 Lilian Calderón-Garcidueñas1,2*
Lilian Calderón-Garcidueñas1,2* Janet V. Cross3
Janet V. Cross3 Maricela Franco-Lira2
Maricela Franco-Lira2 Mariana Aragón-Flores2
Mariana Aragón-Flores2 Michael Kavanaugh1
Michael Kavanaugh1 Ricardo Torres-Jardón4
Ricardo Torres-Jardón4 Chih-kai Chao1
Chih-kai Chao1 Charles Thompson1Jing Chang5
Charles Thompson1Jing Chang5 Hongtu Zhu5
Hongtu Zhu5 Amedeo D'Angiulli6
Amedeo D'Angiulli6- 1Department of Biomedical Sciences, The Center for Structural and Functional Neurosciences, The University of Montana, Missoula, MT, USA
- 2Hospital Central Militar, Secretaria de la Defensa Nacional, Mexico City, Mexico
- 3Department of Pathology, School of Medicine, University of Virginia, Charlottesville, VA, USA
- 4Centro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 5Department of Biostatistics, Gillings School of Global Public Health, University of North Carolina, Chapel Hill, NC, USA
- 6Department of Neuroscience, Carleton University, Ottawa, ON, Canada
Mexico City Metropolitan Area children chronically exposed to high concentrations of air pollutants exhibit an early brain imbalance in genes involved in oxidative stress, inflammation, innate and adaptive immune responses along with accumulation of misfolded proteins observed in the early stages of Alzheimer and Parkinson's diseases. A complex modulation of serum cytokines and chemokines influences children's brain structural and gray/white matter volumetric responses to air pollution. The search for biomarkers associating systemic and CNS inflammation to brain growth and cognitive deficits in the short term and neurodegeneration in the long-term is our principal aim. We explored and compared a profile of cytokines, chemokines (Multiplexing LASER Bead Technology) and Cellular prion protein (PrPC) in normal cerebro-spinal-fluid (CSF) of urban children with high vs. low air pollution exposures. PrPC and macrophage inhibitory factor (MIF) were also measured in serum. Samples from 139 children ages 11.91 ± 4.2 years were measured. Highly exposed children exhibited significant increases in CSF MIF (p = 0.002), IL6 (p = 0.006), IL1ra (p = 0.014), IL-2 (p = 0.04), and PrPC (p = 0.039) vs. controls. MIF serum concentrations were higher in exposed children (p = 0.009). Our results suggest CSF as a MIF, IL6, IL1Ra, IL-2, and PrPC compartment that can possibly differentiate air pollution exposures in children. MIF, a key neuro-immune mediator, is a potential biomarker bridge to identify children with CNS inflammation. Fine tuning of immune-to-brain communication is crucial to neural networks appropriate functioning, thus the short and long term effects of systemic inflammation and dysregulated neural immune responses are of deep concern for millions of exposed children. Defining the linkage and the health consequences of the brain / immune system interactions in the developing brain chronically exposed to air pollutants ought to be of pressing importance for public health.
Introduction
Air pollution is a significant health problem in megacities around the world (Molina and Molina, 2004; Chen and Kan, 2008; Bloom, 2011). In the first 50 years of this century (Bloom, 2011), the projected world population will have a further increase of 2–4.5 billion making the issue of deteriorating environments and their health impact critical.
Mexico City Metropolitan Area (MCMA) children with no known risk factors for neurological or cognitive disorders exhibit significant deficits in a combination of fluid and crystallized cognition tasks vs. low air pollution exposed children (Calderón-Garcidueñas et al., 2008a). Brain structural and volumetric changes are seen in both MCMA children and young animal facility dogs (Calderón-Garcidueñas et al., 2008a, 2011a, 2012a). The canine frontal white matter lesions are characterized by vascular sub-cortical pathology associated with neuroinflammation, gliosis, and ultrafine particulate matter deposition (Calderón-Garcidueñas et al., 2008a). Young MCMA residents exhibit the neuropathological hallmarks of Alzheimer and Parkinson's diseases i.e., amyloid beta42 (Aβ42) plaques, tau hyperphosphorylation with pre-tangles and α-synuclein accumulation (Calderón-Garcidueñas et al., 2012b).
The complex modulation of cytokines and chemokines that influence children's central nervous system (CNS) structural and volumetric responses to air pollution obligate the search for biomarkers associating systemic and CNS inflammation to brain growth and cognitive deficits in the short term and neurodegeneration in the long-term (Calderón-Garcidueñas et al., 2008a,b, 2011a,b, 2012b). A good candidate biomarker, macrophage migration inhibitory factor (MIF) is a cytokine expressed in the CNS and involved in innate and adaptive immune responses (Bernhagen et al., 1993; Bacher et al., 2010; Edwards et al., 2010; Moon et al., 2012; Savaskan et al., 2012; Bucala, 2013; Cox et al., 2013; Freiria-Oliveira et al., 2013; Turtzo et al., 2013). MIF participates in the induction of neural stem/progenitor cells (Ohta et al., 2012), the protection of mice female brains after experimental stroke (Turtzo et al., 2013), the mediation of the antidepressant action of exercise (Moon et al., 2012), and of key importance for this work, MIF plays a key role in neuroinflammatory responses (Savaskan et al., 2012; Cox et al., 2013).
In clinical practice and in the experimental setting, MIF has been used as a marker of neurological worsening in progressive neurodegenerative processes including multiple sclerosis and autoimmune-mediated neuroinflammation (Hagman et al., 2011; Cox et al., 2013), as a key neuro-immune mediator linking depressive symptoms with inflammation and HPA dysregulation (Edwards et al., 2010), and as a predictor of lack of response to antidepressants (Cattaneo et al., 2013). Moreover, in the scenario of air pollution and the early hallmarks of Alzheimer's disease (AD) in Mexico City children (Calderón-Garcidueñas et al., 2008a, 2012a), two key issues are of utmost interest to us: the increased concentrations of MIF in the cerebro-spinal-fluid (CSF) of AD patients and the capacity of MIF to enhance the ability of the tau/Aβ42 ratio to discriminate cognitively normal vs. mildly demented patients (Lee et al., 2008; Bacher et al., 2010; Craig-Schapiro et al., 2011). In addition MIF promotes the production of several inflammatory mediators including IL-6, TNF α, and IFN γ (Popp et al., 2009). Given the wide spectrum of serum and CSF responses to pathological processes including neuroinflammation and neurodegenerative diseases such as AD and Parkinson's disease (PD) (Dziedzic, 2006; Helmy et al., 2012; Yan et al., 2012; Giralt et al., 2013; Martinez et al., 2013; Mooijaart et al., 2013; Smolen et al., 2013), we have selected a 41 cytokine/chemokine panel and MIF to explore the CSF responses to high air pollution residency.
Cellular prion protein (PrPC) is a conserved GPI-anchored membrane protein located on the surface of neurons, at both pre and post-synaptic sites, with a high abundance in the hippocampus, frontal cortex and stratium (Stellato et al., 2011; Ding et al., 2013; Watt et al., 2013). The PRNP gene located in chromosome 20 likely plays key roles in neuronal development, synaptic plasticity, myelin sheath maintenance, cell adhesion, CNS development, and neuroprotection via inhibition of the mitochondrial apoptotic pathway (Mitsios et al., 2007; Altmeppen et al., 2012; Bradford and Mabbott, 2012; Kaiser et al., 2012; Ding et al., 2013).
We are particularly interested in the critical role PrPC may play in the integrity of the CNS under a neuroinflammatory insult, precisely the key marker of air pollution exposure in urban children (Bremer et al., 2010; Popko, 2010; Gourdain et al., 2012; Scalabrino and Veber, 2012). The role of PrPC in cellular immunity (Tsutsui et al., 2008), cognitive functioning (Breitling et al., 2012), oxidative stress, and the impact of prion proteins upon the toxic effect of environmental neurotoxic metals (Choi et al., 2007; Oh et al., 2012) are at the core of our research.
The primary aim of this study was to measure the concentrations of PrPC and selected inflammatory mediators in normal CSF from cohorts of children with high exposures to urban air pollution vs. low pollution control children. Concurrently, we also explored serum MIF and PrPC in clinically healthy children representing cohorts with high and low air pollution exposures.
Our results identify CSF as a MIF, IL6, IL-1Ra, IL-2, and PrPC compartment that differentiates children with high vs. low air pollution exposures. The MIF increases in CSF and serum compartments in highly vs. low exposed children strongly suggest MIF could be a serum biomarker bridge to identify children at risk for brain inflammation. Short and long term effects of dysregulated neural immune responses and systemic inflammation are of pressing importance for public health.
Procedure
Study Areas
Children's cohorts were selected from the MCMA and several small cities in Mexico, characterized by clean environments with concentrations of the six criteria air pollutants (ozone, particulate matter, sulfur dioxide, nitrogen oxides, carbon monoxide and lead) below the current US EPA standards.
Participants
This research was approved by the research ethics committee of the Hospital Central Militar in Mexico City. Children gave active assent and their parents gave written informed consent to participation in the study. This work includes data from 139 children 74F, 65M (Meanage = 11.91 years, SD = 4.2), carefully selected to represent comparable populations from a larger longitudinal cohort research program. There were two groups of children included in this study: Group 1 (n: 28, Meanage = 10.46 years, SD = 4.2, low pollution 8F/6M; high pollution 6F/8M) corresponded to children admitted to the hospital from either MCMA or a low polluted city with a work up diagnosis of acute lymphoblastic leukemia entering a clinical protocol, which included a spinal tap. Only cases with a normal CSF were included in this study. None of the selected CSF samples belonged to children with previous oncological and/or hematological treatments. Group 2 (n: 111, 44 controls (24F/20M), 67 MCMA (33F/34M), Meanage = 13.37 years, SD = 4.2) were clinically healthy children from MCMA and control cities and their serum samples were taken as part of their pediatric examination during a longitudinal follow-up. Inclusion criteria for all participating children were: negative smoking history and environmental tobacco exposure, lifelong residency in MCMA or a control city, residency within 5 miles of the city monitoring stations, full term birth, and unremarkable clinical histories prior to the hospital admission (for the children CSF donors). Low and high pollution exposed children were matched by age, gender, and socioeconomic status.
CSF Samples
Spinal tap was performed in the supine position from lumbar levels using a standard 22 spinal needle. CSF was collected dripping in free air in1 ml aliquot into Nalge Nunc polypropylene CryoTubes. Lumbar puncture samples were collected during non-traumatic, non-complicated procedures. CSF pleocytosis was defined as CSF white blood cell (WBC) counts of >7 cells per mm3. CSF samples were used for the laboratory procedures without dilution steps.
Peripheral Blood Analysis
Blood was collected from an antecubital vein using a 21-G needle. After centrifugation at 3000 rpm for 10 min, aliquots of 1.5 ml serum were transferred to CryoTubes and samples were frozen at −20°C and then transferred to −80°C and stored until further analysis. Blood samples were also collected for a complete blood count (CBC) with differential. Serum samples were processed for Multiplexing LASER Bead Technology (Eve Technologies, Calgary, Canada) that included 41 cytokines and chemokines: EGF, Eotaxin, FGF-2, Flt-3L, Fractalkine, G-CSF, GM-CSF, GRO, IFNα2, IFNγ, IL-1α, IL-1β, IL-1ra, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12 (p40), IL-12 (p70), IL-13, IL-15, IL-17a, IP-10, MCP-1, MCP-3, MDC, MIP-1α, MIP-1β, PDGF-AA, PDGF-AB/BB, RANTES, sCD40L, TGFα, TNFα, TNFβ, and VEGF. An ELISA was done for the quantification of PrPc in serum and CSF, BetaPrion®Human EIA (AJ RoboScreen GmbH, Leipzig, Germany). Macrophage inhibitory factor (MIF) was done using an ELISA procedure from R&D (Minneapolis, MN 55413, USA). Serum samples were run without dilution steps.
Data Analysis
First, we calculated the summary statistics (mean ± standard deviation) of all relevant variables including age, inflammatory mediators and PrPc results. In the 28 CSF samples, we worked with 42 variables and established with independent samples t-tests which variables were statistically significant between children with high exposures to air pollutants (Mexico City) and children residing in low pollution cities (Controls). Spearman correlations were run between the CSF target inflammatory mediators and PrPc. In the 111 serum samples, we worked with 2 variables and established with independent samples t-tests which variables were statistically significant between Mexico City and control children. Significance was assumed at p < 0.05 and data expressed as mean values ± SD. All the statistical analyses described above were performed on statistical software “R” (http://www.r-project.org/).
Results
CSF Data
CSF samples were clear, colorless, with a normal opening pressure, a mean WBC count of 2.1 ± 1 cells per mm3 and no RBC. Glucose was 55.9 ± 6.7 mg/100 ml in controls and 54.8 ± 11.7 mg/100ml in Mexico City children (p = 0.3). Table 1 shows the results of the 42 variables included in the analysis. Six variables were statistically significant, including: MIF, PrPC, IL-6, IL1Ra, IL-2, and MIP1α (this is not an effect of the number of multiple comparisons, i.e., inflation of Type I error, because the binomial probability that 6 comparisons out of 42 would turn out to be significant by chance is less than 0.0001). Relevant Spearman correlations between CSF PrPC and target inflammatory mediators are seen in Table 2. We found significant correlations between PrPC and IL-15 (p = 0.025) and RANTES (p = 0.04), MIF and IL-6 (p < 0.000) and IL-6 with IL-15 and RANTES (p = 0.03).
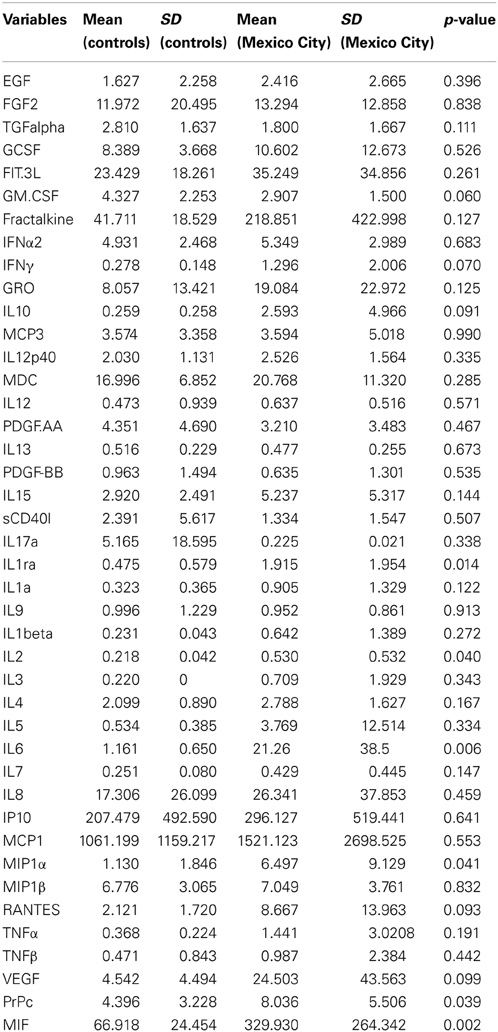
Table 1. Cerebro-spinal- fluid inflammatory mediators in Control vs. Mexico City children (values are shown in pg/ml).
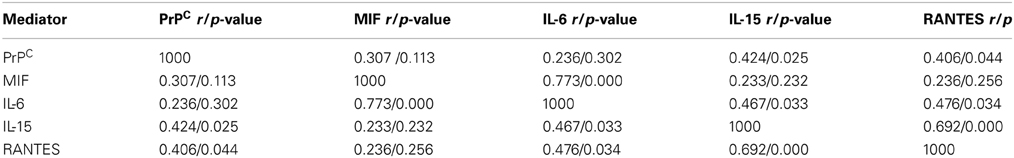
Table 2. Spearman correlations in CSF target inflammatory mediators and PrPC Control v Mexico City children.
Serum Data
MIF and PrPC results in serum are seen in Table 3. MIF was significantly higher (p = 0.009) in Mexico City children, while PrPC showed no differences between cohorts.
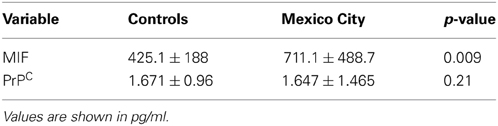
Table 3. Serum MIF and PrPC in Control v Mexico City children.
Discussion
Cerebro-spinal-fluid concentrations of Macrophage migration inhibitory factor MIF, IL-6, IL-1Ra, IL-2, Macrophage Inflammatory Protein 1-α [MIP1α,Chemokine (C-C Motif) Ligand 3] and PrPC differentiate children with high vs. low air pollution exposures and suggest a complex interplay of a network of multipotent cytokines and normal cellular proteins with known neuromodulatory actions participating in neuroinflammatory responses associated with exposures to air pollutants. Our data also suggests that MIF in serum is a potential biomarker bridge to identify children with CNS inflammation.
IL-6, IL-1Ra, IL-2, and MIF are important players in the pathogenesis of neuroinflammation and the systemic inflammation associated with chronic exposures to high concentrations of air pollutants, while the increased CSF concentrations of PrPC could potentially represent a neuroprotective response against oxidative stress (Griffiths et al., 2007; Carnini et al., 2010).
IL-6 is a pleiotropic cytokine with key roles in inflammatory responses and in directing T cell differentiation in adaptive immunity. The elevation of IL-6 in the CSF of highly exposed children and its significant correlation with other potent chemokines and cytokines regulating and coordinating innate and adaptive immune responses is important in the developing CNS.
IL-6 has been implicated in systemic and brain neo-angiogenesis (Gertz et al., 2012). In traumatic brain injury the presence of low concentrations of IL-6 [IL-6 knock-out (KO) mice] is associated with poor behavior performance, and impacts the expression of IL1β, another powerful pro-inflammatory cytokine (Ley et al., 2011). Neuroprotective associations of specific IL-6 SNP (i.e., the G-allele of the SNP rs1800795) on hippocampal volumes (Baune et al., 2012), the protection of midbrain dopaminergic neurons by IL-6 in the model of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) (Spittau et al., 2012), and the significant attenuation of IL-6 concentrations and lower gray volume in old rhesus macaques by a caloric-restricted diet (Willette et al., 2010) support a complex interaction between IL-6, other inflammatory mediators and protective brain responses. The role of IL-6 in long-term potentiation (LTP) and learning is well known (Del Rey et al., 2013). The increased expression of IL-6 is causally related to an increase in synaptic strength since it was abrogated when LTP was interfered by blockade of NMDA-glutamate receptors (Del Rey et al., 2013).
Detrimental effects on the other hand, are described between high concentrations of IL-6, reduced hippocampal volumes and major depressive disorders (Frodl et al., 2012) and in chronic systemic inflammatory conditions with increased risk of stroke (Drake et al., 2011). In advanced HIV infection, CSF concentrations of IL-6 show elevations in patients on suppressive combination antiretroviral therapy regardless of cognitive status, implying the persistence of intra-thecal inflammation even in the absence of clinical manifestations (Kamat et al., 2012).
Of utmost importance for this work and the higher concentrations of CSF IL-6 in Mexico City children is the fact we are seeing the hallmarks of Alzheimer i.e., hyperphosphorilated tau and Aβ42 amyloid plaques in 44 and 51%, respectively, of highly exposed Mexico City teens vs. 0% in low pollution control children. In the literature involving elderly individuals, the associations between IL-6 and dementia are controversial (Papassotiropoulos et al., 2001; Jellinger, 2010; Helmy et al., 2012). Galimberti et al., reported the highest CSF IL-6 concentrations in AD patients with mild cognitive deterioration, suggesting to the authors a key role of IL-6 in the initial phases of neurodegeneration (Galimberti et al., 2008). Combarros et al., suggested that dysregulation of IL-6 in some elderly people contributes to the development of AD (Combarros et al., 2009). Oxidative damage and neuroinflammation are crucial players in neurodegenerative diseases (Nunomura et al., 2001, 2006, 2012a,b), and both are present in the context of air pollution exposures (Calderón-Garcidueñas et al., 2012a). Moreover, under experimental conditions, exposures to different components of air pollutants cause oxidative stress, neuroinflammation and neurodegeneration (Calderón-Garcidueñas et al., 2003, 2008a, 2012a,b; Levesque et al., 2011, 2013). Thus, the CSF and brain inflammatory imbalance observed in highly exposed children could represent an early physiological reaction to chronic environmental stress contributing later to the establishment of neurodegenerative processes with childhood clinical manifestations (Calderón-Garcidueñas et al., 2008b, 2011b, 2012b, 2013). Of critical importance are the portals of entry of air pollutants in the urban setting: (i) the nasal pathway through the olfactory, trigeminal nerves and accessory posterolateral nerve, (ii) the red blood cells and monocytes transporting ultrafine PM and then delivering the particles to distant organs including the brain, (iii) the direct access of PM organic and inorganic components to the systemic circulation through the alveolar-capillary interphase, (iv) the gastrointestinal and vagal pathways (Calderón-Garcidueñas et al., 2007; Block and Calderón-Garcidueñas, 2009; Dhuria et al., 2010; Bleier et al., 2012; Lucchini et al., 2012).
Thus, the association between IL-6 serum increases and deficits in verbal memory (Grassi-Oliveira et al., 2011) obligates us to explore the cognitive consequences of an inflammatory imbalance and cognitive impairment in highly exposed cohorts (Aung et al., 2011; Tsai et al., 2012; Uski et al., 2012; Wittkopp et al., 2013).
Intrathecal inflammation in these children is counterbalanced by the increased production of interleukin 1 receptor antagonist (IL-1Ra), a natural endogenous antagonist of IL-1, critical for a variety of brain effects (Arend, 2002; Gadek-Michalska and Bugajski, 2010; Akash et al., 2013; Peters et al., 2013). IL-1RA exhibits low uptake in the normal brain, with rapid metabolism, a short biological half-life (4–6 h), and excretion via the kidneys (Cawthorne et al., 2011). IL-1Ra higher CSF concentrations in highly exposed children, is a key observation since we have previously described a significant frontal up-regulation of IL-1β in Mexico City children (Calderón-Garcidueñas et al., 2008a). IL-1Ra has clear neuroprotective effects in experimental perinatal inflammation and hypoxic-ischemic injuries (Girard et al., 2012), reduces ischemic brain damage and inflammation in co-morbid rats (Pradillo et al., 2012), prevents postoperative cognitive decline and neuroinflammation in older rats (Barrientos et al., 2012), and plays a key anti-inflammatory role in AD (Rubio-Perez and Morillas-Ruiz, 2012). Interestingly, experimental endotoxemia in healthy male volunteers produces significant transient increases in IL-1Ra and decreases in mood, likely reflecting a compensatory strategy or a greater social cognitive processing as a function of sickness (Kullmann et al., 2013). We interpreted the CSF IL-1Ra increases as a neuroprotective response in Mexico City children.
On the other hand, the increases in CSF IL-2 are in keeping with the critical role of this pro-inflammatory cytokine in both effector T-cell development and FoxP3(+) CD4(+) Treg-cell homeostasis (Shameli et al., 2013). Of utmost interest in the context of air pollution is the role of IL-2 in regulating inflammation in an organ-specific manner through the migration and retention of CD4(+) T-cells (both Th1 and Th2) (Ju et al., 2012). Since IL-2 is also required for regulating the Th2 cytokine response during T-cell activation and participates in multifactorial autoimmune responses (Wang et al., 2009; Baine et al., 2013), the issue of autoimmunity to CNS components warrants full investigation in exposed children especially those with extensive white matter hyperintense lesions (Calderón-Garcidueñas et al., 2008b, 2012b).
Macrophage Inflammatory Protein 1-α [Chemokine (C-C Motif) Ligand 3/CCR3] CSF increases in Mexico City children relate to its well known role in inflammatory responses, angiogenesis and tissue remodeling through binding with receptors to other chemokines (i.e., CCR1, CCR4, CCR5) (Gaspar et al., 2013).
MIF is a newly arrived player in the air pollution scenario, is an active participant in innate and acquired immunity (Savaskan et al., 2012), its association with the production of several inflammatory mediators including IL-6, TNF-α, and IFN-γ, and the identification of CSF MIF as a biomarker in AD and mild cognitive impairment patients (MCI) (Popp et al., 2009; Craig-Schapiro et al., 2011) has a great impact in air pollution-related brain effects.
The role of MIF as a neuro-immune mediator linking inflammation with depressive symptoms and HPA dysregulation (Anisman and Hayley, 2012; Savaskan et al., 2012), and its concomitant increases in CSF and serum makes MIF a potential biomarker to identify children with systemic and CNS inflammation. A facet of MIF of importance for us is the potentiation with autoimmune-mediated neuroinflammation (Cox et al., 2013). Cox et al. demonstrated that MIF is essential for microglial activation and the production of IL-6, IL-1β, TNF-α, and inducible NO synthase in an experimental autoimmune encephalomyelitis model. Cox et al., work is important to us because the target cytokines described in their paper are up-regulated in brain tissues of highly exposed children, and the same cytokines exhibit higher serum concentrations when compared with low pollution exposed children (Calderón-Garcidueñas et al., 2008a,b, 2009, 2011a, 2012a,b).
The CSF increases of the normal cellular isoform of the prion protein (PrPC) in exposed children deserve to be discussed under three important headings:
1. The PrPC neuroprotective role related primarily to its antioxidant properties, its capacity as a radical scavenger, its key role in signal transduction and cell survival, neuronal zinc regulation, and its functional importance in the protection against oxidative stress and metal toxicity (Brown et al., 1997; Kuwahara et al., 1999; Viles et al., 1999; Milhavet and Lehmann, 2002; Hu et al., 2007; Westergard et al., 2007; Carnini et al., 2010; Martins et al., 2010; Bertuchi et al., 2012; Alfaidy et al., 2013; Watt et al., 2013).
2. The PrPC role in animal behavior suggesting that the absence of the prion protein could result in altered neural processing (Massimino et al., 2013).
3. The associations of PrPC with neuropathology apart from the conventional prion diseases (Jiménez-Huete et al., 1998; Ferrer et al., 2001; Voigtländer et al., 2001; Checler and Vincent, 2002; Aguzzi and Haass, 2003; Rezaie et al., 2005; Schwarze-Eicker et al., 2005; Ramljak et al., 2008). Of interest to us are the controversial issues in the literature linking Aβ oligomers and PrPC (Laurén et al., 2009; Nygaard and Strittmatter, 2009; Gunther and Striitmatter, 2010; Kessels et al., 2010; Barry et al., 2011; Saijo et al., 2011; Larson et al., 2012; Um et al., 2012; Chen et al., 2013; Kudo et al., 2013; Whitehouse et al., 2013; Younan et al., 2013). A few specific examples to make the point: Kudo et al. showed that Prnp (-/-) mice are resistant to the neurotoxic effect of oligomeric Aβ in vivo and in vitro (Kudo et al., 2013). Chen et al. have shown PrPC over-expression down-regulated tau protein and PrPC lacking the Aβ oligomer binding site was incapable of rescuing the level of tau reduction. These authors demonstrated that PrPC down-regulated tau via Fyn pathway and the effect can be regulated by Aβ oligomers (Chen et al., 2013). Whitehouse et al. measured PrPC in the frontal cortex of 24 sporadic AD brains vs. 24 age-matched controls and found a significant decreased of PrPC in AD brains. Interestingly, PrPC significantly inversely correlated with BACE1 activity, Aβ load, soluble Aβ, and insoluble Aβ and with the stage of disease, as indicated by Braak tangle stage (Whitehouse et al., 2013). The authors concluded that brain PrPC level may be important in influencing the onset and progression of sporadic AD (Whitehouse et al., 2013).
Why are all these papers relevant to our work? We recently published a paper (Calderón-Garcidueñas et al., 2012a) showing a 15 fold frontal down-regulation of mRNA PrPC in Mexico City residents (18 ± 8.7 years) vs. low air pollution age matched controls. In the same work, we showed frontal tau hyperphosphorylation and Aβ diffuse plaques in the exposed kids, but none in low air pollution controls. Thus, the regional frontal PrPC down-regulation shown by microarray analysis is of deep concern given the age of the autopsy subjects (on average 8 years older) than our CSF children in this study. PrPC might have significant longitudinal variations in mRNA expression by brain region (Rezaie et al., 2005; Velayos et al., 2009), age, gender, and other factors in the setting of air pollution. Moreover, because the CSF is produced and reabsorbed throughout the entire CSF-Interstitial fluid (IF) functional unit (Chikly and Quaghebeur, 2013), our results highlight the need to define PrPC changes both directly in different regions of the brain and in the CSF in low vs. high pollution exposed children and young adults. Furthermore, since PrPC likely plays a key protective role against sporadic AD (Whitehouse et al., 2010; Griffiths et al., 2012) we will expect age-related decreases in specific regional brain areas inversely related to the development of AD hallmarks in highly exposed individuals.
The second reason for concern is the impact of PrPC in cognition. Gimbel et al. showed that AD transgenic mice with intact PrPC expression exhibit deficits in spatial learning and memory, while mice lacking PrPC, but containing Aβ plaque derived from APPswe/PSen1DeltaE9 transgenes, show no detectable impairment of spatial learning and memory (Gertz et al., 2010). Thus, deletion of PrPC expression dissociates Aβ accumulation from behavioral impairment in AD mice, with the cognitive deficits selectively requiring PrPC (Gertz et al., 2010). Interestingly, Schmidt et al. (2013) showed no association between CSF PrPC and cognitive status in 114 AD patients, while Breitling et al. reported in a 5 years follow-up study of 1322 elderly Germans (aged 65+ years at baseline), an inverse association between serum PrPc and cognitive functioning (Breitling et al., 2012). In our study, CSF PrPC was significantly higher in exposed vs. low pollution children, while serum PrPC concentrations showed no differences.
The significant correlations of PrPC with RANTES and IL-15 deserve a comment. Regulated upon Activation, Normal T-cell Expressed and Secreted (RANTES) is a proinflammatory chemokine produced by neurons and capable of upregulating neurotrophic factors and extending cell survival after ischemic stroke (Tokami et al., 2013), while IL-15 instructs the generation of a distinct memory T-lymphocyte subset, intermediate between naive and central memory cells (Cieri et al., 2013). These findings highlight the complex link between normal brain proteins, powerful chemokines, and immune driven cytokines.
It has been clear for a number of years that neuroinflammation has critical implications for cognition, behavior, altered brain growth and neurodegeneration. Brain dysbalances in inflammatory mediators and alterations of essential proteins with key roles in CNS development and neuroprotection will inevitably have detrimental consequences for young brains. We already described some of these consequences in Mexico City youth: differences in white matter volumes involving right parietal and bilateral temporal areas and cognitive deficits consistent with impairment of the targeted lobes, olfaction deficits, auditory and vestibular nuclei accumulating α synuclein and/or β amyloid and significant involvement of the medial superior olive neurons, critically involved in brainstem auditory evoked potentials. Elevation of indices of neuroinflammation and oxidative stress, and AD and PD associated pathology completing the grim picture (Calderón-Garcidueñas et al., 2003, 2008a,b, 2010, 2011a,b, 2012a,b, 2013; Calderón-Garcidueñas and Torres-Jardón, 2012).
At the core of our observations lies the complex dynamic interaction in different compartments of cytokines, chemokines and brain proteins responding to the constant state of inflammation and oxidative stress resulting from unhealthy environmental exposures. The short term consequences are having an impact in the performance of children in school and in their violent behavior (Calderón-Garcidueñas and Torres-Jardón, 2012), while the longer effects are already seen in neuropathological studies of children with accidental deaths (Calderón-Garcidueñas et al., 2008b, 2012b, 2013). There is no doubt of the close linkage between the brain and the immune system particularly in the developing brain (Bilbo and Schwarz, 2012). Brisk responses are at hand both systemically and in the brain upon endogenous and exogenous environmental signals, however, the developing brain is delicately sensitive to chronic stimuli and the end result could be a pathological process with deleterious outcomes for the individual.
Looking Forward and Limitations
Despite controversy regarding the mechanistic pathways involved in the CNS damage associated with exposure to air pollutants, we agree that oxidative stress, endothelial dysfunction vascular damage and neuroinflammation are at the core of the pathology, and the same factors play a key role in neurodegenerative diseases (Qin et al., 2007; Rivest, 2009; Levesque et al., 2011; Castellani and Perry, 2012; de la Torre, 2012; Nunomura et al., 2012a,b; Calderón-Garcidueñas et al., 2013). We are looking forward to bridging the gap between early neuroinflammation and its neurodegenerative consequences, an issue of importance in childhood and adolescence when cognitive performance is critical.
There is a need for looking into the CSF early responses to air pollution because we can connect with the knowledge available for elderly populations, i.e., the progression of Alzheimer core biomarkers [Aβ(1-42), total tau and phosphorylated tau] and inflammatory markers useful in the clinical practice to evaluate patients with mild cognitive impairment and their course to AD. This is critical, given that implementation of early preventive pathways (Castellani and Perry, 2012) in the developing brain could ameliorate or drastically modified the pathology in later years.
Our results are potentially limited by the fact our CSF normal samples were taken from a population of children with a hematological workup for a neoplastic process, albeit no CNS involvement, while our serum samples were from clinically healthy children. As pediatricians we are totally aware of the ethical issues in taking CSF samples from healthy children, so the readily available normal CSF samples destined to be discarded are a good initial source of exploratory samples. Nevertheless, the robust CSF increase in the described inflammatory mediators in highly exposed children warrants extensive investigations using available normal CSF samples in Mexico and around the world. Two issues are important here: the selection of multiplex platforms and knowledge of their ability to accurately and sensitively detect cytokines in CSF and blood (serum/plasma) (Malekzadeh et al., 2012). The second one, the use of standardized operating procedures for the recollection of CSF samples, a key point if we want to decrease the variability attributed to different pre-analytical procedures between laboratories (del Campo et al., 2012).
Summary
We have quantified CSF responses for a selected panel de inflammatory mediators and PrPC in two groups of children with contrasting air pollution lifetime exposures. Our results identify MIF, IL6, IL1Ra, IL-2, and PrPC as potential CSF biomarkers differentiating the high vs. low air pollution exposed children, supporting the previous evidence of an ongoing neuroinflammatory process in Mexico City children. Serum MIF was identified as a robust biomarker in Mexico City children.
Fine tuning of immune-to-brain communication is crucial to neural networks appropriate functioning, thus the short and long term effects of systemic inflammation and dysregulated neural immune responses are of deep concern for millions of highly exposed children.
A large body of work on PrPc, IL-6, MIF, IL-2, and the IL-1 family already exists, thus expanding this knowledge in the scenario of air pollution pediatric effects could greatly facilitate our understanding of the downstream mechanisms of the complex interaction of cytokines/chemokines/ PrPc immune responses in the developing brain and the resulting short and long term brain effects.
The role of PrPc in highly exposed children is an issue worth investigating further as the field moves forward, since we believe knowledge on PrPc responses in a developing brain will shed light on neuroprotective mechanisms against the onset of Alzheimer disease.
Although the two objectives of this study were accomplished: (i) To evaluate the concentrations of PrPC and selected inflammatory mediators in normal CSF from cohorts of children with high vs. low pollution controls and (ii) To explore serum PrPC and MIF in clinically healthy children representing cohorts with high and low air pollution exposures, our immediate goal will be to assess whether the identified biomarkers will be useful in the identification of children at higher risk for cognitive deficits, behavioral disorders, and structural and volumetric brain alterations.
Our ultimate goal is that following the identification of children at the highest risk for detrimental brain effects, we protect them through multidimensional interventions yielding both impact and reach. The prospective of our efforts is readily apparent.
Defining the linkage and the health consequences of the brain/immune system interactions in the developing brain chronically exposed to air pollutants ought to be of pressing importance for public health.
Author Contributions
Lilian Calderón-Garcidueñas planned and directed the study, wrote the manuscript, performed all experiments and supported the study from personal funds. Janet V. Cross planned the study, performed the MIF determinations and wrote the manuscript. Amedeo D'Angiulli wrote, edited and formatted the manuscript and performed part of the statistic analysis. Maricela Franco-Lira and Mariana Aragón-Flores obtained patient consent and ethical approvals, got the samples and wrote the manuscript. Michael Kavanaugh wrote the manuscript. Chih-kai Chao and Charles Thompson performed the PrPC and wrote the manuscript. Ricardo Torres-Jardón wrote the manuscript and Jing Chang and Hongtu Zhu performed the statistical analysis. All authors approved the final draft of the manuscript for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported in part by the UM-CHPBS Endowment Fund Program, NIMHD grant # S21-MD000236 and a AUCC LACREG grant to Drs. Calderón and D'Angiulli. We thank Dennis Templeton, MD PhD, Chair, Department of Pathology, School of Medicine, University of Virginia, Charlottesville, VA for his generous support of this research while Dr. Lilian Calderón-Garcidueñas was a U Virginia visiting professor. We also thank Lara Ferreira-Azevedo, 5th year Pharmacy visiting student from Brazil for her technical assistance.
References
Aguzzi, A., and Haass, C. (2003). Games played by rogue proteins in prion disorders and Alzheimer's disease. Science 302, 814–818. doi: 10.1126/science.1087348
Akash, M. S., Rehman, K., and Chen, S. (2013). IL-1Ra and its delivery strategies: inserting the association in perspective. Pharm. Res. doi: 10.1007/s11095-013-1118-0. [Epub ahead of print].
Alfaidy, N., Chauvet, S., Donadio-Andrei, S., Salomon, A., Saoudi, Y., Richaud, P., et al. (2013). Prion protein expression and functional importance in developmental angiogenesis: role in oxidative stress and copper homeostasis. Antioxid. Redox Signal. 18, 400–411. doi: 10.1089/ars.2012.4637
Altmeppen, H. C., Puig, B., Dohler, F., Thurm, D. K., Falker, C., Krasemann, S., et al. (2012). Proteolytic processing of the prion protein in health and disease. Am. J. Neurodegener. Dis. 1, 15–31.
Anisman, H., and Hayley, S. (2012). Inflammatory factors contribute to depression and its comorbid conditions. Sci. Signal. 5:pe45. doi: 10.1126/scisignal.2003579
Arend, W. P. (2002). The balance between IL-1 and IL-1Ra in disease. Cytokine Growth Factor Rev. 13, 323–340. doi: 10.1016/S1359-6101(02)00020-5
Aung, H. H., Lame, M. W., Gohil, K., He, G., Denison, M. S., Rutledge, J. C., et al. (2011). Comparative gene responses to collected ambient particles in vitro: endothelial responses. Physiol. Genomics 43, 917–929. doi: 10.1152/physiolgenomics.00051.2011
Bacher, M., Deuster, O., Aljabari, B., Egensperger, R., Neff, F., Jessen, F., et al. (2010). The role of macrophage migration inhibitory factor in Alzheimer's disease. Mol. Med. 16, 116–121. doi: 10.2119/molmed.2009.00123
Baine, I., Basu, S., Ames, R., Sellers, R. S., and Macian, F. (2013). Helios induces epigenetic silencing of IL2 gene expression in regulatory T cells. J. Immunol. 190, 1008–1016. doi: 10.4049/jimmunol.1200792
Barrientos, R. M., Hein, A. M., Frank, M. G., Watkins, L. R., and Maier, S. F. (2012). Intracisternal interleukin-1 receptor antagonist prevents postoperative cognitive decline and neuroinflammatory response in aged rats. J. Neurosci. 32, 14641–14648. doi: 10.1523/JNEUROSCI.2173-12.2012
Barry, A. E., Klyubin, I., McDonald, J. M., Mably, A. J., Farrell, M. A., Scott, M., et al. (2011). Alzheimer's disease brain-derived amyloid-β-mediated inhibition of LTP in vivo is prevented by immunotargeting cellular prion protein. J. Neurosci. 31, 7259–7263. doi: 10.1523/JNEUROSCI.6500-10.2011
Baune, B. T., Konrad, C., Grotegerd, D., Suslow, T., Birosova, E., Ohrmann, P., et al. (2012). Interleukin-6 gene (IL6): a possible role of brain morphology in the healthy adult brain. J. Neuroinflammation 9:125. doi: 10.1186/1742-2094-9-125
Bernhagen, J., Calandra, T., Mitchell, R. A., Martin, S. B., Tracey, K. J., Voelter, W., et al. (1993). MIF is a pituitary-derived cytokine that potentiates lethal endotoxemia. Nature 365, 756–759. doi: 10.1038/365756a0
Bertuchi, F. R., Bourgeon, D. M., Landemberger, M. C., Martins, V. R., and Cerchiaro, G. (2012). PrPC displays an essential protective role from oxidative stress in an astrocyte cell line derived from PrPC knockout mice. Biochim. Biophys. Res. Commun. 418, 27–32. doi: 10.1016/j.bbrc.2011.12.098
Bilbo, S. D., and Schwarz, J. M. (2012). The immune system and developmental programming of brain and behavior. Front. Neuroendocrinol. 33, 267–286. doi: 10.1016/j.yfrne.2012.08.006
Bleier, B. S., Feldman, R., Sadow, P. M., Wu, A., Ting, J., and Metson, R. (2012). The accessory posterolateral nerve: an immunohistological analysis. Am. J. Rhinol. Allergy 26, 271–273. doi: 10.2500/ajra.2012.26.3804
Block, M., and Calderón-Garcidueñas, L. (2009). Air Pollution: mechanisms of neuroinflammation and CNS disease. Trends Neurosci. 32, 506–516. doi: 10.1016/j.tins.2009.05.009
Bradford, B. M., and Mabbott, N. A. (2012). Prion disease and the innate immune system. Viruses 4, 3389–3419. doi: 10.3390/v4123389
Breitling, L. P., Muller, H., Stegmaier, C., Kliegel, M., and Brenner, H. (2012). Association of prion protein with cognitive functioning in humans. Exp. Gerontol. 47, 919–24. doi: 10.1016/j.exger.2012.08.001
Bremer, J., Baumann, F., Tiberi, C., Wessig, C., Fischer, H., Schwarz, P., et al. (2010). Axonal prion protein is required for peripheral myelin maintenance. Nat. Neurosci. 13, 310–318. doi: 10.1038/nn.2483
Brown, D. R., Schmidt, B., and Kretzschmar, H. A. (1997). Effects of oxidative stress on prion protein expression in PC12 cells. Int. J. Dev. Neurosci. 15, 961–972. doi: 10.1016/S0736-5748(97)00042-7
Bucala, R. (2013). MIF, MIF alleles, and prospects for therapeutic intervention in autoimmunity. J. Clin. Immunol. 33(Suppl. 1), S72–S78. doi: 10.1007/s10875-012-9781-1
Calderón-Garcidueñas, L., Engle, R., Mora-Tiscareño, A., Styner, M., Gomez-Garza, G., Zhu, H., et al. (2011a). Exposure to severe urban pollution influences cognitive outcomes, brain volume and systemic inflammation in clinically healthy children. Brain Cognition 77, 345–355. doi: 10.1016/j.bandc.2011.09.006
Calderón-Garcidueñas, L., D'Angiulli, A., Kulesza, R. J., Torres-Jardón, R., Romero, L., Keefe, S., et al. (2011b). Air pollution is associated with brainstem auditory nuclei pathology and delayed brainstem auditory evoked potentials. Int. J. Dev. Neurosci. 29, 365–375. doi: 10.1016/j.ijdevneu.2011.03.007
Calderón-Garcidueñas, L., Franco-Lira, M., Henríquez-Roldán, C., González-Maciel, A., Reynoso-Robles, R., Villarreal-Calderon, R., et al. (2010). Urban air pollution: influences on olfactory function and pathology in exposed children and young adults. Exp. Toxicol. Pathol. 62, 91–102. doi: 10.1016/j.etp.2009.02.117
Calderón-Garcidueñas, L., Franco-Lira, M., Mora-Tiscareño, A., Medina-Cortina, H., Torres-Jardón, R., and Kavanaugh, M. (2013). Early Alzheimer's and Parkinson's disease pathology in urban children: friend versus foe responses-it is time to face the evidence. Biomed. Res. Int. doi: 10.1155/2013/161687
Calderón-Garcidueñas, L., Franco-Lira, M., Torres-Jardón, R., Henriquez-Roldán, C., Valencia-Salazar, G., González-Maciel, A., et al. (2007). Pediatric and systemic effects of chronic air pollution exposure: nose, lung, heart, and brain pathology. Toxicol. Pathol. 35, 154–162. doi: 10.1080/01926230601059985
Calderón-Garcidueñas, L., Kavanaugh, M., Block, M. L., D'Angiulli, A., Delgado-Chávez, R., Torres-Jardón, R., et al. (2012a). Neuroinflammation, hyperphosphorilated tau, diffuse amyloid plaques and down- regulation of the cellular prion protein in air pollution exposed children and adults. J. Alzheimers Dis. 28, 93–107.
Calderón-Garcidueñas, L., Mora-Tiscareño, A., Styner, M., Gómez-Garza, G., Zhu, H., Torres-Jardón, R., et al. (2012b). White matter hyperintensities, systemic inflammation, brain growth and cognitive functions in children exposed to air pollution. J. Alzheimers Dis. 31, 183–91.
Calderón-Garcidueñas, L., Macías-Parra, M., Hoffmann, H. J., Valencia-Salazar, G., Henríquez-Roldán, C., Monte, O. C., et al. (2009). Immunotoxicity and environment: immunodysregulation and systemic inflammation in children. Toxicol. Pathol. 37, 161–169. doi: 10.1177/0192623308329340
Calderón-Garcidueñas, L., Maronpot, R. R., Torres-Jardón, R., Henriquez-Roldán, C., Schoonhoven, R., Acuña-Ayala, H., et al. (2003). DNA damage in nasal and brain tissues of canines exposed to air pollutants is associated with evidence of chronic brain inflammation and neurodegeneration. Toxicol. Pathol. 31, 524–538. doi: 10.1080/01926230390226645
Calderón-Garcidueñas, L., Solt, A. C., Henríquez-Roldán, C., Torres-Jardón, R., Nuse, B., Herritt, L., et al. (2008a). Long-term air pollution exposure is associated with neuroinflammation, an altered innate immune response, disruption of the blood-brain-barrier, ultrafine particulate deposition, and accumulation of amyloid beta-42 and alpha-synuclein in children and young adults. Toxicol. Pathol. 36, 289–310.
Calderón-Garcidueñas, L., Mora-Tiscareño, A., Gómez-Garza, G., Broadway, J., Chapman, S., Valencia-Salazar, G., et al. (2008b). Air pollution, cognitive deficits and brain abnormalities: a pilot study with children and dogs. Brain Cogn. 68, 117–127. doi: 10.1016/j.bandc.2008.04.008
Calderón-Garcidueñas, L., and Torres-Jardón, R. (2012). Air pollution, socioeconomic status and children's cognition in megacities: the Mexico City scenario. Front. Psychol. 3:217. doi: 10.3389/fpsyg.2012.00217
Carnini, A., Casha, S., Yong, V. W., Hurlbert, R. J., and Braun, J. E. (2010). Reduction of PrPC in human cerebrospinal fluid after spinal cord injury. Prion 4, 80–86. doi: 10.4161/pri.4.2.11756
Castellani, R. J., and Perry, G. (2012). Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch. Med. Res. 43, 694–698. doi: 10.1016/j.arcmed.2012.09.009
Cattaneo, A., Gennarelli, M., Uher, R., Breen, G., Farmer, A., Aitchison, K. J., et al. (2013). Candidate genes expression profile aasociated with antidepressants response in the GENDEP study: differentiating between baseline predictors and longitudinal targets. Neuropsychopharmacology 38, 377–385. doi: 10.1038/npp.2012.191
Cawthorne, C., Prenant, C., Smigova, A., Julvan, P., Maroy, R., Herholz, K., et al. (2011). Biodistribution, pharmacokinetics and metabolism of interleukin-1 receptor antagonist (IL-1RA) using [18F] IL1RA and PET imaging in rats. Br. J. Pharmacol. 162, 659–672. doi: 10.1111/j.1476-5381.2010.01068.x
Checler, F., and Vincent, B. (2002). Alzheimer's and prion diseases: distinct pathologies, common proteolytic denominators. Trends Neurosci. 25, 616–620. doi: 10.1016/S0166-2236(02)02263-4
Chen, B., and Kan, H. (2008). Air pollution and population health: a global challenge. Environ. Health Prev. Med. 13, 94–101. doi: 10.1007/s12199-007-0018-5
Chen, R. J., Chang, W. W., Lin, Y. C., Cheng, P. L., and Chen, Y. R. (2013). Alzheimer's amyloid-β oligomers rescue cellular prion protein induced Tau reduction via Fyn pathways. ACS Chem. Neurosci. 4, 1287–1296. doi: 10.1021/cn400085q
Chikly, B., and Quaghebeur, J. (2013). Reassessing cerebrospinal fluid (CSF) hydrodynamics: a literature review presenting a novel hypothesis for CSF physiology. J. Bodyw. Mov. Ther. 17, 344–354. doi: 10.1016/j.jbmt.2013.02.002
Choi, C. J., Anantharam, V., Saetveit, N. J., Houk, R. S., Kanthasamy, A., and Kanthasamy, A. G. (2007). Normal cellular prion protein protects against manganese-induced oxidative stress and apoptotic cell death. Toxicol. Sci. 98, 495–509. doi: 10.1093/toxsci/kfm099
Cieri, N., Camisa, B., Cocchiarella, F., Forcato, M., Oliveira, G., Provasi, E., et al. (2013). IL-7 and IL-15 instruct the generation of human memory stem T cells from naive precursors. Blood 121, 573–584. doi: 10.1182/blood-2012-05-431718
Combarros, O., van Duijn, C. M., Hammond, N., Belbin, O., Arias-Vásquez, A., Cortina-Borja, M., et al. (2009). Replication by the epistasis project of the interaction between the genes for IL-6 and IL-10 in the risk of Alzheimer's disease. J. Neuroinflammation 6:22. doi: 10.1186/1742-2094-6-22
Cox, G. M., Kithcart, A. P., Pitt, D., Guan, Z., Alexander, J., Williams, J. L., et al. (2013). Macrophage migration inhibitory factor potentiates autoimmune-mediated neuroinflammation. J. Immunol. 191, 1043–1054. doi: 10.4049/jimmunol.1200485
Craig-Schapiro, R., Kuhn, M., Xiong, C., Pickering, E. H., Liu, J., Misko, T. P., et al. (2011). Multiplex immunoassay panel identifies novel CSF biomarkers for Alzheimer's disease diagnosis and prognosis. PLoS ONE 6:e18850. doi: 10.1371/journal.pone.0018850
de la Torre, J. C. (2012). A turning point for Alzheimer's disease? Biofactors 38, 78–83. doi: 10.1002/biof.200
del Campo, M., Mollenhauer, B., Bertolotto, A., Engelborghs, S., Hampel, H., Simonsen, A. H., et al. (2012). Recommendations to standardize preanalytical confounding factors in Alzheimer's and Parkinson's disease cerebrospinal fluid biomarkers: an update. Biomark. Med. 6, 419–430. doi: 10.2217/bmm.12.46
Del Rey, A., Balschun, D., Wetzel, W., Randolf, A., and Besedovsky, H. O. (2013). A cytokine network involving brain-borne IL-1β, IL-1Ra, IL-18, IL-6 and TNF-α operates during long-term potentiation and learning. Brain Behav. Immun. 33, 15–23. doi: 10.1016/j.bbi.2013.05.011
Dhuria, S. V., Hanson, L. R., and Frey, W. H. 2nd. (2010). Intranasal delivery to the central nervous system: mechanisms and experimental considerations. J. Pharm. Sci. 99, 1654–1673. doi: 10.1002/jps.21924
Ding, T., Zhou, X., Kouadir, M., Shi, F., Yang, Y., Liu, J., et al. (2013). Cellular prion protein participates in the regulation of inflammatory response and apoptosis in BV2 microglia during infection with Mycobacterium bovis. J. Mol. Neurosci. 51, 118–126. doi: 10.1007/s12031-013-9962-2
Drake, C., Boutin, H., Jones, M. S., Denes, A., McColl, B. W., Selvarajah, J. R., et al. (2011). Brain inflammation is induced by co-morbidities and risk factors for stroke. Brain Behav. Immun. 25, 1113–1122. doi: 10.1016/j.bbi.2011.02.008
Dziedzic, T. (2006). Systemic inflammatory markers and risk of dementia. Am. J. Alzheimers Dis. Other Demen. 21, 258–262. doi: 10.1177/1533317506289260
Edwards, K. M., Bosch, J. A., Engeland, C. G., Cacioppo, J. T., and Marucha, P. T. (2010). Elevated macrophage migration inhibitory factor (MIF) is associated with depressive symptoms, blunted cortisol reactivity to acute stress, and lowered morning cortisol. Brain Behav. Immun. 24, 1202–1208. doi: 10.1016/j.bbi.2010.03.011
Ferrer, I., Blanco, R., Carmona, M., Puig, B., Ribera, R., Rey, M. J., et al. (2001). Prion protein expression in senile plaques in Alzheimer's disease. Acta Neuropathol. 101, 49–56.
Freiria-Oliveira, A. H., Blanch, G. T., Li, H., Colombari, E., Colombari, D. S., and Sumners, C. (2013). Macrophage inhibitory factor in the nucleus of solitary tract decreases blood pressure in SHRs. Cardiovasc. Res. 97, 153–160. doi: 10.1093/cvr/cvs297
Frodl, T., Carballedo, A., Hughes, M. M., Saleh, K., Fagan, A., Skokauskas, N., et al. (2012). Reduced expression of glucocorticoid-inducible genes GILZ and SGK-1: high IL6 levels are associated with reduced hippocampal volumes in major depressive disorder. Transl. Psychiatry 2:e88. doi: 10.1038/tp.2012.14
Gadek-Michalska, A., and Bugajski, J. (2010). Interleukin-1 (IL-1) in stress-induced activation of limbic-hypothalamic-pituitary adrenal axis. Pharmacol. Rep. 62, 969–982.
Galimberti, D., Venturelli, E., Fenoglio, C., Guidi, I., Villa, C., Bergamaschini, L., et al. (2008). Intratechal levels of IL6, IL11 and LIF in Alzheimer's disease and frontotemporal lobar degeneration. J. Neurol. 255, 539–544. doi: 10.1007/s00415-008-0737-6
Gaspar, K., Kukova, G., Bunemann, E., Buhren, B. A., Sonkolv, E., Szollosi, A. G., et al. (2013). The chemokine receptor CCR3 participates in tissue remodeling during atopic skin inflammation. J. Dermatol. Sci. 71, 12–21. doi: 10.1016/j.jdermsci.2013.04.011
Gertz, K., Kronenberg, G., Kälin, R. E., Baldinger, T., Werner, C., Balkaya, M., et al. (2012). Essential role of interleukin-6 in post-stroke angiogenesis. Brain 135, 1964–1980. doi: 10.1093/brain/aws075
Gimbel, D. A., Nygaard, H. B., Coffey, E. E., Gunther, E. C., Laurén, J., Gimbel, Z. A., et al. (2010). Memory impairment in transgenic Alzheimer mice requires cellular prion protein. J. Neurosci. 30, 6367–6374. doi: 10.1523/JNEUROSCI.0395-10.2010
Giralt, M., Ramos, R., Quintana, A., Ferrer, B., Erta, M., Castro-Freire, M., et al. (2013). Induction of atypical EAE mediated by transgenic production of IL6 in astrocytes in the absence of systemic IL6. Glia 61, 587–600. doi: 10.1002/glia.22457
Girard, S., Sébire, H., Brochu, M. E., Briota, S., Sarret, P., and Sébire, G. (2012). Postnatal administration of IL-1Ra exerts neuroprotective effects following perinatal inflammation and/or hypoxic-ischemic injuries. Brain Behav. Immun. 26, 1331–1339. doi: 10.1016/j.bbi.2012.09.001
Gourdain, P., Ballerini, C., Nicot, A. B., and Carnaud, C. (2012). Exacerbation of experimental autoimmune encephalomyelitis in prion protein PrPc-null mice: evidence for a critical role of the central nervous system. J. Neuroinflammation 9:25. doi: 10.1186/1742-2094-9-25
Grassi-Oliveira, R., Bauer, M. E., Pezzi, J. C., Teixeira, A. L., and Brietzke, E. (2011). Interleukin-6 and verbal memory in recurrent major depressive disorder. Neuro Endocrinol. Lett. 32, 540–544.
Griffiths, H. H., Whitehouse, I. J., and Hooper, N. M. (2012). Regulation of amyloid-β production by the prion protein. Prion 6, 217–222. doi: 10.4161/pri.18988
Griffiths, M., Neal, J. W., and Gasque, P. (2007). Innate immunity and protective neuroinflammation: new emphasis on the role of neuroimmune regulatory proteins. Int. Rev. Neurobiol. 82, 29–55. doi: 10.1016/S0074-7742(07)82002-2
Gunther, E. C., and Striitmatter, S. M. (2010). Beta-amyloid oligomers and cellular prion protein in Alzheimer's disease. J. Mol. Med. 88, 331–338. doi: 10.1007/s00109-009-0568-7
Hagman, S., Raunio, M., Rossi, M., Dastidar, P., and Elovaara, I. (2011). Disease-associated inflammatory biomarker profiles in blood in different subtypes of multiple sclerosis: prospective clinical and MRI follow-up Study. J. Neuroimmunol. 234, 141–147. doi: 10.1016/j.jneuroim.2011.02.009
Helmy, A. A., Naseer, M. M., Shafie, S. E., and Nada, M. A. (2012). Role of interleukin 6 and alpha-globulins in differentiating Alzheimer and vascular dementias. Neurodegener. Dis. 9, 81–86. doi: 10.1159/000329568
Hu, W., Rosenberg, R. N., and Stüve, O. (2007). Prion proteins: a biological role beyond prion diseases. Acta Neurol. Scand. 116, 75–82. doi: 10.1111/j.1600-0404.2007.00868.x
Jellinger, A. K. (2010). Intrathecal levels of IL6 in Alzheimer's disease. J. Neurol. 257, 142. doi: 10.1007/s00415-009-5312-2
Jiménez-Huete, A., Lievens, P. M., Vidal, R., Piccardo, P., Ghetti, B., Tagliavini, F., et al. (1998). Endogenous proteolytic cleavage of normal and disease-associated isoforms of the human prion protein in neural and non-neural tissues. Am. J. Pathol. 153, 1561–1572. doi: 10.1016/S0002-9440(10)65744-6
Ju, S. T., Sharma, R., Gaskin, F., and Fu, S. M. (2012). IL-2 controls trafficking receptor gene expression and Th2 response for skin and lung inflammation. Clin. Immunol. 145, 82–88. doi: 10.1016/j.clim.2012.07.015
Kaiser, D. M., Acharya, M., Leighton, P. L. A., Wang, H., Daude, N., Wohlgemuth, S., et al. (2012). Amyloid beta precursor protein and prion protein have a conserved interaction affecting cell adhesion and CNS development. PLoS ONE 7:e51305. doi: 10.1371/journal.pone.0051305
Kamat, A., Lyons, J. L., Misra, V., Uno, H., Morgello, S., Singer, E. J., et al. (2012). Monocyte activation markers in cerebrospinal fluid associated with impaired neurocognitive testing in advanced HIV infection. J. Acquir. Immune Defic. Syndr. 60, 234–243. doi: 10.1097/QAI.0b013e318256f3bc
Kessels, H. W., Nguyen, L. N., Nabavi, S., and Malinow, R. (2010). The prion protein as a receptor for amyloid-beta. Nature 466, E3–E4. doi: 10.1038/nature09217
Kudo, W., Petersen, R. B., and Lee, H. G. (2013). Cellular prion protein and Alzheimer disease: link to oligomeric amyloid-β and neuronal cell death. Prion 7, 114–116. doi: 10.4161/pri.22848
Kullmann, J. S., Grigoleit, J. S., Wolf, O. T., Engler, H., Oberbeck, R., Elsenbruch, S., et al. (2013). Experimental human endotoxemia enhances brain activity during social cognition. Soc. Cogn. Affect. Neurosci. doi: 10.1093/scan/nst049. [Epub ahead of print].
Kuwahara, C., Takeuchi, A. M., Nishimura, T., Haraguchi, K., Kubosaki, A., Matsumoto, Y., et al. (1999). Prions prevent neuronal cell-line death. Nature 400, 225–226. doi: 10.1038/22241.
Larson, M., Sherman, M. A., Amar, F., Nuvolone, M., Schneider, J. A., Bennett, D. A., et al. (2012). The complex PrP(c)-Fyn couples human oligomeric Aβ with pathological tau changes in Alzheimer's disease. J. Neurosci. 32, 16857–16871. doi: 10.1523/JNEUROSCI.1858-12.2012
Laurén, J., Gimbel, D. A., Nygaared, H. B., Gilbert, J. W., and Strittmatter, S. M. (2009). Cellular prion protein mediates impairment of synaptic plasticity by amyloid-β oligomers. Nature 457, 1128–1132. doi: 10.1038/nature07761
Lee, K. S., Chung, J. H., Lee, K. H., Shin, M. J., Oh, B. H., and Hong, C. H. (2008). Bioplex analysis of plasma cytokines in Alzheimer's disease and mild cognitive impairment. Immunol. Lett. 121, 105–109. doi: 10.1016/j.imlet.2008.09.004
Levesque, S., Surace, M. J., McDonald, J., and Block, M. L. (2011). Air pollution and the brain: subchronic diesel exhaust exposure causes neuroinflammation and elevates early markers of neurodegenerative disease. J. Neuroinflammation 8:105. doi: 10.1186/1742-2094-8-105
Levesque, S., Taetzsch, T., Lull, M. E., Johnson, J. A., McGraw, C., and Block, M. L. (2013). The role of MAC1 in diesel exhaust particle-induced microglial activation and loss of dopaminergic neuron function. J. Neurochem. 125, 756–765. doi: 10.1111/jnc.12231
Ley, E. J., Clond, M. A., Singer, M. B., and Salim, A. (2011). IL6 deficiency affects function after traumatic brain injury. J. Surg. Res. 170, 253–256. doi: 10.1016/j.jss.2011.03.006
Lucchini, R. G., Dorman, D. C., Elder, A., and Veronesi, B. (2012). Neurological impacts from inhalation of pollutants and the nose-brain connection. Neurotoxicology 33, 838–841. doi: 10.1016/j.neuro.2011.12.001
Malekzadeh, A., de Groot, V., Beckerman, H., van Oosten, B. W., Blankenstein, M. A., and Teunissen, C. (2012). Challenges in multiplex and monoplex platforms for the discovery of inflammatory profiles in neurodegenerative diseases. Methods 56, 508–513. doi: 10.1016/j.ymeth.2012.03.017
Martinez, A. N., Mehra, S., and Kaushal, D. (2013). Role of Interleukin-6 (IL6) in innate immunity to Mycobacterium tuberculosis infection. J. Infect. Dis. 207, 1253–1261. doi: 10.1093/infdis/jit037
Martins, V. R., Beraldo, F. H., Hajj, G. N., Lopes, M. H., Lee, K. S., Prado, M. A., et al. (2010). Prion protein: orchestrating neurotrophic activities. Curr. Issues Mol. Biol. 12, 63–86.
Massimino, M. L., Redaelli, M., Bertoli, A., Sorgato, M. C., and Mucignat-Caretta, C. (2013). Altered behavioral aspects of aged mice lacking the cellular prion protein. Physiol. Behav. 119C, 86–91. doi: 10.1016/j.physbeh.2013.06.006
Milhavet, O., and Lehmann, S. (2002). Oxidative stress and the prion protein in transmissible spongiform encephalopathies. Brain Res. Brain Res. Rev. 38, 328–339. doi: 10.1016/S0165-0173(01)00150-3
Mitsios, N., Saka, M., Krupinski, J., Pennucci, R., Sanfeliu, C., Turu, M. M., et al. (2007). Cellular prion protein is increased in the plasma and peri-infarcted brain tissue after acute stroke. J. Neurosci. Res. 85, 602–611. doi: 10.1002/jnr.21142
Molina, L. T., and Molina, M. J. (2004). Improving air quality in megacities: Mexico City case study. Ann. N.Y. Acad. Sci. 1023, 142–158. doi: 10.1196/annals.1319.006
Mooijaart, S. P., Sattar, N., Trompet, S., Lucke, J., Stott, D. J., Ford, I., et al. (2013). Circulating interleukin-6 concentration and cognitive decline in old age: the PROSPER study. J. Intern. Med. 274, 77–85. doi: 10.1111/joim.12052
Moon, H. Y., Kim, S. H., Yang, Y. R., Song, P., Yu, H. S., Park, H. G., et al. (2012). Macrophage migration inhibitory factor mediates the antidepressant actions of voluntary exercise. Proc. Natl. Acad. Sci. U.S.A. 109, 13094–13099. doi: 10.1073/pnas.1205535109
Nunomura, A., Castellani, R. J., Lee, H. G., Moreira, P. I., Zhu, X., Perry, G., et al. (2006). Neuropathology in Alzheimer's disease: awakening from a hundred-year old dream. Sci. Aging Knowledge Environ. 2006, pe10. doi: 10.1126/sageke.2006.8.pe10
Nunomura, A., Perry, G., Aliev, G., Hirai, K., Takeda, A., Balraj, E. K., et al. (2001). Oxidative damage is the earliest event in Alzheimer's disease. J. Neuropathol. Exp. Neurol. 60, 759–767.
Nunomura, A., Tamaoki, T., Motohashi, N., Nakamura, M., McKeel, D. W. Jr., Tabaton, M., et al. (2012a). The earliest stage of cognitive impairment in transition from normal aging to Alzheimer's disease is marked by prominent RNA oxidation in vulnerable neurons. J. Neuropathol. Exp. Neurol. 71, 233–241. doi: 10.1097/NEN.0b013e318248e614
Nunomura, A., Moreira, P. I., Castellani, R. J., Lee, H. G., Zhu, X., Smith, M. A., et al. (2012b). Oxidative damage to RNA in aging and neurodegenerative disorders. Neurotox. Res. 22, 231–248. doi: 10.1007/s12640-012-9331-x
Nygaard, H. B., and Strittmatter, S. M. (2009). Cellular protein mediates the toxicity of beta-amyloid oligomers: implications for Alzheimer disease. Arch. Neurol. 66, 1325–1328. doi: 10.1001/archneurol.2009.223
Oh, J. M., Choi, E. K., Carp, R. I., and Kim, Y. S. (2012). Oxidative stress impairs autophagic flux in prion protein-deficient hippocampal cells. Autophagy 8, 1448–1461. doi: 10.4161/auto.21164
Ohta, S., Misawa, A., Fukaya, R., Inoue, S., Kanemura, Y., Okano, H., et al. (2012). Macrophage migration inhibitory factor (MIF) promotes cell survival and proliferation of neural stem/progenitor cells. J. Cell Sci. 125, 3210–3220. doi: 10.1242/jcs.102210
Papassotiropoulos, A., Hock, C., and Nitsch, R. M. (2001). Genetics of Interleukin 6: implications for Alzheimer's disease. Neurobiol. Aging 22, 863–871. doi: 10.1016/S0197-4580(01)00294-9
Peters, V. A., Joesting, J. J., and Freund, G. G. (2013). IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain Behav. Immun. 32, 1–8. doi: 10.1016/j.bbi.2012.11.006
Popko, B. (2010). Myelin maintenance: axonal support required. Nat. Neurosci. 13, 275–277. doi: 10.1038/nn0310-275
Popp, J., Bacher, M., Kölsch, H., Noelker, C., Deuster, O., Dodel, R., et al. (2009). Macrophage inhibitory factor in mild cognitive impairment and Alzheimer's disease. J. Psychiatr. Res. 43, 749–753. doi: 10.1016/j.jpsychires.2008.10.006
Pradillo, J. M., Denes, A., Greenhalgh, A. D., Boutin, H., Drake, C., McColl, B. W., et al. (2012). Delayed administration of interleukin-1 receptor antagonist reduces ischemic brain damage and inflammation in comorbid rats. J. Cereb. Blood Flow Metab. 32, 1810–1819. doi: 10.1038/jcbfm.2012.101
Qin, L., Wu, X., Block, M. L., Liu, Y., Breese, G. R., Hong, J. S., et al. (2007). Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 55, 453–462. doi: 10.1002/glia.20467
Ramljak, S., Asif, A. R., Armstrong, V. W., Wrede, A., Groschup, M. H., Buschmann, A., et al. (2008). Physiological role of the cellular prion protein (PrPc): protein profiling study in two cell culture systems. J. Proteome Res. 7, 2681–2695. doi: 10.1021/pr7007187
Rezaie, P., Pontikis, C. C., Hudson, L., Caims, N. J., and Lantos, P. L. (2005). Expression of cellular prion protein in the frontal and occipital lobe in Alzheimer's disease, diffuse Lewy body disease, and in normal brain: an immunohistochemical study. J. Histochem. Cytochem. 53, 929–940. doi: 10.1369/jhc.4A6551.2005
Rivest, S. (2009). Regulation of innate immune responses in the brain. Nat. Rev. Immunol. 9, 429–439. doi: 10.1038/nri2565
Rubio-Perez, J. M., and Morillas-Ruiz, J. M. (2012). A review: inflammatory process in Alzheimer's disease, role of cytokines. ScientificWorldJournal 2012:756357. doi: 10.1100/2012/756357
Saijo, E., Scheff, S. W., and Telling, G. C. (2011). Unaltered prion protein expression in Alzheimer disease patients. Prion 5, 109–116. doi: 10.4161/pri.5.2.16355
Savaskan, N. E., Fingerle-Rowson, G., Buchfelder, M., and Eyüpoglu, I. Y. (2012). Brain miffed by macrophage inhibitory factor. Int. J. Cell Biol. 2012:139573. doi: 10.1155/2012/139573
Scalabrino, G., and Veber, D. (2012). Normal prions as a new target of cobalamin (vitamin B12) in rat central nervous system. Clin. Chem. Lab. Med. 51, 601–606. doi: 10.1515/cclm-2012-0474
Schmidt, C., Artjomova, S., Hoeschel, M., and Zerr, I. (2013). CSF prion protein concentration and cognition in patients with Alzheimer disease. Prion 7, 229–234. doi: 10.4161/pri.23904
Schwarze-Eicker, K., Keyvani, K., Görtz, N., Westaway, D., Sachser, N., and Paulus, W. (2005). Prion protein (PrPc) promotes beta-amyloid plaque formation. Neurobiol. Aging 26, 1177–1182. doi: 10.1016/j.neurobiolaging.2004.10.004
Shameli, A., Yamanouchi, J., Tsai, S., Yang, Y., Clemente-Casares, X., Moore, A., et al. (2013). IL-2 promotes the function of memory-like autoregulatory CD8+T cells but suppresses their development via FoxP3+ Treg cells. Eur. J. Immunol. 43, 394–403. doi: 10.1002/eji.201242845
Smolen, J. S., Schoels, M. M., Nishmoto, N., Breedveld, F. C., Burmester, G. R., Dougados, M., et al. (2013). Consensus statement on blocking the effects of interleukin-6 and in particular by interleukin-6 receptor inhibition in rheumatoid arthritis and other inflammatory conditions. Ann. Rheum. Dis. 72, 482–492. doi: 10.1136/annrheumdis-2012-202469
Spittau, B., Zhou, X., Ming, M., and Krieglstein, K. (2012). IL6 protects MN9D cells and midbrain dopaminergic neurons from MPP+-induced neurodegeneration. Neuromolecular Med. 14, 317–327. doi: 10.1007/s12017-012-8189-7
Stellato, F., Spevacek, A., Proux, O., Minicozzi, V., Millhauser, G., and Morante, S. (2011). Zinc modulates copper coordination mode in prion protein octa-repeats subdomains. Eur. Biophys. J. 40, 1259–1270. doi: 10.1007/s00249-011-0713-4
Tokami, H., Ago, T., Sugimori, H., Kuroda, J., Awano, H., Suzuki, K., et al. (2013). RANTES has a potential to play a neuroprotective role in an autocrine/paracrine manner after ischemic stroke. Brain Res. 1517, 122–132. doi: 10.1016/j.brainres.2013.04.022
Tsai, D. H., Amvai, N., Marques-Vidal, P., Wang, J. L., Riediker, M., Mooser, V., et al. (2012). Effects of particulate matter on inflammatory markers in the general adult population. Part. Fibre Toxicol. 9, 24. doi: 10.1186/1743-8977-9-24
Tsutsui, S., Hahn, J. N., Johnson, T. A., Ali, Z., and Jirik, F. R. (2008). Absence of the cellular prion protein exacerbates and prolongs neuroinflammation in experimental autoimmune encephalomyelitis. Am. J. Pathol. 173, 1029–1041. doi: 10.2353/ajpath.2008.071062
Turtzo, L. C., Li, J., Persky, R., Benashski, S., Weston, G., Bucala, R., et al. (2013). Deletion of macrophage migration inhibitory factor worsens stroke outcome in female mice. Neurobiol. Dis. 54, 421–431. doi: 10.1016/j.nbd.2013.01.016
Um, J. W., Nygaard, H. B., Heiss, J. K., Kostylev, M. A., Stagi, M., Vortmeyer, A., et al. (2012). Alzheimer amyloid-beta oligomer bound to postsynaptic prion protein activates Fyn to impair neurons. Nat. Neurosci. 15, 1227–1235. doi: 10.1038/nn.3178
Uski, O. J., Happo, M. S., Jalava, P. I., Brunner, T., Kelz, J., Obemberger, I., et al. (2012). Acute systemic and lung inflammation in C57BI/6J mice after intratracheal aspiration of particulate matter from small-scale biomass combustion appliances based on old and modern technologies. Inhal. Toxicol. 24, 952–965. doi: 10.3109/08958378.2012.742172
Velayos, J. L., Irujo, A., Cuadrado-Tejedor, M., Paternain, B., Moleres, F. J., and Ferrer, V. (2009). The cellular prion protein and its role in Alzheimer disease. Prion 3, 110–117. doi: 10.4161/pri.3.2.9135
Viles, J. H., Cohen, F. E., Prusiner, S. B., Goodin, D. B., Wright, P. E., and Dyson, H. J. (1999). Copper binding to the prion protein: structural implications of four identical cooperative binding sites. Proc. Natl. Acad. Sci. U.S.A. 96, 2042–2047. doi: 10.1073/pnas.96.5.2042
Voigtländer, T., Klöppel, S., Birner, P., Jarius, C., Flicker, H., Verghese-Nikolakaki, S., et al. (2001). Marked increase of neuronal prion protein immunoreactivity in Alzheimer's disease and human prion diseases. Acta Neuropathol. 101, 417–423.
Wang, J., Wicker, L. S., and Santamaria, P. (2009). IL-2 and its high-affinity receptor: genetic control of immunoregulation and autoimmunity. Semin. Immunol. 21, 363–371. doi: 10.1016/j.smim.2009.04.004
Watt, N. T., Griffiths, H. H., and Hooper, N. M. (2013). Neuronal zinc regulation and the prion protein. Prion 7, 203–208. doi: 10.4161/pri.24503
Westergard, L., Christensen, H. M., and Harris, D. A. (2007). The cellular prion protein (PrPC): its physiological function and role in disease. Biochim. Biophys. Acta 1772, 629–644. doi: 10.1016/j.bbadis.2007.02.011
Whitehouse, I. J., Jackson, C., Turner, A. J., and Hooper, N. M. (2010). Prion protein is reduced in aging and in speoradic but not in familial Alzheimer's disease. J. Alzheimers Dis. 22, 1023–1031.
Whitehouse, I. J., Miners, J. S., Glennon, E. B., Kehoe, P. G., Love, S., Kellet, K. A., et al. (2013). Prion protein is decreased in Alzheimer's brain and inversely correlates with BACE1 activity, amyloid-β levels and Braak stage. PLoS ONE 8:e59554 doi: 10.1371/journal.pone.0059554
Willette, A. A., Bendlin, B. B., McLaren, D. G., Canu, E., Kastman, E. K., Kosmatka, K. J., et al. (2010). Age-related changes in neural volume and microstructure associated with interleukin-6 are ameliorated by a calorie-restricted diet in old rhesus monkeys. Neuroimage 51, 987–994. doi: 10.1016/j.neuroimage.2010.03.015
Wittkopp, S., Staimer, N., Tjoa, T., Gillen, D., Daher, N., Shafer, M., et al. (2013). Mitochondrial genetic background modifies the relationship between traffic-related air pollution exposure and systemic biomarkers of inflammation. PLoS ONE 8:e64444. doi: 10.1371/journal.pone.0064444
Yan, J., Liu, J., Lin, C. Y., Anzgene, A. N., Csurhes, P. A., Pender, M. P., et al. (2012). Interleukin-6 gene promoter-572 C allele may play a role in rate of disease progression in multiple sclerosis. Int. J. Mol. Sci. 13, 13667–13679. doi: 10.3390/ijms131013667
Keywords: Alzheimer, air pollution, children, innate immunity, neurodegeneration, neuroinflammation, particulate matter, prion cellular protein
Citation: Calderón-Garcidueñas L, Cross JV, Franco-Lira M, Aragón-Flores M, Kavanaugh M, Torres-Jardón R, Chao C-k, Thompson C, Chang J, Zhu H and D'Angiulli A (2013) Brain immune interactions and air pollution: macrophage inhibitory factor (MIF), prion cellular protein (PrPC), Interleukin-6 (IL-6), interleukin 1 receptor antagonist (IL-1Ra), and interleukin-2 (IL-2) in cerebrospinal fluid and MIF in serum differentiate urban children exposed to severe vs. low air pollution. Front. Neurosci. 7:183. doi: 10.3389/fnins.2013.00183
Received: 10 July 2013; Accepted: 23 September 2013;
Published online: 10 October 2013.
Edited by:
Adam Denes, University of Manchester, UKReviewed by:
Zsuzsanna Környei, Institute of Experimental Medicine of the Hungarian Academy of Sciences, HungaryMuhammad Sajjad, University of Bonn, Germany
Jesus M. Pradillo, University Complutense of Madrid, Technical University of Madrid, Spain
Copyright © 2013 Calderón-Garcidueñas, Cross, Franco-Lira, Aragón-Flores, Kavanaugh, Torres-Jardón, Chao, Thompson, Chang, Zhu and D'Angiulli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lilian Calderón-Garcidueñas, The Center for Structural and Functional Neurosciences, The University of Montana, 32 Campus Drive, Skaggs Building 287, Missoula, MT 59812, USA e-mail:bGlsaWFuLmNhbGRlcm9uLWdhcmNpZHVlbmFzQHVtb250YW5hLmVkdQ==