 Taishi Kawamoto
Taishi Kawamoto Mitsuhiro Ura
Mitsuhiro Ura Hiroshi Nittono
Hiroshi Nittono- 1Japan Society for the Promotion of Science, Tokyo, Japan
- 2Faculty of Integrated Arts and Sciences, Hiroshima University, Higashi-Hiroshima, Japan
- 3Department of Psychology, Otemon-Gakuin University, Ibaraki, Japan
People have a fundamental need to belong with others. Social exclusion impairs this need and has various effects on cognition, affect, and the behavior of excluded individuals. We have previously reported that activity in the dorsal anterior cingulate cortex (dACC) and right ventrolateral prefrontal cortex (rVLPFC) could be a neurocognitive index of social exclusion (Kawamoto et al., 2012). In this article, we provide an integrative framework for understanding occurrences during and after social exclusion, by reviewing neuroimaging, electrophysiological, and behavioral studies of dACC and rVLPFC, within the framework of intrapersonal and interpersonal processes of social exclusion. As a result, we have indicated directions for future studies to further clarify the phenomenon of social exclusion from the following perspectives: (1) constructional elements of social exclusion, (2) detection sensitivity and interpretation bias in social exclusion, (3) development of new methods to assess the reactivity to social exclusion, and (4) sources of social exclusion.
Introduction
Humans are abundantly social animals, and have a fundamental need to belong (Baumeister and Leary, 1995). Maintaining good and lasting relationships with others is therefore quite vital for us (Baumeister and Leary, 1995; Macdonald and Leary, 2005). Social exclusion breaks the relationships we have with others and influences our physical and mental health in a wide range of ways. For example, previous studies have revealed that social exclusion possibly promotes suicide (Van Orden and Joiner, 2013), increases depression (Nolan et al., 2003), and even decreases survival rates in mammals generally (Kling et al., 1970; Silk et al., 2003) and humans specifically (Holt-Lunstad et al., 2010). Social exclusion also causes aggression in humans (e.g., Twenge et al., 2001; Warburtona et al., 2006; Gaertner et al., 2008; DeWall et al., 2009b; Wesselmann et al., 2010) and is thought to be one of the causes of school shootings (Leary et al., 2003). Furthermore, social exclusion is not rare or unusual in daily life events (Nezlek et al., 2012), and has the potential to occur in a variety of places including text messages on cell phones (Smith and Williams, 2004) and on Facebook (Tobin et al., 2015).
KEY CONCEPT 1. Social exclusion
In this article, we refer to social exclusion as events and situations that signal a lack of social connections with others. Therefore, this article includes studies referring to ostracism, devaluation, and social rejection.
In light of the severe and wide ranging effects of social exclusion on the psychological adaptation of individuals, it is important to identify neural correlates, cognitions, and behaviors that occur during and after social exclusion. We have previously investigated the intrapersonal process of social exclusion by using functional magnetic resonance imaging (fMRI), and identified neural correlates of social exclusion by comparing social exclusion and expectancy violation. As reported in Frontiers in Evolutionary Neuroscience (Kawamoto et al., 2012), we identified dorsal anterior cingulate cortex (dACC) and right ventrolateral prefrontal cortex (rVLPFC) activities as a possible neurocognitive index of social exclusion. In this focused review article, we have reviewed studies on the involvement of these two brain regions in intrapersonal and interpersonal processes of social exclusion. We provide an integrative framework for understanding intrapersonal and interpersonal processes of social exclusion (Figure 1), and review the literature of social exclusion using neuroimaging, electrophysiological, and behavioral methods within this framework. First, we review three stages—detection, appraisal, and regulation—which occur during social exclusion. Second, we review how social exclusion changes perception, attention, and cognition in response to social cues (social monitoring system: SMS). Third, we review how people behave following social exclusion and the way they recover from the aversive impact of social exclusion (interpersonal processes). Fourth, we review how intrapersonal and interpersonal processes of social exclusion are stored as experiences, and how these influence responses to future social exclusion. Finally, we suggest some directions for future studies that we believe are needed for a better understanding of social exclusion.
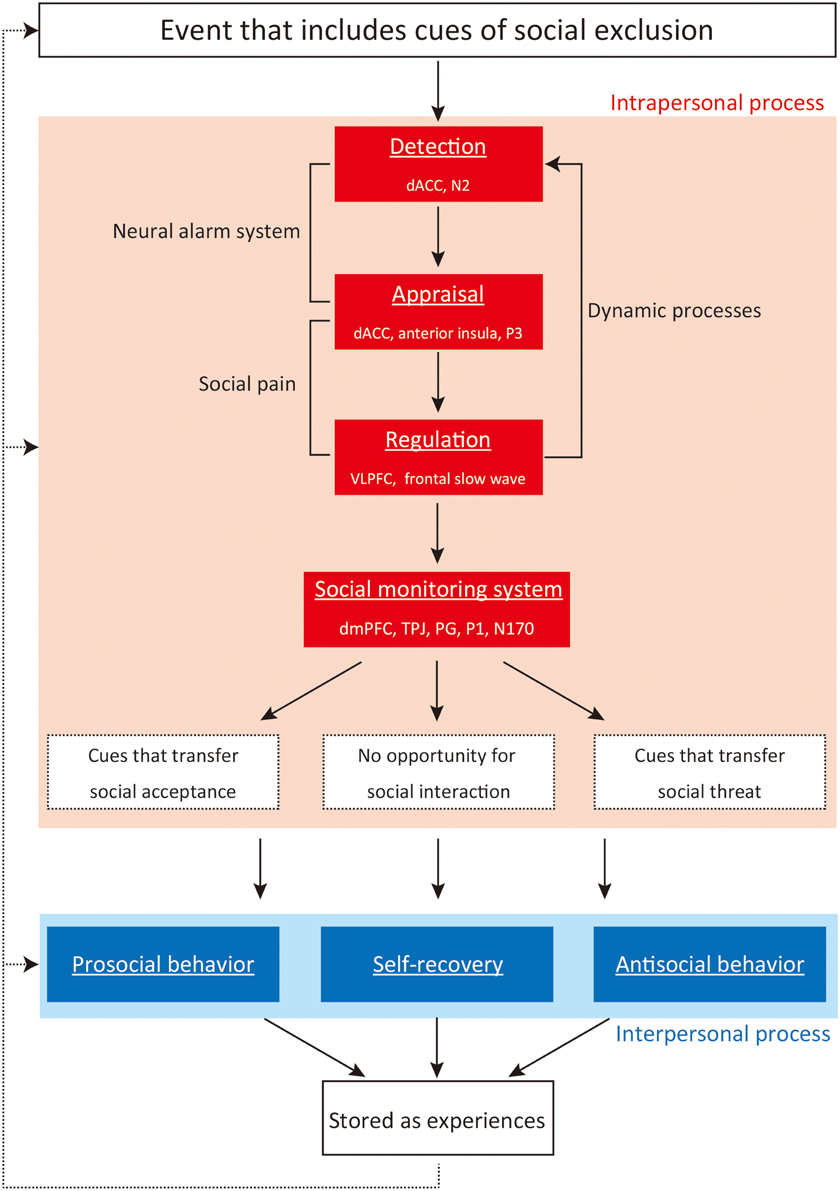
Figure 1. Intrapersonal and interpersonal processes of social exclusion. Note: dACC, dorsal anterior cingulate cortex; VLPFC, ventrolateral prefrontal cortex; dmPFC, dorsomedial prefrontal cortex; TPJ, temporoparietal junction; PG, precentral gyrus.
Intrapersonal Processes During Social Exclusion: Detection, Appraisal, and Regulation
Table 1 summarizes the key studies that have investigated the role of intrapersonal processes of social exclusion. People can detect social exclusion in any situation (Williams, 2009; Wesselmann et al., 2012a). Previous research using “Cyberball” software has revealed that we feel excluded even if our excluders are online (Williams et al., 2000), computer-generated opponents (Zadro et al., 2004; Kawamoto et al., 2013b), or disliked others (Gonsalkorale and Williams, 2007). In the Cyberball paradigm, participants play a simple ball-tossing game with two or three opponents through the computer screen. In the exclusion condition, participants experience social exclusion by receiving few ball tosses from others. Individuals feel social pain during social exclusion (Eisenberger et al., 2003). Prior research has investigated the neural correlates of detection, appraisal, and regulation processes of social exclusion. Three brain regions—dACC, anterior insula (AI), and VLPFC—play a particularly important role in cognitive and affective processing during social exclusion. In a highly influential study, Eisenberger et al. (2003) conducted an fMRI experiment to identify the brain regions associated with processing social exclusion during Cyberball play. They found that the dACC, AI, and right VLPFC were more activated during the social exclusion condition relative to the fair play condition (i.e., when participants caught the ball equally as often as the other players). In addition, dACC activation was positively correlated with the amount of social pain participants felt during social exclusion, whereas right VLPFC activation showed the opposite pattern.

Table 1. Key studies on intrapersonal processes of social exclusion.
KEY CONCEPT 2. Social pain
The distressing experiences arising from the perception of actual or potential psychological distance from close others, or social groups (Eisenberger and Lieberman, 2004, p. 294). We often use the term “painful” when we are referring to social exclusion and social exclusion is known to activate brain regions similar to those that are activated during physical pain (e.g., dACC).
We often use the term “painful” even when we are referring to non-physical injuries. In fact, the use of the words pain and painful to describe when individuals are being excluded from partner or group relationships is common across many different languages (Macdonald and Leary, 2005). Social pain has been defined as “the distressing experience arising from the perception of actual or potential psychological distance from close others or a social group (Eisenberger and Lieberman, 2004, p. 294).” Social pain is more than just a metaphor as it shares common neural correlates with physical pain (Eisenberger and Lieberman, 2004; Macdonald and Leary, 2005; Eisenberger, 2012a,b, 2015). In fact, social exclusion activates the similar brain regions that have been associated with appraisal—the dACC and AI—and regulation—the VLPFC—of the unpleasantness of physical pain (e.g., Eisenberger et al., 2003; Eisenberger and Lieberman, 2004; Eisenberger, 2012a,b, 2015). In addition, previous studies have revealed the parallel nature of increasing physical and social pain (DeWall and Baumeister, 2006; Eisenberger et al., 2006), further supporting their commonality. Thus, both the dACC and Al seem to be related to appraisal of social exclusion whereas the VLPFC seems to be related to regulation of social pain evoked by social exclusion.
Although dACC involvement in processing social exclusion has been replicated by multiple studies (e.g., Eisenberger et al., 2007, 2011; Onoda et al., 2009, 2010; DeWall et al., 2012), social exclusion is a complex phenomena which inherently includes multiple other components such as expectancy violation Previous studies have been challenged to differentiate social exclusion and mere expectancy violation (Somerville et al., 2006; Bolling et al., 2011b; Kawamoto et al., 2012). Our group compared brain activation during social exclusion and during over-inclusion—participants received many throws—using Cyberball (Kawamoto et al., 2012). We found that the dACC was more activated for exclusion-related events than over-inclusion-related events, but failed to show any relation between the dACC and self-rated social pain. On the other hand, rVLPFC was negatively correlated with social pain, supporting the notion of the regulatory function of rVLPFC on social pain (Eisenberger et al., 2003). Note that we did not observe any significant relation between the dACC and self-reported expectancy violation scores (e.g., surprise feeling), suggesting that dACC activation in response to social exclusion is not merely due to expectancy violation. Other studies have found a role of the dACC in expectancy violation (Somerville et al., 2006), and both social exclusion and expectancy violation (Bolling et al., 2011b). How do we interpret this seemingly inconsistent evidence? One possible explanation is to conceptualize dACC function in social exclusion as a “neural alarm system” (Eisenberger and Lieberman, 2004). Eisenberger and Lieberman argued that detection and appraisal processes involved in the dACC were complementary processes underlying the function of the neural alarm system. Therefore, the dACC plays an important role in both detection and appraisal of social exclusion.
KEY CONCEPT 3. Expectancy violation
A situation when individuals' expectations are violated. In exclusion studies using Cyberball, participants are often included before being excluded, resulting in expectations of social inclusion during social exclusion. Thus, some research has argued that responses to social exclusion not only include painful experiences but also cognitive conflict that comes from expectancy violation.
KEY CONCEPT 4. Neural alarm system
Neural systems that signal relational threats. This system was proposed in order to explain the role of the dACC in response to social exclusion. Two sub-systems are considered to be needed for adequate operation of the neural alarm system. The first is a discrepancy monitoring system, which serves to detect deviations from desired standards. The second is a sounding mechanism that signals a problem that needs to be addressed. The discrepancy detection function is considered to be associated with the detection of social exclusion, whereas social pain is thought to be the product of the sounding system.
However, results of meta-analyses have been inconclusive. One recent meta-analysis of fMRI studies related to social exclusion showed the involvement of dACC in processing information related to social exclusion (Rotge et al., 2015), whereas another meta-analysis failed to show an association between social exclusion and dACC (Cacioppo et al., 2013). These contrary findings suggest that neural correlates of social exclusion might be complex (Cacioppo et al., 2013), and need further investigation focusing on the duration and temporal dynamics of social exclusion (Rotge et al., 2015). Supporting this notion, recent electrophysiological and neuroimaging studies have suggested that intrapersonal processes of social exclusion dynamically change with time, and occur not only in whole exclusionary situations but also during specific events within social interactions (e.g., Crowley et al., 2009, 2010; Kawamoto et al., 2012, 2013b; Moor et al., 2012; Themanson et al., 2013). For example, prior findings using event-related brain potentials (ERPs) indicated that N2, P3b, and frontal slow wave in response to each exclusionary cue (i.e., throws among opponents in Cyberball) were closely related to the detection, appraisal, and regulation processes of social exclusion, respectively (Crowley et al., 2009, 2010; Kawamoto et al., 2013b; Themanson et al., 2013). In addition, our group found that attention decreased with time, negative affect accumulated with time, and motivation shifted to a withdrawal pattern during social exclusion (Kawamoto et al., 2013b). Given the previous findings, it is possible that individuals repeatedly process each exclusionary cue, and additive and complex intrapersonal processes determine the final emotional response (e.g., social pain). Thus, social exclusion is an uncertain and complex circumstance in which the excluded individual interprets the situation and source of threat (i.e., the excluder) in dramatically different ways over time. Future research would benefit from investigating temporal change during social exclusion for a better understanding of psychological and neural correlates during social exclusion.
Intrapersonal Processes Following Social Exclusion: Social Monitoring System
Once people are excluded by others, they activate an outer monitoring system, called the “social monitoring system (SMS)” (Pickett and Gardner, 2005). According to the conceptualization of SMS, this system enhances perceptive and cognitive responses to social cues and social information (e.g., facial expressions and vocal tone). SMS is considered an adaptive system that attunes excluded people to information that will help them navigate the social environment more successfully (e.g., Pickett et al., 2004; Gardner et al., 2005a,b; Pickett and Gardner, 2005). Supporting this supposition, prior studies have revealed that social exclusion causes increased attention and perception in response to social information (e.g., Gardner et al., 2000; Pickett et al., 2004; Bernstein et al., 2008; DeWall et al., 2009a). Although relatively few studies have focused on neural correlates of the SMS, prior studies have shown that the brain regions related to mentalizing and mirror neuron networks—the dorsomedial prefrontal cortex (dmPFC), temporoparietal junction (TPJ), and precentral gyrus (PG)—and ERP components related to attention and facial encoding—P1 and N170—might be involved in the SMS (e.g., Moor et al., 2012; Powers et al., 2013; Beyer et al., 2014; Kawamoto et al., 2014; Will et al., 2015). Future studies should strive toward a deeper understanding by further clarifying the neural correlates of the SMS.
KEY CONCEPT 5. Mentalizing
The ability to extract and understand others' thoughts and beliefs.
Interpersonal Processes Following Social Exclusion: Behaviors and Self-Recovery
Table 2 summarizes the key studies that have investigated the role of interpersonal processes of social exclusion. Social exclusion not only shifts an individual's cognitive responses but also influences him to change his behaviors in an attempt to regain social acceptance and/or to avoid further social exclusion. Previous findings have indicated that social exclusion causes both prosocial and antisocial behaviors (e.g., Twenge et al., 2001; Warburtona et al., 2006; Maner et al., 2007; Chow et al., 2008; DeWall et al., 2009b; Wesselmann et al., 2010; DeWall and Bushman, 2011; DeWall and Richman, 2011; Kawamoto et al., 2013a, 2014). Social reconnection theory proposes that excluded people behave prosocially only in the presence of cues that promise transfer of social affiliation (Maner et al., 2007). Supporting this notion, prior studies have revealed that people behave prosocially following social exclusion even when they experience cues that indicate only a remote possibility of social affiliation (e.g., Maner et al., 2007; Lakin et al., 2008; Kawamoto et al., 2014). For example, prior findings have indicated that excluded individuals show heightened willingness to make new friends via student services (Maner et al., 2007), increased behavioral mimicry of a new interaction partner (Lakin et al., 2008), and enhanced facial mimicry in response to pictures of strangers' smiles (Kawamoto et al., 2014). In addition, social exclusion induces increased progesterone production—a hormone that reflects an individual's motivation to affiliate (Frye et al., 2000)—when excluded individuals anticipated interaction with a new group (Maner et al., 2010). Thus, excluded people behave prosocially if there are cues of social acceptance—typically toward those who are not involved in the social exclusion.
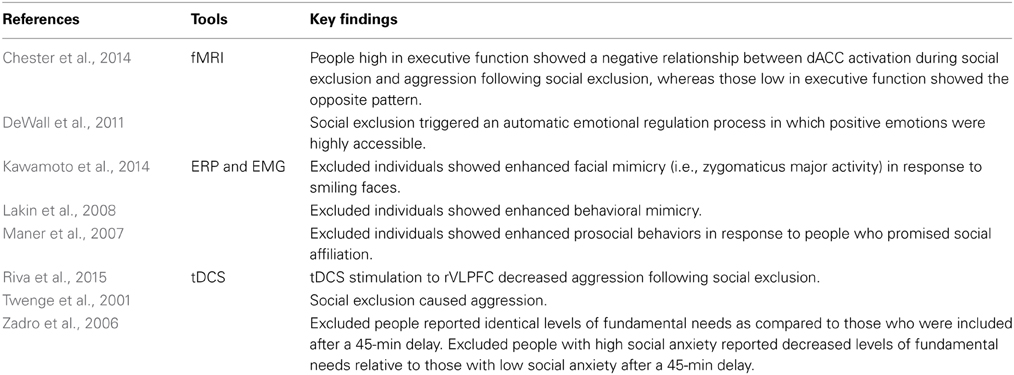
Table 2. Key studies on interpersonal processes of social exclusion.
KEY CONCEPT 6. Prosocial behavior
Behaviors performed with the intention of helping others and being included by others. Social exclusion studies deal with prosocial behavior across a wide range, such as willingness to participate in a group, voting, relational evaluation, and behavioral mimicry.
KEY CONCEPT 7. Antisocial behavior
Behaviors performed with the intention of harming others. Social exclusion studies typically refer to antisocial behavior as aggression. Aggression is measured by highly valid methods such as the volume of loud white noise and amount of hot spicy sauce given to others.
In contrast, excluded individuals tend to behave antisocially toward their excluder. Previous studies have revealed that social exclusion causes aggression toward the excluder (e.g., Buckley et al., 2004; Chow et al., 2008; Kawamoto et al., 2013a). Thus, excluded individuals seem to behave aggressively rather than prosocially toward others that convey cues of social threat. Furthermore, excluded individuals lash out at others those who are not involved in social exclusion (e.g., Gaertner et al., 2008; DeWall et al., 2009b; Wesselmann et al., 2010). How can we buffer the influence of social exclusion on aggressive behavior? Previous research has sought after variables that moderate the link between social exclusion and aggression. For instance, a previous study indicated that individual differences in executive function—cognitive ability that regulates goal oriented behaviors—and dACC activity in response to social exclusion interacted to predict the relation between social exclusion and aggressive behavior (Chester et al., 2014). This study indicated that people with better executive function are less aggressive when they feel social pain (i.e., greater dACC and AI activity), whereas those with poor executive function are more aggressive. In addition, recent studies have revealed that stimulation of the right VLPFC by a transcranical direct current (tDCS) during social exclusion reduced social pain (Riva et al., 2012), and aggressive behavior following social exclusion (Riva et al., 2015). Thus, trait and situational behavioral and emotional regulation seem to play a key role in reducing aggressive behavior following social exclusion.
Excluded people seem to be able to recover from the aversive impact of social exclusion without any direct interaction with others (e.g., prosocial or antisocial behavior toward others). A prior study found that after a 45 min delay, socially excluded participants recovered their primary needs, including the sense of self-esteem, belonging, control, and meaningful existence (Zadro et al., 2006). This finding implies that people have a self-recovery system that can effectively buffer against the aversive influences of social exclusion. One possible mechanism of this system is that excluded people use an inner representation of social connection (e.g., memories of their family, mental images of favorite characters from novels). According to the belonging regulation model, people use indirect strategies to regain a sense of social connection when they feel unconnected (Gardner et al., 2005b). Supporting this notion, previous studies have implied that any entity with which an individual can feel a social connection—a god, comfort food, or a favorite television character—could diminish the aversive impacts of social exclusion (e.g., Derrick et al., 2009; Troisi and Gabriel, 2011; Laurin et al., 2014). In addition, excluded individuals set in motion an automatic emotion regulation process in which positive emotions become highly accessible, which relates to positive mental health (DeWall et al., 2011). Thus, excluded individuals seem to deal flexibly with their mental representations and social environment to regulate and regain the feeling of social connection.
Social Exclusion as Stored Experience: From the Perspective of Individual Differences
Table 3 summarizes the key studies that have investigated individual differences in intrapersonal and interpersonal processes of social exclusion. There are individual differences in both intrapersonal and interpersonal processes of social exclusion (e.g., Kross et al., 2007; Maner et al., 2007, 2010; Masten et al., 2009; Onoda et al., 2010; Yanagisawa et al., 2011a,b; DeWall et al., 2012; Nakashima et al., 2013; Chester et al., 2014, 2015). For instance, prior findings have indicated that people with low self-esteem have more self-reported social pain and show increased dACC activity in response to social exclusion as compared to those with high self-esteem (Onoda et al., 2010). In addition, people with high trait rejection sensitivity and low childhood socioeconomic status exhibited reduced VLPFC activity in response to social exclusion (Kross et al., 2007; Yanagisawa et al., 2013). How are these individual differences in response to social exclusion developed?
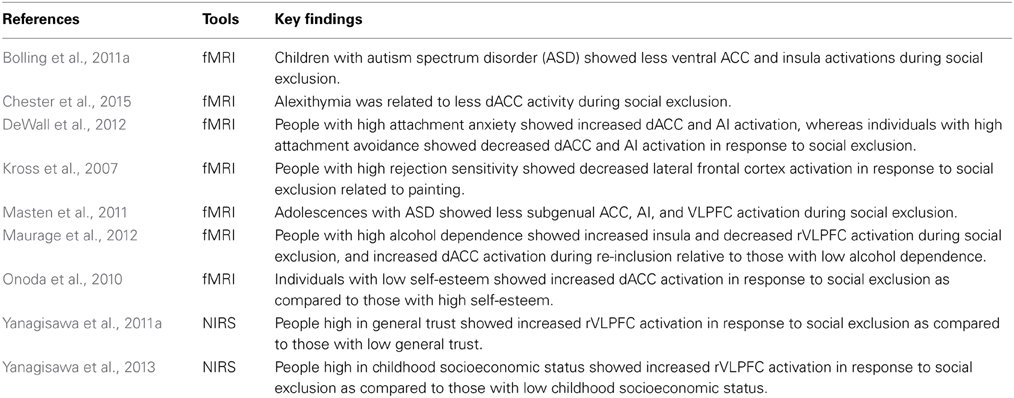
Table 3. Key studies on individual differences of intrapersonal and interpersonal processes of social exclusion.
According to optimal calibration theory, early life history shifts one's neural responses to social exclusion (Chester et al., 2012). Specifically, Chester et al. argued that unpredictable social rejection in early life history causes a “hyper-activated” social pain system (i.e., the dACC and AI) whereas chronic social rejection results in a “deactivated” social pain system. Supporting this notion, a prior study indicated that people with high attachment-anxiety—reflecting higher unpredictable social rejection—showed increases in both dACC and AI activity whereas people with high attachment-avoidance—reflecting higher chronic social rejection—showed decreases in dACC and AI activity in response to social exclusion (DeWall et al., 2012). In addition, a recent study revealed that having a history of chronic victimization increased the magnitude of cardiovascular blunting in response to social exclusion (Newman, 2014), further supporting the notion that past social exclusion experiences carry over to influence responses to future social exclusion. Thus, past social exclusion experiences are possibly stored with individuals' experiences in memory, resulting in an effect on their responses to future social exclusion.
Studies have also been conducted that have investigated intrapersonal and interpersonal processes of social exclusion in clinical populations (e.g., Sebastian et al., 2009; Bolling et al., 2011a; Masten et al., 2011; McPartland et al., 2011; Maurage et al., 2012; Domsalla et al., 2014). One such study for example, has indicated that highly alcohol dependent people have increased insula and decreased rVLPFC activation during social exclusion relative to less alcohol dependent people (Maurage et al., 2012). In addition, highly alcohol dependent people showed increased dACC activation during re-inclusion—being included after being excluded—compared to less alcohol dependent people. Another study that was designed to clarify the mechanisms of social phobia by manipulating attentional bias for threatening faces (Heeren et al., 2012) indicated that the induction of attentional bias for threatening faces resulted in increased anxiety during social exclusion. These findings imply that investigating intrapersonal and interpersonal processes of social exclusion in clinical populations could contribute to understanding the mechanisms of clinical pathologies and to develop effective intervention methods for such conditions.
Conclusion and Future Directions
Over the past few decades, social exclusion has received a lot of attention especially in the social psychology and social neuroscience disciplines. Given that social exclusion is critically vital for humans, it is not surprising that social exclusion affects a wide range of our perceptions, cognitions, affect, behaviors, and psychological adaptations. Although prior studies have revealed a great deal about the intrapersonal and interpersonal processes of social exclusion, some aspects need further clarification. To conclude, we describe four directions for future research, which we believe is worthwhile investigating.
Firstly, future research should explore the constructional elements of social exclusion in more detail, since social exclusion is a complex phenomenon (e.g., Smart Richman and Leary, 2009; Williams, 2009; Cacioppo et al., 2013). In fact, peoples' perceptions, cognitions, emotions, and motivations are changed over time by social exclusion (e.g., Moor et al., 2012; Wesselmann et al., 2012b; Kawamoto et al., 2013b; Themanson et al., 2013). In addition, social exclusion has been studied in several forms, such as participants being ostracized on the computer screen (Cyberball), being told that they would end up alone (future life manipulation), being told that they were not chosen by an experimental partner (get-acquainted task), or evaluating feedback about preference from peers (rejection paradigm). Although all prior manipulations have involved threats to a sense of relational value and need for belonging (Smart Richman and Leary, 2009), differences in the manipulations should be investigated further, as recent studies and reviews have argued they may play a role in the studies' outcomes (e.g., Blackhart et al., 2009; Gerber and Wheeler, 2009; Bernstein and Claypool, 2012). For instance, a recent study found that future life and Cyberball manipulations differed in severity, with one resulting in an increase and the other a decrease in pain tolerance/threshold (Bernstein and Claypool, 2012). More specifically, the authors argued that feedback indicating a future alone was a bigger threat than exclusion in Cyberball, which resulted in decreasing pain sensitivity (numbing) whereas exclusion in Cyberball resulted in increasing pain sensitivity (hypersensitivity). In addition, a previous study indicated that exclusion from a self-resembling in-group resulted in higher dACC activity relative to exclusion from an out-group (Krill and Platek, 2009). Finally, merely being accepted by one other person during social exclusion could reduce aggressive behavior following social exclusion (DeWall et al., 2010b). Thus, intrapersonal and interpersonal processes of social exclusion could be influenced by the severity, source, and situation of social exclusion.
Secondly, future research should study the boundary and interpretation of social exclusion. Previous research has revealed that people are adept at detecting the slightest hint of social exclusion (e.g., Williams et al., 2000; Smith and Williams, 2004; Zadro et al., 2004; Gonsalkorale and Williams, 2007; Kross et al., 2007; Williams, 2009; Wirth et al., 2010; Van Beest et al., 2011), since social exclusion is quite vital for survival and reproduction (Williams, 2009; Wesselmann et al., 2012a). While it seems that accurately detecting any potential cues of social exclusion would be better than missing the cues completely, excessive misinterpretation of non-exclusive social cues would result in maladaptive outcomes. For instance, people high in trait rejection sensitivity—defined as anticipatory anxiety about, a readiness to perceive, and behavioral overreactions to social rejection—tend to interpret ambiguous situations as social rejection (Downey and Feldman, 1996), and show heightened psychological difficulties such as depression and aggression (e.g., Downey and Feldman, 1996; Downey et al., 1998, 2000; Ayduk et al., 1999, 2001; Downey and Romero-Cayas, 2005; Harper et al., 2006; Romero-Canyas et al., 2010). Thus, accurately detecting social exclusion and excessive misinterpretation of social exclusion seem to have different effects on individuals' psychological adaptation.
Signal detection theory (SDT: Green and Swets, 1966; Lynn and Barrett, 2014) would provide a useful framework to investigate both detection sensitivity and interpretation bias of social exclusion. In this framework, two indexes—sensitivity (d′) and response criterion (β)—are calculated independently. Sensitivity reflects the subjects' ability to discriminate noise and signal whereas response criterion reflects the subjects' overall tendency to respond to the signal, independent of whether the actual stimulus is a signal. According to both the evolutionary perspective and rejection sensitivity studies, it is predicted that having a higher detection sensitivity to social exclusion would be related to positive psychological outcomes whereas having a liberal response criterion (e.g., being prone to interpret social cues as social exclusion) is maladaptive. In addition, SDT may benefit from investigation of the pharmacological effect of acesodyne on interpersonal processes of social exclusion. Prior studies have indicated that specific analgesics—acetaminophen and marijuana—can reduce social pain (DeWall et al., 2010a; Deckman et al., 2014). SDT may promote an understanding of the pharmacological effect of acesodyne on excluded individuals, such as whether it influences an individual's sensitivity or response criterion to social exclusion. These studies would be beneficial for developing effective intervention methods.
KEY CONCEPT 8. Signal detection theory
Psychophysical theory that explains the circumstances needed to distinguish signal from noise. In the framework of SDT, two parameters—sensitivity and response criterion—are estimated independently. SDT has been applied across a wide range of studies including memory, facial recognition, pain, and aggression.
Thirdly, in this article we focused mainly on specific responses to social exclusion (e.g., social pain, prosocial behavior, antisocial behavior), however, there is also a need to develop new methods to assess the reactivity to social exclusion, in order to better understanding abnormal responses of clinical populations. For example, a recent study focused on how acoustic responses (i.e., vibrations of the vocal folds during phonation and speech) following social exclusion are modulated by social anxiety (Gilboa-Schechtman et al., 2014). They found that individuals high in social anxiety showed decreased vocal confidence when reading command sentences following social exclusion, whereas those low in social anxiety showed the opposite pattern. Reactions that match specific psychopathologies would help us to better understand the mechanisms of such pathologies, as well as to develop effective interventions.
Finally, it is important to focus on not only targets of social exclusion, but also on the sources of social exclusion. There is a need to investigate the nature and the consequences of the act of excluding, in order to better understand social exclusion (Zadro and Gonsalkorale, 2014). In this focused review, we have mainly focused on targets of social exclusion, and positioned social exclusion at the top of our model. Although some studies have investigated the nature and consequences of the act of excluding (e.g., Poulsen and Kashy, 2012; Bastian et al., 2013; Legate et al., 2013), most social exclusion studies have focused only on the targets of exclusion. Examining the nature and consequence of sources of social exclusion, as well as the interplay between targets and sources would not only help to better understand the nature of social exclusion, but would also have important implications for understanding intimate relationships.
In conclusion, we have provided an integrative framework of the intrapersonal and interpersonal processes of social exclusion, and summarized the findings of prior studies that have made important contributions to understanding what happens during and after social exclusion. We hope that our review and framework provide an effective approach for further understanding the effects of social exclusion on intrapersonal and interpersonal processes.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by a Grant-in-Aid for JSPS Fellows (12J05780) from the Japan Society for Promotion of Science to the first author. This work was also supported by a Grant-in-Aid for Scientific Research (B) 90231183 from the Japan Society for Promotion of Science to the second author.
Author Biography

References
Ayduk, O., Downey, G., and Kim, M. (2001). Rejection sensitivity and depressive symptoms in women. Pers. Soc. Psychol. Bull. 27, 868–878. doi: 10.1177/0146167201277009
Ayduk, O., Downey, G., Testa, A., Yen, Y., and Shoda, Y. (1999). Does rejection elicit hostility in rejection sensitive women? Soc. Cogn. 17, 245–271. doi: 10.1521/soco.1999.17.2.245
Bastian, B., Jetten, J., Chen, H., Radke, H. R. M., Harding, J. F., and Fasoli, F. (2013). Losing our humanity: the self-dehumanizing consequences of social ostracism. Pers. Soc. Psychol. Bull. 19, 156–169. doi: 10.1177/0146167212471205
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Baumeister, R. F., and Leary, M. R. (1995). The need to belong: desire for interpersonal attachments as a fundamental human motivation. Psychol. Bull. 117, 497–529. doi: 10.1037//0033-2909.117.3.497
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bernstein, M. J., and Claypool, H. M. (2012). Social exclusion and pain sensitivity: why exclusion sometimes hurts and sometimes numbs. Pers. Soc. Psychol. Bull. 38, 185–196. doi: 10.1177/0146167211422449
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bernstein, M. J., Young, S. G., Brown, C. M., Sacco, D. F., and Claypool, H. M. (2008). Adaptive responses to social exclusion: social rejection improves detection of real and fake smiles. Psychol. Sci. 19, 981–983. doi: 10.1111/j.1467-9280.2008.02187.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Beyer, F., Munte, T. F., and Kramer, U. M. (2014). Increased neural reactivity to socio-emotional stimuli links social exclusion and aggression. Biol. Psychol. 96, 102–110. doi: 10.1016/j.biopsycho.2013.12.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Blackhart, G. C., Nelson, B. C., Knowles, M. L., and Baumeister, R. F. (2009). Rejection elicits emotional reactions but neither causes immediate distress nor lowers self-esteem: a meta-analytic review of 192 studies on social exclusion. Pers. Soc. Psychol. Rev. 13, 269–309. doi: 10.1177/1088868309346065
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bolling, D. Z., Pitskel, N. B., Deen, B., Crowley, M. J., McPartland, J. C., Kaiser, M. D., et al. (2011a). Enhanced neural responses to rule violation in children with autism: a comparison to social exclusion. Dev. Cogn. Neurosci. 1, 280–294. doi: 10.1016/j.dcn.2011.02.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bolling, D. Z., Pitskel, N. B., Deen, B., Crowley, M. J., Mcpartland, J. C., Mayes, L. C., et al. (2011b). Dissociable brain mechanisms for processing social exclusion and rule violation. Neuroimage 54, 2462–2471. doi: 10.1016/j.neuroimage.2010.10.049
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Buckley, E., Winkel, R., and Lery, M. R. (2004). Reactions to acceptance and rejection: effects of level and sequence of relational evaluation. J. Exp. Soc. Psychol. 40, 14–28. doi: 10.1016/S0022-1031(03)00064-7
Cacioppo, S., Frum, C., Asp, E., Weiss, R. M., Lewis, J. W., and Cacioppo, J. T. (2013). A quantitative meta-analysis of functional imaging studies of social rejection. Sci. Rep. 3:2027. doi: 10.1038/srep02027
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chester, D. S., Eisenberger, N. I., Pond, R. S. Jr., Richman, S. B., Bushman, B. J., and Dewall, C. N. (2014). The interactive effect of social pain and executive functioning on aggression: an fMRI experiment. Soc. Cogn. Affect. Neurosci. 9, 699–704. doi: 10.1093/scan/nst038
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chester, D. S., Pond, R. S., and DeWall, C. N. (2015). Alexithymia is associated with blunted anterior cingulate response to social rejection: Implications for daily rejection. Soc. Cogn. Affect. Neurosci. doi: 10.1093/scan/nsu082. [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chester, D. S., Pond, R. S. Jr., Richman, S. B., and Dewall, C. N. (2012). The optimal calibration hypothesis: how life history modulates the brain's social pain network. Front. Evol. Neurosci. 4:10. doi: 10.3389/fnevo.2012.00010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chow, R. M., Tiedens, L. Z., and Govan, C. L. (2008). Excluded emotions: the role of anger in antisocial responses to ostracism. J. Exp. Soc. Psychol. 44, 896–903. doi: 10.1037/a0029521
Crowley, M. J., Wu, J., Mccarty, E. R., David, D. H., Bailey, C. A., and Mayes, L. C. (2009). Exclusion and micro-rejection: event-related potential response predicts mitigated distress. Neuroreport 20, 1518–1522. doi: 10.1097/WNR.0b013e328330377a
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Crowley, M. J., Wu, J., Molfese, P. J., and Mayes, L. C. (2010). Social exclusion in middle childhood: rejection events, slow-wave neural activity, and ostracism distress. Soc. Neurosci. 5, 483–495. doi: 10.1080/17470919.2010.500169
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Deckman, T., DeWall, N. C., Way, B., Gilman, R., and Richman, S. (2014). Can marijuana reduce social pain? Soc. Psychol. Pers. Sci. 5, 131–139. doi: 10.1177/1948550613488949
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Derrick, J. L., Gabriel, S., and Hugenberg, K. (2009). Social surrogacy: how favored television programs provide the experience of belonging. J. Exp. Soc. Psychol. 45, 352–362. doi: 10.1016/j.jesp.2008.12.003
DeWall, C. N., and Baumeister, R. F. (2006). Alone but feeling no pain: effects of social exclusion on physical pain tolerance and pain threshold, affective forecasting, and interpersonal empathy. J. Pers. Soc. Psychol. 91, 1–15. doi: 10.1037/0022-3514.91.1.1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., and Bushman, B. J. (2011). Social acceptance and rejection: the sweet and the bitter. Curr. Dir. Psychol. Sci. 20, 256–260. doi: 10.1177/0963721411417545
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., Macdonald, G., Webster, G. D., Masten, C. L., Baumeister, R. F., Powell, C., et al. (2010a). Acetaminophen reduces social pain: behavioral and neural evidence. Psychol. Sci. 21, 931–937. doi: 10.1177/0956797610374741
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., Maner, J. K., and Rouby, D. A. (2009a). Social exclusion and early-stage interpersonal perception: selective attention to signs of acceptance. J. Pers. Soc. Psychol. 96, 729–741. doi: 10.1037/a0014634
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., Masten, C. L., Powell, C., Combs, D., Schurtz, D. R., and Eisenberger, N. I. (2012). Do neural responses to rejection depend on attachment style? An fMRI study. Soc. Cogn. Affect. Neurosci. 7, 184–192. doi: 10.1093/scan/nsq107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., and Richman, S. B. (2011). Social exclusion and the desire to reconnect. Soc. Pers. Psychol. Compass 5, 919–932. doi: 10.1111/j.1751-9004.2011.00383.x
DeWall, C. N., Twenge, J. M., Bushman, B., Im, C., and Williams, C. D. (2010b). A little acceptance goes a long way: applying social impact theory to the rejection-aggression link. Soc. Psychol. Pers. Sci. 1, 168–174. doi: 10.1177/1948550610361387
DeWall, C. N., Twenge, J. M., Gitter, S. A., and Baumeister, R. F. (2009b). It's the thought that counts: the role of hostile cognition in shaping aggressive responses to social exclusion. J. Pers. Soc. Psychol. 96, 45–59. doi: 10.1037/a0013196
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
DeWall, C. N., Twenge, J. M., Koole, S. L., Baumeister, R. F., Marquez, A., and Reid, M. W. (2011). Automatic emotion regulation after social exclusion: tuning to positivity. Emotion 11, 623–636. doi: 10.1037/a0023534
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Domsalla, M., Koppe, G., Niedtfeld, I., Vollstädt-Klein, S., Schmahl, C., Bohus, M., et al. (2014). Cerebral processing of social rejection in patients with borderline personality disorder. Soc. Cogn. Affect. Neurosci. 9, 1789–1797. doi: 10.1093/scan/nst176
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Downey, G., Feldman, S., and Ayduk, O. (2000). Rejection sensitivity and male violence in romantic relationships. Pers. Relationships 7, 45–61. doi: 10.1111/j.1475-6811.2000.tb00003.x
Downey, G., and Feldman, S. I. (1996). Implications of rejection sensitivity for intimate relationships. J. Pers. Soc. Psychol. 70, 1327–1343. doi: 10.1037/0022-3514.70.6.1327
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Downey, G., Lebolt, A., Rincon, C., and Freitas, A. L. (1998). Rejection sensitivity and children's interpersonal difficulties. Child Dev. 69, 545–560. doi: 10.1111/j.1467-8624.1998.tb06161.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Downey, G., and Romero-Cayas, R. (2005). “Rejection sensitivity as a predictor of affective and behavioral responses to interpersonal stress: A defensive motivational system,” in The Social Outcast: Ostracism, Social Exclusion, Rejection, and Bullying, eds K. D. Williams, J. P. Forgas, and W. Von Hippel (New York, NY: Psychology Press), 131–154.
Eisenberger, N. I. (2012a). Broken hearts and broken bones: a neural perspective on the similarities between social and physical pain. Curr. Dir. Psychol. Sci. 21, 42–47. doi: 10.1177/0963721411429455
Eisenberger, N. I. (2012b). The neural bases of social pain: evidence for shared representations with physical pain. Psychosom. Med. 74, 126–135. doi: 10.1097/PSY.0b013e3182464dd1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I. (2015). Social pain and the brain: Controversies, questions, and where to go from here. Annu. Rev. Psychol. 66, 601–629. doi: 10.1146/annurev-psych-010213-115146
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I., Inagaki, T. K., Muscatell, K. A., Byrne Haltom, K. E., and Leary, M. R. (2011). The neural sociometer: brain mechanisms underlying state self-esteem. J. Cogn. Neurosci. 23, 3448–3455. doi: 10.1162/jocn_a_00027
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I., Jarcho, J. M., Lieberman, M. D., and Naliboff, B. D. (2006). An experimental study of shared sensitivity to physical pain and social rejection. Pain 126, 132–138. doi: 10.1016/j.pain.2006.06.024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I., and Lieberman, M. D. (2004). Why rejection hurts: a common neural alarm system for physical and social pain. Trends Cogn. Sci. 8, 294–300. doi: 10.1016/j.tics.2004.05.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I., Lieberman, M. D., and Williams, K. D. (2003). Does rejection hurt? An fMRI study of social exclusion. Science 302, 290–292. doi: 10.1126/science.1089134
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eisenberger, N. I., Taylor, S. E., Gable, S. L., Hilmert, C. J., and Lieberman, M. D. (2007). Neural pathways link social support to attenuated neuroendocrine stress responses. Neuroimage 35, 1601–1612. doi: 10.1016/j.neuroimage.2007.01.038
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frye, A. F., Petralia, S. M., and Rhodes, M. E. (2000). Estrous cycle and sex differences in performance on anxiety tasks coincide with increases in hippocampal progesterone and 3α,5α-THP. Pharmacol. Biochem. Behav. 67, 587–596. doi: 10.1016/S0091-3057(00)00392-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gaertner, L., Iuzzini, J., and O'mara, E. M. (2008). When rejection by one fosters aggression against many: multiple-victim aggression as a consequence of social rejection and perceived groupness. J. Exp. Soc. Psychol. 44, 958–970. doi: 10.1016/j.jesp.2008.02.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gardner, W. L., Pickett, C. L., and Brewer, M. B. (2000). Social exclusion and selective memory: how the need to belong influences memory for social events. Pers. Soc. Psychol. Bull. 26, 486–496. doi: 10.1177/0146167200266007
Gardner, W. L., Pickett, C. L., Jefferis, V., and Knowles, M. (2005a). On the outside looking in: loneliness and social monitoring. Pers. Soc. Psychol. Bull. 31, 1549–1560. doi: 10.1177/0146167205277208
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gardner, W. L., Pickett, C. L., and Knowles, M. (2005b). “Social snacking and shielding: using social symbols, selves, and surrogates in the service of belonging needs,” in The Social Outcast: Ostracism, Social Exclusion, Rejection, and Bullying, eds K. D. Williams, J. P. Forgas, and W. Von Hippel. (New York, NY: Psycological Press), 227–242.
Gerber, J., and Wheeler, L. (2009). On being rejected a meta-analysis of experimental research on rejection. Perspect. Psychol. Sci. 4, 468–488. doi: 10.1111/j.1745-6924.2009.01158.x
Gilboa-Schechtman, E., Galili, L., Sahar, Y., and Amir, O. (2014). Being “in” or “out” of the game: subjective and acoustic reactions to exclusion and popularity in social anxiety. Front. Hum. Neurosci. 8:187. doi: 10.3389/fnhum.2014.00147
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gonsalkorale, K., and Williams, K. D. (2007). The KKK won't let me play: ostracism even by a despised outgroup hurts. Eur. J. Soc. Psychol. 37, 1176–1186. doi: 10.1002/ejsp.392
Green, D. M., and Swets, J. A. (1966). Signal Detection Theory and Psychophysics, Vol. 1. New York, NY: Wiley.
Harper, M. S., Dickson, J. W., and Welsh, D. P. (2006). Self-silencing and rejection sensitivity in adolescent romantic relationships. J. Youth Adolescence 35, 435–443. doi: 10.1037/a0026615
Heeren, A., Peschard, V., and Philippot, P. (2012). The causal role of attentional bias for threat cues in social anxiety: a test on a cyber-ostracism task. Cogn. Ther. Res. 36, 512–521. doi: 10.1007/s10608-011-9394-7
Holt-Lunstad, J., Smith, T. B., and Layton, J. B. (2010). Social relationships and mortality risk: a meta-analytic review. PLoS Med. 7:e1000316. doi: 10.1371/journal.pmed.1000316
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kawamoto, T., Araki, M., and Ura, M. (2013a). When a smile changes into evil: pitfalls of smiles following social exclusion. Int. J. Psychol. Stud. 5, 21–27. doi: 10.5539/ijps.v5n3p21
Kawamoto, T., Nittono, H., and Ura, M. (2013b). Cognitive, affective, and motivational changes during ostracism: an ERP, EMG, and EEG study using a computerized Cyberball task. Neurosci. J. 2013:304674. doi: 10.1155/2013/304674
Kawamoto, T., Nittono, H., and Ura, M. (2014). Social exclusion induces early-stage perceptual and behavioral changes in response to social cues. Soc. Neurosci. 9, 174–185. doi: 10.1080/17470919.2014.883325
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kawamoto, T., Onoda, K., Nakashima, K., Nittono, H., Yamaguchi, S., and Ura, M. (2012). Is dorsal anterior cingulate cortex activation in response to social exclusion due to expectancy violation? An fMRI study. Front. Evol. Neurosci. 4:11. doi: 10.3389/fnevo.2012.00011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kling, A., Lancaster, J., and Benitone, J. (1970). Amygdalectomy in the free-ranging vervet (Cercopithecus aethiops). J. Psychiatr. Res. 7, 191–199. doi: 10.1016/0022-3956(70)90006-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Krill, A., and Platek, S. M. (2009). In-group and out-group membership mediates anterior cingulate activation to social exclusion. Front. Evol. Neurosci. 1:1. doi: 10.3389/neuro.18.001.2009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kross, E., Egner, T., Ochsner, K., Hirsch, J., and Downey, G. (2007). Neural dynamics of rejection sensitivity. J. Cogn. Neurosci. 19, 945–956. doi: 10.1162/jocn.2007.19.6.945
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Laurin, K., Schumann, K., and Holmes, J. G. (2014). A relationship with god? Connecting with the divine to assuage fears of interpersonal rejection. Soc. Psychol. Pers. Sci. 5, 769–776. doi: 10.1177/1948550614531800
Lakin, J. L., Chartrand, T. L., and Arkin, R. M. (2008). I am too just like you: nonconscious mimicry as an automatic behavioral response to social exclusion. Psychol. Sci. 19, 816–822. doi: 10.1111/j.1467-9280.2008.02162.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Leary, M. R., Kowalski, R. M., Smith, L., and Phillips, S. (2003). Teasing, rejection, and violence: case studies of the school shootings. Aggressive Behav. 29, 202–214. doi: 10.1002/ab.10061
Legate, N., DeHaan, C. R., Weinstein, N., and Ryan, R. M. (2013). Hurting you hurts me too: the psychological costs of complying with ostracism. Psychol. Sci. 24, 583–588. doi: 10.1177/0956797612457951
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lynn, S. K., and Barrett, L. F. (2014). “Utilizing” signal detection theory. Psychol. Sci. 25, 1663–1673. doi: 10.1177/0956797614541991
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Macdonald, G., and Leary, M. R. (2005). Why does social exclusion hurt? The relationship between social and physical pain. Psychol. Bull. 131, 202–223. doi: 10.1037/0033-2909.131.2.202
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McPartland, J. C., Crowley, M. J., Perszyk, D. R., Naples, A. J., Mukerji, C. E., Wu, J., et al. (2011). Temporal dynamics reveal atypical brain response to social exclusion in autism. Dev. Cogn. Neurosci. 1, 271–279. doi: 10.1016/j.dcn.2011.02.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Maner, J. K., Dewall, C. N., Baumeister, R. F., and Schaller, M. (2007). Does social exclusion motivate interpersonal reconnection? Resolving the “porcupine problem.” J. Pers. Soc. Psychol. 92, 42–55. doi: 10.1037/0022-3514.92.1.42
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Maner, J. K., Miller, S. L., Schmidt, N. B., and Eckel, L. A. (2010). The endocrinology of exclusion: rejection elicits motivationally tuned changes in progesterone. Psychol. Sci. 21, 581–588. doi: 10.1177/0956797610362676
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Masten, C. L., Colich, N. L., Rudie, J. D., Bookheimer, S. Y., Eisenberger, N. I., and Dapretto, M. (2011). An fMRI investigation of responses to peer rejection in adolescents with autism spectrum disorders. Dev. Cogn. Neurosci. 1, 260–270. doi: 10.1016/j.dcn.2011.01.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Masten, C. L., Eisenberger, N. I., Borofsky, L. A., Pfeifer, J. H., Mcnealy, K., Mazziotta, J. C., et al. (2009). Neural correlates of social exclusion during adolescence: understanding the distress of peer rejection. Soc. Cogn. Affect. Neurosci. 4, 143–157. doi: 10.1093/scan/nsp007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Maurage, P., Joassin, F., Philippot, P., Heeren, A., Vermeulen, N., Mahau, P., et al. (2012). Disrupted regulation of social exclusion in alcohol-dependence: an fMRI study. Neuropsychopharmacology 37, 2067–2075. doi: 10.1038/npp.2012.54
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moor, B. G., Guroglu, B., Op De Macks, Z. A., Rombouts, S. A., Van Der Molen, M. W., and Crone, E. A. (2012). Social exclusion and punishment of excluders: neural correlates and developmental trajectories. Neuroimage 59, 708–717. doi: 10.1016/j.neuroimage.2011.07.028
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nakashima, K., Kawamoto, T., Isobe, C., and Ura, M. (2013). Differential responses of independent and interdependent people to social exclusion. Int. J. Psychol. Stud. 5, 22–31. doi: 10.5539/ijps.v5n1p31
Newman, M. L. (2014). Here we go again: bullying history and cardiovascular responses to social exclusion. Physiol. Behav. 133, 76–80. doi: 10.1016/j.physbeh.2014.05.014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nezlek, J. B., Wesselmann, E. D., Wheeler, L., and Williams, K. D. (2012). Ostracism in everyday life. Group Dyn. Theor. Res. 16, 91–104. doi: 10.1037/a0028029
Nolan, S. A., Flynn, C., and Garber, J. (2003). Prospective relations between rejection and depression in young adolescents. J. Pers. Soc. Psychol. 85, 745–755. doi: 10.1037/0022-3514.85.4.745
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Onoda, K., Okamoto, Y., Nakashima, K., Nittono, H., Ura, M., and Yamawaki, S. (2009). Decreased ventral anterior cingulate cortex activity is associated with reduced social pain during emotional support. Soc. Neurosci. 4, 443–454. doi: 10.1080/17470910902955884
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Onoda, K., Okamoto, Y., Nakashima, K., Nittono, H., Yoshimura, S., Yamawaki, S., et al. (2010). Does low self-esteem enhance social pain? The relationship between trait self-esteem and anterior cingulate cortex activation induced by ostracism. Soc. Cogn. Affect. Neurosci. 5, 385–391. doi: 10.1093/scan/nsq002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pickett, C. L., and Gardner, W. L. (2005). “The social monitoring system: enhanced sensitivity to social cues as an adaptive response to social exclusion,” in The Social Outcast: Ostracism, Social Exclusion, Rejection, and Bullying, eds K. D. Williams, J. Forgas, and W. Von Hippel (New York, NY: Psychological Press), 213–226.
Pickett, C. L., Gardner, W. L., and Knowles, M. (2004). Getting a cue: the need to belong and enhanced sensitivity to social cues. Pers. Soc. Psychol. Bull. 30, 1095–1107. doi: 10.1177/0146167203262085
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Poulsen, J. R., and Kashy, D. A. (2012). Two sides of the ostracism coin: how sources and targets of social exclusion perceive themselves and one another. Group Process. Intergr. Relat. 15, 457–470. doi: 10.1177/1368430211430517
Powers, K. E., Wagner, D. D., Norris, C. J., and Heatherton, T. F. (2013). Socially excluded individuals fail to recruit medial prefrontal cortex for negative social scenes. Soc. Cogn. Affect. Neurosci. 8, 151–157. doi: 10.1093/scan/nsr079
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Riva, P., Lauro, L. J. R., DeWall, C. N., and Bushman, B. J. (2012). Buffer the pain away stimulating the right ventrolateral prefrontal cortex reduces pain following social exclusion. Psychol. Sci. 23, 1473–1475. doi: 10.1177/0956797612450894
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Riva, P., Romero Lauro, L. J., Dewall, C. N., Chester, D. S., and Bushman, B. J. (2015). Reducing aggressive responses to social exclusion using transcranial direct current stimulation (tDCS). Soc. Cogn. Affect. Neurosci. doi: 10.1093/scan/nsu053. [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rotge, J. Y., Lemogne, C., Hinfray, S., Huguet, P., Grynszpan, O., Tartour, E., et al. (2015). A meta-analysis of the anterior cingulate contribution to social pain. Soc. Cogn. Affect. Neurosci. 10, 19–27. doi: 10.1093/scan/nsu110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Romero-Canyas, R., Downey, G., Berenson, K., Ayduk, O., and Kang, N. J. (2010). Rejection sensitivity and the rejection-hostility link in romantic relationships. J. Pers. 78, 119–148. doi: 10.1111/j.1467-6494.2009.00611.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sebastian, C., Blakemore, S. J., and Charman, T. (2009). Reactions to ostracism in adolescents with autism spectrum conditions. J. Autism Dev. Disord. 39, 1122–1130. doi: 10.1007/s10803-009-0725-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Silk, J. B., Alberts, S. C., and Altmann, J. (2003). Social bonds of female baboons enhance infant survival. Science 302, 1231–1234. doi: 10.1126/science.1088580
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Smart Richman, L., and Leary, M. R. (2009). Reactions to discrimination, stigmatization, ostracism, and other forms of interpersonal rejection: a multimotive model. Psychol. Rev. 116, 365–383. doi: 10.1037/a0015250
Smith, A., and Williams, K. D. (2004). R U There? Ostracism by cell phone text messages. Group Dyn. Theor. Res. 8, 291–301. doi: 10.1037/1089-2699.8.4.291
Somerville, L. H., Heatherton, T. F., and Kelley, W. M. (2006). Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Nat. Neurosci. 9, 1007–1008. doi: 10.1038/nn1728
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Themanson, J. R., Khatcherian, S. M., Ball, A. B., and Rosen, P. J. (2013). An event-related examination of neural activity during social interactions. Soc. Cogn. Affect. Neurosci. 8, 727–733. doi: 10.1093/scan/nss058
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tobin, S. J., Vanman, E. J., Verreynne, M., and Saeri, A. K. (2015). Threats to belonging on Facebook: lurking and ostracism. Soc. Influence 10, 31–42. doi: 10.1080/15534510.2014.893924
Troisi, J. D., and Gabriel, S. (2011). Chicken soup really is good for the soul: “Comfort food” fulfills the need to belong. Psychol. Sci. 22, 747–753. doi: 10.1177/0956797611407931
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Twenge, J. M., Baumeister, R. F., Tice, D. M., and Stucke, T. S. (2001). If you can't join them, beat them: effects of social exclusion on aggressive behavior. J. Pers. Soc. Psychol. 81, 1058–1069. doi: 10.1037/0022-3514.81.6.1058
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Van Beest, I., Williams, K. D., and Dijk, E. V. (2011). Cyberbomb: effects of being ostracized from a death game. Group Process. Intergr. 14, 581–596. doi: 10.1177/1368430210389084
Van Orden, K. A., and Joiner, T. E. (2013). “Depression and suicide: transactional relations with rejection,” in The Oxford Handbook of Social Exclusion, ed C. N. DeWall (New York, NY: Oxford University Press), 211–219.
Warburtona, W. A., Williams, K. D., and Cairns, D. R. (2006). When ostracism leads to aggression: the moderating effects of control deprivation. J. Exp. Soc. Psychol. 42, 213–220. doi: 10.1016/j.jesp.2005.03.005
Wesselmann, E. D., Butler, F. A., Williams, K. D., and Pickett, C. L. (2010). Adding injury to insult: unexpected rejection leads to more aggressive responses. Aggress. Behav. 36, 232–237. doi: 10.1002/ab.20347
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wesselmann, E. D., Nairne, J. S., and Williams, K. D. (2012a). An evolutionary social psychological approach to studying the effects of ostracism. J. Soc. Evolut. Cult. Psychol. 6, 309–328. doi: 10.1177/1948550612443386
Wesselmann, E. D., Wirth, J. H., Mroczek, D. K., and Williams, K. D. (2012b). Dial a feeling: detecting moderation of affect decline during ostracism. Pers. Individ. Dif. 53, 580–586. doi: 10.1016/j.paid.2012.04.039
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Will, G. J., Crone, E. A., and Guroglu, B. (2015). Acting on social exclusion: neural correlates of punishment and forgiveness of excluders. Soc. Cogn. Affect. Neurosci. 10, 209–218. doi: 10.1093/scan/nsu045
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Williams, K. D. (2009). “Ostracism: a temporal need-threat model,” in Advanced in Experimental Social Psychology, ed M. P. Zanna (New York, NY: Academic Press), 275–314.
Williams, K. D., Cheung, C. K., and Choi, W. (2000). Cyberostracism: effects of being ignored over the Internet. J. Pers. Soc. Psychol. 79, 748–762.
Wirth, J. H., Sacco, D. F., Hugenberg, K., and Williams, K. D. (2010). Eye gaze as relational evaluation: averted eye gaze leads to feelings of ostracism and relational devaluation. Pers. Soc. Psychol. Bull. 36, 869–882. doi: 10.1177/0146167210370032
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yanagisawa, K., Masui, K., Furutani, K., Nomura, M., Ura, M., and Yoshida, H. (2011a). Does higher general trust serve as a psychosocial buffer against social pain? An NIRS study of social exclusion. Soc. Neurosci. 6, 190–197. doi: 10.1080/17470919.2010.506139
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yanagisawa, K., Masui, K., Furutani, K., Nomura, M., Yoshida, H., and Ura, M. (2013). Family socioeconomic status modulates the coping-related neural response of offspring. Soc. Cogn. Affect. Neurosci. 8, 617–622. doi: 10.1093/scan/nss039
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yanagisawa, K., Masui, K., Onoda, K., Furutani, K., Nomura, M., Yoshida, H., et al. (2011b). The effects of the behavioral inhibition and activation systems on social inclusion and exclusion. J. Exp. Soc. Psychol. 47, 502–505. doi: 10.1016/j.jesp.2010.11.014
Zadro, L., Boland, C., and Richardson, R. (2006). How long does it last? The persistence of the effects of ostracism in the socially anxious. J. Exp. Soc. Psychol. 42, 692–697. doi: 10.1016/j.jesp.2005
Zadro, L., and Gonsalkorale, K. (2014). Sources of ostracism: the nature and consequences of excluding and ignoring others. Curr. Dir. Psychol. Sci. 23, 93–97. doi: 10.1177/0963721413520321
Keywords: social exclusion, intrapersonal process, interpersonal process, fMRI, ERPs
Citation: Kawamoto T, Ura M and Nittono H (2015) Intrapersonal and interpersonal processes of social exclusion. Front. Neurosci. 9:62. doi: 10.3389/fnins.2015.00062
Received: 28 May 2014; Accepted: 12 February 2015;
Published online: 06 March 2015.
Edited by:
Karol Osipowicz, Jefferson Neuroscience Hospital, USAReviewed by:
Antonio Pereira, Federal University of Rio Grande do Norte, BrazilAlexandre Heeren, Univesité Catholique de Louvain, Belgium
Copyright © 2015 Kawamoto, Ura and Nittono. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence:dGFpc2hpLmthd2Ftb3RvQGdtYWlsLmNvbQ==