 Eugene Lin
Eugene Lin Po-Hsiu Kuo
Po-Hsiu Kuo Yu-Li Liu
Yu-Li Liu Albert C. Yang
Albert C. Yang Shih-Jen Tsai
Shih-Jen Tsai- 1Department of Biostatistics, University of Washington, Seattle, WA, United States
- 2Department of Electrical & Computer Engineering, University of Washington, Seattle, WA, United States
- 3Graduate Institute of Biomedical Sciences, China Medical University, Taichung, Taiwan
- 4Department of Public Health, Institute of Epidemiology and Preventive Medicine, National Taiwan University, Taipei, Taiwan
- 5Center for Neuropsychiatric Research, National Health Research Institutes, Hsinchu, Taiwan
- 6Division of Interdisciplinary Medicine and Biotechnology, Beth Israel Deaconess Medical Center/Harvard Medical School, Boston, MA, United States
- 7Institute of Brain Science, National Yang-Ming University, Taipei, Taiwan
- 8Department of Psychiatry, Taipei Veterans General Hospital, Taipei, Taiwan
- 9Division of Psychiatry, National Yang-Ming University, Taipei, Taiwan
Evidence suggests that the neuro-inflammation mechanisms associated with interleukin-12 (IL-12) may be linked to Alzheimer's diseases and cognitive aging. In this study, we speculate that single nucleotide polymorphisms (SNPs) in IL-12-associated genes, such as IL12A, IL12B, IL12RB1, and IL12RB2 genes, could be associated with cognitive aging individually and/or via complicated interactions in the elder Taiwanese population. There were totally 3,730 Taiwanese individuals with age ≥60 years from the Taiwan Biobank. Mini-Mental State Examination (MMSE) was analyzed for all participants. We employed MMSE scores to assess cognitive functions. Our analysis revealed that the IL12A gene (including rs116910715, rs78902931, and rs78569420), the IL12B gene (including rs730691), and the IL12RB2 gene (including rs3790558, rs4655538, rs75699623, and rs1874396) were associated with cognitive aging. Among these SNPs, the association with the IL12RB2 rs3790558 SNP remained significant after performing Bonferroni correction (P = 6.87 × 10−4). Additionally, we found that interactions between the IL12A and IL12RB2 genes influenced cognitive aging (P = 0.022). Finally, we pinpointed the effects of interactions between IL12A, IL12B, and IL12RB2 with physical activity (P < 0.001, = 0.002, and < 0.001, respectively). Our study suggests that the IL-12-associated genes may contribute to susceptibility to cognitive aging independently as well as through gene-gene and gene-physical activity interactions.
Introduction
Interleukin-12 (IL-12) is a pro-inflammatory cytokine that builds a key link between adaptive immunity and innate resistance (1). IL-12 is a heterodimer composed of the IL-12α (also known as IL-12p35) and IL-12β (also known as IL-12p40) (1). Furthermore, IL-12 binds to the IL-12 receptor, which is a heterodimeric receptor formed by the IL-12R-β1 and IL-12R-β2 subunits (1). There is growing evidence that IL-12 is regulated in neuro-inflammatory processes associated with neurodegenerative disorders such as Alzheimer's disease (AD) and mild cognitive impairment (2, 3). It has been observed that IL-12 contributes to cognitive decline such as reduced performance in processing speed for elderly individuals aged 70–90 years in an Australia population (4). In the same cohort, a subtype of mild cognitive impairment (that is, non-amnestic multiple domain) was associated with higher levels of IL-12 and IL-12β (5). Moreover, a meta-analysis study revealed that there were significantly higher levels of IL-12 in AD patients when compared with healthy controls in peripheral blood (6). In addition, a recent study simultaneously assessed 242 blood proteins in 80 older adults with remitted major depression and found IL-12β to serve as one of the three proteins to predict cognitive impairment by using a machine learning prediction model (7). It has also been suggested that IL-12β is involved in affecting Mini-Mental State Examination (MMSE) test scores and gray-matter volumes of lateral prefrontal cortex and hippocampus in older adults (8). Furthermore, it has been reported that elderly persons with inadequate physical activity showed higher levels of IL-12β, smaller gray-matter volumes, and more cognitive decline than active elderly persons, suggesting probable gene-physical activity interactions (8).
Among the genes related to IL-12 are the interleukin 12A (IL12A), interleukin 12B (IL12B), interleukin 12 receptor subunit beta 1 (IL12RB1), and interleukin 12 receptor subunit beta 2 (IL12RB2) genes. The IL12A gene is located on chromosome 3q25.33 and encodes the IL-12α subunit (1). The IL12B gene is located on chromosome 5q33.3 and encodes the IL-12β subunit (1). It has been demonstrated that single nucleotide polymorphisms (SNPs) in IL12A (including rs568408) and IL12B (including rs3212227) genes were likely to influence late-onset AD in a Chinese population (9). In addition, Enright et al. (10) reported that the Il12b knockout male mice exhibited a significant increase in the average time to reach the platform in the Morris water navigation task (also known as the Morris water maze; a test of spatial learning for rodents), when compared to the wild-type. In the APP/PS1 mouse model of AD, Vom Berg et al. (11) also found an increased production of the common IL-12 and IL-12β subunit in microglia, the primary immune cells of the central nervous system. Furthermore, Vom Berg et al. (11) demonstrated that the genetic ablation of the Il12b gene or peripheral administration of a neutralizing IL-12β-specific antibody may contribute to a decreased cerebral amyloid load in APP/PS1 mice.
The IL12RB1 gene is located on chromosome 19p13.11 and encodes the IL-12R-β1 subunit (1). Furthermore, the IL12RB2 gene is located on chromosome 1p31.3 and encodes the IL-12R-β2 subunit (1). The IL12RB1 and IL12RB2 genes are thought to contribute to the host defense and inflammatory response (1). de Beaucoudrey et al. reported that loss-of-function mutations in the IL12RB1 gene may debilitate the development of human IL-17-producing T cells in an in vivo study, where IL-17 has been implicated in the pathogenesis of AD-related neuroinflammation (12–14). Additionally, several SNPs (such as rs12119179, rs1495965, and rs924080) near the IL12RB2 gene were found to be associated with Behcet's disease in genome-wide association studies, were the disease can lead to inflammation in the brain and central nervous system (15, 16).
In reference to the aforementioned considerations, it was hypothesized that IL-12 associated genes may play a significant role in the pathogenesis of age-dependent cognitive decline and the development of cognitive aging. Therefore, we presumed that IL-12 relevant genes, namely the IL12A, IL12B, IL12RB1, and IL12RB2 genes, might be associated with cognitive aging. To the best of our knowledge, the effects of these IL-12 relevant genes on cognitive aging are limited with respect to human datasets. Thus, we investigated the interplays between cognitive aging and SNPs in the IL12A, IL12B, IL12RB1, and IL12RB2 genes in the present association study. We also gauged the probable gene-gene and gene-physical activity interactions on cognitive aging.
Materials and Methods
Study Population
This study incorporated Taiwanese participants from the Taiwan Biobank, which collected specimens and relevant information from individuals in recruitment centers across Taiwan (17–22). Our study cohort was comprised of 3,730 subjects. There were the following two inclusion criteria: (1) individuals whose age were 60 years or over; and (2) individuals who were self-reported as being of Taiwanese Han Chinese ancestry (22). We excluded individuals with a history of cancer (22). Ethical approval for the study was granted by the Institutional Review Board of the Taiwan Biobank before performing the study (approval number: 201506095RINC). The approved informed consent form was signed by each subject. All experiments were achieved by means of proper regulations and guidelines.
We defined education according to whether or not high school was attended (20, 21). The definition of physical activity was the measurement of exercise activities at least three times in a week and at least 30 min each time (20, 21).
Cognitive Assessment
We performed global cognitive assessment by using the 30-point MMSE, which encompasses questions according to the five areas of recall, registration, language, attention and calculation, and orientation (18). We evaluated MMSE both as a continuous phenotype and as a binary phenotype according to the following previously defined MMSE thresholds (23): MMSE score ≥24 (normal) and MMSE score < 24 (cognitive impairment). The cognitive assessment was conducted in the local languages (such as Taiwanese and/or Mandarin). The cognitive cut-off score of 24 was derived from previous studies (23) and was based on a Taiwanese version of MMSE.
Genotyping
DNA was isolated from blood samples by employing QIAamp DNA blood kits following the manufacturer's instructions (Qiagen, Valencia, CA, USA). The quality of the isolated genomic DNA was carried out by utilizing agarose gel electrophoresis, and the quantity was completed by spectrophotometry (24). SNP genotyping was evaluated by employing the custom Taiwan BioBank chips, which were accomplished by using the Axiom Genome-Wide Array Plate System (Affymetrix, Santa Clara, CA, USA). The custom Taiwan BioBank chips were created to collect genetic profiles in Taiwanese subjects by utilizing SNPs on the Axiom Genome-Wide CHB 1 Array (Affymetrix, Santa Clara, CA, USA) with minor allele frequencies (MAFs) ≥5% and the Human Exome BeadChip (Illumina, Inc., San Diego, CA, USA) with MAFs > 10% (22). We searched for IL-12-associated variants by referring to the complete list of IL-12-associated genes/SNPs available in the custom Taiwan BioBank chips. The SNP panel consisted of 75 SNPs from the following four genes, namely the IL12A, IL12B, IL12RB1, and IL12RB2 genes (Supplementary Table 1). In addition, we performed quality control procedures for excluding SNPs from subsequent analysis as follows (25). Nine SNPs were excluded because of being unable to achieve Hardy-Weinberg equilibrium (with a P-value < 0.05) or because of a genotyping call rate <95%. Supplementary Table 1 shows the genotyping results, including genotyping call rates, P-values for Hardy-Weinberg equilibrium, and MAFs. Additionally, we filtered SNPs and then selected 35 tag SNPs by using PLINK (26) with a linkage disequilibrium value of r2 = 0.8 as a threshold (Supplementary Table 2).
Statistical Analysis
In this study, linear regression analysis was conducted to assess the relationship between MMSE scores and our variables of interest such as age, gender, and education. In addition, we determined the association of the investigated SNP with MMSE scores by a general linear model using age, gender, and education as covariates (27). The genotype frequencies were weighed for Hardy-Weinberg equilibrium to detect genotyping errors (28) by utilizing a χ2 goodness-of-fit test with one degree of freedom (that is, the number of genotypes minus the number of alleles). Adjustments for multiple testing were performed by using the Bonferroni correction. The criterion for significance was defined as P < 0.05 for all tests. Data were shown by the mean ± standard deviation.
In order to explore gene-gene and gene-physical activity interactions, we used the generalized multifactor dimensionality reduction (GMDR) method (29). We analyzed two-way interactions by utilizing 10-fold cross-validation. The GMDR method generated several output parameters, such as empirical P-values and the testing accuracy, to estimate each chosen interaction. Furthermore, covariates such as age, gender, and education were provided for gene-gene and gene-physical activity interaction analysis in our interaction models. We completed the empirical P-value of the testing accuracy for each chosen interaction by using permutation testing (based on 1,000 shuffles).
In this study, there were missing data in the genotypic data (as shown in Supplementary Table 1) and no missing data in the phenotypic data. In GMDR, a missing genotype is imputed proportional to the frequencies of the SNPs observed at this locus, and insignificant missing data will not affect the analysis (29). Because the selected SNPs possess <0.5% missing genotypic data by excluding SNPs with a genotyping call rate <95% (Supplementary Table 1), we have chosen to use GMDR after assessing the influence of missing data. In order to correct for multiple testing, we employed the Bonferroni correction.
Results
Study Cohort
Table 1 illustrates the clinical and demographic characteristics of our study cohort, which consisted of 3,730 individuals. The median MMSE score was 28 and the interquartile range was 26–29. In this study, we found that correlations between MMSE score with age (P = 8.04 × 10−10), gender (P = 1.83 × 10−7), and education (P = 2.2 × 10−16) were significant.

Table 1. Demographic and clinical characteristics of study subjects.
Association of Cognitive Aging in IL12A, IL12B, and IL12RB2
First, we explored the associations between cognitive aging and four IL-12 related genes, namely the IL12A, IL12B, IL12RB1, and IL12RB2 genes. Among the 35 tag SNPs investigated in the present study (Supplementary Table 2), there were 8 tag SNPs within the IL12A, IL12B, and IL12RB2 genes revealing evidence of associations (P < 0.05) with MMSE scores (Table 2). These 8 tag SNPs were the rs116910715 (recessive model: P = 0.0031), rs78902931 (recessive model: P = 0.0388), and rs78569420 (recessive model: P = 0.0093) SNPs in the IL12A gene; the rs730691 SNP (dominant model: P = 0.0472) in the IL12B gene; and the rs3790558 (dominant model: P = 6.87 × 10−4), rs4655538 (dominant model: P = 0.0454), rs75699623 (dominant model: P = 0.0308), and rs1874396 (dominant model: P = 0.0425) SNPs in the IL12RB2 gene. In this study, we only found the genotyping data for the rs7412 SNP, but not for the rs429358 SNP in the APOE gene. Based on the rs7412 SNP, the frequency of the APOE-ε2 allele in patients showing normal cognition vs. cognitive impairment was 14.2 vs. 18.9%. However, we were unable to assess APOE-ε4 carrier status.
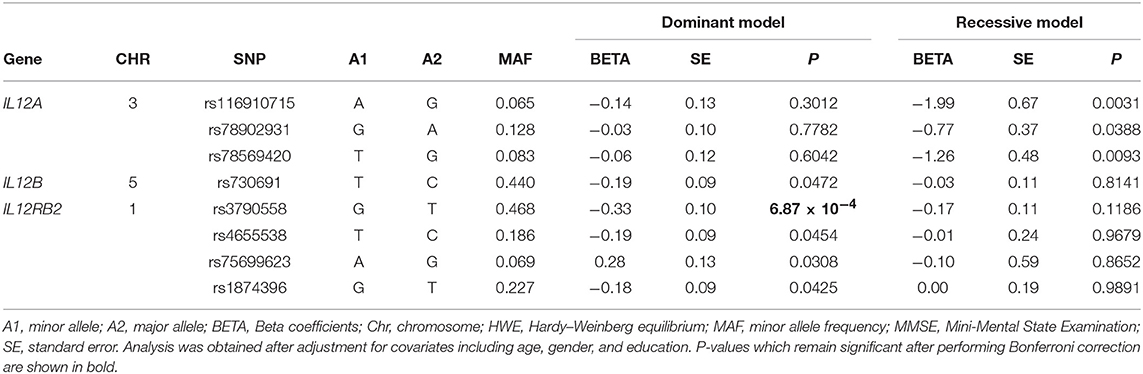
Table 2. Linear regression models of associations between the MMSE scores and 8 tag SNPs within the IL12A, IL12B, and IL12RB2 genes, which have an evidence of association (P < 0.05).
Association Between Cognitive Aging and IL12RB2 rs3790558
Moreover, as illustrated in Table 2, the significance persisted for the association with MMSE scores after employing Bonferroni correction [P < 0.05/(35 × 2) = 7.14 × 10−4] for the rs3790558 SNP (dominant model: P = 6.87 × 10−4) in the IL12RB2 gene.
Gene-Gene Interaction Analysis
Next, we utilized categorized MMSE scores as an outcome (normal: MMSE score ≥24; cognitive impairment: MMSE score < 24) for gene-gene interaction analysis. The GMDR method was employed to estimate the effects of consolidation among the 8 tag SNPs in cognitive aging, incorporating age, gender, and education as covariates. Table 3 describes the results generated from the GMDR method for two-way gene-gene interaction analysis using covariate adjustment. As illustrated in Table 3, there was a significant two-way model concerning IL12RB2 rs4655538 and IL12A rs78902931 (P = 0.022), suggesting a probable gene-gene interaction between IL12RB2 rs4655538 and IL12A rs78902931 in regulating cognitive aging. Likewise, there was a significant two-way gene-gene interaction model concerning IL12RB2 rs4655538 and IL12A rs78569420 (P = 0.036) in regulating cognitive aging. In addition, there was a significant two-way SNP-SNP interaction model concerning IL12RB2 rs3790558 and IL12RB2 rs4655538 (P = 0.017) in influencing cognitive aging. However, the effect of these gene-gene and SNP-SNP interaction models did not remain significant after Bonferroni correction.
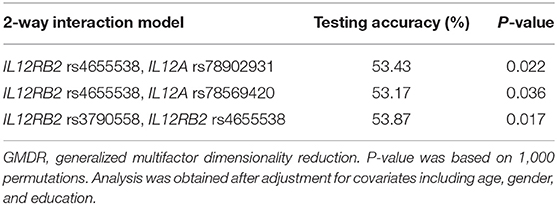
Table 3. Gene-gene interaction models identified by the GMDR method with adjustment for age, gender, and education.
Physical Activity and Gene Interaction Analysis
Table 4 illustrates the GMDR method of physical activity and gene interaction analysis in cognitive aging by utilizing age, gender, and education as covariates. There were significant two-way models concerning IL12A (including rs116910715, rs78902931, and rs78569420) and physical activity (P = 0.002, < 0.001, and 0.001, respectively), indicating potential physical activity and gene interactions between IL12A and physical activity in regulating cognitive aging. Likewise, there was a significant two-way model concerning IL12B rs730691 and physical activity (P = 0.002). Finally, there were significant two-way models concerning IL12RB2 (including rs3790558, rs4655538, s75699623, and rs1874396) and physical activity (P = 0.003, < 0.001, < 0.001, and 0.025, respectively). The effect of these physical activity and gene interaction models remained significant after Bonferroni correction (P < 0.05/8 = 0.006) except the interaction model between physical activity and IL12RB2 rs1874396.
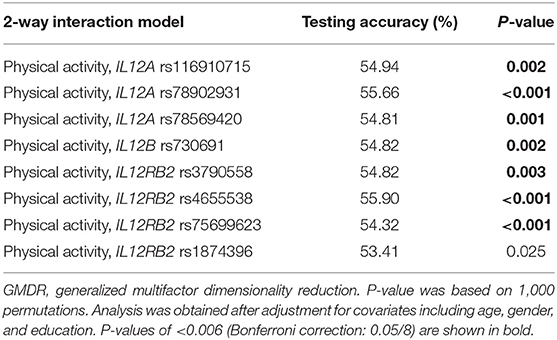
Table 4. Physical activity and gene interaction models identified by the GMDR method with adjustment for age, gender, and education.
Discussion
The present study is the first to date to identify whether the major impacts of 35 tag SNPs within four IL-12-associated genes, namely the IL12A, IL12B, IL12RB1, and IL12RB2 genes, are significantly linked to the risk of cognitive aging individually and via gene-gene and gene-physical activity interactions in elder Taiwanese subjects. Here, we reveal for the first time that the IL12RB2 gene might play a vital role in modulating cognitive aging in elder Taiwanese individuals. Intriguingly, the significant persisted for the association of the key rs3790558 SNP in the IL12RB2 gene with MMSE scores after correcting for multiple testing (P < 7.14 × 10−4). The rs3790558 SNP is located in the intron of IL12RB2 gene. To investigate the possible roles of the rs3790558 SNP as expression quantitative trait locus, we employed HaploReg (http://compbio.mit.edu/HaploReg) to predict a possible functional role of this SNP. We found that the rs3790558 SNP is associated with the regulation of expressions in IL12RB2 gene within transformed fibroblasts/cells and in blood tissues (30, 31). Additionally, our data revealed that gene-gene and gene-physical activity interactions between the IL-12-associated genes may contribute to the etiology of cognitive aging.
To the best of our knowledge, the present study is the first to raise the possibility that the key rs3790558 SNP in the IL12RB2 gene might be associated with cognitive aging. Remarkably, the significant association between this key SNP and MMSE scores persisted even after applying Bonferroni correction. The functional relevance of IL12RB2 rs3790558 on cognitive aging remains to be elucidated. To our knowledge, no other studies have been conducted to pinpoint IL12RB2 rs3790558 with cognitive aging or age-related cognitive decline. Based on the aforementioned implications, we hypothesized that the IL-12 relevant genes such as IL12RB2 might contribute to cognitive aging because IL-12 is implicated as a risk biomarker for AD and cognitive aging (1, 4, 7–9). Our findings further support previous animal studies, which demonstrated that inhibition of IL-12 signaling may reduce cognitive decline (11, 32). In addition, genome-wide association studies by Remmers et al. (16) and by Mizuk et al. (15) have identified an association of the IL12RB2 gene with Behcet's disease at genome-wide significance. It has been shown that patients with Behcet's disease often suffer from irreversible loss of cognitive function in conjunction with various neurological disturbances in the central nervous system (33). In an Il12rb2 knockout mice study, Airoldi et al. (34) also reported that lack of Il12rb2 signaling may result in increased susceptibility to autoimmunity and immunopathology. Moreover, it has been suggested that the immune system and autoimmunity may play a role in the etiology of age-associated cognitive decline and AD (35–37). Furthermore, Li et al. (38) observed that the increased risks of dementia and AD in patients with autoimmune disorders. Interestingly, the IL12RB2 rs3790558 SNP is a strong candidate for autoimmune disorders as this SNP has been previously implicated in autoimmune disorders such as systemic sclerosis. For example, Bossini-Castillo et al. (39) identified IL12RB2 rs3790558 to be associated with systemic sclerosis, a disorder that is characterized by autoimmune dysfunction. It should be mentioned that the G allele frequency of IL12RB2 rs3790558 varies considerably among different ethnic individuals, ranging from 46.8% in the present Taiwanese individuals, 43.7% in European individuals, 47.3% in East Asian individuals, 86.5% in African American individuals, to 48% in South Asian individuals as illustrated in public data from the 1000 Genomes Project (Supplementary Table 4).
Remarkably, we tracked down the interplay between the IL-12-associated genes (including IL12A, IL12B, and IL12RB2) and physical activity. This relationship might functionally manifest itself via epigenetic changes. Our finding is in agreement with other human and animal studies, indicating that physical activity may modulate inflammatory reactions through potential complex gene-physical activity interactions (8, 40, 41). In a previous population-based study of older adults, Papenberg et al. (8) reported that inactive older individuals may exhibit elevated levels of IL-12β, smaller gray-matter volumes, and poorer cognitive performance than older individuals with adequate physical activity.
On another note, our results also indicated the epistatic effects between the IL12RB2 and IL12A genes in modulating cognitive aging by employing the GMDR method. To our knowledge, there are no previous findings available as no other studies have been investigated to assess gene-gene interactions among these genes. The biological effects of synergy between the IL-12-associated genes on cognitive aging remain to be elucidated.
A previous genetic association study found that IL12A rs568408 and IL12B rs3212227 SNP were significantly associated with late-onset AD risk (9). However, the present study showed no association of cognitive aging with these two SNPs (Supplementary Table 3). It is worth pointing out that various possible factors for the conflicting data include sample size, study design, covariate adjustment, phenotype definitions, and diverse ethnicities (21).
Among the strengths of our study is that we were able to utilize the Taiwan Biobank, the largest Taiwanese cohort, to conduct an extensive assessment of the IL-12-associated SNPs in cognitive aging (42, 43). In addition, we conducted the GMDR method to consider gene-gene interaction and gene-physical activity interaction in the model. However, because only the MMSE data were provided in the Taiwan Biobank, a major limitation is that a single measure of cognition (that is, MMSE) used as the cognitive assessment tool limits the rigor and depth of analytic inferences and associations with potential SNPs (44). Moreover, we did not control for other meaningful variables implicated in elderly cognitive decline (for example, insufficient characterization of the cohort including medical comorbidities and/or related medical burden) (44). Future studies are needed to develop a detailed assessment of the associations and interactions of probable SNPs with cognitive aging by leveraging specific cognitive domains (including executive, memory, language, and visuospatial function) in other worldwide populations (42, 44).
In conclusion, the present study completed a comprehensive investigation of the associations of cognitive aging with IL-12 relevant genes, namely the IL12A, IL12B, IL12RB1, and IL12RB2 genes in old adults in the Taiwanese population. Moreover, the present study tested the impacts of gene-gene and gene-physical activity interactions among these genes in relation to cognitive aging. Mainly, if the current results are reproduced in statistically well-powered independent studies, the present study implicates the effects of the IL-12 relevant genes on the risk of cognitive aging individually and via complicated gene-gene and gene-physical activity interactions. This study pinpoints that IL-12 mediated signaling should be the focus of future studies on pathogenesis of age-dependent cognitive decline and a potential target for pharmacologic modulation. Independent studies with larger sample sizes will possibly establish further insights into the role of the IL-12 related genes suggested in this study.
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
This study was approved by the Institutional Review Board of the Taiwan Biobank and complies with the Declaration of Helsinki. Informed written consent was obtained from all participants.
Author Contributions
EL and S-JT: study conception and design. P-HK, Y-LL, and AY: acquisition of data. EL and S-JT: analysis and interpretation of data. EL: draft manuscript. All authors: read and approved the final manuscript.
Funding
This work was supported by grant from Taiwan Ministry of Science and Technology (Grant MOST 107-2634-F-075-002; S-JT) and by the Taipei Veterans General Hospital (Grant V105D17-002-MY2-2; S-JT).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fneur.2019.01065/full#supplementary-material
References
1. Trinchieri G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol. (2003) 3:133–46. doi: 10.1038/nri1001
2. Griffin WS. Neuroinflammatory cytokine signaling and Alzheimer's disease. N Engl J Med. (2013) 368:770–1. doi: 10.1056/NEJMcibr1214546
3. Xu J, Barger SW, Drew PD. The PPAR-gamma agonist 15-deoxy-delta-prostaglandin J(2) attenuates microglial production of IL-12 family cytokines: potential relevance to Alzheimer's disease. PPAR Res. (2008) 2008:349185. doi: 10.1155/2008/349185
4. Trollor JN, Smith E, Agars E, Kuan SA, Baune BT, Campbell L, et al. The association between systemic inflammation and cognitive performance in the elderly: the Sydney Memory and Ageing Study. Age. (2012) 34:1295–308. doi: 10.1007/s11357-011-9301-x
5. Trollor JN, Smith E, Baune BT, Kochan NA, Campbell L, Samaras K, et al. Systemic inflammation is associated with MCI and its subtypes: the Sydney Memory and Aging Study. Dement Geriatr Cogn Disord. (2010) 30:569–78. doi: 10.1159/000322092
6. Swardfager W, Lanctôt K, Rothenburg L, Wong A, Cappell J, Herrmann N. A meta-analysis of cytokines in Alzheimer's disease. Biol Psychiatry. (2010) 68:930–41. doi: 10.1016/j.biopsych.2010.06.012
7. Diniz BS, Sibille E, Ding Y, Tseng G, Aizenstein HJ, Lotrich F, et al. Plasma biosignature and brain pathology related to persistent cognitive impairment in late-life depression. Mol Psychiatry. (2015) 20:594–601. doi: 10.1038/mp.2014.76
8. Papenberg G, Ferencz B, Mangialasche F, Mecocci P, Cecchetti R, Kalpouzos G, et al. Physical activity and inflammation: effects on gray-matter volume and cognitive decline in aging. Hum Brain Mapp. (2016) 37:3462–73. doi: 10.1002/hbm.23252
9. Zhu XC, Tan L, Jiang T, Tan MS, Zhang W, Yu JT. Association of IL-12A and IL-12B polymorphisms with Alzheimer's disease susceptibility in a Han Chinese population. J Neuroimmunol. (2014) 274:180–4. doi: 10.1016/j.jneuroim.2014.06.026
10. Enright BP, Davila DR, Tornesi BM, Blaich G, Hoberman AM, Gallenberg LA. Developmental and reproductive toxicology studies in IL-12p40 knockout mice. Birth Defects Res B Dev Reprod Toxicol. (2011) 92:102–10. doi: 10.1002/bdrb.20287
11. Vom Berg J, Prokop S, Miller KR, Obst J, Kalin RE, Lopategui-Cabezas I, et al. Inhibition of IL-12/IL-23 signaling reduces Alzheimer's disease-like pathology and cognitive decline. Nat Med. (2012) 18:1812–9. doi: 10.1038/nm.2965
12. de Beaucoudrey L, Puel A, Filipe-Santos O, Cobat A, Ghandil P, Chrabieh M, et al. Mutations in STAT3 and IL12RB1 impair the development of human IL-17–producing T cells. J Exp Med. (2008) 205:1543–50. doi: 10.1084/jem.20080321
13. Cristiano C, Volpicelli F, Lippiello P, Buono B, Raucci F, Piccolo M, et al. Neutralization of interleukin-17 rescues amyloid-β-induced neuroinflammation and memory impairment. Br J Pharmacol. (2019) 176:3544–57. doi: 10.1111/bph.14586
14. Saksida T, Koprivica I, Vujičić M, Stošić-Grujičić S, Perović M, Kanazir S, et al. Impaired IL-17 production in gut-residing immune cells of 5xFAD Mice with Alzheimer's disease pathology. J Alzheimer Dis. (2018) 61:619–30. doi: 10.3233/JAD-170538
15. Mizuki N, Meguro A, Ota M, Ohno S, Shiota T, Kawagoe T, et al. Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behçet's disease susceptibility loci. Nat Genet. (2010) 42:703. doi: 10.1038/ng.624
16. Remmers EF, Cosan F, Kirino Y, Ombrello MJ, Abaci N, Satorius C, et al. Genome-wide association study identifies variants in the MHC class I, IL10, and IL23R-IL12RB2 regions associated with Behcet's disease. Nat Genet. (2010) 42:698. doi: 10.1038/ng.625
17. Lin E, Yang AC, Tsai SJ. Association between metabolic syndrome and cognitive function in old adults in a Taiwanese population. Taiwan J Psychiatry. (2017) 31:232–40. doi: 10.29478/tjp
18. Lin E, Tsai SJ, Kuo PH, Liu YL, Yang AC, Kao CF, et al. The ADAMTS9 gene is associated with cognitive aging in the elderly in a Taiwanese population. PLoS ONE. (2017) 12:e0172440. doi: 10.1371/journal.pone.0172440
19. Lin E, Tsai SJ, Kuo PH, Liu YL, Yang AC, Kao CF, et al. The rs1277306 variant of the REST gene confers susceptibility to cognitive aging in an elderly Taiwanese population. Dement Geriatr Cogn Disord. (2017) 43:119–27. doi: 10.1159/000455833
20. Lin E, Tsai SJ, Kuo PH, Liu YL, Yang AC, Kao CF. Association and interaction effects of Alzheimer's disease-associated genes and lifestyle on cognitive aging in older adults in a Taiwanese population. Oncotarget. (2017) 8:24077–87. doi: 10.18632/oncotarget.15269
21. Lin E, Kuo PH, Liu YL, Yang AC, Kao CF, Tsai SJ. Effects of circadian clock genes and environmental factors on cognitive aging in old adults in a Taiwanese population. Oncotarget. (2017) 8:24088–98. doi: 10.18632/oncotarget.15493
22. Chen CH, Yang JH, Chiang CWK, Hsiung CN, Wu PE, Chang LC, et al. Population structure of Han Chinese in the modern Taiwanese population based on 10,000 participants in the Taiwan Biobank project. Hum Mol Genet. (2016) 25:5321–31. doi: 10.1093/hmg/ddw346
23. Sheehan B. Assessment scales in dementia. Ther Adv Neurol Disord. (2012) 5:349–58. doi: 10.1177/1756285612455733
24. Lin E, Kuo PH, Liu YL, Yang AC, Tsai SJ. Transforming growth factor-beta signaling pathway-associated genes SMAD2 and TGFBR2 are implicated in metabolic syndrome in a Taiwanese population. Sci Rep. (2017) 7:13589. doi: 10.1038/s41598-017-14025-4
25. Lin E, Kuo PH, Liu YL, Yang AC, Kao CF, Tsai SJ. Association and interaction of APOA5, BUD13, CETP, LIPA and health-related behavior with metabolic syndrome in a Taiwanese population. Sci Rep. (2016) 6:36830. doi: 10.1038/srep36830
26. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. (2007) 81:559–75. doi: 10.1086/519795
27. Lin E, Kuo PH, Liu YL, Yang AC, Kao CF, Tsai SJ. Effects of circadian clock genes and health-related behavior on metabolic syndrome in a Taiwanese population: evidence from association and interaction analysis. PLoS ONE. (2017) 12:e0173861. doi: 10.1371/journal.pone.0173861
28. Hosking L, Lumsden S, Lewis K, Yeo A, McCarthy L, Bansal A, et al. Detection of genotyping errors by Hardy–Weinberg equilibrium testing. Eur J Human Genet. (2004) 12:395. doi: 10.1038/sj.ejhg.5201164
29. Lou XY, Chen GB, Yan L, Ma JZ, Zhu J, Elston RC, et al. A generalized combinatorial approach for detecting gene-by-gene and gene-by-environment interactions with application to nicotine dependence. Am J Hum Genet. (2007) 80:1125–37. doi: 10.1086/518312
30. Consortium GT. Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: multitissue gene regulation in humans. Science. (2015) 348:648–60. doi: 10.1126/science.1262110
31. Westra HJ, Peters MJ, Esko T, Yaghootkar H, Schurmann C, Kettunen J, et al. Systematic identification of trans eQTLs as putative drivers of known disease associations. Nat Genet. (2013) 45:1238–43. doi: 10.1038/ng.2756
32. Tan MS, Yu JT, Jiang T, Zhu XC, Guan HS, Tan L. IL12/23 p40 inhibition ameliorates Alzheimer's disease-associated neuropathology and spatial memory in SAMP8 mice. J Alzheimer Dis. (2014) 38:633–46. doi: 10.3233/JAD-131148
33. Hirano T, Ohguro N, Hohki S, Hagihara K, Shima Y, Narazaki M, et al. A case of Behçet's disease treated with a humanized anti-interleukin-6 receptor antibody, tocilizumab. Modern Rheumatol. (2012) 22:298–302. doi: 10.3109/s10165-011-0497-5
34. Airoldi I, Di Carlo E, Cocco C, Sorrentino C, Fais F, Cilli M, et al. Lack of Il12rb2 signaling predisposes to spontaneous autoimmunity and malignancy. Blood. (2005) 106:3846–53. doi: 10.1182/blood-2005-05-2034
35. Kipnis J, Derecki NC, Yang C, Scrable H. Immunity and cognition: what do age-related dementia, HIV-dementia and ‘chemo-brain'have in common? Trends Immunol. (2008) 29:455–63. doi: 10.1016/j.it.2008.07.007
36. Mruthinti S, Buccafusco JJ, Hill WD, Waller JL, Jackson TW, Zamrini EY, et al. Autoimmunity in Alzheimer's disease: increased levels of circulating IgGs binding Aβ and RAGE peptides. Neurobiol Aging. (2004) 25:1023–32. doi: 10.1016/j.neurobiolaging.2003.11.001
37. Sardi F, Fassina L, Venturini L, Inguscio M, Guerriero F, Rolfo E, et al. Alzheimer's disease, autoimmunity and inflammation. The good, the bad and the ugly. Autoimmun Rev. (2011) 11:149–53. doi: 10.1016/j.autrev.2011.09.005
38. Li X, Sundquist J, Zöller B, Sundquist K. Dementia and Alzheimer's disease risks in patients with autoimmune disorders. Geriatr Gerontol Int. (2018) 18:1350–5. doi: 10.1111/ggi.13488
39. Bossini-Castillo L, Martin J-E, Broen J, Gorlova O, Simeón CP, Beretta L, et al. A GWAS follow-up study reveals the association of the IL12RB2 gene with systemic sclerosis in Caucasian populations. Human Mol Genet. (2011) 21:926–33. doi: 10.1093/hmg/ddr522
40. Hamer M, Sabia S, Batty GD, Shipley MJ, Tabak AG, Singh-Manoux A, et al. Physical activity and inflammatory markers over 10 years: follow-up in men and women from the Whitehall II cohort study. Circulation. (2012) 126:928–33. doi: 10.1161/CIRCULATIONAHA.112.103879
41. Sim YJ, Yu S, Yoon KJ, Loiacono CM, Kohut ML. Chronic exercise reduces illness severity, decreases viral load, and results in greater anti-inflammatory effects than acute exercise during influenza infection. J Infect Dis. (2009) 200:1434–42. doi: 10.1086/606014
42. Lin E, Kuo PH, Liu YL, Yang AC, Tsai SJ. Detection of susceptibility loci on APOA5 and COLEC12 associated with metabolic syndrome using a genome-wide association study in a Taiwanese population. Oncotarget. (2017) 8:93349–59. doi: 10.18632/oncotarget.20967
43. Lin E, Kuo P-H, Liu Y-L, Yang AC, Tsai S-J. Polymorphisms of the DNA repair gene EXO1 modulate cognitive aging in old adults in a Taiwanese population. DNA Repair. (2019) 78:1–6. doi: 10.1016/j.dnarep.2019.03.013
Keywords: Alzheimer's diseases, cognitive aging, cognitive impairment, interleukin-12, neurodegeneration
Citation: Lin E, Kuo P-H, Liu Y-L, Yang AC and Tsai S-J (2019) Association and Interaction Effects of Interleukin-12 Related Genes and Physical Activity on Cognitive Aging in Old Adults in the Taiwanese Population. Front. Neurol. 10:1065. doi: 10.3389/fneur.2019.01065
Received: 26 April 2019; Accepted: 20 September 2019;
Published: 09 October 2019.
Edited by:
Jennifer S. Yokoyama, University of San Francisco, United StatesReviewed by:
Rufus Olusola Akinyemi, University of Ibadan, NigeriaChung-Feng Kao, National Chung Hsing University, Taiwan
Copyright © 2019 Lin, Kuo, Liu, Yang and Tsai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eugene Lin, bGluZXNAdXcuZWR1; Shih-Jen Tsai, dHNhaTYxMDkxM0BnbWFpbC5jb20=