 Elif Doǧan
Elif Doǧan Hatice Aygün2
Hatice Aygün2 Gökhan Arslan
Gökhan Arslan Emil Rzayev
Emil Rzayev Mustafa Ayyıldız
Mustafa Ayyıldız Erdal Ağar
Erdal Ağar- 1Department of Physiology, Faculty of Medicine, Ondokuz Mayıs University, Samsun, Turkey
- 2Department of Physiology, Faculty of Medicine, Gaziosmanpasa University, Tokat, Turkey
- 3Department of Clinical Biochemistry, Faculty of Medicine, Ondokuz Mayıs University, Samsun, Turkey
P2X7 receptors (P2X7Rs) are ATP sensitive cation channels and have been shown to be effective in various epilepsy models. Absence epilepsy is a type of idiopathic, generalized, non-convulsive epilepsy. Limited data exist on the role of P2X7Rs and no data has been reported regarding the interaction between P2X7Rs and glutamate receptor NMDA in absence epilepsy. Thus, this study was designed to investigate the role of P2X7 and NMDA receptors and their possible interaction in WAG/Rij rats with absence epilepsy. Permanent cannula and electrodes were placed on the skulls of the animals. After the healing period of the electrode and cannula implantation, ECoG recordings were obtained during 180 min before and after drug injections. P2X7R agonist BzATP, at doses of 50 μg and 100 μg (intracerebroventricular; i.c.v.) and antagonist A-438079, at doses of 20 μg and 40 μg (i.c.v.) were administered alone or prior to memantine (5 mg/kg, intraperitoneal; i.p.) injection. The total number (in every 20 min), the mean duration, and the amplitude of spike-wave discharges (SWDs) were calculated and compared. Rats were decapitated and the right and left hemisphere, cerebellum, and brainstem were separated for the measurements of the advanced oxidation protein product (AOPP), malondialdehyde (MDA), superoxide dismutase (SOD), glutathione (GSH), catalase (CAT), glutathione peroxide (GPx), and glutathione reductase (GR). BzATP and A-438079 did not alter measured SWDs parameters, whereas memantine reduced them, which is considered anticonvulsant. BzATP did not alter the anticonvulsant effect of memantine, while A-438079 decreased the effect of memantine. Administration of BzATP increased the levels of SOD and GR in cerebrum hemispheres. A-438079 did not alter any of the biochemical parameters. Memantine reduced the levels of MDA, GSH, and GR while increased the level of CAT in the cerebrum. Administration of BzATP before memantine abolished the effect of memantine on MDA levels. The evidence from this study suggests that P2X7Rs does not directly play a role in the formation of absence seizures. P2X7Rs agonist, reduced the antioxidant activity of memantine whereas agonist of P2X7Rs reduced the anticonvulsant action of memantine, suggesting a partial interaction between P2X7 and NMDA receptors in absence epilepsy model.
Introduction
P2X7 receptors (P2X7Rs) are purinergic cation channels. They are sensitive to high concentrations of ATP in the extracellular space, and they have essential roles in inflammation (Burnstock and Knight, 2018). P2X7Rs could be permeable to small cations when they are stimulated within milliseconds. However, prolonged stimulation (within seconds) of P2X7Rs allows permeation by molecules with a mass of up to 900 Da, leading to the release of inflammatory cytokines and apoptosis (Burnstock and Knight, 2018). Stimulation of P2X7Rs also leads to neuroinflammatory responses, including, ADAM10, and ADAM17 activation, and causes the secretion of prostaglandin E2 and proinflammatory cytokines, such as interleukin-1β (IL-1β), IL-2, IL-4, IL-6, and tumor necrosis factor (TNF) (Engel et al., 2012b; Beamer et al., 2017). P2X7Rs mediate NLRP3 inflammasome-dependent IL-1β secretion following activation of NF-κB in the brain and immune cells (Albalawi et al., 2017; Burnstock and Knight, 2018). Therefore, P2X7Rs are a target for neurodegenerative diseases.
P2X7R expression has been widely shown in the central nervous system, including microglia, oligodendrocytes, and ependymal cells (Jimenez-Mateos et al., 2019). The P2X7R is mainly expressed on microglia, but not on the neurons (Jimenez-Pacheco et al., 2016; Kaczmarek-Hajek et al., 2018). P2X7R is expressed in the CA1 pyramidal and dentate granule neurons, as well as in microglia of epileptic mice (Jimenez-Pacheco et al., 2016) and in the neurons of rat hippocampus (Sperlagh et al., 2002). Moreover, P2X7R expression has been detected in both the astrocyte culture of brain/spinal cord slices (Illes et al., 2012, 2017). This conflicting data about functional expression of P2X7R could be attributed to both brain region-specific expression and the pathological conditions of the brain such as epilepsy (Beamer et al., 2017). P2X7R expression has increased in the hippocampus and neocortex regions of the brain in many epilepsy models (Jimenez-Pacheco et al., 2016; Huang et al., 2017; Rodriguez-Alvarez et al., 2017; Zeng et al., 2017; Jimenez-Mateos et al., 2019).
Absence epilepsy is a common neurological disease in children that affects educational success. Absence epilepsy is a loss of consciousness with a sudden pause in behavior, and if electroencephalography is recorded during seizures, bilateral synchronous 3-Hz frequency spike-wave discharges (SWDs) are observed (Russo et al., 2016; Fisher et al., 2017). Its pathophysiology is unclear, but it has been shown that absence seizures start from a glutamatergic focus located in the perioral region of the somatosensory cortex, and then this area affects the thalamus over time and creates a cortico-thalamo-cortical circuit (Meeren et al., 2005; Pinault and O’Brien, 2005). Wistar Albino Glaxo/Rijswijk (WAG/Rij) rats are also a validated genetic model of absence epilepsy characterized by SWDs on electroencephalography with a spontaneous pause in behavior (Russo et al., 2016). IL-1β increases in the somatosensory cortex and IL-1β antagonist administration reduces SWDs in the Genetic Absence Epilepsy Rat from Strasbourg (GAERS) model, which is also a validated genetic absence epilepsy model (Akin et al., 2011).
The effects of P2X7Rs on epilepsy have been investigated in various experimental epilepsy models (Fischer et al., 2016; Huang et al., 2017; Nieoczym et al., 2017; Rodriguez-Alvarez et al., 2017; Arslan et al., 2019). In a kainic acid-induced status epilepticus (SE) model, P2X7R expression was shown to increase, and seizure severity and neuronal death decreased after pre-treatment or post-treatment with intracerebroventricular (i.c.v.) injection of A-438079 or systemic administration of JNJ-47965567 (Engel et al., 2012a; Jimenez-Pacheco et al., 2013, 2016). Systemic administration of A-438079 reduced convulsions in the kainic acid-induced SE model and reduced neuronal death more than phenobarbital in 10-day-old rats (Mesuret et al., 2014). In a penicillin-induced epilepsy model, a P2X7R agonist showed proconvulsant effects that could be reversed by A-438079 and a T-type calcium channel blocker, whereas a P2X7R antagonist, A-438079, showed an anticonvulsant effect (Arslan et al., 2019). In a pentylenetetrazol (PTZ) kindling model, P2X7R antagonists, Brilliant Blue G (BBG) and tanshinone showed a slight delay in kindling development, and JNJ-47965567 and AFC-5128 showed a strong delay (Soni et al., 2015; Fischer et al., 2016). However, P2X7R antagonists were ineffective in fully kindled rats (Fischer et al., 2016). They also did not affect the number of seizures in the kainic acid-induced kindling model, but they gave rise to less severe chronic seizures (Amhaoul et al., 2016). Systemic administration of A-438079 reduced acute seizures during hypoxia in neonatal mice but had no effect on post-hypoxia seizures (Rodriguez-Alvarez et al., 2017). P2X7R antagonists did not affect the maximal electroshock seizure threshold test or PTZ seizure threshold test, but AFC-5128 or JNJ-47965567 combination with carbamazepine increased the seizure threshold more than carbamazepine alone (Fischer et al., 2016; Nieoczym et al., 2017). BBG showed a week anticonvulsant action on the threshold of 6 Hz induced psychomotor seizures in mice (Nieoczym et al., 2017). However, P2X7R knockout mice were more susceptible to pilocarpine-induced seizures (Kim and Kang, 2011), and blockade of P2X7Rs increased the number and severity of pilocarpine-induced seizures in mice (Rozmer et al., 2017).
On the other hand, NMDA receptors are cation channels, and over-stimulation leads to an increase in intracellular calcium, which could be toxic for cells (Rosini et al., 2019). Memantine, a non-competitive NMDA receptor antagonist, has also shown anticonvulsant effects in many experimental epilepsy models, including a penicillin-induced epilepsy model, an audiogenic seizure model, and a PTZ kindling model (Frey and Voits, 1991; Cakil et al., 2011; Kim et al., 2012; Zaitsev et al., 2015). Only one study has been conducted with WAG/Ola/Hsd rats, which are thought to be a model of genetic absence epilepsy (Frey and Voits, 1991), but there is no much information about this rat substrain. The impairment of gamma-aminobutyric acid (GABA) and glutamate leads to epilepsy. GABAergic antiepileptic drugs worsen absence seizures (Panayiotopoulos, 2001). However, the NMDA receptor NR1 subunit showed a decrease depending on age in WAG/Rij rats (van de Bovenkamp-Janssen et al., 2006). In addition, in both 2-month-old and 6-month-old WAG/Rij rats, the NR2B subunit was lower in various layers of the somatosensory cortex than in the Wistar rats of the same age (Karimzadeh et al., 2013). Stimulation of NMDA receptors increased SWDs in WAG/Rij rats, whereas NMDA receptor reduced SWDs (Peeters et al., 1994).
The P2X7R increases glutamate secretion in a vesicular and non-vesicular manner (Sperlagh et al., 2002; Cho et al., 2010). It also affects GABA and glutamate reuptake in a calcium-dependent manner (Barros-Barbosa et al., 2018). The P2X7R was found to be non-desensitizing, and it allows substantial calcium influx (Fischer et al., 2016). It activates intracellular signaling pathways (Takenouchi et al., 2010). Both P2X7R and NMDA receptor activation can activate pathways of reactive oxygen species (ROS) (Davidson and Duchen, 2006). ROS cause many changes such as aging, cardiovascular disease, cancer, and neurodegenerative diseases, including epilepsy, by damaging proteins, lipids, carbohydrates, and nucleic acids, and they can be controlled by antioxidant systems, preventing the formation of ROS, and damage (Droge, 2002; Terrone et al., 2019).
x There are limited data on the interaction between P2X7Rs and NMDA receptors in epilepsy. Thus, the effects of P2X7Rs and NMDA receptors and their relationship were investigated using a P2X7R agonist, BzATP, which is more selective for P2X7R than for other P2X receptors, a P2X7R antagonist, A-438079, which highly targets the P2X7R, and a selective antagonist of the NMDA receptor, memantine, in WAG/Rij rats with both electrophysiological and biochemical analysis methods.
Materials and Methods
Animals
In this study, 63 male, 6–8 months old, 250–300 g weighing WAG/Rij rats were used with the permission of Ondokuz Mayis University Animal Experiments Local Ethics Committee (2015/56). Wistar Albino Glaxo/Rijswijk (WAG/Rij) rats with spontaneous seizures were purchased from Charles River Lab (Germany). Animals were fed ad libitum and housed in heat-regulated rooms, on a 12 h light-dark cycle. All experimental procedures were conducted under the European Union Directive (2010/63/EU), and Turkish legislation acts concerning animal experiments.
Animals were divided into nine groups (n = 7) randomly as follows:
1. Control group (WAG/Rij rat)
2. Sham Group (WAG/Rij rat, 2 μl sterile distilled water; i.c.v.)
3. BzATP 50 μg (i.c.v.) group
4. BzATP 100 μg (i.c.v.) group
5. A-438079 20 μg (i.c.v.) group
6. A-438079 40 μg (i.c.v.) group
7. Memantine 5 mg/kg (i.p.) group
8. BzATP 100 μg (i.c.v.) + Memantine 5 mg/kg (i.p.) group
9. A-438079 20 μg; (i.c.v.) + Memantine 5 mg/kg (i.p.) group.
Surgery and Electrocorticography Recording
Animals were anesthetized and sedated with ketamine/xylazine (90/10 mg/kg; i.p.) and placed in the stereotaxic apparatus. After the skin was cut off about 3 cm and folded back, subcutaneous tissue was removed from the cranium. According to the rat brain atlas (Paxinos and Watson, 2007), four burr holes were drilled in the skull with a microdrill without damaging the dura mater. For ECoG recordings, three screw electrodes were placed into the holes as coordinates: first electrode; 2 mm anterior and 3.5 mm right lateral to bregma, second electrode; 6 mm posterior and 4 mm right lateral to bregma and earth electrode was placed on the cerebellum. For i.c.v. injections, an external cannula was advanced into the brain as coordinates: 1.1 mm posterior, 1.5 mm right lateral and 3.2 mm vertical to bregma. Afterward, the electrodes and cannula were fixed to the skull with dental cement. The cannula was covered with a dust cup until used. After the surgery, animals were housed individually for a week and habituated to the recording cage (32 cm × 30 cm in width, 50 cm high) for 3 days before the experimental procedure.
On the experiment day, animals were connected to the ECoG recording system (PowerLab, 16/SP, AD Instruments, Australia) by an isolated flexible cable. Baseline electrocorticography (ECoG) recordings were taken for 3 h from all awake animals at the same time of day (10:00 AM). After the drug injection, ECoGs recording continued for another 3 h (Aygun et al., 2019). The number of SWDs and the mean duration and amplitude of SWDs were measured and calculated for every 20 min offline with LabChart 7 Pro (AD Instruments, Australia) (Arslan et al., 2013, 2014).
Drugs and Drug Administration
Ketamine hydrochloride, xylazine hydrochloride, A-438079 hydrochloride hydrate, BzATP [2′(3′)-O-(4-Benzoylbenzoyl) adenosine 5′-triphosphate triethylammonium] and memantine hydrochloride were purchased from Sigma Chemical Co., St. Louis, MO, United States and dissolved in sterile distilled water. After obtaining 180 min of baseline ECoG recordings, A-438079, at the doses of 20 and 40 μg, and BzATP, at the doses of 50 and 100 μg were administered into the lateral ventricle within a thin internal cannula (4.2 mm vertical to the bregma) in a volume of 2 μl. Memantine, at a dose of 5 mg/kg, was injected intraperitoneally in a volume of 0.5 ml. For the interaction groups, memantine was administered 10 min after the chosen doses of BzATP (100 μg) or A-438079 (20 μg) (Arslan et al., 2019). The sham group was given sterile distilled water (2 μl, i.c.v.).
Biochemical Analysis
After the end of the ECoG recordings, all rats were decapitated following ketamine/xylazine anesthesia. Right and left hemisphere, cerebellum, and brainstem were separated in oxygenated artificial cerebrospinal fluid [aCSF containing (mM): NaCl, 124; KCl, 5; KH2PO4, 1.2; CaCl2, 2.4; MgSO4, 1.3; NaHCO3, 26; glucose, 10; HEPES, 10, at pH 7.4 when saturated with 95% O2 and 5% CO2]. The tissues were frozen in liquid nitrogen, homogenized immediately, and soaked in phosphate buffer solution (PBS, 10 mM, and pH 7.2). After 1 min of sonication at +4°C with 220 V (METU Electromechanical, Germany), homogenates were stored at −80°C. On the evaluation day, the homogenates were defrosted at room temperature and were centrifuged at +4°C for 5 min with 15,000 × g (Sigma 3K30, Germany) for biochemical analysis.
Tissue protein concentrations were determined with Lowry’s method (Lowry et al., 1951). The results were expressed per mg protein. Advanced protein oxidation products (AOPP), malondialdehyde (MDA), superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), glutathione peroxidase (GPx) and glutathione reductase (GR) concentrations in the tissues were examined with commercial rat ELISA kits (SunRed Biotechnology Co., Shanghai, China). These kits all use a double-antibody sandwich enzyme-linked immunosorbent assay (Arslan et al., 2019).
Statistical Analysis
Spike-wave discharges parameters were calculated, by using the raw data obtained from LabChart 7-Pro, with an excel program. The total number of SWDs after drug injections was calculated for every 20 min and these data were converted to percentage values by comparing to the baseline values. The mean duration and the amplitude of SWDs during 180 min after injections were calculated as a percentage by comparing to the baseline data.
SPSS 15.0 data analysis software was used for statistical analyses. The normality of the data was tested with the Shapiro–Wilk test. After verifying that the data were normally distributed paired-samples t-test was performed between dependent groups, and one- or two-way ANOVA and then post hoc Tukey tests were used for multiple comparisons. The results are expressed as mean ± standard error (SEM). For all statistical analyses, p < 0.05 was considered statistically significant.
Results
All rats showed SWDs in ECoG characterized by paroxysmal unresponsiveness to environmental stimuli (Figure 1). Paired-samples t-test revealed that the total numbers and the mean duration of SWDs did not significantly change after the injection of solvent compared to baseline values. The total numbers and mean durations of SWDs were 108.4 ± 4.8, 111.0 ± 3.8 and 8.67 ± 0.18, 8.76 ± 0.21 s during 180 min before and after the administration of sterile distilled water, respectively. The mean amplitudes of SWDs were 0.635 ± 0.039 and 0.648 ± 0.048 mV before and after the administration of sterile distilled water, respectively. There was no statistical difference regarding the parameters of SWDs between the control and sham groups (p > 0.05).
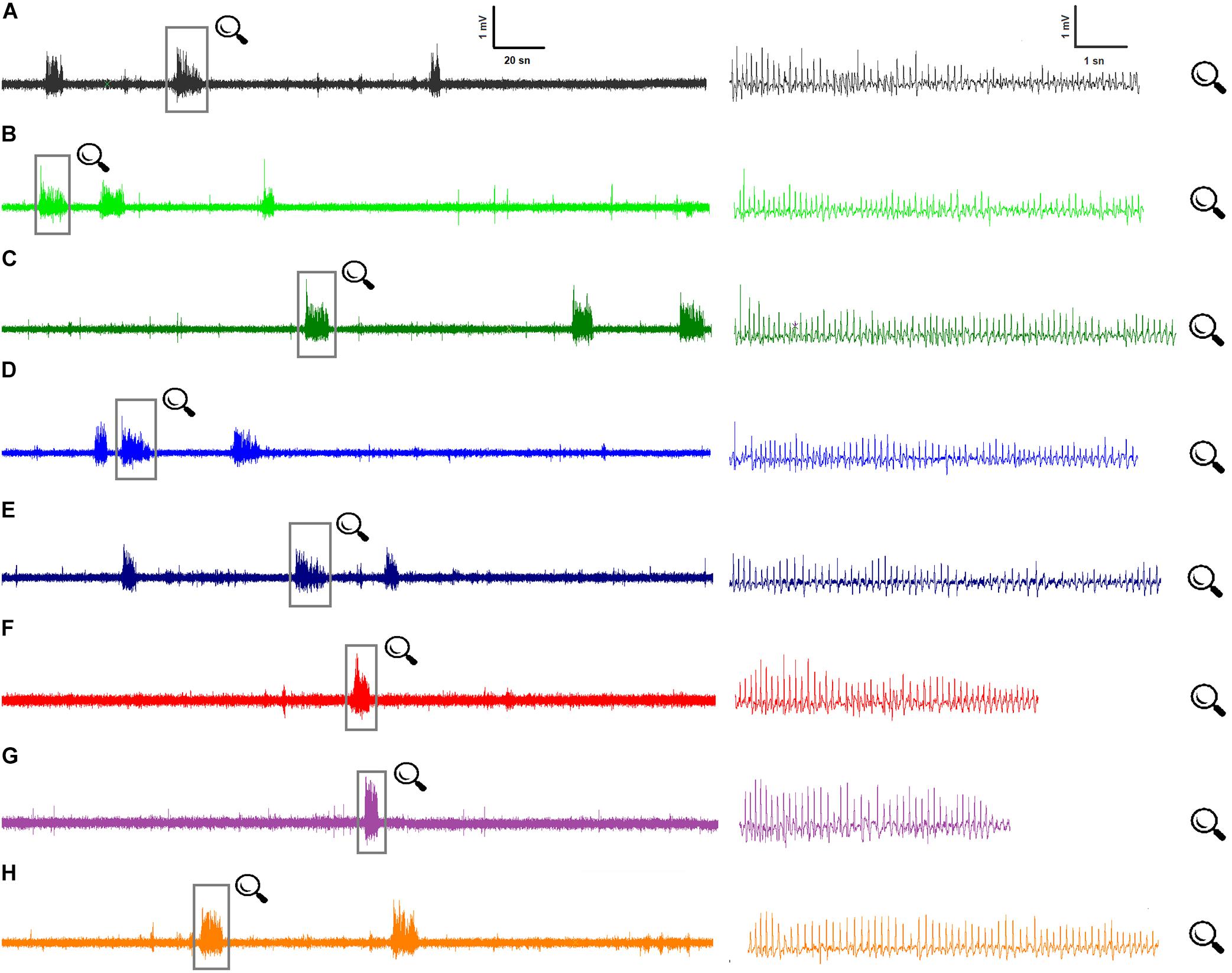
Figure 1. Representative ECoG recordings for all groups at 80th minute: (A) Control ECoG activity in WAG/Rij rats (n = 7); (B) BzATP, at the dose of 50 μg, i.c.v. (n = 7); (C) BzATP, at dose of 100 μg, i.c.v. (n = 7); (D) A-438079, at dose of 20 μg, i.c.v. (n = 7); (E) A-438079, at dose of 40 μg, i.c.v. (n = 7); (F) Memantine administration, at dose of 5 mg/kg, i.p. (n = 7); (G) BzATP (100 μg) administration 10 min before memantine (n = 7); (H) A-438079 (20 μg) administration 10 min before memantine (n = 7).
Table 1 shows the biochemical analysis for the left and right hemispheres, cerebellum, and brainstem of the groups. There was no significant difference between the sham group and the control group in any of the biochemical parameters.
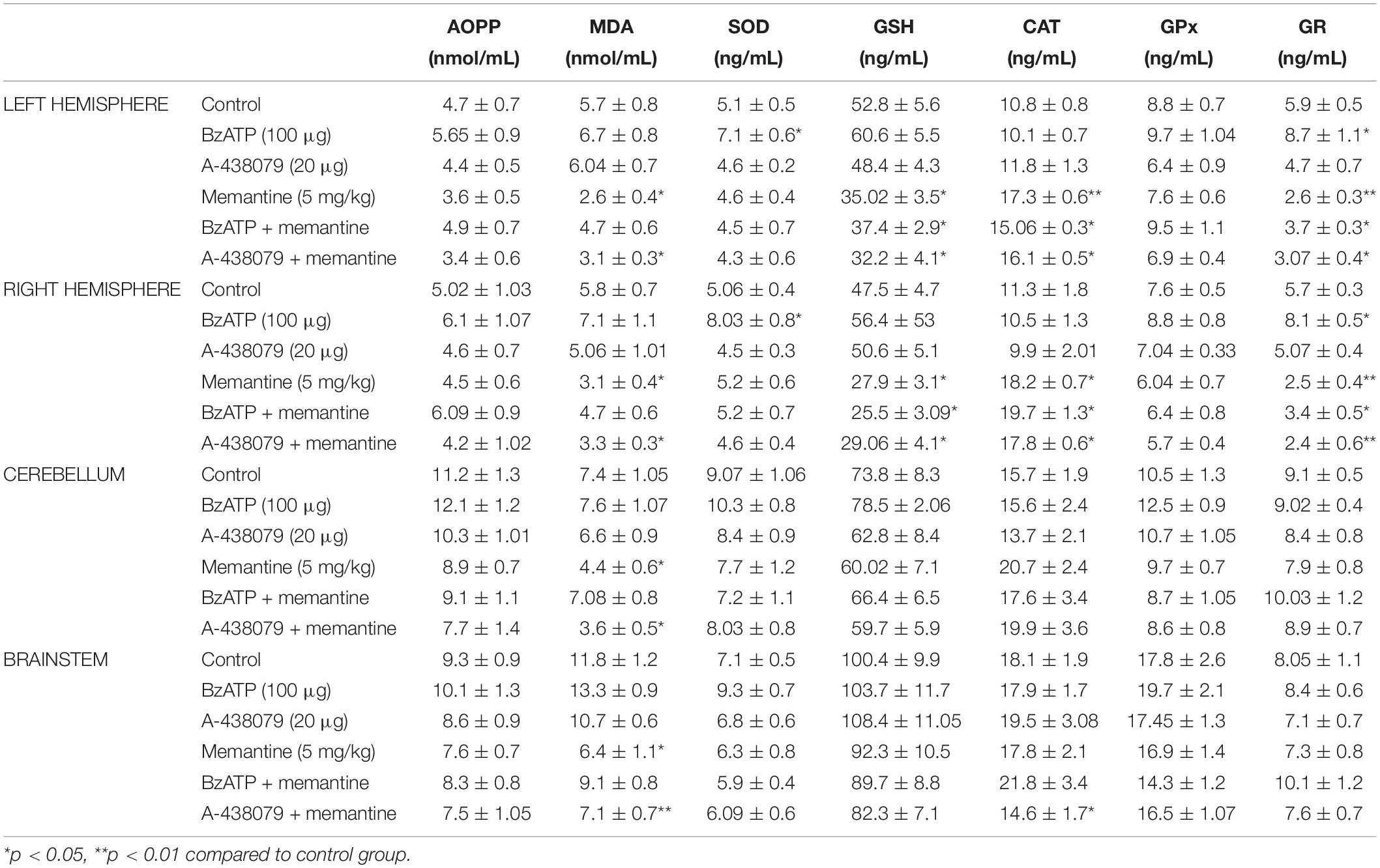
Table 1. The levels of advanced oxidation protein products (AOPP), malondialdehyde (MDA), superoxide dismutase (SOD), glutathione (GSH), catalase (CAT), glutathione peroxide (GPx), and glutathione reductase (GR) in the left and right hemispheres, cerebellum and brainstem of all experimental groups.
The Role of P2X7Rs in WAG/Rij Rats
Administration of P2X7Rs agonist BzATP, at the doses of 50 and 100 μg, did not significantly change any of the total numbers [F(8,162) = 0.12] and durations [F(2,18) = 1.54] of SWDs compared to the control group (Figure 2). Total numbers, mean durations and mean amplitudes of SWDs were 12.2 ± 0.9, 11.1 ± 0.9; 9 ± 0.4, 8.9 ± 0.4 s;0.6490 ± 0.013, 0.6148 ± 0.016 mV in the 80th minute in the BzATP 50 μg and BzATP 100 μg groups, respectively. After the injections of BzATP, at doses of 50 and 100 μg, the total numbers and mean durations of SWDs were 107.3 ± 4.2, 102.7 ± 5.4; 8.82 ± 0.38, 8.52 ± 0.29 s during 180 min, respectively. Injection of A-438079, at the doses of 20 and 40 μg, did not alter the total numbers [F(8,162) = 0.54] and durations [F(2,18) = 6.54] of SWDs compared to the control group (Figure 3). The total numbers and mean durations of SWDs were 107.8 ± 7.7, 102.7 ± 9.4; 8.38 ± 0.28, 8.2 6 ± 0.34 s during 180 min after the injections of 20 and 40 μg A-438079, respectively.
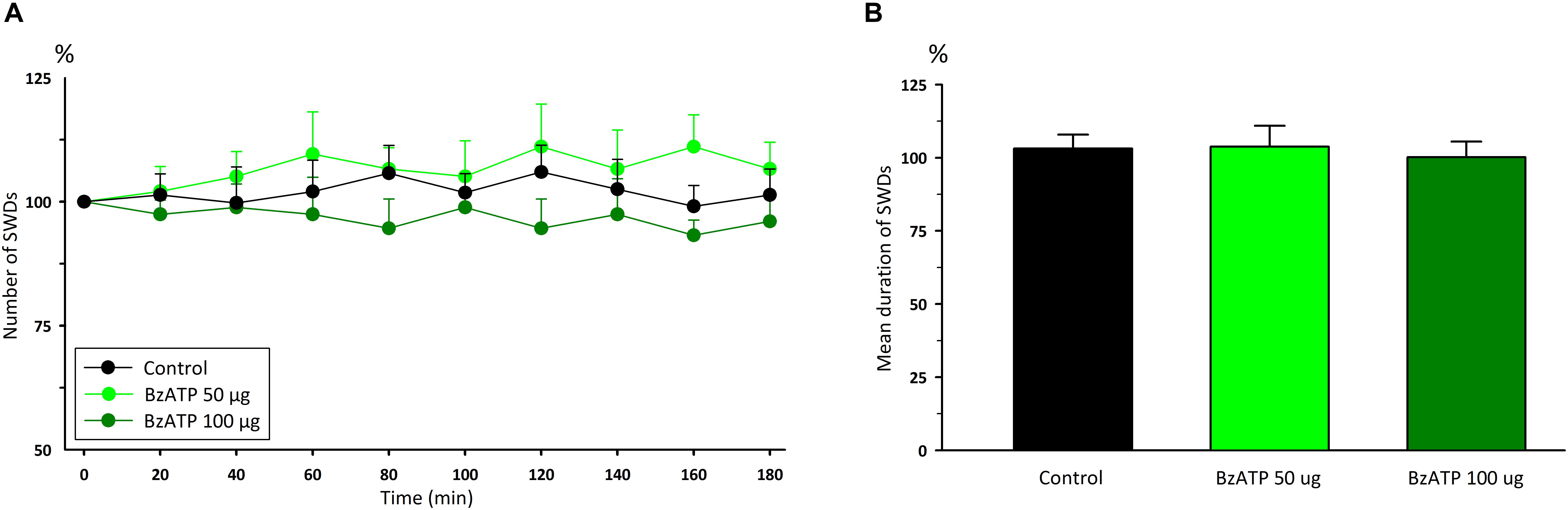
Figure 2. The effects of P2X7R agonist BzATP on (A) the total number of SWDs in every 20 min and (B) the mean duration of SWDs for 180 min.
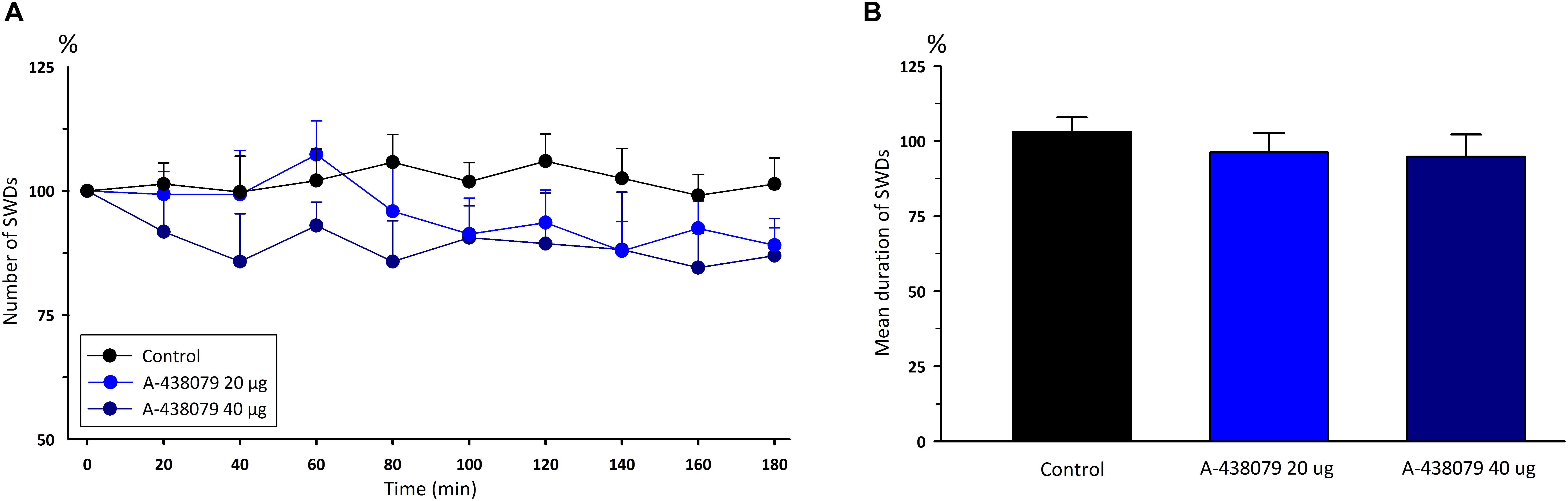
Figure 3. The effects of P2X7R antagonist A-438079 on (A) the total number of SWDs in every 20 min and (B) the mean duration of SWDs for 180 min.
BzATP, at a dose of 100 μg, injection significantly increased SOD and GR levels in the left and right hemispheres (p < 0.05). Other biochemical parameters were not different in the BzATP group compared to the control group (p > 0.05). A-438079, at a dose of 20 μg, did not alter any of the biochemical parameters (Table 1).
The Effect of Memantine on WAG/Rij Rats
Intraperitoneal injection of memantine (5 mg/kg) significantly decreased the total number [F(8,108) = 0.66, p < 0.001] and mean duration of SWDs [F(1,12) = 6.34, p < 0.05] 20 min after injection compared to control group (Figure 4). The total number, mean duration and amplitude of SWDs were 32.5 ± 3.6; 7.74 ± 0.19 s and 0.635 ± 0.03 mV during 180 min after the administration of memantine, respectively.
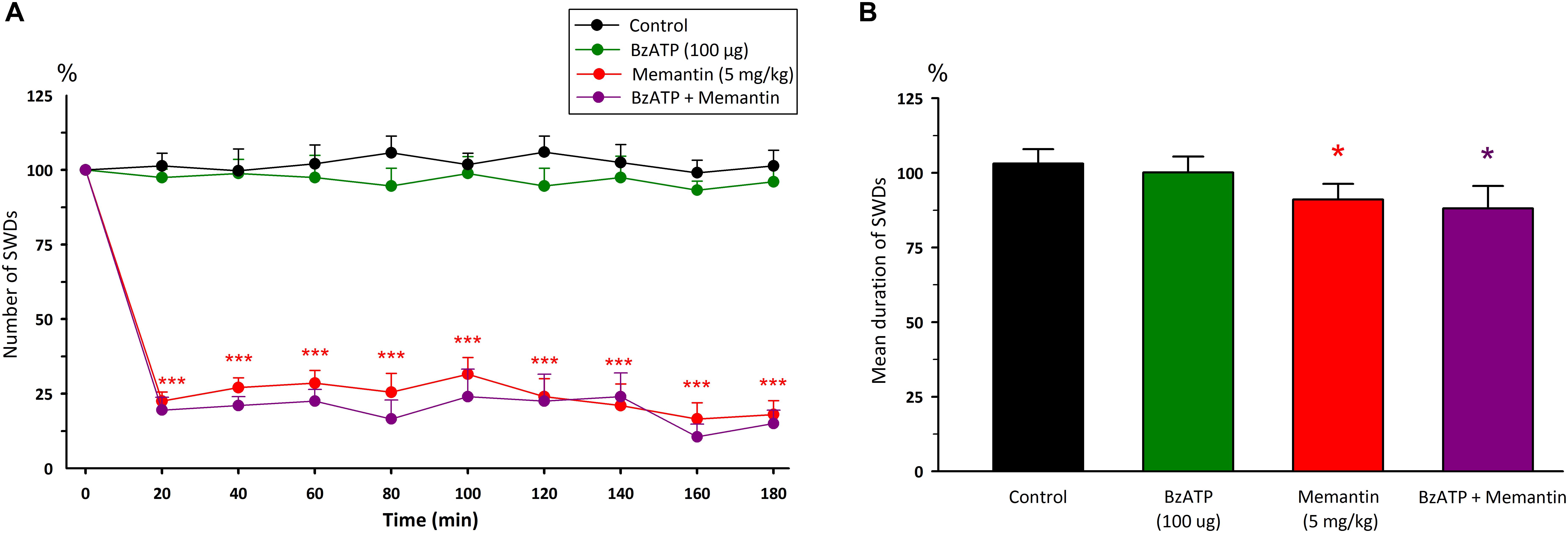
Figure 4. The effects of memantine at a dose of 5mg/kg and its interaction with BzATP on (A) the total number of SWDs in every 20 min and (B) the mean duration of SWDs for 180 min. *p < 0.05, ***p < 0.001 compared to control group.
Memantine significantly decreased MDA levels in all brain regions compared to the control group (p < 0.05). In the cerebrum, injection of memantine decreased GSH and GR levels, and increased CAT levels (p < 0.05). Other biochemical parameters were not different in the memantine group compared to the control group (Table 1).
Interaction Between P2X7R and Memantine in WAG/Rij Rats
Injection of BzATP (100 μg) 10 min before memantine decreased the total number of SWDs after 20 min [F(8,216) = 1.10, p < 0.001] and the mean duration of SWDs [F(3,24) = 10.60, p < 0.05] compared to the control group (Figure 4). However, the total number and the mean duration of SWDs were not found to be different after the co-administration of BzATP with memantine compared to the alone memantine injection. The total number and the mean duration of SWDs were 26.3 ± 2.8 and 7.56 ± 0.42 s for 180 min after the co-administration of BzATP with memantine, respectively.
Co-administration of BzATP with memantine decreased the levels of GSH and GR, and increased CAT levels in the cerebrum compared to the control group (p < 0.05). Other biochemical parameters did not alter with the co-administration of BzATP with memantine compared to the control group.
Although administration of A-438079 10 min prior to memantine decreased the total number of SWDs after 40 min compared to the control group (p < 0.05), but it appears that A-438079 partially reversed the anticonvulsant activity of memantine after 20th minute compared to the memantine group alone [F(8,216) = 1.97, p < 0.001]. The mean duration of SWDs did not alter compared to both the control and memantine groups [F(3,24) = 6.74, Figure 5]. The total number and mean duration of SWDs were 59.7 ± 6.6 and 8.05 ± 0.21 s for 180 min after the co-administration of A-438079 with memantine, respectively.
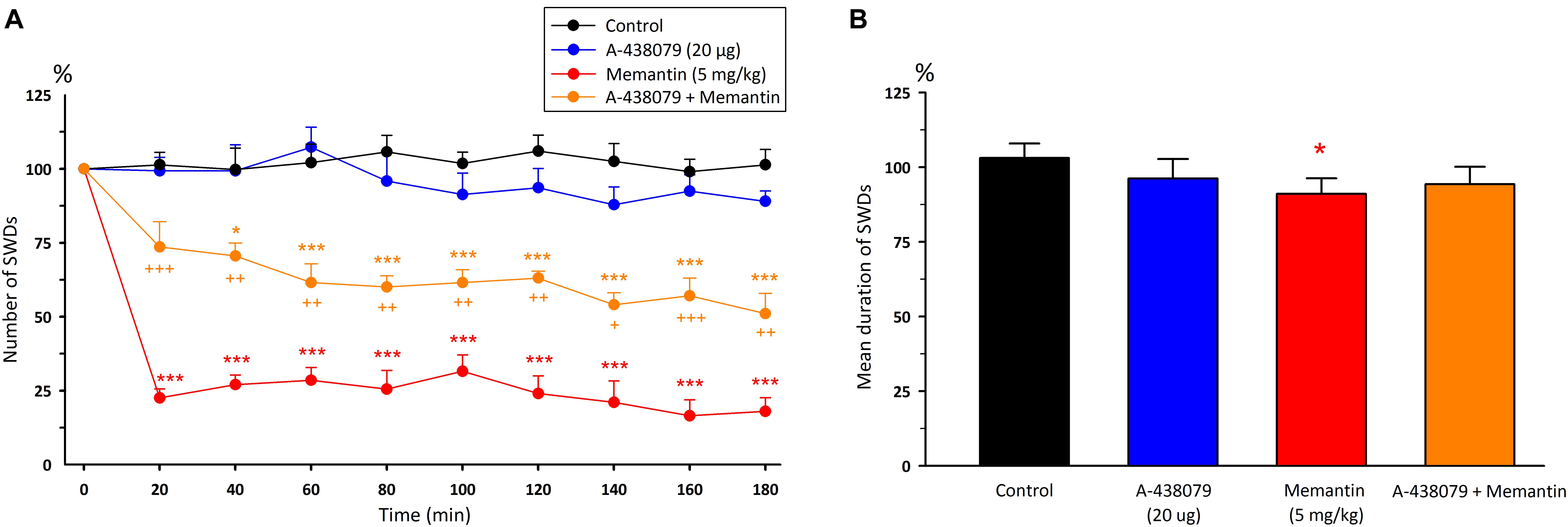
Figure 5. The effects of memantine at a dose of 5mg/kg and its interaction with A-438079 on (A) the total number of SWDs in every 20 min and (B) the mean duration of SWDs for 180 min. *p < 0.05, ***p < 0.001 compared to the control group. +p < 0.05, ++p < 0.01, +++p < 0.001 compared to the – memantine group.
Malondialdehyde levels significantly decreased in all regions after the co-administration of A-438079 with memantine (p < 0.05). In the cerebrum, the combination of A-438079 + memantine decreased GSH and GR levels, and increased CAT levels compared to the control group (p < 0.05). Interestingly, CAT levels were decreased in the brainstem with the injections of A-438079 and memantine (p < 0.05). The rest of the measured biochemical parameters were not changed in the A-438079 + memantine group compared to the control group (Table 1).
Discussion
In the present study, the electrophysiological results revealed that the administration of memantine showed an anticonvulsant effect without changing the mean amplitude of SWDs. Neither BzATP nor A-438079 affected the parameters of SWDs in WAG/Rij rats. BzATP did not reverse the anticonvulsant activity of memantine. However, A-438079 reduced the anticonvulsant activity of memantine.
BzATP increased only the SOD and GSH levels in the cerebrum, whereas A-438079 did not significantly affect any of the biochemical parameters. Memantine showed antioxidant effects by decreasing MDA levels in all tissue samples. Memantine also reduced GSH and GR and increased CAT levels in the cerebrum. BzATP reversed the antioxidant effects of memantine on MDA, while A-438079 enhanced the anticonvulsant effects of memantine in the brainstem.
The Role of P2X7R in WAG/Rij Rats
The P2X7R is the newest member of the purinergic receptor family and has been the subject of epilepsy research with its widespread presence in the nervous system (Jimenez-Mateos et al., 2019). The involvement of the P2X7R in epilepsy has been demonstrated by several researchers with either anticonvulsant, proconvulsant, or no effects in various models of epilepsy (Jimenez-Pacheco et al., 2016; Rodriguez-Alvarez et al., 2017; Arslan et al., 2019). These controversial results depend on experimental differences, such as the selection of the epilepsy model, the dosage of the P2X7R agonist and antagonist used, and the species of the experimental animals. We used an absence epilepsy model in this study, which was a chronic non-convulsive epilepsy model, and there are some unique mechanisms known to create absence seizures (Meeren et al., 2005; D’Antuono et al., 2006; Luttjohann and van Luijtelaar, 2012; Scicchitano et al., 2015). The P2X7R is a non-selective cation channel, and it allows calcium influx, which is essential for the absence epilepsy pathogenesis.
BzATP and A-438079 have high potency for the P2X7R. BzATP is the most potent for the P2X7R, but it is not a selective P2X7R agonist (Donnelly-Roberts and Jarvis, 2007). Thus, BzATP may also be effective in other P2X receptors that may also have an important role in epilepsy (Michel et al., 2001; Henshall et al., 2013). The potent inhibition of the P2X7R with A-438079 confirms that the P2X7R accounts for ATP-triggered Ca2+ entry (Fischer et al., 2016). The moderate bioavailability and moderate plasma elimination half-life were 19% and 1.02 h, respectively, for intraperitoneal A-438079 in rats (McGaraughty et al., 2007). Brain levels of A-438079 rapidly declined after parenteral injection, and Mesuret et al. (2014) suggested that the rapid elimination of the compound restricts the therapeutic window of this compound. However, in our previous study, i.c.v. injection of BzATP and A-438079 showed their effects on the penicillin-induced epileptiform activity within 20 and 60 min after their injections, respectively, and lasted for 120 min (Arslan et al., 2019). BzATP is the most potent agent for the P2X7R, and A-438079 is specific and can almost abolish the effects of BzATP (Anderson and Nedergaard, 2006; Fischer et al., 2016). The multi-targeting of different P2X receptors and their stability in the brain may be considered possible limitations for such studies in epilepsy. In addition, P2X7Rs release cytokines during normal brain function (Sperlagh and Illes, 2014). In particular, P2X7R activation might be linked to the regulation of various aspects of immunocompetent cells through the expression and secretion of many inflammatory mediators, including IL-1b, IL-2, IL-4, IL-6, IL-8, and TNFα (Engel et al., 2012b). However, inflammatory mediators were not measured, which might be another limitation of the present study. Therefore, further studies are required to determine both receptor-specific localization and the level of inflammatory mediators in an absence epilepsy model.
In an Alzheimer’s disease model, P2X7R activity affected nicotinamide adenine dinucleotide phosphate (NADPH) activity and increased the formation of the O.– radical by acting on p38 MAPK (Parvathenani et al., 2003). The antioxidant enzyme that converts the O.– radical into H2O2 is SOD, and an increase in the SOD level promotes the increase of O.– radicals (Younus, 2018). Activation of the P2X7R due to BzATP promotes ROS production through NADPH oxidase in macrophages, microglia, and neurons that can be blocked by P2X7R inhibitors (Hewinson and Mackenzie, 2007; Mead et al., 2012; Munoz et al., 2017). The P2X7R has also been shown to directly affect SOD in amyotrophic lateral sclerosis models and cell culture studies (Gandelman et al., 2013; Fabbrizio et al., 2017). GR is used in the reduction reaction of NADPH to GSH disulfide, which is a way to decrease NADPH. BzATP increases the production and release of ROS in the substantia gelatinosa of the spinal cord by stimulating P2X7Rs in astroglia (Ficker et al., 2014). However, Safiulina et al. (2006) showed ATP-induced ROS generation in CA3 pyramidal neurons due to the stimulation of P2Y1 receptors, not P2X7Rs. ATP treatment increased the expression of Cu/Zn SOD in the RBA-2 astrocyte cell line of cell culture (Chen et al., 2006). In our previous study, BzATP increased lipid peroxidation and the levels of protein oxidation and antioxidant proteins in the brain of penicillin-induced epileptic rats (Arslan et al., 2019). In the present study, BzATP and A-438079 did not affect protein or lipid oxidation, but BzATP increased SOD and GR in the cerebrum. Accordingly, the P2X7R seems to be more effective in the cerebral cortex in WAG/Rij rats with absence epilepsy. Since it has been suggested that oxidative mechanisms and inflammatory mechanisms do not cause the formation of absence epilepsy (Grosso et al., 2011) and there are no inflammatory processes in WAG/Rij rats, it is logical to expect that the low number of P2X7Rs and efficacy may be the reason for the ineffectiveness of BzATP and A-438079 observed in the present study in absence epileptic rats. However, children with absence epilepsy have shown no oxidant markers, while increased lipid peroxidation and protein oxidation levels have been observed in epileptic encephalopathic patients (Grosso et al., 2011).
The Role of NMDA Receptors in WAG/Rij Rats
Although memantine is mostly used in Alzheimer’s disease to improve cognitive function, the anticonvulsant activity of memantine has been demonstrated in various models of experimental epilepsy (Frey and Voits, 1991; Cakil et al., 2011; Kim et al., 2012; Zaitsev et al., 2015). Memantine showed an anticonvulsant effect both in a penicillin-induced epilepsy model and in Krushinsky–Molodkina rats with audiogenic seizures (Cakil et al., 2011; Kim et al., 2012). Memantine was also effective on the tonic component of seizures in a PTZ kindling model, and it prevented neuronal death (Zaitsev et al., 2015). Moreover, memantine showed anticonvulsant effects in WAG/Ola/Hsd rats, which are thought to be a model of genetic absence epilepsy (Frey and Voits, 1991), but there is no sufficient information about this rat substrain. In agreement with these studies, memantine has an anticonvulsant effect on absence epilepsy in WAG/Rij rats, which was used in the present study. Recent studies have shown that the initial focal point of absence epilepsy is in the perioral area of the somatosensory cortex and that the cortex is present in NMDA receptor-mediated glutamatergic pyramidal neurons and is present in layers V and VI (Miras-Portugal et al., 2003; Scicchitano et al., 2015; Russo et al., 2016). Thus, it can be concluded that seizures can be prevented without initially entering the thalamo-cortical circuit by decreasing the activity in this region in the presence of memantine.
As an NMDA receptor blocker, memantine has an antioxidant effect by blocking calcium entry, affecting intracellular signaling pathways (Flores et al., 2011; Annweiler and Beauchet, 2012; Gubandru et al., 2013; Lee et al., 2018). Memantine decreased the level of MDA, which is the final product of polyunsaturated fatty acid peroxidation, in all brain tissue samples and decreased GSH and GR levels but increased CAT levels in the cerebrum in this study.
Functional Interaction Between P2X7Rs and NMDA Receptors
Pre-treatment of organotypic hippocampal slice cultures with ATP reduced NMDA-induced neuronal death in microglia, suggesting microglia-mediated neuroprotection depends on P2X7Rs (Masuch et al., 2016). The glutamatergic agonists NMDA and AMPA increased the BzATP-induced current amplitudes in organotypic hippocampal slice cultures (Khan et al., 2018). Intravitreal injection of A-438079 and BBG significantly reduced NMDA-induced cell loss in the retinae of male Wistar rats, suggesting a strong link between P2X7R and NMDA (Sakamoto et al., 2015). Activation of the P2X7R has been shown to trigger the release of glutamate from neurons and astrocytic cells by vesicular and non-vesicular pathways (Sperlagh et al., 2002; Cho et al., 2010). Stimulation of NMDA receptors also increases ATP release (Cho et al., 2010; Engel et al., 2012a, b). In addition, P2X7Rs have been observed on presynaptic in glutamatergic pyramidal neurons (Sperlagh et al., 2002; Miras-Portugal et al., 2003), suggesting a P2X7-NMDA receptors interaction in pyramidal cells located in the perioral region of the somatosensory cortex, which is considered to be the initial focal point of absence epilepsy. In addition, both receptors are known to increase the intracellular calcium level. Thus, the interaction between P2X7 and NMDA receptors may increase excitability in the brain and may cause neurotoxicity by increasing ROS (Carrasco et al., 2018). In a phencyclidine-induced schizophrenia model, prefrontocortical postsynaptic NMDA currents slightly decreased due to both genetic deletion (P2X7R -/-) and pharmacological blockade with JNJ-47965567 (Kovanyi et al., 2016). However, NMDA currents were not affected in either wild type or P2X7R-deficient mice in in situ cortical astroglia (Oliveira et al., 2011). In contrast, electrophysiological recordings revealed that stimulation of the P2X7R with BzATP did not reverse memantine’s anticonvulsant effect; the anticonvulsant activity of memantine was maintained in the presence of BzATP, but it neutralized the level of MDA. The administration of A-438079 reduced the anticonvulsant activity of memantine, but the net effect remained anticonvulsive in this study. This finding might have been due to increased glutamate through BzATP activity, leading to cytotoxic effects through other glutamate receptors, such as AMPA and kainate (Coyle and Puttfarcken, 1993; Quincozes-Santos et al., 2014). There is evidence that some intracellular pathways affect the interaction of the two receptors. They both affect phosphokinase C (Ortega et al., 2010), pannexin-1 (Bravo et al., 2015), and the MAP kinase pathway (Gandelman et al., 2013; Fabbrizio et al., 2017; Lee et al., 2018). Therefore, it appears that this effect is due to the possible interaction of P2X7 and NMDA receptors in intracellular pathways. In addition, A-438079 and memantine decreased MDA levels in all tissues and changed antioxidant parameters similar to the memantine group in the present study, suggesting a strong interaction between the P2X7R and NMDA receptors in the brainstem.
Conclusion
Electrophysiological data from the present study suggest that P2X7Rs are ineffective for absence epilepsy whereas a biochemical analysis revealed a partial interaction between P2X7 and NMDA receptors in WAG/Rij rats with absence epilepsy. It seems logical to expect this interaction since P2X7R and the NMDA receptor both allow calcium influx. P2X7 and NMDA receptors use common intracellular signal pathways, but this interaction cannot be limited to calcium influx in epilepsy. Many other systems and receptors are involved in calcium influx without P2X7 or NMDA receptors and contribute to epileptogenesis. Besides, the P2X7R has been linked to inflammatory mediators in neurological diseases. Therefore, further studies are required to determine the level of inflammatory mediators and their localization in absence epilepsy.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
The animal study was reviewed and approved by Ondokuz Mayıs University Animal Experiments Local Ethics Committee (2015/56).
Author Contributions
EA planned and supervised all experiments. ED, HA, GA, and MA conducted the experiments. BA and ER provided biochemical measurements. ED, HA, GA, and BA analyzed the data. EA wrote the manuscript.
Funding
This work was supported by grant PYO.TIP.1905.15.002 from OMU BAP.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Akin, D., Ravizza, T., Maroso, M., Carcak, N., Eryigit, T., Vanzulli, I., et al. (2011). IL-1beta is induced in reactive astrocytes in the somatosensory cortex of rats with genetic absence epilepsy at the onset of spike-and-wave discharges, and contributes to their occurrence. Neurobiol. Dis. 44, 259–269. doi: 10.1016/j.nbd.2011.05.015
Albalawi, F., Lu, W., Beckel, J. M., Lim, J. C., McCaughey, S. A., and Mitchell, C. H. (2017). The P2X7 receptor primes IL-1beta and the NLRP3 inflammasome in astrocytes exposed to mechanical strain. Front. Cell. Neurosci. 11:227. doi: 10.3389/fncel.2017.00227
Amhaoul, H., Ali, I., Mola, M., Van Eetveldt, A., Szewczyk, K., Missault, S., et al. (2016). P2X7 receptor antagonism reduces the severity of spontaneous seizures in a chronic model of temporal lobe epilepsy. Neuropharmacology 105, 175–185. doi: 10.1016/j.neuropharm.2016.01.018
Anderson, C. M., and Nedergaard, M. (2006). Emerging challenges of assigning P2X7 receptor function and immunoreactivity in neurons. Trends Neurosci. 29, 257–262. doi: 10.1016/j.tins.2006.03.003
Annweiler, C., and Beauchet, O. (2012). Possibility of a new anti-alzheimer’s disease pharmaceutical composition combining memantine and vitamin D. Drugs Aging 29, 81–91. doi: 10.2165/11597550-000000000-00000
Arslan, G., Alici, S. K., Ayyildiz, M., and Agar, E. (2013). The role of CB1-receptors in the proconvulsant effect of leptin on penicillin-induced epileptiform activity in rats. CNS Neurosci. Ther. 19, 222–228. doi: 10.1111/cns.12075
Arslan, G., Avci, B., Kocacan, S. E., Rzayev, E., Ayyildiz, M., and Agar, E. (2019). The interaction between P2X7Rs and T-type calcium ion channels in penicillin-induced epileptiform activity. Neuropharmacology 149, 1–12. doi: 10.1016/j.neuropharm.2019.01.027
Arslan, G., Ayyildiz, M., and Agar, E. (2014). The interaction between ghrelin and cannabinoid systems in penicillin-induced epileptiform activity in rats. Neuropeptides 48, 345–352. doi: 10.1016/j.npep.2014.09.003
Aygun, H., Ayyildiz, M., and Agar, E. (2019). Effects of vitamin D and paricalcitol on epileptogenesis and behavioral properties of WAG/Rij rats with absence epilepsy. Epilepsy Res. 157:106208. doi: 10.1016/j.eplepsyres.2019.106208
Barros-Barbosa, A. R., Oliveira, A., Lobo, M. G., Cordeiro, J. M., and Correia-de-Sa, P. (2018). Under stressful conditions activation of the ionotropic P2X7 receptor differentially regulates GABA and glutamate release from nerve terminals of the rat cerebral cortex. Neurochem. Int. 112, 81–95. doi: 10.1016/j.neuint.2017.11.005
Beamer, E., Fischer, W., and Engel, T. (2017). The ATP-Gated P2X7 receptor as a target for the treatment of drug-resistant epilepsy. Front. Neurosci. 11:21. doi: 10.3389/fnins.2017.00021
Bravo, D., Maturana, C. J., Pelissier, T., Hernandez, A., and Constandil, L. (2015). Interactions of pannexin 1 with NMDA and P2X7 receptors in central nervous system pathologies: Possible role on chronic pain. Pharmacol. Res. 101, 86–93. doi: 10.1016/j.phrs.2015.07.016
Burnstock, G., and Knight, G. E. (2018). The potential of P2X7 receptors as a therapeutic target, including inflammation and tumour progression. Purinergic Signal. 14, 1–18. doi: 10.1007/s11302-017-9593-0
Cakil, D., Yildirim, M., Ayyildiz, M., and Agar, E. (2011). The effect of co-administration of the NMDA blocker with agonist and antagonist of CB1-receptor on penicillin-induced epileptiform activity in rats. Epilepsy Res. 93, 128–137. doi: 10.1016/j.eplepsyres.2010.11.008
Carrasco, C., Naziroglu, M., Rodriguez, A. B., and Pariente, J. A. (2018). Neuropathic pain: delving into the oxidative origin and the possible implication of transient receptor potential channels. Front. Physiol. 9:95. doi: 10.3389/fphys.2018.00095
Chen, H. B., Chan, Y. T., Hung, A. C., Tsai, Y. C., and Sun, S. H. (2006). Elucidation of ATP-stimulated stress protein expression of RBA-2 type-2 astrocytes: ATP potentiate HSP60 and Cu/Zn SOD expression and stimulates pI shift of peroxiredoxin II. J. Cell. Biochem. 97, 314–326. doi: 10.1002/jcb.20547
Cho, J. H., Choi, I. S., and Jang, I. S. (2010). P2X7 receptors enhance glutamate release in hippocampal hilar neurons. Neuroreport 21, 865–870. doi: 10.1097/wnr.0b013e32833d9142
Coyle, J. T., and Puttfarcken, P. (1993). Oxidative stress, glutamate, and neurodegenerative disorders. Science 262, 689–695. doi: 10.1126/science.7901908
D’Antuono, M., Inaba, Y., Biagini, G., D’Arcangelo, G., Tancredi, V., and Avoli, M. (2006). Synaptic hyperexcitability of deep layer neocortical cells in a genetic model of absence seizures. Genes Brain Behav. 5, 73–84. doi: 10.1111/j.1601-183x.2005.00146.x
Davidson, S. M., and Duchen, M. R. (2006). Calcium microdomains and oxidative stress. Cell Calcium 40, 561–574. doi: 10.1016/j.ceca.2006.08.017
Donnelly-Roberts, D. L., and Jarvis, M. F. (2007). Discovery of P2X7 receptor-selective antagonists offers new insights into P2X7 receptor function and indicates a role in chronic pain states. Br. J. Pharmacol. 151, 571–579. doi: 10.1038/sj.bjp.0707265
Droge, W. (2002). Free radicals in the physiological control of cell function. Physiol. Rev. 82, 47–95. doi: 10.1152/physrev.00018.2001
Engel, T., Gomez-Villafuertes, R., Tanaka, K., Mesuret, G., Sanz-Rodriguez, A., Garcia-Huerta, P., et al. (2012a). Seizure suppression and neuroprotection by targeting the purinergic P2X7 receptor during status epilepticus in mice. FASEB J. 26, 1616–1628. doi: 10.1096/fj.11-196089
Engel, T., Jimenez-Pacheco, A., Miras-Portugal, M. T., Diaz-Hernandez, M., and Henshall, D. C. (2012b). P2X7 receptor in epilepsy; role in pathophysiology and potential targeting for seizure control. Int. J. Physiol. Pathophysiol. Pharmacol. 4, 174–187.
Fabbrizio, P., Amadio, S., Apolloni, S., and Volonte, C. (2017). P2X7 receptor activation modulates autophagy in SOD1-G93A mouse microglia. Front. Cell. Neurosci. 11:249. doi: 10.3389/fncel.2017.00249
Ficker, C., Rozmer, K., Kato, E., Ando, R. D., Schumann, L., Krugel, U., et al. (2014). Astrocyte-neuron interaction in the substantia gelatinosa of the spinal cord dorsal horn via P2X7 receptor-mediated release of glutamate and reactive oxygen species. Glia 62, 1671–1686. doi: 10.1002/glia.22707
Fischer, W., Franke, H., Krugel, U., Muller, H., Dinkel, K., Lord, B., et al. (2016). Critical evaluation of P2X7 receptor antagonists in selected seizure models. PLoS One 11:e0156468. doi: 10.1371/journal.pone.0156468
Fisher, R. S., Cross, J. H., D’Souza, C., French, J. A., Haut, S. R., Higurashi, N., et al. (2017). Instruction manual for the ILAE 2017 operational classification of seizure types. Epilepsia 58, 531–542. doi: 10.1111/epi.13671
Flores, E. M., Cappelari, S. E., Pereira, P., and Picada, J. N. (2011). Effects of memantine, a non-competitive N-methyl-D-aspartate receptor antagonist, on genomic stability. Basic Clin. Pharmacol. Toxicol. 109, 413–417. doi: 10.1111/j.1742-7843.2011.00744.x
Frey, H. H., and Voits, M. (1991). Effect of psychotropic agents on a model of absence epilepsy in rats. Neuropharmacology 30, 651–656. doi: 10.1016/0028-3908(91)90086-q
Gandelman, M., Levy, M., Cassina, P., Barbeito, L., and Beckman, J. S. (2013). P2X7 receptor-induced death of motor neurons by a peroxynitrite/FAS-dependent pathway. J. Neurochem. 126, 382–388. doi: 10.1111/jnc.12286
Grosso, S., Longini, M., Rodriguez, A., Proietti, F., Piccini, B., Balestri, P., et al. (2011). Oxidative stress in children affected by epileptic encephalopathies. J. Neurol. Sci. 300, 103–106. doi: 10.1016/j.jns.2010.09.017
Gubandru, M., Margina, D., Tsitsimpikou, C., Goutzourelas, N., Tsarouhas, K., Ilie, M., et al. (2013). Alzheimer’s disease treated patients showed different patterns for oxidative stress and inflammation markers. Food Chem. Toxicol. 61, 209–214. doi: 10.1016/j.fct.2013.07.013
Henshall, D. C., Diaz-Hernandez, M., Miras-Portugal, M. T., and Engel, T. (2013). P2X receptors as targets for the treatment of status epilepticus. Front. Cell. Neurosci. 7:237. doi: 10.3389/fncel.2013.00237
Hewinson, J., and Mackenzie, A. B. (2007). P2X(7) receptor-mediated reactive oxygen and nitrogen species formation: from receptor to generators. Biochem. Soc. Trans. 35(Pt 5), 1168–1170. doi: 10.1042/bst0351168
Huang, C., Chi, X. S., Li, R., Hu, X., Xu, H. X., Li, J. M., et al. (2017). Inhibition of P2X7 receptor ameliorates nuclear factor-kappa b mediated neuroinflammation induced by status epilepticus in rat hippocampus. J. Mol. Neurosci. 63, 173–184. doi: 10.1007/s12031-017-0968-z
Illes, P., Khan, T. M., and Rubini, P. (2017). Neuronal P2X7 receptors revisited: do they really exist? J. Neurosci. 37, 7049–7062. doi: 10.1523/jneurosci.3103-16.2017
Illes, P., Verkhratsky, A., Burnstock, G., and Franke, H. (2012). P2X receptors and their roles in astroglia in the central and peripheral nervous system. Neuroscientist 18, 422–438. doi: 10.1177/1073858411418524
Jimenez-Mateos, E. M., Smith, J., Nicke, A., and Engel, T. (2019). Regulation of P2X7 receptor expression and function in the brain. Brain Res. Bull. 151, 153–163. doi: 10.1016/j.brainresbull.2018.12.008
Jimenez-Pacheco, A., Diaz-Hernandez, M., Arribas-Blazquez, M., Sanz-Rodriguez, A., Olivos-Ore, L. A., Artalejo, A. R., et al. (2016). Transient P2X7 receptor antagonism produces lasting reductions in Spontaneous Seizures and Gliosis in experimental temporal lobe epilepsy. J. Neurosci. 36, 5920–5932. doi: 10.1523/jneurosci.4009-15.2016
Jimenez-Pacheco, A., Mesuret, G., Sanz-Rodriguez, A., Tanaka, K., Mooney, C., Conroy, R., et al. (2013). Increased neocortical expression of the P2X7 receptor after status epilepticus and anticonvulsant effect of P2X7 receptor antagonist A-438079. Epilepsia 54, 1551–1561. doi: 10.1111/epi.12257
Kaczmarek-Hajek, K., Zhang, J., Kopp, R., Grosche, A., Rissiek, B., Saul, A., et al. (2018). Re-evaluation of neuronal P2X7 expression using novel mouse models and a P2X7-specific nanobody. eLife 7:e36217. doi: 10.7554/eLife.36217
Karimzadeh, F., Soleimani, M., Mehdizadeh, M., Jafarian, M., Mohamadpour, M., Kazemi, H., et al. (2013). Diminution of the NMDA receptor NR2B subunit in cortical and subcortical areas of WAG/Rij rats. Synapse 67, 839–846. doi: 10.1002/syn.21687
Khan, M. T., Liu, J., Nerlich, J., Tang, Y., Franke, H., and Illes, P. (2018). Regulation of P2X7 receptor function of neural progenitor cells in the hippocampal subgranular zone by neuronal activity in the dentate gyrus. Neuropharmacology 140, 139–149. doi: 10.1016/j.neuropharm.2018.08.001
Kim, J. E., and Kang, T. C. (2011). The P2X7 receptor-pannexin-1 complex decreases muscarinic acetylcholine receptor-mediated seizure susceptibility in mice. J. Clin. Invest. 121, 2037–2047. doi: 10.1172/jci44818
Kim, K., Zaitsev, A. V., Lavrent’eva, V. V., Zhabko, E. P., Vataev, S. I., Lukomskaia, N., et al. (2012). [The effect of ionotropic glutamate receptor antagonist on pentylenetetrazole-indused seizures in Krushinsky-Molodkina rats]. Ross. Fiziol. Zh. Im. I M Sechenova 98, 1520–1529.
Kovanyi, B., Csolle, C., Calovi, S., Hanuska, A., Kato, E., Koles, L., et al. (2016). The role of P2X7 receptors in a rodent PCP-induced schizophrenia model. Sci. Rep. 6:36680. doi: 10.1038/srep36680
Lee, K., Park, C., Oh, Y., Lee, H., and Cho, J. (2018). Antioxidant and Neuroprotective Effects of N-((3,4-Dihydro-2H-benzo[h]chromen-2-yl)methyl)-4-methoxyaniline in primary cultured rat cortical cells: involvement of ERK-CREB signaling. Molecules 23:669. doi: 10.3390/molecules23030669
Lowry, O. H., Rosebrough, N. J., Farr, W. L., and Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–272.
Luttjohann, A., and van Luijtelaar, G. (2012). The dynamics of cortico-thalamo-cortical interactions at the transition from pre-ictal to ictal LFPs in absence epilepsy. Neurobiol. Dis. 47, 49–60. doi: 10.1016/j.nbd.2012.03.023
Masuch, A., van der Pijl, R., Funer, L., Wolf, Y., Eggen, B., Boddeke, E., et al. (2016). Microglia replenished OHSC: a culture system to study in vivo like adult microglia. Glia 64, 1285–1297. doi: 10.1002/glia.23002
McGaraughty, S., Chu, K. L., Namovic, M. T., Donnelly-Roberts, D. L., Harris, R. R., Zhang, X. F., et al. (2007). P2X7-related modulation of pathological nociception in rats. Neuroscience 146, 1817–1828. doi: 10.1016/j.neuroscience.2007.03.035
Mead, E. L., Mosley, A., Eaton, S., Dobson, L., Heales, S. J., and Pocock, J. M. (2012). Microglial neurotransmitter receptors trigger superoxide production in microglia; consequences for microglial-neuronal interactions. J. Neurochem. 121, 287–301. doi: 10.1111/j.1471-4159.2012.07659.x
Meeren, H., van Luijtelaar, G., Lopes da Silva, F., and Coenen, A. (2005). Evolving concepts on the pathophysiology of absence seizures: the cortical focus theory. Arch. Neurol. 62, 371–376. doi: 10.1001/archneur.62.3.371
Mesuret, G., Engel, T., Hessel, E. V., Sanz-Rodriguez, A., Jimenez-Pacheco, A., Miras-Portugal, M. T., et al. (2014). P2X7 receptor inhibition interrupts the progression of seizures in immature rats and reduces hippocampal damage. CNS Neurosci. Ther. 20, 556–564. doi: 10.1111/cns.12272
Michel, A. D., Xing, M., and Humphrey, P. P. (2001). Serum constituents can affect 2′-& 3′-O-(4-benzoylbenzoyl)-ATP potency at P2X(7) receptors. Br. J. Pharmacol. 132, 1501–1508. doi: 10.1038/sj.bjp.0703968
Miras-Portugal, M. T., Diaz-Hernandez, M., Giraldez, L., Hervas, C., Gomez-Villafuertes, R., Sen, R. P., et al. (2003). P2X7 receptors in rat brain: presence in synaptic terminals and granule cells. Neurochem. Res. 28, 1597–1605.
Munoz, F. M., Gao, R., Tian, Y., Henstenburg, B. A., Barrett, J. E., and Hu, H. (2017). Neuronal P2X7 receptor-induced reactive oxygen species production contributes to nociceptive behavior in mice. Sci. Rep. 7:3539. doi: 10.1038/s41598-017-03813-7
Nieoczym, D., Socala, K., and Wlaz, P. (2017). Evaluation of the Anticonvulsant Effect of Brilliant Blue G, a Selective P2X7 Receptor Antagonist, in the iv PTZ-, Maximal Electroshock-, and 6 Hz-Induced Seizure Tests in Mice. Neurochem. Res. 42, 3114–3124. doi: 10.1007/s11064-017-2348-z
Oliveira, J. F., Riedel, T., Leichsenring, A., Heine, C., Franke, H., Krugel, U., et al. (2011). Rodent cortical astroglia express in situ functional P2X7 receptors sensing pathologically high ATP concentrations. Cereb. Cortex 21, 806–820. doi: 10.1093/cercor/bhq154
Ortega, F., Perez-Sen, R., Morente, V., Delicado, E. G., and Miras-Portugal, M. T. (2010). P2X7, NMDA and BDNF receptors converge on GSK3 phosphorylation and cooperate to promote survival in cerebellar granule neurons. Cell. Mol. Life Sci. 67, 1723–1733. doi: 10.1007/s00018-010-0278-x
Panayiotopoulos, C. P. (2001). Treatment of typical absence seizures and related epileptic syndromes. Paediatr. Drugs 3, 379–403. doi: 10.2165/00128072-200103050-00006
Parvathenani, L. K., Tertyshnikova, S., Greco, C. R., Roberts, S. B., Robertson, B., and Posmantur, R. (2003). P2X7 mediates superoxide production in primary microglia and is up-regulated in a transgenic mouse model of Alzheimer’s disease. J. Biol. Chem. 278, 13309–13317. doi: 10.1074/jbc.m209478200
Paxinos, G., and Watson, C. (2007). The Rat Brain in Stereotaxic Coordinates. San Diego, CA: Academic Press.
Peeters, B. W., Ramakers, G. M., Ellenbroek, B. A., Vossen, J. M., and Coenen, A. M. (1994). Interactions between NMDA and nonNMDA receptors in nonconvulsive epilepsy in the WAG/Rij inbred strain. Brain Res. Bull. 33, 715–718. doi: 10.1016/0361-9230(94)90237-2
Pinault, D., and O’Brien, T. J. (2005). Cellular and network mechanisms of genetically-determined absence seizures. Thalamus Relat. Syst. 3, 181–203. doi: 10.1017/S1472928807000209
Quincozes-Santos, A., Bobermin, L. D., Tramontina, A. C., Wartchow, K. M., Tagliari, B., Souza, D. O., et al. (2014). Oxidative stress mediated by NMDA, AMPA/KA channels in acute hippocampal slices: neuroprotective effect of resveratrol. Toxicol. In Vitro 28, 544–551. doi: 10.1016/j.tiv.2013.12.021
Rodriguez-Alvarez, N., Jimenez-Mateos, E. M., Engel, T., Quinlan, S., Reschke, C. R., Conroy, R. M., et al. (2017). Effects of P2X7 receptor antagonists on hypoxia-induced neonatal seizures in mice. Neuropharmacology 116, 351–363. doi: 10.1016/j.neuropharm.2017.01.005
Rosini, M., Simoni, E., Caporaso, R., Basagni, F., Catanzaro, M., Abu, I. F., et al. (2019). Merging memantine and ferulic acid to probe connections between NMDA receptors, oxidative stress and amyloid-beta peptide in Alzheimer’s disease. Eur. J. Med. Chem. 180, 111–120. doi: 10.1016/j.ejmech.2019.07.011
Rozmer, K., Gao, P., Araujo, M. G. L., Khan, M. T., Liu, J., Rong, W., et al. (2017). Pilocarpine-Induced Status Epilepticus Increases the Sensitivity of P2X7 and P2Y1 Receptors to Nucleotides at Neural Progenitor Cells of the Juvenile Rodent Hippocampus. Cereb. Cortex 27, 3568–3585. doi: 10.1093/cercor/bhw178
Russo, E., Citraro, R., Constanti, A., Leo, A., Luttjohann, A., van Luijtelaar, G., et al. (2016). Upholding WAG/Rij rats as a model of absence epileptogenesis: hidden mechanisms and a new theory on seizure development. Neurosci. Biobehav. Rev. 71, 388–408. doi: 10.1016/j.neubiorev.2016.09.017
Safiulina, V. F., Afzalov, R., Khiroug, L., Cherubini, E., and Giniatullin, R. (2006). Reactive oxygen species mediate the potentiating effects of ATP on GABAergic synaptic transmission in the immature hippocampus. J. Biol. Chem. 281, 23464–23470. doi: 10.1074/jbc.m601627200
Sakamoto, K., Endo, K., Suzuki, T., Fujimura, K., Kurauchi, Y., Mori, A., et al. (2015). P2X7 receptor antagonists protect against N-methyl-D-aspartic acid-induced neuronal injury in the rat retina. Eur. J. Pharmacol. 756, 52–58. doi: 10.1016/j.ejphar.2015.03.008
Scicchitano, F., van Rijn, C. M., and van Luijtelaar, G. (2015). Unilateral and bilateral cortical resection: effects on spike-wave discharges in a genetic absence epilepsy model. PLoS One 10:e0133594. doi: 10.1371/journal.pone.0133594
Soni, N., Koushal, P., Reddy, B. V., Deshmukh, R., and Kumar, P. (2015). Effect of GLT-1 modulator and P2X7 antagonists alone and in combination in the kindling model of epilepsy in rats. Epilepsy Behav. 48, 4–14. doi: 10.1016/j.yebeh.2015.04.056
Sperlagh, B., and Illes, P. (2014). P2X7 receptor: an emerging target in central nervous system diseases. Trends Pharmacol. Sci. 35, 537–547. doi: 10.1016/j.tips.2014.08.002
Sperlagh, B., Kofalvi, A., Deuchars, J., Atkinson, L., Milligan, C. J., Buckley, N. J., et al. (2002). Involvement of P2X7 receptors in the regulation of neurotransmitter release in the rat hippocampus. J. Neurochem. 81, 1196–1211. doi: 10.1046/j.1471-4159.2002.00920.x
Takenouchi, T., Sekiyama, K., Sekigawa, A., Fujita, M., Waragai, M., Sugama, S., et al. (2010). P2X7 receptor signaling pathway as a therapeutic target for neurodegenerative diseases. Arch. Immunol. Ther. Exp. 58, 91–96. doi: 10.1007/s00005-010-0069-y
Terrone, G., Frigerio, F., Balosso, S., Ravizza, T., and Vezzani, A. (2019). Inflammation and reactive oxygen species in status epilepticus: Biomarkers and implications for therapy. Epilepsy Behav. 101(Pt B), 106275. doi: 10.1016/j.yebeh.2019.04.028
van de Bovenkamp-Janssen, M. C., van der Kloet, J. C., van Luijtelaar, G., and Roubos, E. W. (2006). NMDA-NR1 and AMPA-GluR4 receptor subunit immunoreactivities in the absence epileptic WAG/Rij rat. Epilepsy Res. 69, 119–128. doi: 10.1016/j.eplepsyres.2006.01.003
Zaitsev, A. V., Kim, K., Vasilev, D. S., Lukomskaya, N. Y., Lavrentyeva, V. V., Tumanova, N. L., et al. (2015). N-methyl-D-aspartate receptor channel blockers prevent pentylenetetrazole-induced convulsions and morphological changes in rat brain neurons. J. Neurosci. Res. 93, 454–465. doi: 10.1002/jnr.23500
Keywords: absence, epilepsy, memantine, NMDA, oxidative, P2X7, stress, WAG/Rij
Citation: Doǧan E, Aygün H, Arslan G, Rzayev E, Avcı B, Ayyıldız M and Ağar E (2020) The Role of NMDA Receptors in the Effect of Purinergic P2X7 Receptor on Spontaneous Seizure Activity in WAG/Rij Rats With Genetic Absence Epilepsy. Front. Neurosci. 14:414. doi: 10.3389/fnins.2020.00414
Received: 10 January 2020; Accepted: 06 April 2020;
Published: 06 May 2020.
Edited by:
Mustafa Naziroglu, Süleyman Demirel University, TurkeyReviewed by:
Katarzyna Socała, Marie Curie-Skłodowska University, PolandTobias Engel, Royal College of Surgeons in Ireland, Ireland
Copyright © 2020 Doǧan, Aygün, Arslan, Rzayev, Avcı, Ayyıldız and Ağar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elif Doǧan, ZWxpZnNlbkBvbXUuZWR1LnRy; Erdal Ağar, ZXJhZ2FyQG9tdS5lZHUudHI=