 Filip Černý
Filip Černý Václava Piorecká
Václava Piorecká Monika Kliková2
Monika Kliková2 Jana Kopřivová
Jana Kopřivová Jitka Bušková
Jitka Bušková Marek Piorecký
Marek Piorecký- 1Faculty of Biomedical Engineering, Czech Technical University, Prague, Czechia
- 2Sleep and Chronobiology Research Center, National Institute of Mental Health, Klecany, Czechia
- 3Third Faculty of Medicine, Charles University, Prague, Czechia
The pathophysiology of recurrent isolated sleep paralysis (RISP) has yet to be fully clarified. Very little research has been performed on electroencephalographic (EEG) signatures outside RISP episodes. This study aimed to investigate whether sleep is disturbed even without the occurrence of a RISP episode and in a stage different than conventional REM sleep. 17 RISP patients and 17 control subjects underwent two consecutive full-night video-polysomnography recordings. Spectral analysis was performed on all sleep stages in the delta, theta, and alpha band. EEG microstate (MS) analysis was performed on the NREM 3 phase due to the overall high correlation of subject template maps with canonical templates. Spectral analysis showed a significantly higher power of theta band activity in REM and NREM 2 sleep stages in RISP patients. The observed rise was also apparent in other sleep stages. Conversely, alpha power showed a downward trend in RISP patients' deep sleep. MS maps similar to canonical topographies were obtained indicating the preservation of prototypical EEG generators in RISP patients. RISP patients showed significant differences in the temporal dynamics of MS, expressed by different transitions between MS C and D and between MS A and B. Both spectral analysis and MS characteristics showed abnormalities in the sleep of non-episodic RISP subjects. Our findings suggest that in order to understand the neurobiological background of RISP, there is a need to extend the analyzes beyond REM-related processes and highlight the value of EEG microstate dynamics as promising functional biomarkers of RISP.
Significance statement
We focused on tracking electrophysiological traces of RISP (a REM parasomnia) beyond REM sleep of subjects clinically diagnosed with RISP outside of RISP episodes. We observed a rise of theta band activity in NREM 2 sleep of RISP patients. This may point to a larger dysregulation of sleep mechanism making the person more prone to sudden awakenings in the upcoming REM sleep. Theta band differences were further observed in REM sleep. We additionally utilized the EEG MS methodology on deep sleep to investigate differences in dominant brain topographies. Though dominant brain topographies are consistent with canonical MS, RISP patients show significantly different transitioning between sleep-related topographies suggesting a difference in their sleep regulation mechanisms.
Introduction
Recurrent isolated sleep paralysis (RISP) is a sleep disorder whose mechanism has not yet been fully elucidated due to its random occurrence and difficulty to capture in laboratory conditions (Walther and Schulz, 2004). It manifests by repeated episodes with an inability to perform voluntary movements at sleep onset or on a sudden awakening from sleep. It is often accompanied by hallucinatory experiences, e.g., feeling someone's presence, or feeling pressure against the chest (Stefani and Högl, 2021). Although RISP is not physically harmful, it can leave the victim with anxiety and stress (Denis et al., 2018). Research shows that the experience of an isolated sleep paralysis episode is about 8% (Sateia, 2014) in the general population. However, it was shown to be higher in psychiatric patients (Sharpless and Barber, 2011). A frequent occurrence in the population (Sharpless and Barber, 2011; Stefani and Högl, 2021) and the association with clinically significant distress make RISP an important research topic (Sharpless, 2016). However, very few studies have addressed the electrophysiological correlates of RISP episodes or even examined brain activity outside of them.
The RISP episodes have primarily been described as dissociative states of consciousness, combining a mixture of waking and REM sleep brain states with abundant alpha activity in electroencephalography (EEG) and persistence of muscle atonia (Takeuchi et al., 1992; Terzaghi et al., 2011). Although recently, a predominant theta has been detected during the actual episodes using spectral analysis (Mainieri et al., 2020), which may indicate that the brain is more likely to be in dreaming than waking state. Terzaghi et al. (2011) concluded a case study on a narcolepsy patient, suggesting that sleep paralysis represents a transition state between REM sleep and wakefulness. A study by Takeuchi et al. (1992) shows a possible higher paralysis probability in sudden interruptions of REM sleep.
Patients suffering from sleep apnea also seem to be more prone to sleep paralysis (Anderson et al., 2007). This may have been due to higher REM sleep fragmentation as apneic breaks occur during REM sleep 36% of the time (Mokhlesi and Punjabi, 2012). However, higher REM sleep fragmentation was not present in our previous study of RISP patients (Klikovà et al., 2021) in REM sleep along with differences in other macrostructural sleep parameters. Nevertheless, we observed differences in EEG spectral bands between RISP patients and control subjects. Patients showed higher bifrontal beta activity in the patients' group in REM sleep. Persistent higher cortical activity may represent a predisposing factor for sleep paralysis (Klikovà et al., 2021).
On the other hand, sleep paralysis may not be a matter of REM sleep alone and may reflect global sleep dysregulation. Its incidence is known to increase with elevated homeostatic pressure, i.e., due to sleep deprivation, irregular sleep-wake schedules, and jet lag. Avoiding these predisposing factors is also the most effective therapy for RISP patients (Buškovà and Klikovà, 2019). RISP also often occurs as a symptom of narcolepsy, a disease characterized by disruption of the circadian sleep-wake rhythm (Denis et al., 2015) or interruption of the REM-NREM cycle that may favor sleep paralysis. Changes in EEG NREM sleep oscillations were also found during nightmares (Marquis et al., 2017; Blaskovich et al., 2020), which may be a decisive predictive factor for the frequency of RISP episodes (Liškovà et al., 2016).
Our aim was to determine whether differences in the global functional state of the sleeping brain may play a role in the pathophysiology of RISP regardless of the presence of episodes or even sleep stages and to describe their dynamics. In contrast to our previous work on this dataset (Klikovà et al., 2021), focusing on the REM stage, here we asked whether this REM sleep parasomnia also shows differences in NREM sleep which may suggest a broader, more complex dysregulation of sleep than previously thought.
Materials and methods
Experimental design
All study participants were invited through advertisements on the website of the National Institute of Mental Health website, Klecany, and social media networks. Subjects with a history of sleep paralysis episodes (N = 112) underwent a clinical interview that served as a diagnostic tool to verify that participants fulfilled the criteria for RISP (Sateia, 2014). A total of 20 selected participants with RISP underwent two consecutive full-night video-polysomnography (vPSG) with consecutive multiple sleep latency tests (MSLT) to exclude narcolepsy. Inclusion criteria contained fulfillment of RISP standards defined in the International Classification of Sleep Disorders 3 (ICSD-3; Sateia, 2014) and experience of 2 sleep paralysis (SP) episodes in the past 6 months as recommended by Sharpless (2016). Exclusion criteria contained a diagnosis of other sleep disorders, clinically significant neurological, psychiatric, or somatic disorders that could lead to sleep alteration, and medication affecting sleep (e.g., antidepressants and anxiolytics). The control group comprised 20 healthy age-matched volunteers with no history of SP episodes; who did not experience neurological, psychiatric, or sleep disorders; and who did not receive any psychotropic medication. Before analysis, we excluded one participant from the RISP group due to consuming a benzodiazepine drug before the experimental night and one participant from the control group due to technical issues. Furthermore, we excluded two participants from each group due to uncorrelated MS topographical maps with templates (see Microstate validation).
In total, 17 participants in the RISP group (N = 15 women, two men; mean age = 25.24 years, SD = 7.0) and 17 healthy participants in the control group (N = 13 women, four men; mean age = 25.17, SD = 6.20) were included. The frequencies of SP episodes were several times a year (N = 5), once a month (N = 5), and several times a month (N = 7). Six months before the PSG recording, participants experienced a mean of 10.07 SP episodes (SD 8.04) ranging from 2 to 30 episodes (one outlier stated experience of 70 episodes). There was no between-group difference in gender and age.
The first night of vPSG served as an adaptation night and for the exclusion of other sleep disorders. Therefore, only data from the second night (experimental) were analyzed. VPSG was performed using a digital Brainscope PSG system (M&I spol. s.r.o., Czech Republic). It consisted of 19-channel EEG, electrooculography, electromyography of the bilateral mentalis muscle, the bilateral flexor digitorum superficialis muscle, and the bilateral tibialis anterior muscle, electrocardiography, airflow, thoracic and abdominal respiratory effort, oxygen saturation, microphone and synchronized video monitoring from 10:00 pm to 06:00 am, according to the recommendation of the American Academy of Sleep Medicine (AASM; Sateia, 2014). MSLT measurements were taken in agreement with standard protocol at 09:00 am, 11:00 am, 01:00 pm, 03:00 pm, and 05:00 pm.
After adaptation night, participants were allowed to leave the sleep laboratory in the morning and return at 06:00 pm. They were encouraged to follow their usual daily activities and sleep hygiene rules.
The study was conducted in accordance with the Declaration of Helsinki. All participants provided written informed consent. The study protocol was approved by the local Ethical Committee of the National Institute of Mental Health, Klecany, Czech Republic, and by the Ethical Committee of the Third Faculty of Medicine, Charles University in Prague, Czech Republic. A financial reward of 1,500 CZK (~78$) was provided for participation in polysomnographic recordings.
Data preprocessing
The vPSG was performed using a Brainscope PSG system (M&I spol. s.r.o., Czech Republic). EEG data were recorded on a band-pass filter of 0.1–200.0 Hz and with a 1 kHz sampling rate. For digital signal processing data were downsampled to 250 Hz. Slow drifts of the signal, as well as line noise and other high-frequency noises, were filtered out by a 0.5–30.0 Hz two-way zero-phase bandpass FIR filter with an order of 1000 in the context of spectral analysis and 1–40 Hz in the context of microstate analysis and literature evaluation (Britz et al., 2010; Brodbeck et al., 2012; Fèrat et al., 2020). Each polysomnographic recording was scored for ongoing sleep phases by a somnologist, blinded to the RISP vs. control condition. Each epoch was assigned into groups corresponding to the ongoing sleep phase: NREM 1, NREM 2, NREM 3, REM, and WAKE based on the sleep scoring results. Epochs containing arousals were removed from analysis if the arousal period represented more than 15% of the scored epoch.
EEG spectral analysis
For spectral analysis, EEG signals were evaluated in a montage to the mastoid electrodes (M1, M2). Data were filtered with a two-way zero-phase bandpass type FIR filter in the 0.5–30.0 Hz frequency range.
EEG spectra were evaluated using power spectral density analysis. Power spectra were estimated by frequency analysis using conventional multiple windows based on discrete prolate spheroidal sequences or Slepian sequences (DPSS; Slepian, 1978; Oostenveld et al., 2011). Selected frequency bands of interest included delta (0.5–4.0 Hz), theta (4.0–8.0 Hz), and alpha (8.0–12.0 Hz) EEG bands, which cover the majority of sleep-related activity. Beta band was not analyzed due to previously shown indifferences in RISP vs. healthy sleep (Mainieri et al., 2020). Power spectral densities were further converted to relative power spectra. The normalization of spectra was performed by dividing individual spectral band's powers by the total spectral power of a subject. Relative spectrum therefore represents a percentage of power in a frequency band and enables precise statistical comparison of activity between bands (Inouye et al., 1991).
Results from EEG spectral analysis were statistically evaluated on two levels. For the normalized EEG power spectra, we performed permutation tests of the FieldTrip toolbox (Monte-Carlo estimates of the probability of significance and critical values) with cluster correction (Maris and Oostenveld, 2007; Oostenveld et al., 2011). A T-value was computed for every channel, clustered based on spatial adjacency, and compared between experimental conditions to evaluate its effect. Secondly, false discovery rate (FDR) correction was used to correct for multiple comparisons of spectral analysis results between different spectral ranges and sleep phases. Thus we performed corrections for multiple channels, spectra, and sleep phases.
EEG microstates analysis
As a complementary approach to spectral analysis, we performed EEG microstates (MS) analysis. EEG MS represent short periods of quasi-stability of electric field configurations that reflect a reference-free global functional state of the brain. They are thought to arise due to the coordinated activity of neuronal assemblies originating from large cortical areas and have distinct scalp topographies replicated by numerous studies (Michel and Koenig, 2018). Temporal dynamics of EEG MS are altered in various states of consciousness (Brodbeck et al., 2012). If the brain activity of RISP patients differs during sleep, even outside the paralysis episode, microstates could reflect these differences. Even though RISP is mainly associated with the REM phase, previous studies analyzed the REM and NREM together (De Gennaro et al., 2000; Bettica et al., 2012; Mantua et al., 2016). Furthermore, NREM 3 sleep phase has been associated with REM phase analysis in depression research (Held et al., 2004) and in a sleep quality study (Dongrui et al., 2020).
We performed the analysis only for the NREM 3 stage based on template maps correlations (see Microstate validation chapter). The average electrode reference was used, and data were bandpass filtered in a range of 1–40 Hz (Britz et al., 2010; Brodbeck et al., 2012; Fèrat et al., 2020). Demean and detrend filters were used as well. The FieldTrip toolbox (Oostenveld et al., 2011) was used for preprocessing purposes. The final control's group mean NREM 3 record duration was 94.90 min (SD 21.77), totaling 1,613.22 min. RISP patients' group mean NREM 3 record duration was 88.36 min (SD 23.11), totaling 1,502.20 min.
The modified k-means algorithm was used to cluster the resulting topographical maps in peaks of the global field power (GFP) curve (Pascual-Marqui et al., 1995). This algorithm was restarted 20 times. The polarity of EEG time series data was ignored due to the analysis of spontaneous EEG data segments (Pascual-Marqui et al., 1995). Standard EEG microstate parameters (duration, occurrence, coverage, GFP, and transitions; Tarailis et al., 2021) were evaluated for each microstate class.
Temporal parameters that describe EEG MS maps were statistically evaluated between groups. The non-parametric Wilcoxon rank sum test for two populations, where samples are independent, was used. The resulting p-values were corrected for multiple comparisons with Bonferroni's correction. The correction coefficient equaled 16 for temporal parameters of occurrence, duration, contribution, and GFP (4 MS x 4 parameters). Transition parameters were corrected by the coefficient equal to 16 following both directions of transitions without autocorrelations (Krylova et al., 2021).
Microstate validation
To further support the EEG microstate approach, we tested the degree to which the individual's activity corresponded to the created microstate templates. In other words, we verified whether stable results were obtained for all MS topographies. Individual subject topographies were therefore compared with group template maps (RISP group and control maps separately) and with Grand mean (GM) global maps of the whole dataset. To do this, we used spatial correlation analysis and visual comparison. The spatial correlation was computed using the global dissimilarity index (GDI; Lehmann and Skrandies, 1980), defined as:
where u and v represent the measured voltage under electrodes, i represents individual electrode and n represents the total number of electrodes. This parameter corresponds to the spatial Pearson's product-moment correlation coefficient (Brodbeck et al., 2012).
Statistical analysis
Considering that there are no studies that present robust sleep EEG datasets of RISP patients, we performed power analysis on other sleep disorders.
Narcolepsy is a sleep disorder with a wide range of symptoms, and its patients may experience sleep-wake cycle disorders similar to those in RISP. In a study concentrating on narcolepsy patients, a sample size of 21 narcoleptic individuals was selected to ensure sufficient statistical power (Drissi et al., 2016).
Insomnia also has various spectral characteristics across different EEG bands described. We hypothesized to observe a difference in the theta band, which is typical for the REM phase. We evaluated the relative spectral performances for four sleep stages (REM, NREM 1, NREM 2, and NREM 3) and three EEG bands (delta, theta, and alpha), due to relative spectral power being a more sensitive marker for NREM sleep (Zhao et al., 2021). Studies included in the review of Zhao et al. (2021) use various ranges of spectral bands, therefore the calculation of the sample size was done through the combined mean values and standard deviations. Effect sizes were determined based on Hedges'g (Çooğaltay and Karadağ, 2015). Power analysis estimates for relative spectra were transitioned for MS analysis, as the area of interest was based on spectral analysis results. This review suggests that a group of 30 insomniacs and 20 healthy controls to be necessary to identify a difference in the REM band for relative theta spectral power.
As RISP is not as widespread as narcolepsy or insomnia it is challenging to select age- and gender-matched patients who only suffer from this disease without other neuropsychiatric disorders. We, therefore, chose a dataset size of 20 controls and 20 patients. Due to technical and data acquisition issues, we excluded a few subjects. In total, 17 patients and 17 controls entered the analysis.
Code accessibility
All processing scripts are available on: https://github.com/vaclavapiorecka/sleepo.
Results
Macrostructural vPSG sleep parameters
During the experimental nights, no episodes corresponding to RISP diagnosis were detected. MSLT results excluded narcolepsy in all RISP participants. Table 1 shows the mean values of macrostructural sleep parameters.
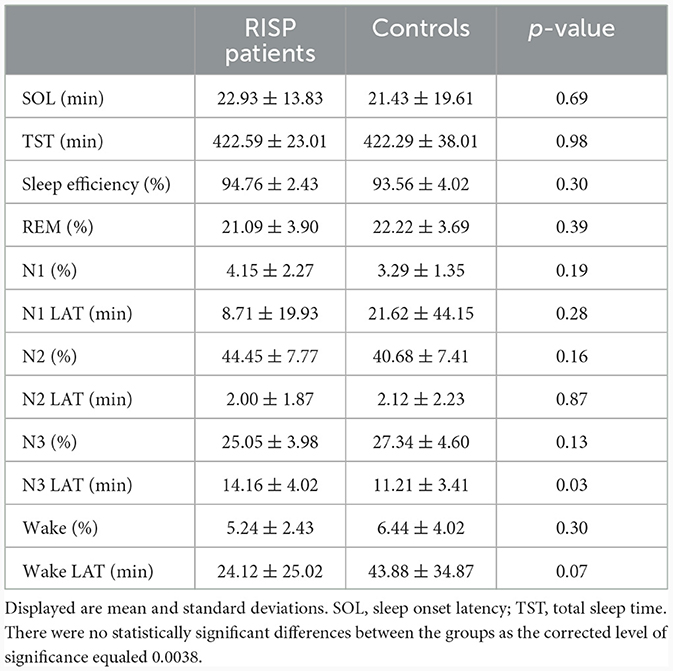
Table 1. Macrostructural sleep parameters of RISP (n = 17) and control group (n = 17).
EEG spectral analysis
Power values of EEG trials were computed for three defined spectral bands of interest. Spectral power was calculated for trials in each sleep stage individually. Figure 1 shows a topographically mapped difference in spectral power between the RISP group and controls. There were no statistically significant differences when using only the Bonferroni correction (19 electrodes x four sleep stages x three spectral bands); this is, however, a very strict correction. Cluster-based and FDR-corrected results showed significantly higher activity in the theta band for the NREM 2 and REM phases. The Bonferroni correction was used only for sleep stages and frequency band group corrections. A rise in theta band activity was apparent across the whole scalp for the NREM 2 phase.
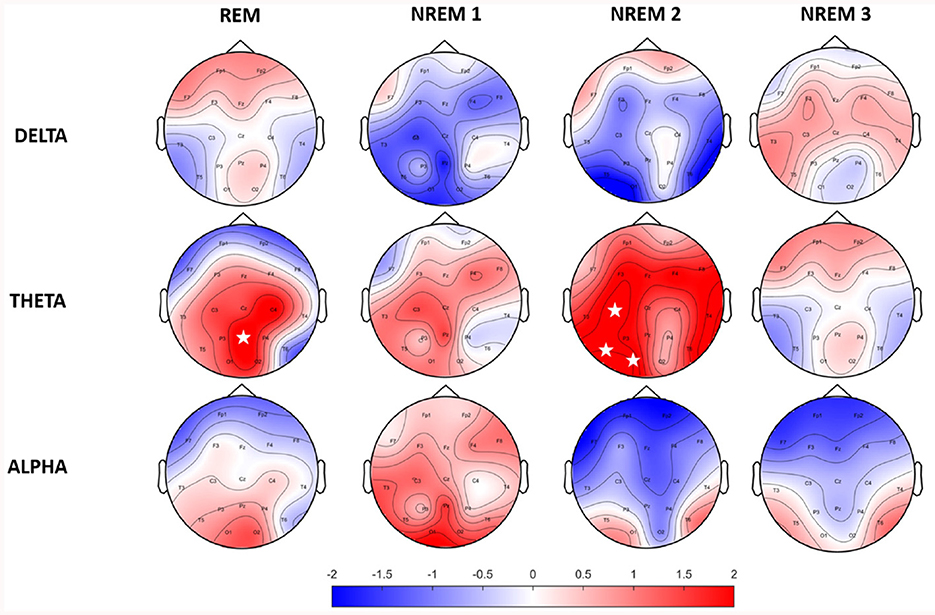
Figure 1. Topographic maps of relative spectral power differences between RISP patients group and healthy controls. Positive values represent higher activity in the RISP patients group. White asterisks mark electrodes with statistically significant differences after FDR correction.
EEG microstate analysis
Identified MS maps represented 80.29 ± 1.56 % GEV for the patients' group and 79.00 ± 1.85 % for the control group in the case of four template maps. Based on literature (Koenig et al., 2002), GEV, and visual inspection of MS templates, this number of maps was chosen as sufficient for analysis. Therefore, this number of template maps was extracted for both groups (controls and RISP patients). The topographies of both groups of microstates and GM topographies are depicted in Figure 2 and compared to canonical MS maps of Koenig et al. (2002) in Figure 3.
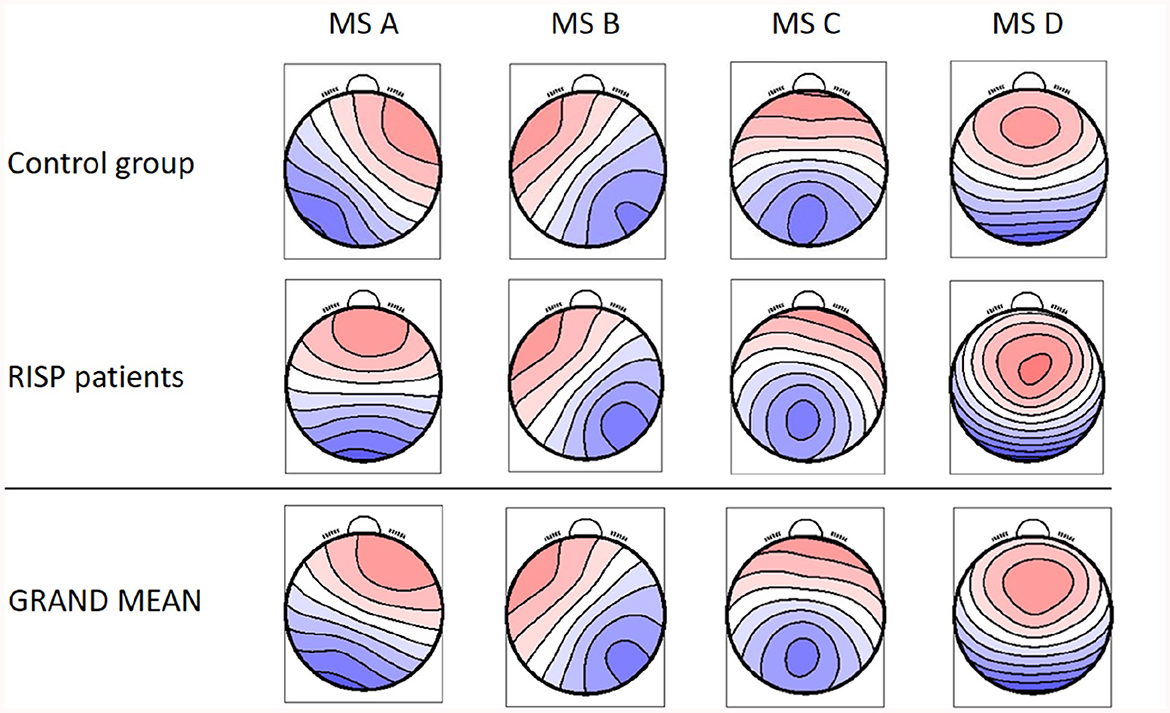
Figure 2. Resulting four MS topographical maps for individual groups and the GM MS maps for the whole dataset. Created with EEG microstates plugin for EEGlab.
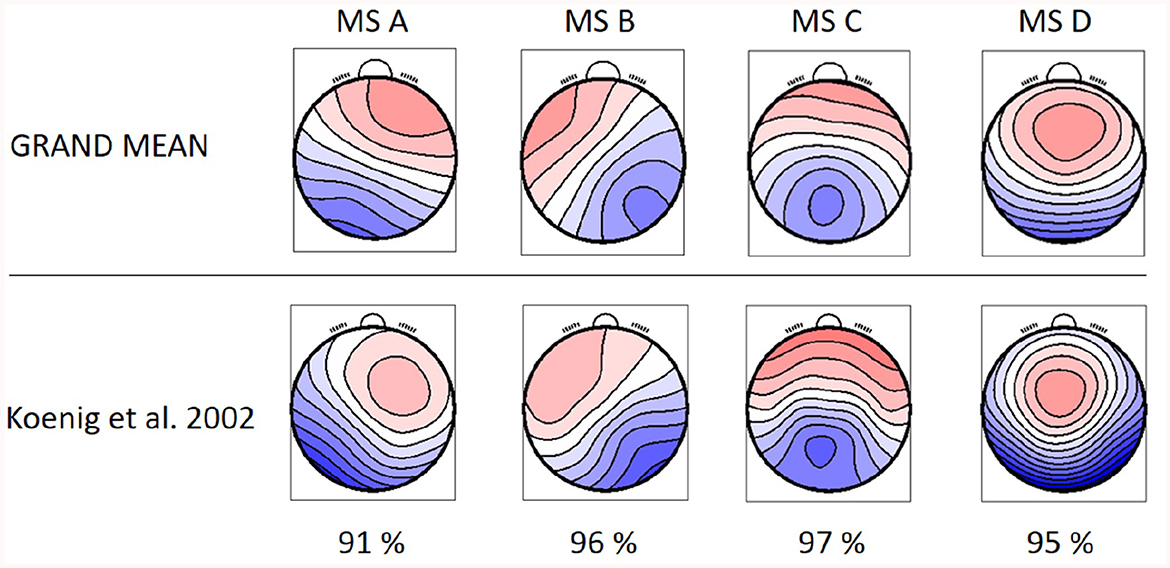
Figure 3. GM topographical maps of RISP and healthy controls in comparison to the topographical maps extracted by Koenig et al. (2002). The percentage of spatial correlation for each microstate topography is shown below.
The resulting MS, derived as an average topography of both groups, have similar topographies to those standardized in the literature (A–D; Koenig et al., 2002; Lehmann et al., 2005; Kikuchi et al., 2011; Kindler et al., 2011) and thus were labeled accordingly. The spatial correlation of the GM maps with the topographies of individual groups (Equation 1) is presented in Figure 4.
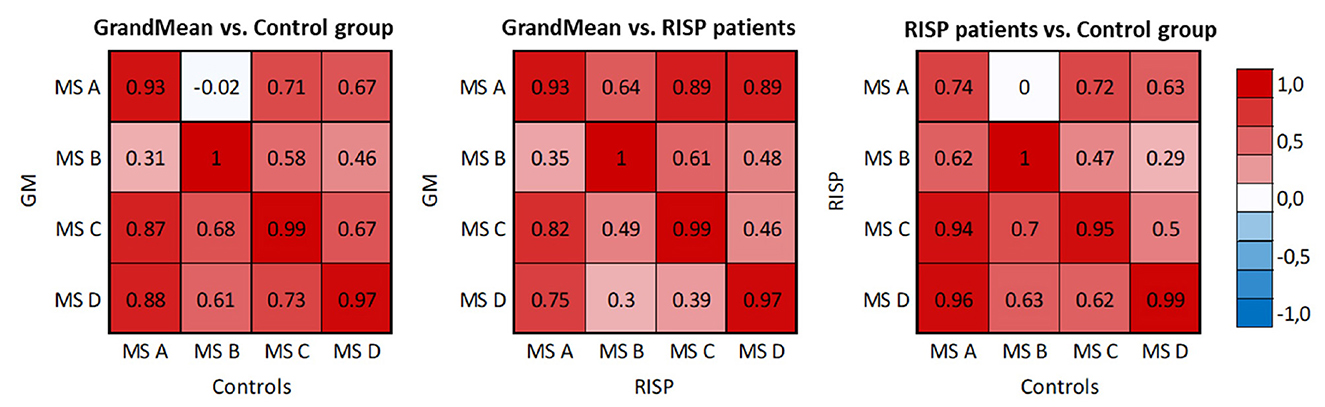
Figure 4. Spatial correlation matrices of identified MS templates of RISP patients, control group, and GM of the whole dataset. The scale was set between –1 and 1.
The correlation matrices describe correlations between the microstate topography of the RISP patients and control group and the correlation of each group and the GM microstate topographies. The correlation of both groups' topographies and GM topographies validated that using the GM template for sorting purposes is adequate.
The mean and SD values of MS parameters duration, occurrence, coverage, and GFP were computed across the groups, see Table 2. The mean values of transitions are visualized in Figure 5. Due to the non-normal probability distribution, the non-parametric test for two independent samples was used, specifically the Wilcoxon rank sum test. The resulting p-values are visualized in Table 3 and Figure 5.
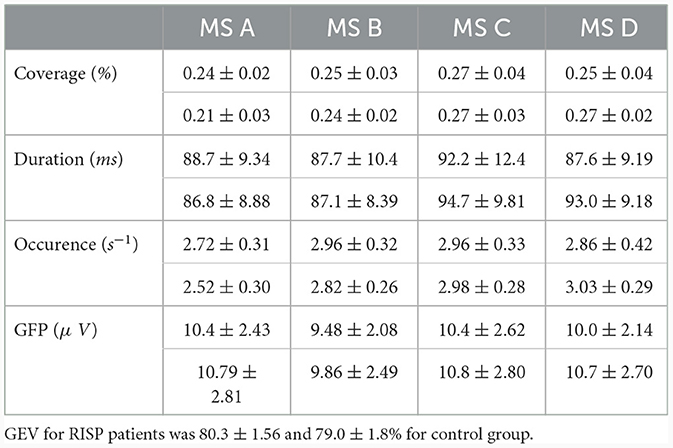
Table 2. MS temporal parameters for RISP patients (upper part of the row) and control group.
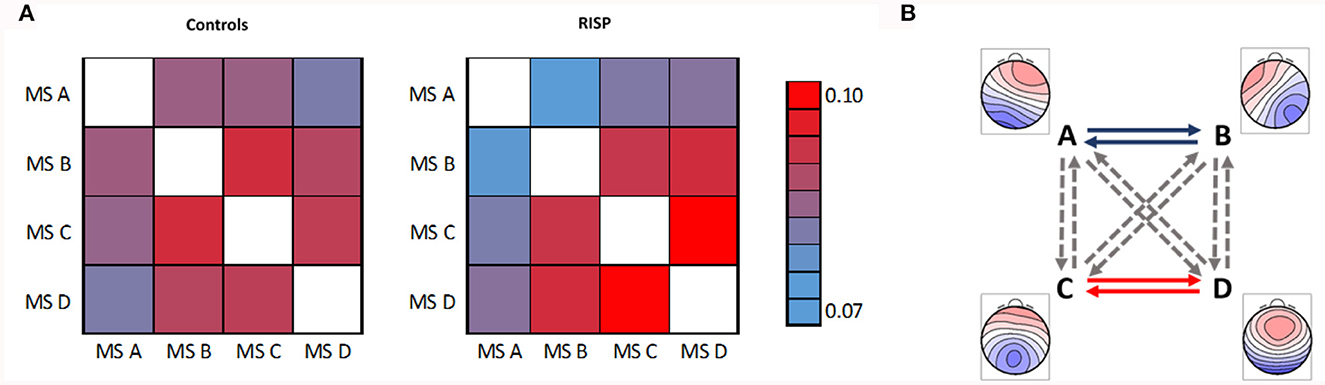
Figure 5. Mean transitions between MS. (A) Left represents mean transitions of control and (A) right RISP group. (B) Statistically significant differences of transition parameters between healthy controls and RISP group, represented by GM microstates topographical maps. The p-value of 0.05 is corrected for the multiple comparisons to 0.0042.
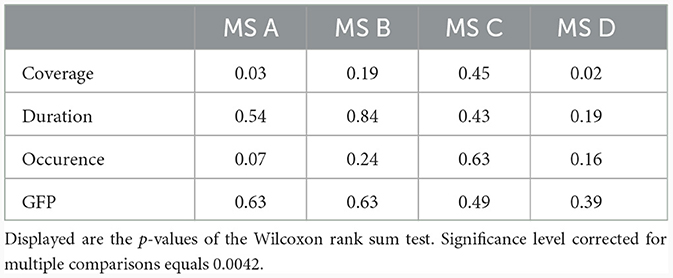
Table 3. Statistical evaluation of MS parameters between controls and RISP group.
Spatial correlations were computed for each MS in single-group analyzes. This way we checked for outlier maps, such as maps from data with artifacts. MS A and B are uncorrelated in the case of normative templates (Koenig et al., 2002) (0.04) and also in our control group (–0.04). At the same time, there is already a weak spatial correlation (0.66) in the RISP group. In RISP, the transition between A and B is reduced. Controls have a higher spatial correlation between MS C and D (0.82), while in RISP subjects MS C and D are uncorrelated (0.28). The C–D transition is elevated in RISP. The transition probability between MS C and D in RISP was negatively related to the degree of the difference in correlation between controls and RISP. When the correlation between these MS is lower, their transitions are more frequent.
In broadband spectrum, we observed significant differences in the transition between MS A and B and between MS C and D in both directions. The RISP group showed a lower transition rate between MS A and B and a higher transition rate between MS C and D than the control group.
Discussion
In this study, we present an analysis of EEG sleep parameters of patients diagnosed with RISP. Rather than analyzing actual RISP episodes, our emphasis was on quantitatively comparing the sleep of RISP patients with healthy subjects. In current literature Stefani and Högl (2021) and Wròbel-Knybel et al. (2022), RISP has mostly been associated with REM sleep, however, the precise mechanism underlying this disorder is unknown. Compared to other studies Klikovà et al. (2021) and Stefani and Högl (2021) focusing primarily on REM sleep, we targeted the NREM sleep stages using multiple analysis methods on a unique dataset obtained with strict entry criteria.
EEG spectral analysis showed a significantly higher theta activity in REM and NREM 2 sleep stages in RISP patients compared to healthy controls. This finding corresponds with the results of Mainieri et al. (2020) describing predominant theta activity during a paralysis episode (Darchia et al., 2018). Our patients did not, however, experience any paralysis episodes during the recording night. A higher occurrence of theta activity over the scalp compared to the control group might thus indicate a predisposition to a RISP episode. We also showed a slight nonsignificant increase in delta activity in REM and NREM 3. Interestingly, a similar finding, i.e., increased delta activity in patients compared to controls in REM and NREM 3 (uncorrected), was reported in a narcolepsy study (Mukai et al., 2003). 50–60% of narcolepsy patients also report experiencing sleep paralysis episodes (Drissi et al., 2016). Further EEG differences in RISP patients' REM sleep were described in our previous study (Klikovà et al., 2021). Our current study shows that there are some unusual spectral power fluctuations in REM and also in NREM sleep. These findings support previous research (Denis, 2018; Benham, 2022) about the relationship between sleep quality and the propensity to RISP episodes.
Next, EEG MS analysis was performed on the NREM stage of sleep. Specifically we opted for NREM 3 stage as it is the most thoroughly understood stage of NREM sleep along with its precise definition in terms of distinction in scoring compared to other NREM sleep stages. REM stage was not analyzed via MS analysis aligning with the practice observed in other studies which also omit REM stage from sleep MS analysis. Resulting topographical maps showed high GEV and visual similarity to canonical MS maps and we thus labeled them accordingly (A–D). The fact that the MS maps are coherent with maps found across studies (Michel and Koenig, 2018) supports the power of this analysis. After the Bonferroni correction, we found lower transition rates between MS A–B and higher transition rates of MS C–D in RISP patients' NREM 3 sleep stage. No significant differences were found in other temporal MS parameters (duration, occurrence, contribution, and GFP) and no other sleep stage was analyzed due to low correlations between template maps and individual subjects' maps.
We've also shown an overall dominance of MS C and D topographies in RISP patients' NREM sleep. MS A, B, and C were previously associated with sleep, but their temporal dynamics vary by states of vigilance and in psychiatric disorders (Krylova et al., 2021). Especially the duration parameter of MS is a frequent differentiating indicator of MS behavior (Antonova et al., 2022). We observed a slightly increased duration of MS C and MS D in RISP patients which may be pointing to a higher vigilance state in RISP patients. This duration difference, although not significant, may also be supported by the observed higher levels of theta activity during deep sleep. These results indicate the greatest stability of microstate C in terms of spatial arrangement. MS C duration parameters were previously also shown to be increased in narcolepsy patients during wake and reduced during sleep (Drissi et al., 2016). This study concluded that these microstructural EEG alterations might reflect the intrusion of brain states characteristic of wakefulness into sleep and instability of the sleep-regulating flip-flop mechanism, resulting not only in pathological switches between REM and NREM sleep but also within NREM sleep itself, which may lead to a microstructural fragmentation of the EEG (Kuhn et al., 2014).
In literature, MS C has the highest correlation across average maps in different datasets (Brodbeck et al., 2012). Transitions to MS C also often mark a pathological condition such as anxiety disorder, migraines, or dementia (Nishida et al., 2013; Al Zoubi et al., 2019; Li et al., 2022). MS C was further associated with the cingulate cortex and cingulate gyrus (Britz et al., 2010). The cingulate cortex was proposed to be the main mediator for switching between all MS classes (Pascual-Marqui et al., 2014; Custo et al., 2017) as it plays an important role in switching between vigilance states (Krylova et al., 2021).
According to a study by Brèchet et al. (2020), MS C and D show the highest occurrences in NREM sleep data. Source localization showed the sources of MS C to be in frontal brain regions and sources of MS D in occipital areas. MS C may signal physiological frontal area deactivation typical for deep sleep. The increased number of transitions between our MS maps could reflect this switching mechanism between activation-inactivation states. Furthermore, increased transitions between MS associated with vigilance may contribute to a manifestation of a RISP episode. However, caution is in order when evaluating EEG-fMRI MS analyzes due to possible artifact residuals caused by MRI which may be a contributor to the MS maps (Piorecky et al., 2019).
In terms of the strict association of sleep paralysis with REM sleep, it is also interesting that the association of another nearby REM parasomnia, as is nightmare disorder with REM sleep, which is a significant predictor of sleep paralysis (Liškovà et al., 2016), has recently become questionable. Literature states that there is no solid evidence indicating that nightmares occur more frequently during REM sleep than during NREM sleep. For instance, nightmares in PTSD patients are evenly distributed across NREM and REM periods (van der Wijk et al., 2020). There are several recent studies investigating neurophysiological correlates of nightmares, irrespective of the influence of comorbid psychopathologies (Simor and Blaskovich, 2019). Subjects who recall frequent nightmares were characterized by increased arousal-related cortical activity (Simor et al., 2013) and enhanced high-frequency electroencephalographic (EEG) oscillations during NREM sleep compared to control participants (Marquis et al., 2017; Blaskovich et al., 2020). The altered arousal-related activity was more pronounced during transitions between NREM and REM sleep states (Simor et al., 2014), suggesting that NREM-REM sleep transitions facilitating microarousals, microsleeps, and sleep interruptions may be specifically disrupted in these individuals. Interestingly, we also found altered microstate dynamics in NREM sleep, again suggesting a more complex sleep dysregulation beyond REM sleep in RISP patients. To validate our findings and further elucidate the RISP pathophysiology it would be convenient to measure subjects during sleep paralysis episodes and then compare the power changes in studied frequency bands and also focus on the characteristics of MS, all in the deep sleep phase. It should also be noted that our study did not explore the NREM sleep beta band due to previously shown indifferences between RISP and healthy subjects in REM sleep. This band could be explored in future studies also for NREM sleep. Furthermore, we acknowledge a bias toward female participants in our dataset. Future studies should try to minimize the difference in the sex of participants in order to remove its influence.
To conclude, our study shows that EEG abnormalities in RISP patients are present outside paralysis episodes and are not necessarily present only in REM sleep. Future studies could therefore further investigate these traces of RISP beyond REM sleep.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation to any qualified researcher.
Ethics statement
The studies involving humans were approved by Ethical Committee of the National Institute of Mental Health, Klecany, Czech Republic and Ethical Committee of the Third Faculty of Medicine, Charles University, Prague, Czech Republic. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
FČ: Investigation, Methodology, Visualization, Writing — original draft. VP: Data curation, Investigation, Methodology, Writing — original draft. MK: Conceptualization, Data curation, Writing — review & editing. JK: Supervision, Writing — review & editing. JB: Supervision, Writing — review & editing. MP: Conceptualization, Methodology, Visualization, Writing — original draft.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Grant Agency of the Czech Technical University in Prague, Reg. No. SGS22/200/OHK4/3T/17 and Reg. No. SGS21/140/OHK4/2T/17, by the Czech Science Foundation, Reg. no. 22-16874S, and by Programme Cooperatio Neurosciences of Charles University.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Al Zoubi, O., Mayeli, A., Tsuchiyagaito, A., Misaki, M., Zotev, V., Refai, H., et al. (2019). EEG microstates temporal dynamics differentiate individuals with mood and anxiety disorders from healthy subjects. Front. Hum. Neurosci. 13:56. doi: 10.3389/fnhum.2019.00056
Anderson, K. N., Pilsworth, S., Sharples, L. D., Smith, I. E., and Shneerson, J. M. (2007). Idiopathic hypersomnia: a study of 77 cases. Sleep 30, 1274–1281. doi: 10.1093/sleep/30.10.1274
Antonova, E., Holding, M., Suen, H. C., Sumich, A., Maex, R., and Nehaniv, C. (2022). EEG microstates: functional significance and short-term test-retest reliability. Neuroimage 2:100089. doi: 10.1016/j.ynirp.2022.100089
Benham, G. (2022). Sleep paralysis in college students. J. Am. Coll. Health 70, 1286–1291. doi: 10.1080/07448481.2020.1799807
Bettica, P., Squassante, L., Groeger, J., Gennery, B., Winsky-Sommerer, R., and Dijk, D.-J. (2012). Differential effects of a dual orexin receptor antagonist (SB-649868) and zolpidem on sleep initiation and consolidation, SWS, REM sleep, and EEG power spectra in a model of situational insomnia. Neuropsychopharmacology 37, 1224–33. doi: 10.1038/npp.2011.310
Blaskovich, B., Reichardt, R., Gombos, F., Spoormaker, V. I., and Simor, P. (2020). Cortical hyperarousal in NREM sleep normalizes from pre- to post- REM periods in individuals with frequent nightmares. Sleep 43:zsz201. doi: 10.1093/sleep/zsz201
Brèchet, L., Brunet, D., Perogamvros, L., Tononi, G., and Michel, C. (2020). EEG microstates of dreams. Sci. Rep. 10:74075. doi: 10.1038/s41598-020-74075-z
Britz, J., Van De Ville, D., and Michel, C. M. (2010). Bold correlates of EEG topography reveal rapid resting-state network dynamics. NeuroImage 52, 1162–1170. doi: 10.1016/j.neuroimage.2010.02.052
Brodbeck, V., Kuhn, A., von Wegner, F., Morzelewski, A., Tagliazucchi, E., Borisov, S., et al. (2012). EEG microstates of wakefulness and nrem sleep. NeuroImage 62, 2129–2139. doi: 10.1016/j.neuroimage.2012.05.060
Buškovà, J., and Klikovà, M. (2019). Recurrent isolated sleep paralysis. Neurologie Pro Praxi 20, 54–56. doi: 10.36290/neu.2018.018
Çoğaltay, N., and Karadağ, E. (2015). “Introduction to meta-analysis,” in Leadership and Organizational Outcomes, ed. E. Karadağ (Cham: Springer International Publishing), 19–28.
Custo, A., Van De Ville, D., Wells, W., Tomescu, M., and Michel, C. (2017). EEG resting-state networks: microstates' source localization. Brain Connect. 7:476. doi: 10.1089/brain.2016.0476
Darchia, N., Oniani, N., Sakhelashvili, I., Supatashvili, M., Basishvili, T., Eliozishvili, M., et al. (2018). Relationship between sleep disorders and health related quality of life–results from the Georgia SOMNUS study. Int. J. Environ. Res. Publ. Health 15:1588. doi: 10.3390/ijerph15081588
De Gennaro, L., Ferrara, M., and Bertini, M. (2000). The relationship between frequency of rapid eye movements in REM sleep and SWS rebound. J. Sleep Res. 9, 155–159. doi: 10.1046/j.1365-2869.2000.00193.x
Denis, D. (2018). Relationships between sleep paralysis and sleep quality: current insights. Nat. Sci. Sleep 10:355. doi: 10.2147/NSS.S158600
Denis, D., French, C. C., and Gregory, A. M. (2018). A systematic review of variables associated with sleep paralysis. Sleep Med. Rev. 38, 141–157. doi: 10.1016/j.smrv.2017.05.005
Denis, D., French, C. C., Rowe, R., Zavos, H. M. S., Nolan, P. M., Parsons, M. J., et al. (2015). A twin and molecular genetics study of sleep paralysis and associated factors. J. Sleep Res. 24, 438–446. doi: 10.1111/jsr.12282
Dongrui, G., Long, S., Yang, H., Cheng, Y., Guo, S., Yu, Y., et al. (2020). SWS brain-wave music may improve the quality of sleep: an EEG study. Front. Neurosci. 14:67. doi: 10.3389/fnins.2020.00067
Drissi, N. M., Szakàcs, A., Witt, S. T., Wretman, A., Ulander, M., Stàhlbrandt, H., et al. (2016). Altered brain microstate dynamics in adolescents with narcolepsy. Front. Hum. Neurosci. 10:369. doi: 10.3389/fnhum.2016.00369
Fèrat, V., Seeber, M., Michel, C., and Ros, T. (2020). Beyond broadband: towards a spectral decomposition of EEG microstates. Hum. Brain Map. 16:342378. doi: 10.1101/2020.10.16.342378
Held, K., Künzel, H., Ising, M., Schmid, D., Zobel, A., Murck, H., et al. (2004). Treatment with the CRH1-receptor-antagonist R121919 improves sleep-EEG in patients with depression. J. Psychiat. Res. 38, 129–136. doi: 10.1016/S0022-3956(03)00076-1
Inouye, T., Shinosaki, K., Sakamoto, H., Toi, S., Ukai, S., Iyama, A., et al. (1991). Quantification of EEG irregularity by use of the entropy of the power spectrum. Electroencephal. Clin. Neurophysiol. 79, 204–210. doi: 10.1016/0013-4694(91)90138-T
Kikuchi, M., Koenig, T., Munesue, T., Hanaoka, A., Strik, W., Dierks, T., et al. (2011). EEG microstate analysis in drug-naive patients with panic disorder. PLoS ONE 6:22912. doi: 10.1371/journal.pone.0022912
Kindler, J., Hubl, D., Strik, W., Dierks, T., and Koenig, T. (2011). Resting-state eeg in schizophrenia: auditory verbal hallucinations are related to shortening of specific microstates. Clin. Neurophysiol. 122, 1179–1182. doi: 10.1016/j.clinph.2010.10.042
Klikovà, M., Pioreckỳ, M., Miletìnovà, E., Janku, K., Dudysovà, D., and Buškovà, J. (2021). Objective rapid eye movement sleep characteristics of recurrent isolated sleep paralysis: a case-control study. Sleep 44:zsab153. doi: 10.1093/sleep/zsab153
Koenig, T., Prichep, L., Lehmann, D., Sosa, P. V., Braeker, E., Kleinlogel, H., et al. (2002). Millisecond by millisecond, year by year: normative EEG microstates and developmental stages. NeuroImage 16, 41–48. doi: 10.1006/nimg.2002.1070
Krylova, M., Alizadeh, S., Izyurov, I., Teckentrup, V., Chang, C., van der Meer, J., et al. (2021). Evidence for modulation of EEG microstate sequence by vigilance level. NeuroImage 224:117393. doi: 10.1016/j.neuroimage.2020.117393
Kuhn, A., Brodbeck, V., Tagliazucchi, E., Morzelewski, A., Wegner, F., and Laufs, H. (2014). Narcoleptic patients show fragmented EEG-microstructure during early NREM sleep. Brain Topogr. 28:1. doi: 10.1007/s10548-014-0387-1
Lehmann, D., Faber, P. L., Galderisi, S., Herrmann, W. M., Kinoshita, T., Koukkou, M., et al. (2005). EEG microstate duration and syntax in acute, medication-naïve, first-episode Schizophrenia: a multi-center study. Psychiat. Res. 138, 141–156. doi: 10.1016/j.pscychresns.2004.05.007
Lehmann, D., and Skrandies, W. (1980). Reference-free identification of components of checkerboard-evoked multichannel potential fields. Electroencephal. Clin. Neurophysiol. 48, 609–621.
Li, Y., Chen, G., Lv, J., Hou, L., Dong, Z., Wang, R., et al. (2022). Abnormalities in resting-state EEG microstates are a vulnerability marker of migraine. J. Headache Pain 23:1414. doi: 10.1186/s10194-022-01414-y
Liškovà, M., Janečkovà, D., Kluzovà Kràčmarovà, L., Mladà, K., and Buškovà, J. (2016). The occurrence and predictive factors of sleep paralysis in university students. Neuropsychiat. Dis. Treat. 12, 2957–2962. doi: 10.2147/NDT.S115629
Mainieri, G., Maranci, J.-B., Champetier, P., Leu-Semenescu, S., Gales, A., Dodet, P., et al. (2020). Are sleep paralysis and false awakenings different from REM sleep and from lucid REM sleep? A spectral EEG analysis. J. Clin. Sleep Med. 17:9056. doi: 10.5664/jcsm.9056
Mantua, J., Gravel, N., and Spencer, R. M. C. (2016). Reliability of sleep measures from four personal health monitoring devices compared to research-based actigraphy and polysomnography. Sensors 16:50646. doi: 10.3390/s16050646
Maris, E., and Oostenveld, R. (2007). Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 164, 177–190. doi: 10.1016/j.jneumeth.2007.03.024
Marquis, L. P., Paquette, T., Blanchette-Carrière, C., Dumel, G., and Nielsen, T. (2017). REM sleep theta changes in frequent nightmare recallers. Sleep 40:zsx110. doi: 10.1093/sleep/zsx110
Michel, C. M., and Koenig, T. (2018). EEG microstates as a tool for studying the temporal dynamics of whole-brain neuronal networks: a review. NeuroImage 180, 577–593. doi: 10.1016/j.neuroimage.2017.11.062
Mokhlesi, B., and Punjabi, N. M. (2012). “REM-related” obstructive sleep apnea: an epiphenomenon or a clinically important entity? Sleep 35, 5–7. doi: 10.5665/sleep.1570
Mukai, J., Uchida, S., Miyazaki, S., Nishihara, K., and Honda, Y. (2003). Spectral analysis of all-night human sleep EEG in narcoleptic patients and normal subjects. J. Sleep Res. 12, 63–71. doi: 10.1046/j.1365-2869.2003.00331.x
Nishida, K., Morishima, Y., Yoshimura, M., Isotani, T., Irisawa, S., Jann, K., et al. (2013). EEG microstates associated with salience and frontoparietal networks in Frontotemporal dementia, Schizophrenia and Alzheimer's disease. Clin. Neurophysiol. 124, 1106–1114. doi: 10.1016/j.clinph.2013.01.005
Oostenveld, R., Fries, P., Maris, E., and Schoffelen, J. M. (2011). Fieldtrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011:156869. doi: 10.1155/2011/156869
Pascual-Marqui, R., Lehmann, D., Faber, P., Milz, P., Kochi, K., Yoshimura, M., et al. (2014). The resting microstate networks (RMN): cortical distributions, dynamics, and frequency specific information flow. ArXiv 1411:1949. doi: 10.48550/arXiv.1411.1949
Pascual-Marqui, R., Michel, C., and Lehmann, D. (1995). Segmentation of brain electrical activity into microstates: model estimation and validation. IEEE Trans. Biomed. Eng. 42, 658–665.
Piorecky, M., Koudelka, V., Strobl, J., Brunovsky, M., and Krajca, V. (2019). Artifacts in simultaneous HDEEG/FMRI imaging: a nonlinear dimensionality reduction approach. Sensors 19:970. doi: 10.3390/s19204454
Sateia, M. J. (2014). International classification of sleep disorders-third edition. Chest 146, 1387–1394. doi: 10.1378/chest.14-0970
Sharpless, B. (2016). A clinician's guide to recurrent isolated sleep paralysis. Neuropsychiat. Dis. Treat. 12:S100307. doi: 10.2147/NDT.S100307
Sharpless, B. A., and Barber, J. P. (2011). Lifetime prevalence rates of sleep paralysis: a systematic review. Sleep Med. Rev. 15, 311–315. doi: 10.1016/j.smrv.2011.01.007
Simor, P., and Blaskovich, B. (2019). The pathophysiology of nightmare disorder: signs of impaired sleep regulation and hyperarousal. J. Sleep Res. 28:e12867. doi: 10.1111/jsr.12867
Simor, P., Horvàth, K., Ujma, P. P., Gombos, F., and Bòdizs, R. (2013). Fluctuations between sleep and wakefulness: wake-like features indicated by increased EEG alpha power during different sleep stages in nightmare disorder. Biol. Psychol. 94, 592–600. doi: 10.1016/j.biopsycho.2013.05.022
Simor, P., Körmendi, J., Horvàth, K., Gombos, F., Ujma, P. P., and Bòdizs, R. (2014). Electroencephalographic and autonomic alterations in subjects with frequent nightmares during pre-and post-REM periods. Brain Cogn. 91, 62–70. doi: 10.1016/j.bandc.2014.08.004
Slepian, D. (1978). Prolate spheroidal wave functions, fourier analysis, and uncertainty—v: the discrete case. Bell Syst. Tech. J. 57, 1371–1430.
Stefani, A., and Högl, B. (2021). Nightmare disorder and isolated sleep paralysis. Neurotherapeutics 18, 100–106. doi: 10.1007/s13311-020-00966-8
Takeuchi, T., Miyasita, A., Sasaki, Y., Inugami, M., and Fukuda, K. (1992). Isolated sleep paralysis elicited by sleep interruption. Sleep 15, 217–225.
Tarailis, P., Šimkutė, D., Koenig, T., and Griškova-Bulanova, I. (2021). Relationship between spatiotemporal dynamics of the brain at rest and self-reported spontaneous thoughts: an EEG microstate approach. J. Personal. Med. 11:111216. doi: 10.3390/jpm11111216
Terzaghi, M., Ratti, P. L., Manni, F., and Manni, R. (2011). Sleep paralysis in narcolepsy: more than just a motor dissociative phenomenon? Neurol. Sci. 33, 169–172. doi: 10.1007/s10072-011-0644-y
van der Wijk, G., Blaskovich, B., Farahzadi, Y., and Simor, P. (2020). Unaltered EEG spectral power and functional connectivity in REM microstates in frequent nightmare recallers: are nightmares really a REM parasomnia? Sleep Med. 75, 192–200. doi: 10.1016/j.sleep.2020.07.014
Walther, B. W., and Schulz, H. (2004). Recurrent isolated sleep paralysis: polysomnographic and clinical findings. Somnologie 8, 53–60. doi: 10.1111/j.1439-054X.2004.00017.x
Wròbel-Knybel, P., Flis, M., Rog, J., Jalal, B., Wołkowski, L., and Karakuła-Juchnowicz, H. (2022). Characteristics of sleep paralysis and its association with anxiety symptoms, perceived stress, ptsd, and other variables related to lifestyle in selected high stress exposed professions. Int. J. Environ. Res. Publ. Health 19:7821. doi: 10.3390/ijerph19137821
Keywords: EEG, polysomnography, recurrent isolated sleep paralysis (RISP), microstate analysis, EEG spectral analysis
Citation: Černý F, Piorecká V, Kliková M, Kopřivová J, Bušková J and Piorecký M (2024) All-night spectral and microstate EEG analysis in patients with recurrent isolated sleep paralysis. Front. Neurosci. 18:1321001. doi: 10.3389/fnins.2024.1321001
Received: 13 October 2023; Accepted: 25 January 2024;
Published: 08 February 2024.
Edited by:
Andrea Romigi, Università Telematica Internazionale Uninettuno, ItalyReviewed by:
Umesh Vivekananda, University College London, United KingdomMahmoud Alipour, Nationwide Children's Hospital, United States
Copyright © 2024 Černý, Piorecká, Kliková, Kopřivová, Bušková and Piorecký. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Filip Černý, ZmlsaXAuY2VybnkuMUBmYm1pLmN2dXQuY3o=