 Farrokh Mansouri1
Farrokh Mansouri1 Alaa Shanbour2,3
Alaa Shanbour2,3 Frank Mazza4
Frank Mazza4 Peter Fettes3
Peter Fettes3 José Zariffa1,5,6,7
José Zariffa1,5,6,7 Jonathan Downar3,8,9,10*
Jonathan Downar3,8,9,10*- 1Institute of Biomaterials and Biomedical Engineering, University of Toronto, Toronto, ON, Canada
- 2Department of Psychiatry, Central Michigan University, Mount Pleasant, MI, United States
- 3Institute of Medical Science, University of Toronto, Toronto, ON, Canada
- 4Department of Physiology, University of Toronto, Toronto, ON, Canada
- 5KITE, Toronto Rehab, University Health Network, Toronto, ON, Canada
- 6Rehabilitation Sciences Institute, University of Toronto, Toronto, ON, Canada
- 7Edward S. Rogers Sr. Department of Electrical and Computer Engineering, University of Toronto, Toronto, ON, Canada
- 8Department of Psychiatry, University of Toronto, Toronto, ON, Canada
- 9Centre for Mental Health, University Health Network, Toronto, ON, Canada
- 10Krembil Research Institute, University Health Network, Toronto, ON, Canada
Non-invasive brain stimulation (NIBS) is emerging as a robust treatment alternative for major depressive disorder, with a potential for achieving higher remission rates by providing targeted stimulation to underlying brain networks, such as the salience network (SN). Growing evidence suggests that these therapeutic effects are dependent on the frequency and phase synchrony between SN oscillations and stimulation as well as the task-specific state of the SN during stimulation. However, the development of phase-synchronized non-invasive stimulation has proved challenging until recently. Here, we use a phase-locked pulsed brain stimulation approach to study the effects of two NIBS methods: transcranial alternating current stimulation (tACS) versus phase-locked transcranial pulsed current stimulation (tPCS), on the SN during an SN activating task. 20 healthy volunteers participated in the study. Each volunteer partook in four sessions, receiving one stimulation type at random (theta-tACS, peak tPCS, trough tPCS or sham) while undergoing a learning game, followed by an unstimulated test based on learned material. Each session lasted approximately 1.5 h, with an interval of at least 2 days to allow for washout and to avoid cross-over effects. Our results showed no statistically significant effect of stimulation on the event related potential (ERP) recordings, resting electroencephalogram (EEG), and the performance of the volunteers. While stimulation effects were not apparent in this study, the nominal performance of the phase-locking algorithm offers a technical foundation for further research in determining effective stimulation paradigms and conditions. Specifically, future work should investigate stronger stimulation and true task-specific stimulation of SN nodes responsible for the task as well as their recording. If refined, NIBS could offer an effective, homebased treatment option.
Introduction
Non-invasive brain stimulation (NIBS) is an emerging alternative when conventional treatment approaches for Major Depressive Disorder (MDD) fail (Fregni and Pascual-Leone, 2007; Daskalakis et al., 2008; Ferrucci et al., 2009; Dayan et al., 2013). It is estimated that one in three MDD patients suffers from Treatment Resistant Depression (TRD), with failure to respond to at least two courses of antidepressant treatment (Nemeroff, 2007). NIBS technologies such as Food and Drug Administration (FDA)-approved repetitive transcranial magnetic stimulation (rTMS) and transcranial direct current stimulation (tDCS) have shown efficacy in helping with TRD in a large number of clinical trials, achieving 15–32% (O’Reardon et al., 2007; Berlim et al., 2014; Gaynes et al., 2014) and 7–43% (Berlim et al., 2013; Shiozawa et al., 2014) remission rates, respectively. There is a potential for achieving higher remission rates by providing a more targeted stimulation to the brain networks involved in MDD. Growing evidence (reviewed below) suggests that this can be achieved by first applying the stimulation at the same oscillatory profile as the underlying brain oscillation, and second by applying the stimulation during the time when the brain network of interest is in the most suitable state to receive the stimulation.
In contrast to tDCS, transcranial alternating current stimulation (tACS) and transcranial pulsed current stimulation (tPCS) use oscillatory waveforms with sinusoidal and square pulses that better match the underlying natural physiological brain activity. In a recent study, it has been shown that tACS can have an effect in spike timing of single neurons elucidating the mechanism of action for these types of NIBS and opening the opportunity for future research into the effects of timing of the stimulation waveform with respect to the underlying brain activity (Krause et al., 2019).
Multiple studies have shown the importance of achieving synchrony between the target brain oscillation and the stimulation frequencies and phase in order to increase the effect of the stimulation. A recent study has shown that adjusting rTMS pulse frequency to individual gamma oscillation resulted in a significant mood elevation compared to unadjusted rTMS stimulation at slightly higher or lower frequencies than their individual gamma oscillation (Chung et al., 2018). In another example, TMS phase-specific modulation of motor evoked potentials has been shown by applying pulses at the peak or trough of the μ-rhythm of the motor cortex (Zrenner et al., 2017). Similarly, the brain stimulation modalities that use energies below the threshold for induction of an action potential (e.g., transcranial electrical stimulation), are more effective when delivered at similar oscillation frequencies as the underlying brain target (Fröhlich and McCormick, 2010; Reato et al., 2013). For example, tACS has been shown effective to modulate alpha power only when delivered at alpha frequency (Zaehle et al., 2010; Vossen et al., 2015). Further, tACS delivered at a specific phase with respect to the underlying brain oscillation modulated the intensity of the tremor when applied to the motor cortex (Brittain et al., 2013) and changed the hearing thresholds when applied to the auditory cortex (Riecke et al., 2015; Wilsch et al., 2018).
The effects of brain stimulation have also consistently been shown to depend on the brain state at the time of stimulation (Silvanto et al., 2008). Current models of brain function posit that brain regions operate as integrated networks bound by coherent activity, and task-specific activation of these networks is seen across various brain states (Seager et al., 2002; Park and Friston, 2013; Pessoa, 2014). The state of the brain during the stimulation can change the outcome of the intervention; an elementary example would be the observation that the active motor threshold is substantially lower than the resting motor threshold for stimulation of the primary motor cortex (Hallett, 2007).
A useful stimulation target is the Salience Network (SN), which is activated when there is a transition between a cognitive task and sensory information (Downar et al., 2000, 2001). SN dysfunction is associated with a wide range of neuropsychiatric disorders, including MDD, post-traumatic stress disorder (PTSD), and schizophrenia (Peters et al., 2016). Targeting the SN has been proven successful when using rTMS for treating MDD (Peters et al., 2016). Considering the importance of brain state during brain stimulation, applying the stimulation during an SN-activating task may potentially improve the effect of the stimulation.
Providing a more targeted brain stimulation may help discover more effective treatments for MDD patients. A closed-loop system enabling phase-locked stimulation could potentially allow more precise control of the stimulation frequency and phase. Such system could achieve a more consistent treatment effect overall, given the findings pointing at the importance of brain state during the stimulation and the synchrony of the stimulation with the underlying brain oscillation. Here, we conducted an experiment to study the effect of two NIBS methods: tACS and transcranial phase-locked pulsed current stimulation (tPCS), on the SN during an SN activating task. The effects of these types of stimulation have previously been shown to be: spectral power density changes in specific frequency bands during rest EEG (Zaehle et al., 2010; Helfrich et al., 2014; Vossen et al., 2015), changes in task-specific activation of the brain (Meinzer et al., 2012; Jaušovec and Jaušovec, 2014; Cabral-Calderin et al., 2016), and behavioral or task-specific performance changes (Santarnecchi et al., 2013; Voss et al., 2014). We, therefore, hypothesized that by providing stimulation in synchrony with the underlying brain activity during an SN activating task, we would achieve more effective stimulation of the SN, as reflected by increases in these previously established electrophysiological and behavioral effects. We further hypothesized that tACS would strengthen the SN activity resulting in faster reaction time and better performance during the task, while producing an increase in resting theta power and ERP.
In this work we attempted to engage the SN by using tACS at theta frequency or phase-locked tPCS synchronized to the frontal theta when the volunteers were engaged in a SN activating task. Successful implementation of tACS in a closed-loop system with electroencephalography (EEG) recording is currently an unsolved problem, because the artifact generated by the stimulation obscures the recording (Neuling et al., 2017; Noury and Siegel, 2017). To study the phase- and frequency-locked brain stimulation method, we previously have developed a phase-locked tPCS brain stimulation technique that can extract phase and frequency of the EEG signal and deliver the stimulation pulses at specific phase and frequency of the EEG signal in real-time (Mansouri et al., 2018). Here we applied this technique to phase-lock tPCS to the activity of the SN recorded through theta EEG. The electrical pulses were delivered either at peak or trough of the recording, with the expected result of generating opposite effects – as was previously shown when using rTMS over motor cortex (Zrenner et al., 2017), electrical stimulation of hippocampal brain slices (Hyman et al., 2003), and electrical stimulation of the somatosensory cortex in monkeys (Zanos et al., 2018). We hypothesized that trough tPCS would strengthen the SN activity, while peak tPCS would have the opposite effect, and that these effects would be exhibited through an increase or decrease of theta power and ERP recording, alongside changes in behavioral performance during the task.
Materials and Methods
Participants and Visits
Twenty healthy adult (older than 18) volunteers free of neurological or psychiatric illnesses participated in the study. All participants gave written informed consent, and the study was approved by the Research Ethics Board of the University Health Network. For each of the four visits, volunteers received one of the stimulation types at random (theta-tACS, peak tPCS, trough tPCS or sham). The order of the visits was assigned at random using a random generator with the participants not informed of the order of the visits; only the experimenter knew the type of stimulation at each visit. The visits were scheduled at least 2 days apart to allow for washout and to avoid cross-over effects. Each session took approximately 1 h and a half.
Study Visit and Computer Task
During each visit, volunteers sat in front of a computer and performed a decision-making task described by Frank et al. (2004, 2005). Each visit consisted of a learning and a test period (Figure 1). During the learning period, the volunteers were presented with three pairs of randomly assigned Japanese characters (AB, CD, and EF) that were not easily verbalized. In each pair, one of the characters was more likely to win a reward (A:80%-B:20%, C:70%-D:30% and E:60%-F:40%). For each trial, the volunteers were presented with one pair (each character was displayed on an image of a jar) and within 1 sec had to select a character by pressing the left or the right arrow keys on a keyboard. Next, the volunteers were presented with a feedback to tell them whether they “won”, “lost”, or were too late to respond. This game continued for 1 h, with 1 min breaks every 4 min.
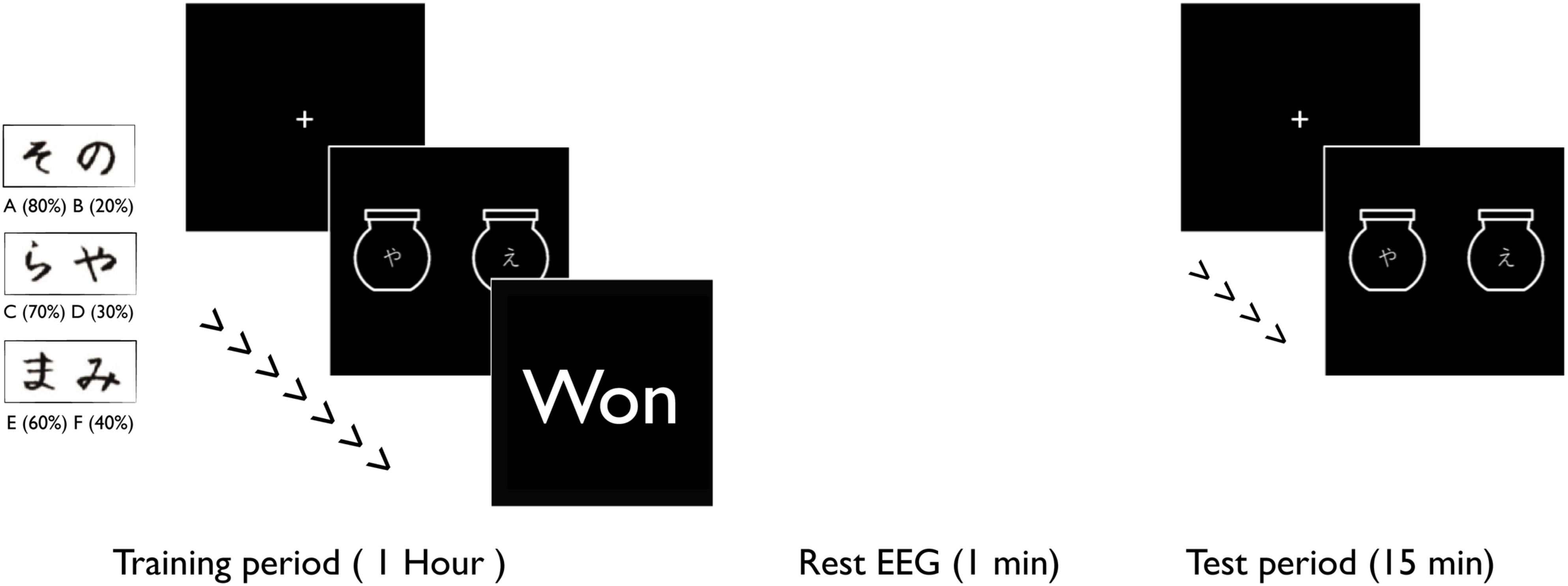
Figure 1. Volunteers were presented with six pairs of Japanese characters. In each pair, one character had a higher probability of “winning”. During the training period, the volunteers were presented with these pairs to learn their probabilities through reinforcement learning. The training task was continued for 1 h. The volunteers completed 1 min of rest EEG recording after the training period. During the test period volunteers, were presented with two Japanese characters but not necessarily paired in the same way, to create conflicting scenarios (win/win or lose/lose).
After completing the learning game, volunteers took a short test with same pairs as the learning game (AB, CD, and EF) and also pairs that they had not previously seen (AC, AE, CE, BD, BE, and DE). During the test, no feedback was presented, to avoid further learning effects.
The same task was performed during each of the four sessions. For each session, six new randomly selected Japanese characters were used. At each session, during the learning task, the volunteers received one of the 4 stimulation types. No stimulation was delivered during the test task.
Phase-Locked Transcranial Pulsed Current Stimulation
A similar method to Mansouri et al. (2018) was used to provide square wave 5 ms pulses of 2 mA amplitude at either peak (90 degree phase) or trough (270 degree phase) of the theta oscillation (4–8 Hz) recorded from the midfrontal part of the scalp. First, the stimulation electrodes (2 cm × 2 cm) were placed on the scalp at F3 (anode) and F4 (cathode); Ten20 conductive gel (Weaver and Company, Aurora, CO., United States) was applied to reduce the impedances of the electrodes to below 5 kΩ. Next, a 16-channel passive-electrode EEG cap (EasyCap GmbH, Germany) was worn by the volunteers on top of the stimulation electrodes and HiCL Abrasive EEG Gel (EasyCap GmbH, Germany) was applied to Fz (recording electrode), Pz (Reference electrode) and right tragus (Ground) to reduce the impedance of each of the recording electrodes to below 5 kΩ (Figure 2).
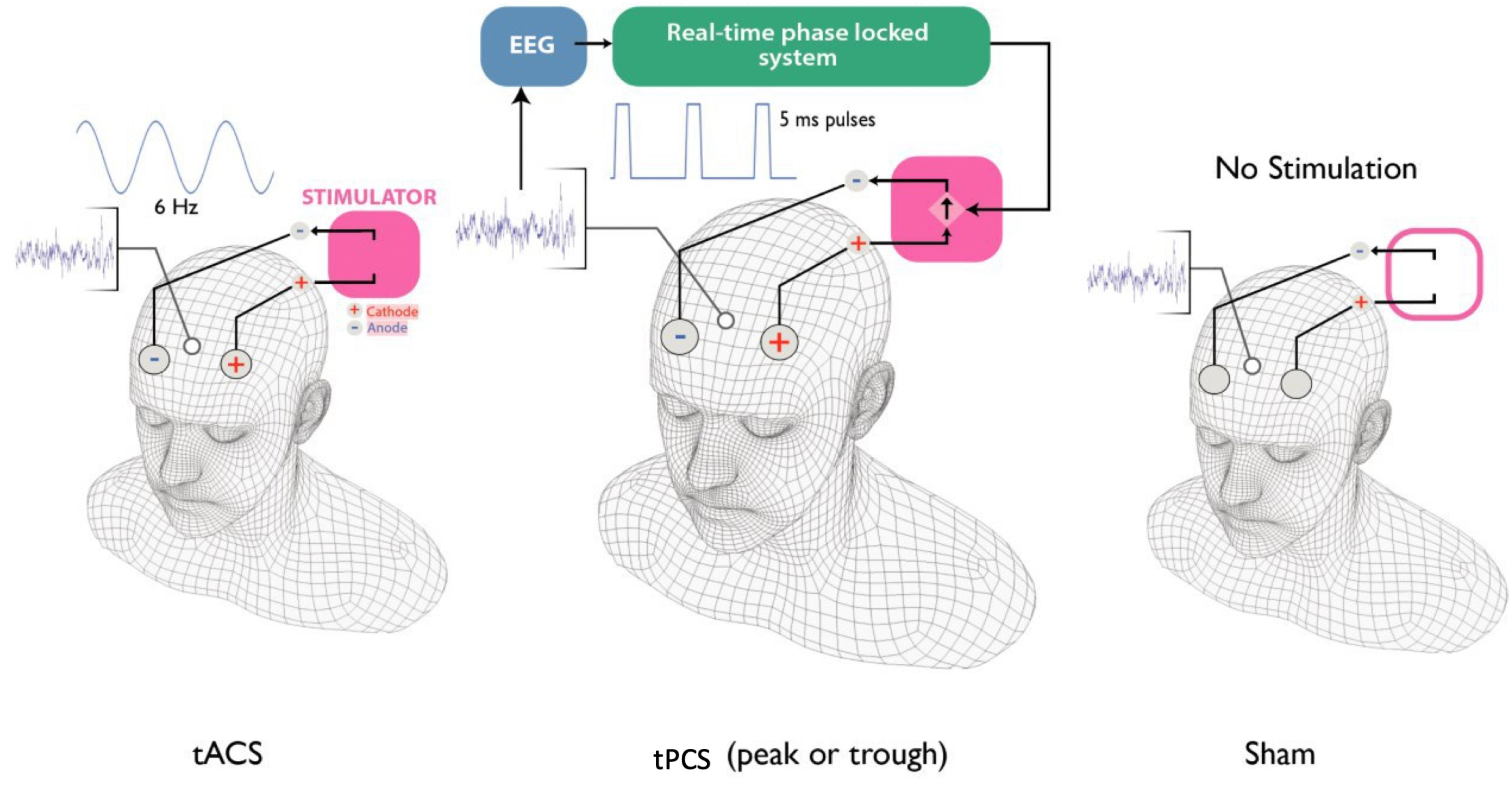
Figure 2. Four different stimulation types were used in this study: tACS, tPCS peak, tPCS trough and sham. During tACS stimulation, a 6 Hz sinusoidal stimulation was applied over F3 and F4. During the tPCS at peak or trough, 5 ms pulses were delivered at peak or trough of the theta oscillation recorded from Fz. During the sham session, no stimulation was delivered.
As previously described in Mansouri et al. (2017), the recorded EEG from Fz using V-AMP16 (Brain Products GmbH, Munich, Germany) was analyzed in MATLAB (MATLAB version 7.9.0) using a Fast Fourier Transform-based forecasting method (Mansouri et al., 2017) to provide the timing of the pulses and this timing was communicated to the ANT Neuro stimulator through an Arduino interface. The stimulation was applied for 1 h during the learning task; no stimulation was delivered during the test task.
Transcranial Alternating Current Stimulation (tACS) Procedure
First, the stimulation electrodes (2 cm × 2 cm) were placed on the scalp at F3 (anode) and F4 (cathode); Ten20 conductive gel (Weaver and Company, Aurora, CO., United States) was applied to reduce the impedances of the electrodes to below 5 kΩ. Then, the stimulation was set to provide a 6 Hz sinusoidal current stimulation with amplitude of 2 mA peak to peak and zero direct current. The stimulation was initiated with a 30-s ramp up from zero to 2 mA amplitude. As in the tpES case, the stimulation was applied for 1 h during the learning task and no stimulation was delivered during the test task.
Sham Stimulation Procedure
The stimulation electrodes were placed in a similar way to the pulsed stimulation and tACS stimulation. An initial tACS for 1 min with 30 s ramp up to 2 mA and 30 s ramp down stimulation was applied to help with the blinding of the participants. No further stimulation was applied during this visit. The participants completed both the learning and the test task.
Resting-State EEG and ERP Recording
The same EEG setup was used for rest EEG and ERP recordings. Pz was used as the reference and right tragus as the ground electrodes. Rest EEG was recorded before the test task for 1 min as the volunteers sat in front of the computer and the recording continued to capture the event-related potential (ERP) during the test task. Trigger signals were provided to the amplifier to capture the events during the game.
Resting-State EEG and ERP Analysis
All the analysis was done in MATLAB (MATLAB R2016b) and EEGLAB EEG analysis toolbox (Delorme and Makeig, 2004) was used for specific analysis. First, the channels were manually inspected and bad channels (large noise or poor connection) were identified and removed from the analysis. An average reference transformation was applied to the data to minimize the effect of reference site. Next a zero-phase 1 Hz high-pass FIR filter was used to remove the baseline drift. We applied a threshold to identify and remove large movement and eyeblink artifacts. Further, a zero-phase shift 1–50 Hz FIR bandpass filter was applied to the data. Response-locked ERP measures were extracted and a zero-phase shift 1–14 Hz bandpass IIR filter was applied to avoid bifurcation (Frank et al., 2005). Welch’s power spectral density estimate with a Hanning window was used to generate the spectral density of the rest EEG.
Analysis
Statistical Methods and Analysis
Considering the design of the study, a repeated-measure method to investigate the variability within the factors (subjects) is suitable for testing the effect of the stimulation. Previous publications, that employed the same task with a larger group of participants, used a parametric test for their statistical evaluations (Frank et al., 2004, 2005). We performed a Shapiro–Wilk test, which refuted that our data is from a normally distributed population. Thus, instead of the parametric repeated-measure analysis of variance (ANOVA), the non-parametric repeated-measure Friedman’s test was used to assess the effect of the stimulation type over the outcome measures. Significance p-value level was set at 0.05 for rejecting the null hypothesis (no effect of stimulation). Considering the 15 statistical tests performed on the data, the significance p-value level with multiple comparison Bonferroni correction was adjusted to 0.003.
Phase Locking Value (PLV) was used to evaluate the performance of the phase-locked tPCS. PLV is a value between 0 and 1; higher PLV shows better phase locking.
Sample Size Justification
A power analysis for repeated-measure ANOVA test within factors was conducted using G∗Power (Faul et al., 2007) assuming an intermediate effect size (f = 0.5) based on previously published tDCS studies (Minarik et al., 2016; Aleman et al., 2018), 95% power and alpha error probability of 5%. This analysis suggested the total sample size of at least 20 participants. We assumed there were no carryover effects between the four sessions.
Results
Recruitment
Twenty volunteers (10/10 male/female; mean age 31.7 ± SD 8.6 years, range 23–53) were recruited to complete four visits. There were no complications after the stimulation sessions; minor tingling was reported during tACS stimulation. Pulsed stimulation was felt only during the beginning of the session as a tapping sensation on the head; the volunteers gradually acclimatized to these effects and reported no sensation of the stimulation afterward.
Closed-Loop Brain Stimulation
Both peak and trough tPCS were applied successfully in terms of phase-locking performance. A minimum of 0.31 PLV and maximum of 0.70 PLV and average of 0.50 ± 0.13 was achieved for peak stimulation. A minimum of 0.32 PLV and maximum of 0.80 PLV and average of 0.53 ± 0.14 was achieved for trough stimulation. The average error in the peak stimulation was 2.5o ± 16o and for trough was 5.2o ± 17o.
Task-Specific Findings
All participants learned the probabilistic reward associations of the task successfully and were able to score more than 65% on AB cases, 60% on CD cases and 50% on EF cases during the learning task in all four visits. However, when comparing learning performance across the four stimulation conditions, no significant effect was apparent: there was no effect of stimulation type on decision of the players to choose A over B (Friedman’s test χ2F(3) = 1.49, p = 0.68), C over D (Friedman’s test χ2F (3) = 4.02, p = 0.26), and E over F (Friedman’s test χ2F(3) = 5.84, p = 0.12).
Next, we examined at the probability of selecting A over all the other characters and probability of avoiding B over all the characters. “Positive learners”, as described by Frank et al., 2004 have stronger tendency to select A, while “negative learners” have stronger tendency to avoid B (Frank et al., 2004). We did not find any statistical effect of stimulation on probability of choosing A (Friedman’s test χ2F(3) = 1.76, p = 0.62), or avoiding B (Friedman’s test χ2F(3) = 1.49, p = 0.68). Also the ratio of probability of choosing A over avoiding B was not affected by the type of stimulation (Friedman’s test χ2F(3) = 0.85, p = 0.84).
In addition, we evaluated the reaction times in no conflict cases where there was winning character paired with a losing character (i.e., AB), lose/lose conflict cases where there was a losing character paired with another losing character (i.e., BD) and win/win conflict cases where a winning character was paired with another winning character (i.e., AC). There was no significant effect of stimulation type on reaction time for no conflict cases (Friedman’s test χ2F(3) = 3.03, p = 0.39), lose/lose conflict cases (Friedman’s test χ2F(3) = 0.52, p = 0.92), or win/win conflict cases (Friedman’s test χ2F(3) = 3.43, p = 0.33) (Figure 3).
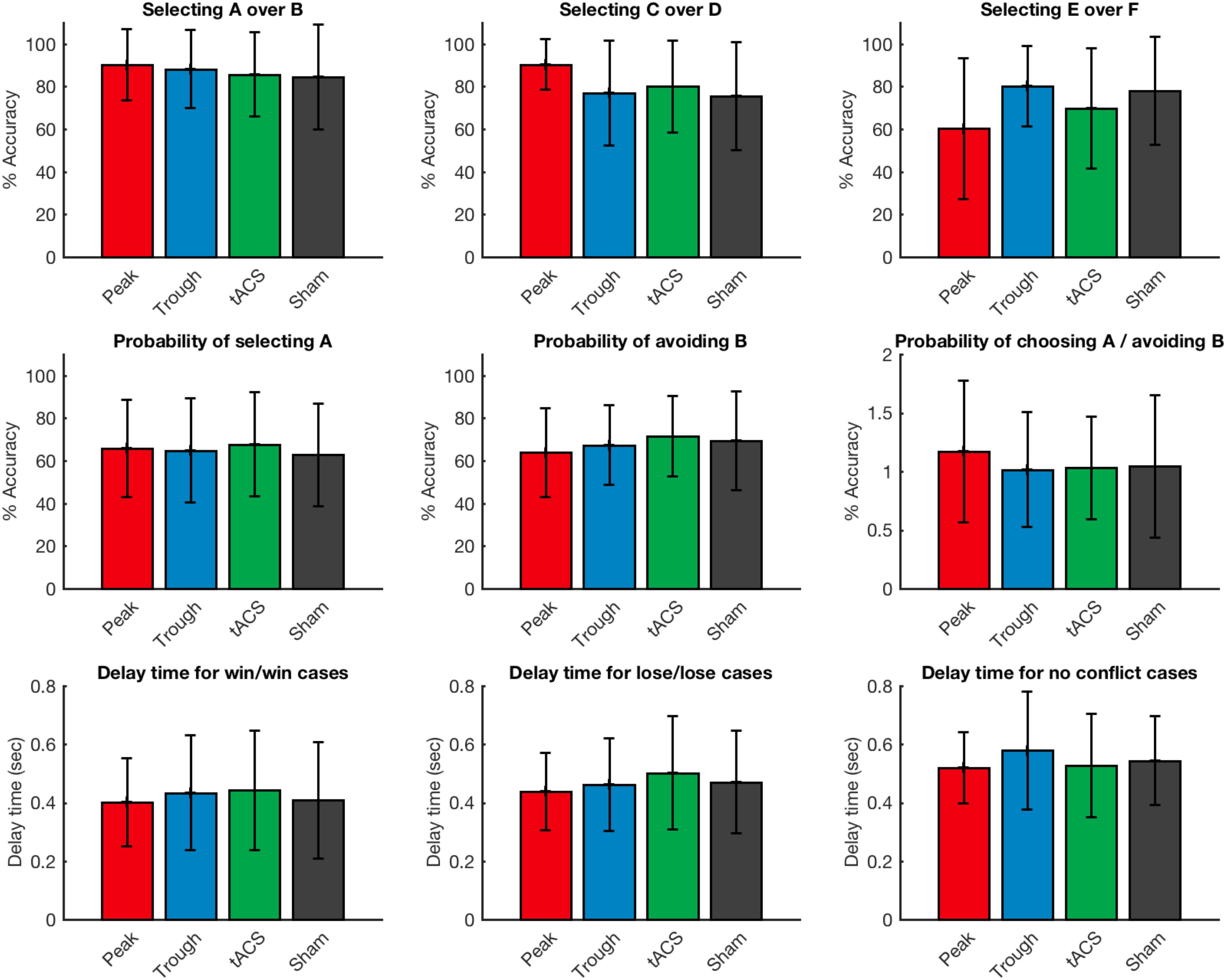
Figure 3. Accuracy and response timing of the volunteers during the test period of the game during the various possible choice scenarios on the task. Error bars represent standard deviation.
ERP Recording
Next, similarly to Frank et al., 2004, we investigated the event-related potentials when time-locked to response (Frank et al., 2004). The analysis showed similar ERPs as previously reported (Frank et al., 2004, 2005). There was no effect of stimulation in negativity amplitude in win/win ERP voltages (Friedman’s test χ2F(3) = 1.03, p = 0.79) and negativity amplitude in lose/lose ERP voltages (Friedman’s test χ2F(3) = 0.34, p = 0.95) (Figure 4).
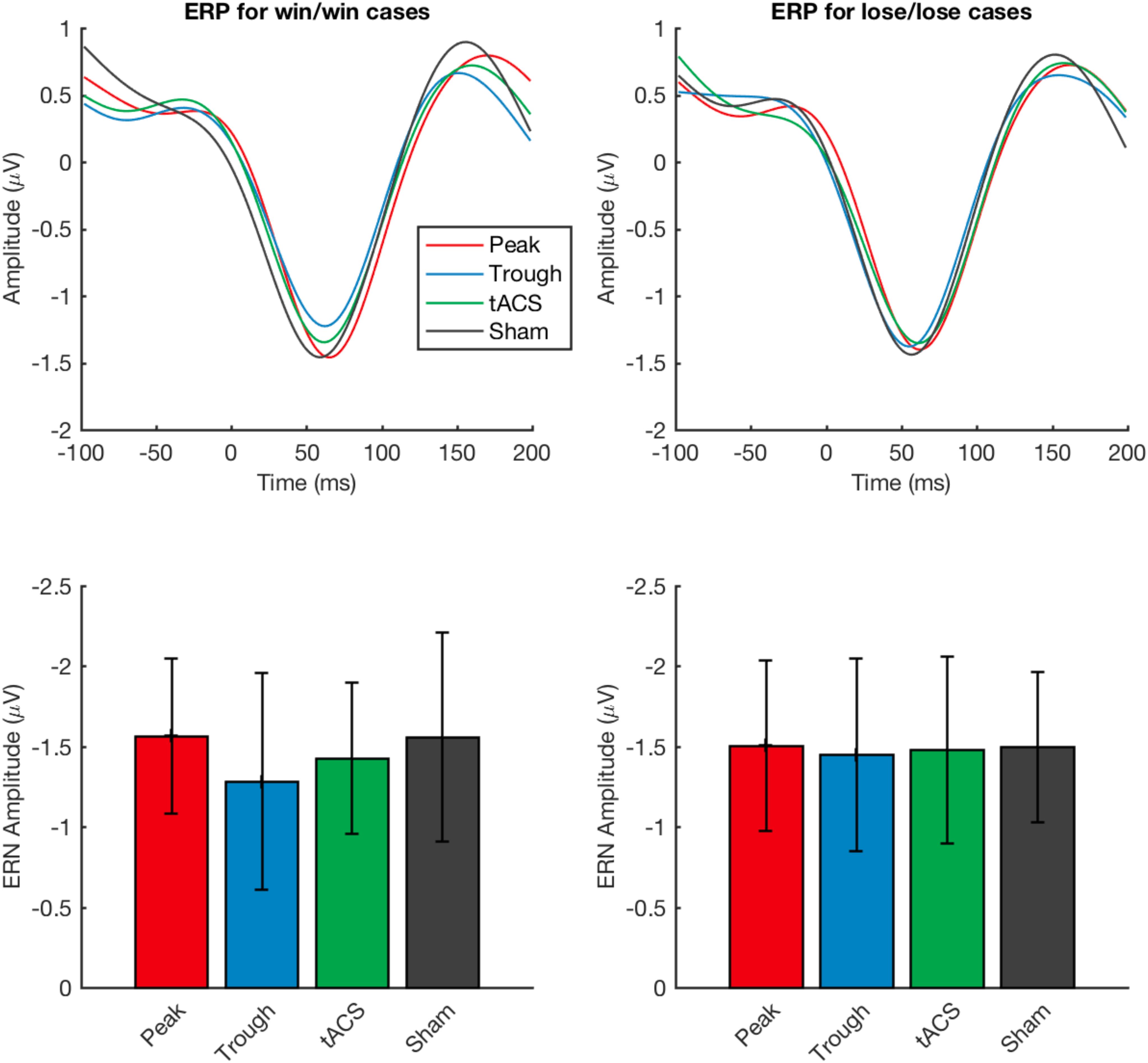
Figure 4. Event-related potential recordings obtained during the test task under the four types of stimulation applied. Waveforms for event-related positive (ERP) and amplitudes for event-related negative (ERN) potentials are shown for tACS applied under the sham, active non-phase-locked, active peak-phase-locked, and active trough phase-locked stimulation conditions are compared. No significant differences were detected across any of the four stimulation types. Error bars represent standard deviation.
EEG Spectral Power
Power spectral analysis of the resting EEG signal also did not show any difference among the 4 stimulation types. There was no statistical significance when measuring the power in delta (1–4 Hz) (Friedman’s test χ2F(3) = 1.12, p = 0.77), theta (4–8 Hz) (Friedman’s test χ2F(3) = 4.54, p = 0.21), alpha (8–13 Hz) (Friedman’s test χ2F(3) = 1.78, p = 0.62), or beta (13–30 Hz) (Friedman’s test χ2F(3) = 1.71, p = 0.64) EEG frequency bands (Figure 5).
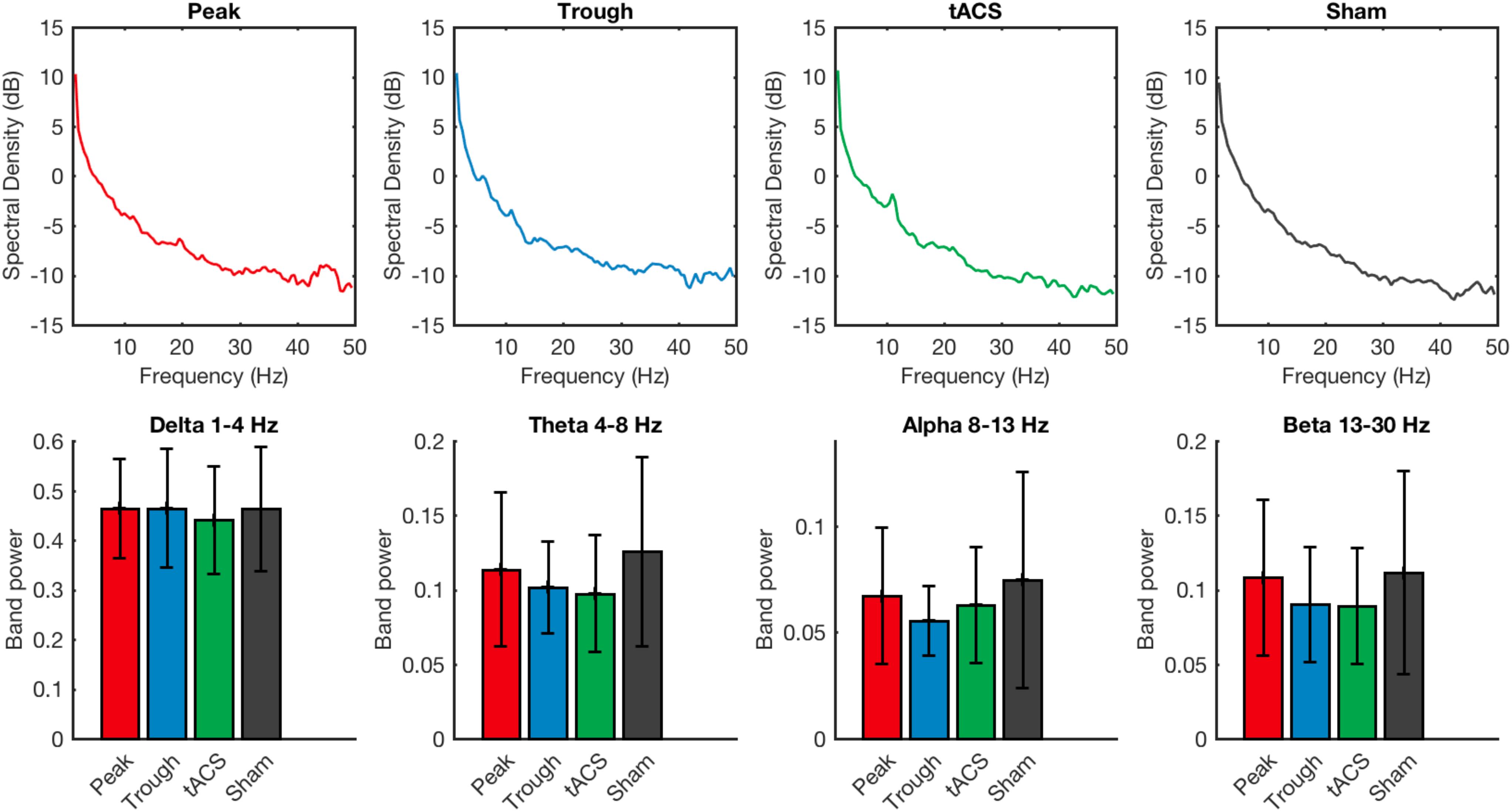
Figure 5. Power spectral densities of the resting EEG recorded after the training session are compared for tACS applied under the sham, active non-phase-locked, active peak-phase-locked, and active trough phase-locked stimulation conditions. No significant differences were detected across any of the four stimulation types. Error bars represent standard deviation.
Discussion
In this work we have attempted to modulate SN activity differentially, by applying theta tACS or phase-locked tPCS synchronized to the frontal theta when the volunteers were engaged in a SN activating task. However, the results of the present study did not demonstrate differential effects of any of the 4 stimulation types on either electrophysiological or behavioral measures. Indeed, no significant effects were observed for any of the active stimulation conditions compared to sham stimulation in this investigation.
Previously, theta tACS applied during rest has been shown to be an effective modulator of frontal theta power (Pahor and Jaušovec, 2014); however, its effects have not been studied when the stimulation is applied during a task. In another study, theta tACS applied over parietal brain regions before a working memory task resulted in increased working memory storage; however, the same stimulation over frontal region had no effect (Jaušovec and Jaušovec, 2014). Cognitive effects of theta tPCS have been shown to be very small and specific to complex mathematical tasks (Morales-Quezada et al., 2015). In a more recent study, theta tACS applied over frontal region showed a decrease in nodal efficiency of dorsal anterior cingulate cortex (dACC) – one of the nodes of SN (Onoda et al., 2017).
The lack of any effect for any of the active stimulation conditions in the present study raises the question of whether the stimulation parameters were adequate to achieve neurophysiological effects in general, notwithstanding the role of phase-locking. Recent publications suggest that stronger transcranial electrical stimulation currents (i.e., >2 mA) may be required in some circumstances to have detectable effects on brain activity and behavior (Vöröslakos et al., 2018). In our study we respected the stimulation limits imposed by the hardware and what has been conventionally used and considered safe for these types of stimulation, which is 2 mA of current. However, it has been shown that in order to produce effective fields in the brain, currents as high as 6 mA are sometimes needed (Vöröslakos et al., 2018). Considering the thickness of the skull and the layers of dura protecting the brain, it is reasonable that stimulation may require higher currents to achieve the desired effect. Previously, other studies have shown that stimulation intensities as low as 2 mA can achieve sufficiently strong fields in the brain through computer simulations (Wagner et al., 2007), and studies using tDCS and tACS have been proven effective (Zaghi et al., 2010; Stagg and Nitsche, 2011; Reato et al., 2013; Kuo et al., 2014); however, these experiments have used different electrode montages and targeted other brain networks rather than SN itself. Moreover, brain stimulation in these studies was generally applied during rest, while in our study the stimulation was applied during a task. Further, some studies could only detect improvement in MDD patients after multiple sessions of tDCS (Brunoni et al., 2011; Alonzo et al., 2012), multiple sessions of stimulations should be considered for future trials. Finally, and importantly, most previous work regarding effective current amplitude applied either ongoing transcranial direct current stimulation (tDCS) or else sinusoidal tACS. In contrast, the present study’s phase-locked stimulation applied short pulses of 5 ms rather than sinusoidal pulses – a feature that may have precluded measurable effects on brain activity due to the short duty cycle of stimulation.
Considering the small sample size and the variability in the outcome measure, we speculate that the methods we used here (specifically, the 2 mA amplitude and brief-pulse waveform of phase-locked stimulation) may have had smaller effects than anticipated, and that the natural variability between the experimental sessions could have been larger than the effects of stimulation. In future work, using a larger sample size, increasing the stimulation intensity, or applying a longer duty cycle for the phase-locked tPCS could possibly lead to unveiling the effect of these types of NIBS.
As an additional factor to consider, the stimulation techniques used in this work may not have been suitable to modulate SN activity due to their low power and low spatial focality. Even though there is evidence of effectiveness of tACS in other models, modulation of SN has been mainly shown using rTMS, which is a much stronger type of brain stimulation in terms of both field intensity and depth. Body tissues have nearly uniform magnetic permeability and do not significantly distort the magnetic field produced by rTMS, allowing effective focusing of the stimulation to a restricted target at higher intensity than tES (Wagner et al., 2004). On the other hand, for non-invasive electrical stimulation, the large differences in electrical conductivity of the various tissues in the overlying scalp and skull not only blocks most of the current from reaching brain tissue, but also disperses the fields, thereby precluding effective focusing of the stimulation. Some work has been done to use multiple electrodes in a high density (HD) fashion to improve the focality of the stimulation (Kuo et al., 2013), which potentially could improve the effectiveness of the stimulation in future work.
It is believed that the electrical stimuli of tES affect the ongoing activity of the brain; thus, task-specific activation of the SN was used as means to assess the effect of the stimulation. However, the task used in our paper has not yet been used in the context of brain stimulation, and thus, perhaps other tasks should be studied with a similar stimulation protocol. While we know that the presently employed task activates SN and produces increased theta oscillation – a hallmark of SN activity – it is possible that in this case the task did not activate the specific nodes of the SN where the stimulation was delivered, or that the activation was not reflected in the outcome measures we studied in this experiment. We have selected these specific outcome measures based on the previous findings (resting EEG power changes, ERPs specific to the task and task specific behavioral changes). The effects of these types of stimulation has previously been shown to result in spectral power density changes in specific frequency bands during rest EEG (Zaehle et al., 2010; Helfrich et al., 2014; Vossen et al., 2015), changes in task specific activation of the brain (Meinzer et al., 2012; Jaušovec and Jaušovec, 2014; Cabral-Calderin et al., 2016), and behavioral or task specific performance changes (Santarnecchi et al., 2013; Voss et al., 2014). It therefore would be valuable to test our methods, tACS and phase-locked tPCS, in a sensorimotor model so we can check for changes in motor or sensory evoked potentials as a result of the stimulation.
Translationally, the search for novel NIBS approaches is much needed, as the current treatment options for TRD are very limited. Electroconvulsive therapy (ECT) has conventionally been the treatment of choice for TRD; however, it is associated with high costs, significant stigma, and cognitive side effects that deter many patients (Gangadhar, 2005). rTMS is a potential alternative; however, its limited availability, higher costs, and requirement for in-clinic rather than at-home application are the main limitations restricting its widespread adoption. A cost-effective, home-based brain stimulation technique may potentially resolve those limitations and provide an alternative treatment option for TRD patients. tPCS/tACS can potentially be made available as a home-based treatment, and the application of the stimulating currents in an oscillatory, phase-locked fashion could provide an opportunity to improve its efficacy by attempting to engage specific brain networks. Primarily, the past few decades of neuropsychiatric and neuroimaging research points toward the SN as the main target for treating MDD and TRD. However, tDCS and other simple electrical stimulation protocols can become more effective when integrated in a closed-loop system (Zrenner et al., 2016). Future work in this direction may potentially help develop a closed-loop electrical based brain stimulation technique.
To summarize, in this study we have tested novel approaches, tACS and phase-locked tPCS, to stimulate the SN network with hopes that such stimulation could be used for therapeutic purposes. If found effective, these techniques could potentially open doors for new treatments of MDD and other neuropsychiatric disorders linked to the SN. Although our results showed no statistically significant effect of stimulation on the ERP recordings, resting EEG recordings, and the performance of the volunteers in a SN activating task, future work will reveal whether such effects could be obtained by either modifying the amplitude or waveform of electrical stimulation, or else applying the same phase-locking algorithm to a more powerful stimulation modality, such as rTMS. Success could lead to more successful outcomes for patients undergoing treatment for TRD and other medically refractory neuropsychiatric illnesses.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
Twenty healthy volunteers free of neurological or psychiatric illnesses participated in the study. All participants gave informed consent, and the study was approved by the Research Ethics Board of the University Health Network.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by the MITACS Accelerate Grant (#IT05799) FM.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aleman, A., Enriquez-Geppert, S., Knegtering, H., and Dlabac-de Lange, J. J. (2018). Moderate effects of noninvasive brain stimulation of the frontal cortex for improving negative symptoms in schizophrenia: meta-analysis of controlled trials. Neurosci. Biobehav. Rev. 89, 111–118. doi: 10.1016/j.neubiorev.2018.02.009
Alonzo, A., Brassil, J., Taylor, J. L., Martin, D., and Loo, C. K. (2012). Daily transcranial direct current stimulation (tDCS) leads to greater increases in cortical excitability than second daily transcranial direct current stimulation. Brain Stimul. 5, 208–213. doi: 10.1016/j.brs.2011.04.006
Berlim, M. T., Van den Eynde, F., and Daskalakis, Z. J. (2013). Clinical utility of transcranial direct current stimulation (tDCS) for treating major depression: a systematic review and meta-analysis of randomized, double-blind and sham-controlled trials. J. Psychiatr. Res. 47, 1–7. doi: 10.1016/j.jpsychires.2012.09.025
Berlim, M. T., Van Den Eynde, F., Tovar-Perdomo, S., and Daskalakis, Z. J. (2014). Response, remission and drop-out rates following high-frequency repetitive transcranial magnetic stimulation (rTMS) for treating major depression: a systematic review and meta-analysis of randomized, double-blind and sham-controlled trials. Psychol. Med. 44, 225–239. doi: 10.1017/S0033291713000512
Brittain, J. S., Probert-Smith, P., Aziz, T. Z., and Brown, P. (2013). Tremor suppression by rhythmic transcranial current stimulation. Curr. Biol. 23, 436–440. doi: 10.1016/j.cub.2013.01.068
Brunoni, A. R., Ferrucci, R., Bortolomasi, M., Vergari, M., Tadini, L., Boggio, P. S., et al. (2011). Transcranial direct current stimulation (tDCS) in unipolar vs. bipolar depressive disorder. Prog. Neuro- Psychopharmacology. Biol. Psychiatry 35, 96–101. doi: 10.1016/j.pnpbp.2010.09.010
Cabral-Calderin, Y., Anne Weinrich, C., Schmidt-Samoa, C., Poland, E., Dechent, P., Bähr, M., et al. (2016). Transcranial alternating current stimulation affects the BOLD signal in a frequency and task-dependent manner. Hum. Brain Mapp. 37, 94–121. doi: 10.1002/hbm.23016
Chung, S. W., Sullivan, C. M., Rogasch, N. C., Hoy, K. E., Bailey, N. W., Cash, R. F. H., et al. (2018). The effects of individualised intermittent theta burst stimulation in the prefrontal cortex: a TMS-EEG study. Hum. Brain Mapp. 40, 608–627. doi: 10.1002/hbm.24398
Daskalakis, Z. J., Levinson, A. J., and Fitzgerald, P. B. (2008). Repetitive transcranial magnetic stimulation for major depressive disorder: a review. Can. J. Psychiatry. 53, 555–566. doi: 10.1177/070674370805300902
Dayan, E., Censor, N., Buch, E. R., Sandrini, M., and Cohen, L. G. (2013). Noninvasive brain stimulation: from physiology to network dynamics and back. Nat. Neurosci. 16, 838–844. doi: 10.1038/nn.3422
Delorme, A., and Makeig, S. (2004). EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods. 134, 9–21. doi: 10.1016/j.jneumeth.2003.10.009
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2000). A multimodal cortical network for the detection of changes in the sensory environment. Nat. Neurosci. 3, 277–283. doi: 10.1038/72991
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2001). The effect of task relevance on the cortical response to changes in visual and auditory stimuli: an event-related fMRI study. Neuroimage 14, 1256–1267. doi: 10.1006/nimg.2001.0946
Faul, F., Erdfelder, E., Lang, A. G., and Buchner, A. (2007). G∗Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior. Research. Methods. 39, 175–191. doi: 10.3758/BF03193146
Ferrucci, R., Bortolomasi, M., Vergari, M., Tadini, L., Salvoro, B., Giacopuzzi, M., et al. (2009). Transcranial direct current stimulation in severe, drug-resistant major depression. J. Affect. Disord. 118, 215–219. doi: 10.1016/j.jad.2009.02.015
Frank, M. J., Seeberger, L. C., and O’Reilly, R. C. (2004). By carrot or by stick: cognitive reinforcement learning in parkinsonism. Science 306, 1940–1943. doi: 10.1126/science.1102941
Frank, M. J., Woroch, B. S., and Curran, T. (2005). Error-related negativity predicts reinforcement learning and conflict biases. Neuron 47, 495–501. doi: 10.1016/j.neuron.2005.06.020
Fregni, F., and Pascual-Leone, A. (2007). Technology insight: noninvasive brain stimulation in neurology-perspectives on the therapeutic potential of rTMS and tDCS. Nat. Clin. Pract. Neurol. 3, 383–393. doi: 10.1038/ncpneuro0530
Fröhlich, F., and McCormick, D. A. (2010). Endogenous electric fields may guide neocortical network activity. Neuron 67, 129–143. doi: 10.1016/j.neuron.2010.06.005
Gaynes, B. N., Lloyd, S. W., Lux, L., Gartlehner, G., Hansen, R. A., Brode, S., et al. (2014). Repetitive transcranial magnetic stimulation for treatment-resistant depression: a systematic review and meta-analysis. J. Clin. Psychiatry. 75, 477–489. doi: 10.4088/JCP.13r08815
Hallett, M. (2007). Transcranial magnetic stimulation: a primer. Neuron 55, 187–199. doi: 10.1016/j.neuron.2007.06.026
Helfrich, R. F., Schneider, T. R., Rach, S., Trautmann-Lengsfeld, S. A., Engel, A. K., and Herrmann, C. S. (2014). Entrainment of brain oscillations by transcranial alternating current stimulation. Curr. Biol. 24, 333–339. doi: 10.1016/j.cub.2013.12.041
Hyman, J. M., Wyble, B. P., Goyal, V., Rossi, C. A., and Hasselmo, M. E. (2003). Stimulation in hippocampal region CA1 in behaving rats yields long-term potentiation when delivered to the peak of theta and long-term depression when delivered to the trough. J. Neurosci. 23, 11725–11731. doi: 10.1523/jneurosci.23-37-11725.2003
Jaušovec, N., and Jaušovec, K. (2014). Increasing working memory capacity with theta transcranial alternating current stimulation (tACS). Biol. Psychol. 96, 42–47. doi: 10.1016/j.biopsycho.2013.11.006
Krause, M. R., Vieira, P. G., Csorba, B. A., Pilly, P. K., and Pack, C. C. (2019). Transcranial alternating current stimulation entrains single-neuron activity in the primate brain. Proc. Natl. Acad. Sci. U. S. A. 116, 5747–5755. doi: 10.1073/pnas.1815958116
Kuo, H., Bikson, M., Datta, A., Minhas, P., Paulus, W., Kuo, M. F., et al. (2013). Comparing cortical plasticity induced by conventional and high-definition 4 × 1 ring tDCS: a neurophysiological study. Brain Stimul. 6, 644–648. doi: 10.1016/j.brs.2012.09.010
Kuo, M.-F., Paulus, W., and Nitsche, M. A. (2014). Therapeutic effects of non-invasive brain stimulation with direct currents (tDCS) in neuropsychiatric diseases. Neuroimage 85, 948–960. doi: 10.1016/j.neuroimage.2013.05.117
Mansouri, F., Dunlop, K., Giacobbe, P., Downar, J., and Zariffa, J. (2017). A fast EEG forecasting algorithm for phase-locked transcranial electrical stimulation of the human brain. Front. Neurosci. 11:401. doi: 10.3389/fnins.2017.00401
Mansouri, F., Fettes, P., Schulze, L., Giacobbe, P., Zariffa, J., and Downar, J. (2018). A real-time phase-locking system for non-invasive brain stimulation. Front. Neurosci. 12:877. doi: 10.3389/fnins.2018.00877
Meinzer, M., Antonenko, D., Lindenberg, R., Hetzer, S., Ulm, L., Avirame, K., et al. (2012). Electrical brain stimulation improves cognitive performance by modulating functional connectivity and task-specific activation. J. Neurosci. 32, 1859–1866. doi: 10.1523/JNEUROSCI.4812-11.2012
Minarik, T., Berger, B., Althaus, L., Bader, V., Biebl, B., Brotzeller, F., et al. (2016). The importance of sample size for reproducibility of tDCS effects. Front. Hum. Neurosci. 10:453. doi: 10.3389/fnhum.2016.00453
Morales-Quezada, L., Cosmo, C., Carvalho, S., Leite, J., Castillo-Saavedra, L., Rozisky, J. R., et al. (2015). Cognitive effects and autonomic responses to transcranial pulsed current stimulation. Exp. Brain Res. 233, 701–709. doi: 10.1007/s00221-014-4147-y
Nemeroff, C. B. (2007). Prevalence and management of treatment-resistant depression. J. Clin. Psychiatry 68(Suppl. 8), 17–25. doi: 10.1186/1471-2288-11-29
Neuling, T., Ruhnau, P., Weisz, N., Herrmann, C. S., and Demarchi, G. (2017). Faith and oscillations recovered: on analyzing EEG/MEG signals during tACS. Neuroimage 147, 960–963. doi: 10.1016/j.neuroimage.2016.11.022
Noury, N., and Siegel, M. (2017). Phase properties of transcranial electrical stimulation artifacts in electrophysiological recordings. Neuroimage 158, 406–416. doi: 10.1016/j.neuroimage.2017.07.010
Onoda, K., Kawagoe, T., Zheng, H., and Yamaguchi, S. (2017). Theta band transcranial alternating current stimulations modulates network behavior of dorsal anterior cingulate cortex. Sci. Rep. 7:, 3607. doi: 10.1038/s41598-017-03859-3857
O’Reardon, J. P., Solvason, H. B., Janicak, P. G., Sampson, S., Isenberg, K. E., Nahas, Z., et al. (2007). Efficacy and safety of transcranial magnetic stimulation in the acute treatment of major depression: a multisite randomized controlled trial. Biol. Psychiatry. 62, 1208–1216. doi: 10.1016/j.biopsych.2007.01.018
Pahor, A., and Jaušovec, N. (2014). The effects of theta transcranial alternating current stimulation (tACS) on fluid intelligence. Int. J. Psychophysiol. 93, 322–331. doi: 10.1016/j.ijpsycho.2014.06.015
Park, H.-J., and Friston, K. (2013). Structural and functional brain networks: from connections to cognition. Science 80:, 342. doi: 10.1126/science.1238411
Pessoa, L. (2014). Understanding brain networks and brain organization. Phys. Life Rev. 11, 400–435. doi: 10.1016/j.plrev.2014.03.005
Peters, S. K., Dunlop, K., and Downar, J. (2016). Cortico-Striatal-Thalamic loop circuits of the salience network: a central pathway in psychiatric disease and treatment. Front. Syst. Neurosci. 10:104. doi: 10.3389/fnsys.2016.00104
Reato, D., Rahman, A., Bikson, M., and Parra, L. C. (2013). Effects of weak transcranial alternating current stimulation on brain activity—a review of known mechanisms from animal studies. Front. Hum. Neurosci. 7:687. doi: 10.3389/fnhum.2013.00687
Riecke, L., Formisano, E., Herrmann, C. S., and Sack, A. T. (2015). 4-Hz transcranial alternating current stimulation phase modulates hearing. Brain Stimul. 8, 777–783. doi: 10.1016/j.brs.2015.04.004
Santarnecchi, E., Polizzotto, N. R., Godone, M., Giovannelli, F., Feurra, M., Matzen, L., et al. (2013). Frequency-dependent enhancement of fluid intelligence induced by transcranial oscillatory potentials. Curr. Biol. 23, 1449–1453. doi: 10.1016/j.cub.2013.06.022
Seager, M. A., Johnson, L. D., Chabot, E. S., Asaka, Y., and Berry, S. D. (2002). Oscillatory brain states and learning: impact of hippocampal theta-contingent training. Proc. Natl. Acad. Sci. U.S.A. 99, 1616–1620. doi: 10.1073/pnas.032662099
Shiozawa, P., Fregni, F., Benseñor, I. M., Lotufo, P. A., Berlim, M. T., Daskalakis, J. Z., et al. (2014). Transcranial direct current stimulation for major depression: an updated systematic review and meta-analysis. Int. J. Neuropsychopharmacol. 17, 1443–1452. doi: 10.1017/S1461145714000418
Silvanto, J., Muggleton, N., and Walsh, V. (2008). State-dependency in brain stimulation studies of perception and cognition. Trends Cogn. Sci. 12, 447–454. doi: 10.1016/j.tics.2008.09.004
Stagg, C. J., and Nitsche, M. A. (2011). Physiological basis of transcranial direct current stimulation. Neuroscientist. 17, 37–53. doi: 10.1177/1073858410386614
Vöröslakos, M., Takeuchi, Y., Brinyiczki, K., Zombori, T., Oliva, A., Fernández-Ruiz, A., et al. (2018). Direct effects of transcranial electric stimulation on brain circuits in rats and humans. Nat. Commun. 9:, 483. doi: 10.1038/s41467-018-02928-2923
Voss, U., Holzmann, R., Hobson, A., Paulus, W., Koppehele-Gossel, J., Klimke, A., et al. (2014). Induction of self awareness in dreams through frontal low current stimulation of gamma activity. Nat. Neurosci. 17, 810–814. doi: 10.1038/nn.3719
Vossen, A., Gross, J., and Thut, G. (2015). Alpha power increase after transcranial alternating current stimulation at alpha frequency (a-tACS) reflects plastic changes rather than entrainment. Brain Stimul. 8, 499–508. doi: 10.1016/j.brs.2014.12.004
Wagner, T., Fregni, F., Fecteau, S., Grodzinsky, A., Zahn, M., and Pascual-Leone, A. (2007). Transcranial direct current stimulation: a computer-based human model study. Neuroimage 35, 1113–1124. doi: 10.1016/j.neuroimage.2007.01.027
Wagner, T., Zahn, M., Grodzinsky, A. J., and Pascual-Leone, A. (2004). Three-dimensional head model simulation of transcranial magnetic stimulation. IEEE Trans. Biomed. Eng. 51, 1586–1598. doi: 10.1109/TBME.2004.827925
Wilsch, A., Neuling, T., Obleser, J., and Herrmann, C. S. (2018). Transcranial alternating current stimulation with speech envelopes modulates speech comprehension. Neuroimage 172, 766–774. doi: 10.1016/j.neuroimage.2018.01.038
Zaehle, T., Rach, S., and Herrmann, C. S. (2010). Transcranial alternating current stimulation enhances individual alpha activity in human EEG. PLoS One 5:e13766. doi: 10.1371/journal.pone.0013766
Zaghi, S., Acar, M., Hultgren, B., Boggio, P. S., and Fregni, F. (2010). Noninvasive brain stimulation with low-intensity electrical currents: putative mechanisms of action for direct and alternating current stimulation. Neuroscientist 16, 285–307. doi: 10.1177/1073858409336227
Zanos, S., Rembado, I., Chen, D., and Fetz, E. E. (2018). Phase-locked stimulation during cortical beta oscillations produces bidirectional synaptic plasticity in awake monkeys. Curr. Biol. 28, 2515.e–2526.e4. doi: 10.1016/j.cub.2018.07.009
Zrenner, C., Belardinelli, P., Müller-Dahlhaus, F., and Ziemann, U. (2016). Closed-Loop neuroscience and non-invasive brain stimulation: a tale of two loops. Front. Cell. Neurosci. 10:92. doi: 10.3389/fncel.2016.00092
Keywords: brain stimulation, transcranial alternating current stimulation, transcranial pulsed current stimulation, transcranial electrical stimulation, phase-locked brain stimulation, closed-loop brain stimulation, salience network
Citation: Mansouri F, Shanbour A, Mazza F, Fettes P, Zariffa J and Downar J (2019) Effect of Theta Transcranial Alternating Current Stimulation and Phase-Locked Transcranial Pulsed Current Stimulation on Learning and Cognitive Control. Front. Neurosci. 13:1181. doi: 10.3389/fnins.2019.01181
Received: 06 May 2019; Accepted: 18 October 2019;
Published: 14 November 2019.
Edited by:
Mikhail Lebedev, Duke University, United StatesReviewed by:
Leandro Da Costa Lane Valiengo, University of São Paulo, BrazilMaarten A. Frens, Sophia Children’s Hospital, Netherlands
Anthony Terrence O’Brien, Massachusetts Institute of Technology, United States
Copyright © 2019 Mansouri, Shanbour, Mazza, Fettes, Zariffa and Downar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jonathan Downar, am9uYXRoYW4uZG93bmFyQHVobi5jYQ==