 Ida A. K. Nilsson1,2,3*
Ida A. K. Nilsson1,2,3*- 1Department of Molecular Medicine and Surgery, Karolinska Institutet, Stockholm, Sweden
- 2Center for Molecular Medicine, Karolinska University Hospital, Stockholm, Sweden
- 3Centre for Eating Disorders Innovation, Karolinska Institutet, Stockholm, Sweden
Animal models are invaluable resources in research concerning the neurobiology of anorexia nervosa (AN), to a large extent since valid clinical samples are rare. None of the existing models can capture all aspects of AN but they are able to mirror the core features of the disorder e.g., elective starvation, emaciation and premature death. The anorectic anx/anx mouse is of particular value for the understanding of the abnormal response to negative energy balance seen in AN. These mice appear normal at birth but gradually develops starvation and emaciation despite full access to food, and die prematurely around three weeks of age. Several changes in hypothalamic neuropeptidergic and -transmitter systems involved in regulating food intake and metabolism have been documented in the anx/anx mouse. These changes are accompanied by signs of inflammation and degeneration in the same hypothalamic regions; including activation of microglia cells and expression of major histocompatibility complex I by microglia and selective neuronal populations. These aberrances are likely related to the dysfunction of complex I (CI) in the oxidative phosphorylation system of the mitochondria, and subsequent increased oxidative stress, which also has been revealed in the hypothalamus of these mice. Interestingly, a similar CI dysfunction has been shown in leukocytes from patients with AN. In addition, a higher expression of the Neurotrophic Receptor Tyrosine Kinase 3 gene has been shown in the anx/anx hypothalamus. This agrees with AN being associated with specific variants of the genes for brain derived neurotrophic factor and Neurotrophic Receptor Tyrosine Kinase 2. The anx/anx mouse is also glucose intolerant and display pancreatic dysfunction related to increased levels of circulating free fatty acids (FFA) and pancreatic inflammation. An increased incidence of eating disorders has been reported for young diabetic women, and as well has increased levels of circulating FFAs in AN. Also similar to individuals with AN, the anx/anx mouse has reduced leptin and increased cholesterol levels in serum. Thus, the anx/anx mouse shares several characteristics with patients with AN, including emaciation, starvation, premature death, diabetic features, increased FFA and low leptin, and is therefore a unique resource in research on the (neuro)biology of AN.
Introduction – Anorexia Nervosa
Anorexia nervosa (AN) is a complex psychiatric disorder affecting around 1% of females and 0.1% of males, of which as many as 10% die as a result of the disorder (Bulik et al., 2006; Keski-Rahkonen et al., 2007; Papadopoulos et al., 2009). The diagnostic criteria, according to the Diagnostic and statistical manual of mental disorders (DSMV), include persistent food intake restriction leading to significantly low body weight, combined with persistent behaviors that interfere with weight gain, and body image distortion (Schaumberg et al., 2017). One central and yet unexplained part of AN is the contradictory response to negative energy balance and the inability to ingest adequate energy, leading to severe underweight. It is indeed paradoxical that while most individuals quickly regain the weight lost from dieting (Pietilainen et al., 2012), individuals with AN stay in an emaciated state commonly for many years, some even until death. It has been speculated that hunger signals are diminished or even absent in individuals with AN, and that satiety signals on the other hand are exaggerated (DeBoer, 2011; Oberndorfer et al., 2013). Supporting this hypothesis, a genome wide association study (GWAS), as well as genetic correlation data, indicate that individuals with AN are genetically predisposed to a lower body weight set point (Duncan et al., 2017; Hinney et al., 2017). However, in order to understand the complex biology of AN, in particular the illogical response to starvation and underweight, we need to learn more about the neurobiological pathways and molecular mechanisms that are associated with severe dysregulation of food intake. This is something that is technically difficult and to some extent impossible to do in humans, since post-mortem tissues rarely are available. On the other hand, animal models cannot capture all aspects of AN but they are able to mirror the core features of the disorder e.g., elective starvation, emaciation and premature death (Siegfried et al., 2003). Animal models have therefore proved to be invaluable resources for researchers in the field. One such model is the anx/anx mouse.
The anx/anx Mouse
The homozygous anx-mouse appears normal at birth, meaning that it is indistinguishable from their homozygous and heterozygous wildtype (wt) siblings. However, during the first postnatal weeks they gradually develop the core symptoms of AN; starvation and emaciation (Figure 1). The anx/anx mouse dies prematurely around 3 weeks of age, and by then weigh around half as much as their siblings. They are able to eat, but despite full access to milk from the mother, eat significantly less already from postnatal day (P) 5. Worth to note is that the diurnal patterns in food intake seen in their healthy siblings are mirrored in the anx/anx mouse, even though the amount ingested is significantly smaller (Maltais et al., 1984). Neurological/behavioral deviations such as head weaving, hyperactivity, body tremors and uncoordinated gait, were described in the original paper by Maltais et al. (1984). When corrected for body weight, brain and thymus weights are increased compared to their healthy siblings, both at P5 and P15, while the weight of spleen is reduced (Maltais et al., 1984). See Table 1 for a summary of the aberrances in the anx/anx mouse discussed here and below.

Figure 1. An anx/anx mouse and wildtype (+/+) littermate, age 17 days.
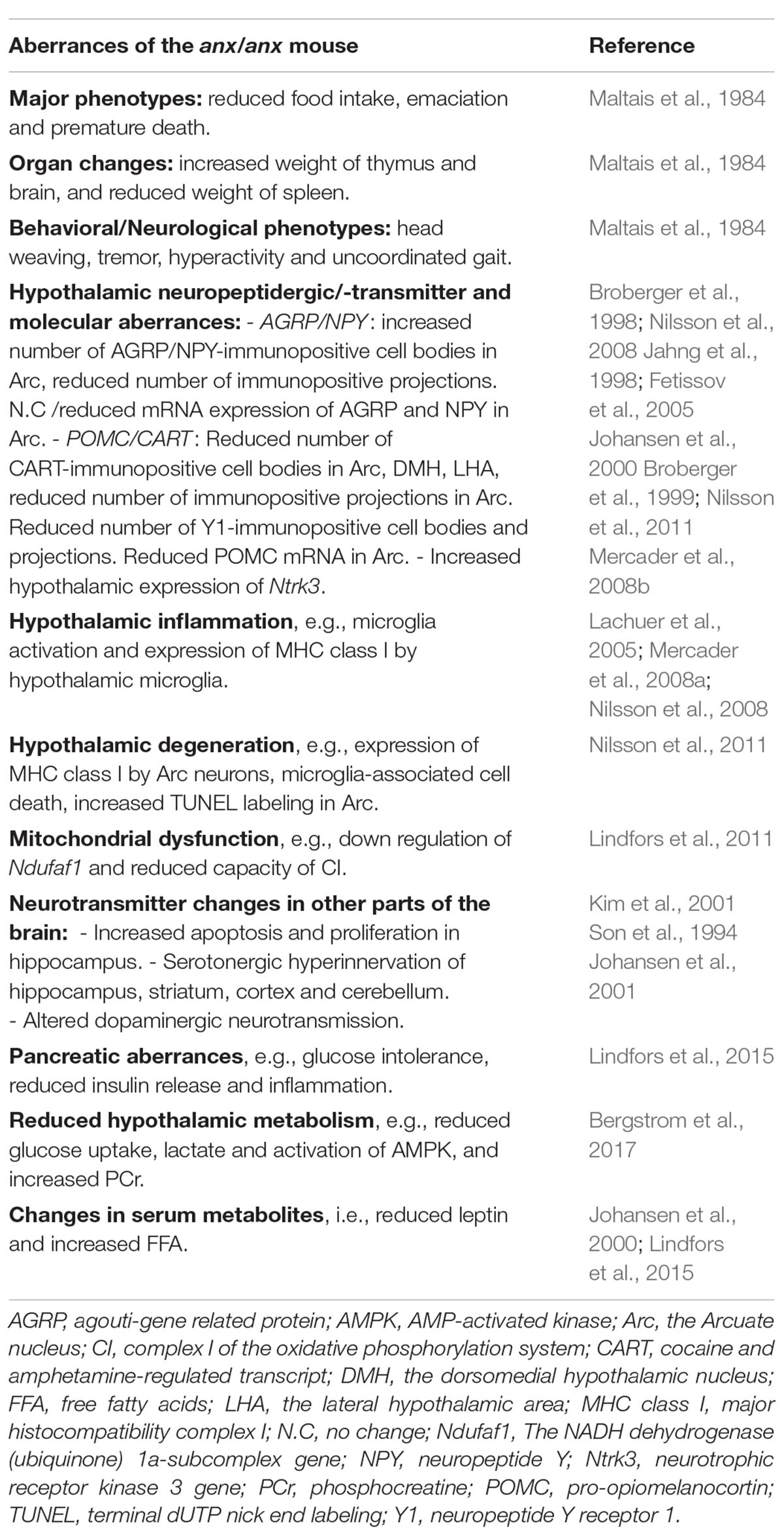
Table 1. Main characteristics of the anx/anx mouse.
The anx Mutation
The anx mutation arose spontaneously at the Jackson laboratory in Bar Harbor, Maine, already in 1976 in the F2 generation of a cross between DW/J and an inbred strain, the latter was derived from a cross between M.m.poschiavinus and an inbred Swiss strain. The male anx carrier was crossed to a female B6C3H-a/a F1 mouse, and the mutation has since then been conserved on this background (Maltais et al., 1984). We have mapped the mutation to a 0.2 cM interval residing between the markers D2Mit133 and Jojo5 chromosome 2 (Chr 2: bp 118, 889, 896–120, 175, 1081) (Lindfors et al., 2011). So far, no sequencing attempts have been able to show any unique sequence alteration. However, one needs to keep in mind that the background of the anx/anx mouse includes five different strains (see above) which makes de novo assembly difficult. The lack of unique finding could also mean that the mutation is located in a regulatory element outside the interval. The NADH dehydrogenase (ubiquinone) 1a-subcomplex (Ndufaf1) gene, shown to be closely associated with several of the anx/anx phenotypes, is however, located in the short interval of the mutation (see section on mitochondrial dysfunction below) (Lindfors et al., 2011). Ndufaf1 is an assembly factor for complex I (CI) in the mitochondrial oxidative phosphorylation system (OXPHOS) (Vogel et al., 2005). In addition, work by Kim et al. (2017) identified a point mutation in Tyro3 which they conclude is not the anx-mutation but a strain specific modifier of anx-phenotypes (Kim et al., 2017). Thus, despite that the anx/anx mouse model recently turned 40 years, the mutation is still unknown. Hopefully modern techniques within e.g., sequencing will be able to shed light on this mystery.
Neurochemistry of the anx/anx Mouse
Several changes in neuropeptidergic and -transmitter systems, in particular systems in the hypothalamus known to regulate food intake and metabolism (energy homeostasis), have been documented in the anx/anx brain (Broberger et al., 1997, 1999; Johansen et al., 2000, 2003; Nilsson et al., 2013). A part of the hypothalamus, called the Arcuate nucleus (Arc), is of particular importance concerning energy homeostasis. The Arc harbors among others a neuronal population co-expressing two orexigenic neuropeptides; agouti-gene related protein (AGRP) and neuropeptide Y (NPY), and a neuronal population co-expressing the anorexigenic peptide/precursor; pro-opiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART) (Chronwall, 1985; Cone et al., 2001; Schwartz, 2001). Aberrances have been documented in both these neuronal populations in the anx/anx mouse. Immunohistochemistry revealed increased number of NPY and AGRP immunopositive cell bodies within the Arc, combined with a reduction in AGRP/NPY immunopositive projections in the hypothalamic and extra-hypothalamic target areas of these neurons (Broberger et al., 1997, 1998; Fetissov et al., 2005; Nilsson et al., 2008). In situ hybridization studies have with regard to these neuropeptides been inconsistent, which most likely is attributed to overexposure of the labeled glass slides in the earlier studies. Thus, while initial studies documented no change in mRNA levels of NPY in the Arc of the anx/anx mouse (Broberger et al., 1997; Jahng et al., 1998), a later study showed increased mRNA for both NPY and AGRP in the anx/anx Arc (Fetissov et al., 2005). With regard to the POMC/CART population, significantly decreased levels of CART mRNA, as well as CART immunopositive cell bodies and fibers in Arc have been shown in the anx/anx hypothalamus. Also, a lower number of detectable CART-expressing cells in the dorsomedial hypothalamic nucleus/lateral hypothalamic area is seen (Johansen et al., 2000). In situ hybridization demonstrated decreased numbers of POMC-expressing neurons in the anx/anx Arc (Broberger et al., 1999). Using the neuropeptide Y receptor 1 (Y1) which outlines the soma and dendrites of POMC/CART neurons (Zhang et al., 1994; Kopp et al., 2002), markedly reduced immunoreactivity in Arc and the paraventricular nucleus of hypothalamus was revealed (Broberger et al., 1999; Nilsson et al., 2011). Clinically, genetic variants of AGRP have been associated with AN (Dardennes et al., 2007) or with lowest BMI during AN illness (Yilmaz et al., 2014). Increased plasma levels of the peptide have been documented in AN (Moriya et al., 2006), but it is so far unknown if this change remains after weight recovery. The changed cerebrospinal fluid levels of NPY seen in AN is however, known to be secondary to the illness (Gendall et al., 1999).
In addition, an increased expression of the neurotrophic receptor kinase 3 (Ntrk3) gene has been shown in the anx/anx hypothalamus (Mercader et al., 2008b). This agrees with AN being associated with specific variants of the genes for brain derived neurotrophic factor (BDNF) and neurotrophic receptor tyrosine kinase 2 (NTRK2) (Ribases et al., 2003, 2005).
Changes have been documented also in other brain regions than the hypothalamus. Increased apoptosis and proliferation in the dentate gyrus of the hippocampus (Kim et al., 2001), serotonergic hyperinnervation in hippocampus, cortex, olfactory bulb and cerebellum (Son et al., 1994), as well as altered dopaminergic transmission in the striatum (Johansen et al., 2001), have been demonstrated. Genetic variants as well as deviant levels of metabolites and receptors related to dopamine and serotonin have been linked to the AN pathology (Kaye et al., 1999, 2005; Kaye, 2008).
Neuroinflammation and Degeneration in the anx/anx Hypothalamus
The hypothalamic neurochemical aberrances of the anx/anx mouse are accompanied by signs of inflammation and degeneration (Lachuer et al., 2005; Mercader et al., 2008a; Nilsson et al., 2008, 2011). Microglia cells are immunocompetent cells that are activated in the central nervous system in response to e.g., inflammation, neurodegeneration or injury (Nakajima and Kohsaka, 2004; Streit et al., 2005). In the anx/anx brain, microglia are activated selectively in the hypothalamic regions where the neurons, both cell bodies and projections, expressing the orexigenic neuropeptide AGRP are located (Nilsson et al., 2008). The first appearance of activated microglia overlaps in time with the loss of AGRP immunoreactive projections, i.e., P12–15 (Nilsson et al., 2008). Similarly, chemical ablation of the AGRP neurons results in starvation in both normal weight and obese mice, and results in glia (microglia and astroglia) activation in the target areas (Wu et al., 2008, 2012). Major histocompatibility complex I is expressed by the activated microglia, but also by the AGRP and POMC expressing neurons in the anx/anx brain (Nilsson et al., 2011). This latter finding combined with increased hypothalamic terminal dUTP nick end labeling (TUNEL) labeling and so called microglia-associated cell death (Ribak et al., 2009), made us conclude that hypothalamic degeneration is associated with the anorexia of the anx/anx mouse (Nilsson et al., 2011). In addition, two microarray studies of the anx/anx hypothalamus revealed changed expression of an enrichment of genes involved in inflammation and cell death (Lachuer et al., 2005; Mercader et al., 2008a). While it is unknown if hypothalamic inflammation occurs in AN, it has been linked to cachexia, the anorexia that often accompanies chronic illnesses such as cancer and HIV (Durham et al., 2009; Dwarkasing et al., 2016).
Mitochondrial CI Dysfunction and Reduced Hypothalamic Metabolism
A dysfunction selective of CI in OXPHOS, and subsequent increased oxidative stress, have been revealed in the hypothalamus of the anx/anx mouse (Lindfors et al., 2011). This CI dysfunction is connected to down regulation of the gene Ndufaf1 which in fact is located in the anx interval (see section on the anx mutation above). The down regulation has been confirmed at the protein level at P21 (Lindfors et al., 2011). Ndufaf1 encodes one of several proteins crucial for the correct assembly of the 44–46 proteins that build up CI (Smeitink et al., 2001; Ugalde et al., 2004a,b; Guerrero-Castillo et al., 2017). Selective neuronal damage and glia activation, as shown in the anx/anx mouse (Nilsson et al., 2008, 2011), has been shown in another animal model with CI deficiencies, i.e., the Ndufs4-KO mouse (Quintana et al., 2010). The NDUFAF1 gene, as well as other players in CI biogenesis, have been implicated in human pathology; resulting in e.g., leukodystrophy and failure to thrive in young children (Vogel et al., 2005, 2007; Dunning et al., 2007; Distelmaier et al., 2009). In fact, CI dysfunction has been shown in leukocytes from patients with AN (Victor et al., 2014), but it remains to be explored if this is a cause or consequence of the disorder. This far, the NDUFAF1 gene has not been associated with AN, but it would be worth exploring genetics variants related to OXPHOS function and a potential association with AN, similar to what has been shown in other psychiatric disorders e.g., autism spectrum disorder (Giulivi et al., 2010). With this saying the anx/anx model is a model of value for research on all human conditions with loss of appetite i.e., anorexia, including the anorexia seen in cachexia and failure to thrive, as well as AN. The anx/anx mouse is unique in the sense that few other models exist were the mice, similarly to the human conditions just mentioned, eat insufficient despite having full access to food. This in contrast to models were the researcher in one way or another limits the access of food (Siegfried et al., 2003).
Diseases associated with mitochondrial dysfunction are commonly associated with a stressed metabolic profile, and hypermetabolism (Wredenberg et al., 2006; Jeppesen et al., 2007; Milone and Wong, 2013). Supposedly such metabolic responses occur in order to safeguard adequate levels of ATP. In some cases, conversely, mitochondrial dysfunction is associated with reduced glucose uptake and hypometabolism, e.g., in Alzheimer’s disease and epilepsy (Chandrasekaran et al., 1996; Tenney et al., 2014). This resembles what we saw in the anx/anx hypothalamus, i.e., lower glucose uptake rate, decreased lactate content, as well as elevated phosphocreatine (PCr) content and reduced activation of AMP-activated kinase (AMPK) in the basal state (Bergstrom et al., 2017). This is similar to the hypometabolic state seen in hibernation (Healy et al., 2011) and could be reflecting lower neuronal activity (Cunnane et al., 2011). Different neuronal populations respond differently to this type of metabolic stress (Schreiber and Baudry, 1995), which has been ascribed to the subtype of ATP-sensitive potassium channel (K-ATP) they express. A specific subtype of K-ATP channel that consists of Kir 6.2 and SUR1 subunits becomes activated by mitochondrial CI dysfunction, i.e., by increased ROS levels and/or reduced levels of ATP. This leads to ceased electrical activity, hyperpolarization and reduced firing, in a means of protecting the cell from the energy deficiency and increased oxidative stress (Liss et al., 1999). Kir6.2/SUR1 K-ATP channels are expressed by the hypothalamic POMC/CART and AGRP/NPY neurons, and by a limited number of other cell populations including the pancreatic beta-cells and dopaminergic neurons in Substantia Nigra (Miki et al., 2001; Ibrahim et al., 2003; van den Top and Spanswick, 2006; van den Top et al., 2007). Firing of action potentials and release of neurotransmitters are processes that require high amounts of energy. Therefore, inhibition of these processes would conserve energy during conditions when energy is scarce (Attwell and Laughlin, 2001; Sengupta et al., 2010). In addition, uncontrolled generation of ROS, commonly accompanying CI dysfunction, can also cause diminished firing of the AGRP/NPY neurons, thus resulting in a reduced orexigenic drive (Andrews et al., 2008; Horvath et al., 2009).
Pancreatic Dysfunction and Aberrant Levels of Fat Derived Molecules
The anx/anx mouse also displays a pancreatic dysfunction (Lindfors et al., 2015). More specifically, they are markedly glucose intolerant, and show reduced insulin release upon glucose tolerance test. This is associated with elevated serum concentrations of free fatty acids (FFAs) in the anx/anx mouse and increased macrophage infiltration [indicative of inflammation (Imai et al., 1996; Ka et al., 2015)] of anx/anx islets. Increased levels of FFAs have been connected to inhibition of glucose-induced insulin secretion (Eguchi et al., 2012). Interestingly, isolated anx/anx islets cultured in the absence of FFAs show increased insulin release upon glucose stimulation and show no signs of inflammation. Thus, the diabetic phenotype of the anx/anx mouse seems to be related to the elevated FFAs and inflammation in pancreatic islets. This finding is interesting in the light of the increased incidence of eating disorders that has been reported in young women with diabetes (Hudson et al., 1985; Meltzer et al., 2001), and documented increased levels of circulating FFAs in AN (Pinter et al., 1975; Curatola et al., 2004). Also similar to individuals with AN, the anx/anx mouse has low levels of the fat derived and food intake regulating hormone leptin, and high levels of cholesterol in serum (Maltais et al., 1984; Schorr and Miller, 2017).
Conclusion and Future Perspective
The anx/anx mouse shares several characteristics with patients with AN, including emaciation, starvation, premature death, diabetic features, increased FFA and low leptin, and is therefore a unique and very valuable resource in order to explore and understand the (neuro)biology of AN. Future research should explore if hypothalamic inflammation and/or degeneration, as seen in the anx/anx mouse, are mechanisms involved also in AN. Further studies are also needed in order to clarify if the mitochondrial dysfunction seen in AN (Victor et al., 2014) is a cause or consequence of the disorder. Finally, it would be of value to be able to define the anx-mutation, as well as explore other brain areas related to food intake regulation, e.g., nucleus tractus solitarius and the parabrachial nucleus in the anx/anx mouse.
Author Contributions
IN reviewed the literature, wrote, and edited the manuscript.
Funding
The work presented in this paper was supported by funding from the Karolinska Institutet (KI), the Karolinska University Hospital, Hjärnfonden/Swedish Brain Foundation, a NARSAD YI Grant from The Brain and Behavior Research Foundation and the following Foundations: Magnus Bergvall, Tore Nilsson, Thuring, Mjölkdroppen Märtha Lundqvist and Eva and Oscar Ahrén.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
References
Andrews, Z. B., Liu, Z. W., Walllingford, N., Erion, D. M., Borok, E., Friedman, J. M., et al. (2008). UCP2 mediates ghrelin’s action on NPY/AgRP neurons by lowering free radicals. Nature 454, 846–851. doi: 10.1038/nature07181
Attwell, D., and Laughlin, S. B. (2001). An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 21, 1133–1145. doi: 10.1097/00004647-200110000-00001
Bergstrom, U., Lindfors, C., Svedberg, M., Johansen, J. E., Haggkvist, J., Schalling, M., et al. (2017). Reduced metabolism in the hypothalamus of the anorectic anx/anx mouse. J. Endocrinol. 233, 15–24. doi: 10.1530/JOE-16-0383
Broberger, C., Johansen, J., Brismar, H., Johansson, C., Schalling, M., and Hokfelt, T. (1999). Changes in neuropeptide Y receptors and pro-opiomelanocortin in the anorexia (anx/anx) mouse hypothalamus. J. Neurosci. 19, 7130–7139. doi: 10.1523/JNEUROSCI.19-16-07130.1999
Broberger, C., Johansen, J., Johansson, C., Schalling, M., and Hokfelt, T. (1998). The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice. Proc. Natl. Acad. Sci. U.S.A. 95, 15043–15048. doi: 10.1073/pnas.95.25.15043
Broberger, C., Johansen, J., Schalling, M., and Hokfelt, T. (1997). Hypothalamic neurohistochemistry of the murine anorexia (anx/anx) mutation: altered processing of neuropeptide Y in the arcuate nucleus. J. Comp. Neurol. 387, 124–135. doi: 10.1002/(SICI)1096-9861(19971013)387:1<124::AID-CNE10>3.0.CO;2-U
Bulik, C. M., Sullivan, P. F., Tozzi, F., Furberg, H., Lichtenstein, P., and Pedersen, N. L. (2006). Prevalence, heritability, and prospective risk factors for anorexia nervosa. Arch. Gen. Psychiatry 63, 305–312. doi: 10.1001/archpsyc.63.3.305
Chandrasekaran, K., Hatanpaa, K., Brady, D. R., and Rapoport, S. I. (1996). Evidence for physiological down-regulation of brain oxidative phosphorylation in Alzheimer’s disease. Exp. Neurol. 142, 80–88. doi: 10.1006/exnr.1996.0180
Chronwall, B. M. (1985). Anatomy and physiology of the neuroendocrine arcuate nucleus. Peptides 6(Suppl. 2), 1–11. doi: 10.1016/0196-9781(85)90128-7
Cone, R. D., Cowley, M. A., Butler, A. A., Fan, W., Marks, D. L., and Low, M. J. (2001). The arcuate nucleus as a conduit for diverse signals relevant to energy homeostasis. Int. J. Obes. Relat. Metab. Disord. 25(Suppl. 5), S63–S67. doi: 10.1038/sj.ijo.0801913
Cunnane, S., Nugent, S., Roy, M., Courchesne-Loyer, A., Croteau, E., Tremblay, S., et al. (2011). Brain fuel metabolism, aging, and Alzheimer’s disease. Nutrition 27, 3–20. doi: 10.1016/j.nut.2010.07.021
Curatola, G., Camilloni, M. A., Vignini, A., Nanetti, L., Boscaro, M., and Mazzanti, L. (2004). Chemical-physical properties of lipoproteins in anorexia nervosa. Eur. J. Clin. Invest. 34, 747–751. doi: 10.1111/j.1365-2362.2004.01415.x
Dardennes, R. M., Zizzari, P., Tolle, V., Foulon, C., Kipman, A., Romo, L., et al. (2007). Family trios analysis of common polymorphisms in the obestatin/ghrelin, BDNF and AGRP genes in patients with Anorexia nervosa: association with subtype, body-mass index, severity and age of onset. Psychoneuroendocrinology 32, 106–113. doi: 10.1016/j.psyneuen.2006.11.003
DeBoer, M. D. (2011). What can anorexia nervosa teach us about appetite regulation? Nutrition 27, 405–406. doi: 10.1016/j.nut.2011.02.001
Distelmaier, F., Koopman, W. J., van den Heuvel, L. P., Rodenburg, R. J., Mayatepek, E., Willems, P. H., et al. (2009). Mitochondrial complex I deficiency: from organelle dysfunction to clinical disease. Brain 132(Pt 4), 833–842. doi: 10.1093/brain/awp058
Duncan, L., Yilmaz, Z., Gaspar, H., Walters, R., Goldstein, J., Anttila, V., et al. (2017). Significant locus and metabolic genetic correlations revealed in genome-wide association study of anorexia nervosa. Am. J. Psychiatry 174, 850–858. doi: 10.1176/appi.ajp.2017.16121402
Dunning, C. J., McKenzie, M., Sugiana, C., Lazarou, M., Silke, J., Connelly, A., et al. (2007). Human CIA30 is involved in the early assembly of mitochondrial complex I and mutations in its gene cause disease. EMBO J. 26, 3227–3237. doi: 10.1038/sj.emboj.7601748
Durham, W. J., Dillon, E. L., and Sheffield-Moore, M. (2009). Inflammatory burden and amino acid metabolism in cancer cachexia. Curr. Opin. Clin. Nutr. Metab. Care 12, 72–77. doi: 10.1097/MCO.0b013e32831cef61
Dwarkasing, J. T., Marks, D. L., Witkamp, R. F., and van Norren, K. (2016). Hypothalamic inflammation and food intake regulation during chronic illness. Peptides 77, 60–66. doi: 10.1016/j.peptides.2015.06.011
Eguchi, K., Manabe, I., Oishi-Tanaka, Y., Ohsugi, M., Kono, N., Ogata, F., et al. (2012). Saturated fatty acid and TLR signaling link beta cell dysfunction and islet inflammation. Cell Metab. 15, 518–533. doi: 10.1016/j.cmet.2012.01.023
Fetissov, S. O., Bergstrom, U., Johansen, J. E., Hokfelt, T., Schalling, M., and Ranscht, B. (2005). Alterations of arcuate nucleus neuropeptidergic development in contactin-deficient mice: comparison with anorexia and food-deprived mice. Eur. J. Neurosci. 22, 3217–3228. doi: 10.1111/j.1460-9568.2005.04513.x
Gendall, K. A., Kaye, W. H., Altemus, M., McConaha, C. W., and La Via, M. C. (1999). Leptin, neuropeptide Y, and peptide YY in long-term recovered eating disorder patients. Biol. Psychiatry 46, 292–299. doi: 10.1016/S0006-3223(98)00292-3
Giulivi, C., Zhang, Y. F., Omanska-Klusek, A., Ross-Inta, C., Wong, S., Hertz-Picciotto, I., et al. (2010). Mitochondrial dysfunction in autism. JAMA 304, 2389–2396. doi: 10.1001/jama.2010.1706
Guerrero-Castillo, S., Baertling, F., Kownatzki, D., Wessels, H. J., Arnold, S., Brandt, U., et al. (2017). The assembly pathway of mitochondrial respiratory chain complex I. Cell Metab. 25, 128–139. doi: 10.1016/j.cmet.2016.09.002
Healy, J. E., Gearhart, C. N., Bateman, J. L., Handa, R. J., and Florant, G. L. (2011). AMPK and ACCchange with fasting and physiological condition in euthermic and hibernating golden-mantled ground squirrels (Callospermophilus lateralis). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 159, 322–331. doi: 10.1016/j.cbpa.2011.03.026
Hinney, A., Kesselmeier, M., Jall, S., Volckmar, A. L., Focker, M., Antel, J., et al. (2017). Evidence for three genetic loci involved in both anorexia nervosa risk and variation of body mass index. Mol. Psychiatry 22, 321–322. doi: 10.1038/mp.2016.126
Horvath, T. L., Andrews, Z. B., and Diano, S. (2009). Fuel utilization by hypothalamic neurons: roles for ROS. Trends Endocrinol. Metab. 20, 78–87. doi: 10.1016/j.tem.2008.10.003
Hudson, J. I., Wentworth, S. M., Hudson, M. S., and Pope, H. G. Jr (1985). Prevalence of anorexia nervosa and bulimia among young diabetic women. J. Clin. Psychiatry 46, 88–89.
Ibrahim, N., Bosch, M. A., Smart, J. L., Qiu, J., Rubinstein, M., Ronnekleiv, O. K., et al. (2003). Hypothalamic proopiomelanocortin neurons are glucose responsive and express K(ATP) channels. Endocrinology 144, 1331–1340. doi: 10.1210/en.2002-221033
Imai, Y., Ibata, I., Ito, D., Ohsawa, K., and Kohsaka, S. (1996). A novel gene iba1 in the major histocompatibility complex class III region encoding an EF hand protein expressed in a monocytic lineage. Biochem. Biophys. Res. Commun. 224, 855–862. doi: 10.1006/bbrc.1996.1112
Jahng, J. W., Houpt, T. A., Kim, S. J., Joh, T. H., and Son, J. H. (1998). Neuropeptide Y mRNA and serotonin innervation in the arcuate nucleus of anorexia mutant mice. Brain Res. 790, 67–73. doi: 10.1016/S0006-8993(98)00049-3
Jeppesen, T. D., Quistorff, B., Wibrand, F., and Vissing, J. (2007). 31P-MRS of skeletal muscle is not a sensitive diagnostic test for mitochondrial myopathy. J. Neurol. 254, 29–37. doi: 10.1007/s00415-006-0229-5
Johansen, J. E., Broberger, C., Lavebratt, C., Johansson, C., Kuhar, M. J., Hokfelt, T., et al. (2000). Hypothalamic CART and serum leptin levels are reduced in the anorectic (anx/anx) mouse. Brain Res. Mol. Brain Res. 84, 97–105. doi: 10.1016/S0169-328X(00)00228-X
Johansen, J. E., Fetissov, S., Fischer, H., Arvidsson, S., Hokfelt, T., and Schalling, M. (2003). Approaches to anorexia in rodents: focus on the anx/anx mouse. Eur. J. Pharmacol. 480, 171–176. doi: 10.1016/j.ejphar.2003.08.104
Johansen, J. E., Teixeira, V. L., Johansson, C., Serrao, P., Berggren, P. O., Soares-Da-Silva, P., et al. (2001). Altered dopaminergic transmission in the anorexic anx/anx mouse striatum. Neuroreport 12, 2737–2741. doi: 10.1097/00001756-200108280-00029
Ka, S. O., Song, M. Y., Bae, E. J., and Park, B. H. (2015). Myeloid SIRT1 regulates macrophage infiltration and insulin sensitivity in mice fed a high-fat diet. J. Endocrinol. 224, 109–118. doi: 10.1530/JOE-14-0527
Kaye, W. (2008). Neurobiology of anorexia and bulimia nervosa. Physiol. Behav. 94, 121–135. doi: 10.1016/j.physbeh.2007.11.037
Kaye, W. H., Frank, G. K., Bailer, U. F., Henry, S. E., Meltzer, C. C., Price, J. C., et al. (2005). Serotonin alterations in anorexia and bulimia nervosa: new insights from imaging studies. Physiol. Behav. 85, 73–81. doi: 10.1016/j.physbeh.2005.04.013
Kaye, W. H., Frank, G. K., and McConaha, C. (1999). Altered dopamine activity after recovery from restricting-type anorexia nervosa. Neuropsychopharmacology 21, 503–506. doi: 10.1016/S0893-133X(99)00053-6
Keski-Rahkonen, A., Hoek, H. W., Susser, E. S., Linna, M. S., Sihvola, E., Raevuori, A., et al. (2007). Epidemiology and course of anorexia nervosa in the community. Am. J. Psychiatry 164, 1259–1265. doi: 10.1176/appi.ajp.2007.06081388
Kim, D. Y., Yu, J., Mui, R. K., Niibori, R., Taufique, H. B., Aslam, R., et al. (2017). The tyrosine kinase receptor Tyro3 enhances lifespan and neuropeptide Y (Npy) neuron survival in the mouse anorexia (anx) mutation. Dis. Model. Mech. 10, 581–595. doi: 10.1242/dmm.027433
Kim, M. J., Kim, Y., Kim, S. A., Lee, H. J., Choe, B. K., Nam, M., et al. (2001). Increases in cell proliferation and apoptosis in dentate gyrus of anorexia (anx/anx) mice. Neurosci. Lett. 302, 109–112. doi: 10.1016/S0304-3940(01)01684-6
Kopp, J., Xu, Z. Q., Zhang, X., Pedrazzini, T., Herzog, H., Kresse, A., et al. (2002). Expression of the neuropeptide Y Y1 receptor in the CNS of rat and of wild-type and Y1 receptor knock-out mice. Focus on immunohistochemical localization. Neuroscience 111, 443–532. doi: 10.1016/S0306-4522(01)00463-8
Lachuer, J., Ouyang, L., Legras, C., Del Rio, J., and Barlow, C. (2005). Gene expression profiling reveals an inflammatory process in the anx/anx mutant mice. Brain Res. Mol. Brain Res. 139, 372–376. doi: 10.1016/j.molbrainres.2005.06.003
Lindfors, C., Katz, A., Selander, L., Johansen, J. E., Marconi, G., Schalling, M., et al. (2015). Glucose intolerance and pancreatic beta-cell dysfunction in the anorectic anx/anx mouse. Am. J. Physiol. Endocrinol. Metab. 309, E418–E427. doi: 10.1152/ajpendo.00081.2015
Lindfors, C., Nilsson, I. A., Garcia-Roves, P. M., Zuberi, A. R., Karimi, M., Donahue, L. R., et al. (2011). Hypothalamic mitochondrial dysfunction associated with anorexia in the anx/anx mouse. Proc. Natl. Acad. Sci. U.S.A. 108, 18108–18113. doi: 10.1073/pnas.1114863108
Liss, B., Bruns, R., and Roeper, J. (1999). Alternative sulfonylurea receptor expression defines metabolic sensitivity of K-ATP channels in dopaminergic midbrain neurons. EMBO J. 18, 833–846. doi: 10.1093/emboj/18.4.833
Maltais, L. J., Lane, P. W., and Beamer, W. G. (1984). Anorexia, a recessive mutation causing starvation in preweanling mice. J. Hered. 75, 468–472. doi: 10.1093/oxfordjournals.jhered.a109987
Meltzer, L. J., Johnson, S. B., Prine, J. M., Banks, R. A., Desrosiers, P. M., and Silverstein, J. H. (2001). Disordered eating, body mass, and glycemic control in adolescents with type 1 diabetes. Diabetes Care 24, 678–682. doi: 10.2337/diacare.24.4.678
Mercader, J. M., Lozano, J. J., Sumoy, L., Dierssen, M., Visa, J., Gratacos, M., et al. (2008a). Hypothalamus transcriptome profile suggests an anorexia-cachexia syndrome in the anx/anx mouse model. Physiol. Genom. 35, 341–350. doi: 10.1152/physiolgenomics.90255.2008
Mercader, J. M., Saus, E., Aguera, Z., Bayes, M., Boni, C., Carreras, A., et al. (2008b). Association of NTRK3 and its interaction with NGF suggest an altered cross-regulation of the neurotrophin signaling pathway in eating disorders. Hum. Mol. Genet. 17, 1234–1244. doi: 10.1093/hmg/ddn013
Miki, T., Liss, B., Minami, K., Shiuchi, T., Saraya, A., Kashima, Y., et al. (2001). ATP-sensitive K+ channels in the hypothalamus are essential for the maintenance of glucose homeostasis. Nat. Neurosci. 4, 507–512. doi: 10.1038/87455
Milone, M., and Wong, L. J. (2013). Diagnosis of mitochondrial myopathies. Mol. Genet. Metab. 110, 35–41. doi: 10.1016/j.ymgme.2013.07.007
Moriya, J., Takimoto, Y., Yoshiuchi, K., Shimosawa, T., and Akabayashi, A. (2006). Plasma agouti-related protein levels in women with anorexia nervosa. Psychoneuroendocrinology 31, 1057–1061. doi: 10.1016/j.psyneuen.2006.06.006
Nakajima, K., and Kohsaka, S. (2004). Microglia: neuroprotective and neurotrophic cells in the central nervous system. Curr. Drug Targets Cardiovasc. Haematol. Disord. 4, 65–84.
Nilsson, I., Lindfors, C., Fetissov, S. O., Hokfelt, T., and Johansen, J. E. (2008). Aberrant agouti-related protein system in the hypothalamus of the anx/anx mouse is associated with activation of microglia. J. Comp. Neurol. 507, 1128–1140. doi: 10.1002/cne.21599
Nilsson, I. A., Lindfors, C., Schalling, M., Hokfelt, T., and Johansen, J. E. (2013). Anorexia and hypothalamic degeneration. Vitam. Horm. 92, 27–60. doi: 10.1016/B978-0-12-410473-0.00002-7
Nilsson, I. A., Thams, S., Lindfors, C., Bergstrand, A., Cullheim, S., Hokfelt, T., et al. (2011). Evidence of hypothalamic degeneration in the anorectic anx/anx mouse. Glia 59, 45–57. doi: 10.1002/glia.21075
Oberndorfer, T. A., Frank, G. K., Simmons, A. N., Wagner, A., McCurdy, D., Fudge, J. L., et al. (2013). Altered insula response to sweet taste processing after recovery from anorexia and bulimia nervosa. Am. J. Psychiatry 170, 1143–1151. doi: 10.1176/appi.ajp.2013.11111745
Papadopoulos, F. C., Ekbom, A., Brandt, L., and Ekselius, L. (2009). Excess mortality, causes of death and prognostic factors in anorexia nervosa. Br. J. Psychiatry 194, 10–17. doi: 10.1192/bjp.bp.108.054742
Pietilainen, K. H., Saarni, S. E., Kaprio, J., and Rissanen, A. (2012). Does dieting make you fat? A twin study. Int. J. Obes. 36, 456–464. doi: 10.1038/ijo.2011.160
Pinter, E. J., Tolis, G., and Friesen, H. G. (1975). L-dopa, growth hormone and adipokinesis in the lean and the obese. Int. J. Clin. Pharmacol. Biopharm. 12, 277–280.
Quintana, A., Kruse, S. E., Kapur, R. P., Sanz, E., and Palmiter, R. D. (2010). Complex I deficiency due to loss of Ndufs4 in the brain results in progressive encephalopathy resembling Leigh syndrome. Proc. Natl. Acad. Sci. U.S.A. 107, 10996–11001. doi: 10.1073/pnas.1006214107
Ribak, C. E., Shapiro, L. A., Perez, Z. D., and Spigelman, I. (2009). Microglia-associated granule cell death in the normal adult dentate gyrus. Brain Struct. Funct. 214, 25–35. doi: 10.1007/s00429-009-0231-7
Ribases, M., Gratacos, M., Armengol, L., de Cid, R., Badia, A., Jimenez, L., et al. (2003). Met66 in the brain-derived neurotrophic factor (BDNF) precursor is associated with anorexia nervosa restrictive type. Mol. Psychiatry 8, 745–751. doi: 10.1038/sj.mp.4001281
Ribases, M., Gratacos, M., Badia, A., Jimenez, L., Solano, R., Vallejo, J., et al. (2005). Contribution of NTRK2 to the genetic susceptibility to anorexia nervosa, harm avoidance and minimum body mass index. Mol. Psychiatry 10, 851–860. doi: 10.1038/sj.mp.4001670
Schaumberg, K., Welch, E., Breithaupt, L., Hubel, C., Baker, J. H., Munn-Chernoff, M. A., et al. (2017). The science behind the academy for eating disorders’ nine truths about eating disorders. Eur. Eat. Disord. Rev. 25, 432–450. doi: 10.1002/erv.2553
Schorr, M., and Miller, K. K. (2017). The endocrine manifestations of anorexia nervosa: mechanisms and management. Nat. Rev. Endocrinol. 13, 174–186. doi: 10.1038/nrendo.2016.175
Schreiber, S. S., and Baudry, M. (1995). Selective neuronal vulnerability in the hippocampus–a role for gene expression? Trends Neurosci. 18, 446–451.
Schwartz, M. W. (2001). Brain pathways controlling food intake and body weight. Exp. Biol. Med. 226, 978–981. doi: 10.1177/153537020122601103
Sengupta, B., Stemmler, M., Laughlin, S. B., and Niven, J. E. (2010). Action potential energy efficiency varies among neuron types in vertebrates and invertebrates. PLoS Comput. Biol. 6:e1000840. doi: 10.1371/journal.pcbi.1000840
Siegfried, Z., Berry, E. M., Hao, S., and Avraham, Y. (2003). Animal models in the investigation of anorexia. Physiol. Behav. 79, 39–45. doi: 10.1016/S0031-9384(03)00103-3
Smeitink, J., van den Heuvel, L., and DiMauro, S. (2001). The genetics and pathology of oxidative phosphorylation. Nat. Rev. Genet. 2, 342–352. doi: 10.1038/35072063
Son, J. H., Baker, H., Park, D. H., and Joh, T. H. (1994). Drastic and selective hyperinnervation of central serotonergic neurons in a lethal neurodevelopmental mouse mutant, Anorexia (anx). Brain Res. Mol. Brain Res. 25, 129–134. doi: 10.1016/0169-328X(94)90287-9
Streit, W. J., Conde, J. R., Fendrick, S. E., Flanary, B. E., and Mariani, C. L. (2005). Role of microglia in the central nervous system’s immune response. Neurol. Res. 27, 685–691. doi: 10.1179/016164105X49463
Tenney, J. R., Rozhkov, L., Horn, P., Miles, L., and Miles, M. V. (2014). Cerebral glucose hypometabolism is associated with mitochondrial dysfunction in patients with intractable epilepsy and cortical dysplasia. Epilepsia 55, 1415–1422. doi: 10.1111/epi.12731
Ugalde, C., Janssen, R. J., van den Heuvel, L. P., Smeitink, J. A., and Nijtmans, L. G. (2004a). Differences in assembly or stability of complex I and other mitochondrial OXPHOS complexes in inherited complex I deficiency. Hum. Mol. Genet. 13, 659–667. doi: 10.1093/hmg/ddh071
Ugalde, C., Vogel, R., Huijbens, R., Van Den Heuvel, B., Smeitink, J., and Nijtmans, L. (2004b). Human mitochondrial complex I assembles through the combination of evolutionary conserved modules: a framework to interpret complex I deficiencies. Hum. Mol. Genet. 13, 2461–2472.
van den Top, M., Lyons, D. J., Lee, K., Coderre, E., Renaud, L. P., and Spanswick, D. (2007). Pharmacological and molecular characterization of ATP-sensitive K(+) conductances in CART and NPY/AgRP expressing neurons of the hypothalamic arcuate nucleus. Neuroscience 144, 815–824. doi: 10.1016/j.neuroscience.2006.09.059
van den Top, M., and Spanswick, D. (2006). Integration of metabolic stimuli in the hypothalamic arcuate nucleus. Prog. Brain Res. 153, 141–154. doi: 10.1016/S0079-6123(06)53008-0
Victor, V. M., Rovira-Llopis, S., Saiz-Alarcon, V., Sanguesa, M. C., Rojo-Bofill, L., Banuls, C., et al. (2014). Altered mitochondrial function and oxidative stress in leukocytes of anorexia nervosa patients. PLoS One 9:e106463. doi: 10.1371/journal.pone.0106463
Vogel, R. O., Janssen, R. J., Ugalde, C., Grovenstein, M., Huijbens, R. J., Visch, H. J., et al. (2005). Human mitochondrial complex I assembly is mediated by NDUFAF1. FEBS J. 272, 5317–5326. doi: 10.1111/j.1742-4658.2005.04928.x
Vogel, R. O., Janssen, R. J., van den Brand, M. A., Dieteren, C. E., Verkaart, S., Koopman, W. J., et al. (2007). Cytosolic signaling protein Ecsit also localizes to mitochondria where it interacts with chaperone NDUFAF1 and functions in complex I assembly. Genes Dev. 21, 615–624. doi: 10.1101/gad.408407
Wredenberg, A., Freyer, C., Sandstrom, M. E., Katz, A., Wibom, R., Westerblad, H., et al. (2006). Respiratory chain dysfunction in skeletal muscle does not cause insulin resistance. Biochem. Biophys. Res. Commun. 350, 202–207. doi: 10.1016/j.bbrc.2006.09.029
Wu, Q., Howell, M. P., and Palmiter, R. D. (2008). Ablation of neurons expressing agouti-related protein activates fos and gliosis in postsynaptic target regions. J. Neurosci. 28, 9218–9226. doi: 10.1523/JNEUROSCI.2449-08.2008
Wu, Q., Whiddon, B. B., and Palmiter, R. D. (2012). Ablation of neurons expressing agouti-related protein, but not melanin concentrating hormone, in leptin-deficient mice restores metabolic functions and fertility. Proc. Natl. Acad. Sci. U.S.A. 109, 3155–3160. doi: 10.1073/pnas.1120501109
Yilmaz, Z., Kaplan, A. S., Tiwari, A. K., Levitan, R. D., Piran, S., Bergen, A. W., et al. (2014). The role of leptin, melanocortin, and neurotrophin system genes on body weight in anorexia nervosa and bulimia nervosa. J. Psychiatr. Res. 55, 77–86. doi: 10.1016/j.jpsychires.2014.04.005
Zhang, X., Bao, L., Xu, Z. Q., Kopp, J., Arvidsson, U., Elde, R., et al. (1994). Localization of neuropeptide Y Y1 receptors in the rat nervous system with special reference to somatic receptors on small dorsal root ganglion neurons. Proc. Natl. Acad. Sci. U.S.A. 91, 11738–11742. doi: 10.1073/pnas.91.24.11738
Keywords: hypothalamus, anorexia, inflammation, neurodegeneration, neuropeptide, AGRP, microglia
Citation: Nilsson IAK (2019) The anx/anx Mouse – A Valuable Resource in Anorexia Nervosa Research. Front. Neurosci. 13:59. doi: 10.3389/fnins.2019.00059
Received: 29 November 2018; Accepted: 21 January 2019;
Published: 05 February 2019.
Edited by:
Virginie Tolle, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Andreas Stengel, University of Tübingen, GermanyNicolas Chartrel, Institut National de la Santé et de la Recherche Médicale (INSERM), France
Copyright © 2019 Nilsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ida A. K. Nilsson, aWRhLm5pbHNzb25Aa2kuc2U=