide emodel, ionization penalty, and topological polar surface area (TPSA) were used to select the docking poses in the output. The output from the HTVS contained the top 10% of the best scoring compound states and were analyzed for use in the extra precise (XP) screen via their docking scores.
2.4.3 XP Screening
Top scoring compounds from the HTVS were then studied through high-precision docking calculations which was performed using the XP Glide module. As with the HTVS protocol, 10 poses of the short-listed ligands were docked flexibly in their respective receptor site within the generated grids. A post-docking minimization was performed and the top 20% of the best scoring ligands were retained. XP Glide uses two key features that impact the XP Glide scoring: the recognition of structural motifs that provide large contributions to binding affinity and the application of large desolvation penalties to ligand and protein polar and charged groups wherever appropriate. To accomplish this, the sampling algorithm and scoring functions have been simultaneously optimized in XP. Ligands making it through the XP screen were organized by their docking scores and analyzed for ligand/receptor interactions. Selected ligands for each receptor were investigated through manual docking based on the automatic docking score, binding mode, as well as reported activity.
2.4.4 Additional Criteria
Manual Docking Identification of Potential PAINS Off-Targets Evaluation. Selected compounds were subjected to manual docking at CB2 and TRPV1 for further investigation of key interactions. Docking at CB2 was performed following the protocols previously reported by us for cannabinoid and related GPCRs (Morales et al., 2017a). In the case of TRPV1, select ligands were positioned within the respective binding site with steric clashes being removed via ligand and/or receptor adjustment using a graphical interface. Minimization of the ligand and surrounding 6 Å of residues (due to complex size) was performed using Prime version 19.3 (Schrödinger Inc.) with the OPLS3e forcefield in an implicit membrane.
In silico calculation of ADME properties. A set of 34 physico-chemical descriptors was computed using QikProp version 3.5 integrated in Maestro (Schrödinger, LLC, New York, United States). The QikProp descriptors are shown in Supplementary Tables S2, S3. The 3D conformations used in the calculation of QikProp descriptors were generated using LigPrep as previously detailed.
Identification of Potential PAINS. In the search of potential candidates, it is crucial to avoid the presence of potential promiscuous moieties or PAINS (pan-assay interference compounds) (Baell and Holloway, 2010; Capuzzi et al., 2017). Therefore, the selected molecules were subjected to a PAINS identification study using the swissADME webserver (Daina et al., 2017).
Off-Targets evaluation. XP Glide docks at potential off-target receptors including cannabinoid-related GPCRs such as CB1 (Shao et al., 2016; Hua et al., 2020), GPR55 (Kotsikorou et al., 2013; Lingerfelt et al., 2017), GPR18 (Sotudeh et al., 2019) and TRP channels such as TRPV2 (Pumroy et al., 2019), TRPV3 (Singh et al., 2018; Zubcevic et al., 2018), TRPA1 (Suo et al., 2020; Zhao et al., 2020), and TRPM8 (Diver et al., 2019; Yin et al., 2019). For this purpose, the cited available structures, whether crystal, cryoEMs, or models previously developed in our group, have been used. Results of these additional dockings can be found in Supplementary Tables S4, S5.
3 Results and Discussion
Polypharmacological approaches targeting the ECS have already shown successful results in diverse disease models (Fernández-Fernández et al., 2014; Malek and Starowicz, 2016; Barutta et al., 2017; Lago-Fernandez et al., 2021). However, drug discovery strategies primarily targeting CB2 and TRPV1 have not yet been explored. As previously detailed, activation of these targets participates in diverse therapeutic effects including analgesia and neuroprotection, which both offer interesting polypharmacological prospects.
3.1 Structural Understanding of Compounds With Reported Activity at Both Targets
To computationally identify promising chemotypes with a CB2/TRPV1 dual agonist profile we have first analyzed reported compounds exhibiting activity at both receptors. As detailed in Supplementary Table S1, endocannabinoids, phytocannabinoids, and their respective synthetic derivatives have so far shown the best promise in this field.
The well-known endogenous ligands 2-arachidonoylglycerol (2-AG) and anandamide (AEA) exhibit agonist effects at both targets with low micromolar potency. As observed in diverse in vitro and in vivo models, these endocannabinoids also display activity at other cannabinoid-related GPCRs including CB1, GPR55, and GPR18 (Morales and Jagerovic, 2016; Morales and Reggio, 2017; Morales et al., 2020) as well as other TRP channels including TRPA1 and TRPM8 (Muller et al., 2019).
Synthetic endocannabinoid-like derivatives have also shown dual activity (Supplementary Table S1). For instance, Appendino and coworkers reported a series of conformationally constrained fatty-acid ethanolamides with CB1, CB2, and TRPV1 activity (Appendino et al., 2009). An example from this series is ACPA-OH (Supplementary Table S1), which introduces a hydroxycyclopropyl in the amide head group forcing a specified stereochemistry and rigidity. This compound is a potent TRPV1 agonist that exerts low micromolar CB2 affinity and nanomolar binding at CB1 (Appendino et al., 2009). Further synthetic efforts from Di Marzo’s research group led to the identification of hybrid cannabinoid-vanilloid ligands with a highly CB1 selective profile (Melck et al., 1999; Szallasi and Di Marzo, 2000; Di Marzo et al., 2001, Di Marzo et al., 2002). Among these fatty-acid derivatives, one of the few compounds that binds to CB2 is O-1811 (Supplementary Table S1), which presents a substituted dimethyl-hydroxyhexanyl tail (Di Marzo et al., 2001). Despite targeting CB2, O-1811 displays over 6-fold CB1 selectivity.
Interestingly, molecules combining the polyunsaturated fatty-acid chain with the vanillyl-amide head group of capsaicin behave as CB1/TRPV1 agonists that potently inhibit anandamide accumulation (Melck et al., 1999; Szallasi and Di Marzo, 2000; Di Marzo et al., 2001, 2002). One such molecule, arvanil (Supplementary Table S1), has shown therapeutic potential in the treatment of dyskinesia associated to Huntington’s disease (De Lago et al., 2005) and inhibition of spasticity and persistent pain (Brooks et al., 2002).
Structural modifications in the long chain of endocannabinoid-like molecules led to the identification of the first series of CB2 selective/TRPV1 dual ligands (Appendino et al., 2006). Combination of non-polyunsaturated fatty acid-derived chains with 12-acylgroups yielded compounds such as 12-phenylacetylricinoleyl cyclopropylamide (PhAR derivative 12, Supplementary Table S1) which behaves as a potent TRPV1 agonist and CB2 inverse agonist.
Diverse phytocannabinoids have also shown activity at CB2 and TRPV1 (De Petrocellis et al., 2011; Zagzoog et al., 2020). For instance, the main non-psychotropic component of Cannabis sativa, cannabidiol (CBD), is a CB2 partial agonist/TRPV1 agonist (De Petrocellis et al., 2011; Tham et al., 2018). It is worth mentioning that at CB2, CBD has been reported to act as negative allosteric modulator in the presence of orthosteric full agonists (Martínez-Pinilla et al., 2017; Navarro et al., 2021). The acidic CBD derivative, cannabidiolic acid (CBDA), and its propyl counterpart, cannabivarin (CBDV), also exhibited TRPV1 agonism while being CB2 partial agonists (De Petrocellis et al., 2011; Zagzoog et al., 2020). The phytogenic compound cannnabigerol (CBG) presents the same functional profile at both targets (De Petrocellis et al., 2011; Zagzoog et al., 2020). On the other hand, the well-known psychoactive compound tetrahydrocannabinol (THC) is not active at TRPV1 (De Petrocellis et al., 2011), whereas its propyl derivative tetrahydrocannabivarin (THCV) behaves as a TRPV1/CB2 agonist (Supplementary Table S1). It is important to note that all these phytocannabinoids also display activity at CB1 receptors.
Synthetic phytocannabinoid-like derivatives have also shown interesting dual activity. The CB2 selective agonists HU308 and JWH133 could be considered dual ligands due to their activity at TRPV1 being HU308 a weaker agonist at this channel (Soethoudt et al., 2017). The widely used aminoalkylindole WIN55212-2, which is a potent CB1/CB2 synthetic agonist, has also been reported to activate and desensitize TRPV1 (Soethoudt et al., 2017).
In the search of novel structures with CB2/TRPV1 activity, we aim to minimize off-target effects at CB1 or related receptors. Therefore, considering the aforementioned reported activity, we selected JWH133 as a molecular basis for the identification of potential dual CB2/TRPV1 agonists. The therapeutic potential of this ligand has been recently reviewed elsewhere (Agonist et al., 2021). As a first step we rationalized its interactions at both receptors using molecular docking. At CB2 JWH133 sits in the orthosteric pocket with the same orientation as the CB2 agonist resolved in the cryoEM structure AM12033 (Supplementary Figure S3A). The tricycle stablishes π-π stacking with residues F2.61, F2.57 and F183 (extracellular loop 2) while residues I3.29, F2.64 and V3.32 stabilize the molecule through van der Waals interactions. The orientation of the distal aliphatic tail of JWH133 differs from that of AM12033 due to the lack of a functional group at the end. As in the case of AM12033 the so-called twin toggle switch residues F3.36 and W6.48 (Hua et al., 2020) are stabilized in their active conformation as shown in Supplementary Figure S3A. In TRPV1, JWH133 cleared both HTVS and XP screening in what is thought to be the CBD binding site. CBD has yet to be co-resolved with TRPV1, though it has with TRPV2 and an analysis of the putative CBD binding site was performed across all ICRs (Muller et al., 2020). The CBD structure in TRPV2 displays a different orientation than the CBD screen at TRPV1, though the differences cited above could be responsible. CBD and JWH133 show similar π-π stacking with Y584, though CBD is also stabilized by F639, likely due to the central constraint of JWH133 which angles the ligand slightly outward (Supplementary Figure S3B).
3.2 Towards the Identification of Potential Dual Ligands
In order to identify potential chemotypes with a yet unexplored TRPV1/CB2 dual profile two different in silico approaches have been followed. These strategies are described in the following subsections and the workflows are depicted in Supplementary Figure S2.
3.2.1 Virtual Screening of JWH133 Structurally Related Chemical Databases
A chemical library of compounds with >0.85 Tanimoto similarity index with JWH133 was curated and screened at CB2 and TRPV1 using the methods described above (workflow depicted in Supplementary Figure S2A). Analysis of docking interactions of top-ranked XP results from the CB2 site and the TRPV1 sites revealed seventeen common ligands between the CB2 site and the VBP and CBD sites (Table 1). No common ligands were identified between the CB2 site and the TRPV1 tunnel. The selection of novel potential CB2/TRPV1 chemotypes includes key structural features and ligand-receptor site interactions at both targets, as well as the absence of previously reported activity at these receptors. This strategy allows for prioritization of molecularly diverse and novel compounds. To ensure the VBP and CBD sites were explored equally, two ligands were selected that targeted CB2 and the VBP (57756957 and 151332252), two ligands were selected that targeted CB2 and the CBD site (59824268 and 123533625), and one ligand was selected that targeted CB2, the VBP, and the CBD site (153641693), resulting in the selection of five ligands for further investigation via manual docking and pharmacokinetic profiling (Supplementary Table S2). The selected chemotypes have not been yet explored at CB2/TRPV1 and their reported activity is not significant, providing novel opportunities for the investigation of the endocannabinoid system.
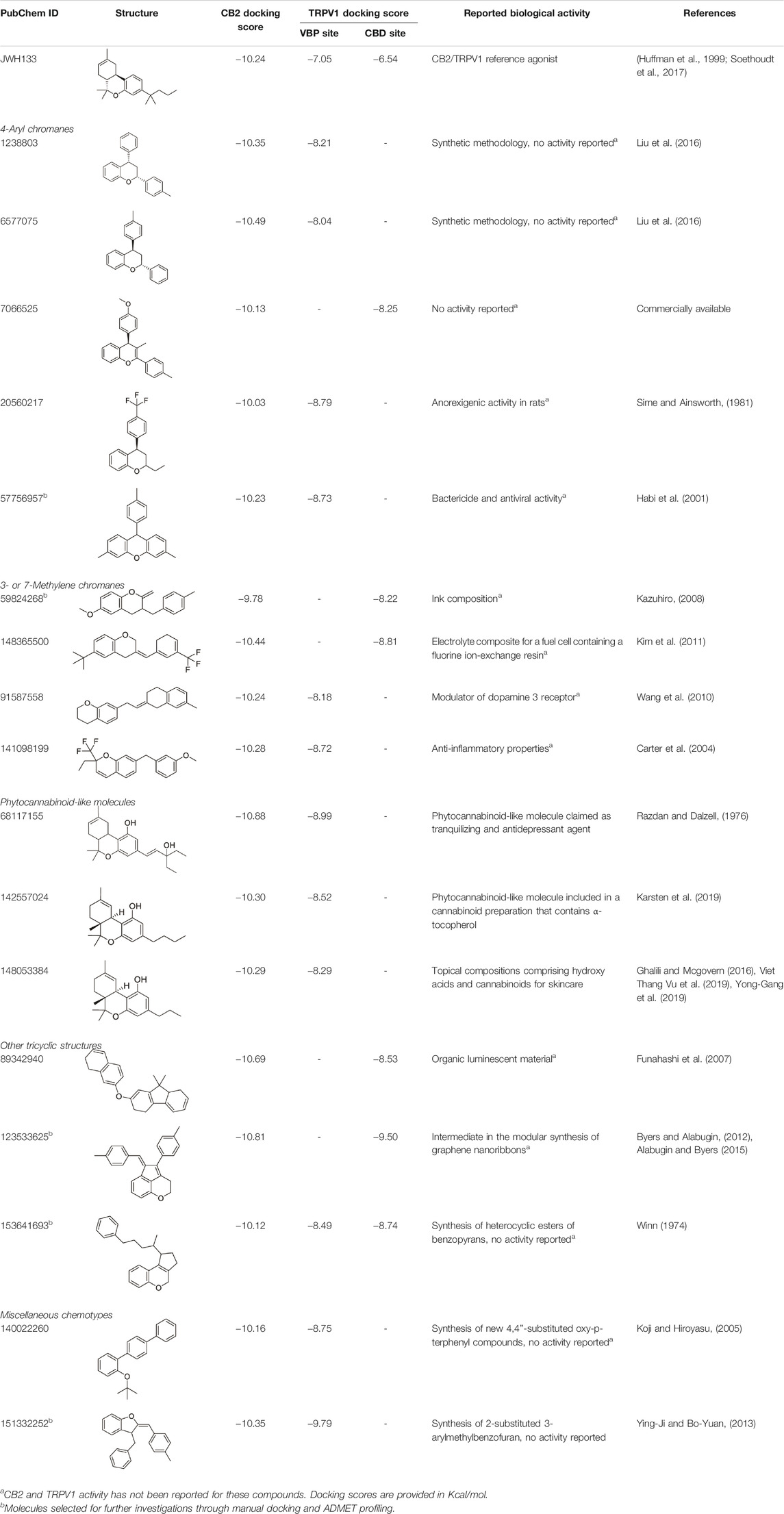
TABLE 1. Potential dual CB2/TRPV1 candidates obtained upon screening of a JWH133 structurally related chemical database. Selected hits have been classified according to common structural moieties.
Among the five selected candidates, 3-(4-methylbenzyl)-chromane 59824268 presented a better druggability profile (Supplementary Table S2) being therefore prioritized for future in vitro testing as CB2/TRPV1 dual modulator. As shown in Supplementary Table S2, candidates 57756957, 123533625, 153641693, and 151332252 exhibit HERG values that fall outside the range of approved drugs. Docking studies of 59824268 at CB2 and TRPV1 are shown in Figure 1. At CB2 this chromane sits a bit higher than JWH133 in the binding crevice being stabilized by hydrophobic and aromatic interactions with residues F2.64, F2.61, F2.57, F183 and F7.35. Regarding TRPV1, 59824268 orients itself in a way similar to JWH133 in the pocket maintaining overlap with the aromatic ring. While JWH133 appears to have primary interactions with Y584, 59824268 has interactions with Y584 in addition to Y632 and F639, further stabilizing the chromane in this pocket.
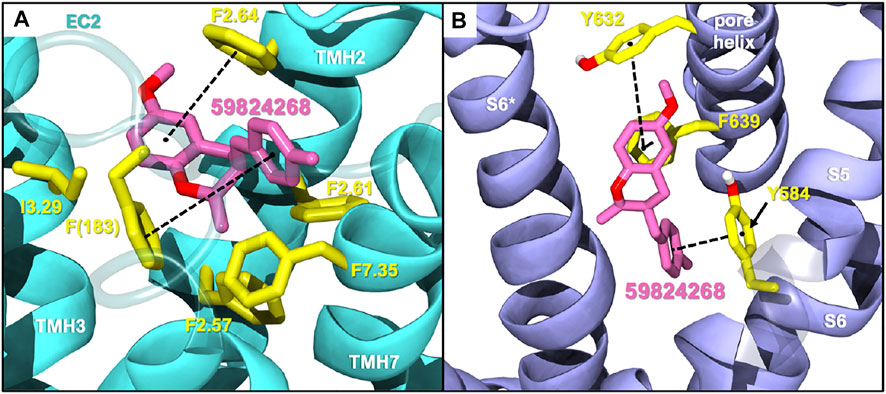
FIGURE 1. Selected compound 59824268 docked in CB2 (A) and TRPV1 (B). EC2: Extracellular loop 2; TMH, transmembrane helix.
3.2.2 Cross-Agonist Virtual Screening
The second strategy for the identification of dual compounds is based on a HTVS of reported CB2 and TRPV1 agonists. CB2 agonists indexed in the ChEMBL database have been retrieved and studied in the three known TRPV1 binding sites as detailed in section 2.4. Likewise, TRPV1 ligands indexed in the ChEMBL database have been retrieved and studied in the CB2 binding site. Following the workflow depicted in Supplementary Figure S2B, five candidates were selected for further analysis at each receptor (Table 2). Reported activity at the known target, docking score (Table 2) and druggability profile (Supplementary Table S3) led us to select compounds 1288208, 1288239 (TRPV1 virtual screening) 1508577 and 1508215 (CB2 virtual screening).
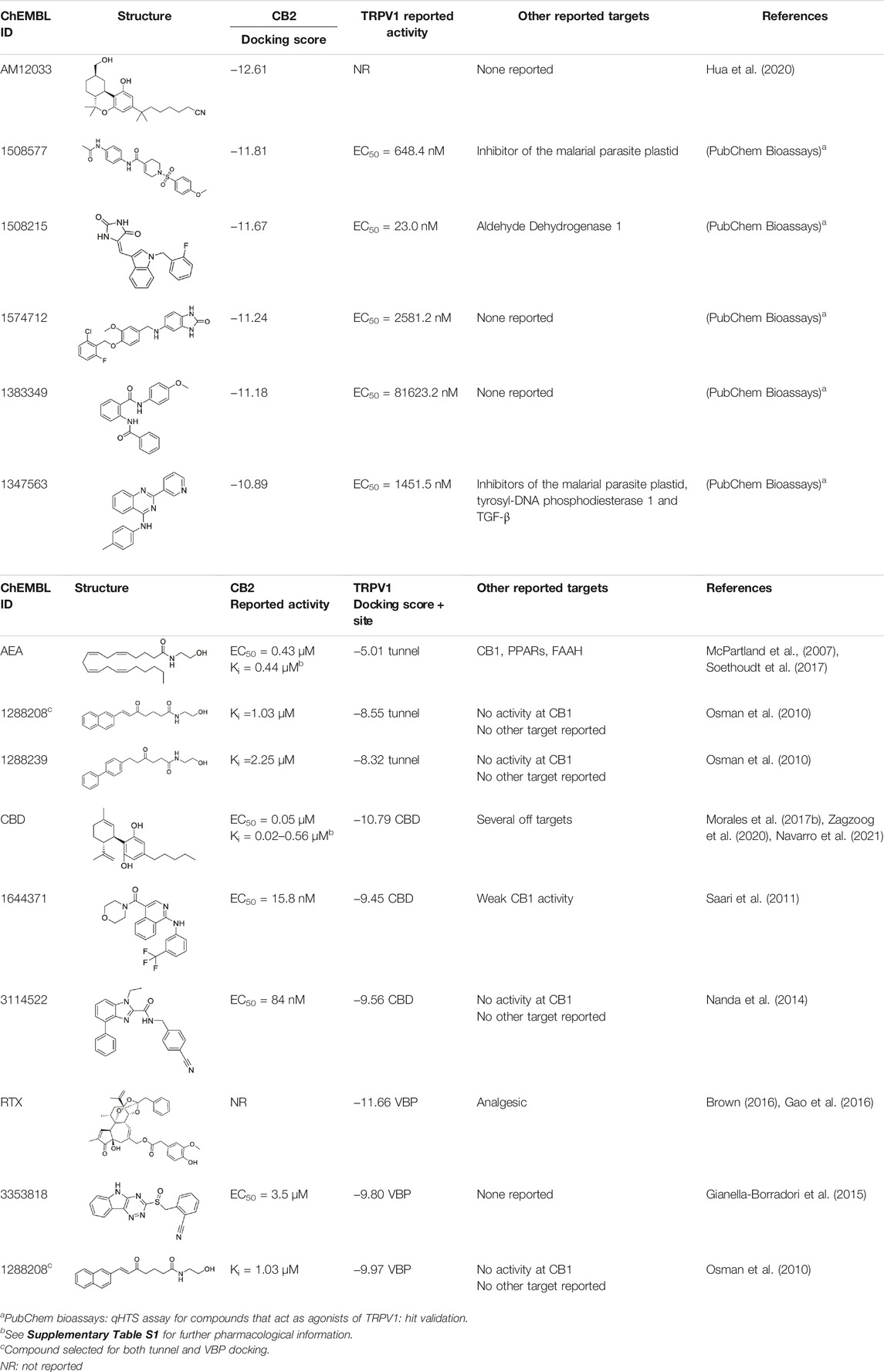
TABLE 2. Potential dual CB2/TRPV1 candidates obtained through the crossed-agonist strategy.
1288208 passed the screening as a potential modulator of TRPV1 at two sites: the VBP and tunnel. While there is argument for the elimination of this ligand due to lack of site specificity, it was selected for exactly this reason. With the abundance of ligands reported to modulate TRPV1, and the variability in reported and putative binding locations, a ligand that shows the potential for interaction at multiple locations, both putative and confirmed, within the channel is worthy of further study to better understand why this is. In the VBP, the headgroup of 1288208 forms H-bonds with R557 and S512 via the backbone and hydroxy group, both reachable from within the tunnel. The α,β-unsaturated ketone oxygen H-bonds with Y511, and the addition of the naphthyl moiety at the tail end of the ligand provides pi-stacking capabilities farther up in the VBP with F543 and F591 (Figure 2A).
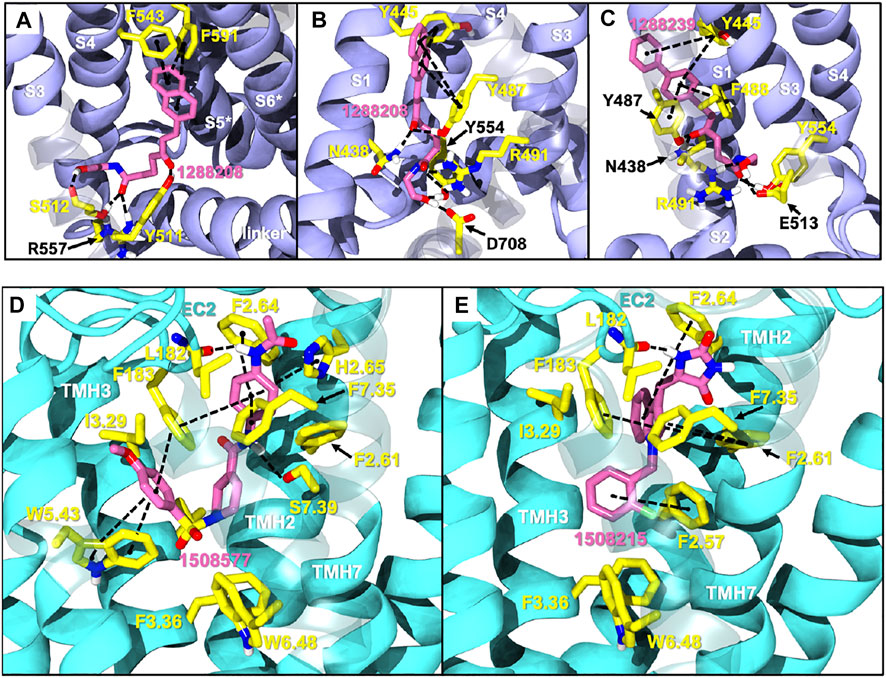
FIGURE 2. Docks of selected potential dual candidates: TRPV1 in purple cartoon ribbons (A–C) and CB2 in cyan cartoon ribbons (D,E). Molecules are displayed in pink tubes; all interactions are shown via dashed lines and each helix and residue is labeled. (A) shows 1288208 in the VBP. A portion of S3 is transparent to aid in visibility; (B) shows 1288208 in the tunnel. Helix S2 is shown completely transparent to aid in the visibility of the tunnel; (C) shows 1288239 in the tunnel with a portion of helix S2 transparent to aid in visibility. (D) shows a lipid view of the 1508577/CB2 complex; (E) shows a lipid view of the 1508215/CB2 complex; TMH6 and 7 are displayed with transparency for a clearer view of the binding site.
AEA docking in the tunnel shows headgroup interactions with several residues including Y554, Y555, Y487, D708, and N438 (Supplementary Figure S4). Like AEA, the hydroxyl portion of the headgroup of 1288208 maintains interactions with Y554 and D708, while additional H-bonding between R491 and the amide oxygen is present. The inclusion of an α,β-unsaturated ketone mid-tail allows for more H-bonding via Y487 and N438 near the entrance of the tunnel. The naphthyl moiety at the end of the tail displays pi-pi interactions with both Y487 and Y445 (Figure 2B). The additional interactions of 1288208 could aid in the stability of the ligand in the tunnel from an external standpoint, allowing the headgroup more time in the tunnel, potentially triggering channel activation as previously hypothesized from MD simulations (Muller et al., 2020, 2021).
The other selected TRPV1 ligand, 1288239 (Figure 2C), shows headgroup interactions with R491 and Y554, like 1288208, with an additional interaction with E513. The ketone found midway down the tail of ligand H-bonds with N438 and Y487, again similar to 1288208. One feature that differentiates 1288239 from 1288208 is a biphenyl moiety in place of a naphthyl moiety. The lower ring of the biphenyl moiety has aromatic interactions with Y487 and F488, and both rings interact with Y445.
Because of their high potency at TRPV1, their interaction pattern at the CB2 orthosteric pocket and their optimal drug-like properties, compounds 1508577 and 1508215 were selected as potential candidates in the in silico search of dual ligands. Compounds like ACPA-OH and JWH133 also ranked at the top, however, since we are looking for unexplored dual chemotypes, they were not selected in this in silico study. Consistent with the hydrophobic nature of the CB2 orthosteric pocket, compound 1508577 is mainly stabilized by aromatic and van der Waals interactions. As displayed in Figure 2D, π-π stackings are stablished between the methoxybenzene group with W5.43 and F183 and the phenylacetamide group with F183, F7.35, F2.64 and H2.65. Moreover, the acetamide hydrogen engages with the backbone carbonyl oxygen of V182 in a H-bond while the central amide H-bonds with S7.39. Compound 1508215 orients similarly in the binding crevice stablishing aromatic π-π interactions between the central indole core and F183, F2.61 and F2.64, and the fluorobenzyl group with F2.57. In addition, the imidazolidinedione group H-bonds with the backbone carbonyl oxygen of V182.
In summation, from this approach, compounds 1288208, 1288239, 1508577, and 1508215 have been selected for future in vitro appraisal as dual CB2/TRPV1 agonists. Other compounds such as 1644371 could also be remarkable candidates for testing at TRPV1 due to its nanomolar agonist potency at CB2.
3.2.3 Off-Target Evaluation
The selected hits (59824268, 1288208, 1288239, 1508577 and 1508215) have also been docked in related receptors in order to identify potential off-target effects. These molecules have been screened at CB1 and the cannabinoid-related GPCRs GPR55 and GPR18 in their active and inactive states. In addition, cannabinoid-related channels including TRPV2, TRPV3, TRPA1, and TRPM8 have also been assessed. As shown in Supplementary Table S5, by comparing docking scores to their reference orthosteric ligands we can conclude that at cannabinoid GPCRs compounds 1288208 and 1288239 might be more promiscuous showing high interaction energies at the GPR55 active and GPR18 inactive models. Moreover, compounds 1508577 and 1508215 may moderately act at CB1 whereas 1288208 and 1288239 were reported to lack binding affinity (Osman et al., 2010). 59824268 may be less selective with higher energies for the apo TRPV3 structure as well as both TRPA1 structures. 1288208 and 1288239 both show energies that are either comparable to or better than the reference ligand for each respective receptor, perhaps suggesting that the ethanolamide head group may be too promiscuous of a moiety to include when aiming to develop ligands for selective dual targeting. 1508577 shows variable activity across the TRP channels with comparable energies to the reference compounds of TRPV3 and TRPA1 in both states, with 1508215 displaying the potential for promiscuity at TRPA1.In light of these results, compounds 59824268, 1508577 and 1508215 could be prioritized as TRPV1/CB2 dual modulators. However, compounds with moderate activity at other cannabinoid targets could also be beneficial when targeting specific pathologies in which the ECS is involved or avoided when searching for more selective cannabinoid modulators.
Nonetheless, off-targets cannot be completely ruled out and not only cannabinoid-related but also other receptor families should be tested experimentally at further stages of this project.
4 Conclusion
Three-dimensional crystal and cryo-EM structures of GPCRs and TRP channels are being resolved at a rapid pace in the last years. The resolution of these structures are showing great impact in the field of drug discovery facilitating the emergence of successful in silico strategies for the identification of potential drugs targeting complex physiopathological processes.
The ECS is composed by a variety of receptors including GPCRs, TRP channels, nuclear receptors such as the PPARs (Morales et al., 2017b). Polypharmacological approaches targeting this system have already shown successful results (Fernández-Fernández et al., 2014; Malek and Starowicz, 2016; Barutta et al., 2017; Lago-Fernandez et al., 2021). For instance, a PPARγ-CB2 molecule has entered clinical trials for the treatment of systemic and multiple sclerosis (EHP-101, 2020; Palomares et al., 2018).
In this context, synergistic effects between TRPV1 and CBRs offer novel avenues for the management of pain or neurodegenerative pathologies among others. While CB1/TRPV1 dual modulators have been further studied, CB2/TRPV1 agonists have not been yet exploited. Therefore, this brief research article addresses the computational search of novel potential dual candidates for further in vitro and in vivo exploration.
Using two different virtual screening approaches we have identified hits with potential dual agonistic activity taking into account reported data and docking and druggability results. From this study, compounds 59824268, 1288208, 1288239, 1508577 and 1508215 (Supplementary Figure S5) are proposed as main candidates for future experimental appraisal. Other selected molecules reported in this article also present interesting profiles and might be worth exploring. These results provide insights into understudied scaffolds that potentially modulate CB2 and TRPV1 providing novel tools for further studies.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: PubChem: https://pubchem.ncbi.nlm.nih.gov/ChEMBL: https://www.ebi.ac.uk/chembl.
Author Contributions
Development of study: CM and PM. Data analysis: CM and PM. Interpretation of results: CM, PM, NJ, and PR Manuscript preparation and review: CM, PM, NJ, and PR All authors reviewed, revised, and approved the manuscript for submission.
Funding
This research was funded by the Spanish Ministry of Science and Innovation, grant number MICIU/FEDER: RTI 2018-095544-B-I00 and the Spanish National Research Council (CSIC), grant number PIE-201580E033, and the National Institute on Drug Abuse grant R01 DA003934 (PR) and F31 DA53022 (CM).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
PM acknowledges the Spanish Ministry of Science and Innovation for her postdoctoral fellow from the Juan de la Cierva Incorporación Programme-MICIU (IJC 2019-042182-I).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2022.841190/full#supplementary-material
References
Adamczyk, P., Miszkiel, J., McCreary, A. C., Filip, M., Papp, M., and Przegaliński, E. (2012). The Effects of Cannabinoid CB1, CB2 and Vanilloid TRPV1 Receptor Antagonists on Cocaine Addictive Behavior in Rats. Brain Res. 1444, 45–54. doi:10.1016/j.brainres.2012.01.030
Alabugin, I. V., and Byers, P. M. (2015). Modular Synthesis of Graphene Nanoribbons and Graphene Substructures from Oligo-Alkynes.
Aly, E., and Masocha, W. (2021). Targeting the Endocannabinoid System for Management of HIV-Associated Neuropathic Pain: A Systematic Review. IBRO Neurosci. Rep. 10, 109–118. doi:10.1016/j.ibneur.2021.01.004
Anthony, A. T., Rahmat, S., Sangle, P., Sandhu, O., and Khan, S. (2020). Cannabinoid Receptors and Their Relationship with Chronic Pain: A Narrative Review. Cureus 12 (9), e10436. doi:10.7759/cureus.10436
Appendino, G., Cascio, M. G., Bacchiega, S., Moriello, A. S., Minassi, A., Thomas, A., et al. (2006). First "hybrid" Ligands of Vanilloid TRPV1 and Cannabinoid CB2receptors and Non-polyunsaturated Fatty Acid-Derived CB2-Selective Ligands. FEBS Lett. 580, 568–574. doi:10.1016/j.febslet.2005.12.069
Appendino, G., Ligresti, A., Minassi, A., Cascio, M. G., Allarà, M., Taglialatela-Scafati, O., et al. (2009). Conformationally Constrained Fatty Acid Ethanolamides as Cannabinoid and Vanilloid Receptor Probes. J. Med. Chem. 52, 3001–3009. doi:10.1021/jm900130m
Arnold, J. C., Hone, P., Holland, M. L., and Allen, J. D. (2012). CB2 and TRPV1 Receptors Mediate Cannabinoid Actions on MDR1 Expression in Multidrug Resistant Cells. Pharmacol. Rep. 64, 751–757. doi:10.1016/S1734-1140(12)70871-X
Arnold, W. R., Carnevale, L. N., Xie, Z., Baylon, J. L., Tajkhorshid, E., Hu, H., et al. (2021). Anti-inflammatory Dopamine- and Serotonin-Based Endocannabinoid Epoxides Reciprocally Regulate Cannabinoid Receptors and the TRPV1 Channel. Nat. Commun. 12. doi:10.1038/s41467-021-20946-6
Aso, E., and Ferrer, I. (2016). CB2 Cannabinoid Receptor as Potential Target against Alzheimer's Disease. Front. Neurosci. 10, 1–10. doi:10.3389/fnins.2016.00243
Assimakopoulou, M., Pagoulatos, D., Nterma, P., and Pharmakakis, N. (2017). Immunolocalization of Cannabinoid Receptor Type 1 and CB2 Cannabinoid Receptors, and Transient Receptor Potential Vanilloid Channels in Pterygium. Mol. Med. Rep. 16, 5285–5293. doi:10.3892/mmr.2017.7246
Avraham, Y., Davidi, N., Porat, M., Chernoguz, D., Magen, I., Vorobeiv, L., et al. (2010). Leptin Reduces Infarct Size in Association with Enhanced Expression of CB2, TRPV1, SIRT-1 and Leptin Receptor.doi:10.2174/156720210791184943
Baell, J. B., and Holloway, G. A. (2010). New Substructure Filters for Removal of pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 53, 2719–2740. doi:10.1021/jm901137j
Barutta, F., Grimaldi, S., Gambino, R., Vemuri, K., Makriyannis, A., Annaratone, L., et al. (2017). Dual Therapy Targeting the Endocannabinoid System Prevents Experimental Diabetic Nephropathy. Nephrol. Dial. Transpl. 32, 1655–1665. doi:10.1093/ndt/gfx010
Behl, T., Kaur, G., Bungau, S., Jhanji, R., Kumar, A., Mehta, V., et al. (2020). Distinctive Evidence Involved in the Role of Endocannabinoid Signalling in Parkinson's Disease: A Perspective on Associated Therapeutic Interventions. Ijms 21, 1–27. doi:10.3390/ijms21176235
Bellini, G., Torella, M., Manzo, I., Tortora, C., Luongo, L., Punzo, F., et al. (2017). Pkcβii-Mediated Cross-Talk of TRPV1/CB2 Modulates the Glucocorticoid-Induced Osteoclast Overactivity. Pharmacol. Res. 115, 267–274. doi:10.1016/j.phrs.2016.11.039
Berry, A. J., Zubko, O., Reeves, S. J., and Howard, R. J. (2020). Endocannabinoid System Alterations in Alzheimer's Disease: A Systematic Review of Human Studies. Brain Res. 1749, 147135. doi:10.1016/j.brainres.2020.147135
Bhatta, P., Dhukhwa, A., Sheehan, K., Al Aameri, R. F. H., Borse, V., Ghosh, S., et al. (2019). Capsaicin Protects against Cisplatin Ototoxicity by Changing the STAT3/STAT1 Ratio and Activating Cannabinoid (CB2) Receptors in the Cochlea. Sci. Rep. 9, 1–16. doi:10.1038/s41598-019-40425-9
Brooks, J. W., Pryce, G., Bisogno, T., Jaggar, S. I., Hankey, D. J., Brown, P., et al. (2002). Arvanil-induced Inhibition of Spasticity and Persistent Pain: Evidence for Therapeutic Sites of Action Different from the Vanilloid VR1 Receptor and Cannabinoid CB(1)/CB(2) Receptors. Eur. J. Pharmacol. 439, 83–92. Available at: www.elsevier.com/locate/ejphar (Accessed April 28, 2021). doi:10.1016/s0014-2999(02)01369-9
Brown, D. (2016). Resiniferatoxin: The Evolution of the “Molecular Scalpel” for Chronic Pain Relief. Pharmaceuticals 9, 47–11. doi:10.3390/ph9030047
Bryk, M., and Starowicz, K. (2021). Cannabinoid-based Therapy as a Future for Joint Degeneration. Focus on the Role of CB2 Receptor in the Arthritis Progression and Pain: an Updated Review. Pharmacol. Rep. 73, 681–699. doi:10.1007/s43440-021-00270-y
Byers, P. M., and Alabugin, I. V. (2012). Polyaromatic Ribbons from Oligo-Alkynes via Selective Radical Cascade: Stitching Aromatic Rings with Polyacetylene Bridges. J. Am. Chem. Soc. 134, 9609–9614. doi:10.1021/ja3023626
Capuzzi, S. J., Muratov, E. N., and Tropsha, A. (2017). Phantom PAINS: Problems with the Utility of Alerts for Pan-Assay Interference Compounds. J. Chem. Inf. Model. 57, 417–427. doi:10.1021/acs.jcim.6b00465
Carter, J. S., William, A. K., Brown, D. L., Deprow, A., Fletcher, T., Hallinan, E. A., et al. (2004). Benzopyran Compounds Useful for Treating Inflammatory Conditions.
Cassano, T., Calcagnini, S., Pace, L., De Marco, F., Romano, A., and Romano, S. (2017). Cannabinoid Receptor 2 Signaling in Neurodegenerative Disorders: From Pathogenesis to a Promising Therapeutic Target. Front. Neurosci. 11, 1–10. doi:10.3389/fnins.2017.00030
Caterina, M. J., Schumacher, M. A., Tominaga, M., Rosen, T. A., Levine, J. D., and Julius, D. (1997). The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 389, 816–824. doi:10.1038/39807
Cécyre, B., Bachand, I., Papineau, F., Brochu, C., Casanova, C., and Bouchard, J.-F. (2020). Cannabinoids Affect the Mouse Visual Acuity via the Cannabinoid Receptor Type 2. Sci. Rep. 10, 1–13. doi:10.1038/s41598-020-72553-y
Chávez, A. E., Chiu, C. Q., and Castillo, P. E. (2010). TRPV1 Activation by Endogenous Anandamide Triggers Postsynaptic Long-Term Depression in Dentate Gyrus. Nat. Neurosci. 13, 1511–1518. doi:10.1038/nn.2684
Cheong, S. L., Federico, S., Spalluto, G., Klotz, K.-N., and Pastorin, G. (2019). The Current Status of Pharmacotherapy for the Treatment of Parkinson's Disease: Transition from Single-Target to Multitarget Therapy. Drug Discov. Today 24, 1769–1783. doi:10.1016/j.drudis.2019.05.003
Cristino, L., de Petrocellis, L., Pryce, G., Baker, D., Guglielmotti, V., and Di Marzo, V. (2006). Immunohistochemical Localization of Cannabinoid Type 1 and Vanilloid Transient Receptor Potential Vanilloid Type 1 Receptors in the Mouse Brain. Neuroscience 139, 1405–1415. doi:10.1016/j.neuroscience.2006.02.074
Daina, A. O., Olivier, M., and Michielin, V. (2017). SwissADME: a Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 7, 42717. doi:10.1038/srep42717
De Lago, E., Urbani, P., Ramos, J. A., Di Marzo, V., and Fernández-Ruiz, J. (2005). Arvanil, a Hybrid Endocannabinoid and Vanilloid Compound, Behaves as an Antihyperkinetic Agent in a Rat Model of Huntington's Disease. Brain Res. 1050, 210–216. doi:10.1016/j.brainres.2005.05.024
De Petrocellis, L., Ligresti, A., Moriello, A. S., Allarà, M., Bisogno, T., Petrosino, S., et al. (2011). Effects of Cannabinoids and Cannabinoid-enrichedCannabisextracts on TRP Channels and Endocannabinoid Metabolic Enzymes. Br. J. Pharmacol. 163, 1479–1494. doi:10.1111/j.1476-5381.2010.01166.x
Di Marzo, V., Bisogno, T., De Petrocellis, L., Brandi, I., Jefferson, R. G., Winckler, R. L., et al. (2001). Highly Selective CB1 Cannabinoid Receptor Ligands and Novel CB1/VR1 Vanilloid Receptor "Hybrid" Ligands. Biochem. Biophysical Res. Commun. 281, 444–451. doi:10.1006/bbrc.2001.4354
Di Marzo, V., Griffin, G., De Petrocellis, L., Brandi, I., Bisogno, T., Williams, W., et al. (2002). A Structure/activity Relationship Study on Arvanil, an Endocannabinoid and Vanilloid Hybrid. J. Pharmacol. Exp. Ther. 300, 984–991. doi:10.1124/jpet.300.3.984
Diver, M. M., Cheng, Y., and Julius, D. (2019). Structural Insights into TRPM8 Inhibition and Desensitization. Science 365, 1434–1440. doi:10.1126/science.aax6672
Du, Y., Fu, M., Huang, Z., Tian, X., Li, J., Pang, Y., et al. (2020). TRPV1 Activation Alleviates Cognitive and Synaptic Plasticity Impairments through Inhibiting AMPAR Endocytosis in APP23/PS45 Mouse Model of Alzheimer's Disease. Aging Cell 19, e13113. doi:10.1111/acel.13113
EHP-101 (2020). Available at https://clinicaltrials.gov/ct2/results?cond=&term=EHP-101&cntry=&state=&city=&dist= (Accessed December 20, 2020).
Fowler, C. J. (2020). The Endocannabinoid System - Current Implications for Drug Development. J. Intern. Med. 290, 2–26. doi:10.1111/joim.13229
Gao, Y., Cao, E., Julius, D., and Cheng, Y. (2016). TRPV1 Structures in Nanodiscs Reveal Mechanisms of Ligand and Lipid Action. Nature 534, 347–351. doi:10.1038/nature17964
Ghalili, B., and Mcgovern, K. (2016). Topical Compositions Comprising Hydroxy Acids and Cannabinoids for Skin Care.Available at. https://lens.org/089-999-413-301-246%0Ahttps://lens.org/150-825-718-678-650%0Ahttps://lens.org/148-818-296-970-602.
Gianella-Borradori, M., Christou, I., Bataille, C. J. R., Cross, R. L., Wynne, G. M., Greaves, D. R., et al. (2015). Ligand-based Virtual Screening Identifies a Family of Selective Cannabinoid Receptor 2 Agonists. Bioorg. Med. Chem. 23, 241–263. doi:10.1016/j.bmc.2014.11.002
Gibson, H. E., Edwards, J. G., Page, R. S., Van Hook, M. J., and Kauer, J. A. (2008). TRPV1 Channels Mediate Long-Term Depression at Synapses on Hippocampal Interneurons. Neuron 57, 746–759. doi:10.1016/j.neuron.2007.12.027
Gontijo, V. S., Viegas, F. P. D., Ortiz, C. J. C., de Freitas Silva, M., Damasio, C. M., Rosa, M. C., et al. (2020). Molecular Hybridization as a Tool in the Design of Multi-Target Directed Drug Candidates for Neurodegenerative Diseases. Cn 18, 348–407. doi:10.2174/1385272823666191021124443
González-Aparicio, R., and Moratalla, R. (2014). Oleoylethanolamide Reduces L-DOPA-Induced Dyskinesia via TRPV1 Receptor in a Mouse Model of Parkinson´s Disease. Neurobiol. Dis. 62, 416–425. doi:10.1016/j.nbd.2013.10.008
Guindon, J., and Hohmann, A. G. (2008). Cannabinoid CB2 Receptors: a Therapeutic Target for the Treatment of Inflammatory and Neuropathic Pain. Br. J. Pharmacol. 153, 319–334. doi:10.1038/sj.bjp.0707531
Habi, A., Gravel, D., Villenueve, L., Su, H., and Vaillancourt, M. (2001). Halogenated Rhodamine Derivatives and Applications Thereof.
Hashiesh, H. M., Sharma, C., Goyal, S. N., Jha, N. K., and Ojha, S. (2021). Pharmacological Properties, Therapeutic Potential and Molecular Mechanisms of JWH133, a CB2 Receptor-Selective Agonist. Front. Pharmacol. 12, 1–34. doi:10.3389/fphar.2021.702675
Ho, K. W., Ward, N. J., and Calkins, D. J. (2012). TRPV1: a Stress Response Protein in the central Nervous System. Am. J. Neurodegener. Dis. 1, 1–14. Available at www.AJND.us (Accessed April 27, 2021).
Hua, T., Li, X., Wu, L., Iliopoulos-Tsoutsouvas, C., Wang, Y., Wu, M., et al. (2020). Activation and Signaling Mechanism Revealed by Cannabinoid Receptor-Gi Complex Structures. Cell 180, 655–665.e18. doi:10.1016/j.cell.2020.01.008
Huffman, J. W., Liddle, J., Yu, S., Aung, M. M., Abood, M. E., Wiley, J. L., et al. (1999). 3-(1′,1′-Dimethylbutyl)-1-deoxy-Δ 8 -THC and Related Compounds: Synthesis of Selective Ligands for the CB 2 Receptor. Bioorg. Med. Chem. 7, 2905–2914. doi:10.1016/S0968-0896(99)00219-9
Jagerovic, N., Fernández-Fernández, C., Erdozain, A. M., Girón, R., Sánchez, E., López-Moreno, J. A., et al. (2014). Combining Rimonabant and Fentanyl in a Single Entity: Preparation and Pharmacological Results. Dddt 8, 263–277. doi:10.2147/DDDT.S55045
Javed, H., Azimullah, S., Haque, M. E., and Ojha, S. K. (2016). Cannabinoid Type 2 (CB2) Receptors Activation Protects against Oxidative Stress and Neuroinflammation Associated Dopaminergic Neurodegeneration in Rotenone Model of Parkinson's Disease. Front. Neurosci. 10, 1–14. doi:10.3389/fnins.2016.00321
Karsten, Mäder., Weiss, Verena., and Jörg Kressler, T. N. (2019). Cannabinoid Composition and Products Including Alpha-Tocopherol. Available at https://patentscope.wipo.int/search//iasr?ia=DE2015000204&PAGE=PDF&ACCESS=screen&TOK=OLj7dxliHXGZmPch65OIlysSXCg.
Kazuhiro, Y. (2008). Ink Composition, Inkjet Recording Method, Printed Material, Production Method of a Planographic Printng Plate and Planographic Printing Plate. 1, 11–14.
Kim, K., Heo, P., Pak, C., and Choi, S. (2011). Composition, Composite Prepared from the Composition, Electrode Using the Composition or the Composite, Composite Membrane Including the Composite, and Fuel Cell Including the Composite Membrane. Available at https://patents.google.com/patent/EP2595229B1.
Kong, W.-L., Peng, Y.-Y., and Peng, B.-W. (2017). Modulation of Neuroinflammation: Role and Therapeutic Potential of TRPV1 in the Neuro-Immune axis. Brain Behav. Immun. 64, 354–366. doi:10.1016/j.bbi.2017.03.007
Kotsikorou, E., Sharir, H., Shore, D. M., Hurst, D. P., Lynch, D. L., Madrigal, K. E., et al. (2013). Identification of the GPR55 Antagonist Binding Site Using a Novel Set of High-Potency GPR55 Selective Ligands. Biochemistry 52, 9456–9469. Available at. http://pubs.acs.org/doi/abs/10.1021/bi4008885 (Accessed January 28, 2014)doi:10.1021/bi4008885
Lago-Fernandez, A., Zarzo-Arias, S., Jagerovic, N., and Morales, P. (2021). Relevance of Peroxisome Proliferator Activated Receptors in Multitarget Paradigm Associated with the Endocannabinoid System. Ijms 22, 1001. doi:10.3390/ijms22031001
Lastres-Becker, I., De Miguel, R., De Petrocellis, L., Makriyannis, A., Di Marzo, V., and Fernández-Ruiz, J. (2003). Compounds Acting at the Endocannabinoid And/or Endovanilloid Systems Reduce Hyperkinesia in a Rat Model of Huntington's Disease. J. Neurochem. 84, 1097–1109. doi:10.1046/j.1471-4159.2003.01595.x
Li, M., Zhu, M., Xu, Q., Ding, F., Tian, Y., and Zhang, M. (2020). Sensation of TRPV1 via 5-hydroxytryptamine Signaling Modulates Pain Hypersensitivity in a 6-hydroxydopamine Induced Mice Model of Parkinson's Disease. Biochem. Biophysical Res. Commun. 521, 868–873. doi:10.1016/j.bbrc.2019.10.204
Lingerfelt, M. A., Zhao, P., Sharir, H. P., Hurst, D. P., Reggio, P. H., and Abood, M. E. (2017). Identification of Crucial Amino Acid Residues Involved in Agonist Signaling at the GPR55 Receptor. Biochemistry 56, 473–486. doi:10.1021/acs.biochem.6b01013
Liu, J., Wang, X., Xu, L., Hao, Z., Wang, L., and Xiao, J. (2016). One Step Synthesis of 2-alkenylchromanes via Inverse Electron-Demand Hetero-Diels-Alder Reaction of O -quinone Methide with Unactivated Dienes. Tetrahedron 72, 7642–7649. doi:10.1016/j.tet.2016.10.027
Lowin, T., Pongratz, G., and Straub, R. H. (2016). The Synthetic Cannabinoid WIN55,212-2 Mesylate Decreases the Production of Inflammatory Mediators in Rheumatoid Arthritis Synovial Fibroblasts by Activating CB2, TRPV1, TRPA1 and yet Unidentified Receptor Targets. J. Inflamm. 13, 1–10. doi:10.1186/s12950-016-0114-7
Lowin, T., and Straub, R. H. (2015). Cannabinoid-based Drugs Targeting CB1 and TRPV1, the Sympathetic Nervous System, and Arthritis. Arthritis Res. Ther. 17, 226. doi:10.1186/s13075-015-0743-x
Maione, S., Cristino, L., Migliozzi, A. L., Georgiou, A. L., Starowicz, K., Salt, T. E., et al. (2009). TRPV1 Channels Control Synaptic Plasticity in the Developing superior Colliculus. J. Physiol. 587, 2521–2535. doi:10.1113/jphysiol.2009.171900
Malek, N., and Starowicz, K. (2016). Dual-acting Compounds Targeting Endocannabinoid and Endovanilloid Systems-A Novel Treatment Option for Chronic Pain Management. Front. Pharmacol. 7, 1–9. doi:10.3389/fphar.2016.00257
Maramai, S., Benchekroun, M., Gabr, M. T., and Yahiaoui, S. (2020). Multitarget Therapeutic Strategies for Alzheimer's Disease: Review on Emerging Target Combinations. Biomed. Res. Int. 2020, 1–27. doi:10.1155/2020/5120230
Marinelli, S., Di Marzo, V., Berretta, N., Matias, I., Maccarrone, M., Bernardi, G., et al. (2003). Presynaptic Facilitation of Glutamatergic Synapses to Dopaminergic Neurons of the Rat Substantia Nigra by Endogenous Stimulation of Vanilloid Receptors. J. Neurosci. 23, 3136–3144. doi:10.1523/jneurosci.23-08-03136.2003
Martínez-Pinilla, E., Varani, K., Reyes-Resina, I., Angelats, E., Vincenzi, F., Ferreiro-Vera, C., et al. (2017). Binding and Signaling Studies Disclose a Potential Allosteric Site for Cannabidiol in Cannabinoid CB2 Receptors. Front. Pharmacol. 8, 1–10. doi:10.3389/fphar.2017.00744
Martins, D., Tavares, I., and Morgado, C. (2014). “hotheaded”: The Role of TRPV1 in Brain Functions. Neuropharmacology 85, 151–157. doi:10.1016/j.neuropharm.2014.05.034
McPartland, J. M., Glass, M., and Pertwee, R. G. (2007). Meta-analysis of Cannabinoid Ligand Binding Affinity and Receptor Distribution: Interspecies Differences. Br. J. Pharmacol. 152, 583–593. doi:10.1038/sj.bjp.0707399
Melck, D., Bisogno, T., De Petrocellis, L., Chuang, H.-h., Julius, D., Bifulco, M., et al. (1999). Unsaturated Long-Chain N-Acyl-Vanillyl-Amides (N-AVAMs): Vanilloid Receptor Ligands that Inhibit Anandamide-Facilitated Transport and Bind to CB1 Cannabinoid Receptors. Biochem. Biophysical Res. Commun. 262, 275–284. doi:10.1006/bbrc.1999.1105
Mlost, J., Kostrzewa, M., Borczyk, M., Bryk, M., Chwastek, J., Korostyński, M., et al. (2021). CB2 Agonism Controls Pain and Subchondral Bone Degeneration Induced by Mono-Iodoacetate: Implications GPCR Functional Bias and Tolerance Development. Biomed. Pharmacother. 136, 111283. doi:10.1016/j.biopha.2021.111283
Morales, P., and Jagerovic, N. (2016). Advances towards the Discovery of GPR55 Ligands. Curr. Med. Chem. 23, 2087–2100. Available at: http://benthamscience.com/journals/current-medicinal-chemistry/article/141455/(Accessed May 9, 2016). doi:10.2174/0929867323666160425113836
Morales, P., Hurst, D. P., and Reggio, P. H. (2017a). Methods for the Development of In Silico GPCR Models. Methods Enzymol., 405–448. doi:10.1016/bs.mie.2017.05.005
Morales, P., Hurst, D. P., and Reggio, P. H. (2017b). “Molecular Targets of the Phytocannabinoids: A Complex Picture Phytocannabinoids,” in Unraveling the Complex Chemistry and Pharmacology of Cannabis Sativa. Editors A. D. Kinghorn, and S. Gibbons (Springer International Publishing), 103–131. doi:10.1007/978-3-319-45541-9
Morales, P., Lago-Fernandez, A., Hurst, D. P., Sotudeh, N., Brailoiu, E., Reggio, P. H., et al. (2020). Therapeutic Exploitation of GPR18: Beyond the Cannabinoids? J. Med. Chem. 63, 14216–14227. doi:10.1021/acs.jmedchem.0c00926
Morales, P., and Reggio, P. H. (2017). An Update on Non-CB1, Non-CB2 Cannabinoid Related G-Protein-Coupled Receptors. Cannabis Cannabinoid Res. 2, 265–273. doi:10.1089/can.2017.0036
Morgese, M. G., Cassano, T., Cuomo, V., and Giuffrida, A. (2007). Anti-dyskinetic Effects of Cannabinoids in a Rat Model of Parkinson's Disease: Role of CB1 and TRPV1 Receptors. Exp. Neurol. 208, 110–119. doi:10.1016/j.expneurol.2007.07.021
Muller, C., Lynch, D. L., Hurst, D. P., and Reggio, P. H. (2020). A Closer Look at Anandamide Interaction with TRPV1. Front. Mol. Biosci. 7, 1–8. doi:10.3389/fmolb.2020.00144
Muller, C., Lynch, D. L., Hurst, D. P., and Reggio, P. H. (2021). TRPV1 Activation by Anandamide via a Unique Lipid Pathway. J. Chem. Inf. Model. 61, 5742–5746. doi:10.1021/acs.jcim.1c00893
Muller, C., Morales, P., and Reggio, P. H. (2019). Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 11, 1–15. doi:10.3389/fnmol.2018.00487
Nam, J. H., Park, E. S., Won, S.-Y., Lee, Y. A., Kim, K. I., Jeong, J. Y., et al. (2015). TRPV1 on Astrocytes Rescues Nigral Dopamine Neurons in Parkinson's Disease via CNTF. Brain 138, 3610–3622. doi:10.1093/brain/awv297
Nanda, K. K., Henze, D. A., Della Penna, K., Desai, R., Leitl, M., Lemaire, W., et al. (2014). Benzimidazole CB2 Agonists: Design, Synthesis and SAR. Bioorg. Med. Chem. Lett. 24, 1218–1221. doi:10.1016/j.bmcl.2013.12.068
Navarro, G., Gonzalez, A., Sánchez-Morales, A., Casajuana-Martin, N., Gómez-Ventura, M., Cordomí, A., et al. (2021). Design of Negative and Positive Allosteric Modulators of the Cannabinoid CB2 Receptor Derived from the Natural Product Cannabidiol. J. Med. Chem. 64, 9354–9364. doi:10.1021/acs.jmedchem.1c00561
Navarro, G., Morales, P., Rodríguez-Cueto, C., Fernández-Ruiz, J., Jagerovic, N., and Franco, R. (2016). Targeting Cannabinoid CB2 Receptors in the Central Nervous System. Medicinal Chemistry Approaches with Focus on Neurodegenerative Disorders. Front. Neurosci. 10, 1–11. doi:10.3389/fnins.2016.00406
Osman, N. A., Mahmoud, A. H., Allarà, M., Niess, R., Abouzid, K. A., Marzo, V. D., et al. (2010). Synthesis, Binding Studies and Molecular Modeling of Novel Cannabinoid Receptor Ligands. Bioorg. Med. Chem. 18, 8463–8477. doi:10.1016/j.bmc.2010.10.050
Palomares, B., Ruiz-Pino, F., Navarrete, C., Velasco, I., Sánchez-Garrido, M. A., Jimenez-Jimenez, C., et al. (2018). VCE-004.8, A Multitarget Cannabinoquinone, Attenuates Adipogenesis and Prevents Diet-Induced Obesity. Sci. Rep. 8, 1–15. doi:10.1038/s41598-018-34259-0
Peppin, J. F., and Pappagallo, M. (2014). Capsaicinoids in the Treatment of Neuropathic Pain: A Review. Ther. Adv. Neurol. Disord. 7, 22–32. doi:10.1177/1756285613501576
PubChem Bioassays (2021). Available at https://www.ebi.ac.uk/chembl/assay_report_card/CHEMBL1963979/(Accessed December 16, 2021).
Pumroy, R. A., Samanta, A., Liu, Y., Hughes, T., Zhao, S., Yudin, Y., et al. (2019). Molecular Mechanism of TRPV2 Channel Modulation by Cannabidiol. doi:10.7554/eLife.48792.001
Punzo, F., Manzo, I., Tortora, C., Pota, E., Angelo, V. D., Bellini, G., et al. (2018). Effects of CB2 and TRPV1 Receptors' Stimulation in Pediatric Acute T-Lymphoblastic Leukemia. Oncotarget 9, 21244–21258. doi:10.18632/oncotarget.25052
Ramírez‐López, A., Pastor, A., Torre, R., La Porta, C., Ozaita, A., Cabañero, D., et al. (2021). Role of the Endocannabinoid System in a Mouse Model of Fragile X Undergoing Neuropathic Pain. Eur. J. Pain 25, 1316–1328. doi:10.1002/ejp.1753
Rossi, F., Bellini, G., Tortora, C., Bernardo, M. E., Luongo, L., Conforti, A., et al. (2015). CB2 and TRPV1 Receptors Oppositely Modulate In Vitro Human Osteoblast Activity. Pharmacol. Res. 99, 194–201. doi:10.1016/j.phrs.2015.06.010
Saari, R., Törmä, J.-C., and Nevalainen, T. (2011). Microwave-assisted Synthesis of Quinoline, Isoquinoline, Quinoxaline and Quinazoline Derivatives as CB2 Receptor Agonists. Bioorg. Med. Chem. 19, 939–950. doi:10.1016/j.bmc.2010.11.059
Sawamura, S., Shirakawa, H., Nakagawa, T., Mori, Y., and Kaneko, S. (2017). TRP Channels in the BrainT. L. R. Emir (Boca Raton, FL: CRC Press/Taylor & Francis). Neurobiol. TRP Channels, 295–322. doi:10.4324/9781315152837-16
Shao, Z., Yin, J., Chapman, K., Grzemska, M., Clark, L., Wang, J., et al. (2016). High-resolution crystal Structure of the Human CB1 Cannabinoid Receptor. Nature 540, 602–606. doi:10.1038/nature20613
Shuba, Y. M. (2021). Beyond Neuronal Heat Sensing: Diversity of TRPV1 Heat-Capsaicin Receptor-Channel Functions. Front. Cel. Neurosci. 14, Article 612480. doi:10.3389/fncel.2020.612480
Singh, A. K., McGoldrick, L. L., and Sobolevsky, A. I. (2018). Structure and Gating Mechanism of the Transient Receptor Potential Channel TRPV3. Nat. Struct. Mol. Biol. 25, 805–813. doi:10.1038/s41594-018-0108-7
Soethoudt, M., Grether, U., Fingerle, J., Grim, T. W., Fezza, F., de Petrocellis, L., et al. (2017). Cannabinoid CB2 Receptor Ligand Profiling Reveals Biased Signalling and Off-Target Activity. Nat. Commun. 8, 13958. doi:10.1038/ncomms13958
Sotudeh, N., Morales, P., Hurst, D. P., Lynch, D. L., and Reggio, P. H. (2019). Towards A Molecular Understanding of the Cannabinoid Related Orphan Receptor GPR18: A Focus on its Constitutive Activity. Ijms 20, 2300. doi:10.3390/ijms20092300
Suo, Y., Wang, Z., Zubcevic, L., Hsu, A. L., He, Q., Borgnia, M. J., et al. (2020). Structural Insights into Electrophile Irritant Sensing by the Human TRPA1 Channel. Neuron 105, 882–894.e5. doi:10.1016/j.neuron.2019.11.023
Szallasi, A., and Di Marzo, V. (2000). New Perspectives on Enigmatic Vanilloid Receptors. Trends Neurosciences 23, 491–497. doi:10.1016/S0166-2236(00)01630-1
Tham, M., Yilmaz, O., Alaverdashvili, M., Kelly, M. E. M., Denovan-Wright, E. M., and Laprairie, R. B. (2018). Allosteric and Orthosteric Pharmacology of Cannabidiol and Cannabidiol-Dimethylheptyl at the Type 1 and Type 2 Cannabinoid Receptors. Br. J. Pharmacol. 176, 1455–1469. doi:10.1111/bph.14440
Uddin, M. S., Mamun, A. A., Sumsuzzman, D. M., Ashraf, G. M., Perveen, A., Bungau, S. G., et al. (2020). Emerging Promise of Cannabinoids for the Management of Pain and Associated Neuropathological Alterations in Alzheimer's Disease. Front. Pharmacol. 11, 1–13. doi:10.3389/fphar.2020.01097
Viet Thang Vu, M., Thacker, L. N., O’Malley, D. R., and Ogishi, T. (2019). Multi-use Cartridge for Ingestion of Cannabis-Based Products. 1. PATENT US20190275268A1.
Wang, S., Chen, J., Collins, G., and Levant, B. (2010). Selective Ligands for the Dopamine 3 (D3) Receptor and Methods of Using the Same.
Wi, R., Chung, Y. C., Jin, B. K., and Duan, L. (2020). Functional Crosstalk between CB and TRPV1 Receptors Protects Nigrostriatal Dopaminergic Neurons in the MPTP Model of Parkinson's Disease. J. Immunol. Res. 2020, 1–11. doi:10.1155/2020/5093493
Wilkerson, J. L., Alberti, L. B., Thakur, G. A., Makriyannis, A., and Milligan, D. (2022). Peripherally administered cannabinoid receptor 2 (CB2R) agonists lose anti-allodynic effects in TRPV1 knockout mice, while intrathecal administration leads to anti-allodynia and reduced GFAP, CCL2 and TRPV1 expression in the dorsal spinal cord and DRG. Brain Res. 1774, 147721. doi:10.1016/j.brainres.2021.147721
Yin, Y., Le, S. C., Hsu, A. L., Borgnia, M. J., Yang, H., and Lee, S.-Y. (2019). Structural Basis of Cooling Agent and Lipid Sensing by the Cold-Activated TRPM8 Channel. Science 363. doi:10.1126/science.aav9334
Ying-Ji, W., and Bo-Yuan, C. (2013). Preparation Method for 2-substituted 3-arylmethylbenzofuran. Org. Lett. 15(18), 4742–4745. doi:10.1021/ol4021095
Yong-Gang, W., Guan-Peng, Q., Bo-Lin, L., Giu-Zhuan, S., Guo-Zhi, Z., Yong-Hua, L., et al. (2019). Phosphonium Derivative, Preparation Method and Use Thereof.
Zagzoog, A., Mohamed, K. A., Kim, H. J. J., Kim, E. D., Frank, C. S., Black, T., et al. (2020). In Vitro and In Vivo Pharmacological Activity of Minor Cannabinoids Isolated from Cannabis Sativa. Sci. Rep. 10, 1–13. doi:10.1038/s41598-020-77175-y
Zhang, M., Chi, M., Zou, H., Tian, S., Zhang, Z., and Wang, G. (2017). Effects of Coadministration of Low Dose Cannabinoid Type 2 Receptor Agonist and Morphine on Vanilloid Receptor 1 Expression in a Rat Model of Cancer Pain. Mol. Med. Rep. 16, 7025–7031. doi:10.3892/mmr.2017.7479
Zhao, J., Lin King, J. V., Cheng, Y., Julius, D., and Julius, D. (2020). Irritant-evoked Activation and Calcium Modulation of the TRPA1 Receptor. Nature 585, 141–145. doi:10.1038/s41586-020-2480-9
Keywords: cannabinoids, ionotropic receptors, CB2, TRPV1, dual ligands, multitargeting
Citation: Morales P, Muller C, Jagerovic N and Reggio PH (2022) Targeting CB2 and TRPV1: Computational Approaches for the Identification of Dual Modulators. Front. Mol. Biosci. 9:841190. doi: 10.3389/fmolb.2022.841190
Received: 22 December 2021; Accepted: 03 February 2022;
Published: 25 February 2022.
Edited by:
Grzegorz Wegrzyn, University of Gdansk, PolandReviewed by:
Qing-Rong Liu, National Institute on Aging (NIH), United StatesKuo-Hui Su, University of Toledo, United States
Copyright © 2022 Morales, Muller, Jagerovic and Reggio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paula Morales, cGF1bGEubW9yYWxlc0BpcW0uY3NpYy5lcw==; Chanté Muller, Y2FtdWxsZXJAdW5jZy5lZHU=
†These authors have contributed equally to this work