 Maria-Paz Viveros1*
Maria-Paz Viveros1* Adriana Mendrek2
Adriana Mendrek2 Tomáš Paus3,4
Tomáš Paus3,4 Ana Belén López-Rodríguez1
Ana Belén López-Rodríguez1 Eva Maria Marco1
Eva Maria Marco1 Rachel Yehuda5,6
Rachel Yehuda5,6 Hagit Cohen6
Hagit Cohen6 Amy Lehrner7
Amy Lehrner7 Edward J. Wagner8
Edward J. Wagner8
- 1 Physiology Department (Animal Physiology II), Biology Faculty, Health Research Institute of the Hospital Clínico San Carlos, Complutense University of Madrid, Madrid, Spain
- 2 Department of Psychology, Bishop’s University, Sherbrooke, QC, Canada
- 3 Rotman Research Institute, University of Toronto, Toronto, ON, Canada
- 4 Montreal Neurological Institute, McGill University, Montreal, QC, Canada
- 5 Mount Sinai School of Medicine, New York, NY, USA
- 6 Anxiety and Stress Research Unit, Ministry of Health Mental Health Center, Faculty of Health Sciences, Ben-Gurion University of the Negev, Beer-Sheva, Israel
- 7 Traumatic Stress Studies Division, James J. Peters Veterans Affairs Medical Center, Bronx, NY, USA
- 8 Department of Basic Medical Sciences, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA, USA
Women and men differ in a wide variety of behavioral traits and in their vulnerability to developing certain mental disorders. This review endeavors to explore how recent preclinical and clinical research findings have enhanced our understanding of the factors that underlie these disparities. We start with a brief overview of some of the important genetic, molecular, and hormonal determinants that contribute to the process of sexual differentiation. We then discuss the importance of animal models in studying the mechanisms responsible for sex differences in neuropsychiatric disorders (e.g., drug dependence) – with a special emphasis on experimental models based on the neurodevelopmental and “three hits” hypotheses. Next, we describe the most common brain phenotypes observed in vivo with magnetic resonance imaging. We discuss the challenges in interpreting these phenotypes vis-à-vis the underlying neurobiology and revisit the known sex differences in brain structure from birth, through adolescence, and into adulthood. This is followed by a presentation of pertinent clinical and epidemiological data that point to important sex differences in the prevalence, course, and expression of psychopathologies such as schizophrenia, and mood disorders including major depression and posttraumatic stress disorder. Recent evidence implies that mood disorders and psychosis share some common genetic predispositions and neurobiological bases. Therefore, modern research is emphasizing dimensional representation of mental disorders and conceptualization of schizophrenia and major depression as a continuum of cognitive deficits and neurobiological abnormalities. Herein, we examine available evidence on cerebral sexual dimorphism to verify if sex differences vary quantitatively and/or qualitatively along the psychoses-depression continuum. Finally, sex differences in the prevalence of posttraumatic disorder and drug abuse have been described, and we consider the genomic and molecular data supporting these differences.
Introduction
Psychiatric disorders and drug dependence are very important issues for healthcare systems that affect millions of people each year, and according to the National Institute of Mental Health (NIMH) only a small fraction of those patients receive an adequate treatment. These afflictions include anxiety and posttraumatic stress disorder (PTSD), attention-deficit hyperactivity disorder, autism, eating disorders, mood disorders (depression, bipolar disorder, BP), personality disorder, schizophrenia and drug abuse, and dependence1. As explained in the sections that follow many of them begin early in life (Kessler et al., 2007), and their high prevalence and social impact, especially for caregivers, demand a major effort for their prevention, correct diagnosis, and treatment. Thus, understanding their nature and origin is one of the most challenging enterprises of modern biomedical research. Clinical observations and epidemiological data reveal important sex differences in the prevalence and expression of various mental health problems and drug abuse.
A significant weakness in the research of psychiatric disorders and drug abuse is that despite all of the evidence suggesting that the susceptibility, prevalence, progression, and/or severity of this common disorders are sex-dependent, most studies with animals have been focused on one sex (usually males) or failed to report the sex of the animals (Beery and Zucker, 2011). In addition, females are underrepresented or even excluded in clinical and preclinical studies (Rubino and Parolaro, 2011). However, the majority of investigations examining estrogenic effects in the brain (e.g., neuroprotection, neuroinflammation, neurogenesis, migration) have been performed in females, which contrasts with the extended use of males in basic neuroscience research (Gillies and McArthur, 2010). This traditional view should be replaced with a new one that recognizes the sexual dimorphisms in the susceptibility to psychiatric disorders, which will help advance and evaluate the effectiveness of treatments depending on the sex. Here we provide more detailed information of these sexual dimorphisms to emphasize the urgent need to study both sexes to order to gain a proper understanding of these disorders.
Mechanisms Involved in Sexual Differentiation of the Reproductive and Central Nervous Systems
Sexual Differentiation of the Reproductive System
Sexual differentiation in mammals is initiated in utero during early gestation and is triggered, in large part, by the sex-determining region of the Y chromosome (SRY) gene located on the Y chromosome. The SRY gene encodes a protein termed testis-determining factor (TDF) that, along with SRY box-containing proteins like Sox9, directs the differentiation of the primordial gonadal cells into the testes (Maatouk and Capel, 2008; Bhandari et al., 2011). TDF inhibits the expression of a X-linked gene named DAX1, whose duplication within Xp21.3 causes XY sex reversal and therefore represses male-specific gene expression (Vilain and McCabe, 1998). SOX proteins stimulate the expression of steroidogenic factor 1 (SF-1; Shen and Ingraham, 2002), and Sox9 works together with SF-1 to stimulate the expression of Mullerian-inhibiting substance (MIS; a.k.a. Mullerian regression factor and anti-Mullerian hormone) in developing Sertoli cells (De Santa et al., 1998; Arango et al., 1999). Missense mutations in the Sox9 high mobility group (HMG) domain lead to XY sex reversal, whereas insertion of a color coat transgene into the Sox9 regulatory region upregulates expression and leads to XX sex reversal in mice (Bernard et al., 2003; Canastar et al., 2008). Human chorionic gonadotropin (hCG) secreted by the trophoblast also increases the expression of SF-1 (Manna et al., 1999), thereby stimulating the Leydig and Sertoli cells within the fetal testes to synthesize and secrete testosterone and MIS, respectively. Testosterone stimulates the Wolffian duct to develop into the male reproductive tract. In the primordial external genital tissue, testosterone is converted to dihydrotestosterone (DHT) via 5α-reductase, and DHT thus formed stimulates the development of accessory sex glands, penis, and scrotum. This androgen-induced masculinization of the external genitalia can be attributed, in part, to decreased Wnt/β-catenin signaling (Miyagawa et al., 2009). The testicular secretion of MIS elicits the apoptotic regression of the Mullerian duct by activating a Wnt4/β-catenin pathway in the Mullerian duct mesenchyme (Kobayashi et al., 2011). Without the hCG-induced MIS and testosterone secretion the Mullerian duct will develop into the female reproductive tract, while the Wolffian duct declines. This may also be attributed, in part, to Wnt/β-catenin signaling in the genetic/gonadal female that decreases the expression of Sox9 and MIS while promoting the expression of the female cell markers Bmp2 and Rspo1 (Bernard et al., 2012). The organizational effects imparted by a combination of in utero and neonatal testosterone surges are required for full expression of male characteristics (Naftolin et al., 1996; Resko and Roselli, 1997).
Sexual Differentiation of the Nervous System
The SRY/MIS/testosterone triad produces changes in the central nervous system (CNS) that parallel the divergence of the primordial gonads and external genitalia. SRY transcripts can be detected in murine brain as early as E11, and SRY expression continues on through postnatal day (PND) 30 in the midbrain, diencephalon, and cortex (Mayer et al., 2000). The SRY gene also regulates the expression of the X-linked monoamine oxidase type A gene in human male neuroblastoma cells, which may help explain sex differences in the prevalence of developmental neuropsychiatric disorders like autism and attention-deficit hyperactivity disorder (Wu et al., 2009). The expression of the SRY gene and the subsequent formation of the testes and secretion of testosterone masculinize the pattern of progesterone receptor expression in the brain (Wagner et al., 2004). In utero testosterone elicits an increase in the size of the sexually dimorphic nucleus of the preoptic area during late gestation (Roselli et al., 2011). In addition, cerebellar neurons express MIS receptors and the testicular secretion of MIS, along with testosterone, increases Purkinje cell number and cerebellar size in males (Wittmann and McLennan, 2011). However, recent evidence suggests that neuronal steroidogenesis may contribute to sexual differentiation of the rat brain as removal of peripheral sources of sex steroids at birth failed to abolish the sex- and region-specific changes in the brain concentrations of these hormones (Konkle and McCarthy, 2011).
There are also sex differences in gene expression occurring in the murine brain as early as 10.5 days post-coitum, which precedes the in utero testosterone surge (Dewing et al., 2003). Many of these genes reside on autosomes, and include Cyp7b (involved in neurosteroid metabolism) and Rorα4 (implicated in Purkinje cell differentiation). Others are located on either the X or Y chromosome, and among them are Xist (involved in X inactivation), Dby, and Eif2s3y (both of which are implicated in male fertility and other neural and behavioral phenotypes). Similar observations were made by Xu et al. (2002), who found that six X-linked murine genes (Usp9x, Ube1x, Smcx, Eif2s3x, Utx, and Dbx) were expressed to a higher degree in the brains of adult females, regardless of inactivation status. For some of these genes (Smcx, Eif2s3x, and Dbx), it appears that this differential expression is encountered as early as 13.5 days post-coitus. Not unexpectedly, the six homologs on the Y chromosome (Usp9y, Ube1y, Smcy, Eif2s3y, Uty, and Dby) are expressed in the brains of male but not female mice. Their expression was evident in genetic males lacking the SRY gene; indicating that testicular secretions are not required. The expression of at least some of these genes (Smcy, Eif2s3y, and Dby) occurs in a developmentally regulated fashion. Interestingly, the sex difference in the expression of the X-linked genes, at least in the adult murine brain, is not compensated by expression of the Y chromosome homologs in the brains of male mice. Given that these genes encode proteins involved in functions ranging from ubiquitination to protein translation; this suggests that the disparate dosages of these sex chromosomal genes may also contribute to sex differences in neural development, at least at some points along the developmental continuum.
Studies using the four-core genotype (FCG) mice, in which sex chromosome complement (XX versus XY) is unrelated to the animal’s gonadal sex, have revealed that inequities in sex chromosomal complement can also contribute to sexually dimorphic traits that would include the number of embryonic mesencephalic dopamine neurons and the density of vasopressinergic innervation of the lateral septum, both of which are greater in XY animals than in XX animals (Arnold and Chen, 2009). Other sexually differentiated neurobehavioral parameters influenced by sex chromosomal complement include aggression (greater in XY than XX females), pup retrieval (highest in XX females), social interaction (less in XX than XY females in response to an intruder), nociception (shorter response latencies in XX than in XY animals as assessed via hotplate and formalin tests), habit formation (greater in XX than in XY animals), as well as susceptibility to neural tube and autoimmune disorders (higher in XX mice; Arnold and Chen, 2009). The disparities in many of these neurobehavioral parameters are clearly shaped by organizational as well as activational effects of gonadal steroids (e.g., vasopressin expression in the septum, aggression, parental behavior, and nociception), whereas sex differences in other characteristics occur prior to, and therefore independent of, the in utero/neonatal testosterone surge (e.g., mesencephalic dopaminergic cell number, incidence of neural tube disorders; de Vries et al., 1986; Han and de Vries, 2003; Craft et al., 2004; Gatewood et al., 2006; Arnold and Chen, 2009). Regardless, SRY gene expression and the testosterone surges that occur as a result precludes a number of estrogenic responses including, but not limited to, calcium signaling in hypothalamic astrocytes and the generation of a preovulatory gonadotropin-releasing hormone (GnRH) surge, both of which happen only in gonadal females (Resko and Roselli, 1997; Kuo et al., 2010). However, while male brains are unresponsive to many estrogenic effects, aromatase is expressed in many brain regions throughout the critical period of brain sexual differentiation (Mura et al., 2010), which suggests that estrogens are instrumental for at least some aspects of male neurogenic development including cell proliferation, survival and death, synaptogenesis and dendritic branching, CpG methylation and histone deacetylation in the hypothalamus and limbic system (McCarthy and Arnold, 2011), as well as cognitive function and affective behaviors regulated by the frontal cortex (Peruffo et al., 2011).
Like the SRY gene product TDF, SOX proteins play a significant role in brain development and sexual differentiation of the CNS. Of these, SOX9 and SOX11 are expressed primarily within the CNS. SOX9 is highly conserved across phyla, and is expressed at high levels during the embryonic development of marsupial, murine, and human brain (Pask et al., 2002). SOX9 expression can be detected as early as 7.75 days post-coitus within cranial neural crest cells of the hindbrain neural folds, and its expression within the first pharyngeal arch is dependent upon the canonical Wnt/β-catenin signaling pathway (Bagheri-Fam et al., 2006). By 11.5 days post-coitum its expression extends to the ventricular zone of the neuroectoderm that gives rise to the forebrain, midbrain, hindbrain, and spinal cord (Bagheri-Fam et al., 2006). As early as E18.5 SOX9 expression is reported to extend out into the granular cell and Purkinje cell layers of the rodent cerebellum, and co-localize with the Bergmann glial cell marker vimentin (Pompolo and Harley, 2001; Scotting et al., 2005). In adult rodent forebrain SOX9 is co-expressed with the astrocyte marker S-100 (Pompolo and Harley, 2001). Collectively, this indicates that SOX9 is intimately involved in directing the fate neural crest and neural tube precursor cells toward a glial cell phenotype. As mentioned above, overexpression of SOX9 in the fetal gonads leads to XX sex reversal, and these XX males exhibit the same degree of aggression and the same pattern of mating behavior as their XY male counterparts (Canastar et al., 2008). This demonstrates that SOX9 makes an important contribution to the development of male-specific behaviors.
The transcription factor Sox11 and one of its binding partners required for transactivation of downstream target genes Brn1 also are upregulated during the late embryonic and neonatal periods of rat brain development (Kim et al., 2008). The expression of SOX11 and other SOX proteins like SOX2 and SOX4 is seen at E8.5 of murine in utero development, around the time of neurulation (Cheung et al., 2000). By E12, strong expression is observed in the sub-ventricular zone, where precursor cells have begun to differentiate (Cheung et al., 2000). The level of brain expression of Sox11 observed at E15.5–17.5 are ∼10× higher than at PND 7, and over 100× above that observed in the adult – with the highest levels of expression found in the cortex and other areas enriched with neuronal progenitors and immature neurons (Kim et al., 2008; Han et al., 2009; Ling et al., 2009). This expression temporally coincides with the heightened expression of components of the Wnt/β-catenin, p53, and tight junction signaling pathways (Ling et al., 2009). It is also observed in differentiating and proliferating P19 embryonic carcinoma cells (Ling et al., 2009). Sox11 expression in cultured dorsal root ganglia neurons and Neuro2a neuroblastoma cells elicits neurite outgrowth and differentiation (Jankowski et al., 2006). Conversely, knockdown of Sox11 expression in these cells reduces neurite outgrowth, and causes increased expression of the pro-apoptotic gene BNIP3 as well as decreased expression of the anti-apoptotic gene TANK and the gene encoding the actin organizing protein Arpc3 that is involved in axon growth (Jankowski et al., 2006). Thus, it is clear that the CNS is an organ that, much like the cells and tissues of the gonads, reproductive tract and external genitalia, is targeted by the SRY gene product TDF, MIS, testicular hormones, neurosteroids, and also possibly SOX proteins during the in utero and neonatal periods for the development of male patterns of behavior.
Contribution, Advantages, and Limitations of Animal Models in the Study of Psychiatric Disorders and Drug Abuse
The most accurate way to study psychiatric disorders and drug abuse would be using patients, but obviously this is not possible for ethical and practical reasons. Fortunately, animal models allow us to study molecular, protein and tissue changes, as well as behavioral alterations, that shed light on the origins and pathophysiologic sequelae of these diseases. They also provide high-value information about the effectiveness of medicines and the possible development and testing of treatments. As with different research tools, animal models have their pros and cons. One of the advantages of animal models is that the experiments performed on them are conducted under well-controlled conditions (Viveros et al., 2011b); with the expressed purpose of minimizing the inherent variability. The great majority of animal models used for psychiatric disorders and drug addiction involve rats and mice. The advantages of using these two species are that we have a very good knowledge of their anatomy, physiology, biochemistry, genetics, and behavior. Moreover, in recent years their genomes have been sequenced, which give us the opportunity to construct KO animals, analyze single-nucleotide polymorphisms (SNPs), target specific sequences within disease-associated alleles, and finally to compare their genomes with that of the human genome.
The major limitation of animal models for psychiatric disorders and drug dependence is that it is impossible to reproduce the syndrome in its entirety (Viveros et al., 2011b), because many of the symptoms used to diagnose psychiatric disorders in patients (e.g., hallucinations, suicide feelings, sadness, or guilt) are uniquely human. Another difficulty of these models is that despite the similarities between rodent brain and human brain, there are still some differences in functions and behaviors that must be taken into account in order to avoid misinterpreting the results. Thus, the generation of reliable animal models to study neuropsychiatric disorders continues to present a challenge to modern biomedical research (see Nestler and Hyman, 2010 for review).
Insights on Drug Abuse and Dependence from Experimental Models
The most common animal model or “standard tool” to study drug addiction is the drug self-administration paradigm that normally entails training an animal to self-administer a drug (intravenous or oral administration) by pressing a bar (Lynch et al., 2010). Along with other standardized behavioral assays (see Table 1), they collectively provide useful information on receptor pharmacology, neurotoxic lesions, or hormonal effects (Roberts et al., 2007).

Table 1. Standardized animal models to study different aspects of drug addiction.
As explained in the introduction, drug abuse, and dependence are sex-dependent and also present a general pattern – with men presenting higher rates of dependence and abuse and women being more vulnerable to drug negative effects. However, there are some particularities for the most commonly abused drugs.
Alcohol
Studies of alcohol abuse in human and animal models present a lack of correlation that is frequently explained by educational and sociocultural factors. It is known that women are less prone to develop alcohol abuse (Kalaydjian et al., 2009) whereas females mice present a higher degree of alcohol consumption than males (Lancaster et al., 1996). The use of the FCG mouse model described above has enabled Barker et al. (2010) to determine that sex chromosome complement affects habit formation depending on the reinforcer (possibly explaining the differences between men and women for at least some drugs or palatable foods), and confirm that females have greater free alcohol consumption. This suggests that there are differences in motivation to work for alcohol or disparities in the ability to track reinforcer availability.
Psychostimulants
Since the first preclinical research on cocaine sensitization was published, sex differences in response to the drug in adult rats have been noted. Many studies with animals have reported that females are more responsive, sensitize more readily than males to cocaine, and self-administer greater quantities of cocaine than males (Dow-Edwards, 2010). Other results showed that male and gonadectomized female rats had enhanced behavioral responses and brain metabolic activity following repeated cocaine administration, while changes in intact females were substantially less prominent (Dow-Edwards et al., 2005). These differences have been attributed historically to social or hormonal factors. Indeed it has been shown that gonadal steroid hormones exert organizational and activational effects within the CNS (Dluzen and McDermott, 2006), both of which could interact with cocaine abuse. However, recent studies involving sex chromosome complement suggest that genetic factors are also decisive in addiction (Barker et al., 2010). As described above these differences stem from disparate doses of genes residing on the X chromosome. Often the phenotypes differentiated by direct sex chromosome effects are also influenced by gonadal hormones, so understanding the interaction of sex-specific hormonal and sex chromosomal effects is an urgent need (McCarthy and Arnold, 2011). The effects of sex hormones and sex chromosomes have been investigated in behavioral, molecular, and immunohistochemical studies, but further evidence is needed, for example, with neuroimaging tools that show sex-dependent cocaine-induced changes in brain function (Andersen et al., 2012).
Marijuana
Preclinical studies have recently identified many of the factors that determine vulnerability to cannabis dependence. These include genetic and biological factors, animal strain differences (Deiana et al., 2007), as well as sex and ovarian hormones (Deiana et al., 2007; Fattore et al., 2007; Fattore and Fratta, 2010). Animal models suggest that there are sex-dependent differences in developing and maintaining cannabinoid self-administration (SA) behavior. Fattore et al. (2007) have shown that females acquire cannabinoid agonist intake at a higher rate than males, and that they are more susceptible to cannabinoid-induced rewarding effects and more reactive than males to saline substitution during the extinction period. In addition, the chronic use of tetrahydrocannabinol (THC) in adolescent females (but not males) subsequently induces depressive-like behaviors in adulthood (Rubino et al., 2008). Other preclinical studies show that sub-chronic adolescent exposure to CP (a CB1 receptor agonist) induces sex-dependent, long-term deleterious effects on memory function. Mateos et al. (2010) have shown that the detrimental effects of chronic cannabinoid use are sex-dependent, and that the diverse aspects of memory function may be differentially affected in each sex. CP also produces sexually dimorphic effects in other behaviors like motor activity (decreased in treated females), exploratory parameters (increased in treated females), and anxiety-like behavior (decreased in treated females; Biscaia et al., 2003; Llorente-Berzal et al., 2011). Cannabinoid treatment also increases the number of GFAP (a marker of astrocytes)-positive cells in the dentate gyrus of males but not females, and this was associated with decreased CB1 receptor immunoreactivity in CP-treated males (López-Gallardo et al., 2011). Many of the long-term cognitive and behavioral effects of adolescent cannabinoid treatment might be related to less synaptic contacts in the hippocampus (Rubino et al., 2008), and it is tempting to speculate that this disparate synaptic effect is sex-dependent and occurs in parallel with the differential behavioral impact (Viveros et al., 2011b). Early cannabinoid consumption alters the endocannabinoid system, which is an important component of reward system that may predispose individuals to the misuse of addictive drugs later in life as proposed in the “gateway hypothesis” (Kandel et al., 2006). Adolescent treatment with CP exerts sex-dependent effects on morphine reinforcement in adulthood. CP-treated males show a higher SA rate of morphine under a fixed-ratio schedule and decreased mu-opioid receptor functionality in the nucleus accumbens (NAc) shell, whereas females are not affected (Biscaia et al., 2008). This same effect was reported by Higuera-Matas et al. (2008) – female rats exposed to chronic CP treatment during puberty showed higher adult cocaine-SA and a higher reduction in anxiety when compared with males.
Animal Models of Psychiatric Disorders
As explained at the beginning of this section, although animal models do not reproduce every symptom of psychiatric disorders, they allow us to mimic some biological and behavioral changes. We will focus on experimental models based on the neurodevelopmental theory and the related “three-hit” hypothesis, where genetic factors, early life events and later life events should be considered as a “hit” separately. The neurodevelopmental theory states that abnormalities and disturbances during brain development, caused by both genetic and environmental factors, are likely to affect behavior later in life and lead to the emergence of psychiatric disorders such as schizophrenia, BP, or autism (Rehn and Rees, 2005; Fatemi and Folsom, 2009). Based on this theory, Maynard et al. (2001) developed the “two-hit” hypothesis of schizophrenia that could also be applied to other neurodevelopmental psychiatric disorders that begin in childhood or adolescence (Sanches et al., 2008). According to this hypothesis, genetic, and early environmental factors (first-hit) disrupt CNS development and together act as a vulnerability factor that produces a long-term susceptibility to an additional adverse event (second-hit), usually around puberty, that then precipitates the onset of schizophrenic symptoms (see Fernández-Espejo et al., 2009 for review). Other studies propose the three-hit hypothesis. The effects of these “hits” appear to be sexually dimorphic. However, there is very scarce information regarding sex differences in these models (Malone et al., 2010; Llorente et al., 2011; Llorente-Berzal et al., 2011). It would be especially interesting to include the factor “sex” in the so-called “match-mismatch hypothesis” that extends the concept of the neurodevelopmental theory by proposing that many diseases characterized by developmental risk factors reflect a mismatch between early environment and later environment. Thus, some human psychiatric disorders would appear when the physiologic, homeostatic, and stress systems do not adapt (i.e., match) with the current adult environment. In this context, sexually dimorphisms could be due to differences in adaptive plasticity and to the ability to respond properly to new environment conditions (Schmidt, 2011).
(1) “Early neurodevelopmental models” focus on genetic and environmental factors during the critical prenatal and perinatal period that produce brain developmental abnormalities, thereby allowing the study of the long-term physiological and behavioral consequences of early life stress. The predisposition toward sex-biased disorders has been associated with stress-related fetal antecedents (Pardo and Eberhart, 2007; Mueller and Bale, 2008). In many preclinical studies, prenatal stress (PS) has been shown to produce alterations in the HPA axis of the offspring that may give rise to anxiety, depressive-like behaviors, and cognitive deficits, all of which are endophenotypes associated with neuropsychiatric disease (Dunn et al., 2011). PS influences development through reprogramming and epigenetic processes; modifying key determinants of stress responses such as the expression of corticotrophin releasing factor (CRF) and glucocorticoid receptors (GR) by DNA methylation. As the brain is not completely developed during this period of early stress, it has been hypothesized that sex-specific changes in fetal development and long-term stress sensitivity may occur through interactions between the maternal environment, placental changes, and epigenetic programming of the embryo (Mueller and Bale, 2008; Dunn et al., 2011). Indeed, the placenta gene pattern differs in males and females; with lower expression of DNMT1, the enzyme responsible for methylation maintenance, observed in males than in females. Male offspring exposed to PS present altered stress coping responses and depression-like behaviors, whereas females are not affected (Mueller and Bale, 2008). This is in line with clinical studies showing a sex-dependent risk to develop schizophrenia in males due to stress early in pregnancy. Furthermore, Morgan and Bale (2011) have shown that these abnormalities can persist in subsequent generations by transmitting a stress-sensitive phenotype, suggesting the possibility of heritable epigenetic mechanisms.
Another interesting animal model of early life stress with which we have been working during the past several years consists of submitting rats to a single 24-h episode of maternal deprivation (MD) at PND 9. As adults, these MD rats exhibit abnormalities resembling psychotic-like symptoms, including disturbances in pre-pulse inhibition, latent inhibition, auditory sensory gating, and startle habituation. Possible underlying neurochemical correlates in adult MD animals include reduced hippocampal levels of neuropeptide Y (NPY), calcitonin-gene-related peptide, polysialylated neural cell adhesion molecule (NCAM), and brain-derived neurotrophic factor (BDNF), as well as a decrease in NMDA receptor subunits NR-2A and NR-2B (see for review Ellenbroek and Riva, 2003; Ellenbroek et al., 2004). In addition, adolescent MD rats showed depressive-like behaviors (Llorente et al., 2007), as well as a trend toward increased impulsivity (Marco et al., 2007). According to the neurodevelopmental hypothesis, it has been hypothesized that at least certain behavioral abnormalities observed in MD animals might be related to altered neurodevelopmental processes. Immunohistochemical results appeared to support this hypothesis, as it has been shown that MD-induced neuronal degeneration increased the number of GFAP-positive cells in the hippocampus and cerebellar cortex of neonatal rats, and altered the hippocampal endocannabinoid system. Interestingly, these effects were often more marked in males (Llorente et al., 2007; Suarez et al., 2009, 2010; Viveros et al., 2009). These results are in line with some epidemiological studies showing a higher degree of dysfunctional brain activity and neuroanatomy in SZ men (see Schizophrenia).
(2) “Late neurodevelopmental models” suggest that even in the absence of these early abnormalities in brain development, later life events could lead to the emergence of psychiatric disorders. These “late models” focus on adolescence that is a period of unique vulnerability for persistent effects of chronic stress. Experimental animal studies support that adolescence is a critical period in development, a “window of vulnerability” with respect to the onset of major neuropsychiatric disorders. Adverse events (e.g., drug exposure), during this period may serve a risk factor for suffering psychotic symptoms in adulthood (Adriani and Laviola, 2004; Fernández-Espejo et al., 2009). Although the effects of adolescent stress are not as well characterized as early postnatal stress models, the available literature on late neurodevelopmental models suggests that exposure to chronic stress during adolescence can be more detrimental than similar exposure in adulthood (Bourke and Neigh, 2011). Furthermore, studies with rats (Doremus-Fitzwater et al., 2009) and mice (Adriani and Laviola, 2000; Laviola et al., 2002) show that stress during adolescence modifies the reactivity of the HPA axis, which is disrupted in many psychiatric disorders (Dunn et al., 2011). The work of Bourke and Neigh (2011) demonstrates a sustained increase in depressive-like behaviors in female but not male rats following chronic adolescent stress. The results of this model contrast with those obtained from early developmental models where males are more affected by stress; thus suggesting that the sensitive periods also vary between males and females. Temporal differences in HPA axis development between males and females (Pignatelli et al., 2006) may account for prolonged sex-dependent effects induced by adolescent stress on physiology and behavior (Bourke and Neigh, 2011). Results from late development animal models are in line with clinical studies showing that girls react more strongly to stressful events than boys, and experience greater depressive symptomatology (Hankin et al., 2007).
Unfortunately, there is limited animal research data that addresses sex differences in PTSD. The animal model of PTSD that characterizes behavioral patterns in response to a stressor (i.e., predator scent stress) was used to investigate sex differences in rats (Mazor et al., 2009). Analyses did not find differences in prevalence rates of PTSD-like behaviors – although baseline levels of anxiety were higher for females and their peak levels of response to stress were lower (Mazor et al., 2009). Sex differences in stress response, endocrinology, and morphology at baseline have also been widely reported in the animal literature. Females generally demonstrate higher plasma and adrenal corticosterone at rest (Iwasaki-Sekin et al., 2009) that are believed to result from the effects of estrogen. Estrus cycle fluctuations in estrogens can significantly affect the morphology of the hippocampus in female rats, with higher estrogen resulting in increased dendritic spine density (Leranth et al., 2003). This is in line with the dimorphisms found in adaptive plasticity and the ability to cope and adapt to stressors, thus potentially influencing vulnerability to stress-related disease and disorder.
Furthermore, many examples from the literature document sex-related differences in behavior and neurobiology at baseline and in response to stress. In contrast with epidemiological patterns in humans, female rodents are more resilient following acute and chronic stress, whereas males are more vulnerable. This inverse relationship may in part reflect baseline differences in glucocorticoids. Female rodents have higher basal levels of corticosterone and more adaptive stress responses, whereas men have higher basal cortisol levels. These findings may imply that higher levels of cortisol could be protective against the development of PTSD. This hypothesis is consistent with studies showing an association between the development of PTSD and low peri-traumatic cortisol levels (McFarlane et al., 2011), which has been proposed as a pretraumatic risk factor for PTSD and stress-related psychopathology (Yehuda et al., 2010). Regardless, the development of PTSD following trauma requires further investigations to help reconcile the contradictory findings of sex differences in animal versus human research. These will undoubtedly help elucidate the true biological indicators of risk or pathophysiology, which will inform efforts at prophylaxis and intervention.
Sex Differences and the Adolescent Brain in Humans
As described above, shaping of the mammalian brain by sex hormones begins in utero (see Thornton et al., 2009 for a review of the organizational effects of androgens in non-human primates). Sexual dimorphism in the human brain, as assessed in vivo with magnetic resonance imaging (MRI), is present at birth. For example, the brain size of male neonates is about 6% larger than that of female neonates (Gilmore et al., 2007). This might be, at least in part, related to prenatal exposure to androgens; girls with male co-twins have slightly larger brains than girls with female co-twins (Peper et al., 2009). As reviewed recently, sex differences in global and regional brain volumes vary in their magnitude over the lifespan, with values of the effect size (Cohen’s d) ranging between 0.5 and 1.6 for global brain size and around 0.2 for regional volumes adjusted for brain size (Paus, 2010a).
Here, we will focus on sex differences in brain structure during human adolescence. A surge of sex hormones during puberty represents one hallmark of adolescence. But as pointed out by others, adolescence does not equal puberty: “puberty refers to the activation of the hypothalamic-pituitary-gonadal axis that culminates in gonadal maturation” while “adolescence refers to the maturation of adult social and cognitive behaviors” (Sisk and Foster, 2004). When interpreting sex differences in brain structure (and function) during adolescence, we should keep in mind the possibility that they reflect rather than drive sex differences in behavior. In other words, they may reflect experience-based changes in brain structure akin to those seen in other contexts (e.g., Draganski et al., 2004; Driemeyer et al., 2008). This possibility is of particular importance during this period of human development, which is characterized by significant changes in the daily life of an adolescent (over those of a child), such as the need to establish new social hierarchies in the context of (large) peer groups.
As pointed out above, MRI is the tool of choice for in vivo studies of developmental changes in the structure (and function) of the human brain (see Table 2 for an overview). The following are the most common MR sequences used in studies of brain development. High resolution, isotropic T1-weighted (T1W) images provide excellent contrast between gray and white matter and, as such, are well suited for morphometric studies of the human brain; T1W images yield a large number of outcome measures, including volumes of different brain compartments (e.g., gray and white matter volumes) and brain regions (e.g., volume of the hippocampus), as well as features of the cerebral cortex (e.g., thickness and folding). Diffusion Tensor Imaging (DTI) enables a virtual dissection of key white matter tracts (i.e., anatomic connectivity) that are necessary for communication amongst brain regions, as well as the assessment of micro-structural features (fractional anisotropy and mean diffusivity) that depend on axon packing, axon caliber, and myelination. Magnetization Transfer Ratio (MTR) provides an indirect index of the regional content of myelin. Functional MRI allows investigators to measure the brain response associated with the processing of various stimuli, cognitive operations, or emotional states; this measure is based on the regional coupling between brain activity and vasodilation. Readers interested in more detail information about these techniques are referred to other publications (e.g., Paus, 2010b).
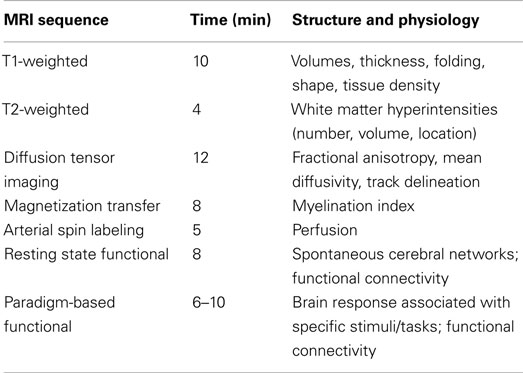
Table 2. Example of a 60-min MR protocol enabling one to characterize structural and functional properties of the human brain.
These techniques have been employed in various combinations in a number of large-scale studies of brain development, including (1) a cohort established in the Child Psychiatry Branch of the National Institutes of Mental Health (NIMH-CHPB); (2) NIH Pediatric MRI Database (NIH-PD; Evans and Brain Development Cooperative Group, 2006); (3) the Saguenay Youth Study (SYS; Pausova et al., 2007); and (4) the Imagen Study (Schumann et al., 2010). As described in more detail elsewhere (Paus, 2010c), these studies of typically developing children and adolescents have collected a wealth of data on the developmental trajectories of gray and white matter growth (NIMH-CHPB, NIH-PD, SYS), as well as on the functional engagement of brain networks during face processing, response inhibition, and reward (IMAGEN). Two of the studies (SYS, IMAGEN) are beginning to work on the identification of genes associated with various brain phenotypes in a genome-wide manner. Next, we will summarize two sets of findings obtained in the SYS. The first concerns sexual dimorphism in age-related changes in the volumes of white and gray matter, and the role of the androgen-receptor gene (AR) in moderating the influence of testosterone on these volumes in male adolescents. The second set describes apparent differences in the vulnerability of female (versus male) offspring to prenatal exposure to maternal cigarette smoking vis-à-vis several brain phenotypes.
As shown in Figure 1, both absolute and relative (brain size adjusted) volumes of white matter increase with age more steeply in male versus female adolescents (Perrin et al., 2008). In male adolescents, we also observed a moderating effect of a functional polymorphism in AR on the relationship between plasma levels of bioavailable testosterone and WM volume; this relationship was strong in boys with an “efficient” version of AR and weak in boys with an “inefficient” version of AR (Perrin et al., 2008). We also found that MTR, an indirect index of myelination, decreased with age in the white matter of male adolescents, suggesting that the observed testosterone-related changes might reflect an increase in axonal caliber instead (Perrin et al., 2008). We concluded that, during male adolescence, there might be a disproportionate growth of the axon and myelin sheath that would result in possible sex differences in the g ratio (defined as the ratio between axon caliber and fiber diameter). Next, we predicted that white matter regions occupied by large-diameter axons would have a lower concentration of myelin (Paus and Toro, 2009). We confirmed this prediction indirectly by observing age-related changes in the “whiteness” of the large-axon cortico-spinal tract (Herve et al., 2009) and, more directly, by showing that this fiber tract, assessed at the level of the internal capsule, has lower MTR values and longer T2 relaxation times than the adjacent (smaller axon) white matter (Herve et al., 2011). We speculated that the presumed testosterone-induced changes in axonal diameter during male adolescence increase the probability of a suboptimal g ratio in the large-diameter fibers and, in turn, decrease conduction velocity in these fibers, as well as perturb axonal transport in these axons (Paus and Toro, 2009). Such disturbances might interfere with the timing of inter-regional communication and contribute to the known earlier onset of schizophrenia in men versus women (Hafner et al., 1998).
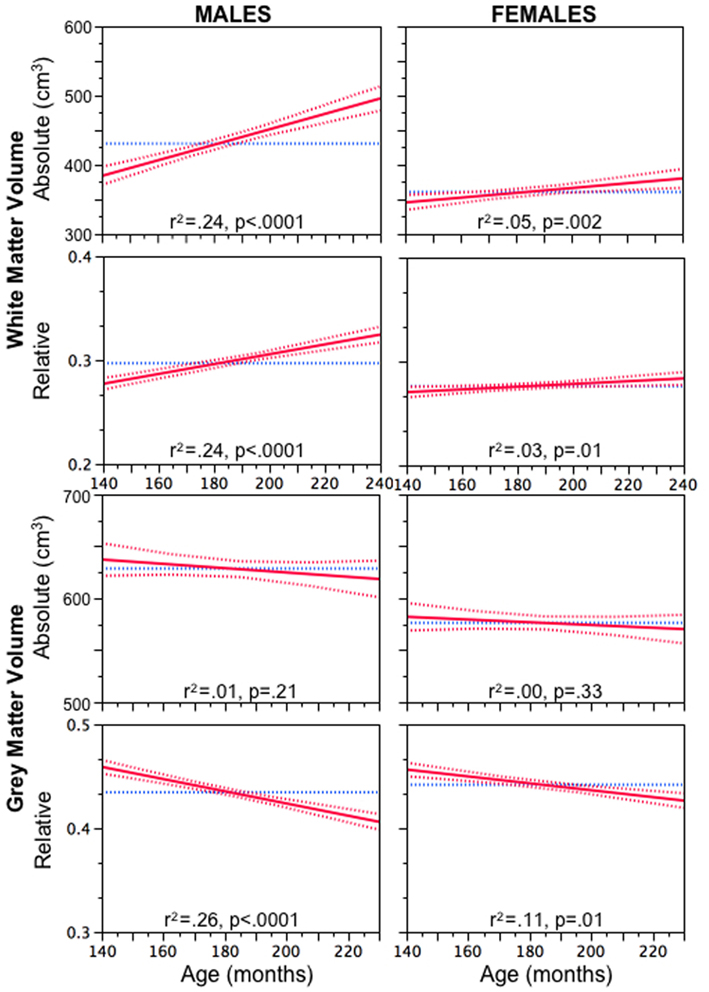
Figure 1. Age-related increase in absolute (top) and relative (brain size adjusted) volume of white matter in male and female adolescents. Reprinted with permission from Paus and Toro (2009).
The above age- and testosterone-related changes in white matter are mirrored in gray matter. As can be seen in Figure 2, regional age-related decreases in cortical gray matter are more pronounced in male versus female adolescents. Furthermore, the negative relationship between plasma levels of bioavailable testosterone and cortical gray matter is more pronounced in male adolescents with an “efficient” AR versus those with an “inefficient” AR (Paus et al., 2010). Although the neurobiological underpinnings of these age-related decreases in gray matter are unknown, they are unlikely to be related to synaptic pruning as have been commonly suggested (see Paus et al., 2008a for a discussion of this topic). At present, we are testing the possibility that these changes in gray matter might be also associated with testosterone-associated changes in intra-cortical axons.
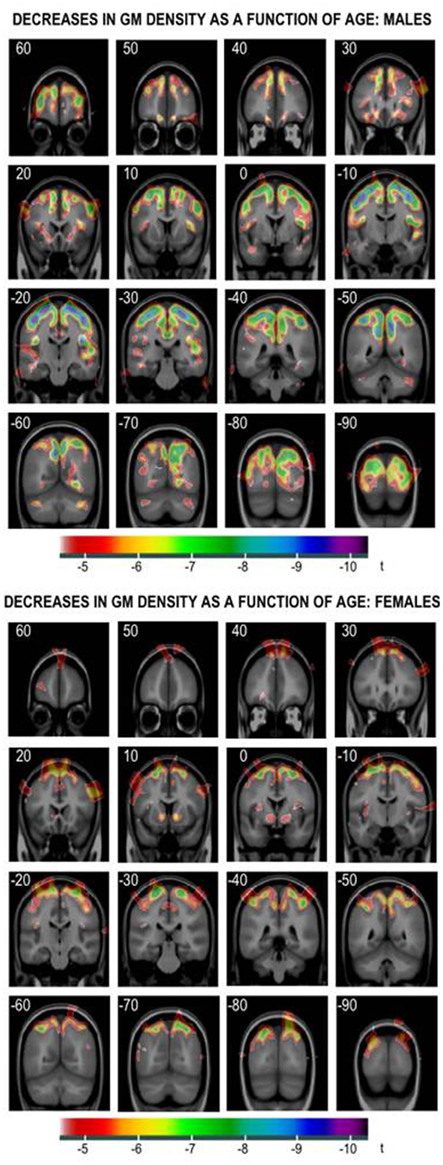
Figure 2. Age-related decreases in cortical gray matter in male (top) and female (bottom) adolescents. Reprinted with permission from Paus and Toro (2009).
The SYS has been designed to evaluate the possible long-term consequences of prenatal exposure to maternal cigarette smoking on the adolescent brain (Pausova et al., 2007). Given the limitations of epidemiological studies in examining associations between exposures and outcomes, we are, of course, unable to draw any causal conclusions from simply observing such associations. Nonetheless, results presented below raise interesting questions about the role of sex in moderating individuals’ vulnerability to an adverse intrauterine environment. When we compared “exposed” and “non-exposed” adolescents, the latter having been matched to the former by maternal education to minimize possible differences in socio-economic status, we observed that the exposed (versus non-exposed) adolescents had a thinner orbitofrontal cortex (Toro et al., 2008) and smaller corpus callosum (Paus et al., 2008b). Importantly, these effects appear more pronounced in female adolescents, especially in the case of the corpus callosum. More recently, we have carried out the first genome-wide scan for genes associated with brain size and identified KCTD8 at genome-wide significance in the sample of female (but not male) adolescents (Paus et al., 2011). When testing the most significant SNP of this gene (rs716890, p = 5.40E−09) vis-à-vis the total cortical area, we discovered an interaction of this polymorphism with prenatal exposure to maternal cigarette smoking: this SNP explained 21% of the variance in cortical area in the exposed female adolescents and only 4.5% (not significant) in the non-exposed females (Paus et al., 2011). We speculated that the absence of an interaction between KCTD8 genotype and exposure with respect to the area (and folding) of the cerebral cortex in male adolescents might be related to a protective effect of androgens in utero. These two sets of findings from the SYS illustrate the possible effects of sex hormones on the developing brain, effects that might either increase (testosterone, AR, and white matter) or decrease (testosterone, KCTD8, and cortical area) individuals’ vulnerability to brain disorders.
Sexual Dimorphisms in Psychiatric Disorders and Drug Abuse
Clinical observations and epidemiological data reveal important sex differences in the prevalence and symptomatology of various mental health problems and drug abuse. For example, the lifetime prevalence of major depressive disorder (MDD), as well as anxiety disorders combined, in women is approximately twice that of men (American Psychiatry Association (APA), 1994; Blazer et al., 1994; Scott, 2006; Smith et al., 2008), while the onset of schizophrenia is typically 3–5 years earlier and the course of the illness is more severe in men than in women (Flor-Henry, 1985; Hafner et al., 1998a; McGrath, 2007; Keshavan et al., 2008; McGrath and Susser, 2009). Interestingly, when it comes to the neurodevelopmental disorders, diagnosed in children, such as autism and attention-deficit hyperactivity disorder, the situation is reversed (i.e., they are more commonly diagnosed in boys). Concerning drug dependence and abuse, the general pattern of sex differences is that men have higher rates of dependence and drug abuse whereas women progress to dependence faster and are more vulnerable to the negatives effects of drugs. This is true for almost every drug, but some specific aspects for each will be explained below.
Overall, it is recognized that factors that interfere with the influence of sex hormones on the developing brain may permanently influence not only later behavior (e.g., cognition, emotion, empathy, aggression, etc.), but may also carry the risk of neuropsychiatric disorders. For example, the “extreme-male brain” hypothesis of autism proposes that the condition represents an extreme variant of male-typical neurocognitive profile (Baron-Cohen, 2009). Specifically, in the general population males tend to score higher on systemizing (defined as the tendency to analyze the rules of the system and predict its behavior), while females are better at empathizing (defined as the tendency to infer mental and emotional states of others and respond accordingly; a critical component of social cognition), but overall there is a significant overlap between the two sexes. In comparison, in autism-spectrum disorders the discrepancy between systemizing and empathizing is skewed in a male-typical direction and this profile may be a result of exposure to elevated levels of testosterone during fetal neurodevelopment (Baron-Cohen, 2002).
Schizophrenia is also considered to be a neurodevelopmental disorder with origins during second and third trimester of the fetal development (Szulc et al., 2005; Powell, 2010; Keshavan et al., 2011), which coincides with sexual differentiation of the brain. Interestingly, it has been recently suggested that autism and psychosis represent two extremes on a cognitive spectrum, with social cognition being underdeveloped (gaze-avoidance, “literal-mindedness,” etc.) in autistic-spectrum disorders and over-developed (“hyper-mentalizing,” over-interpretation of mental states and intentions etc.) in psychotic-spectrum disorders (the most severe being schizophrenia; Crespi and Badcock, 2008). It has been further proposed that as autism involves traits representative of an “extreme-male brain,” psychotic-spectrum disorders may be characterized by a “female-typical” neuroanatomy, cognition, and behaviors (Crespi and Badcock, 2008). However, while the “extreme-male brain” theory of autism works relatively well with males, it is harder to apply to females affected with autism-spectrum disorders (Barbeau et al., 2009). Similarly, while it is likely that males affected with psychosis may be de-masculinized/feminized, the situation may be different for females, as our data in chronic schizophrenia patients as well as the available literature (reviewed below) imply. It should be noted that the theories discussed in this section are not universally accepted and there are discordant data and views; nevertheless they provide interesting examples of how sex steroid hormones may interact with and contribute to psychiatric symptom.
For the purpose of the present review we narrowed our focus to sex differences in primarily adult psychopathologies, including mood disorders, schizophrenia, autism, and PTSD as well as drug dependence, as all of them present with striking sexually dimorphic features in the brain and clinical expression.
Schizophrenia
Schizophrenia (SZ) remains one of the most complex and debilitating disorders of the CNS. Its complexity is reflected in the heterogeneous clinical presentation with symptoms ranging from hallucinations and delusions, disorganized speech and behavior, to flat affect and lack of motivation (Rund and Borg, 1999; Green, 2006). The seriousness of SZ lies in the fact that its lifetime prevalence is approximately 1%, it begins typically during late adolescence or early adulthood, often leads to a social and economic impoverishment and a great distress for both patients and their families, and ends in suicide in about 10–15% of cases (50% of patients will attempt suicide at some point of their illness; Andreasen and Carpenter, 1993; Saha et al., 2007). The most recent studies suggest that the rate of 1% is not universal and while it is a good estimate in the western society, the prevalence of schizophrenia is lower in the developing countries, but higher in migrant groups than in native-born populations (Saha et al., 2005). The genetic basis of schizophrenia has been well documented in several twin and adoption studies (Kendler and Diehl, 1993; McGuffin et al., 1995), however the fact that approximately 50% of monozygotic twins are discordant for schizophrenia, indicates the importance of environmental factors (Takei et al., 1995; Mortensen et al., 1999). The etiology of SZ remains obscure.
Sex differences in clinical expression
The lifetime risk to develop schizophrenia is higher in young men, while beyond the age of 40, the risk is higher in women (Leung and Chue, 2000). This effect contributes to the earlier (approximately 3–5 years) age-at-onset of schizophrenia in men relative to women (Jablenksy, 1992; Hafner et al., 1998). Moreover, there is evidence that compared with women, men exhibit poorer premorbid functioning and greater IQ deficits (Flor-Henry, 1985). Finally, sex differences in the clinical expression of the disorder have been documented with men exhibiting on average more pronounced negative symptoms such as social withdrawal, blunted affect, poverty of speech, and abolition (Kay et al., 1986; Ring et al., 1991; Schultz et al., 1997), while women displaying more affective symptoms such as dysphoria, impulsivity, inappropriate affect, and more atypical psychotic symptoms (Shtasel et al., 1992; Andia et al., 1995; Szymanski et al., 1995).
Hormonal implications
Numerous epidemiological, neurochemical, and clinical studies imply a protective role of estrogen in schizophrenia (Seeman, 1996, 1997; Grigoriadis and Seeman, 2002). For example, inverse relationships have been found between menarche (i.e., age of first period in girls) and ages at the first psychotic symptoms and the first hospitalization (Cohen et al., 1999). Clinical reports show a correlation between estrogen plasma concentration and intensity of psychotic symptoms. During pregnancy, when estrogen levels are high, low rates of relapse have been observed in women with schizophrenia, while exacerbation of symptoms usually occurs postpartum (Chang and Renshaw, 1986; Kendell et al., 1987). In a similar fashion, symptoms have been reported to fluctuate across the menstrual cycle in women with schizophrenia, with clinical deterioration during follicular phase (low estrogen) and amelioration during mid-luteal phase (high estrogen; Hallonquist et al., 1993; Gattaz et al., 1994; Hendrick et al., 1996; Bergemann et al., 2007).
Although to a lesser degree than estrogen, testosterone has also been implicated in the pathophysiology of schizophrenia (Moffat, 2005; Mendrek, 2007). Early studies reported delayed puberty and low testosterone levels in males with schizophrenia (Kline et al., 1968), whereas more recent investigations have demonstrated an inverse relationship between the plasma level of testosterone and the severity of negative symptoms in male patients (Shirayama et al., 2002; Akhondzadeh et al., 2006; Ko et al., 2007).
Neuroanatomical data
Consistently with the direction of normal sexual dimorphism, neuroimaging, and post-mortem studies in SZ have shown that men have larger ventricles (Andreasen et al., 1990), and smaller overall frontal and temporal lobe volumes relative to women (Andreasen et al., 1994; Reite et al., 1997; Gur et al., 2000), although not all the studies have found this effect (Flaum et al., 1995; Lauriello et al., 1997). Goldstein and associates have found an interesting effect in the cingulate gyrus. Specifically, while the structure is typically larger in healthy women than in men (Paus et al., 1996; Goldstein et al., 2001), in SZ patients the opposite pattern was observed (Goldstein et al., 2002). These findings were subsequently replicated in SZ patients (Takahashi et al., 2003) and in individuals with schizotypal personality disorder (Takahashi et al., 2004). An equally interesting reversal of normal sexual dimorphism has been reported by Gur et al. (2002, 2004) in two other structures: orbitofrontal cortex and amygdala. In a different study, Frazier et al. (2008) reported that early onset SZ boys had smaller amygdala relative to girls, while the opposite pattern has been found in the general population. Preliminary results from our laboratory also imply a disturbed normal sexual dimorphism in brain structure in SZ. Thus, using the voxel-based morphometry (VBM) we have found that both male and female patients had significantly diminished prefrontal gray matter density compared with healthy controls, but only male patients showed preserved gray matter in the orbital surface of the inferior frontal cortex. In the parietal cortex, on the other hand, while again both male and female patients had significantly lower gray matter density compared to controls, only women patients had more preserved parietal cortex in the supramarginal gyrus and precuneus (Jiménez et al., 2008; Mancini-Marïe et al., 2008a,b; Figure 3). A recent study by Irle et al. (2011) suggests that the hippocampal volume is reduced in male but not in female patients at the onset of SZ (relative to the same-sex controls), but with progressing years SZ women “catch up” with men. This study emphasizes the importance of investigating neuroanatomical changes in a longitudinal manner in both sexes.
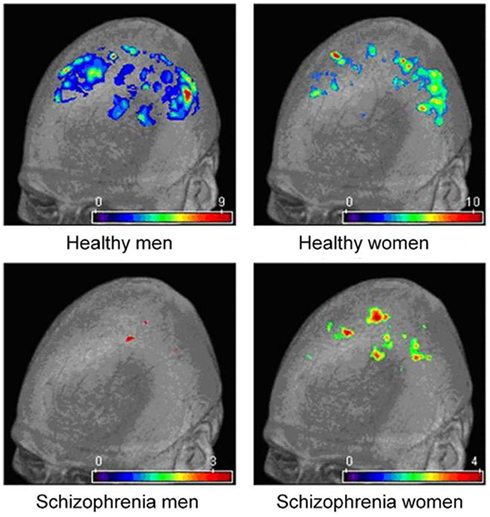
Figure 3. Voxel-based morphometry (VBM) analysis of parietal gray matter densities.
Functional neuroimaging studies
Despite numerous neuroimaging studies of cognitive function and emotion processing in schizophrenia, sex differences have not typically been examined. We have addressed this issue in a series of studies. Our initial investigation consisted of the re-analysis of already existing fMRI data sets collected in schizophrenia patients during processing of negative emotions. We have found a very different pattern of cerebral activations between men and women patients, but the absence of a healthy control group prevented us from drawing any definitive conclusions regarding the nature of this sexual dimorphism (Mendrek, 2007). Thus, we collected data in an independent sample of 42 patients (21 women) and 42 healthy controls during processing of emotional material, as well as during performance of a purely cognitive task of visuo-spatial processing, which involved mental rotation of 3D-figures (known to elicit robust sex differences in the general population). The behavioral data alone revealed an interesting reversal of normal sexual dimorphism, with superior performance of healthy males relative to healthy females, and the reverse pattern in patients (Jiménez et al., 2009, 2010). The fMRI results were consistent and showed more extensive cerebral activations (in the parietal and lateral prefrontal cortex) in healthy men relative to healthy women, while in patients it was women who exhibited more significant activations than men (Jiménez et al., 2009, 2010). Interestingly brain activations during the performance of mental rotation task by healthy males and schizophrenia females (but not in schizophrenia males and control females) were positively correlated with the plasma levels of testosterone (Mendrek et al., 2011). The brain activation during processing of emotional stimuli and during emotional memory also revealed disturbed sexual dimorphism in patients relative to controls (in preparation for publication). Altogether, the reversal of sexual dimorphism in brain processing in schizophrenia patients suggest that brain processing in male SZ patients may be feminized, while brain processing in female SZ patients may be masculinized. The mechanisms underlying this effect need to be further examined (e.g., organizational versus activational effects of sex steroid hormones, genetic versus environmental factors).
Depression
Major depressive disorder is characterized by affective, cognitive, and somatic symptoms such as: depressed mood, diminished interest and pleasure in most activities, significant weight loss (or weight gain), insomnia (or hypersomnia), fatigue, loss of energy, feelings of guilt, and worthlessness, diminished ability to concentrate, recurrent thoughts of death, and suicidal ideation. More than half of suicides occur in the context of depressive disorder (Barraclough et al., 1974; Hagnell and Rorsman, 1979). Twin and other family studies indicate that genetic factors contribute substantially (but less than in SZ or BP) to the liability for developing MDD, particularly in cases characterized by recurrent illness and early age-at-onset (30% in monozygotic twins; Sullivan et al., 2000). Stressful events are thought to interact with genetic susceptibility in the development of mood disorders and may contribute to the neurochemical imbalance. The disturbance in the serotonin and norepinephrine activity has been strongly implicated, as most antidepressant drugs exert their primary pharmacological actions through these systems (Goodwin, 1993; Trivedi et al., 2006; Ansorge et al., 2007; Dunlop and Nemeroff, 2007; Gartlehner et al., 2008; Kirsch et al., 2008).
Sex differences in clinical expression
The overall lifetime prevalence of MDD in women is approximately twice that of men (Kessler, 2003; Smith et al., 2008); ranging anywhere from 10 to 25% for women and 5 to 12% for men (Barraclough et al., 1974). This difference however is not apparent prior to puberty and in the years after menopause (Weissman and Olfson, 1995). Moreover, women are two to four times more likely than men to present with a seasonal component (Leibenluft, 1999; Leibenluft et al., 1999), atypical features (increased appetite, weight gain, hypersomnia), and higher levels of somatic complains, ruminations, feelings of worthlessness, and guilt (Hirschfeld et al., 1984). As the number of symptoms increases, so does the female/male prevalence ratio (Kornstein et al., 2000). Several explanations have been advanced to account for the differences between men and women in the prevalence and expression of MDD, including hormonal influences and psychosocial factors.
Hormonal implications
Sex differences in the prevalence of MDD have been typically attributed to the endocrine system, specifically the influence of estrogen in women. This explanation has been put forth because: (1) the rates of MDD are similar in girls and boys before puberty and the sex difference is less prominent in elderly people (Sprock and Yoder, 1997; Angst, 1998); (2) mood often appears to fluctuate with the change of hormones; times of low estrogen – such as the premenstrual and postpartum periods – are associated with increased risk for mood disorder (Fink et al., 1996). Approximately 13% of new mothers experience clinically significant depression during the first few months postpartum (O’Hara and Swain, 1996) with rates as high as 26% among adolescent mothers (Troutman and Cutrona, 1990) and 38% among low-income mothers (Hobfoll et al., 1995).
Psychosocial factors
In addition to the sex steroid hormones several psychosocial factors have been identified as potential contributors to the differential prevalence and expression of MDD in men and women. For example, the female sex role socialization has been associated with low self-esteem, low perceived control, pessimistic attributional styles, and dependency, all of which are vulnerability factors for developing depression (Grigoriadis and Robinson, 2007). In comparison, the masculine sex role socialization, which puts a high-value on assertiveness and independence, may play a protective role in men. However, the encouragement of stoicism and suppression of emotions may also prevent men from properly identifying their depressed mood and seeking treatment (Accortt et al., 2008).
Neuroanatomical data
Sex differences in the brains of MDD patients remain almost completely unexamined. In a study by Hastings et al. (2004) a different pattern of volumetric changes was observed in depressed men and women. Thus, relative to sex-matched controls, the anterior cingulate was smaller in depressed men compared with depressed women (this effect was in the opposite direction to that found in schizophrenia; result discussed above) – a difference consistent with the sexual dimorphism present in the general population, but even more pronounced. In addition, depressed women, but not depressed men, had a reduced volume of the amygdala compared with controls (another result in the opposite direction to what has been observed in schizophrenia patients). In the general population it is males who have larger amygdala and therefore the effect of a further reduction of the amygdala in depressed women points again to exaggerated sexual dimorphism in MDD patients. Overall, these results suggest that the biology of mood disorders in females may differ in some aspects from males, and may contribute to the higher rate of depression in women (Hastings et al., 2004).
Overall, the existing findings of sex differences in SZ (and to some extent BD) appear to be in the opposite direction to what has been reported in MDD. For example, we know that being a woman predisposes to the development of MDD and is associated with a more severe course of the illness. In schizophrenia, on the other hand, it is being a man that is related to the presence of more neuroanatomical and cognitive deficits, and a poorer treatment outcome. In addition, the evidence is emerging that the neuroanatomical sex differences in MDD are in the opposite direction to those observed in SZ and BD, though the effect has never been systematically investigated.
Posttraumatic Stress Disorder
Posttraumatic stress disorder is a psychiatric disorder that can occur following exposure to life threatening events that elicit overwhelming fear, helplessness, and horror. The disorder is characterized by the presence of three symptom groups: reexperiencing, avoidance, and hyperarousal. Reexperiencing symptoms are intrusions of the traumatic memory in the form of distressing images, nightmares, or dissociative experiences. These recollections serve to make the trauma-exposed person feel “haunted” by the memory of the event. This is not only because the memory is uncontrollable, but also because it generates physical and emotional responses associated with fear and terror. These responses result in avoidance behaviors, which include active attempts to avoid reminders of the traumatic event (including people, places, or activities associated with the trauma), and more passive attempts to avoid through emotional numbing and constriction of affect. Hyperarousal symptoms, such as insomnia, irritability, impaired concentration, hypervigilance, and increased startle responses reflect more “physiologic” manifestations of trauma exposure. Together, these symptoms impair social, occupational, or interpersonal functioning, and persist for at least a month following the trauma in order to meet criteria for PTSD according to DSM-IV. The U. S. National Comorbidity Survey Replication (NCS-R) estimates the lifetime prevalence of PTSD among adults to be 6.8% (Kessler et al., 2005).
Sex differences in clinical expression
Although men report significantly more exposure to potentially traumatic events than do women (with the exception of childhood trauma), epidemiological studies find higher rates of PTSD among women than men (Helzer et al., 1987; Davidson et al., 1991; Breslau et al., 1998; Breslau and Anthony, 2007). In the U.S., lifetime prevalence rates of PTSD are estimated at 3.6% among men and 9.7% among women (National Comorbidity Survey, 2005). Furthermore, women with PTSD report more reexperiencing symptoms and more severe symptoms (Zlotnick et al., 2001; Tolin and Foa, 2006), and worse quality of life outcomes than do men with the disorder (Holbrook et al., 2002). Numerous theories regarding women apparently heightened vulnerability to the development of PTSD have been proposed, including the intimate, interpersonal nature of potentially traumatic events that women are more likely to experience (i.e., higher rates of severe intimate partner violence and sexual assault across the lifespan), the profoundly corrosive, and damaging meanings inherent in these kinds of violations, the possibility that women may be prone to heightened levels of emotional reactivity which could confer additional risk for PTSD, and the potential that sex differences in behavioral and psychological sequelae to trauma influence diagnostic presentation (e.g., internalizing versus externalizing symptoms; Pratchett et al., 2010).
Hormonal implications
Biologically informed research on PTSD has particularly focused on the hypothalamic-pituitary-adrenal (HPA) axis, the major constituent of the neuroendocrine response to acute and chronic stress (Yehuda, 2006). Although the findings have not always been consistent, studies of the HPA axis and PTSD document a distinct neuroendocrine profile in that catecholamine and CRF levels appear to be increased in this disorder but urinary and plasma levels of cortisol have been found to be lower than in normal (for review, see Yehuda, 2002). Such findings have led to the development of a model of PTSD in which increased glucocorticoid receptor sensitivity leads to reduced levels of circulating cortisol, contributing to a failure of the autoregulation system to contain sympathetic activation and re-establish homeostasis following removal of a threat.
Data on sex differences in PTSD-related biology are limited. However, research on sex differences in the neuroendocrinology of PTSD has documented some differences in cortisol and catecholamines among subjects with PTSD. For example, among children with PTSD, girls had significantly elevated cortisol compared to boys (Carrion et al., 2002), and a cortisol study of adults with PTSD found that women’s salivary cortisol levels decreased throughout the day whereas men’s levels increased (Freidenberg et al., 2010). Epinephrine and norepinephrine have been associated with PTSD symptoms at 1 and 5 months following trauma exposure for men but not for women (Hawk et al., 2000). CRF receptor functions have also been found to render women more sensitive to low CRF and less adaptable to high levels, potentially contributing to women’s disproportionate development of PTSD (Bangasser et al., 2010).
Neuroanatomical data
Neuroanatomical differences in human males and females with PTSD have also been identified. For example, while MRI studies of children with maltreatment-related PTSD found overall differences in the brains of children with PTSD versus controls, they also found sex differences within the PTSD group. Males had greater corpus callosum reduction (De Bellis et al., 1999), larger prefrontal lobe cerebrospinal fluid volumes and smaller splenium, smaller cerebral volumes and corpus callosum regions 1 (rostrum) and 6 (isthmus), and greater lateral ventricular volume increases (De Bellis and Kehavan, 2003). Functional MRI research has also found that enhanced brainstem activity is associated with a diagnosis of PTSD for men but not for women, and that men with PTSD show greater hippocampal activity in response to fear than their female counterparts. However, observed differences may reflect normative sex differences on these parameters rather than sex differences in pathophysiology or risk for PTSD. For example, adult males have higher basal cortisol levels than women (e.g., Vierhapper et al., 1998; Takai et al., 2007).
Molecular biologic data
Recent data from emerging molecular biologic studies of PTSD have also identified sex differences. For example, a recent study of gene expression in PTSD found differential patterns of monocyte gene expression in men and women (Neylan et al., 2011). Whereas men with PTSD (n = 24) were found to have three under-expressed genes, women PTSD subjects (n = 10) had both increased and decreased gene expression. Furthermore, men demonstrated no evidence of chronic inflammation, as hypothesized, but women did demonstrate elevated inflammatory activity. Separate analyses of these data (O’Donovan et al., 2011) also found that men and women differed on regulation of target genes for CREB/ATF, with men showing up-regulation and women showing down-regulation. Males and females both showed up-regulation of target genes for the NF-KB/Rel family of transcription factors and down-regulation of target genes for the glucocorticoid receptor. Sex differences have also been identified in the associations of PTSD with pituitary adenylate cyclase-activating polypeptide (PACAP), involved in cellular stress response, and the PAC1 receptor pathway (Ressler et al., 2011). PACAP blood levels were associated with PTSD symptoms and diagnosis, and with conditioned fear response, for women only. PACAP levels predicted response on the three symptom clusters of PTSD for women but not for men. A SNP in an estrogen response element in the PAC1 receptor predicted PTSD symptoms and diagnosis for women only, implicating estrogen in the regulation of pathways relevant to sex differences in rates of PTSD symptomatology and diagnosis.
Drug Abuse
The National Institute on Drug Abuse has described drug addiction as a chronic, relapsing brain disease that is characterized by compulsive drug seeking and use, despite harmful consequences2. Addiction is the progression from impulsive to compulsive behavior of drug taking and it occurs in three stages: (1) binge/intoxication; (2) withdrawal/negative affect; and (3) preoccupation/anticipation (Viveros et al., 2011a). Substance abuse and addiction are enormous public health concerns in many areas and they are often linked to other psychiatric disorders (Lynch et al., 2010). The results of animal models and clinical studies show that there are sex differences at every phase of the drug reinforcement process. For the majority of drugs of abuse, women are more vulnerable (Becker and Hu, 2008) and exhibit higher rates of self-administration (Fattore et al., 2007; Biscaia et al., 2008). Sexual dimorphisms in drug abuse patterns are observed in humans in the intake of psychoactive stimulants, alcohol, opioids, and marijuana (Becker and Hu, 2008; Gillies and McArthur, 2010). Such differences have been attributed historically to sociocultural and hormonal factors (Barker et al., 2010); however, an increasing body of evidence indicates that these cultural influences are changing and that the use, abuse, and dependence are increasing faster in women.
Sex differences in epidemiological studies
Epidemiological studies show that men have higher rates of alcohol dependence and abuse (Kalaydjian et al., 2009); however, animal models show that females are more prone to consume alcohol than males (Lancaster et al., 1996; Middaugh et al., 1999). This lack of correlation between humans and animals models could be due to sociocultural factors, but this trend in alcohol consumption and abuse is changing as well as for other drugs as cocaine. Studies with psychostimulants show that cocaine abuse has particularly increased among women, and mounting evidence shows that women are more vulnerable to some aspects of psychostimulants abuse. Epidemiological studies in the late 1980s (Griffin et al., 1989; Mendelson et al., 1991) indicated that women begin using cocaine and amphetamines earlier than men, that they have more severe cocaine use and that women progress to dependence faster than do men (known as a “telescoped course”). Additionally, addicted women present with a higher incidence of comorbid psychiatric conditions, such as depression or PTSD (Najavits and Lester, 2008) and also women show higher levels of craving and longer periods of use after abstinence than men (Becker and Hu, 2008). There are also evidences proving that humans present sex differences in biological and behavioral effects of cannabis use (Fattore et al., 2008), but the mechanisms or origin of these differences are not well known yet. Men present higher circulating levels of cannabis psychoactive compound, the tetrahydrocannabinol (THC), are more sensitive to the subjective effects of THC (Haney, 2007), present more withdrawal symptoms (Crowley et al., 1998), and are more likely to be polysubstance users and show a higher prevalence in panic and personality disorders after abuse (Hasin et al., 2008).
Hormonal implications
Some studies found that the effects of psychostimulants vary across the menstrual cycle – they are potentiated during the late luteal phase (when estradiol and progesterone levels are declining; Becker and Hu, 2008). Thus, the basis for the sex difference in response to cocaine is generally believed to be due to estrogen (Dow-Edwards, 2010). Animal models of cocaine self-administration reveal sex differences in the acquisition phase that are independent of hormonal factors, with females being more vulnerable than males. However, the motivation to take cocaine is hormone dependent; estradiol enhances the motivation of cocaine intake and it plays a key role in the reinforcing effects of psychostimulants (Becker and Hu, 2008). Clinical studies show that women are more vulnerable than men to the transition from recreational use to drug abuse (Randall et al., 1999), and that they are more sensitive to the addictive properties of drugs (Lynch et al., 2002). However, there is some evidence suggesting that women experience weaker effects of THC. This could be due to sex differences in drug disposition and the distribution of fat tissue (this is a hormone dependent factor) that helps sequester lipophilic compounds like cannabinoids, as well as the possibility that women may metabolize cannabinoid compounds faster than men. The lack of correlation with some preclinical studies may be because male rodents have a higher percentage of fat tissue than females. As a result, more THC would be retained by fat cells (Fattore and Fratta, 2010), which would diminish the amount of THC reaching the brain and thereby affect the behavioral effects of cannabinoids (Rubino and Parolaro, 2011). It is also important to notice that there is an overlap between stress-system responses and the effects of drugs because the majority of them are able to stimulate the hypothalamic-pituitary-adrenal axis. In laboratory animals, some stress models have been shown to increase the acquisition and reinstatement of stimulants, opiates, and alcohol (Fox and Sinha, 2009). This same relationship has been observed in humans, where chronic stress plays a very important role in the motivation to abuse drugs and alcohol (Sinha, 2001). HPA axis responses are also sex-dependent, thus providing another possible explanation for why women are more vulnerable than men to the negative effects of drugs (Fox and Sinha, 2009).
Neuroanatomical data
The reward system is a group of brain structures which regulate and control behavior by inducing pleasurable effects. The major rewarding pathway in the brain is the mesolimbic pathway and goes from the ventral tegmental area (VTA) via the medial forebrain bundle to the NAc, which is the primary release site for the neurotransmitter dopamine (Viveros et al., 2011a). Epidemiological studies suggest that some differences in vulnerability to drug abuse could be due to dimorphisms in reward pathways or dopamine release as well as in reward-related structures like NAc, PFC, or amygdale; this last has been shown to be influenced by pubertal hormones in structural and organizational events (see Gillies and McArthur, 2010 for review).
Concluding Remarks
In conclusion, what experimental and epidemiological studies tell us is that sex differences in the brain appear to be predominantly established during developmentally sensitive time intervals (i.e., in utero, perinatal, and adolescence periods). According to the classical view of brain sexual differentiation, organizational effects are permanent structural changes arising from the actions of in utero and perinatal sex hormones, whereas activational effects are direct and temporary influences of circulating gonadal hormones on function and behavior following puberty. However, we now know that the SRY can begin directing sexual differentiation prior to onset of hormonal secretions from the fetal testes. In addition, sex chromosomal complement and transcription factors like Sox9 make important contributions to the process of sexual differentiation. Moreover, it has been recently proposed that steroid-dependent organization of behavior may also occur during adolescence, which indicates the possibility of a reassessment of the developmental time-frame for organizational effects of gonadal steroids (Weinberg et al., 2008; Schulz et al., 2009). Both preclinical and clinical studies show that the effects of stressful events are sex and time-dependent. The susceptibility to disease or dysfunction and the effect of injury/stress can be as much as twofold to fivefold greater in one sex; depending on the psychiatric disorder. These include higher rates of neuropsychiatric and learning disorders with developmental origins in males and higher rates of aging related neurodegenerative diseases and mental health dysfunctions in females (McCarthy and Arnold, 2011). For a long time, some of these differences has been explained by the influence of genetic background, circulating levels of gonadal hormones, or educational and psychosocial factors in humans, however epigenetic and reprogramming processes are becoming more and more important regarding the sexually dimorphisms of psychiatric disorders. The combination of genomics, proteomics, metabolomics, behavioral, neuroimaging, and neuroendocrinological techniques, as well as the performance of studies with both sexes, could lead to a better understanding of these disorders and to the development of effective treatments depending on sex because, as both human and animal studies suggest: sex matters.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by Instituto de Salud Carlos III, Redes temáticas de Investigación Cooperativa en salud (ISCIII y FEDER): Red de trastornos adictivos RD06/0001/1013; GRUPOS UCM-BSCH – 951579; Ministerio de Ciencia e Innovación BFU2009-10109, and Plan Nacional sobre Drogas at SAS/1250/2009 official notification.
Footnotes
References
Accortt, E. E., Freeman, M. P., and Allen, J. J. (2008). Women and major depressive disorder: clinical perspectives on causal pathways. J. Womens Health (Larchmt.) 17, 1583–1590.
Adriani, W., and Laviola, G. (2000). A unique hormonal and behavioral hyporesponsivity to both forced novelty and d-amphetamine in periadolescent mice. Neuropharmacology 39, 334–346.
Adriani, W., and Laviola, G. (2004). Windows of vulnerability to psychopathology and therapeutic strategy in the adolescent rodent model. Behav. Pharmacol. 15, 341–352.
Akhondzadeh, S., Rezaei, F., Larijani, B., Nejatisafa, A. A., Kashani, L., and Abbasi, S. H. (2006). Correlation between testosterone, gonadotropins and prolactin and severity of negative symptoms in male patients with chronic schizophrenia. Schizophr. Res. 84, 405–410.
American Psychiatry Association (APA). (1994). Diagnostic and Statistical Manual of Mental Disorders. Washington, DC: American Psychiatric Press.
Andersen, M. L., Sawyer, E. K., and Howell, L. L. (2012). Contributions of neuroimaging to understanding sex differences in cocaine abuse. Exp. Clin. Psychopharmacol. 20, 2–15.
Andia, A. M., Zisook, S., Heaton, R. K., Hesselink, J., Jernigan, T., Kuck, J., Morganville, J., and Braff, D. L. (1995). Gender differences in schizophrenia. J. Nerv. Ment. Dis. 183, 522–528.
Andreasen, N. C., and Carpenter, W. T. Jr. (1993). Diagnosis and classification of schizophrenia. Schizophr. Bull. 19, 199–214.
Andreasen, N. C., Ehrhardt, J. C., Swayze, V. W. II, Alliger, R. J., Yuh, W. T., Cohen, G., and Ziebell, S. (1990). Magnetic resonance imaging of the brain in schizophrenia. The pathophysiologic significance of structural abnormalities. Arch. Gen. Psychiatry 47, 35–44.
Andreasen, N. C., Flashman, L., Flaum, M., Arndt, S., Swayze, V. II, O’Leary, D. S., Ehrhardt, J. C., and Yuh, W. T. (1994). Regional brain abnormalities in schizophrenia measured with magnetic resonance imaging. JAMA 272, 1763–1769.
Ansorge, M. S., Hen, R., and Gingrich, J. A. (2007). Neurodevelopmental origins of depressive disorders. Curr. Opin. Pharmacol. 7, 8–17.
Arango, N. A., Lovell-Badge, R., and Behringer, R. R. (1999). Targeted mutagenesis of the endogenous mouse Mis gene promoter: in vivo definition of genetic pathways of vertebrate sexual development. Cell 4, 409–419.
Arnold, A. P., and Chen, X. (2009). What does the four core genotypes mouse model tell us about sex differences in the brain and other tissues? Front. Neuroendocrinol. 30, 1–9.
Bagheri-Fam, S., Barrionuevo, F., Dohrmann, U., Gunther, T., Schule, R., Kemler, R., Mallo, M., Kanzler, B., and Scherer, G. (2006). Long-range upstream and downstream enhancers control distinct subsets of the complex spatiotemporal Sox9 expression pattern. Dev. Biol. 291, 382–397.
Bangasser, D. A., Curtis, A., Reyes, B. A., Bethea, T. T., Parastatidis, I., Ischiropoulos, H., Van Bockstaele, E. J., and Valentino, R. J. (2010). Sex differences in corticotropin-releasing factor receptor signaling and trafficking: potential role in female vulnerability to stress-related psychopathology. Mol. Psychiatry 15, 896–904.
Barbeau, E. B., Mendrek, A., and Mottron, L. (2009). Are autistic traits autistic? Br. J. Psychol. 100, 23–28.
Barker, J. M., Torregrossa, M. M., Arnold, A. P., and Taylor, J. R. (2010). Dissociation of genetic and hormonal influences on sex differences in alcoholism-related behaviors. J. Neurosci. 30, 9140–9144.
Baron-Cohen, S. (2002). The extreme male brain theory of autism. Trends Cogn. Sci. (Regul. Ed.) 6, 248–254.
Baron-Cohen, S. (2009). Autism: the empathizing-systemizing (E-S) theory. Ann. N. Y. Acad. Sci. 1156, 68–80.
Barraclough, B., Bunch, J., Nelson, B., and Sainsbury, P. (1974). A hundred cases of suicide: clinical aspects. Br. J. Psychiatry 125, 355–373.
Beery, A. K., and Zucker, I. (2011). Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 35, 565–572.
Bergemann, N., Parzer, P., Runnebaum, B., Resch, F., and Mundt, C. (2007). Estrogen, menstrual cycle phases, and psychopathology in women suffering from schizophrenia. Psychol. Med. 37, 1427–1436.
Bernard, P., Ryan, J., Sim, H., Czech, D. P., Sinclair, A. H., Koopman, P., and Harley, V. R. (2012). Wnt signaling in ovarian development inhibits Sf1 activation of Sox9 via the Tesco enhancer. Endocrinology 153, 901–912.
Bernard, P., Tang, P., Liu, S., Dewing, P., Harley, V. R., and Vilain, E. (2003). Dimerization of SOX9 is required for chondrogenesis, but not for sex determination. Hum. Mol. Genet. 12, 1755–1765.
Bhandari, R. K., Sadler-Riggleman, I., Clement, T. M., and Skinner, M. K. (2011). Basic helix-loop-helix transcription factor TCF21 is a downstream target of the male sex determining gene SRY. PLoS ONE 6, e19935.
Biscaia, M., Fernandez, B., Higuera-Matas, A., Miguens, M., Viveros, M. P., Garcia-Lecumberri, C., and Ambrosio, E. (2008). Sex-dependent effects of periadolescent exposure to the cannabinoid agonist CP-55,940 on morphine self-administration behaviour and the endogenous opioid system. Neuropharmacology 54, 863–873.
Biscaia, M., Marin, S., Fernandez, B., Marco, E. M., Rubio, M., Guaza, C., Ambrosio, E., and Viveros, M. P. (2003). Chronic treatment with CP 55,940 during the peri-adolescent period differentially affects the behavioural responses of male and female rats in adulthood. Psychopharmacology (Berl.) 170, 301–308.
Blazer, D. G., Kessler, R. C., McGonagle, K. A., and Swartz, M. S. (1994). The prevalence and distribution of major depression in a national community sample: the National Comorbidity Survey. Am. J. Psychiatry 151, 979–986.
Bourke, C. H., and Neigh, G. N. (2011). Behavioral effects of chronic adolescent stress are sustained and sexually dimorphic. Horm. Behav. 60, 112–120.
Breslau, N., and Anthony, J. C. (2007). Gender differences in the sensitivity to posttraumatic stress disorder: an epidemiological study of urban young adults. J. Abnorm. Psychol. 116, 607–611.
Breslau, N., Kessler, R. C., Chilcoat, H. D., Schultz, L. R., Davis, G., and Andreski, P. (1998). Trauma and posttraumatic stress disorder: the 1996 Detroit area survey of trauma. Arch. Gen. Psychiatry 55, 626–632.
Campbell, U. C., and Carroll, M. E. (2000). Acquisition of drug self-administration: environmental and pharmacological interventions. Exp. Clin. Psychopharmacol. 8, 312–325.
Canastar, A., Maxson, S. C., and Bishop, C. E. (2008). Aggressive and mating behaviors in two types of sex reversed mice: XY females and XX males. Arch. Sex. Behav. 37, 2–8.
Carrion, V., Weems, C., Ray, R., Glaser, B., Hessl, B., and Reiss, A. (2002). Diurnal salivary cortisol in pediatric posttraumatic stress disorder. Biol. Psychiatry 51, 575–582.
Cheung, M., Abu-Elmagd, M., Clevers, H., and Scotting, P. J. (2000). Roles of Sox4 in central nervous system development. Brain Res. Mol. Brain Res. 79, 180–191.
Cohen, R. Z., Seeman, M. V., Gotowiec, A., and Kopala, L. (1999). Earlier puberty as a predictor of later onset of schizophrenia in women. Am. J. Psychiatry 156, 1059–1064.
Craft, R. M., Mogil, J. S., and Aloisi, A. M. (2004). Sex differences in pain and analgesia: the role of gonadal hormones. Eur. J. Pain 8, 397–411.
Crespi, B., and Badcock, C. (2008). Psychosis and autism as diametrical disorders of the social brain. Behav. Brain Sci. 31, 241–261.
Crowley, T. J., MacDonald, M. J., Whitmore, E. A., and Mikulich, S. K. (1998). Cannabis dependence, withdrawal, and reinforcing effects among adolescents with conduct symptoms and substance use disorders. Drug Alcohol Depend. 50, 27–37.
Davidson, J. R., Hughes, D., Blazer, D. G., and George, L. K. (1991). Post-traumatic stress disorder in the community: an epidemiological study. Psychol. Med. 21, 713–721.
De Bellis, M., and Kehavan, M. (2003). Sex differences in brain maturation in maltreatment-related pediatric posttraumatic stress disorder. Neurosci. Biobehav. Rev. 27, 103–117.
De Bellis, M., Keshavan, M., Clark, C., Casey, B., Giedd, J., Boring, A., Frustaci, K., and Ryan, N. (1999). A.E. Bennett Research Award. Developmental traumatology. Part II: Brain development. Biol. Psychiatry 45, 1271–1284.
De Santa, B. P., Bonneaud, N., Boizet, B., Desclozeaux, M., Moniot, B., Sudbeck, P., Scherer, G., Poulat, F., and Berta, P. (1998). Direct interaction of SRY-related protein SOX9 and steroidogenic factor 1 regulates transcription of the human anti-Mullerian hormone gene. Mol. Cell. Biol. 18, 6653–6665.
de Vries, G. J., Duetz, W., Buijs, R. M., van Heerikhuize, J., and Vreeburg, J. T. (1986). Effects of androgens and estrogens on the vasopressin and oxytocin innervation of the adult rat brain. Brain Res. 399, 296–302.
Deiana, S., Fattore, L., Spano, M. S., Cossu, G., Porcu, E., Fadda, P., and Fratta, W. (2007). Strain and schedule-dependent differences in the acquisition, maintenance and extinction of intravenous cannabinoid self-administration in rats. Neuropharmacology 52, 646–654.
Dewing, P., Shi, T., Horvath, S., and Vilain, E. (2003). Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Brain Res. Mol. Brain Res. 118, 82–90.
Dluzen, D. E., and McDermott, J. L. (2006). Estrogen, testosterone, and methamphetamine toxicity. Ann. N. Y. Acad. Sci. 1074, 282–294.
Doremus-Fitzwater, T. L., Varlinskaya, E. I., and Spear, L. P. (2009). Social and non-social anxiety in adolescent and adult rats after repeated restraint. Physiol. Behav. 97, 484–494.
Dow-Edwards, D. (2010). Sex differences in the effects of cocaine abuse across the life span. Physiol. Behav. 100, 208–215.
Dow-Edwards, D., Dasari, V. M., Menendez, R., Lugo-Escobar, N., Torres-Reveron, A., and Segarra, A. C. (2005). Alterations in brain glucose metabolism following repeated cocaine in adult rats: sex and gonadal status. Abstr. Soc. Neurosci.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., and May, A. (2004). Neuroplasticity: changes in grey matter induced by training. Nature 427, 311–312.
Driemeyer, J., Boyke, J., Gaser, C., Buchel, C., and May, A. (2008). Changes in gray matter induced by learning – revisited. PLoS ONE 3, e2669.
Dunlop, B. W., and Nemeroff, C. B. (2007). The role of dopamine in the pathophysiology of depression. Arch. Gen. Psychiatry 64, 327–337.
Dunn, G. A., Morgan, C. P., and Bale, T. L. (2011). Sex-specificity in transgenerational epigenetic programming. Horm. Behav. 59, 290–295.
Ellenbroek, B. A., de Bruin, N. M., van Den Kroonenburg, P. T., van Luijtelaar, E. L., and Cools, A. R. (2004). The effects of early maternal deprivation on auditory information processing in adult Wistar rats. Biol. Psychiatry 55, 701–707.
Ellenbroek, B. A., and Riva, M. A. (2003). Early maternal deprivation as an animal model for schizophrenia. Clin. Neurosci. 3, 297–302.
Evans, A. C., and Brain Development Cooperative Group. (2006). The NIH MRI study of normal brain development. Neuroimage 1, 184–202.
Fatemi, S. H., and Folsom, T. D. (2009). The neurodevelopmental hypothesis of schizophrenia, revisited. Schizophr. Bull. 35, 528–548.
Fattore, L., Altea, S., and Fratta, W. (2008). Sex differences in drug addiction: a review of animal and human studies. Womens Health (Lond. Engl.) 4, 51–65.
Fattore, L., and Fratta, W. (2010). How important are sex differences in cannabinoid action? Br. J. Pharmacol. 160, 544–548.
Fattore, L., Spano, M. S., Altea, S., Angius, F., Fadda, P., and Fratta, W. (2007). Cannabinoid self-administration in rats: sex differences and the influence of ovarian function. Br. J. Pharmacol. 152, 795–804.
Fernández-Espejo, E., Viveros, M. P., Nunez, L., Ellenbroek, B. A., and Rodriguez de, F. F. (2009). Role of Cannabis and endocannabinoids in the genesis of schizophrenia. Psychopharmacology (Berl.) 206, 531–549.
Fink, G., Sumner, B. E., Rosie, R., Grace, O., and Quinn, J. P. (1996). Estrogen control of central neurotransmission: effect on mood, mental state, and memory. Cell. Mol. Neurobiol. 16, 325–344.
Flaum, M., O’Leary, D. S., Swayze, V. W. II, Miller, D. D., Arndt, S., and Andreasen, N. C. (1995). Symptom dimensions and brain morphology in schizophrenia and related psychotic disorders. J. Psychiatr. Res. 29, 261–276.
Fox, H. C., and Sinha, R. (2009). Sex differences in drug-related stress-system changes: implications for treatment in substance-abusing women. Harv. Rev. Psychiatry 17, 103–119.
Frazier, J. A., Hodge, S. M., Breeze, J. L., Giuliano, A. J., Terry, J. E., Moore, C. M., Kennedy, D. N., Lopez-Larson, M. P., Caviness, V. S., Seidman, L. J., Zablotsky, B., and Makris, N. (2008). Diagnostic and sex effects on limbic volumes in early-onset bipolar disorder and schizophrenia. Schizophr. Bull. 34, 37–46.
Freidenberg, B., Gusmano, R., Hickling, E., Blanchard, E., Bremner, E., and Frye, C. (2010). Women with PTSD have lower basal salivary levels later in the day than do men with PTSD: a preliminary study. Physiol. Behav. 99, 234–236.
Gartlehner, G., Thieda, P., Hansen, R. A., Gaynes, B. N., Deveaugh-Geiss, A., Krebs, E. E., and Lohr, K. N. (2008). Comparative risk for harms of second-generation antidepressants: a systematic review and meta-analysis. Drug Saf. 31, 851–865.
Gatewood, J. D., Wills, A., Shetty, S., Xu, J., Arnold, A. P., Burgoyne, P. S., and Rissman, E. F. (2006). Sex chromosome complement and gonadal sex influence aggressive and parental behaviors in mice. J. Neurosci. 26, 2335–2342.
Gattaz, W. F., Vogel, P., Riecher-Rossler, A., and Soddu, G. (1994). Influence of the menstrual cycle phase on the therapeutic response in schizophrenia. Biol. Psychiatry 36, 137–139.
Gillies, G. E., and McArthur, S. (2010). Estrogen actions in the brain and the basis for differential action in men and women: a case for sex-specific medicines. Pharmacol. Rev. 62, 155–198.
Gilmore, J. H., Lin, W., Prastawa, M. W., Looney, C. B., Vetsa, Y. S., Knickmeyer, R. C., Evans, D. D., Smith, J. K., Hamer, R. M., Lieberman, J. A., and Gerig, G. (2007). Regional gray matter growth, sexual dimorphism, and cerebral asymmetry in the neonatal brain. J. Neurosci. 27, 1255–1260.
Goldstein, J. M., Seidman, L. J., Horton, N. J., Makris, N., Kennedy, D. N., Caviness, V. S. Jr., Faraone, S. V., and Tsuang, M. T. (2001). Normal sexual dimorphism of the adult human brain assessed by in vivo magnetic resonance imaging. Cereb. Cortex 11, 490–497.
Goldstein, J. M., Seidman, L. J., O’Brien, L. M., Horton, N. J., Kennedy, D. N., Makris, N., Caviness, V. S. Jr., Faraone, S. V., and Tsuang, M. T. (2002). Impact of normal sexual dimorphisms on sex differences in structural brain abnormalities in schizophrenia assessed by magnetic resonance imaging. Arch. Gen. Psychiatry 59, 154–164.
Green, M. F. (2006). Cognitive impairment and functional outcome in schizophrenia and bipolar disorder. J. Clin. Psychiatry 67, e12.
Griffin, M. L., Weiss, R. D., Mirin, S. M., and Lange, U. (1989). A comparison of male and female cocaine abusers. Arch. Gen. Psychiatry 46, 122–126.
Grigoriadis, S., and Robinson, G. E. (2007). Gender issues in depression. Ann. Clin. Psychiatry 19, 247–255.
Grigoriadis, S., and Seeman, M. V. (2002). The role of estrogen in schizophrenia: implications for schizophrenia practice guidelines for women. Can. J. Psychiatry 47, 437–442.
Gur, R. C., Gunning-Dixon, F. M., Turetsky, B. I., Bilker, W. B., and Gur, R. E. (2002). Brain region and sex differences in age association with brain volume: a quantitative MRI study of healthy young adults. Am. J. Geriatr. Psychiatry 10, 72–80.
Gur, R. E., Cowell, P. E., Latshaw, A., Turetsky, B. I., Grossman, R. I., Arnold, S. E., Bilker, W. B., and Gur, R. C. (2000). Reduced dorsal and orbital prefrontal gray matter volumes in schizophrenia. Arch. Gen. Psychiatry 57, 761–768.
Gur, R. E., Kohler, C., Turetsky, B. I., Siegel, S. J., Kanes, S. J., Bilker, W. B., Brennan, A. R., and Gur, R. C. (2004). A sexually dimorphic ratio of orbitofrontal to amygdala volume is altered in schizophrenia. Biol. Psychiatry 55, 512–517.
Hafner, H., an der Heiden, W., Behrens, S., Gattaz, W. F., Hambrecht, M., Loffler, W., Maurer, K., Munk-Jorgensen, P., Nowotny, B., Riecher-Rossler, A., and Stein, A. (1998). Causes and consequences of the gender difference in age at onset of schizophrenia. Schizophr. Bull. 24, 99–113.
Hafner, H., Hambrecht, M., Loffler, W., Munk-Jorgensen, P., and Riecher-Rossler, A. (1998a). Is schizophrenia a disorder of all ages? A comparison of first episodes and early course across the life-cycle. Psychol. Med. 28, 351–365.
Hagnell, O., and Rorsman, B. (1979). Suicide in the Lundby study: a comparative investigation of clinical aspects. Neuropsychobiology 5, 61–73.
Hallonquist, J. D., Seeman, M. V., Lang, M., and Rector, N. A. (1993). Variation in symptom severity over the menstrual cycle of schizophrenics. Biol. Psychiatry 33, 207–209.
Han, T. M., and de Vries, G. J. (2003). Organizational effects of testosterone, estradiol, and dihydrotestosterone on vasopressin mRNA expression in the bed nucleus of the stria terminalis. J. Neurobiol. 54, 502–510.
Han, X., Wu, X., Chung, W. Y., Li, T., Nekrutenko, A., Altman, N. S., Chen, G., and Ma, H. (2009). Transcriptome of embryonic and neonatal mouse cortex by high-throughput RNA sequencing. Proc. Natl. Acad. Sci. U.S.A. 106, 12741–12746.
Haney, M. (2007). Opioid antagonism of cannabinoid effects: differences between marijuana smokers and nonmarijuana smokers. Neuropsychopharmacology 32, 1391–1403.
Hankin, B. L., Mermelstein, R., and Roesch, L. (2007). Sex differences in adolescent depression: stress exposure and reactivity models. Child Dev. 78, 279–295.
Hasin, D. S., Keyes, K. M., Alderson, D., Wang, S., Aharonovich, E., and Grant, B. F. (2008). Cannabis withdrawal in the United States: results from NESARC. J. Clin. Psychiatry 69, 1354–1363.
Hastings, R. S., Parsey, R. V., Oquendo, M. A., Arango, V., and Mann, J. J. (2004). Volumetric analysis of the prefrontal cortex, amygdala, and hippocampus in major depression. Neuropsychopharmacology 29, 952–959.
Hawk, L., Dougall, A., Ursano, R., and Baum, A. (2000). Urinary catecholamines and recent onset posttraumatic stress disorder after motor vehicle accidents. Psychosom. Med. 62, 423–434.
Helzer, J. E., Robins, L. N., and McEvoy, L. (1987). Post-traumatic stress disorder in the general population: findings of the epidemiologic catchment area survey. N. Engl. J. Med. 317, 1630–1634.
Hendrick, V., Altshuler, L. L., and Burt, V. K. (1996). Course of psychiatric disorders across the menstrual cycle. Harv. Rev. Psychiatry 4, 200–207.
Herve, P. Y., Cox, E. F., Lotfipour, A. K., Mougin, O. E., Bowtell, R. W., Gowland, P. A., and Paus, T. (2011). Structural properties of the corticospinal tract in the human brain: a magnetic resonance imaging study at 7 Tesla. Brain Struct. Funct. 216, 255–262.
Herve, P. Y., Leonard, G., Perron, M., Pike, G. B., Pitiot, A., Richer, L., Veillette, S., Pausova, Z., and Paus, T. (2009). Handedness, motor skills and maturation of the corticospinal tract in the adolescent brain. Hum. Brain Mapp. 30, 3151–3162.
Higuera-Matas, A., Soto-Montenegro, M. L., del, O. N., Miguens, M., Torres, I., Vaquero, J. J., Sanchez, J., Garcia-Lecumberri, C., Desco, M., and Ambrosio, E. (2008). Augmented acquisition of cocaine self-administration and altered brain glucose metabolism in adult female but not male rats exposed to a cannabinoid agonist during adolescence. Neuropsychopharmacology 33, 806–813.
Hirschfeld, R. M., Klerman, G. L., Clayton, P. J., Keller, M. B., and Andreasen, N. C. (1984). Personality and gender-related differences in depression. J. Affect. Disord. 7, 211–221.
Hobfoll, S. E., Ritter, C., Lavin, J., Hulsizer, M. R., and Cameron, R. P. (1995). Depression prevalence and incidence among inner-city pregnant and postpartum women. J. Consult. Clin. Psychol. 63, 445–453.
Holbrook, T. L., Hoyt, D. B., Stein, M. B., and Sieber, W. J. (2002). Gender differences in long-term posttraumatic stress disorder outcomes after major trauma: women are at higher risk of adverse outcomes than men. J. Trauma. 53, 882–888.
Holtzman, S. G. (1990). “Discriminative stimulus effects of drugs: relationship to potential for abuse,” in Testing and Evaluation of Drugs of Abuse. Modern Methods in Pharmacology, Vol. 6, eds M. W. Adler and A. Cowan (New York: Wiley-Liss), 197–210.
Irle, E., Lange, C., Ruhleder, M., Exner, C., Siemerkus, J., and Weniger, G. (2011). Hippocampal size in women but not men with schizophrenia relates to disorder duration. Psychiatry Res. 3, 133–139.
Iwasaki-Sekin, A., ManoOtagiri, A., Ohata, H., Yamauchi, N., and Shibasaki, T. (2009). Gender differences in corticotrophin and corticosterone secretion and corticotrophin-releasing factor mRNA expression in the paraventricular nucleus of the hypothalamus and the central nucleus of the amygdale in response to footshock stress or psychological stress in rats. Psychoendocrinology 34, 226–237.
Jablenksy, A. (1992). Schizophrenia: manifestations, incidence and course in different cultures. A World Health Organization ten-country study. Psychol. Med. Monogr. Suppl. 20, 1–97.
Jankowski, M. P., Cornuet, P. K., McIlwrath, S., Koerber, H. R., and Albers, K. M. (2006). SRY-box containing gene 11 (Sox11) transcription factor is required for neuron survival and neurite growth. Neuroscience 143, 501–514.
Jiménez, J., Mancini-Marïe, A., Corcoran, C., Fahim, C., Karama, S., Beauregard, M., Lévesque, J., Mensour, M., Stip, E., and Mendrek, A. (2008). Laterality in grey matter densities of the frontal lobe in men and women with schizophrenia: a voxel-based morphometry study. Schizophr. Res. 98, 38–39.
Jiménez, J., Mancini-Marie, A., and Mendrek, A. (2009). The case for not combining men and women in neurocognitive studies for schizophrenia. Schizophr. Res. 108, 293–294.
Jiménez, J. A., Mancini-Marïe, A., Lakis, N., Rinaldi, M., and Mendrek, A. (2010). Disturbed sexual dimorphism of brain activation during mental rotation in schizophrenia. Schizophr. Res. 122, 53–62.
Kalaydjian, A., Swendsen, J., Chiu, W. T., Dierker, L., Degenhardt, L., Glantz, M., Merikangas, K. R., Sampson, N., and Kessler, R. (2009). Sociodemographic predictors of transitions across stages of alcohol use, disorders, and remission in the National Comorbidity Survey Replication. Compr. Psychiatry 50, 299–306.
Kandel, D. B., Yamaguchi, K., and Klein, L. C. (2006). Testing the gateway hypothesis. Addiction 101, 470–472.
Katz, J. L., and Higgins, S. T. (2003). The validity of the reinstatement model of craving and relapse to drug use. Psychopharmacology (Berl.) 168, 21–30.
Kay, S. R., Fiszbein, A., Lindenmayer, J. P., and Opler, L. A. (1986). Positive and negative syndromes in schizophrenia as a function of chronicity. Acta Psychiatr. Scand. 74, 507–518.
Kendell, R. E., Chalmers, J. C., and Platz, C. (1987). Epidemiology of puerperal psychoses. Br. J. Psychiatry 150, 662–673.
Kendler, K. S., and Diehl, S. R. (1993). The genetics of schizophrenia: a current, genetic-epidemiologic perspective. Schizophr. Bull. 19, 261–285.
Keshavan, M. S., Nasrallah, H. A., and Tandon, R. (2011). Schizophrenia, “just the facts” 6. Moving ahead with the schizophrenia concept: from the elephant to the mouse. Schizophr. Res. 127, 3–13.
Keshavan, M. S., Tandon, R., Boutros, N. N., and Nasrallah, H. A. (2008). Schizophrenia, just the facts: what we know in 2008 Part 3: neurobiology. Schizophr. Res. 106, 89–107.
Kessler, R. C., Amminger, G. P., Guilar-Gaxiola, S., Alonso, J., Lee, S., and Ustun, T. B. (2007). Age of onset of mental disorders: a review of recent literature. Curr. Opin. Psychiatry 20, 359–364.
Kessler, R. C., Berglund, P., Delmer, O., Jin, R., Merikangas, K. R., and Walters, E. E. (2005). Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch. Gen. Psychiatry 6, 593–602.
Kim, D. K., Han, S. B., Hong, S. T., Choi, Y. J., Sun, W., Geum, D., and Kim, H. (2008). Expression of Sox11 and Brn transcription factors during development and following transient forebrain ischemia in the rat. Neurosci. Lett. 433, 259–264.
Kirsch, I., Deacon, B. J., Huedo-Medina, T. B., Scoboria, A., Moore, T. J., and Johnson, B. T. (2008). Initial severity and antidepressant benefits: a meta-analysis of data submitted to the Food and Drug Administration. PLoS Med. 5, e45.
Kline, N. S., Blair, J., Cooper, T. B., Esser, A. H., Hackett, E., and Vestergaard, P. (1968). A controlled seven year study of endocrine and other indices in drug treated chronic schizophrenics. Acta Psychiatr. Scand. Suppl. 206, 7–75.
Ko, Y. H., Jung, S. W., Joe, S. H., Lee, C. H., Jung, H. G., Jung, I. K., Kim, S. H., and Lee, M. S. (2007). Association between serum testosterone levels and the severity of negative symptoms in male patients with chronic schizophrenia. Psychoneuroendocrinology 32, 385–391.
Kobayashi, A., Stewart, C. A., Wang, Y., Fujioka, K., Thomas, N. C., Jamin, S. P., and Behringer, R. R. (2011). beta-Catenin is essential for Mullerian duct regression during male sexual differentiation. Development 138, 1967–1975.
Konkle, A. T., and McCarthy, M. M. (2011). Developmental time course of estradiol, testosterone, and dihydrotestosterone levels in discrete regions of male and female rat brain. Endocrinology 152, 223–235.
Kornstein, S. G., Schatzberg, A. F., Thase, M. E., Yonkers, K. A., McCullough, J. P., Keitner, G. I., Gelenberg, A. J., Ryan, C. E., Hess, A. L., Harrison, W., Davis, S. M., and Keller, M. B. (2000). Gender differences in chronic major and double depression. J. Affect. Disord. 60, 1–11.
Kuo, J., Hamid, N., Bondar, G., Dewing, P., Clarkson, J., and Micevych, P. E. (2010). Sex differences in hypothalamic astrocyte response to estradiol stimulation. Biol. Sex Differ. 1, 7.
Lancaster, F. E., Brown, T. D., Coker, K. L., Elliott, J. A., and Wren, S. B. (1996). Sex differences in alcohol preference and drinking patterns emerge during the early postpubertal period. Alcohol. Clin. Exp. Res. 20, 1043–1049.
Lauriello, J., Hoff, A., Wieneke, M. H., Blankfeld, H., Faustman, W. O., Rosenbloom, M., DeMent, S., Sullivan, E. V., Lim, K. O., and Pfefferbaum, A. (1997). Similar extent of brain dysmorphology in severely ill women and men with schizophrenia. Am. J. Psychiatry 154, 819–825.
Laviola, G., Adriani, W., Morley-Fletcher, S., and Terranova, M. L. (2002). Peculiar response of adolescent mice to acute and chronic stress and to amphetamine: evidence of sex differences. Behav. Brain Res. 130, 117–125.
Leibenluft, E. (1999). Gender differences in major depressive disorder and bipolar disorder. CNS Spectr. 4, 25–33.
Leibenluft, E., Ashman, S. B., Feldman-Naim, S., and Yonkers, K. A. (1999). Lack of relationship between menstrual cycle phase and mood in a sample of women with rapid cycling bipolar disorder. Biol. Psychiatry 46, 577–580.
Leranth, C., Petnehazy, O., and MacLusky, N. J. (2003). Gonadal hormones affect spine synaptic density in the CA1 hippocampal subfield of male rats. J. Neurosci. 23, 1588–1592.
Leung, A., and Chue, P. (2000). Sex differences in schizophrenia, a review of the literature. Acta Psychiatr. Scand. Suppl. 401, 3–38.
Ling, K. H., Hewitt, C. A., Beissbarth, T., Hyde, L., Banerjee, K., Cheah, P. S., Cannon, P. Z., Hahn, C. N., Thomas, P. Q., Smyth, G. K., Tan, S. S., Thomas, T., and Scott, H. S. (2009). Molecular networks involved in mouse cerebral corticogenesis and spatio-temporal regulation of Sox4 and Sox11 novel antisense transcripts revealed by transcriptome profiling. Genome Biol. 10, R104.
Llorente, R., Arranz, L., Marco, E. M., Moreno, E., Puerto, M., Guaza, C., De la, F. M., and Viveros, M. P. (2007). Early maternal deprivation and neonatal single administration with a cannabinoid agonist induce long-term sex-dependent psychoimmunoendocrine effects in adolescent rats. Psychoneuroendocrinology 32, 636–650.
Llorente, R., Miguel-Blanco, C., Aisa, B., Lachize, S., Borcel, E., Meijer, O. C., Ramirez, M. J., de Kloet, E. R., and Viveros, M. P. (2011). Long term sex-dependent psychoneuroendocrine effects of maternal deprivation and juvenile unpredictable stress in rats. J. Neuroendocrinol. 23, 329–344.
Llorente-Berzal, A., Fuentes, S., Gagliano, H., Lopez-Gallardo, M., Armario, A., Viveros, M. P., and Nadal, R. (2011). Sex-dependent effects of maternal deprivation and adolescent cannabinoid treatment on adult rat behaviour. Addict. Biol. 16, 624–637.
López-Gallardo, M., Lopez-Rodriguez, A. B., Llorente-Berzal, A., Rotllant, D., Mackie, K., Armario, A., Nadal, R., and Viveros, M. P. (2011). Maternal deprivation and adolescent cannabinoid exposure impact hippocampal astrocytes, CB1 receptors and brain-derived neurotrophic factor in a sexually dimorphic fashion. Neuroscience 204, 90–103.
Lynch, W. J., Nicholson, K. L., Dance, M. E., Morgan, R. W., and Foley, P. L. (2010). Animal models of substance abuse and addiction: implications for science, animal welfare, and society. Comp. Med. 60, 177–188.
Lynch, W. J., Roth, M. E., and Carroll, M. E. (2002). Biological basis of sex differences in drug abuse: preclinical and clinical studies. Psychopharmacology (Berl.) 164, 121–137.
Maatouk, D. M., and Capel, B. (2008). Sexual development of the soma in the mouse. Curr. Top. Dev. Biol. 83, 151–183.
Malone, D. T., Hill, M. N., and Rubino, T. (2010). Adolescent Cannabis use and psychosis: epidemiology and neurodevelopmental models. Br. J. Pharmacol. 160, 511–522.
Mancini-Marïe, A., Corcoran, C., Jimenez, J., Fahim, C., Karama, S., Beauregard, M., Lévesque, J., Mensour, B., Stip, E., and Mendrek, A. (2008a). Differences in grey matter density in frontal lobe regions in men and women with schizophrenia. Schizophr. Res. 98, 39–40.
Mancini-Marïe, A., Jimenez, J., Corcoran, C., Stip, E., Melissa, R., and Mendrek, A. (2008b). Reversed sexual dimorphism: parietal cortex grey matter density in women and men with schizophrenia compared to matched healthy controls using 3 tesla MRI. Schizophr. Res. 102, 71.
Manna, P. R., Pakarinen, P., El-Hefnawy, T., and Huhtaniemi, I. T. (1999). Functional assessment of the calcium messenger system in cultured mouse Leydig tumor cells: regulation of human chorionic gonadotropin-induced expression of the steroidogenic acute regulatory protein. Endocrinology 140, 1739–1751.
Marco, E. M., Adriani, W., Canese, R., Podo, F., Viveros, M. P., and Laviola, G. (2007). Enhancement of endocannabinoid signalling during adolescence: modulation of impulsivity and long-term consequences on metabolic brain parameters in early maternally deprived rats. Pharmacol. Biochem. Behav. 86, 334–345.
Mateos, B., Borcel, E., Loriga, R., Luesu, W., Bini, V., Llorente, R., Castelli, M., and Viveros, M. P. (2010). Adolescent exposure to nicotine and/or the cannabinoid agonist CP 55,940 induces gender-dependent long-lasting memory impairments and changes in brain nicotinic and CB1 cannabinoid receptors. J. Psychopharmacol. 12, 1676–1690.
Mayer, A., Mosler, G., Just, W., Pilgrim, C., and Reisert, I. (2000). Developmental profile of Sry transcripts in mouse brain. Neurogenetics 3, 25–30.
Maynard, T. M., Sikich, L., Lieberman, J. A., and LaMantia, A. S. (2001). Neural development, cell-cell signaling, and the “two-hit” hypothesis of schizophrenia. Schizophr. Bull. 27, 457–476.
Mazor, A., Matar, M. A., Kaplan, Z., Kozlovsky, N., Zohar, J., and Cohen, H. (2009). Gender related qualitative differences in baseline and post stress anxiety responses are not reflected in the incidence of criterion based PTSD like behaviour patterns. World J. Biol. Psychiatry 10, 856–869.
McCarthy, M. M., and Arnold, A. P. (2011). Reframing sexual differentiation of the brain. Nat. Neurosci. 14, 677–683.
McFarlane, A. C., Barton, C. A., Yehuda, R., and Wittert, G. (2011). Cortisol response to acute trauma and risk of posttraumatic stress disorder. Psychoneuroendocrinology 36, 720–727.
McGrath, J. J. (2007). The surprisingly rich contours of schizophrenia epidemiology. Arch. Gen. Psychiatry 64, 14–16.
McGrath, J. J., and Susser, E. S. (2009). New directions in the epidemiology of schizophrenia. Med. J. Aust. 190(Suppl. 4), S7–S9.
McGuffin, P., Owen, M. J., and Farmer, A. E. (1995). Genetic basis of schizophrenia. Lancet 346, 678–682.
Mendelson, J. H., Weiss, R., Griffin, M., Mirin, S. M., Teoh, S. K., Mello, N. K., and Lex, B. W. (1991). Some special considerations for treatment of drug abuse and dependence in women. NIDA Res. Monogr. 106, 313–327.
Mendrek, A. (2007). Reversal of normal cerebral sexual dimorphism in schizophrenia: evidence and speculations. Med. Hypotheses 69, 896–902.
Mendrek, A., Lakis, N., and Jimenez, J. (2011). Associations of sex steroid hormones with cerebral activations during mental rotation in men and women with schizophrenia. Psychoneuroendocrinology 36, 1422–1426.
Middaugh, L. D., Kelley, B. M., Bandy, A. L., and McGroarty, K. K. (1999). Ethanol consumption by C57BL/6 mice: influence of gender and procedural variables. Alcohol 17, 175–183.
Miyagawa, S., Satoh, Y., Haraguchi, R., Suzuki, K., Iguchi, T., Taketo, M. M., Nakagata, N., Matsumoto, T., Takeyama, K., Kato, S., and Yamada, G. (2009). Genetic interactions of the androgen and Wnt/beta-catenin pathways for the masculinization of external genitalia. Mol. Endocrinol. 23, 871–880.
Moffat, S. D. (2005). Effects of testosterone on cognitive and brain aging in elderly men. Ann. N. Y. Acad. Sci. 1055, 80–92.
Morgan, C. P., and Bale, T. L. (2011). Early prenatal stress epigenetically programs dysmasculinization in second-generation offspring via the paternal lineage. J. Neurosci. 31, 11748–11755.
Mortensen, P. B., Pedersen, C. B., Westergaard, T., Wohlfahrt, J., Ewald, H., Mors, O., Andersen, P. K., and Melbye, M. (1999). Effects of family history and place and season of birth on the risk of schizophrenia. N. Engl. J. Med. 340, 603–608.
Mueller, B. R., and Bale, T. L. (2008). Sex-specific programming of offspring emotionality after stress early in pregnancy. J. Neurosci. 28, 9055–9065.
Mura, A., Gadau, S., Lepore, G., Balzano, F., Zedda, M., Mura, E., and Farina, V. (2010). Expression and distribution of P450-aromatase in the ovine hypothalamus at different stages of fetal development. Neuro Endocrinol. Lett. 31, 690–699.
Naftolin, F., Leranth, C., Horvath, T. L., and Garcia-Segura, L. M. (1996). Potential neuronal mechanisms of estrogen actions in synaptogenesis and synaptic plasticity. Cell. Mol. Neurobiol. 16, 213–223.
Najavits, L. M., and Lester, K. M. (2008). Gender differences in cocaine dependence. Drug Alcohol Depend. 97, 190–194.
National Comorbidity Survey. (2005). NCS-R appendix tables: Table 1. Lifetime Prevalence of DSM-IV/WMH-CIDI Disorders by Sex and Cohort. Table 2. Twelve-Month Prevalence of DSM-IV/WMH-CIDI Disorders by Sex and Cohort. Available at: http://www.hcp.med.harvard.edu/ncs/publications.php
Nestler, E. J., and Hyman, S. E. (2010). Animal models of neuropsychiatric disorders. Nat. Neurosci. 13, 1161–1169.
Neylan, T. C., Sun, B., Rempel, H., Ross, J., Lenoci, M., O’Donovan, A., and Pulliam, L. (2011). Suppressed monocyte gene expression profile in men versus women with PTSD. Brain Behav. Immun. 25, 524–531.
O’Donovan, A., Sun, B., Cole, S., Rempel, H., Lenoci, M., Pulliam, L., and Neylan, T. (2011). Transcriptional control of monocyte gene expression in post-traumatic stress disorder. Dis. Markers 30, 123–132.
O’Hara, M. W., and Swain, A. M. (1996). Rates and risk of postpartum depression – a meta-analysis. Int. Rev. Psychiatry 8, 37–54.
Pask, A. J., Harry, J. L., Graves, J. A., O’Neill, R. J., Layfield, S. L., Shaw, G., and Renfree, M. B. (2002). SOX9 has both conserved and novel roles in marsupial sexual differentiation. Genesis 33, 131–139.
Paus, T. (2010a). “Sex differences in the human brain: a developmental perspective,” in Progress in Brain Research, Vol. 186, ed. I. Savic (Stockholm: Academic Press), 13–28.
Paus, T. (2010b). Growth of white matter in the adolescent brain: myelin or axon. Brain Cogn. 72, 26–35.
Paus, T., Bernard, M., Chakravarty, M. M., Davey Smith, G., Gillis, J., Lourdusamy, A., Leonard, G., Melka, M. G., Pavlidis, P., Perron, M., Pike, G. B., Richer, L., Schumann, G., Timpson, N., Toro, R., Veillette, S., and Pausova, Z. (2011). KCTD8 gene and brain growth in adverse intrauterine environment: a genome-wide association study. Cereb. Cortex.
Paus, T., Keshavan, M., and Giedd, J. N. (2008a). Why do many psychiatric disorders emerge during adolescence? Nat. Rev. Neurosci. 9, 947–957.
Paus, T., Nawazkhan, I., Leonard, G., Perron, M., Pike, G. B., Pitiot, A., Richer, L., Veillette, S., and Pausova, Z. (2008b). Corpus callosum in adolescent offspring exposed prenatally to maternal cigarette smoking. Neuroimage 40, 435–441.
Paus, T., Nawaz-Khan, I., Leonard, G., Perron, M., Pike, G. B., Pitiot, A., Richer, L., Susman, E., Veillette, S., and Pausova, Z. (2010). Sexual dimorphism in the adolescent brain: role of testosterone and androgen receptor in global and local volumes of grey and white matter. Horm. Behav. 57, 63–75.
Paus, T., Otaky, N., Caramanos, Z., MacDonald, D., Zijdenbos, A., D’Avirro, D., Gutmans, D., Holmes, C., Tomaiuolo, F., and Evans, A. C. (1996). In vivo morphometry of the intrasulcal gray matter in the human cingulate, paracingulate, and superior-rostral sulci: hemispheric asymmetries, gender differences and probability maps. J. Comp. Neurol. 376, 664–673.
Paus, T., and Toro, R. (2009). Could sex differences in white matter be explained by g ratio? Front. Neuroanat. 3:14.
Pausova, Z., Paus, T., Abrahamowicz, M., Almerigi, J., Arbour, N., Bernard, M., Gaudet, D., Hanzalek, P., Hamet, P., Evans, A. C., Kramer, M., Laberge, L., Leal, S., Leonard, G., Lerner, J., Lerner, R. M., Mathieu, J., Perron, M., Pike, B., Pitiot, A., Richer, L., Séguin, J. R., Syme, C., Toro, R., Tremblay, R. E., Veillette, S., and Watkins, K. (2007). Genes, maternal smoking and the offspring brain and body during adolescence: design of the saguenay youth study. Hum. Brain Mapp. 28, 502–518.
Peper, J. S., Brouwer, R. M., Van Baal, G. C., Schnack, H. G., van, L. M., Boomsma, D. I., Kahn, R. S., and Hulshoff Pol, H. E. (2009). Does having a twin brother make for a bigger brain? Eur. J. Endocrinol. 160, 739–746.
Perrin, J. S., Herve, P. Y., Leonard, G., Perron, M., Pike, G. B., Pitiot, A., Richer, L., Veillette, S., Pausova, Z., and Paus, T. (2008). Growth of white matter in the adolescent brain: role of testosterone and androgen receptor. J. Neurosci. 28, 9519–9524.
Peruffo, A., Giacomello, M., Montelli, S., Corain, L., and Cozzi, B. (2011). Expression and localization of aromatase P450arom, estrogen receptor-α, and estrogen receptor-β in the developing bovine frontal cortex. Gen. Comp. Endocrinol. 172, 211–217.
Pickens, R., Meisch, R. A., and Thompson, T. (1978). “Drug self-administration: an analysis of the reinforcing effects of drugs,” in Handbook of Psychopharmacology, Vol. 12, eds L. L. Iversen, S. P. Iversen and S. H. Snyder (New York: Plenum Press), 1–37.
Pignatelli, D., Xiao, F., Gouveia, A. M., Ferreira, J. G., and Vinson, G. P. (2006). Adrenarche in the rat. J. Endocrinol. 191, 301–308.
Pompolo, S., and Harley, V. R. (2001). Localisation of the SRY-related HMG box protein, SOX9, in rodent brain. Brain Res. 906, 143–148.
Powell, S. B. (2010). Models of neurodevelopmental abnormalities in schizophrenia. Curr. Top. Behav. Neurosci. 4, 435–481.
Pratchett, L. C., Pelcovitz, M. R., and Yehuda, R. (2010). Trauma and violence: are women the weaker sex? Psychiatr. Clin. North Am. 33, 465–474.
Randall, C. L., Roberts, J. S., Del Boca, F. K., Carroll, K. M., Connors, G. J., and Mattson, M. E. (1999). Telescoping of landmark events associated with drinking: a gender comparison. J. Stud. Alcohol 60, 252–260.
Rehn, A. E., and Rees, S. M. (2005). Investigating the neurodevelopmental hypothesis of schizophrenia. Clin. Exp. Pharmacol. Physiol. 32, 687–696.
Reite, M., Sheeder, J., Teale, P., Adams, M., Richardson, D., Simon, J., Jones, R. H., and Rojas, D. C. (1997). Magnetic source imaging evidence of sex differences in cerebral lateralization in schizophrenia. Arch. Gen. Psychiatry 54, 433–440.
Resko, J. A., and Roselli, C. E. (1997). Prenatal hormones organize sex differences of the neuroendocrine reproductive system: observations on guinea pigs and nonhuman primates. Cell. Mol. Neurobiol. 17, 627–648.
Ressler, K. J., Mercer, K. B., Bradley, B., Jovanovic, T., Mahan, A., Kerley, K., Norrholm, S. D., Kilaru, V., Smith, A. K., Myers, A. J., Ramirez, M., Engel, A., Hammack, S. E., Toufexis, D., Braas, K. M., Binder, E. B., and May, V. (2011). Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 470, 492–497.
Ring, N., Tantam, D., Montague, L., Newby, D., Black, D., and Morris, J. (1991). Gender differences in the incidence of definite schizophrenia and atypical psychosis – focus on negative symptoms of schizophrenia. Acta Psychiatr. Scand. 84, 489–496.
Roberts, D. C., Morgan, D., and Liu, Y. (2007). How to make a rat addicted to cocaine. Prog. Neuropsychopharmacol. Biol. Psychiatry 31, 1614–1624.
Roselli, C. E., Estill, C. T., Stadelman, H. L., Meaker, M., and Stormshak, F. (2011). Separate critical periods exist for testosterone-induced differentiation of the brain and genitals in sheep. Endocrinology 152, 2409–2415.
Rubino, T., and Parolaro, D. (2011). Sexually dimorphic effects of cannabinoid compounds on emotion and cognition. Front Behav. Neurosci. 5:64.
Rubino, T., Vigano, D., Realini, N., Guidali, C., Braida, D., Capurro, V., Castiglioni, C., Cherubino, F., Romualdi, P., Candeletti, S., Sala, M., and Parolaro, D. (2008). Chronic delta 9-tetrahydrocannabinol during adolescence provokes sex-dependent changes in the emotional profile in adult rats: behavioral and biochemical correlates. Neuropsychopharmacology 33, 2760–2771.
Rund, B. R., and Borg, N. E. (1999). Cognitive deficits and cognitive training in schizophrenic patients: a review. Acta Psychiatr. Scand. 100, 85–95.
Saha, S., Chant, D., and McGrath, J. (2007). A systematic review of mortality in schizophrenia: is the differential mortality gap worsening over time? Arch. Gen. Psychiatry 64, 1123–1131.
Saha, S., Chant, D., Welham, J., and McGrath, J. (2005). A systematic review of the prevalence of schizophrenia. PLoS Med. 2, e141.
Sanches, M., Keshavan, M. S., Brambilla, P., and Soares, J. C. (2008). Neurodevelopmental basis of bipolar disorder: a critical appraisal. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 1617–1627.
Schmidt, M. V. (2011). Animal models for depression and the mismatch hypothesis of disease. Psychoneuroendocrinology 36, 330–338.
Schultz, S. K., Miller, D. D., Oliver, S. E., Arndt, S., Flaum, M., and Andreasen, N. C. (1997). The life course of schizophrenia: age and symptom dimensions. Schizophr. Res. 23, 15–23.
Schulz, K. M., Molenda-Figueira, H. A., and Sisk, C. L. (2009). Back to the future: the organizational-activational hypothesis adapted to puberty and adolescence. Horm. Behav. 55, 597–604.
Schumann, G., Loth, E., Banaschewski, T., Barbot, A., Barker, G., Büchel, C., Conrod, P. J., Dalley, J. W., Flor, H., Gallinat, J., Garavan, H., Heinz, A., Itterman, B., Lathrop, M., Mallik, C., Mann, K., Martinot, J.-L., Paus, T., Poline, J.-B., Robbins, T. W., Rietschel, M., Reed, L., Smolka, M., Spanagel, R., Speiser, C., Stephens, D. N., Ströhle, A., Struve, M., and IMAGEN Consortium. (2010). The IMAGEN study: reinforcement-related behaviour in normal brain function and psychopathology. Mol. Psychiatry 15, 1128–1139.
Scotting, P. J., Kordes, U., and Cheng, Y.-C. (2005). Sox group E gene expression distinguishes different types and maturation stages of glial cells in developing chick and mouse. Brain Res. Dev. Brain Res. 157, 209–213.
Seeman, M. V. (1997). Psychopathology in women and men: focus on female hormones. Am. J. Psychiatry 154, 1641–1647.
Shen, J. H., and Ingraham, H. A. (2002). Regulation of the orphan nuclear receptor steroidogenic factor 1 by Sox proteins. Mol. Endocrinol. 16, 529–540.
Shirayama, Y., Hashimoto, K., Suzuki, Y., and Higuchi, T. (2002). Correlation of plasma neurosteroid levels to the severity of negative symptoms in male patients with schizophrenia. Schizophr. Res. 58, 69–74.
Shtasel, D. L., Gur, R. E., Gallacher, F., Heimberg, C., and Gur, R. C. (1992). Gender differences in the clinical expression of schizophrenia. Schizophr. Res. 7, 225–231.
Sinha, R. (2001). How does stress increase risk of drug abuse and relapse? Psychopharmacology (Berl.) 158, 343–359.
Sisk, C. L., and Foster, D. L. (2004). The neural basis of puberty and adolescence. Nat. Neurosci. 7, 1040–1047.
Smith, D. J., Kyle, S., Forty, L., Cooper, C., Walters, J., Russell, E., Caesar, S., Farmer, A., McGuffin, P., Jones, I., Jones, L., and Craddock, N. (2008). Differences in depressive symptom profile between males and females. J. Affect. Disord. 108, 279–284.
Sprock, J., and Yoder, C. Y. (1997). Women and depression: an update on the report of the APA task force. Sex Roles 36, 269–303.
Suarez, J., Llorente, R., Romero-Zerbo, S. Y., Mateos, B., Bermudez-Silva, F. J., de Fonseca, F. R., and Viveros, M. P. (2009). Early maternal deprivation induces gender-dependent changes on the expression of hippocampal CB(1) and CB(2) cannabinoid receptors of neonatal rats. Hippocampus 19, 623–632.
Suarez, J., Rivera, P., Llorente, R., Romero-Zerbo, S. Y., Bermudez-Silva, F. J., de Fonseca, F. R., and Viveros, M. P. (2010). Early maternal deprivation induces changes on the expression of 2-AG biosynthesis and degradation enzymes in neonatal rat hippocampus. Brain Res. 1349, 162–173.
Sullivan, P. F., Neale, M. C., and Kendler, K. S. (2000). Genetic Epidemiology of major depression: review and meta-analysis. Am. J. Psychiatry 157, 1552–1562.
Szulc, A., Galinska, B., and Czernikiewicz, A. (2005). Schizophrenia – neurodevelopmental and neurodegenerative theory revisited. Psychiatr. Pol. 39, 249–258.
Szymanski, S., Lieberman, J. A., Alvir, J. M., Mayerhoff, D., Loebel, A., Geisler, S., Chakos, M., Koreen, A., Jody, D., Kane, J., et al.. (1995). Gender differences in onset of illness, treatment response, course, and biologic indexes in first-episode schizophrenic patients. Am. J. Psychiatry 152, 698–703.
Takahashi, T., Suzuki, M., Kawasaki, Y., Hagino, H., Yamashita, I., Nohara, S., Nakamura, K., Seto, H., and Kurachi, M. (2003). Perigenual cingulate gyrus volume in patients with schizophrenia: a magnetic resonance imaging study. Biol. Psychiatry 53, 593–600.
Takahashi, T., Suzuki, M., Zhou, S. Y., Hagino, H., Kawasaki, Y., Yamashita, I., Nohara, S., Nakamura, K., Seto, H., and Kurachi, M. (2004). Lack of normal gender differences of the perigenual cingulate gyrus in schizophrenia spectrum disorders. A magnetic resonance imaging study. Eur. Arch. Psychiatry Clin. Neurosci. 254, 273–280.
Takai, N., Yamaguchi, M., Aragaki, T., Eto, K., Uchihashi, K., and Nishikawa, K. (2007). Gender-specific differences in salivary biomarker responses to acute psychological stress. Ann. N. Y. Acad. Sci. 1098, 510–515.
Takei, N., Van Os, J., and Murray, R. M. (1995). Maternal exposure to influenza and risk of schizophrenia: a 22 year study from The Netherlands. J. Psychiatr. Res. 29, 435–445.
Thornton, J., Zehr, J. L., and Loose, M. D. (2009). Effects of prenatal androgens on rhesus monkeys: a model system to explore the organizational hypothesis in primates. Horm. Behav. 55, 633–645.
Tolin, D. F., and Foa, E. B. (2006). Sex differences in trauma and posttraumatic stress disorder: a quantitative review of 25 years of research. Psychol. Bull. 132, 959–992.
Toro, R., Leonard, G., Lerner, J. V., Lerner, R. M., Perron, M., Pike, G. B., Richer, L., Veillette, S., Pausova, Z., and Paus, T. (2008). Prenatal exposure to maternal cigarette smoking and the adolescent cerebral cortex. Neuropsychopharmacology 33, 1019–1027.
Trivedi, M. H., Rush, A. J., Wisniewski, S. R., Nierenberg, A. A., Warden, D., Ritz, L., Norquist, G., Howland, R. H., Lebowitz, B., McGrath, P. J., Shores-Wilson, K., Biggs, M. M., Balasubramani, G. K., and Fava, M. (2006). Evaluation of outcomes with citalopram for depression using measurement-based care in STAR D: implications for clinical practice. Am. J. Psychiatry 163, 28–40.
Troutman, B. R., and Cutrona, C. E. (1990). Nonpsychotic postpartum depression among adolescent mothers. J. Abnorm. Psychol. 99, 69–78.
Vierhapper, H., Nowotny, P., and Waldhausl, W. (1998). Sex-specific differences in cortisol production rates in humans. Metab. Clin. Exp. 47, 974–976.
Vilain, E., and McCabe, E. R. (1998). Mammalian sex determination: from gonads to brain. Mol. Genet. Metab. 65, 74–84.
Viveros, M. P., Bermúdez-Silva, F. J., López-Rodríguez, A. B., and Wagner, E. J. (2011a). The endocannabinoid system as pharmacological target derived from its CNS role in energy homeostasis and reward. Applications in eating disorders and addiction. Pharmaceuticals 8, 1101–1136.
Viveros, M. P., Llorente, R., Suarez, J., Llorente-Berzal, A., Lopez-Gallardo, M., and Rodriguez de, F. F. (2011b). The endocannabinoid system in critical neurodevelopmental periods: sex differences and neuropsychiatric implications. J. Psychopharmacol. (Oxford) 26, 164–176.
Viveros, M. P., Llorente, R., Lopez-Gallardo, M., Suarez, J., Bermudez-Silva, F., De la Fuente, M., Rodriguez de Fonseca, F., and Garcia-Segura, L. M. (2009). Sex-dependent alterations in response to maternal deprivation in rats. Psychoneuroendocrinology 34(Suppl. 1), S217–S226.
Wagner, C. K., Xu, J., Pfau, J. L., Quadros, P. S., de Vries, G. J., and Arnold, A. P. (2004). Neonatal mice possessing an Sry transgene show a masculinized pattern of progesterone receptor expression in the brain independent of sex chromosome status. Endocrinology 145, 1046–1049.
Weinberg, J., Sliwowska, J. H., Lan, N., and Hellemans, K. G. (2008). Prenatal alcohol exposure: foetal programming, the hypothalamic-pituitary-adrenal axis and sex differences in outcome. J. Neuroendocrinol. 20, 470–488.
Weissman, M. M., and Olfson, M. (1995). Depression in women: implications for health care research. Science 269, 799–801.
Wittmann, W., and McLennan, I. S. (2011). The male bias in the number of Purkinje cells and the size of the murine cerebellum may require Müllerian inhibiting substance/anti-Müllerian hormone. J. Neuroendocrinol. 23, 831–838.
Wu, J. B., Chen, K., Li, Y., Lau, Y.-F. C., and Shih, J. C. (2009). Regulation of monoamine oxidase A by the SRY gene on the Y chromosome. FASEB J. 23, 4029–4038.
Xu, J., Burgoyne, P. S., and Arnold, A. P. (2002). Sex differences in sex chromosome gene expression in mouse brain. Hum. Mol. Genet. 11, 1409–1419.
Yehuda, R. (2002). Current status of cortisol findings in post-traumatic stress disorder. Psychiatr. Clin. North Am. 25, 341–368.
Yehuda, R. (2006). Advances in understanding neuroendocrine alterations in PTSD and their therapeutic implications. Ann. N. Y. Acad. Sci. 1071, 137–166.
Yehuda, R., Flory, J., Pratchett, L., Buxbaum, J., Ising, M., and Holsboer, F. (2010). Putative biological mechanisms for the association between early life adversity and the subsequent development of PTSD. Psychopharmacology (Berl.) 212, 405–417.
Keywords: sexual differentiation, adolescence, schizophrenia, depression, bipolar disorders, posttraumatic disorders, addiction, animal models
Citation: Viveros M-P, Mendrek A, Paus T, López-Rodríguez AB, Marco EM, Yehuda R, Cohen H, Lehrner A and Wagner EJ (2012) A comparative, developmental, and clinical perspective of neurobehavioral sexual dimorphisms. Front. Neurosci. 6:84. doi: 10.3389/fnins.2012.00084
Received: 06 December 2011; Accepted: 18 May 2012;
Published online: 12 June 2012.
Edited by:
Elena Jazin, Uppsala University, SwedenReviewed by:
Jun Xu, Tufts University, USABirger Scholz, Uppsala University, Sweden
Khyobeni Mozhui, University of Tennessee Health Science Center, USA
Copyright: © 2012 Viveros, Mendrek, Paus, López-Rodríguez, Marco, Yehuda, Cohen, Lehrner and Wagner. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Maria-Paz Viveros, Departamento de FisiologÍa (FisiologÍa Animal II), Facultad de BiologÍa, Universidad Complutense, C/José Antonio Nováis no 2, 28040 Madrid, Spain. e-mail:cGF6dml2ZXJAYmlvLnVjbS5lcw==