1
Laboratory for Perception and Memory, Department of Neuroscience, Centre National de la Recherche Scientifi que, Institut Pasteur, Paris, France
2
Department of Anatomy, Pharmacology and Forensic Medicine, National Institute of Neuroscience-Italy, University of Turin, Turin, Italy
A commentary on
Interneurons produced in adulthood are required for the normal functioning of the olfactory bulb network and for the execution of selected olfactory behaviors.
by Breton-Provencher, V., Lemasson, M., Peralta, M.R. 3rd, Saghatelyan, A. (2009). J. Neurosci. 29(48), 15245–15257.
Constitutive neurogenesis clearly occurs within the adult central nervous system of almost all mammals. This process generates new neurons in at least two areas of the brain: the olfactory bulb (OB) and the dentate gyrus (DG) of the hippocampus. However, it remains unknown why new neurons specifically populate those two neural structures rather than other areas. In empirical experiments, adult neurogenesis was impaired and the consequences of this impairment for the neural circuitry and behavior of animals were investigated with a view to determining the role of new neurons in the adult brain.
In the December issue of the Journal of Neuroscience, Breton-Provencher et al. addressed this issue, by using drugs to establish a transient blockade of adult neurogenesis and combining several techniques to evaluate the consequences of this blockade for OB circuits and particular types of behavior. Here, we summarize and discuss some of their findings in light of other recently published data.
Several experimental approaches have been developed for blocking adult neurogenesis. Most were inspired by tools originally designed to reduce cell proliferation in the treatment of cancers. For example, cytosine-β-D-arabinofuranoside (AraC) is an antimitotic drug used in chemotherapy to treat hematological malignancies in humans. Breton-Provencher et al. used osmotic pumps to infuse AraC into the lateral ventricle (LV) of adult mice for 28 days. The neurogenic niches of the neural precursors that migrate to the OB circuit are located in the wall of the LV, in the subventricular zone (SVZ). AraC infusion into the LV is thus a highly effective method for decreasing OB neurogenesis. After AraC-treatment, the authors reported a 75% decrease in Dcx levels on western blots, this protein being a marker for immature neuroblasts; they also showed an almost complete absence of Dcx immunostaining in the OB (see Breton-Provencher, Figure 1A,B). This infusion also affected neurogenesis in the DG, but to lesser extent (−50%; see Breton-Provencher, Figure 1C). Different methods for blocking adult neurogenesis may have different effects on the proportion of new neurons in the OB or DG (see Table 1
). Focused irradiation tends to be more specific than pharmacological methods (Clelland et al., 2009
; Lazarini et al., 2009
; Valley et al., 2009
). Various genetic approaches have also been used to decrease the size of newly generated neuron population in a transient manner (Imayoshi et al., 2008
; Deng et al., 2009
). However, these genetic approaches have, to date, proved even less specific than irradiation or pharmacological approaches, affecting adult neurogenesis to similar extents in the OB and DG. Adult neurogenesis may serve different functions in these two structures, but may also underlie common patterns of behavior, such as exploratory or learning behavior. It is therefore important to check the specificity of the blockade of adult neurogenesis in both neurogenic areas, particularly when trying to determine the contribution of new neurons to specific types of behavior.
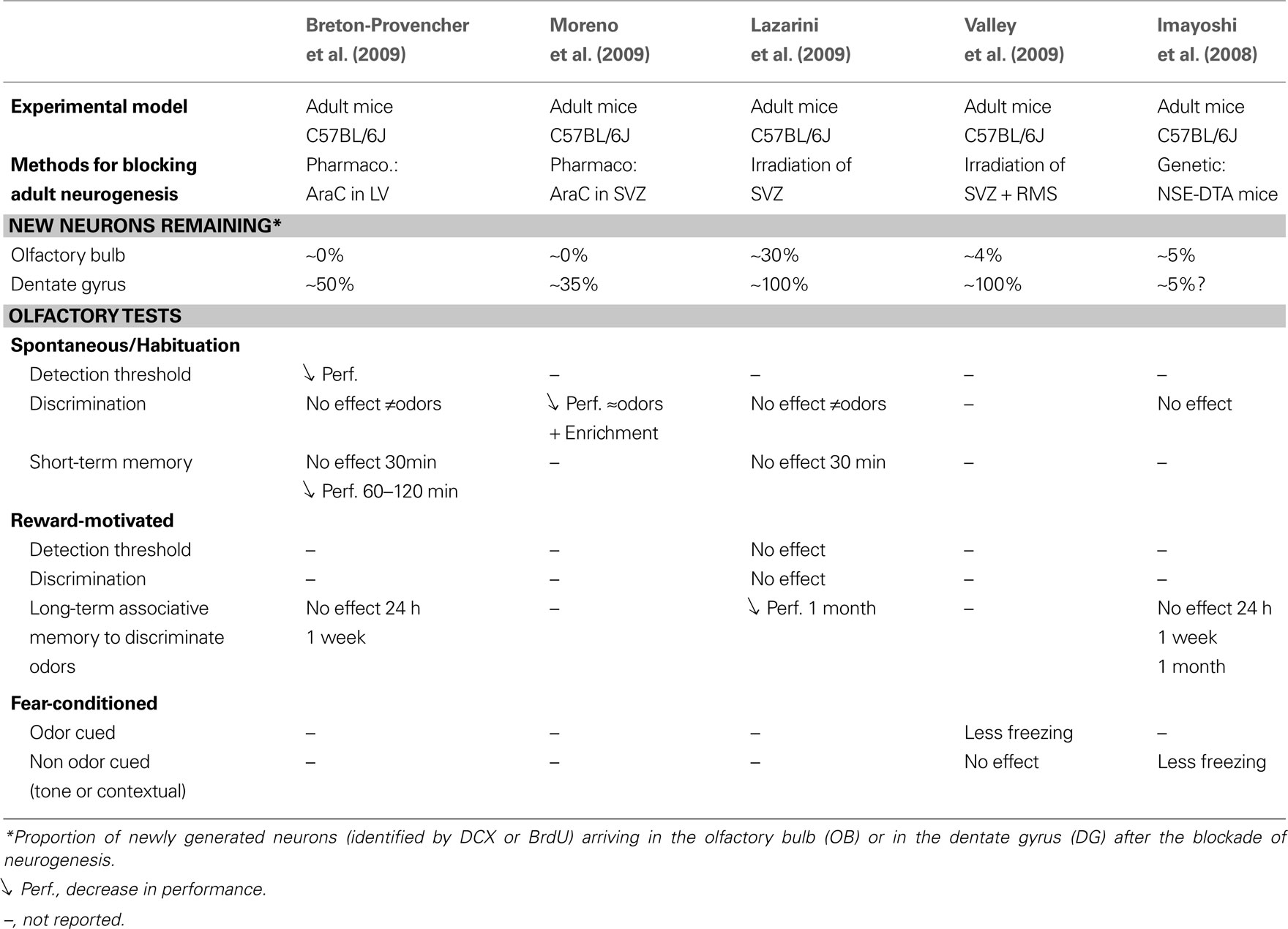
Table 1. Comparison of recent studies blocking adult neurogenesis and examining the consequences of the blockade for olfactory behavior.
Breton-Provencher et al. investigated the role of neurons generated in adult animals, in several behavioral tests. They first checked that the effects observed were specifically due to the ablation of new neurons in the OB, by demonstrating that AraC-treatment had no effect on the performance of the mice in diverse activities that might indirectly influence their performance in specific olfactory tests (Breton-Provencher, Figure 6). In particular, they reported that AraC-treated mice performed well in exercises involving spontaneous locomotor activity, object exploration and recognition. They also tested the treated mice in tail suspension and novelty suppressed feeding tests, to demonstrate that the treatment did not induce anxious behavior (see also (Kitamura et al., 2009
; Moreno et al., 2009
). These controls were necessary and entirely appropriate. Nevertheless, the role of the hippocampus in olfactory behavior cannot be excluded (Gourevitch et al., 2010
). In our opinion, for the complete exclusion of possible effects of a partial blockade of DG neurogenesis on the olfactory tests, a straightforward control would have been to evaluate the olfactory behavior of the mice whilst impairing DG neurogenesis alone.
The olfactory behavior experiments showed that continuous neurogenesis in the OB was required only for certain non reinforced olfactory tasks (Breton-Provencher, Figure 7). The authors showed that AraC-treated mice had higher detection thresholds for odorants. The authors therefore used high odor concentrations for their subsequent evaluation of olfactory memory in the mice. They found that brief, non reinforced exposure to an odor was remembered for only 30 min by the AraC-treated mice, whereas the same odor was remembered for at least 2 h by the saline-treated mice. Finally, the authors assessed the long-term associative olfactory memory of the mice with impaired adult neurogenesis. In particular, they tested the capacity of the mice to remember an odor associated with a reward (food) and to discriminate this odor from an odor not associated with a reward. They reported that 24 h or 1 week after learning, mice with impaired adult neurogenesis could still remember and discriminate the reward-associated odor.
Four other independent research groups have also recently studied the effect of blocking adult neurogenesis on olfactory behavior (Imayoshi et al., 2008
; Lazarini et al., 2009
; Moreno et al., 2009
; Valley et al., 2009
). However, no clear consensus emerges from their results. Different methods to block adult neurogenesis and to evaluate olfactory behavior have been used, making it difficult to compare and to interpret the various results obtained. We have tried to summarize the results obtained in these studies in Table 1
. Overall, these independent studies seem to show, consistently, that blocking adult neurogenesis in the OB does not affect spontaneous discrimination between different odors (Imayoshi et al., 2008
; Breton-Provencher et al., 2009
; Lazarini et al., 2009
). However, another recent study showed that blocking adult neurogenesis resulted in a dramatic decrease in performance in a task involving discrimination between two perceptually similar odorants after passive exposure (Moreno et al., 2009
). Unlike Breton-Provencher et al., Lazarini et al. reported that the blockade of adult neurogenesis had no effect on odor detection thresholds. This difference in the results obtained by these two studies may be accounted for by differences in the nature of the tests, which were reward-motivated in Lazarini’s study but not in Breton-Provencher’s study. However, spontaneous olfactory tasks do not seem to be the only tasks requiring adult neurogenesis. The blockade of adult neurogenesis in the OB by focused irradiation, results in a decrease in freezing behavior conditioned by odor cues (Valley et al., 2009
), and an impairment of the long-term memory (1 month) of reward-associated odor (Lazarini et al., 2009
; but see Imayoshi et al., 2008
).
Interestingly, although mice with impaired neurogenesis may perform well in learning tests, the amount of time for which the learned information is stored in their memory seems to depend on adult neurogenesis. Adult neurogenesis seems to play a key role in various types of specific olfactory behavior and does not appear to be restricted to spontaneous, reward-motivated or fear-conditioned learning (see Table 1
).
Breton-Provencher et al. tried to associate the results obtained in their behavior experiments with putative changes in the OB circuitry, by investigating whether the blockade of adult neurogenesis induced significant changes in OB activity. By combining immunohistochemistry and patch-clamping techniques, the authors convincingly demonstrated that the number of functional GABAergic contacts received by mitral cells was much smaller in the AraC-treated mice than in the controls (Breton-Provencher, Figures 3 and 4). Surprisingly, despite the prevention of new neuron production over a period of 28 days, the overall number of granule cells was not modified (Breton-Provencher, Figures 2A,B). The authors of the study also demonstrated that the dendrite length and spine density of the neurons produced 1 month before AraC-treatment were not affected by the shortage of new neurons (Breton-Provencher, Figures 2C–E). It has been reported on several occasions that a rigorous selection of newly generated neurons for survival occurs around 2 weeks after their arrival in the OB (Mouret et al., 2008
). This raises questions as to whether the blockade of new neuron production increases the survival of neurons generated just before AraC-treatment, thus accounting for the lack of change in the overall number of granule cells. This hypothesis could be tested experimentally by counting the number of neurons generated shortly before the beginning of the treatment, and by showing that, 1 month later, a larger proportion of these neurons survived in AraC-treated than in saline-treated mice.
The observation that mitral cells received about half as many spontaneous GABAergic synaptic events in AraC-treated animals than in controls, despite the presence of similar numbers of granule cells, is also rather surprising. The authors interpreted this result as a consequence of new neurons being prone to plastic excitatory centrifugal inputs unlike older pre-existing granule cells (Nissant et al., 2009
). However, this conclusion is probably invalidated by the observation that, even in the presence of TTX or Kynurenate, the frequency of IPSCs is significantly lower in mitral cells of AraC-treated animals. Thus, to account for these results, it still need to be demonstrated that GABA synaptic release is much more efficient in newly generated neurons than in older neurons, or that newly generated neurons simply make far more functional GABAergic contacts.
The GABAergic interneurons in the granule cell layer are known to synchronize the activity of neurons in the mitral cell layer. The authors recorded local field potentials (LFPs) around the mitral cell layer of brain slices from acutely treated mice and analyzed the gamma oscillations (20–80 Hz) evoked by electrical stimulation of the olfactory nerve (Breton-Provencher, Figure 5). Blocking the supply of new interneurons to the adult OB did not affect the magnitude of the LFP oscillations, but tended to reduce their frequency.
Similar results for oscillation frequencies have been obtained for knockout mice lacking GABAA α1-subunit receptors (Lagier et al., 2007
). In α1-knockout mice, the remaining GABAA α3-subunit receptors in mitral cells were shown to mediate less frequent spontaneous IPSCs of unchanged amplitude, but with a slower decay, resulting in slower gamma oscillations (Lagier et al., 2007
). However, it should be borne in mind that Lagier et al. studied mice with a constitutive KO, which may have affected OB development or long-term compensation by other GABAergic mechanisms. Nevertheless, the striking similarities in the effects on oscillations between the studies of Lagier and Breton-Provencher lead us to think that the properties of the synaptic contacts established with mitral cells may differ between newly generated and pre-existing neurons, particularly in terms of post-synaptic GABAA receptor subunits, for example. If this hypothesis were correct though, the kinetics of the spontaneous GABAergic events measured in the mitral cells of AraC-treated mice should display a slower decay, similar to that for α3-subunit receptors.
By contrast, another study showed that progressively blocking every subtype of GABAA receptor with various concentrations of Gabazine reduced the amplitude of evoked LFP oscillations, but not their frequencies (Bathellier et al., 2006
). Bathelier et al. induced a broad blockade of post-synaptic receptors, whereas Breton-Provencher et al. eliminated a subpopulation of interneurons generated in adult animals. The contrasting results obtained by Bathelier et al. and Breton-Provencher et al. again suggest that newly generated neurons do not perform the same function as the majority of OB interneurons, which are presumably more mature. Nevertheless, the challenge remains to find a clear explanation of the mechanisms underlying the qualitative changes in network dynamics observed in the absence of newly generated neurons.
Determining the value of new neurons to the adult OB and DG is an important and timely challenge in the field of neurobiology. The work of Breton-Provencher et al., together with recently published analyses of animals with impaired adult neurogenesis provides us with precious food for thought concerning the possible role of new neurons (Imayoshi et al., 2008
; Clelland et al., 2009
; Deng et al., 2009
; Kitamura et al., 2009
; Lazarini et al., 2009
; Moreno et al., 2009
; Valley et al., 2009
). The contrasting results obtained in different studies blocking adult neurogenesis and assessing olfactory behavior highlight the need for further investigations to decipher the precise function of adult neurogenesis for the sense of smell. It remains an open question whether newly generated interneurons in the OB play roles different from those of their older, pre-existing counter-parts. We also cannot rule out the possibility that newly generated neurons have different roles at different stages of maturation. This process is probably even more complex than this, in that adult neurogenesis populates the OB with several subtypes of interneurons, which probably have different functions. However, despite the many questions that remain unresolved, the efforts of these authors to associate behavioral effects and changes in OB circuit activity are remarkable, and will no doubt represent a major step toward a greater understanding of the role of adult neurogenesis in olfaction.
We thank the members of the laboratory, including, in particular, Mariana Alonso, Gabriel Lepousez, Françoise Lazarini, Antoine Nissant, Gilles Gheusi and Pierre-Marie Lledo, for fruitful discussions. This work was supported by the Agence Nationale de la Recherche (ANR-2007 SEST-01411), the Fondation pour la Recherche Médicale “Equipe FRM”, the Groupe AG2R, the Ecole des Neurosciences de Paris (ENP), and a doctoral fellowship from Università Italo-Francese (Progetto Vinci).