 Daniel Saiz-Sanchez*
Daniel Saiz-Sanchez* Isabel Ubeda-Bañon
Isabel Ubeda-Bañon Alicia Flores-Cuadrado
Alicia Flores-Cuadrado Melania Gonzalez-Rodriguez
Melania Gonzalez-Rodriguez Sandra Villar-Conde
Sandra Villar-Conde Veronica Astillero-Lopez
Veronica Astillero-Lopez Alino Martinez-Marcos
Alino Martinez-Marcos
- Neuroplasticity and Neurodegeneration Laboratory, Ciudad Real Medical School, CRIB, University of Castilla-La Mancha, Ciudad Real, Spain
Alzheimer’s and Parkinson’s diseases are the most prevalent neurodegenerative disorders in aging. Hyposmia has been described as an early symptom that can precede cognitive and motor deficits by decades. Certain regions within the olfactory system, such as the anterior olfactory nucleus, display the neuropathological markers tau and amyloid-β or α-synuclein from the earliest stages of disease progression in a preferential manner. Specific neuronal subpopulations, namely those expressing somatostatin (SST), are preferentially affected throughout the olfactory and limbic systems. SST is a neuropeptide present in a subpopulation of GABAergic interneurons throughout the brain and its main function is to inhibit principal neurons and/or other interneurons. It has been reported that SST expression is reduced by 50% in Alzheimer’s disease and that it is related to the formation of Aβ oligomers. The mechanisms underlying the preferential vulnerability of SST-expressing neurons in Alzheimer’s disease (and, to a minor extent, in Parkinson’s disease) are not known but analysis of the available data could shed light on their etiology. This short review aims to update the knowledge of functional features of somatostatin within the olfactory system and its role in olfactory deficits during neurodegeneration.
Somatostatin and Olfactory System
Somatostatin-14 (SST-14), a peptide composed of 14 amino acids, was first detected in the hypothalamus and was shown to inhibit the secretion of growth hormone (Brazeau et al., 1973). Later, SST-28 was identified in the intestine (Pradayrol et al., 1980). Both were derived from the same prohormone and showed similar affinity to at least six SST receptors (SSTR1–5, two isoforms SSTR2A–2B). These receptors are members of the G-protein-coupled seven-transmembrane domain receptor family, which are broadly expressed in the brain, including all olfactory structures (Olias et al., 2004).
Approximately 20% of neurons in the cerebral cortex are interneurons, most of them expressing γ-amino-butyric acid (GABA) and divided into three non-overlapping classes: parvalbumin-expressing, 5-HT3A receptor-expressing and somatostatin-expressing populations (Riedemann, 2019). This latter population can be, in turn, morphologically divided into Martinotti cells and non-Martinotti cells (among others, long-range projecting interneurons, basket cells and double-bouquet cells). Their axons typically target the distal dendrites of pyramidal cells (contrary to parvalbumin-expressing cells), creating a dense wiring into the local network with high basal firing activity that continues in the absence of synaptic input (Urban-Ciecko and Barth, 2016). The pivotal role of somatostatin-positive interneurons on disinhibition (inhibiting GABAergic interneurons and promoting pyramidal cell activation) has been recently demonstrated, and it appears to be essential for long-term regulation and network metaplasticity, which may be important for hippocampal-dependent learning and memory (Artinian and Lacaille, 2018). During development, cortical interneurons including somatostatin-positive cells derive from the medial ganglionic eminence (Hu et al., 2017), while those interneurons reaching the olfactory bulb originate in the lateral ganglionic eminence (Wichterle et al., 2001). Importantly, olfactory system together with the hippocampus constitutes a neurogenic niche during adulthood and specific interneuron population are periodically replaced (Lledo et al., 2008).
Olfactory sensory neurons are placed in the olfactory epithelium in the nasal cavity and send their axons to the olfactory bulb where primary olfactory information is processed. From here, the main cells (mitral and tufted cells) then send their projections to the rest of the olfactory cortices: anterior olfactory nucleus, olfactory tubercle, piriform cortex, amygdala, and entorhinal cortex (Martinez-Marcos, 2009). The anterior olfactory nucleus is a key structure that constitutes the first relay of olfactory information and it also sends ipsilateral and contralateral projections to the rest of olfactory areas (Brunjes et al., 2005). The piriform cortex is considered as the principal olfactory cortex and it is mainly involved in odor perception (Courtiol and Wilson, 2017). Specifically, the amygdala and the entorhinal cortices are multimodal areas that receive olfactory information, among others, and in turn send projections to the hippocampus (McDonald and Mott, 2017). This latter circuit participates in olfactory emotion and olfactory memory formation (Kadohisa, 2013).
Somatostatin -expressing cells are present in all olfactory areas as well as SSTRs. In mouse, SST is expressed in the granule cell layer and within the inner part of the external plexiform layer in the olfactory bulb (Lepousez et al., 2010a) and within the different subregions of the anterior olfactory nucleus (Brunjes et al., 2011). Regarding the olfactory cortices, SST-expressing cells are located mostly in layers II and III, while terminal axons target dendrites in layer I of the piriform cortex (Suzuki and Bekkers, 2010). In the amygdala, SST is present in deeper layers of olfactory cortical subregions, although non-olfactory central nucleus accumulates the highest levels (Real et al., 2009). The olfactory entorhinal cortex, thus corresponding to the lateral entorhinal cortex, contains large number of SST interneurons and fibers in all layers (Witter et al., 2017). On the other hand, SSTRs1–4 are specifically distributed within the olfactory bulb (Nocera et al., 2019) and they are highly expressed in the piriform cortex, olfactory amygdala, namely anterior cortical and posteromedial cortical nuclei and within the entorhinal cortex (Breder et al., 1992; Fehlmann et al., 2000; Hannon et al., 2002). Interestingly, SSTR5 is absent within the olfactory system with the exception of the olfactory tubercle (Feuerbach et al., 2000). In human brain, SST interneurons are present in all olfactory areas (Figure 1). In the olfactory bulb SST is sparse within the EPL while its expression is very abundant around the anterior olfactory nucleus (Figures 1A,A’,A”,B,B’; Smith et al., 1993). SST is also present in the piriform cortex, particularly in layers II and III (Saiz-Sanchez et al., 2015). Interestingly, the piriform cortex is divided into two different portions, namely the anterior portion placed at the frontal cortex and a posterior portion located at the temporal lobe (Figures 1C,C’). Temporal lobe also includes key olfactory areas involved in olfactory memory formation such as the amygdala and the entorhinal cortex (LaBar and Cabeza, 2006). SST is present in all amygdaloid nuclei including cortical olfactory amygdala (Unger et al., 1988) and in all layers of the entorhinal cortex (Figures 1D,D’,D”; Chan-Palay, 1987). SSTRs studies in human brain mostly show distribution patterns regarding neocortex and/or limbic system including the amygdala and the hippocampal formation, while olfactory system is still poorly examined (Reubi et al., 1986; Schindler et al., 1998).
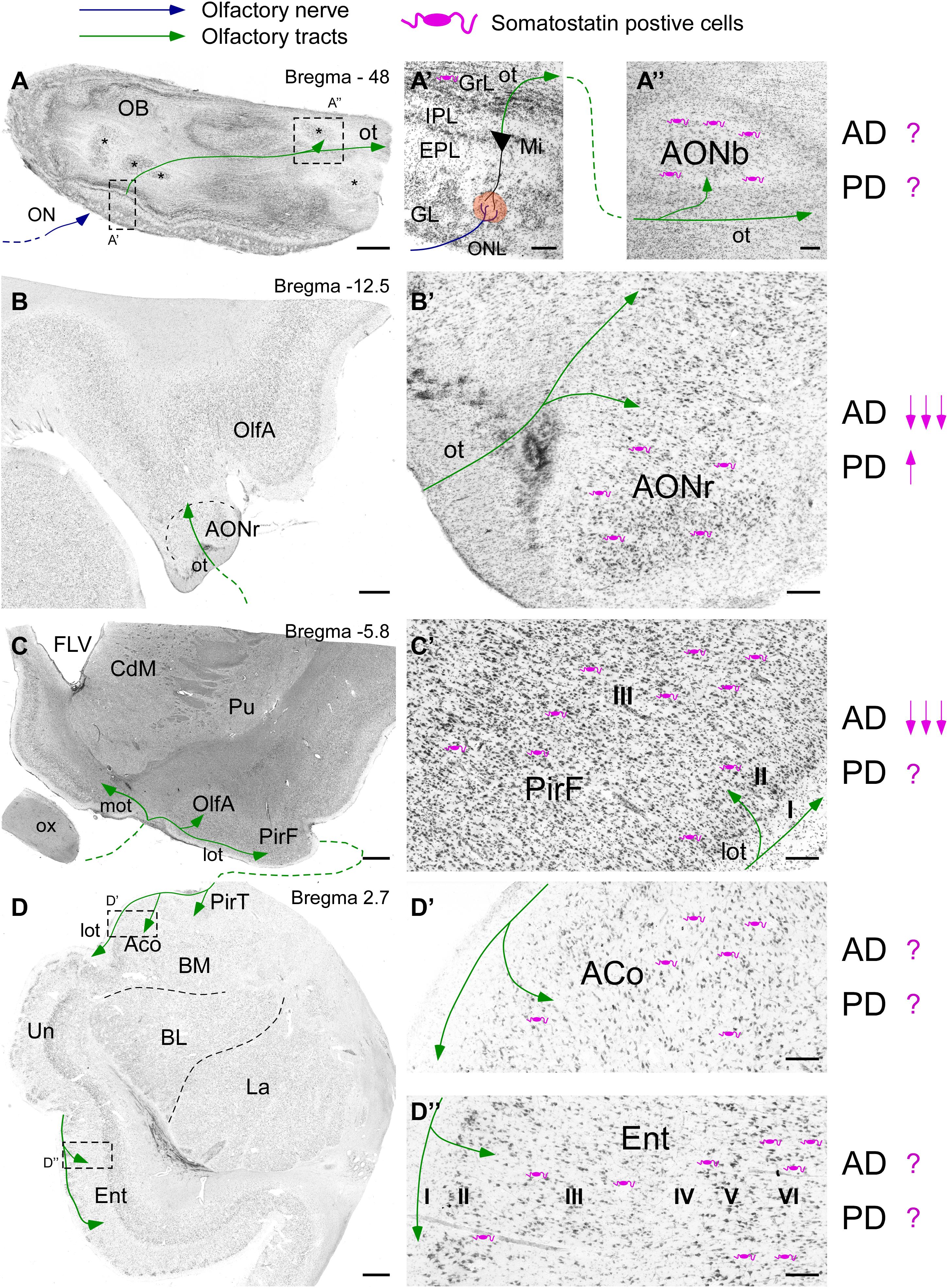
Figure 1. Scheme of the human olfactory system based on Nissl staining including the olfactory bulb (A) including different portions of the anterior olfactory nucleus (asterisks), the olfactory peduncle (B), the piriform cortex (C), the amygdala and the entorhinal cortex (D). Bregma levels are indicated based on human brain atlas (Mai et al., 2015). Somatostatin cells (purple) localization and their variation regarding Alzheimer’s and Parkinson’s diseases are specified as described in the literature. High magnification images show (A’) the different layers and (A”) the anterior olfactory nucleus within olfactory bulb. Note the representation of the first olfactory relay placed in the glomeruli (orange circle) between the axons coming from the olfactory neurons forming the olfactory nerve (blue line) and the dendrites of mitral cells. Then, the mitral cells form the olfactory tract (green line), which projects over all olfactory areas. (B’) represents the site of entry of the references from mitral cells into cortex in the frontal lobe, where the retrobulbar portion of the anterior olfactory nucleus can be observed. Once the olfactory peduncle contacts with cortex two different olfactory tracts appear (C); the medial olfactory tract (contralateral projections) and the lateral olfactory tract (ipsilateral projections). (C’) Represents the piriform cortex at its frontal subdivision. The typical three-layer histology is indicated. Note that somatostatin cells are in layer II and mainly in layer III. Afterward, olfactory tract reaches the temporal lobe (D), including the temporal subdivision of the piriform cortex, the cortical amygdala (D’) and the most rostral portion of the entorhinal cortex (D”). Note that no specific layer topography exists within the amygdala. On the contrary well defined six layers can be observed in the entorhinal cortex. Scale bars for A,B = 1000 μm; C,D = 2000 μm; and A’–D” = 200 μm.
Beside its cognitive and neuroendocrine functions (Viollet et al., 2008), central SST is involved in olfactory information processing such as olfactory detection and discrimination behaviors (Lepousez et al., 2010b; Nocera et al., 2019). Alternatively, SST contributes to additional actions related to olfaction, namely anxiety or fear-related behaviors (Yang et al., 2016). During the first relay of olfactory information SST regulates odor discrimination through SSTR2 receptor expressed by mitral cells (Lepousez et al., 2010b; Nocera et al., 2019). Furthermore, SST function related to the anterior olfactory nucleus, which constitutes the first relay from mitral cells, remains unknown. Piriform cortex is the main olfactory cortical area involved in the formation of odor percepts (Howard et al., 2009) and it regulates both excitation and inhibition network by inhibiting other interneurons, including other somatostatin cells and principal cells and they may help to discriminate odor responses from background cortical activity (Large et al., 2016). Moreover, SST cells regulate neurons by subtractive inhibition, which enhances the threshold for sensory input to trigger a response, that is independent of odor identity and intensity (Sturgill and Isaacson, 2015). Entorhinal cortex is involved in olfactory associative learning and recognition abilities (Nilssen et al., 2019). However, SST participation on olfactory processing within these areas is still poorly unknown. Nevertheless, SST distribution differs between mouse and rat (Brunjes et al., 2011).
Somatostatin and Parkinson’s Disease
Olfactory deficits have been reported as a preclinical risk factor for the development of PD (Ponsen et al., 2004). Moreover, the main pathological marker (accumulation of intracellular aggregates of α-synuclein forming Lewy bodies) is present in key olfactory structures, such as the anterior olfactory nucleus, from the earliest stages of disease progression (Del Tredici and Braak, 2016).
Postmortem studies have reported that around 8% of cells containing Lewy pathology in the olfactory bulb are somatostatinergic cells, as compared to more than 50% of calcium-binding protein-expressing cells, particularly those expressing calbindin (CB) and calretinin (CR) and to a lesser extent parvalbumin-expressing cells (PV) (Ubeda-Banon et al., 2010). Studies comparing control vs. PD cases have revealed a significant decrease of SST and an increase of PV in the AON (Ubeda-Banon et al., 2017). These low percentages of co-localization of SST with Lewy pathology were also observed in the amygdala (Flores-Cuadrado et al., 2017). Studies carried out in control, non-demented and demented PD cases also suffering from Alzheimer’s disease demonstrated that only comorbid cases showed a 40% reduction of SST in the frontal and temporal cortices (Beal et al., 1986).
Somatostatin and Alzheimer’s Disease
The two main hallmarks of AD are extracellular deposition of amyloid-β (Aβ1–42) forming senile plaques and intracellular aggregates of tau protein forming neurofibrillary tangles (Selkoe, 2001). Both have been described as affecting the olfactory system, while tau is involved early, especially in the entorhinal cortex (Braak et al., 2006) and within the anterior olfactory nucleus (Attems and Jellinger, 2006). These features make the olfactory system especially vulnerable to early stage disease progression.
Reduction of SST in AD was reported 40 years ago (Davies et al., 1980). Nowadays, SST reduction in the early stages and its involvement in memory formation is well established (Epelbaum et al., 2009). Other interneuron subpopulations such as CR or CB cells are reduced as well. Interestingly, as in PD, PV cells are not reduced in olfactory cortex (Saiz-Sanchez et al., 2015). However, most studies have focused on the hippocampal formation, and olfactory system studies are scarce. Moreover, olfactory deficits, namely hyposmia and anosmia, appear in patients with mild cognitive impairment (MCI) and correlate with later evolution to AD dementia (Devanand et al., 2000). Despite SST reduction being common within most olfactory areas (Saiz-Sanchez et al., 2010, 2015) olfactory explicit memory (namely odor identification and odor recognition, which are hippocampus dependent) is thought to be more deeply involved in AD as compared with olfactory threshold detection or implicit memory tasks such as habituation or sensitization (Quarmley et al., 2017). Nonetheless, the use of standardized tests remains controversial.
Somatostatin dysfunction is involved early in memory deficits observed in mouse models and may be affected by Aβ1–42 deposition (Schmid et al., 2016). Remarkably, SST is the main binder of Aβ1–42 and can encourage the formation of different Aβ1–42 oligomers by acquiring amyloid properties (Wang et al., 2017; Solarski et al., 2018). In agreement with this, SST and Aβ1–42 levels in the cerebrospinal fluid seem to be correlated (Duron et al., 2018) and co-localization of both in the human brain, including the olfactory cortex, are widespread histological features (Saiz-Sanchez et al., 2010, 2015). In fact, SST has been highlighted as a regulator of Aβ1–42 deposition (Saito et al., 2005). On the other hand, positron emission tomography results link olfactory impairment with tau rather than amyloid deposition (Risacher et al., 2017). However, we cannot rule out the aging influence on olfactory impairment and not only tau accumulation itself (Martel et al., 2015). Interestingly, cortistatin (a neuropeptide related to SST) can induce the phosphorylation of tau and may be linked with AD pathophysiology (Rubio et al., 2008).
Overview
The olfactory system is early and severely affected by pathologic proteins in both AD and PD. On the other hand, SST is unequally involved, being strongly involved in AD and having a weaker effect in PD. SST is reduced during the early stages of AD, including the olfactory areas, and may be related to Aβ1–42 and/or tau pathophysiology. However, the knowledge of both SST and SSTRs involvement in the human olfactory system is very scarce. Olfactory deficits may be related to SST deficiencies and to memory impairment due to tau deposition. In fact, SST is preserved in non-demented PD cases. Finally, anterior olfactory nucleus highlights as a key olfactory area; it is one of the earliest affected by tau accumulation and contains high quantities of SST. Further studies may indicate which olfactory deficiencies are more accurate for early diagnoses and help to refine the scope of SST as a potential therapeutic target.
Author Contributions
DS-S has coordinated all information, has written the manuscript, and has performed the figure. IU-B, AF-C, and SV-C have conducted the literature search focused on Parkinson’s disease and have written the related draft. MG-R and VA-L have conducted the literature search focused on Alzheimer’s disease and have written the related draft. AM-M has supervised all procedures.
Funding
Sponsored by the Spanish Ministry of Economy and Competitiveness-FEDER (Grant no. SAF2016-75768-R) and the Autonomous Government of Castilla-La Mancha/FEDER (Grant no. SBPLY/17/180501/000430) to AM-M.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The human tissues used to generate Figure 1 were provided by biobanks IDIBAPS (Barcelona), BT-CIEN (Madrid), and BIOBANC-MUR (Murcia). Experimental procedures for these samples were approved by the Ethical Committee for Clinical Research of the Ciudad Real University Hospital.
Abbreviations
ACo, cortical amygdala; AD, Alzheimer’s disease; AONb/AONr, anterior olfactory nucleus bulbar/retrobulbar portion; BL/BM, basolateral/basomedial amygdala; CdM, caudate nucleus medial; Ent, entorhinal cortex; EPL, external plexiform later; FLV, lateral ventricle frontal part; GL, glomerular layer; GrL, granule cell layer; IPL, internal plexiform layer; La, lateral amygdala; lot, lateral olfactory tract; Mi, mitral cell layer; mot, medial olfactory tract; OB, olfactory bulb; OlfA, olfactory area; ON, olfactory nerve; ot, olfactory tract; ox, optic chiasm; PirF/PirT, piriform cortex frontal/temporal subdivision; Pu, putamen; SST, somatostatin/somatostatin cells; Un, uncus.
References
Artinian, J., and Lacaille, J. C. (2018). Disinhibition in learning and memory circuits: new vistas for somatostatin interneurons and long-term synaptic plasticity. Brain. Res. Bull. 141, 20–26. doi: 10.1016/j.brainresbull.2017.11.012
Attems, J., and Jellinger, K. A. (2006). Olfactory tau pathology in Alzheimer disease and mild cognitive impairment. Clin. Neuropathol. 25, 265–271.
Beal, M. F., Mazurek, M. F., and Martin, J. B. (1986). Somatostatin immunoreactivity is reduced in Parkinson’s disease dementia with Alzheimer’s changes. Brain Res. 397, 386–388. doi: 10.1016/0006-8993(86)90644-x
Braak, H., Alafuzoff, I., Arzberger, T., Kretzschmar, H., and Del Tredici, K. (2006). Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol. 112, 389–404. doi: 10.1007/s00401-006-0127-z
Brazeau, P., Vale, W., Burgus, R., Ling, N., Butcher, M., Rivier, J., et al. (1973). Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179, 77–79. doi: 10.1126/science.179.4068.77
Breder, C. D., Yamada, Y., Yasuda, K., Seino, S., Saper, C. B., and Bell, G. I. (1992). Differential expression of somatostatin receptor subtypes in brain. J. Neurosci. 12, 3920–3934. doi: 10.1523/jneurosci.12-10-03920.1992
Brunjes, P. C., Illig, K. R., and Meyer, E. A. (2005). A field guide to the anterior olfactory nucleus (cortex). Brain Res. Brain Res. Rev. 50, 305–335. doi: 10.1016/j.brainresrev.2005.08.005
Brunjes, P. C., Kay, R. B., and Arrivillaga, J. P. (2011). The mouse olfactory peduncle. J. Comp. Neurol. 519, 2870–2886. doi: 10.1002/cne.22662
Chan-Palay, V. (1987). Somatostatin immunoreactive neurons in the human hippocampus and cortex shown by immunogold/silver intensification on vibratome sections: coexistence with neuropeptide Y neurons, and effects in Alzheimer-type dementia. J. Comp. Neurol. 260, 201–223. doi: 10.1002/cne.902600205
Courtiol, E., and Wilson, D. A. (2017). The olfactory mosaic: bringing an olfactory network together for odor perception. Perception 46, 320–332. doi: 10.1177/0301006616663216
Davies, P., Katzman, R., and Terry, R. D. (1980). Reduced somatostatin-like immunoreactivity in cerebral cortex from cases of Alzheimer disease and Alzheimer senile dementa. Nature 288, 279–280. doi: 10.1038/288279a0
Del Tredici, K., and Braak, H. (2016). Review: Sporadic Parkinson’s disease: development and distribution of alpha-synuclein pathology. Neuropathol. Appl. Neurobiol. 42, 33–50. doi: 10.1111/nan.12298
Devanand, D. P., Michaels-Marston, K. S., Liu, X., Pelton, G. H., Padilla, M., Marder, K., et al. (2000). Olfactory deficits in patients with mild cognitive impairment predict Alzheimer’s disease at follow-up. Am. J. Psychiatry 157, 1399–1405. doi: 10.1176/appi.ajp.157.9.1399
Duron, E., Vidal, J. S., Grousselle, D., Gabelle, A., Lehmann, S., Pasquier, F., et al. (2018). Somatostatin and neuropeptide Y in cerebrospinal fluid: correlations with amyloid peptides Abeta1-42 and tau proteins in elderly patients with mild cognitive impairment. Front. Aging Neurosci. 10:297. doi: 10.3389/fnagi.2018.00297
Epelbaum, J., Guillou, J. L., Gastambide, F., Hoyer, D., Duron, E., and Viollet, C. (2009). Somatostatin, Alzheimer’s disease and cognition: an old story coming of age? Prog. Neurobiol. 89, 153–161. doi: 10.1016/j.pneurobio.2009.07.002
Fehlmann, D., Langenegger, D., Schuepbach, E., Siehler, S., Feuerbach, D., and Hoyer, D. (2000). Distribution and characterisation of somatostatin receptor mRNA and binding sites in the brain and periphery. J. Physiol. Paris 94, 265–281. doi: 10.1016/s0928-4257(00)00208-4
Feuerbach, D., Fehlmann, D., Nunn, C., Siehler, S., Langenegger, D., Bouhelal, R., et al. (2000). Cloning, expression and pharmacological characterisation of the mouse somatostatin sst(5) receptor. Neuropharmacology 39, 1451–1462. doi: 10.1016/s0028-3908(00)00063-0
Flores-Cuadrado, A., Ubeda-Banon, I., Saiz-Sanchez, D., and Martinez-Marcos, A. (2017). Alpha-synucleinopathy in the human amygdala in Parkinson Disease: differential vulnerability of somatostatin- and parvalbumin-expressing neurons. J. Neuropathol. Exp. Neurol. 76, 754–758. doi: 10.1093/jnen/nlx054
Hannon, J. P., Petrucci, C., Fehlmann, D., Viollet, C., Epelbaum, J., and Hoyer, D. (2002). Somatostatin sst2 receptor knock-out mice: localisation of sst1-5 receptor mRNA and binding in mouse brain by semi-quantitative RT-PCR, in situ hybridisation histochemistry and receptor autoradiography. Neuropharmacology 42, 396–413. doi: 10.1016/s0028-3908(01)00186-1
Howard, J. D., Plailly, J., Grueschow, M., Haynes, J. D., and Gottfried, J. A. (2009). Odor quality coding and categorization in human posterior piriform cortex. Nat. Neurosci. 12, 932–938. doi: 10.1038/nn.2324
Hu, J. S., Vogt, D., Sandberg, M., and Rubenstein, J. L. (2017). Cortical interneuron development: a tale of time and space. Development 144, 3867–3878. doi: 10.1242/dev.132852
Kadohisa, M. (2013). Effects of odor on emotion, with implications. Front. Syst. Neurosci. 7:66. doi: 10.3389/fnsys.2013.00066
LaBar, K. S., and Cabeza, R. (2006). Cognitive neuroscience of emotional memory. Nat. Rev. Neurosci. 7, 54–64. doi: 10.1038/nrn1825
Large, A. M., Kunz, N. A., Mielo, S. L., and Oswald, A. M. (2016). Inhibition by somatostatin interneurons in olfactory cortex. Front. Neural. Circuits 10:62. doi: 10.3389/fncir.2016.00062
Lepousez, G., Csaba, Z., Bernard, V., Loudes, C., Videau, C., Lacombe, J., et al. (2010a). Somatostatin interneurons delineate the inner part of the external plexiform layer in the mouse main olfactory bulb. J. Comp. Neurol. 518, 1976–1994. doi: 10.1002/cne.22317
Lepousez, G., Mouret, A., Loudes, C., Epelbaum, J., and Viollet, C. (2010b). Somatostatin contributes to in vivo gamma oscillation modulation and odor discrimination in the olfactory bulb. J. Neurosci. 30, 870–875. doi: 10.1523/JNEUROSCI.4958-09.2010
Lledo, P. M., Merkle, F. T., and Alvarez-Buylla, A. (2008). Origin and function of olfactory bulb interneuron diversity. Trends Neurosci. 31, 392–400. doi: 10.1016/j.tins.2008.05.006
Mai, J., Majtanik, M., and Paxinos, G. (2015). Atlas of the Human Brain. Cambridge, MA: Academic Press.
Martel, G., Simon, A., Nocera, S., Kalainathan, S., Pidoux, L., Blum, D., et al. (2015). Aging, but not tau pathology, impacts olfactory performances and somatostatin systems in THY-Tau22 mice. Neurobiol. Aging 36, 1013–1028. doi: 10.1016/j.neurobiolaging.2014.10.033
Martinez-Marcos, A. (2009). On the organization of olfactory and vomeronasal cortices. Prog. Neurobiol. 87, 21–30. doi: 10.1016/j.pneurobio.2008.09.010
McDonald, A. J., and Mott, D. D. (2017). Functional neuroanatomy of amygdalohippocampal interconnections and their role in learning and memory. J. Neurosci. Res. 95, 797–820. doi: 10.1002/jnr.23709
Nilssen, E. S., Doan, T. P., Nigro, M. J., Ohara, S., and Witter, M. P. (2019). Neurons and networks in the entorhinal cortex: a reappraisal of the lateral and medial entorhinal subdivisions mediating parallel cortical pathways. Hippocampus 29, 1238–1254. doi: 10.1002/hipo.23145
Nocera, S., Simon, A., Fiquet, O., Chen, Y., Gascuel, J., Datiche, F., et al. (2019). Somatostatin serves a modulatory role in the mouse olfactory bulb: neuroanatomical and behavioral evidence. Front. Behav. Neurosci. 13:61. doi: 10.3389/fnbeh.2019.00061
Olias, G., Viollet, C., Kusserow, H., Epelbaum, J., and Meyerhof, W. (2004). Regulation and function of somatostatin receptors. J. Neurochem. 89, 1057–1091. doi: 10.1111/j.1471-4159.2004.02402.x
Ponsen, M. M., Stoffers, D., Booij, J., van Eck-Smit, B. L., Wolters, E., and Berendse, H. W. (2004). Idiopathic hyposmia as a preclinical sign of Parkinson’s disease. Ann. Neurol. 56, 173–181. doi: 10.1002/ana.20160
Pradayrol, L., Jornvall, H., Mutt, V., and Ribet, A. (1980). N-terminally extended somatostatin: the primary structure of somatostatin-28. FEBS Lett. 109, 55–58. doi: 10.1016/0014-5793(80)81310-x
Quarmley, M., Moberg, P. J., Mechanic-Hamilton, D., Kabadi, S., Arnold, S. E., Wolk, D. A., et al. (2017). Odor Identification Screening Improves Diagnostic Classification in Incipient Alzheimer’s Disease. J. Alzheimers Dis. 55, 1497–1507. doi: 10.3233/JAD-160842
Real, M. A., Heredia, R., Labrador Mdel, C., Davila, J. C., and Guirado, S. (2009). Expression of somatostatin and neuropeptide Y in the embryonic, postnatal, and adult mouse amygdalar complex. J. Comp. Neurol. 513, 335–348. doi: 10.1002/cne.21970
Reubi, J. C., Cortes, R., Maurer, R., Probst, A., and Palacios, J. M. (1986). Distribution of somatostatin receptors in the human brain: an autoradiographic study. Neuroscience 18, 329–346. doi: 10.1016/0306-4522(86)90158-2
Riedemann, T. (2019). Diversity and function of somatostatin-expressing interneurons in the cerebral cortex. Int J. Mol. Sci. 20:E2952. doi: 10.3390/ijms20122952
Risacher, S. L., Tallman, E. F., West, J. D., Yoder, K. K., Hutchins, G. D., Fletcher, J. W., et al. (2017). Olfactory identification in subjective cognitive decline and mild cognitive impairment: association with tau but not amyloid positron emission tomography. Alzheimers Dement. 9, 57–66. doi: 10.1016/j.dadm.2017.09.001
Rubio, A., Perez, M., de Lecea, L., and Avila, J. (2008). Effect of cortistatin on tau phosphorylation at Ser262 site. J. Neurosci. Res. 86, 2462–2475. doi: 10.1002/jnr.21689
Saito, T., Iwata, N., Tsubuki, S., Takaki, Y., Takano, J., Huang, S. M., et al. (2005). Somatostatin regulates brain amyloid beta peptide Abeta42 through modulation of proteolytic degradation. Nat. Med. 11, 434–439. doi: 10.1038/nm1206
Saiz-Sanchez, D., De la Rosa-Prieto, C., Ubeda-Banon, I., and Martinez-Marcos, A. (2015). Interneurons, tau and amyloid-beta in the piriform cortex in Alzheimer’s disease. Brain Struct. Funct. 220, 2011–2025. doi: 10.1007/s00429-014-0771-3
Saiz-Sanchez, D., Ubeda-Banon, I., de la Rosa-Prieto, C., Argandona-Palacios, L., Garcia-Munozguren, S., Insausti, R., et al. (2010). Somatostatin, tau, and beta-amyloid within the anterior olfactory nucleus in Alzheimer disease. Exp. Neurol. 223, 347–350. doi: 10.1016/j.expneurol.2009.06.010
Schindler, M., Holloway, S., Humphrey, P. P., Waldvogel, H., Faull, R. L., Berger, W., et al. (1998). Localization of the somatostatin sst2(a) receptor in human cerebral cortex, hippocampus and cerebellum. Neuroreport 9, 521–525. doi: 10.1097/00001756-199802160-00027
Schmid, L. C., Mittag, M., Poll, S., Steffen, J., Wagner, J., Geis, H. R., et al. (2016). Dysfunction of Somatostatin-Positive Interneurons Associated with Memory Deficits in an Alzheimer’s Disease Model. Neuron 92, 114–125. doi: 10.1016/j.neuron.2016.08.034
Selkoe, D. J. (2001). Alzheimer’s disease: genes, proteins, and therapy. Physiol. Rev. 81, 741–766. doi: 10.1152/physrev.2001.81.2.741
Smith, R. L., Baker, H., and Greer, C. A. (1993). Immunohistochemical analyses of the human olfactory bulb. J. Comp. Neurol. 333, 519–530. doi: 10.1002/cne.903330405
Solarski, M., Wang, H., Wille, H., and Schmitt-Ulms, G. (2018). Somatostatin in Alzheimer’s disease: a new role for an old player. Prion 12, 1–8. doi: 10.1080/19336896.2017.1405207
Sturgill, J. F., and Isaacson, J. S. (2015). Somatostatin cells regulate sensory response fidelity via subtractive inhibition in olfactory cortex. Nat. Neurosci. 18, 531–535. doi: 10.1038/nn.3971
Suzuki, N., and Bekkers, J. M. (2010). Inhibitory neurons in the anterior piriform cortex of the mouse: classification using molecular markers. J. Comp. Neurol. 518, 1670–1687. doi: 10.1002/cne.22295
Ubeda-Banon, I., Flores-Cuadrado, A., Saiz-Sanchez, D., and Martinez-Marcos, A. (2017). Differential effects of parkinson’s disease on interneuron subtypes within the human anterior olfactory nucleus. Front. Neuroanat. 11:113. doi: 10.3389/fnana.2017.00113
Ubeda-Banon, I., Saiz-Sanchez, D., de la Rosa-Prieto, C., Argandona-Palacios, L., Garcia-Munozguren, S., and Martinez-Marcos, A. (2010). Alpha-Synucleinopathy in the human olfactory system in Parkinson’s disease: involvement of calcium-binding protein- and substance P-positive cells. Acta Neuropathol. 119, 723–735. doi: 10.1007/s00401-010-0687-9
Unger, J. W., McNeill, T. H., Lapham, L. L., and Hamill, R. W. (1988). Neuropeptides and neuropathology in the amygdala in Alzheimer’s disease: relationship between somatostatin, neuropeptide Y and subregional distribution of neuritic plaques. Brain Res. 452, 293–302. doi: 10.1016/0006-8993(88)90033-9
Urban-Ciecko, J., and Barth, A. L. (2016). Somatostatin-expressing neurons in cortical networks. Nat. Rev. Neurosci. 17, 401–409. doi: 10.1038/nrn.2016.53
Viollet, C., Lepousez, G., Loudes, C., Videau, C., Simon, A., and Epelbaum, J. (2008). Somatostatinergic systems in brain: networks and functions. Mol. Cell. Endocrinol. 286, 75–87. doi: 10.1016/j.mce.2007.09.007
Wang, H., Muiznieks, L. D., Ghosh, P., Williams, D., Solarski, M., Fang, A., et al. (2017). Somatostatin binds to the human amyloid beta peptide and favors the formation of distinct oligomers. Elife 6:e28401. doi: 10.7554/eLife.28401
Wichterle, H., Turnbull, D. H., Nery, S., Fishell, G., and Alvarez-Buylla, A. (2001). In utero fate mapping reveals distinct migratory pathways and fates of neurons born in the mammalian basal forebrain. Development 128, 3759– 3771.
Witter, M. P., Doan, T. P., Jacobsen, B., Nilssen, E. S., and Ohara, S. (2017). Architecture of the entorhinal cortex a review of entorhinal anatomy in rodents with some comparative notes. Front. Syst. Neurosci. 11:46. doi: 10.3389/fnsys.2017.00046
Keywords: α-synuclein, amyloid-β, anterior olfactory nucleus, hyposmia, tau
Citation: Saiz-Sanchez D, Ubeda-Bañon I, Flores-Cuadrado A, Gonzalez-Rodriguez M, Villar-Conde S, Astillero-Lopez V and Martinez-Marcos A (2020) Somatostatin, Olfaction, and Neurodegeneration. Front. Neurosci. 14:96. doi: 10.3389/fnins.2020.00096
Received: 14 October 2019; Accepted: 23 January 2020;
Published: 19 February 2020.
Edited by:
Anna Menini, International School for Advanced Studies (SISSA), ItalyReviewed by:
Cécile Viollet, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceDaniel Vogt, Michigan State University, United States
Copyright © 2020 Saiz-Sanchez, Ubeda-Bañon, Flores-Cuadrado, Gonzalez-Rodriguez, Villar-Conde, Astillero-Lopez and Martinez-Marcos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Saiz-Sanchez, RGFuaWVsLlNhaXpAdWNsbS5lcw==