 Willams Oliveira1†
Willams Oliveira1† Oswaldo Cruz-Neto2†
Oswaldo Cruz-Neto2† Jéssica L. S. Silva2†
Jéssica L. S. Silva2† Marcelo Tabarelli2†
Marcelo Tabarelli2† Carlos A. Peres3,4†
Carlos A. Peres3,4† Ariadna V. Lopes2*†
Ariadna V. Lopes2*†- 1Programa de Pós-Graduação em Biologia Vegetal, Departamento de Botânica, Universidade Federal de Pernambuco, Recife, Pernambuco, Brazil
- 2Departamento de Botânica, Universidade Federal de Pernambuco, Recife, Pernambuco, Brazil
- 3School of Environmental Sciences, University of East Anglia, Norwich, United Kingdom
- 4Instituto Juruá, Manaus, Brazil
Climate change is one of the main drivers of biological reorganization, population decline of pollinators, and disruption of species interactions. These impacts represent a major threat to crop pollination and human food security. Here, we tested the hypothesis that the spatial mismatches between Neotropical food plant species and their bee pollinators are exacerbated under scenarios of projected climate change. To investigate this hypothesis we performed species distribution modeling to simulate the effects of climate change on suitable habitats for the occurrence of both native food plants and their main pollinators. We selected three economically important food plants native to Brazil bearing a self-incompatible reproductive system that is strictly dependent on pollinators: (1) Bertholletia excelsa, (2) Eugenia uniflora, and (3) Passiflora edulis; and we selected the main effective bee pollinators of each plant species: (1) Apis mellifera (i.e., pollinator of E. uniflora), (2) Eulaema mocsaryi (i.e., pollinator of B. excelsa), and (3) Xylocopa frontalis (i.e., pollinator of P. edulis). We documented that climate change will likely distinctly affect areas of suitable habitats for food plants and their main bee pollinators across Brazil, in which all species will likely experience contractions in their ecological niches. In addition, we also documented that suitable habitats were reduced for the co-occurrence of all food plants and their pollinators. Specifically, 51.5% for P. edulis and X. frontalis, 76% for B. excelsa and E. mocsaryi, and 54% for E. uniflora and A. mellifera. Therefore, these findings underscore that plausible climate change scenarios can act as a potential driver of spatial mismatches between food plants and their main pollinators, disrupting the pollination of these food plants. Our results show that plant and pollinator species respond negatively to the impacts of climate change under all scenarios, which can result in alarming projections for strictly bee-pollinated self-incompatible plant species. This study reaffirms that bees are sensitive to climate change, highlighting the negative impact even for the exotic European honeybee, Apis mellifera. Finally, climate change could impact crop pollination, with detrimental implications for food production and food security.
1 Introduction
Climate change is rapidly altering biodiversity at all levels, from genes to ecosystems (Scheffers et al., 2016; IPCC, 2021), with significant consequences for essential ecological services like pollination. It is expected that by 2050 climate change will lead to an average temperature increase of up to 4˚C (IPCC, 2014). Overall, climate change has been associated with modifications at different levels of biological organization, affecting the maintenance of ecological processes and the provision of ecosystem services (Houghton et al., 2001; Malhi and Wright, 2004; Malhi et al., 2008; Scheffers et al., 2016; Ferreira et al., 2021), such as pollination (e.g., Botkin et al., 2007; Loarie et al., 2008; Giannini et al., 2017, 2020; Silva et al., 2019, 2022; Sales et al., 2021; Millard et al., 2023). In response to climate change, some species move to new more suitable habitats, as a way of adjusting their climatic envelope (e.g., Urban, 2015; Chen et al., 2011; Pecl et al., 2017). In this way, climate change acts as a strong environmental driver capable of reorganizing biological assemblages, affecting many species interactions, leading to incompatible geographic distributions between interacting species (Schweiger et al., 2008; Valiente-Banuet et al., 2015; Giannini et al., 2017, 2020; Sales et al., 2021). Then, climate change can cause spatiotemporal mismatches, changes in ecological networks with the formation of new interactions, and shifts in dispersal capacity (Memmott et al., 2007; Schleuning et al., 2020). In addition, incompatibilities in plant-animal interactions are also driven by changes in phenology, such as flowering time or animal emergence and migration (Fitter and Fitter, 2002; Kharouba and Vellend, 2015; Cohen et al., 2018; Kharouba et al., 2018).
Bees are widely reported to play a key role in ecosystem functioning, contributing to the pollination of most wild plants and crops (Klein et al., 2007; Ollerton et al., 2011; Oliveira et al., 2024). However, bees have experienced globally declines in abundance and richness due to land use change, biological invasions, pesticides, and agricultural expansion (Potts et al., 2010; Dicks et al., 2016; Zattara and Aizen, 2021; Albacete et al., 2023). In addition to these stressors, climate change has also been widely reported as one of the main causes of pollinator declines across the globe (IPBES, 2016; Potts et al., 2010, 2016; Giannini et al., 2017; Lima and Marchioro, 2021; Sales et al., 2021), as observed for honeybees and bumblebees (Becher et al., 2013; Kerr et al., 2015; Soroye et al., 2020). Warmer temperatures, changes in precipitation regimes, and higher frequency of extreme weather events affect the behavior and survival of bee species (Goulson et al., 2015; Soroye et al., 2020; IPCC, 2021). In general, rising temperatures can negatively impact plant-pollinator interactions through reduced visitation rates (e.g., Rader et al., 2013; Maluf et al., 2022). For example, watermelon crops under extreme climate change scenarios incur a 14.5% decline in pollination services provided by managed bees (Rader et al., 2013). Then, plant responses to climate change are also influenced by how animals respond to these changes (Schleuning et al., 2020). Thus, disruptions in plant-animal interactions intensified by the effects of climate change can impact biodiversity and the status quo of species interactions.
As a major threat to the maintenance of natural and managed populations globally (Pacifici et al., 2017; Potts et al., 2016; Rafferty, 2017), climate change is a major potential driver of reduced agricultural yields (IPBES, 2016; BPBES/REBIPP, 2019; Giannini et al., 2017, 2020; Sales et al., 2021; Millard et al., 2023). The reduction in pollinator populations driven by climate change affects the provision of pollination services for many agricultural crops that are pollinator-dependent and economically important, such as tomato, guava, coffee and avocado (Costanza et al., 2014; IPBES, 2016; BPBES/REBIPP, 2019; Giannini et al., 2017, 2020; Sales et al., 2021), all of which could reduce the productivity of agricultural crops and negatively affect the global economy (IPBES, 2016). Since pollinators play an important role in the production of many fruits and seeds that are consumed by humans, making a large and strong contribution to global agricultural production (Giannini et al., 2015; Dicks et al., 2016; Novais et al., 2018), compromising the pollination process in pollinator-dependent crops can result in low economic yields (Garibaldi et al., 2013), which directly affects human food security (IPBES, 2016; Oliveira et al., 2023).
The total area cultivated with pollinator-dependent crops has expanded in recent decades (Aizen et al., 2019; Oliveira et al., 2023), and, consequently, the need for pollinators and pollination has also increased. Thus, it is urgent to understand how to mitigate the effects of climate change on crop pollination to ensure biodiversity persistence and safeguard food sovereignty. In Brazil, for example, 60% of all crops depend on pollinators (Giannini et al., 2015). Furthermore, Brazil ranks 5th in its human population, is the 3rd largest agricultural producer and exporter, and is arguably Earth’s most megadiverse country (e.g., Lopes et al., 2021; Porto et al., 2020; Oliveira et al., 2023a). However, food insecurity and hunger are still a major problem in Brazil, which returned to the hunger map in 2021, with over 60 million people facing moderate to severe food insecurity in 2020 (FAO, 2021). Understanding crop-pollinator interactions occurring in Brazil thus appears as an excellent opportunity to examine the effects of climate change on selected cases of self-incompatible or highly pollinator-dependent crops and discuss the implications for crop pollination and food security in light of the future scenarios.
In this study, we seek to understand how climate change simultaneously affects the range and spatial distribution of three economically important food plants native to Brazil and their respective main pollinators. Here we focus on Passiflora edulis (Passionfruit), Bertholletia excelsa (Brazil-nut), and Eugenia uniflora (Pitanga or Brazilian cherry) as model species to investigate the impacts of climate change on plant-pollinator interactions. These species are key in agroforestry systems, which restore ecosystems by enhancing soil productivity and supporting biodiversity while mitigating climate change (Urruth et al., 2022; Lima et al., 2023). In addition, these three food plant species are also excellent models because they are self-incompatible (i.e., rely on cross-pollination to set fruits and seeds), and essentially dependent on pollinators; in other words, yields are reduced by at least 90% in the absence of pollinators (Klein et al., 2007). We tested the hypothesis that the current spatial distribution of self-incompatible food plant species and their main pollinating bees will be highly modified under different scenarios of projected climate change, resulting in a mismatch in their new regions of occurrence, thereby threatening crop pollination services. Therefore, we expected a reconfiguration of the areas of occurrence leading to a spatial mismatch in plant-pollinator interactions.
2 Materials and methods
2.1 Selection of food plant and pollinator species
To conduct our climate modeling, we selected the three native Brazilian food plants bearing self-incompatible reproductive system (i.e., obligatory cross-pollination) and with the highest Economic Value of Pollination (EVP) (BPBES/REBIPP, 2019; Oliveira et al., 2024); these species are strictly dependent on pollinators to set seed (sensu Klein et al., 2007): (1) Passiflora edulis (EVP= US$284,549,580/yr); (2) Bertholletia excelsa (EVP= US$75,586,750/yr); and (3) Eugenia uniflora (EVP= US$264,600/yr). Additionally, we selected their main effective pollinators: (1) the native carpenter bee Xylocopa frontalis, for P. edulis, (2) the native orchid bee Eulaema mocsaryi, for B. excelsa, and (3) the introduced honeybee Apis mellifera, for Eugenia uniflora (BPBES/REBIPP, 2019; Diniz and Buschini, 2016; Cavaltante et al., 2018; Sales et al., 2021).
2.2 Occurrence data
We compiled a comprehensive database on the georeferenced range data of the three food plants and their pollinators. For plant and pollinators presence-only occurrence we used the Global Biodiversity Information Facility (GBIF), an international platform that provides open access to data on species occurrence (GBIF, 2021) (https://www.gbif.org, last accessed May 2023). For plant occurrence, we also used the REFLORA - Herbário Virtual, a virtual herbarium catalogue containing information on Brazilian plants (http://reflora.jbrj.gov.br/reflora/herbarioVirtual, last accessed May 2023); and the Botanical Information and Ecology Network Platform (BIEN), a global database that provides data on plant diversity, trait records, and species distribution, including georeferenced plant observation data from herbarium, field plot, and survey inventory records (https://bien.nceas.ucsb.edu/bien/biendata, last accessed May 2023). To access the BIEN database, we use the BIEN package (Maitner et al., 2018; Silva et al., 2022) available from the R 4.1.1 platform (R Core Team, 2021). To perform our analysis, we excluded all repeated and mismatched occurrence data for each plant and pollinator species. We obtained a total of 3,479 and 2,241 geographic coordinates for plant and pollinator species, respectively. Supplementary Table S1 contains the DOI of each occurrence search by species from GBIF (e.g., Taheri et al., 2021; Silva et al., 2019, 2022).
2.3 Current and future environmental variables
We downloaded climate data summarizing aspects of precipitation, temperature, and elevation, at 2.5 min spatial resolution, from WorldClim project 2.1 for contemporary times (average for the years 1970 - 2000) and the future in 2090 (average 2081 - 2100) (Fick and Hijmans, 2017; Silva et al., 2022). In our analysis of future climate projections spanning the period from 2081 to 2100, we relied upon data derived from the Coupled Model Intercomparison Project Phase 6 (CMIP6) (Eyring et al., 2016). The CMIP6 models are currently accessible and exhibit a heightened sensitivity to climate compared to their predecessors (for more details see Eyring et al., 2016). For this study, we deliberately selected three distinct Shared Socioeconomic Pathways (SSPs) scenarios (Riahi et al., 2017; IPCC, 2021).
The three SSPs we considered here are as follows: (1) the “middle of the road” scenario, SSP2-4.5, which embodies optimism in terms of mitigation and adaptation, envisaging that social, economic, and technological trends will remain relatively consistent with historical patterns. It assumes moderate population growth, reduced income inequality, and limited environmental degradation; (2) the “regional rivalry–rocky road” scenario, SSP3-7.0, which presents a more pragmatic view of the challenges associated with mitigation and adaptation. It assumes limited investments in education, healthcare, and technological progress in the world’s poorest regions, coupled with rapid population growth, rising inequality, and significant environmental degradation. This pathway aims for radiative forcing stabilization by 2100; and (3) the “fossil-fueled development—taking the highway” scenario, SSP5-8.5, which is pessimistic assuming substantial challenges for mitigation, while those for adaptation are minimal. SSP5-8.5 is characterized by the exploitation of abundant fossil fuel reserves, resulting in the highest greenhouse gas emissions and a significant increase in terrestrial temperatures (Riahi et al., 2017; IPCC, 2021). Our selection of SSP scenarios was based on the BCCCSM2-MR (Wu et al., 2021), CanESM5 (Swart et al., 2019), and MIROC6 (Tatebe et al., 2019) general circulation models, which have shown superior performance in Neotropical regions, as corroborated by previous studies (e.g., Cai et al., 2020; Fuentes-Castillo et al., 2020; Menéndez-Guerrero et al., 2020; Sales et al., 2021; Silva et al., 2019, 2022). Furthermore, we incorporated elevation data as an essential edaphic variable, sourced from the WorldClim project 2.1 (Fick and Hijmans, 2017).
To reduce issues related to overfitting and collinearity among the 19 climatic variables and elevation, our approach involved conducting separate modeling for each species based on uncorrelated variables within both the present and future climate scenarios. The assessment of correlations among predictor variables was achieved through principal component analysis (PCA) within R 4.1.1 (R Core Team, 2021), and we opted to use the principal components responsible for 95% of the overall variation in environmental variables (De Marco and Nóbrega, 2018). Ultimately, selected predictor variables for our models encompassed the following: (1) Temperature seasonality (BIO4); (2) Annual precipitation (BIO12); (3) Precipitation of the wettest quarter (BIO16); (4) Precipitation of the warmest quarter (BIO18); (5) Precipitation of the coldest quarter (BIO19); and (6) Elevation. These specific variables have been established as significant factors in modeling plant species distributions, as substantiated by previous studies (Marengo et al., 2017; Silva et al., 2019, 2022; Cavalcante et al., 2020).
2.4 Species distribution modeling
We applied three distinct algorithms to predict habitat suitability to food plant species native to Brazil and their main pollinators. These algorithms included: (1) The maximum entropy method, the most widely used method for predicting species distributions (e.g., Fonseca et al., 2015; Silva et al., 2019), based on the principle of maximum entropy, in which the best approximation to an unknown probability distribution is the one that satisfies any constraint on the distribution (Phillips et al., 2006); (2) The support vector machine method, which presents performance compatible with or superior to methods that consider only the presence data and with moderate amounts of data, avoiding common problems and limitations (Drake et al., 2006); and (3) The random forest method, non-parametric analysis, which uses a low number of test samples, and provides high performance compared to other traditional regression algorithms (Sahragard et al., 2018). These selected algorithms are well-suited for working with presence and pseudo-absence data, an approach that aligns with the characteristics of the data collected within the localities surveyed in this study (Andrade et al., 2020). The combined use of these three algorithms improves the quality of the generated distribution models (e.g., Silva et al., 2022). To execute these analyses, we utilized the “ENMTML” R package (Andrade et al., 2020). A total of 15 replicates were generated for each algorithm, corresponding to each climate scenario (current, SSP-2.45, SSP-3.70, and SSP5-8.5) for each food plant and pollinator species. Each occurrence dataset was partitioned into two groups: a training set, comprising 70% of the sampled occurrence data for each plant and pollinator species, and a test or validation set, encompassing the remaining 30% of the sampled occurrence data. This partitioning was achieved through the bootstrap method. Given that the algorithms applied are predicated on the basis of presence and pseudo-absence data, we configured the distribution models to identify 500 pseudo-absences in grid cells characterized by lower climatic suitability for the presence of each target species (Barbet-Massin et al., 2012).
The evaluation of each model’s performance involved the use of two widely recognized metrics: True Skill Statistics (TSS) and the Area Under the Curve (AUC). These metrics have previously been applied in studies assessing plant distribution models (Sales et al., 2021; Silva et al., 2019, 2022). TSS and AUC are established measures for gauging the goodness-of-fit of species distribution models (Allouche et al., 2006). The TSS offers a comprehensive evaluation of model accuracy by considering its performance relative to random accuracy. It yields a score within the range of -1 to 1, with values close to 1 indicating optimal model performance. Typically, TSS values exceeding 0.5 are considered indicative of satisfactory model performance. On the other hand, the AUC values fall within the 0 to 1 range, with the accuracy of the model being classified as: (1) low accuracy (0.5-0.7), (2) moderate accuracy (0.7-0.9), and (3) high accuracy (>0.9) (Wakie et al., 2014). Based on the final models, we created binary maps using the suitability that gives the highest values of TSS threshold, in which the sum of sensitivity and specificity is maximum. The binary maps for each species in each climatic scenario were cut for the Brazil boundaries. Only areas of high suitability appear on binary maps.
Utilizing an ensemble method, we harnessed consensus maps to depict habitat suitability for both food plant and pollinator species across various software packages and climate scenarios, a process informed by the principal component analysis (PCA). To merge the three maps pertaining to each species within a given future scenario, we leveraged the functionality of the “Spatial Analyst Tools” followed by “Cell Statistics - Overlay Statistic (Mean)” within the ArcGIS 10.0 software (ESRI, 2019). Subsequently, we subjected all consensus maps to a cutting process, enabling a detailed examination of the repercussions of climate change on the distribution of food plants and pollinators within the entire Brazilian domain. The extent of suitable habitat was calculated individually for each species within each scenario. To ensure the accuracy and reliability of our modeling outcomes, we meticulously scrutinized and refined all results using ArcGIS 10.0 software (ESRI, 2019). Notably, our models strongly underscored the pivotal influence of climatic variables in delineating the limits of species occurrence, meaning that the models generated aptly captured the underlying biological factors governing population persistence within this environment (Searcy and Shaffer, 2016).
3 Results
Considering food plants, model performance experienced robust support from the occurrence data given that mean values for AUC and TSS were high at 0.84 and 0.62, respectively (Table 1). The model performance for pollinating bees was also robust with 0.82 and 0.81 mean values for AUC and TSS, respectively (Table 2). The ecological niche models generated showed that the variables BIO12 (annual precipitation) and elevation had the highest contribution to the current, SSP-3.70, and SSP-5.85 scenarios for all food plant and pollinator species compared to any other predictor. In addition, elevation and BIO19 (coldest quarter precipitation) explained more of the variation in species distribution under the SSP-2.45 scenario.
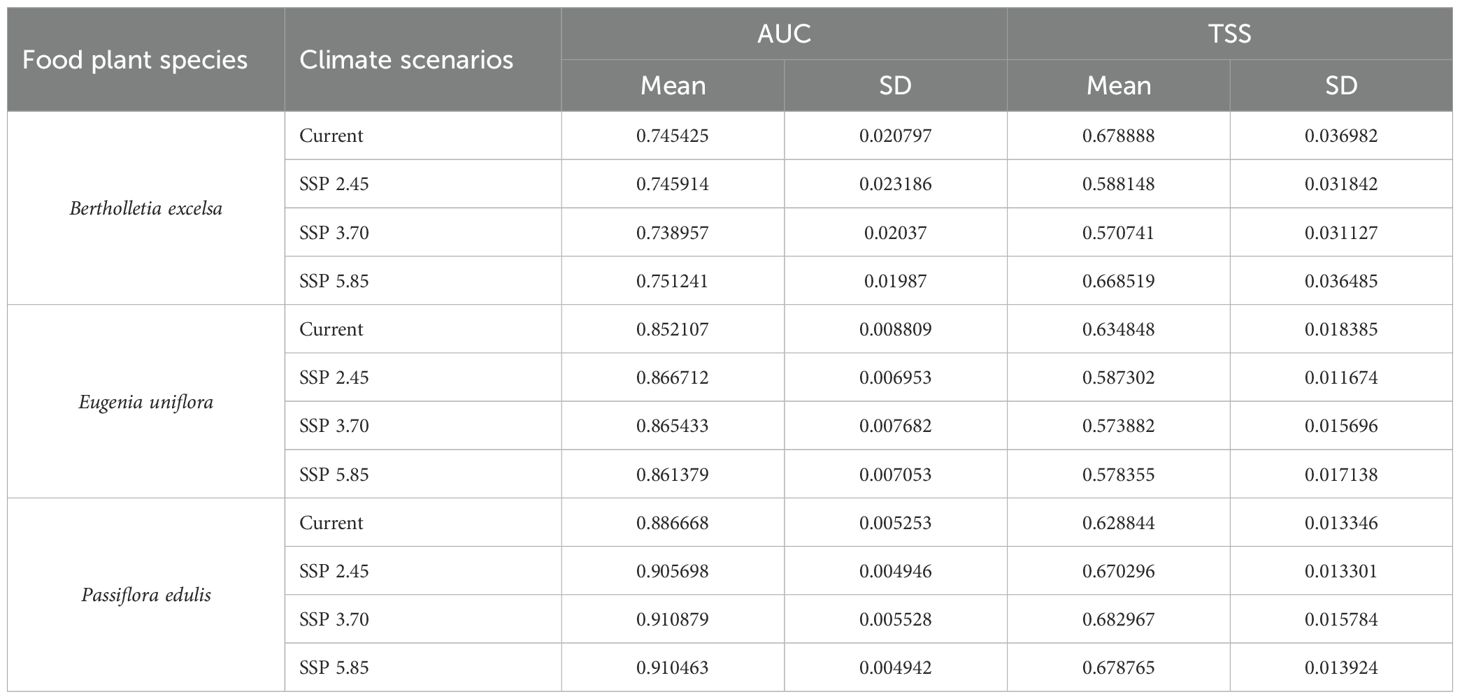
Table 1. Evaluation of species distribution model performance considering the adjustments between the occurrence data of food plant species native to Brazil and climatic scenarios for the current period (averaged for 1970 - 2000) and three future scenarios for 2090 (averaged for 2081 - 2100): SSP-2.45, SSP-3.70, and SSP-5.85.
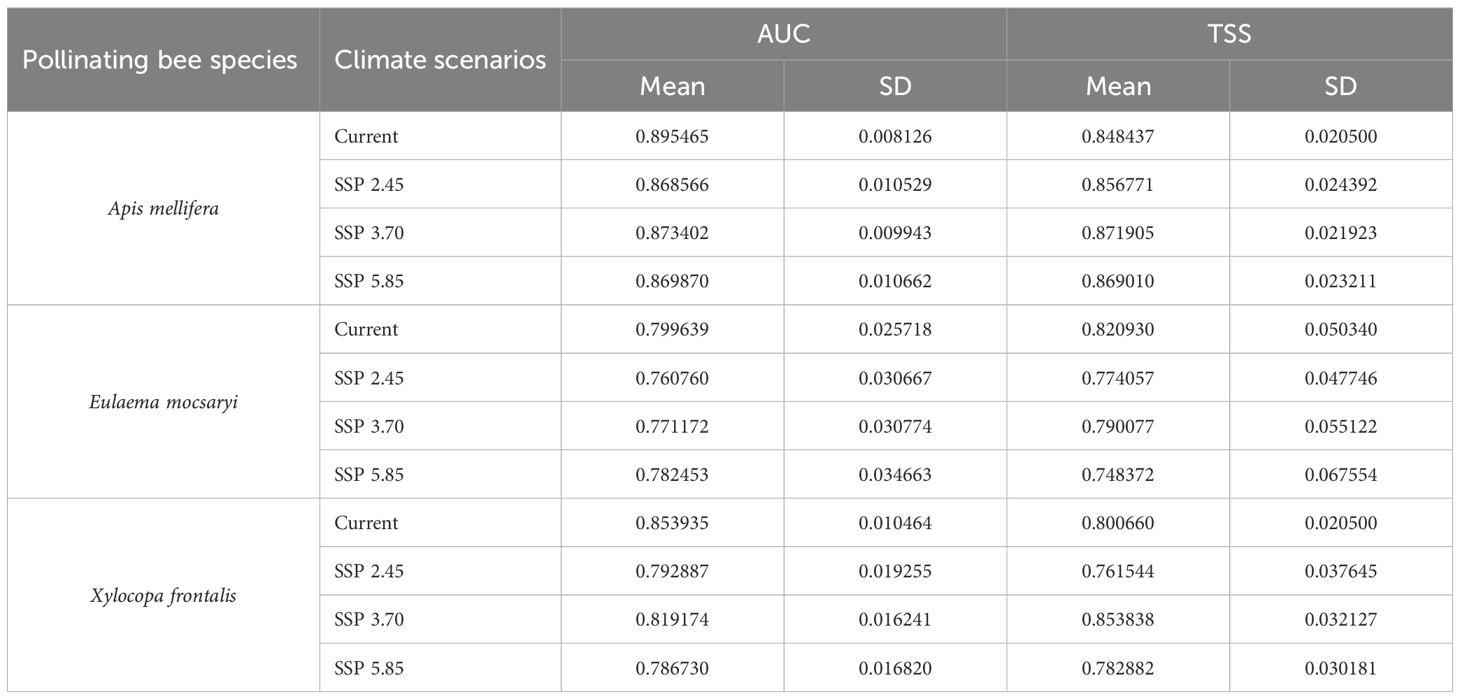
Table 2. Evaluation of species distribution model performance considering the adjustments between the occurrence data of pollinating bee species and climatic scenarios for the current period (averaged for 1970 - 2000) and three future scenarios for 2090 (averaged for 2081 - 2100): SSP-2.45, SSP-3.70, and SSP-5.85.
We observed that climate change was associated with contractions of suitable habitats for all plant and pollinator species under any climate scenario examined here. Comparing the current and any future scenarios, climate change was associated with a reduction in suitable habitat areas for all three food plant species. Specifically, suitable habitats may be reduced by up to 52.5% for Passiflora edulis, 70% for Bertholletia excelsa, and 54.5% for Eugenia uniflora (Figure 1). We documented that reductions in suitable habitat areas were greatest under the SSP-5.85 scenario for P. edulis and B. excelsa (Figure 1), and the SSP-3.70 scenario for E. uniflora (Figure 1). Considering the pollinators, climate change also marked reduced suitable habitat areas for all three pollinator species. Overall, habitats could be reduced by up to 39.6% for Xylocopa frontalis, 51.4% for Eulaema mocsaryi, and 44.1% for Apis mellifera (Figure 2). The highest reductions were under the SSP-2.45 scenario for X. frontalis and E. mocsaryi and the SSP-3.70 scenario for A. mellifera (Figure 2).
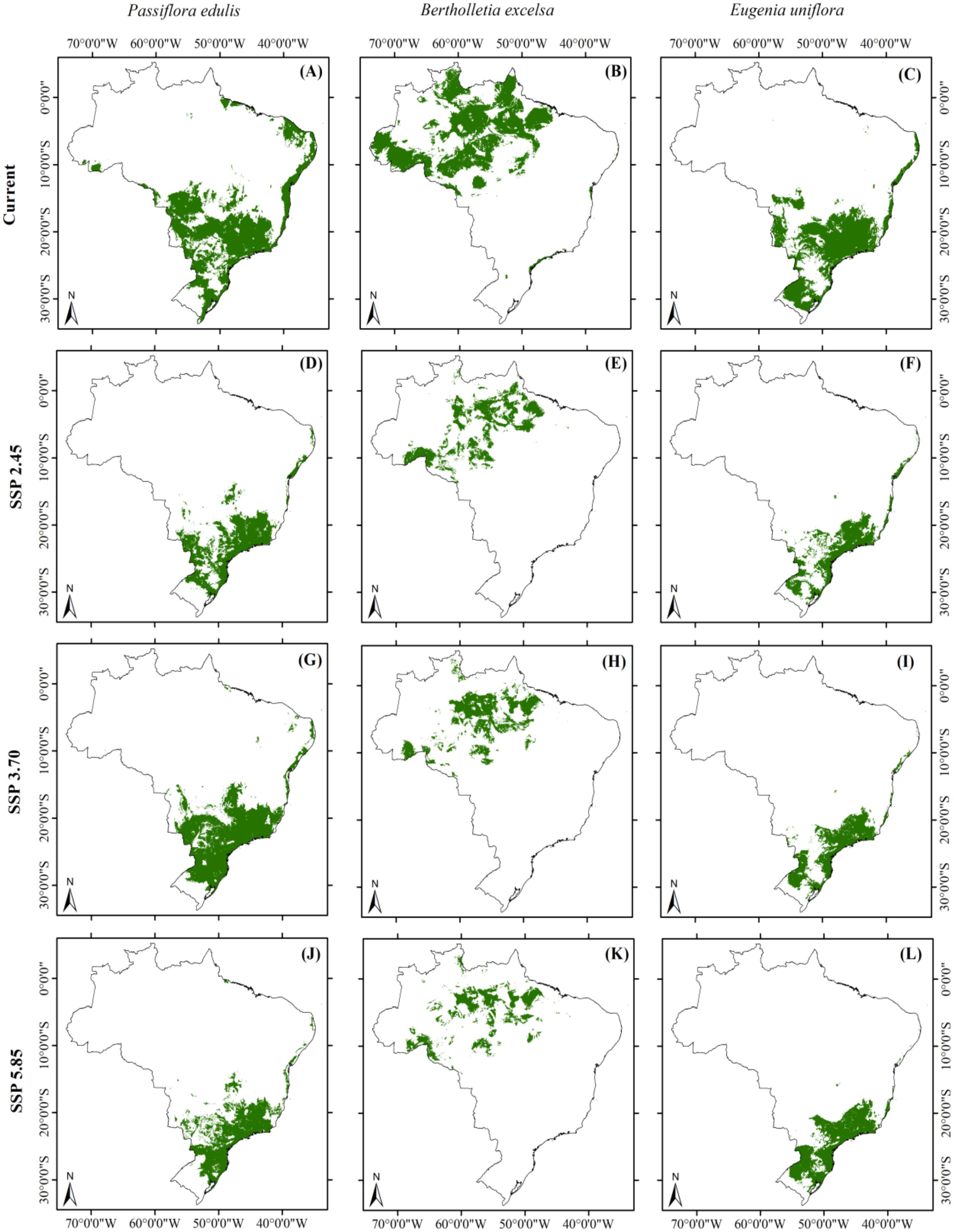
Figure 1. Geographic projection of suitable habitat areas for food plant species native to Brazil under both one contemporary (A–C) and three future scenarios [SSP-2.45 (D–F), SSP-3.70 (G–I), and SSP-5.85 (J–L)] for 2090 (averaged for 2081-2100).
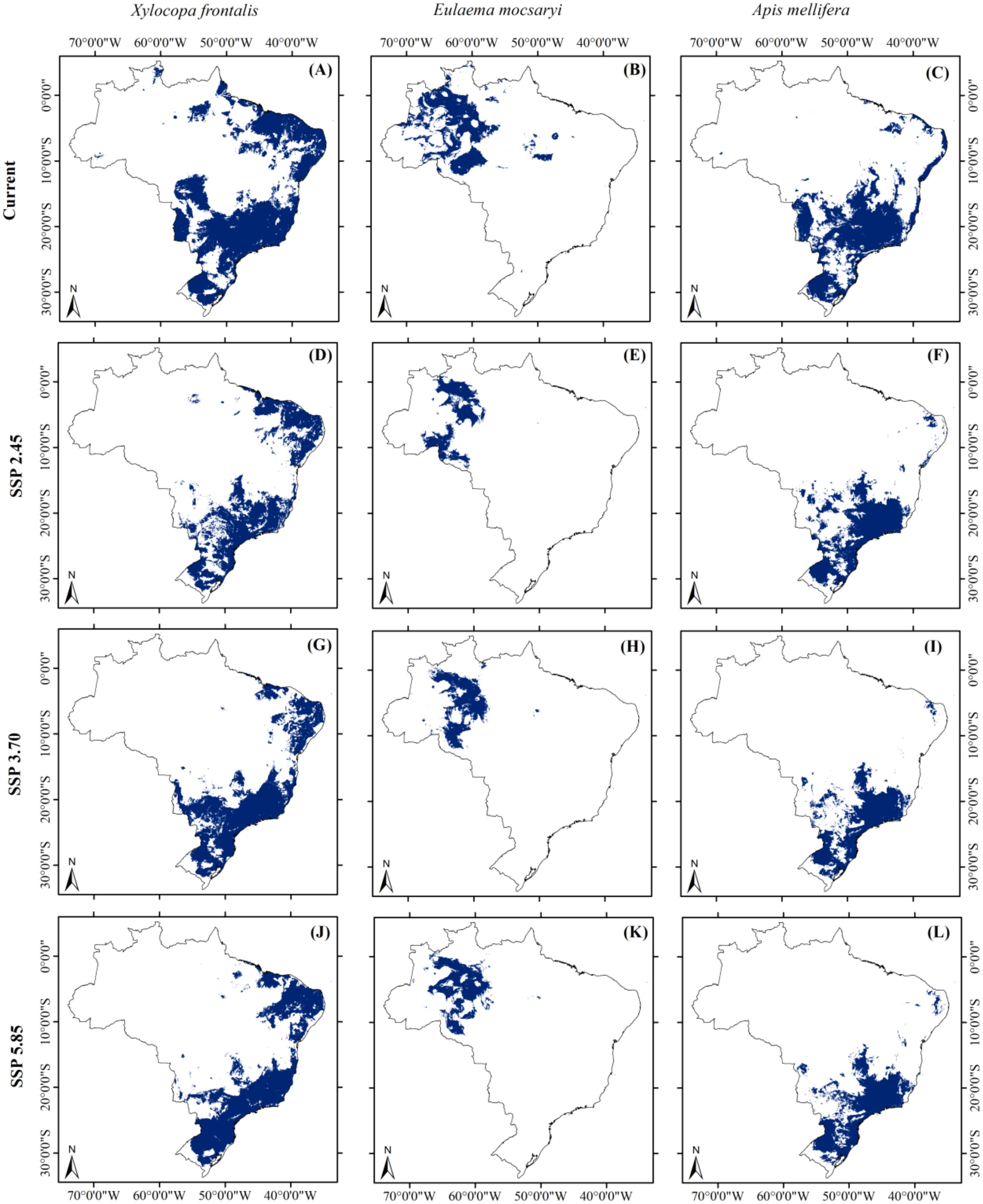
Figure 2. Geographic projection of suitable habitat areas for pollinators under both one contemporary (A–C) and three future scenarios [SSP-2.45 (D–F), SSP-3.70 (G–I), and SSP-5.85 (J–L)] for 2090 (averaged for 2081-2100).
Overall, suitable habitats for P. edulis will likely be across the northern and southern regions of Brazil within the Atlantic forest. For B. excelsa, relictual suitable habitats contracted from the seasonally-dry peripheral to the core portions of the species distribution within the Amazon. For E. uniflora, we observed that suitable habitats also moved towards the central-southern portions of the species range, with species distributions within the Atlantic forest. Regarding pollinators, suitable habitats for X. frontalis moved to the northeastern and southeastern range limits, with marked losses in suitable habitats central-western Brazil, whereas suitable habitats for E. mocsaryi shifted to northwestern Brazil. For honeybees, remaining habitat patches shifted to southeastern and southern Brazil, following marked loss of suitable habitats in the semi-arid northeast.
Overlaying species distribution maps, we observed that climate change is clearly likely to induce marked niche mismatches between food plants and their pollinators. Specifically, suitable habitats for the co-occurrence of plant-pollinator interactions could be reduced by up to 51.5% for P. edulis and X. frontalis, 76% for B. excelsa and E. mocsaryi, and 54% for E. uniflora and A. mellifera (Figure 3). The niche mismatches between the food plants and their pollinating bees will impact the plant-pollinator interactions and, consequently, plant reproduction. These reductions in co-occurrence were most exacerbated under the SSP-2.45 scenario for P. edulis and X. frontalis, and for E. uniflora and A. mellifera. The worst predicted co-occurrence forecast for Brazil-nut trees and their orchid bee pollinators was under the SSP-5.85 scenario (Figure 3). Our findings highlight not only how climate change can impact ecological networks by disrupting plant-pollinator interactions, but also their potential to pose threats to food security by reducing pollination and, consequently, the productivity of pollinator-dependent food plants that essentially rely on pollinators to set fruits and seeds.
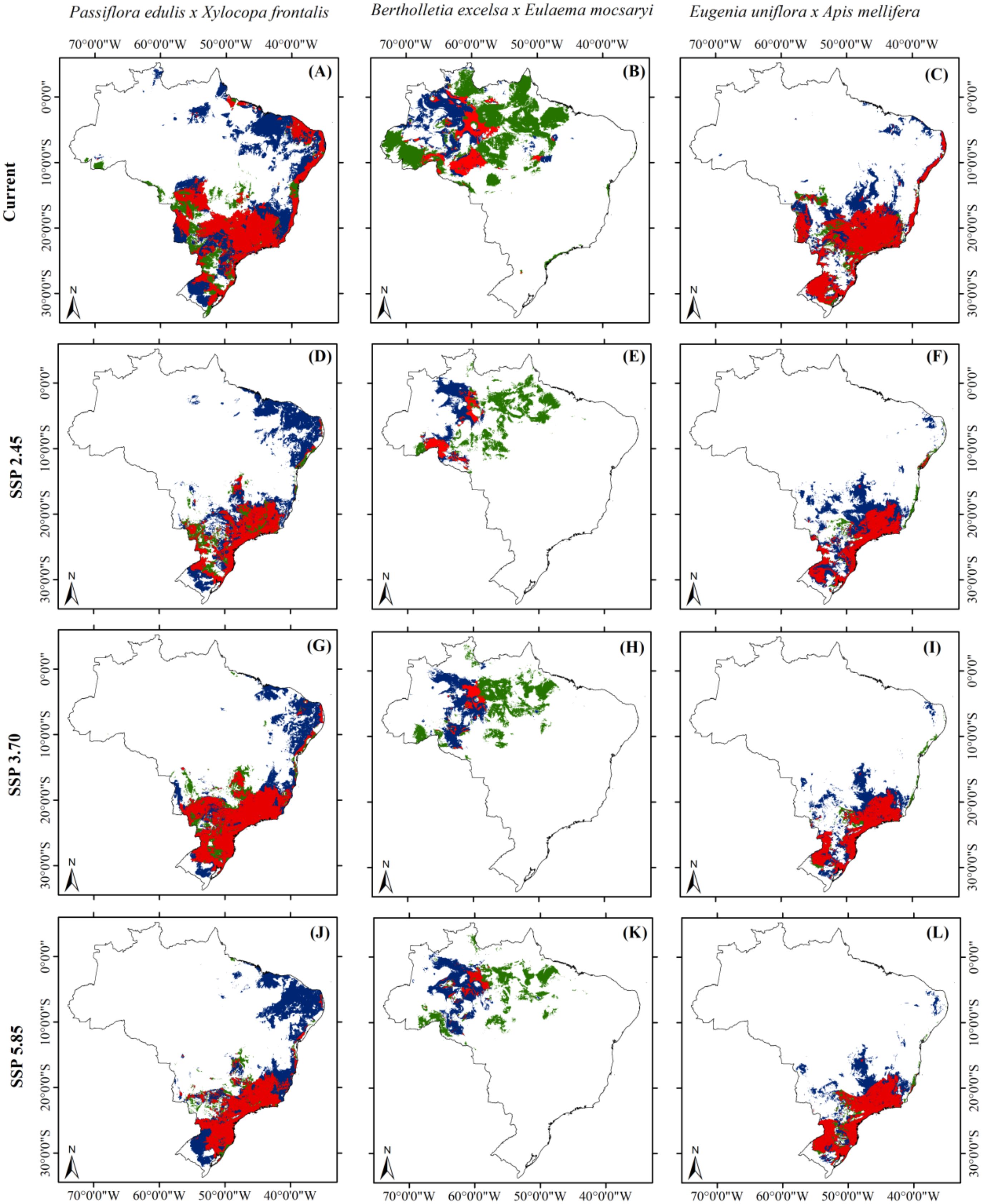
Figure 3. Geographic projection of suitable habitat areas for the co-occurrence of food plant species native to Brazil and their main pollinators under both one contemporary (A–C) and three future scenarios [SSP-2.45 (D–F), SSP-3.70 (G–I), and SSP-5.85 (J–L)] for 2090 (averaged for 2081-2100). Green areas indicate the occurrence of food plants; blue areas indicate the occurrence of pollinators; and red areas indicate the spatial overlap between food plants and pollinators.
4 Discussion
Our results clearly indicate that climate change will likely reduce suitable habitats and alter the spatial distribution for pollinator-dependent food plants native to Brazil and their primary bee pollinators. Our findings are evidenced by the pattern of contractions in occurrence areas, which were most severe under some climate change scenarios, with divergent responses among the species examined here. Furthermore, our findings show that climate change can act as a potential driver of decoupling between each crop and its main pollinating bee, leading to co-occurrence mismatches. Reductions in suitable habitat areas were documented for all food plants and their pollinators under all climate change scenarios, indicating that climate change can potentially affect the occurrence and co-occurrence of plants and their pollinators, which corroborates our hypotheses regardless of the scenario we used. Therefore, shifts in habitat suitability induced by climate change can result in severe spatial mismatches, potentially exacerbating the risks of pollination failure.
Our results reinforce now widely observed and predicted patterns of climate-induced shifts species ranges in terms of suitability, some of which involving potential impacts on pollination (e.g., Giannini et al., 2013, 2017; Settele et al., 2016; Elias et al., 2017; Millard et al., 2023). The effects of climate change have been linked to significant reductions in pollination services (Settele et al., 2016; Sales et al., 2021; Millard et al., 2023). This poses intractable challenges to food production and food security as biotic pollination is a major ecosystem service ensuring crop yields in a large number of food plants (Klein et al., 2007; Giannini et al., 2015; Oliveira et al., 2024). From this perspective, our results are consistent with previously documented effects of climate change on agricultural crops and pollinators in Brazil. Specifically, projected climate change could reduce suitable areas for the occurrence of bees, which pollinate some crops, including tomato, persimmon, and passionfruit (e.g., Giannini et al., 2013, 2017; Elias et al., 2017). In addition, the interaction between agricultural land use and climate change is associated with population declines in insect pollinators, and these impacts are more severe in tropical regions (Millard et al., 2023). Watermelon production is also expected to decline due to climate change when crops are pollinated by managed honeybees, but the opposite was shown for native bee pollinators, reinforcing the importance of the native biota in mitigating climate change (Rader et al., 2013). Some species could therefore respond positively to climate change. In contrast to this study, it has been suggested that Brazil-nut trees could expand their distribution area by up to 6%, depending on the climate change scenario (Sales et al., 2020). However, when interactions with pollinators were examined, declines in pollinator diversity may still lead to a spatial mismatch and threaten the pollination of Brazil-nut tree stands (Sales et al., 2020). Regardless of the climate scenario, the distribution of wild Brazil-nut tree populations across the Brazilian, Peruvian and the Bolivian Amazon is likely to be constrained, not least because of mounting threats from widespread forest degradation through logging and wildfires (C.A. Peres, unpubl. data).
Climate change could be a strong environmental driver altering ecosystem functioning (Malhi and Wright, 2004), often leading to an ecological imbalance that favors generalist species over specialists (e.g., Burkle et al., 2013; Silva et al., 2019). The latter, in turn, have narrower ecological niches and become more susceptible to suitable habitat contraction or undergo local extinction (Blois et al., 2013; Silva et al., 2019). We observed a marked decline in suitable habitats. For Passiflora edulis and Eugenia uniflora, we observed a range contraction in the Atlantic forest region, whereas Bertholletia excelsa succumbs to substantial habitat loss across the Amazon. The Atlantic forest is one of the most susceptible biodiversity hotspots to climate change (Béllard et al., 2014), which is intensified by deforestation in this biome (Scarano and Ceotto, 2015). The Atlantic forest is expected to face pervasive biotic reorganization followed by a savannization process (Joly et al., 2014; Scarano and Ceotto, 2015), particularly in the Brazilian northeast where aridity is a major issue, which could explain the patterns we find for P. edulis and E. uniflora, but also for their pollinators (Xylocopa frontalis and Apis mellifera). Additionally, the risk of pollinator shortages for native crops has been documented in some counties in northern Brazil, although this is mainly concentrated in the northeast and the southeast, with the Atlantic forest and the Caatinga dry forest at higher risk of pollinator shortages (Oliveira et al., 2024). Considering Brazil-nut trees, reduced precipitation and increasing frequency of drought are expected in the eastern and southeast Amazon, the regions most affected by deforestation (Malhi et al., 2008). This is aggravated given that Brazil-nut tree populations have been widely affected by deforestation, habitat fragmentation, wildfires, and overexploitation of their seed crops (Peres et al., 2003; Brandão et al., 2021). Therefore, deforestation is a major factor boosting the effects of climate change on pollinators, which is reinforced by climate projections in the eastern Amazon indicating that 95% of all bee species will face declines in their area of occurrence, affecting crop pollination (Giannini et al., 2020).
Our projections indicate that climate change is a potential driver of niche mismatch between food plants and their pollinators. Brazil is one of the countries at highest risk of crop yield losses due to the impacts of climate change on the abundance of insect pollinators (Millard et al., 2023). The contraction in areas of biotic overlap may therefore represent a severe threat to Brazilian agriculture, given that the tropics will likely experience the highest risks of pollinator loss, which threatens the production of key pollinator-dependent crops, such as cocoa, mango, watermelon, passionfruit, tomato, and coffee (Giannini et al., 2013, 2017; Elias et al., 2017; Millard et al., 2023). As such, our worst-case scenario highlights the severe impacts on ecosystem functioning and food production, with downstream socioeconomic implications. Such negative effects of climate change on the pollination of food plants affect not only biodiversity but also subsistence and cash-crop yields, the national economy, and the livelihoods of local communities. The decline in local food production tends to increase the value of national trade, which may benefit the national economy, but reinforces inequality and rural poverty by eroding per capita purchase power (Cornelsen et al., 2015; Murphy et al., 2022). These effects will be even more intense in Brazil, which is a leading producer and exporter of tropical fruit crops (Oliveira et al., 2023) and one of the countries most vulnerable to climate change (Millard et al., 2023). A decline in pollination interactions induced by climate change heightens food insecurity (Millard et al., 2023). Furthermore, the reduction in crop diversity driven by the expansion of monoculture lands is a key driver of pollinator declines, which in turn could threaten the resilience of ecosystems and agricultural yields (e.g., Aizen et al., 2019; Oliveira et al., 2023). Overall, the combined impacts of land-use and climate change reduce long-term agricultural sustainability by increasing the likelihood of disrupted plant-pollinator interactions, which can ultimately lead to food shortages. Accordingly, the loss of pollinators will increase trade risks for major food importers such as China, western Europe and the USA, as the lack of pollinators will disrupt agricultural yields of pollinator-dependent cash crops, such as coffee and cocoa (Millard et al., 2023).
Therefore, decision-makers need to focus on developing public policies that prioritize climate change mitigation and encourage sustainable agricultural practices to protect key mutualistic interactions that benefit human societies, such as pollination. Resilient ecosystems not only benefit plants and their pollinators but are also vital in ensuring agricultural productivity and food security (IPBES, 2016). Thus, we need to rethink our agricultural practices, implementing biodiversity conservation as a crucial pillar to mitigate the ecological challenges imposed by climate change, while also ensuring resilience and food sovereignty. To achieve this, policy makers should urgently include the remaining areas where key mutualists are expected to co-occur into action plans for conservation, while maintaining species abundance and richness, and building more sustainable agricultural production pathways (Altieri and Nicholls, 2008, 2017; Garibaldi et al., 2016; Nicholls and Altieri, 2018; Oliveira et al., 2023). Furthermore, sustainable agriculture practices should be adaptable to a variety of production methods and conditions, and consider the entire agricultural system in their design (Muhie, 2022 and references therein). These practices can include climate-smart agriculture (a relatively new approach that ensures improved agricultural practices while lowering greenhouse gas emissions to assist underprivileged people in increasing agricultural productivity and income), organic farming (which promotes resource sustainability, environmental protection, animal welfare, food quality and safety, social justice, and market-driven payments for internalized costs), sustainable intensification (which increases crop yields without threatening the environment or converting natural areas into farmland), regenerative agriculture (a food and agricultural system focused on rehabilitation and preservation), and integrated nutrient management (a technique for safely disposing of agricultural wastes by incorporating inorganic and organic fertilizers in a balanced and integrated manner to preserve soil fertility and give plants the right amount of nutrients). These practices can achieve the goals of life on land and zero-hunger of the Sustainable Development Goals, especially in major emergent agricultural economies such as Brazil.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
WO: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. OC: Conceptualization, Data curation, Formal analysis, Methodology, Supervision, Validation, Visualization, Writing – review & editing. JS: Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – review & editing. MT: Funding acquisition, Validation, Visualization, Writing – review & editing. CP: Writing – review & editing, Supervision. AL: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) through PhD studentship #141954/2020-5 awarded to WO and #175071/2023-3 awarded to JS; PQ #308832/2014-0 and #309505/2018-6 awarded to AL; the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (for all authors grant code: 001; postdoc fellowship awarded to OC-N, grant number: APQ-0789-2.05/16), and the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) (postdoc fellowship to OC-N, grant number: BCT-34 0208-2.05/17).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frbee.2025.1510451/full#supplementary-material
Supplementary Table 1 | Download information of occurrence data of three food plants native to Brazil and two pollinators obtained from the GBIF repository (source: The Global Biodiversity Information Facility 2021 - GBIF; https://www.gbif.org).
References
Aizen M. A., Aguiar S., Biesmeijer J. C., Garibaldi L. A., Inouye D. W., Jung C., et al. (2019). Global agricultural productivity in threatened by increasing pollinator dependence without a parallel increase in crop diversification. Global Change Biol. 25, 3516–3527.
Albacete S., Sancho G., Azpiazu C., Rodrigo A., Molowny-Horas R., Sgolastra F., et al. (2023). Bees exposed to climate change are more sensitive to pesticides. Glob. Change Biol. 29, 6248–6260. doi: 10.1111/gcb.16928
Altieri M. A. and Nicholls C. I. (2008). Scaling up agroecological approaches for food sovereignty in Latin America. Development 51, 472–480. doi: 10.1057/dev.2008.68
Altieri M. A. and Nicholls C. I. (2017). Agroecology: a brief account of its origins and currents of thought in Latin America. Agroecol Sustain. Food Syst. 41, 231–237. doi: 10.1080/21683565.2017.1287147
Andrade A. F. A., Velazco S. J. E., and de Marco Júnior P. (2020). ENMTML: An r package for a straightforward construction of complex ecological niche models. Environ. Model. Software 125, 104615.
Barbet-Massin M., Jiguet F., Albert C. H., and Thuiller W. (2012). Selecting pseudo-absences for species distribution models: how, where and how many? Methods Ecol. Evol. 3, 327–338. doi: 10.1111/j.2041-210X.2011.00172.x
Becher M. A., Osborne J. L., Thorbek P., Kennedy P. J., and Grimm V. (2013). Towards a systems approach for understanding honeybee decline: a stocktaking and synthesis of existing models. J. Appl. Ecol. 50, 868–880. doi: 10.1111/1365-2664.12112
Béllard C., Leclerc C., Leroy B., Bakkenes M., Veloz S., Thuiller W., et al. (2014). Vulnerability of biodiversity hotspots to global change. Glob. Ecol. Biogeogr. 23, 1376–1386. doi: 10.1111/geb.12228
Blois J. L., Zarnetske P. L., Fitzpatrick M. C., and Finnegan S. (2013). Climate change and the past, present, and future of biotic interactions. Science 341, 499–504. doi: 10.1126/science.1237184
Botkin D. B., Saxe H., Araujo M. B., Betts R., Bradshaw R. H. W., Cedhagen T., et al. (2007). Forecasting the effects of global warming on biodiversity. Bioscience 57, 227–236. doi: 10.1641/B570306
BPBES/REBIPP (Brazilian Platform on Biodiversity and Ecosystem Services/Rede Brasileira de Interações Planta-Polinizador) (2019). Relatório temático sobre Polinizacão, Polinizadores e Producão de Alimentos no Brasil. São Paulo: Editora Cubo. 184.
Brandão D. O., Barata L. E. S., Nobre I., and Nobre C. A. (2021). The effects of Amazon deforestation on non-timber forest products. Reg Environ. Change 21, 122. doi: 10.1007/s10113-021-01836-5
Burkle L. A., Marlin J. C., and Knight T. M. (2013). Plant-pollinator interactions over 120 years: Loss of species, co-occurrence, and function. Science 339, 1611–1615. doi: 10.1126/science.1232728
Cai I. S., Foujols M. A., Gardoll S., Gastineau G., Ghattas J., Grandpeix J. Y., et al. (2020). Climate impacts of the el niño – southern oscillation on south america. Nat. Rev. Earth Environ. 1, 215–231. doi: 10.1038/s43017-020-0040-3
Cavalcante A. M. B., Duarte A. S., and Ometto J. P. H. B. (2020). Modelling the potential distribution of Epiphyllum phyllanthus (L.) Haw. under future climate scenarios in the Caatinga biome. An Acad. Bras. Cienc. 92, e20180836. doi: 10.1590/0001-3765202020180836
Cavaltante M. C., Galetto L., Maués M. M., Pacheco-Filho A. J. S., Bomfim I. G. A., and Freitas B. M. (2018). Nectar production dynamics and daily pattern of pollinator visits in Brazil nut (Bertholletia excelsa Bonpl.) plantations in Central Amazon: implications for fruit production. Apidologie 49, 505–516. doi: 10.1007/s13592-018-0578-y
Chen I.-C., Hill J. K., Ohlemuller R., Roy D. B., and Thomas C. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026. doi: 10.1126/science.1206432
Cohen J. M., Lajeunesse M. J., and Rohr J. R. (2018). A global synthesis of animal phenological responses to climate change. Nat. Clim. Change 8, 224–228. doi: 10.1038/s41558-018-0067-3
Cornelsen L., Green R., Turner R., Dangour A. D., Shankar B., Mazzocchi M., et al. (2015). What happens to patterns of food consumption when food prices change? evidence from a systematic review and meta-analysis of food price elasticities globally. Health Economics 24, 1548–1559.
Costanza R., de Groot R., Sutton P., van der Ploeg S., Anderson S. J., Kubiszewski I., et al. (2014). Changes in the global value of ecosystem services. Global Environ. Change 26, 152–158.
De Marco P. and Nóbrega C. C. (2018). Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PloS One 0202403, e0202403. doi: 10.1371/journal.pone.0202403
Dicks L. V., Viana B., Bommarco R., Brosi B., Arizmendi M. C., Cunningham S. A., et al. (2016). Ten policies for pollinators. Science 354, 14–15. doi: 10.1126/science.aai9226
Diniz M. E. R. and Buschini M. L. T. (2016). Diversity of flower visiting bees of Eugenia uniflora L. (Myrtaceae) in fragments of Atlantic Forest in South Brazil. Sociobiology 63, 982–990. doi: 10.13102/sociobiology.v63i3.982
Drake J. M., Randin C., and Guisan A. (2006). Modelling ecological niches with support vector machines. J. Appl. Ecol. 43, 424–432. doi: 10.1111/j.1365-2664.2006.01141.x
Elias M. A. S., Borges F. J. A., Bergamini L. L., Franceschinelli E. V., and Sujii E. R. (2017). Climate change threatens pollination services in tomato crops in Brazil. Agric. Ecosyst. Environ. 239, 257–264. doi: 10.1016/j.agee.2017.01.026
Eyring V., Bony S., Meehl G. A., Senior C. A., Stevens B., Stouffer R. J., et al. (2016). Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model. Devt 9, 1837–1958. doi: 10.5194/gmd-9-1937-2016
FAO, IFAD, UNICEF, WFP, and WHO (2021). “The state of food security and nutrition in the world 2021,” in Transforming food systems for food security, improved nutrition and affordable healthy diets for all (Rome: FAO).
Ferreira R. B., Parreira M. R., and Nabout J. C. (2021). The impact of global climate change on the number and replacement of provisioning ecosystem services of Brazilian Cerrado plants. Environ. Monit. Assess 193, 731. doi: 10.1007/s10661-021-09529-6
Fick S. E. and Hijmans R. J. (2017). Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4301–4315. doi: 10.1002/joc.5086
Fitter A. H. and Fitter R. S. R. (2002). Rapid changes in flowering time in british plants. Science 296, 1689–1691. doi: 10.1126/science.1071617
Fonseca M. G., Aragão L. E. O. C., Lima A., Shimabukuro Y. E., Arai E., and Anderson L. O. (2015). Modelling fire probability in the Brazilian Amazon using the maximum entropy method. Int. J. Wildland Fire 25, 955–969. doi: 10.1071/WF15216
Fuentes-Castillo T., Hernández H. J., and Pliscoff P. (2020). Hotspots and ecoregion vulnerability driven by climate change velocity in southern south america. Regional Environ. Change 20, 27.
Garibaldi L. A., Carvalheiro L. G., Vaissière B. E., et al. (2016). Mutually beneficial pollinator diversity and crop yield outcomes in small and large farms. Science 351, 388–391.
Garibaldi L. A., Steffan-Dewenter I., Winfree R., Aizen M. A., Bommarco R., Cunningham S. A., et al. (2013). Wild pollinators enhance fruit set of crops regardless of honeybee abundance. Science 340, 1608–1611. doi: 10.1126/science.1230200
GBIF (2021) GBIF home page. Available at: https://www.gbif.org/pt/citation-guidelineshttps://www.gbif.org.
Giannini T. C., Acosta A. L., Silva C. I., Oliveira P. E. A. M., Imperatriz-Fonseca V. L., and Saraiva A. M. (2013). Identifying the areas to preserve passion fruit pollination service in Brazilian Tropical Savannas under climate change. Agr Ecosyst. Environ. 171, 39–46. doi: 10.1016/j.agee.2013.03.003
Giannini T. C., Boff S., Cordeiro G. D., Cartolano E. A., Veiga A. K., Imperatriz-Fonseca V. L., et al. (2015). Crop pollinators in Brazil: a review of reported interactions. Apidologie 46, 209–223. doi: 10.1007/s13592-014-0316-z
Giannini T. C., Costa W. F., Borges R. C., Miranda L., Costa C. P. W., Saraiva A. M., et al. (2020). Climate change in the Eastern Amazon: crop-pollinator and occurrence-restricted bees are potentially more affected. Reg Environ. Change 20, 9. doi: 10.1007/s10113-020-01611-y
Giannini T. C., Costa W. F., Cordeiro G. D., Imperatriz-Fonseca V. L., Saraiva A. M., Biesmeijer J., et al. (2017). Projected climate change threatens pollinators and crop production in Brazil. PloS One 12, e0182274. doi: 10.1371/journal.pone.0182274
Goulson D., Nicholls E., Botias C., and Rotheray E. L. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, 1255957. doi: 10.1126/science.1255957
Houghton J. T., Ding Y., Griggs D. J., Noguer M., Winden P. J., and Dai X. (2001). Climate Change 2001: The Scientific Basis. Contributions of Working Group 1 to the Third Assessment Report of the Intergovernmental Panel on Climate Change (Cambridge: Cambridge University Press).
IPBES - Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (2016). Summary for policymakers of the assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on pollinators, pollination and food production (Germany: Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services).
IPCC - Intergovernmental Panel on Climate Change (2014). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Core Writing Team, Pachauri R. K., and Meyer L. A. (Geneva, Switzerland: IPCC).
IPCC- Intergovernmental Panel on Climate Change (2021). Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change (Cambridge: Cambridge University Press).
Joly C., Metzger J. P., and Tabarelli M. (2014). Experiences from the Brazilian Atlantic Forest: ecological findings and conservation initiatives. New Phytol. 204, 459–473. doi: 10.1111/nph.12989
Kerr J. T., Pindar A., Galpern P., Packer L., Potts S. G., Roberts S. M., et al. (2015). Climate change impacts on bumblebees converge across continents. Science 349, 177–180. doi: 10.1126/science.aaa7031
Kharouba H. M., Ehrlén J., Gelman A., Bolmgren K., Allen J. M., Travers S. E., et al. (2018). Global shifts in the phenological synchrony of species interactions over recent decades. Proc. Natl. Acad. Sci. U.S.A. 115, 5211–5216. doi: 10.1073/pnas.1714511115
Kharouba H. M. and Vellend M. (2015). Flowering time of butterfly nectar food plants is more sensitive to temperature than the timing of butterfly adult flight. J. Anim Ecol. 84, 1311–1321. doi: 10.1111/1365-2656.12373
Klein A. M., Vaissière B. E., Cane J. H., Steffan-Dewenter I., Cunningham S. A., Kremen C., et al. (2007). Importance of pollinators in changing landscapes for world crops. Proc. R. Soc B: Biol. Sci. 274, 303–313. doi: 10.1098/rspb.2006.3721
Lima V. P., Lima R. A. F., Joner F., D’Orangeville L., Raes N., Siddique I., et al. (2023). Integrating climate change into agroforestry conservation: A case study on native plant species in the Brazilian Atlantic Forest. J. Appl. Ecol. 60, 1977–1994. doi: 10.1111/1365-2664.14464
Lima V. P. and Marchioro C. A. (2021). Brazilian stingless bees are threatened by habitat conversion and climate change. Reg Environ. Change 21, 14. doi: 10.1007/s10113-021-01751-9
Loarie S. R., Carter B. E., Hayhoe K., McMahon S., Moe R., Knight C. A., et al. (2008). Climate change and the future of California’s endemic flora. PloS One 3, 2502. doi: 10.1371/journal.pone.0002502
Lopes A. V., Porto R. G., Cruz-Neto O., Peres C. A., Viana B. F., Giannini T. C., et al. (2021). Neglected diversity of crop pollinators: Lessons from the world’s largest tropical country. Perspect. Ecol. Conserv. 19, 500–504. doi: 10.1016/j.pecon.2021.06.004
Maitner B. S., Boyle B., Casler N., Condit R., Donoghue J. II, Durán S. M., et al. (2018). The BIEN R package: A toll to access the Botanical Information and Ecology Network (BIEN) database. Methods Ecol. Evol. 9, 373–379. doi: 10.1111/2041-210X.12861
Malhi Y., Roberts J. T., Betts R. A., Killeen T. J., Li W., and Nobre C. A. (2008). Climate change, deforestation, and the fate of the amazon. Science 319, 169–172. doi: 10.1126/science.114696
Malhi Y. and Wright J. (2004). Spatial patterns and recent trends in the climate of tropical rainforest regions. Phil Trans. R. Soc Lond. 365, 311–329. doi: 10.1098/rstb.2003.1433
Maluf R. P., Alzate-Marin A. L., Silva C. C., Pansarin L. M., Bonifácio-Anacleto F., Schuster I., et al. (2022). Warming and soil water availability affect plant-flower visitor interactions for Stylosanthes capitata, a tropical forage legume. Sci. Total Environ. 15, 152982. doi: 10.1016/j.scitotenv.2022.152982
Marengo J. A., Torres R. R., and Alves L. M. (2017). Drought in Northeast Brazil – past, present and future. Theor. Appl. Climatol. 129, 1189–1200. doi: 10.1007/s00704-016-1840-8
Memmott J., Craze P. G., Waser N. M., and Price M. V. (2007). Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 10, 710–717.
Menéndez-Guerrero P. A., Green D. M., and Davies J. (2020). Climate change and the future restructuring of Neotropical anuran biodiversity. Ecography 43, 222–235. doi: 10.1111/ecog.04510
Millard J., Outhwaite C. L., Ceauşu S., Carvalheiro L. G., Silva e Silva F. D., Dicks L. V., et al. (2023). Key tropical crops at risk from pollinator loss due to climate change and land use. Sci. Adv. 9, eadh075. doi: 10.1126/sciadv.adh0756
Muhie S. H. (2022). Novel approaches and practices to sustainable agriculture. J. Agric. Food Res. 10, 100446. doi: 10.1016/j.jafr.2022.100446
Murphy J. T., Breeze T. D., Willcox B., Kavanagh S., and Stout J. C. (2022). Globalisation and pollinators: pollinator declines are an economic threat to global food systems. People Nat. 4, 773–785. doi: 10.1002/pan3.10314
Nicholls C. I. and Altieri M. A. (2018). Pathways for the amplification of agroecology. Agroecology Sustain. Food System 42, 1170–1193.s.
Novais S. M. A., Nunes C. A., Santos N. B., D`Amico A. R., Fernandes G. W., Quesada M., et al. (2018). Effects of a possible pollinator crisis on food crop production in Brazil. PloS One 13, e0197396. doi: 10.1371/journal.pone.0197396
Oliveira W., Colares L. F., Porto R. G., Viana B. F., Tabarelli M., and Lopes A. V. (2024). Food plants in Brazil: Origin, economic value of pollination and pollinator shortage risk. Sci. Total Environ. 912, 169147. doi: 10.1016/j.scitotenv.2023.169147
Oliveira W., Cruz-Neto O., Tabarelli M., Galetto L., Peres C. A., and Lopes A. V. (2023). Markedly declining reproductive functional diversity of food plants in the world’s largest tropical country despite rapid cropland expansion. Agric. Ecosyst. Environ. 357, 108673. doi: 10.1016/j.agee.2023.108673
Ollerton J., Winfree R., and Tarrant S. (2011). How many flowering plants are pollinated by animals? Oikos 120, 321–326.
Pacifici M., Visconti P., Butchart S. H. M., Watson J. E. M., Cassola F. M., and Rondinini C. (2017). Species’ traits influenced their response to recent climate change. Nat. Clim. Change 7, 205–208. doi: 10.1038/nclimate3223
Pecl G. T., Araújo M. B., Bell J. D., Blanchard J., Bonebrake T. C., Chen I.-C., et al. (2017). Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, eaai9214. doi: 10.1126/science.aai9214
Peres C. A., Baider C., Zuidema P. A., Wadt L. H. O., Kainer K. A., Gomes-Silva D. A. P., et al. (2003). Demographic threats to the sustainability of Brazil nut exploitation. Science 302, 2112–2114. doi: 10.1126/science.1091698
Phillips S. J., Anderson R. P., and Shapire R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. doi: 10.1016/J.ECOLMODEL.2005.03.026
Porto R. G., Almeida R. F., Cruz-Neto O., Tabarelli M., Viana B. F., Peres C. A., et al. (2020). Pollination ecosystem services: A comprehensive review of economic values, research funding and policy action. Food Secur 12, 1425–1442. doi: 10.1007/s12571-020-01043-w
Potts S. G., Biesmeijer J. C., Kremen C., Neumann P., Schweiger O., and Kunin W. E. (2010). Global pollinator declines: trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353. doi: 10.1016/j.tree.2010.01.007
Potts S. G., Imperatriz-Fonseca V., Ngo H. T., Aizen M. A., Biesmeijer J. C., Breeze T. D., et al. (2016). Safeguarding pollinators and their values to human well-being. Nature 540, 220–229. doi: 10.1038/nature20588
Rader R., Reilly J., Bartomeus I., and Winfree R. (2013). Native bees buffer the negative impact of climate warming on honey bee pollination of watermelon crops. Glob. Change Biol. 19, 3103–3110. doi: 10.1111/gcb.12264
Rafferty N. E. (2017). Effects of global change on insect pollinators: multiple drivers lead to novel communities. Curr. Opin. Insect Sci. 23, 22–27.
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org (Accessed November, 2023).
Riahi K., van Vuuren D. P., Kriegler E., Edmonds J., O’Neill B. C., Fujimori S., et al. (2017). The Shared Socioeconomic Pathways and their energy, land use, and greenhouse emissions implication: An overview. Glob. Environ. Change 42, 153–168. doi: 10.1016/j.gloenvcha.2016.05.009
Sales L., Rodrigues L., and Masiero R. (2021). Climate change drives spatial mismatch and threatens the biotic interactions of the Brazil nut. Global Ecol. Biogeography 30, 117–127.
Sahragard H. P., Ajorlo M., and Karami P. (2018). Modeling habitat suitable of range plant species using random forest method in arid mountainous rangelands. J. Mt Sci. 15, 2159–2171. doi: 10.1007/s11629-018-4898-1
Sales L., Rodrigues L., and Masiero R. (2020). Climate change drives spatial mismatch and threatens the biotic interactions of the Brazil nut. Glob. Ecol. Biogeogr. 30, 117–127. doi: 10.1111/geb.13200
Scarano F. R. and Ceotto P. (2015). Brazilian Atlantic forest: impact, vulnerability, and adaptation to climate change. Biodiversity and Conservation 24, 2319–31.
Scheffers B. R., Meester L. D., Bridge T. C. L., Hoffmann A. A., Pandolfi J. M., Corlett R. T., et al. (2016). The broad footprint of climate change from genes to biomes to people. Science 354, 719–732. doi: 10.1126/science.aaf7671
Schleuning M., Neuschulz E. L., Albrecht. J., Bender I. M. A., Bowler D. E., and Dehling D. M. (2020). Trait-based assessments of climate-change impacts on interacting species. Trends Ecol. Evol. 35, 319–328. doi: 10.1016/j.tree.2019.12.010
Schweiger O., Settele J., Kudrna O., Klotz S., and Kühn I. (2008). Climate change can cause spatial mismatch of trophically interacting species. Ecology 89, 3472–3479.
Searcy C. A. and Shaffer H. B. (2016). Do ecological niche models accurately identify climatic determinants of species ranges? Am. Nat. 187, 423–435. doi: 10.1086/685387
Settele J., Bishop J., and Potts S. G. (2016). Climate change impacts on pollination. Nat. Plants 2, 16092. doi: 10.1038/nplants.2016.92
Silva J. L. S., Cruz-Neto O., Peres C. A., Tabarelli M., and Lopes A. V. (2019). Climate change will reduce suitable Caatinga dry forest habitat for endemic plants with disproportionate impacts on specialized reproductive strategies. PloS One 14, e0217028. doi: 10.1371/journal.pone.0217028
Silva J. L. S., Cruz-Neto O., Tabarelli M., Albuquerque U. P., and Lopes A. V. (2022). Climate change will likely threaten areas of suitable habitats for the most relevant medicinal plants native to the Caatinga dry forest. Ethnobiol Conserv. 11, 15. doi: 10.15451/ec2022-06-11.15-1-24
Soroye P., Newbold T., and Kerr J. (2020). Climate change contributes to widespread declines among bumble bees across continents. Science 367, 685–688. doi: 10.1126/science.aax8591
Swart N. C., Cole J. N. S., Kharin V. V., Lazare M., Scinocca J. F., Gillett N. P., et al. (2019). The canadian earth system model version 5 (CanESM5.0.3). Geosci. Model. Dev. 12, 4823–4873. doi: 10.5194/gmd-12-4823-2019
Taheri S., Naimi B., Rahbek C., and Araújo M. B. (2021). Improvements in reports of species redistribution under climate change are required. Sci. Adv. 7, eabe1110. doi: 10.1126/sciadv.abe1110
Tatebe H., Ogura T., Nitta T., Komuro Y., Ogochi K., Takemura T., et al. (2019). Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model. Dev. 12, 2727–2765. doi: 10.5194/gmd-12-2727-2019
Urban M. C. (2015). Accelerating extinction risk from climate change. Science 348, 571–573. doi: 10.1126/science.aaa4984
Urruth L. M., Bassi J. B., and Chemello D. (2022). Policies to encourage agroforestry in the Southern Atlantic Forest. Land Use Policy 112, 105–802. doi: 10.1016/j.landusepol.2021.105802
Valiente-Banuet A., Aizen M. A., Alcántara J. M., Arroyo. J., Cocucci A., Galetti M., et al. (2015). Beyond species loss: the extinction of ecological interactions in a changing world. Funct. Ecol. 29, 299–307. doi: 10.1111/1365-2435.12356
Wakie T. T., Evangelista P. H., Jarnevich C. S., and Laituri M. (2014). Mapping current and potential distribution of non-native prosopis juliflora in the afar region of Ethiopia. PloS One 9, e112854. doi: 10.1371/journal.pone.0112854
Wu T., Yu R., Lu Y., Jie W., Fang Y., Zhang J., et al. (2021). BCC-CSM2-HR: a high-resolution version of the Beijing Climate Center Climate System Model. Geosci. Model. Dev. 14, 2977–3006. doi: 10.5194/gmd-2020-284
Keywords: Apis mellifera, Brazil-nut, food plants, food security, distribution modeling, mismatched mutualism, passion fruit, Pitanga
Citation: Oliveira W, Cruz-Neto O, Silva JLS, Tabarelli M, Peres CA and Lopes AV (2025) Climate change will lead to local extinctions and mismatched range contractions disrupting bee-dependent crop pollination. Front. Bee Sci. 3:1510451. doi: 10.3389/frbee.2025.1510451
Received: 13 October 2024; Accepted: 05 May 2025;
Published: 29 May 2025.
Edited by:
Vera Lucia Imperatriz- Fonseca, University of São Paulo, BrazilReviewed by:
Robert Chlebo, Slovak University of Agriculture, SlovakiaRose Sagwe, Institute of Primate Research, Kenya
Ricardo Caliari Oliveira, Autonomous University of Barcelona, Spain
Copyright © 2025 Oliveira, Cruz-Neto, Silva, Tabarelli, Peres and Lopes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ariadna V. Lopes, YXJpYWRuYS5sb3Blc0B1ZnBlLmJy
†ORCID: Willams Oliveira, orcid.org/0000-0001-8345-7986
Oswaldo Cruz-Neto, orcid.org/0000-0002-6625-7568
Jéssica L. S. Silva, orcid.org/0000-0001-8519-0891
Marcelo Tabarelli, orcid.org/0000-0001-7573-7216
Carlos A. Peres, orcid.org/0000-0002-1588-8765
Ariadna V. Lopes, orcid.org/0000-0001-5750-5913