 Valérie Dufour
Valérie Dufour Cristian Pasquaretta
Cristian Pasquaretta Pierre Gayet
Pierre Gayet Elisabeth H. M. Sterck
Elisabeth H. M. Sterck- 1Ethology Evolutive Team, Institute Pluridisciplinaire Hubert Curien (IPHC), University of Strasbourg, CNRS, Strasbourg, France
- 2Research Center on Animal Cognition (CRCA), Center for Integrative Biology (CBI), Centre National de la Recherche Scientifique, University of Toulouse, UPS, Toulouse, France
- 3Ethology Research, Animal Science Department, Biomedical Primate Research Center, Rijswijk, Netherlands
- 4Animal Ecology, Utrecht University, Utrecht, Netherlands
In a previous study (Dufour et al., 2015) we reported the unusual characteristics of the drumming performance of a chimpanzee named Barney. His sound production, several sequences of repeated drumming on an up-turned plastic barrel, shared features typical for human musical drumming: it was rhythmical, decontextualized, and well controlled by the chimpanzee. This type of performance raises questions about the origins of our musicality. Here we recorded spontaneously occurring events of sound production with objects in Barney's colony. First we collected data on the duration of sound making. Here we examined whether (i) the context in which objects were used for sound production, (ii) the sex of the producer, (iii) the medium, and (iv) the technique used for sound production had any effect on the duration of sound making. Interestingly, duration of drumming differed across contexts, sex, and techniques. Then we filmed as many events as possible to increase our chances of recording sequences that would be musically similar to Barney's performance in the original study. We filmed several long productions that were rhythmically interesting. However, none fully met the criteria of musical sound production, as previously reported for Barney.
Introduction
The universality of music across human cultures is an undisputable fact: all humans make music, sing, dance, and gather to enjoy sharing emotions elicited by musical performances (Merker et al., 2015). By contrast, there is very limited evidence that our closest living relatives, the great apes, make music in the same way (Fitch, 2006). From an evolutionary perspective, this leaves us wondering about the origins of our musical skills. Archeological remains do not provide sufficient evidence of instruments being used to create music prior to 40,000 BC, nor do they suggest the presence of any other musical behaviors (Kunej and Turk, 2000). Great apes probably possess some prerequisites for musical productions (Fitch, 2006; Honing et al., 2015). Drumming in chimpanzees and chest beating in gorillas is considered a homolog to human music: a shared ancestral trait not found in less closely related species (Fitch, 2006). Thus, taking a closer look at great ape drumming behavior may enlighten our understanding of our own musicality.
Chimpanzees drum on tree buttresses or other resonant structures as part of their dominance displays (Goodall, 1986). The chimpanzee Mike was observed repeatedly charging higher-ranking males whilst propelling kerosene cans in front of him (Goodall, 1986). Drumming can accompany vocal signals such as long-distance calls (e.g., climax of a pant hoot), sometimes even replacing part of the vocal phrase (Boesch, 1991; Arcadi, 1996; Babiszewska et al., 2015). This combination of vocalizations and noise making using objects is generally associated with social tension or high levels of arousal within the group (Goodall, 1986; Nishida et al., 1999). Other individuals can join the initiator by vocalizing, drumming, or both (Fedurek et al., 2013). Also gorillas drum on their own bodies and objects during displays (Schaller, 1963). Generally, drumming in display contexts is likely to be perceived by others as a demonstration of strength.
Drumming, leaf clipping, and breaking or shaking branches are examples of how chimpanzee males create sound with objects to communicate sexual interest (Nishida, 1980; Goodall, 1986). After demonstrating such intent, it is not rare to see the targeted female approach and present for a mount. Similarly, musically qualified humans may be more successful at seduction than their non-musical counterparts, indicating that musical ability could be a sign of male health and fertility (Miller, 2000; Sluming and Manning, 2000; Charlton, 2014).
Finally, making noise can be associated with pleasurable emotion, both in humans and great apes. Humans play music. Young chimpanzees often drag branches noisily along the ground and were seen, on one occasion, to repeatedly hit a clay pot (Matsusaka, 2012), seemingly enjoying the noise. Thus, on both contextual and emotional levels, there are several links between sound making with objects in great apes and instrumental music in humans.
However, many other properties of noise making with objects by great apes do not meet criteria for music. Indeed, drumming in great apes is generally contextualized (sex, play, or display context), whereas humans produce music outside any particular context or function (Arom, 2000). Music is very often rhythmical, while drumming sequences of apes are generally too short to allow for rhythm to be detected (but see Dufour et al., 2015).
Another key component of music production is the performers' capacity to synchronize their actions to an external beat (Arom, 2000). We know that pinnipeds (Cook et al., 2013), cockatoos (Patel et al., 2009), parrots (Schachner et al., 2009), and budgerigars (Hasegawa et al., 2011) can learn or be trained—with varying levels of precision—to synchronize their body movement to a rhythm. A form of action entrainment by motor mimicry of pounding gestures has been reported in young chimpanzees watching others cracking nuts (Fuhrmann et al., 2014). The female chimpanzee Aï spontaneously pressed two keys on a keyboard in synchrony to a rhythmical auditory stimulus without any previous training (Hattori et al., 2013). Recently, a bonobo was found to occasionally match its own drumming tempo to the one of a human drummer (Large and Gray, 2015), even when the tempo differed slightly from the natural pace of the bonobo. However, the tempo matching disappeared quickly despite the bonobo being encouraged and rewarded for drumming. By comparison, human children can synchronize to external drumming at the age of around 3 years, increasing in accuracy as they grow older (Honing et al., 2012). Kanzi, a language-trained bonobo, was reported to perform rhythmical drumming (Kugler and Savage-Rumbaugh, 2002), but there are no published data describing this event. In a recent study, we described a long drumming solo on an upturned plastic barrel by a chimpanzee named Barney. This solo was rhythmic, decontextualized, and fitted several criteria for human music (Dufour et al., 2015). It would be justifiable to question the significance of this unique observation: was Barney's performance a “once in a lifetime” event, i.e., a chimpanzee accidentally “discovering music”? Or was it a rare behavior that had gone unnoticed by chimpanzee specialists and had not been given the consideration it deserved, remaining unpublished due to its anecdotal nature? Most importantly, can Barney do it again?
To identify the factors leading to Barney's performance, we conducted a 2 months-long survey on sound making with objects in the chimpanzee facility where Barney was living. Our first aim was to gather information on factors influencing the duration of sound making using objects found in the environment. To that end we recorded the context, the medium used, and the complexity of sound production techniques. Furthermore, we noted the sex of the individual producing sound with objects. Reports on chimpanzee drumming in the wild (Nishida, 1980; Goodall, 1986; Arcadi, 1996; Nishida et al., 1999) suggest that—in particular male chimpanzees—drum primarily in socio- or emotionally negative contexts, such as displays. However, since studies on chimpanzee drumming are rather scarce so far, formulating informed predictions for the effects of context, sex, technique, and medium on drumming durations is not straightforward. Therefore, we adapted an explorative approach to tackle potential effects of these predictor variables on the duration of sound production. Secondly, we aimed at recording as many sound production events as possible in order to detect any performances resembling Barney's original drumming (i.e., long, rhythmically interesting, and decontextualized sequences). Any such cases were checked for evidence for musicality.
Methods
Subjects and Study Site
This study took place at the Biomedical Primate Research Centre (BPRC) at Rijswijk, the Netherlands, in July and August 2005. The facility held a total of 54 individuals of which 28 were adult females and 26 adult males. The population was composed of six groups living in enclosures with outside areas facing the same courtyard (see Figure 1). Thus, some groups (those who faced other groups) could see each other and all groups could hear each other. Note that this colony was moved to the Safari Park Beekse Bergen in 2006, and that the BPRC no longer houses chimpanzees. The survey of sound production using objects or other enclosure elements as a resonating medium (sound-object use, hereafter referred to as So-U) was carried out in two phases. In Phase 1 (from 13th July to 12th August 2005) data were collected for the four groups that had at least two adult males (Dirk's group, Barney's group, Bob's group and Dennis' group). Due to low rates of So-U in one of the groups (Bob's group), this group was removed from Phase 1 data collection after a few days. In Phase 2 data collection on So-U sequence recordings took place on all six groups from the 22nd of July to the 25th of August 2005.
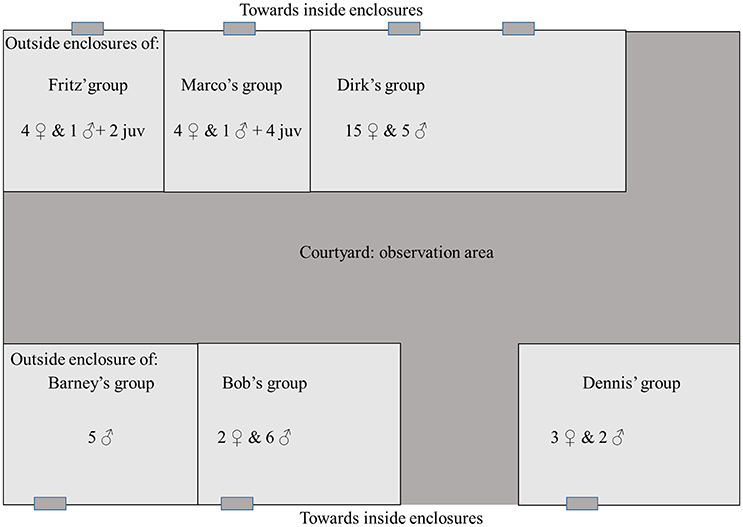
Figure 1. Disposition and group composition of the chimpanzee's colony. Some group can see each other, and all groups can hear each other.
Phase 1: Contexts of So-U
Data Collection
In Phase 1 the goal was to gather as much information about the factors affecting the duration of So-U as possible. One observer (PG) stood in the central courtyard and focused on one group for 15 min at a time. We conducted a total of 162 focal observations (54 per group). Focal observations were spread equally throughout the day (from 8.30 to 10 am: 15 observations per group, 10.30–12.20: 20 observations per group, and 1.20–5 p.m.: 19 observations per group) and the groups were observed in random order within each time period. During focal group observations the observer wrote down all occurrences of So-U in the group, noting the identity of the individual producing the sound, the duration of So-U, the type of medium used, the technique used, and any information (individual posture, behavior, or external factors) that could indicate the context of occurrence (Table 1). Some contexts were easily identifiable (i.e., nest-building, play, sexual activity, intimidation display without aggression, aggression, see Goodall, 1986). Others were not always clearly discernable in the absence of any obvious contextual information. An example for this is “tension,” which was defined as So-U when the animal had its hair erect (hunched back posture) without escalation into a display or an aggression and in the absence of any other contextual element. Indeed, hunching can sometimes also occur in courtship (but lead then to copulation or to attempted copulation, accompanied by an erected penis), greetings (involving a “friendly reunion”), or excitement (upon seeing enriching food, hearing other chimpanzees, etc.) (Nishida et al., 1999). When courtship and greeting could be excluded and no external stimuli potentially triggering excitement were apparent, we assigned the context “tension” to the So-U episode.

Table 1. Description of the behavioral units recorded live by the observer for contexts, type of medium, techniques, vocalizations, and postures.
Data Re-coding for Phase 1
To ensure a sufficiently large and reliable dataset allowing for So-U analysis we introduced categories for contexts, drumming techniques, and objects used for sound production respectively.
As So-U occurred frequently when individuals were tense, aggressive, or displayed we regrouped these contexts into one category that we termed “socio- or emotionally negative contexts.” Socio- or emotionally positive contexts comprised sexual activity, playing, teasing, nest-building, and attention seeking. Any outside-group noise and unidentified contexts (Figure 2) were excluded from data analyses. Note that comparing So-U durations between contexts when working with data that were not controlled for durations of the behavioral category itself might be questionable. We took this limitation into account when discussing the results.
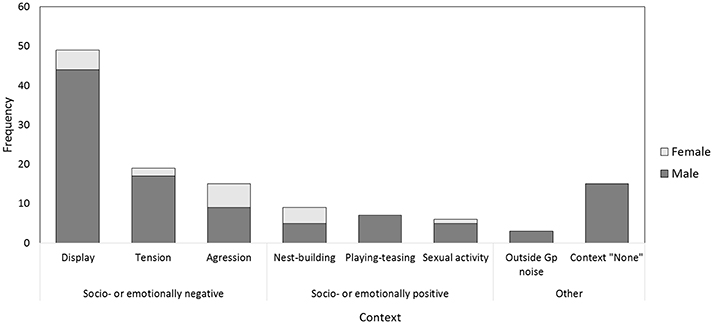
Figure 2. Contexts of So-U according to sex. Main contexts are aggression, display, tension (socio- or emotionally negative contexts). Other contexts are sexual activity, teasing, playing, attention seeking, and nest-building (considered as socio- or emotionally positive). Note that most So-U in sexual activity are initiated by males, except one initiated by a female who sought the male attention by knocking on a door. So-U with unidentified contexts (context “none”) are also recorded.
For the noise production techniques we distinguished between “simple” (only one technique used, i.e., hitting, shaking, trailing, half-circling, pushing, or throwing) and “complex” (combination of two or more techniques) techniques.
Regarding the objects used to produce noise we differentiated between (i) metallic media (doors, fence, metallic ground), (ii) plastic containers (small plastic containers, half and large plastic barrels), and (iii) small or non-resonating objects (small plastic bottles, enclosure furniture, cardboard tubes).
Statistical Models for Phase 1
First we used log likelihood ratio test (LRT) to check for individual differences in the frequency and handling duration of So-U. We compared the full model with individual nested in group as a random effect against the full model with only group as a random effect. We build a model to study the response variable “duration of So-U.” We used multi-model inference and model averaging methods to calculate the weight of evidence (Wi) for each predictor involved in the model (Burnham and Anderson, 2002). We standardized the binary predictors to a common scale both to correctly estimate their influence (Engqvist, 2005) and to control for some possible degree of collinearity among them Schielzeth (2010). We carried out an information theoretical approach: all the possible models were run and ranked based on their AIC scores (Akaike, 1985) after correction for small sample sizes (AICc) and their normalized Akaike weights (AICw, Burnham and Anderson, 2002). We computed estimates, standard errors and 95% confidence intervals (CI) for models whose cumulative model weights reached 90% of the total weights. We ran model selections and averages using the R Package MuMIn (Barton, 2009). The model was run following a negative binomial error distribution using the R package glmmADMB (Fournier et al., 2012).
The full model included the following predictor variables: (i) the medium, (ii) the complexity of the technique, (iii) the context, (iv) the sex, and (v) all possible interactions of these main effects except for the interaction “technique-medium.” Indeed, among all the So-U performed with mixed techniques (33 events) 32 occurred with the same medium (plastic container) preventing us to investigate the effect of this interaction. Note that only one female used a small medium (and in only one occasion). Therefore, we also excluded the interaction “sex-medium” from the model.
Phase 2: Video Recording of So-U for Further Rhythmical Analysis
In Phase 2 we conducted all occurrence sampling of So-U in all six groups to increase our chances of filming a long, decontextualized, and rhythmically complex So-U resembling Barney's original performance. We recorded the identity of the performer and the context of occurrence. For interesting bouts rhythmical analysis was conducted in the same way as in Dufour et al. (2015), i.e., by checking the possibility of rhythmical patterns in a series of impacts (Ljung-box test analysis), and then checking the predictability of the next section using autocorrelation analysis (see Supplementary Methods). For each sequence data were analyzed using R (R Core Team, 2013). P level of significance was set at 0.05.
Ethics Statement
The study was conducted in compliance with all relevant Dutch laws and respected international and scientific standards and guidelines. All analyses were based on the recording of spontaneous behavioral sequences initiated by the chimpanzees. Due to the observational nature of this study and the absence of discomfort for the animals no additional permission was required from the institute's animal experiment committee, as assessed by the Biomedical Primate Research Centre Animal welfare officer.
Results
Phase 1: Contexts of So-U
During focal observations in Phase 1 we monitored a total of 123 So-U with various media (Supplementary Figure 1). So-U occurred in several contexts including aggression, display, tension, sexual activity, teasing, playing, attention seeking, nest-building, and outside group noise. 15 (of the 123) bouts could not be assigned to any context. Sound production in unidentified contexts occurred only in males (Figure 2). While So-U occurred both in males and in females, males did so more often than females (males: 105 times, females: 18 times), all contexts included. We recorded 88 So-U with simple techniques and 35 with complex techniques (16 individuals out of 24 never used complex techniques). Focusing on the two main contextual categories (thus excluding context “none” and context “outside group noise”), we recorded more So-U in socio- or emotionally negative contexts (83) than in socio- or emotionally positive contexts (22). There were significant differences between individuals both in the frequency and duration of So-U (LRT frequency model: df = 1, Δ deviance = 4.25, P = 0.039; LRT duration model: df = 1, Δ Deviance = 6.254, P = 0.012).
The time spent handling a medium in So-U varied from 1 to 720 s, with a median duration of 6 s. Model averaging on the duration of So-U as the response variable revealed that context, sex, the interaction between context and sex, and the technique used had higher relative importance than other variables in the model (Table 2). So-U in socio- or emotionally positive contexts lasted longer than in socio- or emotionally negative ones (Estimate = 1.053, 95% IC: 0.052–2.055; Table 2). Males had longer So-U than females but this was only significant in negative contexts (in negative contexts: Estimate = −3.253, 95% IC: −4.891 to −1.615; in positive contexts: Estimate = −1.991, 95% IC: −4.151–0.167; Table 2, Figure 3). Complex techniques lasted longer than simple ones (Estimate = 1.053, 95% IC: 0.132–1.161; Figure 4).
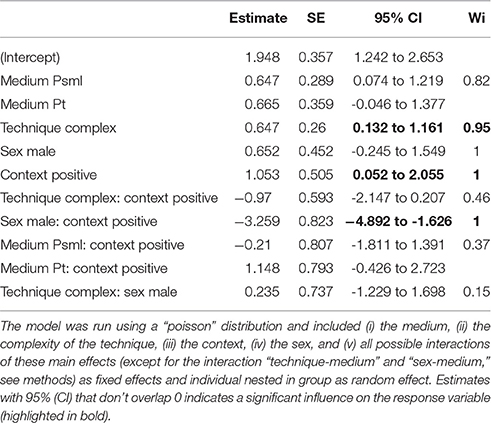
Table 2. Model average using AICc-based selection approach, showing estimate, standard error (SE), 95% confidence interval (95% CI) and relative weight of evidence (Wi) for each variable both for the handling duration model.
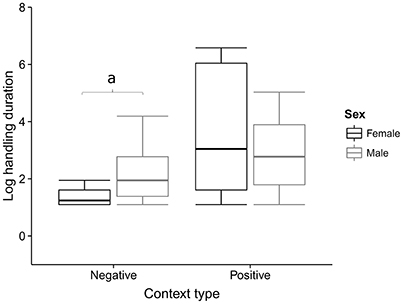
Figure 3. So-U duration according to the context for males and females. The average model performed on the duration of So-U indicates that males had significantly longer So-U than females in the negative contexts (“a” indicates this significant influence in the figure).
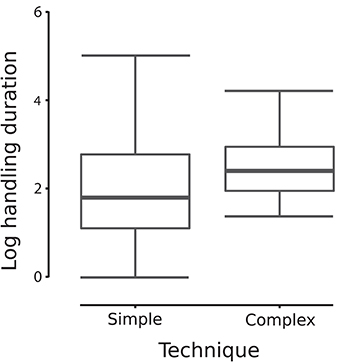
Figure 4. So-U duration according to each type of technique (complex or simple). The average model indicates that complex techniques involved longer durations than simple techniques (“a” indicates this significant influence in the figure).
Are there any Similarities with Barney's Drumming?
In Phase 1, we observed 27 So-U that lasted longer than 20 s. Eight were performed with either a half barrel or a large barrel. Only one of these performances was not clearly associated to a context, as the performer alternated between hoots and a relaxed face while shaking the barrel. The seven remaining bouts of sound production were contextualized (4 displays, 1 tension, 1 nesting, 1 sexual activity). Barney performed only six So-U, two of which involved using a small container, and four of which involved hitting a door. Five of these events occurred in a tension context and one was associated to a display.
In Phase 2, we filmed 262 events of So-U by various individuals, including 90 So-U that lasted longer than 20 s. Eighty-four of these occurred in a clearly recognizable context: socially negative (46), nest-building (10), sexual activity (9), attention-seeking (3), or play (16). Note that several sequences could sometimes be detected within one So-U bout. In total we analyzed the rhythmical properties of 10 So-U, producing a total of 20 sequences, i.e., those in which successive beats could be clearly identified from the background noise (Table 3). Rhythmical analysis concerned only seven contextualized So-U (Table 3). Two of these were socially negative So-U (Oscar seq 1; Oscar seq 2.1 and 2.2, Table 3). However, the technique used in these cases (holding a barrel and hitting it against the wall with half-circling trajectories, Supplementary Video 1), could potentially explain their rhythmicity. Three sexual displays (Paul seq 1.1, 1.2, and 1.3: Supplementary Video 2, Dennis seq 1 and Dennis seq 2.1 and 2.2) showed interesting and complex rhythmical patterns (Ljung-box portmanteau test, Supplementary Figure 2). In Paul seq 1.2, for example, there were alternating series of short and long inter-beat durations with a remarkably long-term dependency (autocorrelation test, Table 3 and Supplementary Figure 3). The tempo was independent of the technique or medium used, and seemed to be controlled by the chimpanzees. Finally, among the six So-U without clear contexts, two showed analyzable rhythmical properties (Table 3). All of them could also be rhythmically highly dependent on the technique used (see Supplementary Videos 3 and 4).
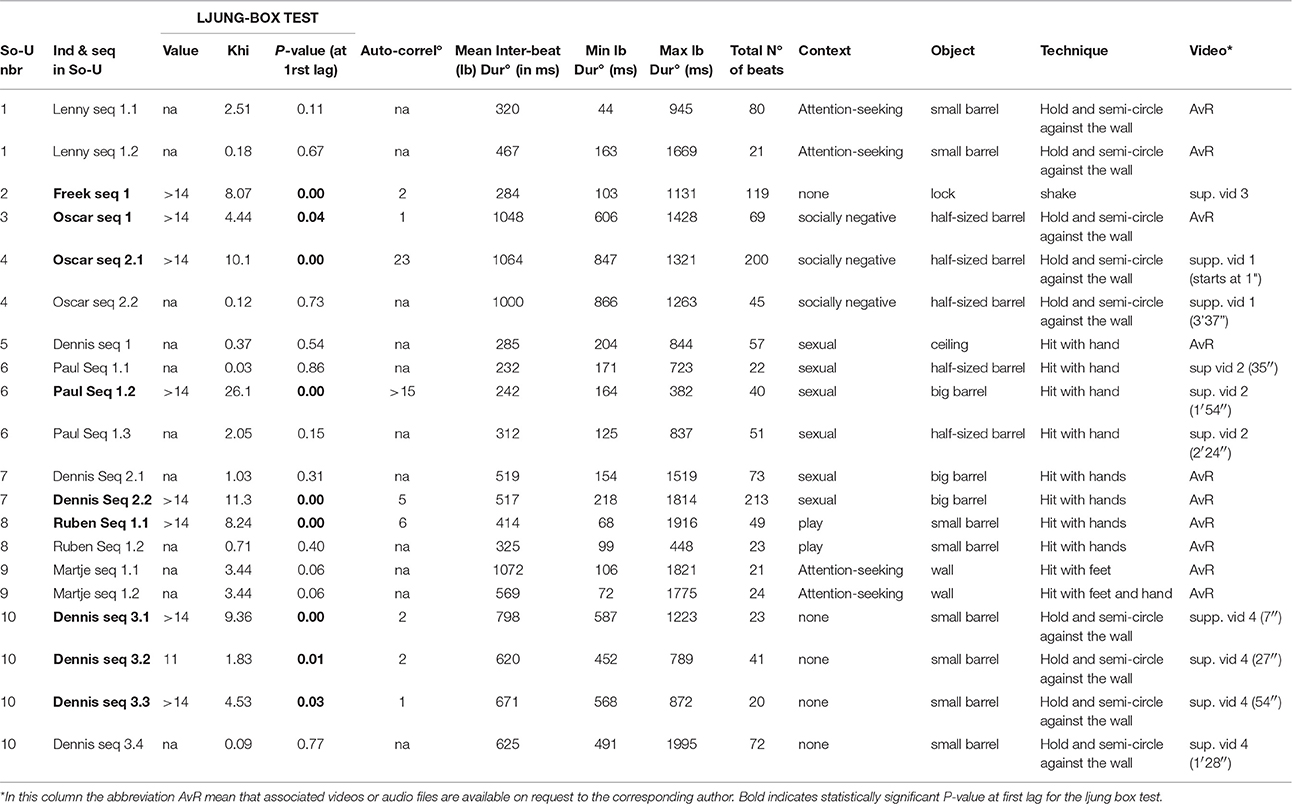
Table 3. List of the 10 So-U in which a total of 20 sequences longer than 20 inter-beat intervals could be detected.
Discussion and Conclusion
The evolution of musicality is shrouded in the mists of our past, but the production of sounds with objects by chimpanzees may reveal the presence of some of its prerequisites in our common ancestor. Although we did not record a second instance of decontextualized and rhythmic drumming like the one event recorded for Barney (Dufour et al., 2015), the chimpanzees of the studied colony frequently incorporated objects when making sound. Sound production occurred in a diversity of contexts. So-U produced in socially or emotionally positive contexts lasted longer than in negative contexts. The primary explanation could be that our target behaviors for socio- or emotionally negative contexts (e.g., displays) may simply not last as long as the target behaviors in socio- or emotionally positive contexts (e.g., play bouts or building a nest)—with or without So-U. Therefore, the difference in duration of So-U across contexts may likely be an artifact of socially or emotionally positive behaviors lasting longer than socio- and emotionally-negative ones. An alternative hypothesis (that may be considered in further studies) is that So-U might have lasted longer in this context because individuals were involved in a relaxing activity like nesting and playing. Their attention was maybe better focused on the production of sounds and its pleasurable aspects.
More than 85% of So-U were produced by males, which is in line with what we expected based on the literature (Nishida, 1980; Goodall, 1986; Arcadi, 1996; Nishida et al., 1999). As So-U in socio- or emotionally negative contexts lasted longer when produced by males than by females, we could speculate that male chimpanzees are more motivated than females to produce sound with object in this context, but we cannot generalized at the population level. This observation fits well with the intimidating function of buttress drumming described in wild male chimpanzees (Goodall, 1986). Further work should aim at a more detailed assessment of the communicative function of So-U by assessing responses from the audience in these contexts. In socially or emotionally positive contexts, there was no difference in object handling durations between females and males.
Finally, So-U could involve complex techniques that lasted longer than So-U with simple techniques. We hypothesize that shifting from one technique to another could be a way to counter tiredness arising from multiple repetitions of the same gestures. This illustrates how chimpanzees actively engaged in producing sounds with objects, a prerequisite for the evolution of music.
Given the diversity of contexts recorded, we cannot conclude about the main driving force in the production of So-U in our colony (388 events in <2 months). We cannot therefore pinpoint which factor most probably led to the discovery and spreading of music by our ancestors: the need to demonstrate strength, to attract females, or the pleasurable aspects of making noise.
One objective of this study was to check if Barney or the other chimpanzees of the colony were capable of producing a performance similar to the one reported in Dufour et al. (2015) on a regular basis. Some individuals produced long and/or elaborated So-U bouts. Most So-U were contextualized, short and “unremarkable,” except maybe for some sexual displays reminiscent of human and bird courtship displays as illustrated in Supplementary Video 3. In this video, the male successfully attracted a female's attention by repeatedly hitting a large barrel with a rather slow and clearly audible tempo (with multiple repositioning of the barrel toward the female). Note that in the wild, sexual displays are more likely to involve branch breaking or leaf clipping (Nishida, 1980) rather than demonstrative drumming per se (Crockford and Boesch, 2005). In this respect, this video illustrates, potentially, an innovative use of drumming compared to wild chimpanzees. When focusing on the longest and most decontextualized So-U, we found interesting rhythmical patterns. However, most of these manipulations were constrained by the general configuration of the sound production: like, for example, hitting a barrel with a semi-circle trajectory against the wall (see Supplementary Video 1, for an example). The rhythmical element was not therefore entirely controlled by the chimpanzee, as it was in Barney's case. If Barney's solitary drumming bout had not been recorded by chance, this unique evidence of potential rhythmicity in chimpanzees would never have been brought to light. He did not repeat this feat during the study and may never do so again, making this recording all the more valuable.
At this point, we may question the adequacy of Arom's “decontextualization” criterion when evaluating musicality in animals (Arom, 2000). Indeed, human music is often contextualized (associated to rituals and social functions). The inclusion of this criteria sets the bar very high for sound production in animals to be considered music, and excludes many vocal sophistications heard in some bird songs. It also excludes some of the rhythmical sexual displays we recorded here. A more flexible use of Arom's criteria might therefore be needed to widen our understanding of animal musicality. Nevertheless, the structure of Barney's initial performance remains undeniably and intuitively recognizable as drumming, and conforms with the “higher order” criteria proposed by Arom (2000).
The many studies that explore the origins of music (e.g., including research presented in this special issue), are hampered by the limited amount of documentation describing instrumental sound production in apes. Although music appears to be within the grasp of chimpanzees, they have not yet taken the step to music per se. This modest contribution was designed to provide additional information about the use of objects and various media for sound production by chimpanzees, thus providing a starting point for further work along these lines (see also Ravignani et al., 2013). Further studies should attempt to investigate the type of attraction that instrumental noise making has on chimpanzees, including the refinement and leisureliness expressed while doing so. This should contribute to a better understanding of how music evolved.
Author Contributions
ES and VD designed the study, analyzed the data and wrote the manuscript. PG and VD collected the data. CP supervised and designed the data analysis.
Funding
CP was funded by an ANR programme blanc (ANR 12 BSV7 0013 02).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fnins.2017.00002/full#supplementary-material
References
Akaike, H. (1985). “Prediction and entropy,” in A Celebration of Statistics, eds A. C. Atkinson and S. E. Fienberg (New York, NY: Springer), 1–24.
Arcadi, A. C. (1996). Phrase structure of wild chimpanzee pant hoots: patterns of production and interpopulation variability. Am. J. Primatol. 39, 159–178. doi: 10.1002/(SICI)1098-2345(1996)39:3<159::AID-AJP2>3.0.CO;2-Y
Arom, S. (2000). “Prolegomena to a biomusicology,” in The Origins of Music, eds N. L. Wallin, B. Merker, and S. Brown (London: MIT Press), 27–29.
Babiszewska, M., Schel, A. M., Wilke, C., and Slocombe, K. E. (2015). Social, contextual, and individual factors affecting the occurrence and acoustic structure of drumming bouts in wild chimpanzees (Pan troglodytes). Am. J. Phys. Anthropol. 156, 125–134. doi: 10.1002/ajpa.22634
Barton, K. (2009). MuMin: Multi-Model Inference. R package version 1.13.4. Available online at: http://cran.r-project.org/package=MuMIn
Boesch, C. (1991). Symbolic communication in wild chimpanzees? Hum. Evol. 6, 81–89. doi: 10.1007/BF02435610
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd Edn. New York, NY: Springer.
Charlton, B. D. (2014). Menstrual cycle phase alters women's sexual preferences for composers of more complex music. Proc. R. Soc. B Biol. Sci. 281:20140403. doi: 10.1098/rspb.2014.0403
Cook, P., Rouse, A., Wilson, M., and Reichmuth, C. (2013). A California sea lion (Zalophus californianus) can keep the beat: motor entrainment to rhythmic auditory stimuli in a non-vocal mimic. J. Comp. Psychol. 127, 412–427. doi: 10.1037/a0032345
Crockford, C., and Boesch, C. (2005). Call combinations in wild chimpanzees. Behaviour 142, 397–421. doi: 10.1163/1568539054012047
Dufour, V., Poulin, N., Curé, C., and Sterck, E. H. M. (2015). Chimpanzee drumming: a spontaneous performance with characteristics of human musical drumming. Sci. Rep. 5:11320. doi: 10.1038/srep11320
Engqvist, L. (2005). The mistreatment of covariate interaction terms in linear model analyses of behavioural and evolutionary ecology studies. Anim. Behav. 70, 967–971. doi: 10.1016/j.anbehav.2005.01.016
Fedurek, P., Machanda, Z. P., Schel, A. M., and Slocombe, K. E. (2013). Pant hoot chorusing and social bonds in male chimpanzees. Anim. Behav. 86, 189–196. doi: 10.1016/j.anbehav.2013.05.010
Fitch, W. T. (2006). The biology and evolution of music: a comparative perspective. Cognition 100, 173–215. doi: 10.1016/j.cognition.2005.11.009
Fournier, D. A., Skaug, H. J., Ancheta, J., Ianelli, J., Magnusson, A., Maunder, M., et al. (2012). AD Model Builder: using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optim. Methods Softw. 27, 243–249. doi: 10.1080/10556788.2011.597854
Fuhrmann, D., Ravignani, A., Marshall-Pescini, S., and Whiten, A. (2014). Synchrony and motor mimicking in chimpanzee observational learning. Sci. Rep. 4:5283. doi: 10.1038/srep05283
Goodall, J. (1986). “Grooming,” in The Chimpanzees of Gombe: Patterns of Behavior, (Cambridge, MA: The Belkna), 387–408.
Hasegawa, A., Okanoya, K., Hasegawa, T., and Seki, Y. (2011). Rhythmic synchronization tapping to an audio–visual metronome in budgerigars. Sci. Rep. 1, 1–8. doi: 10.1038/srep00120
Hattori, Y., Tomonaga, M., and Matsuzawa, T. (2013). Spontaneous synchronized tapping to an auditory rhythm in a chimpanzee. Sci. Rep. 3:1566. doi: 10.1038/srep01566
Honing, H., Merchant, H., Haden, G. P., Prado, L., and Bartolo, R. (2012). Rhesus monkeys (Macaca mulatta) detect rhythmic groups in music, but not the beat. PLoS ONE 7:e51369. doi: 10.1371/journal.pone.0051369
Honing, H., ten Cate, C., Peretz, I., and Trehub, S. E. (2015). Without it no music: cognition, biology and evolution of musicality. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:20140088. doi: 10.1098/rstb.2014.0088
Kugler, K., and Savage-Rumbaugh, S. (2002). “Rhythmic drumming by Kanzi an adult male bonobo (Pan paniscus) at the language research center,” in Paper Presented at the 25th Meeting of the American Society of Primatologists, Norman: Oklahoma University.
Kunej, D., and Turk, I. (2000). “New perspectives on the beginnings of music: Archeological and musicological analysis of a middle Paleolithic bone ‘flute’,” in Origins of Music, eds N. L. Wallin and B. Merker (Cambridge: MIT Press), 235–268.
Large, E. W., and Gray, P. M. (2015). Spontaneous tempo and rhythmic entrainment in a Bonobo. J. Comp. Psychol. 129, 317–328. doi: 10.1037/com0000011
Matsusaka, T. (2012). Playful drumming by immature wild chimpanzees at mahale : do they enjoy making sounds ? Pan Africa News 19, pp. 9–11.
Merker, B., Morley, I., and Zuidema, W. (2015). Five fundamental constraints on theories of the origins of music. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:20140095. doi: 10.1098/rstb.2014.0095
Miller, G. F. (2000). “Evolution of human music through sexual selection,” in The Origins of Music, eds N. L. Wallin and B. Merker (Cambridge: MIT Press), 329–360.
Nishida, T. (1980). The leaf-clipping display: a newly-discovered expressive gesture in wild chimpanzees. J. Hum. Evol. 9, 117–128. doi: 10.1016/0047-2484(80)90068-8
Nishida, T., Kano, T., Goodall, J., McGrew, W. C., and Nakamura, M. (1999). Ethogram and ethnography of Mahale chimpanzees. Anthropol. Sci. 107, 141–188. doi: 10.1537/ase.107.141
Patel, A. D., Iversen, J. R., Bregman, M. R., and Schulz, I. (2009). Studying synchronization to a musical beat in nonhuman animals. Ann. N.Y. Acad. Sci. 1169, 459–469. doi: 10.1111/j.1749-6632.2009.04581.x
Ravignani, A., Matellan Olivera, V., Gingras, B., Hofer, R., Rodriguez Hernandez, C., Sonnweber, R. S., et al. (2013). Primate drum kit: a system for studying acoustic pattern production by non-human primates using acceleration and strain sensors. Sensors (Basel) 13, 9790–9820. doi: 10.3390/s130809790
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/
Schachner, A., Brady, T. F., Pepperberg, I. M., and Hauser, M. D. (2009). Spontaneous motor entrainment to music in multiple vocal mimicking species. Curr. Biol. 19, 831–836. doi: 10.1016/j.cub.2009.03.061
Schielzeth, H. (2010). Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 1, 103–113. doi: 10.1111/j.2041-210X.2010.00012.x
Keywords: music, drumming, object manipulation, chimpanzees, Barney's colony
Citation: Dufour V, Pasquaretta C, Gayet P and Sterck EHM (2017) The Extraordinary Nature of Barney's Drumming: A Complementary Study of Ordinary Noise Making in Chimpanzees. Front. Neurosci. 11:2. doi: 10.3389/fnins.2017.00002
Received: 31 March 2016; Accepted: 03 January 2017;
Published: 19 January 2017.
Edited by:
Andrea Ravignani, Vrije Universiteit Brussel, BelgiumReviewed by:
Bjorn Hellmut Merker, Self, SwedenRuth Sonnweber, Max Planck Institute for Evolutionary Anthropology (MPG), Germany
Copyright © 2017 Dufour, Pasquaretta, Gayet and Sterck. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Valérie Dufour, dmFsZXJpZS5kdWZvdXJAaXBoYy5jbnJzLmZy