 Rosario Galarza-Seeber1
Rosario Galarza-Seeber1 Juan D. Latorre1
Juan D. Latorre1 Lisa R. Bielke2
Lisa R. Bielke2 Vivek A. Kuttappan1
Vivek A. Kuttappan1 Amanda D. Wolfenden1
Amanda D. Wolfenden1 Xochitl Hernandez-Velasco3
Xochitl Hernandez-Velasco3 Ruben Merino-Guzman3
Ruben Merino-Guzman3 Jose L. Vicente4
Jose L. Vicente4 Annie Donoghue5David Cross1Billy M. Hargis1
Annie Donoghue5David Cross1Billy M. Hargis1 Guillermo Tellez1*
Guillermo Tellez1*
- 1Department of Poultry Science, University of Arkansas, Fayetteville, AR, USA
- 2Department of Animal Sciences, The Ohio State University, Columbus, OH, USA
- 3Departamento de Medicina y Zootecnia de Aves, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 4Pacific Vet Group-USA, Inc., Fayetteville, AR, USA
- 5Poultry Production and Product Safety Research Unit, USDA Agricultural Research Service, Poultry Science Center, University of Arkansas, Fayetteville, AR, USA
Previous studies conducted in our laboratory have demonstrated that intestinal barrier function can be adversely affected by diet ingredients or feed restriction, resulting in increased intestinal inflammation-associated permeability. Two experiments were conducted in broilers to evaluate the effect of three concentrations of Aflatoxin B1 (AFB1; 2, 1.5, or 1 ppm) on gastrointestinal leakage and liver bacterial translocation (BT). In experiment 1, 240 day-of-hatch male broilers were allocated in two groups, each group had six replicates of 20 chickens (n = 120/group): Control feed or feed + 2 ppm AFB1. In experiment 2, 240 day-of-hatch male broilers were allocated in three groups, each group had five replicates of 16 chickens (n = 80/group): Control feed; feed + 1 ppm AFB1; or feed + 1.5 ppm AFB1. In both experiments, chickens were fed starter (days 1–7) and grower diets (days 8–21) ad libitum and performance parameters were evaluated every week. At day 21, all chicks received an oral gavage dose of FITC-d (4.16 mg/kg) 2.5 h before collecting blood samples to evaluate gastrointestinal leakage of FITC-d. In experiment 2, a hematologic analysis was also performed. Liver sections were aseptically collected and cultured using TSA plates to determine BT. Cecal contents were collected to determine total colony-forming units per gram of Gram-negative bacteria, lactic acid bacteria (LAB), or anaerobes by plating on selective media. In experiment 2, liver, spleen, and bursa of Fabricius were removed to determine organ weight ratio, and also intestinal samples were obtained for morphometric analysis. Performance parameters, organ weight ratio, and morphometric measurements were significantly different between Control and AFB1 groups in both experiments. Gut leakage of FITC-d was not affected by the three concentrations of AFB1 evaluated (P > 0.05). Interestingly, a significant reduction in BT was observed in chickens that received 2 and 1 ppm AFB1. An increase (P < 0.05) in total aerobic bacteria, total Gram negatives, and total LAB were observed in chickens fed with 2 and 1.5 ppm of AFB1 when compared with Control and 1 ppm chickens. The integrity of gut epithelial barrier was not compromised after exposure to the mycotoxin.
Introduction
In the winter of 1959, the British cargo ship Rosetti, unloaded a shipment of peanut meal from Brazil to England, which was utilized as a protein supplement in the diets of poultry and other domestic animals. By summer of 1960, an outbreak of an unknown disease killed several species of poultry including turkeys, ducklings, and pheasants. In all, 500 cases were reported involving the deaths of more than 100,000 turkeys. This was the first report of Turkey “X” Disease (1, 2). Exhaustive research led to the discovery of aflatoxins, secondary metabolites of Aspergillus flavus and Aspergillus parasiticus, as the etiological agents and the development of mycotoxicology (3–5). More recent studies demonstrated that aflatoxins are potent carcinogenic compounds (6–11). About 14 different types of aflatoxins are produced in nature (10, 12), but aflatoxin B1 (AFB1) produced by A. flavus and A. parasiticus is considered the most toxic (13, 14). In spite of 55 years of continuous research on aflatoxins, several areas of aflatoxicosis remain yet to be investigated. It is particularly interesting that studies on poultry aflatoxicosis have not kept pace with the research in mammals, and there still exists an incomplete description of aflatoxicosis in avian species, especially when searching for scientific publications related to the effect(s) of aflatoxins on the gastrointestinal tract (GIT).
The GIT is the first organ coming into contact with mycotoxins from the diet and should be expected to be affected by AFB1 with greater potency as compared to other organs. Nevertheless, literature regarding the effects of AFB1 on the GIT is particularly confusing. Few researchers have look at morphometric changes following dietary administration of aflatoxins in chickens, turkeys, and ducks, but results from those studies contradict each other, particularly when looking at villi high and villi to crypt ratio (10, 11, 15–20). Similarly, contradictive results arise from the effects of AFB1 on digestibility of amino acids, energy utilization, and absorption of macronutrients (18, 20–27).
Aflatoxins are absorbed very quickly into the blood from the GIT, followed by an extensive transformation into metabolites primarily in the liver (9, 28, 29). Contrary to the studies on mucosal damage and nutrient absorption caused by AFB1, there is an universal agreement that beside the carcinogenic and hepatotoxic effects on the liver, dietary aflatoxins reduce weight gain, feed intake (FI), increase feed conversion ratio (FCR), and are immunosuppressive (12, 30, 31).
Today, only a few reports could be found in databanks, in which the issue of barrier function and intestinal permeability has been reported. From recent studies by Yunus et al. (19) in broilers, it has been suggested that the absorptive surface of small intestine declines during a chronic exposure to low levels of AFB1. However, in that study, broilers compensated for the reduced absorptive surface by increasing the length of the small intestine (19). In the second study, transepithelial electrical resistance (TEER), used as an important indicator of barrier function of intestinal epithelial cells (IEC), showed that AFB1 was only moderately affect during acute exposure to the toxin (10). As far as we can tell, the only study of the effect of AFB1 on possible damage to tight junctions (TJs) was performed by Caloni et al. (32) who demonstrated that AFB1 does not affect the integrity of TJ proteins or barrier damage in vitro.
We have previously shown that intestinal barrier function can be adversely affected by poorly digested diets, feed restriction, or dexamethasone resulting in increased intestinal inflammation-associated permeability in poultry (33–36). The purpose of the present investigation was to evaluate the effect of three doses of aflatoxin B1 on growth, physiological parameters, and gut permeability in broiler chickens.
Materials and Methods
Animal Source, Diets, and Experimental Design
Two experiments were conducted several weeks apart using two hundred and forty 1-day-old male broiler chicks (Cobb-Vantress, Silom Springs, AR, USA) raised in floor pens. Unmedicated corn-soybean-based broiler starter and medicated (with coccidiostat) corn-soybean-based broiler grower diets were prepared according to the broiler’s recommendations (37). Experiments were conducted to evaluate the effect of three concentrations of AFB1 (2 ppm in experiment 1 and 1.5 or 1 ppm in experiment 2) on systemic fluorescein isothiocyanate-dextran (FITC-d; 3–5 kDa) levels and liver bacterial translocation (BT) as indicators of increased gut epithelial leakage. AFB1 was provided by Dr. George E. Rottinghaus, Veterinary Medical Diagnostic Laboratory, University of Missouri, Columbia, MO, USA. AFB1 was produced through the fermentation of rice and the aflatoxin content was measured by spectrophotometric analysis. The aflatoxin within the rice powder consisted of 74.62% AFB1, 22.38% AFG1, 2.48% AFB2, and 0.49% AFG2, based on total aflatoxin in the rice powder. Diets containing AFB1 were analyzed, and the presence of parent AF was confirmed by high-performance liquid chromatography with fluorescence detection (HPLC-FLD) method by using a Romer Derivatization Unit (Romer Labs, Inc., MO, USA). AFB1 was added to the diets and mixed thoroughly in a graded sequence to specified concentrations. The birds were given diets with or without supplemental AFB1 and water ad libitum. All animal handling procedures were in compliance with the Institutional Animal Care and Use Committee at the University of Arkansas. In experiment 1, broilers were allocated randomly to two groups, each group had six replicates of 20 chickens (n = 120/group): Control feed or feed + 2 ppm AFB1. In experiment 2, broilers were allocated randomly to three groups, each group had five replicates of 16 chickens (n = 80/group): Control feed; feed + 1 ppm AFB1; or feed + 1.5 ppm AFB1. In both experiments, chickens were fed starter (days 1–7) and grower diet (days 8–21) ad libitum until the end of the experiment at day 21. In each experiment, each pen was used as a replicate and also as an experimental unit per treatment to evaluate body weight (BW), body weight gain (BWG), FI, and FCR. These growth performance parameters were obtained every week. At the end of experiment 2, blood samples were collected from the wing vein into tubes with heparin as anticoagulant for differential cell counts. In both experiments, 21-day-old chickens received an oral gavage dose of FITC-d (4.16 mg/kg) 2.5 h before collecting blood samples to evaluate passage of FITC-d. Chickens were humanely killed by CO2 asphyxiation. Blood was collected from the femoral vein to obtain serum for FITC-d determination (as described below) and serum clinical chemistry (in experiment 2 only) with a Corning clinical chemistry analyzer (Chiron Corporation, San Jose, CA, USA). Liver sections (n = 12 chickens/treatment) were aseptically collected to determine BT, and cecal contents were collected to determine total colony-forming units per gram of Gram-negative bacteria, lactic acid bacteria (LAB), or anaerobes by plating on a selective media as described below.
Determination of Hematological Parameters
Differential counts of blood samples collected from experiment 2 were determined using a Cell-Dyne 3500 System (Abbott Laboratories, Chicago, IL, USA) that had been standardized for differential counts of poultry blood cells. Hematologic measurements of heparin anticoagulated blood included total numbers of white blood cells (WBC), heterophils, lymphocytes, monocytes, eosinophils, and basophils. Heterophil/lymphocyte ratios (H/L), an indicator of stress in birds (38), were calculated by dividing the number of heterophils in 1 mL of peripheral blood by the number of lymphocytes. Total counts of red blood cells, hemoglobin (HGB), hematocrit (HCT)%, mean corpuscular volume (MVC), and mean corpuscular hemoglobin (MCH) were also determined. Additionally, in experiment 2, liver, spleen, and bursa of Fabricius were removed and cleaned of adherent tissues. The weight of these organs was measured and expressed as percentage of BW (organs weight/final BW) × 100.
Serum Determination of FITC-d
Blood samples were kept at room temperature for 3 h and centrifuged (1,000 × g for 15 min) to separate the serum from the red blood cells. FITC-d levels of undiluted serum were measured at excitation wavelength of 485 nm and emission wavelength of 528 nm (Synergy HT, Multi-mode microplate reader, BioTek Instruments, Inc., VT, USA). Fluorescence measured was then compared to a standard curve with known FITC-d concentrations. Gut leakage for each bird was reported as microgram of FITC-d/mL of serum (35, 36).
Bacterial Translocation
The number of birds used was based on published studies, in which similar variables were measured (34, 39). Briefly, the right half of the liver was removed from each chicken, collected in sterile bags, homogenized, weighed, and 1:4 wt/vol dilutions were made with sterile 0.9% saline. Ten-fold dilutions of each sample from each group were made in a sterile 96-well Bacti flat bottom plate, and the diluted samples were plated on tryptic soy agar plates (TSA, catalog no. 211822, Becton Dickinson, Sparks, MD, USA).
Determination of Microbial Level in Ceca
Both ceca were aseptically removed, placed into sterile bags, and homogenized. Samples were weighed, and 1:4 wt/vol dilutions were made with sterile 0.9% saline. Ten fold dilutions of each sample from each group were made in a sterile 96-well Bacti flat bottom plate, and the diluted samples were plated on four different culture media to evaluate the total number of LAB in deMan Rogosa Sharpe (Difco™ Lactobacilli MRS Agar VWR Cat. No. 90004-084 Suwanee, GA, USA); total recovered Gram-negative bacteria in MacConkey; total anaerobes in tryptic soy agar with sodium thioglycolate plates (TSA, catalog no. 211822, Becton Dickinson, Sparks, MD, USA); and total yeast in Sabouraud Glucose Agar Base with antibiotics, tetraciclina, and 100-mg sodium benzyl penicillin (HiMedia Laboratories Pvt. Ltd., Mumbai, India).
Histology and Morphometric Analysis of Intestine
Intestinal sections from duodenum (~1-cm section was collected from the middle of the descending duodenum), and ileum (0.5-cm section was obtained from the mid-ileum at Meckel’s diverticulum) were fixed in 10% neutral buffered formalin and embedded in paraffin, sectioned (5 μm thick), set on a glass slide, and stained with hematoxylin and eosin (H&E), and then examined by light microscopy. Photomicrographs of randomly selected fields of each intestinal sample were acquired using a microscope equipped with a Leica DFC450C camera and Leica v.3.8.Software (Leica Application Suite) and used for morphometric analysis. ImageJ 1.47v software (http://rsb.info.nih.gov/ij/) was used for the morphometric measurements of villus length, villus width, and crypt depth. Under a magnification of 2×, 10 villi per bird per section were measured, with a total of five birds per group. Villus length was measured from the top of the villus to the upper part of the lamina propria. Crypt depth was measured from the base upwards to the region of transition between the crypt and villus (40). Villus width was measured at the widest area of each villus, whereas villus:crypt ratio was determined dividing villus height into crypt depth values. Villus surface area was calculated using the formula (2π)(VW/2)(VL), where VW = villus width and VL = villus length (41).
Statistical Analysis
All data were subjected to analysis of variance as an entirely randomized design using the General Linear Models procedure of SAS (42). Data were expressed as mean ± SE. Significant differences among the means were determined by using Duncan’s multiple-range test at P < 0.05.
Results
Performance Parameters
Body weight of chickens fed 2 ppm of AFB1 was not affected in the first week; however, BW was significantly (P < 0.05) reduced by 18.36 and 34.89% during the second and third weeks of age, respectively, when compared with Controls (Table 1). BW gain and FI were also affected by AFB1 consumption with a reduction of 20% for both variables during the second week and 37 and 49%, respectively, in the third week. FCR only showed a significant difference in the third week with an improvement in the AFB1 group when compared with Controls (Table 1). Administration of 1 and 1.5 ppm of AFB1 also decreased BW by 8 and 11% during the second week and 16 and 26% in the third week, respectively, compared with Controls. This reduction was proportionally similar in BWG being 10 and 13% lower for 1 and 1.5 ppm of AFB1 during the second week, and 17 and 28% for 1 and 1.5 ppm during the third week. FI was not affected by AFB1 consumption during the first 2 weeks; however, there was a reduction of 15 and 28% in fed intake in chickens that consumed 1 and 1.5 ppm of AFB1, respectively, during the last week (Table 1). FCR varied accordingly in the three diet groups during the whole experiment except the second week where Control group had a more efficient ratio compared to the AFB1 groups (Table 1). In experiment 2, the liver weight ratio was significantly increased in chickens that received 1.5 ppm when compared with Control (Table 2). However, spleen ratio was increased in both groups of chickens that received 1 or 1.5 ppm of AFB1 when compared with Controls. Bursa ratio was increased only in chickens that received 1 ppm (Table 2).
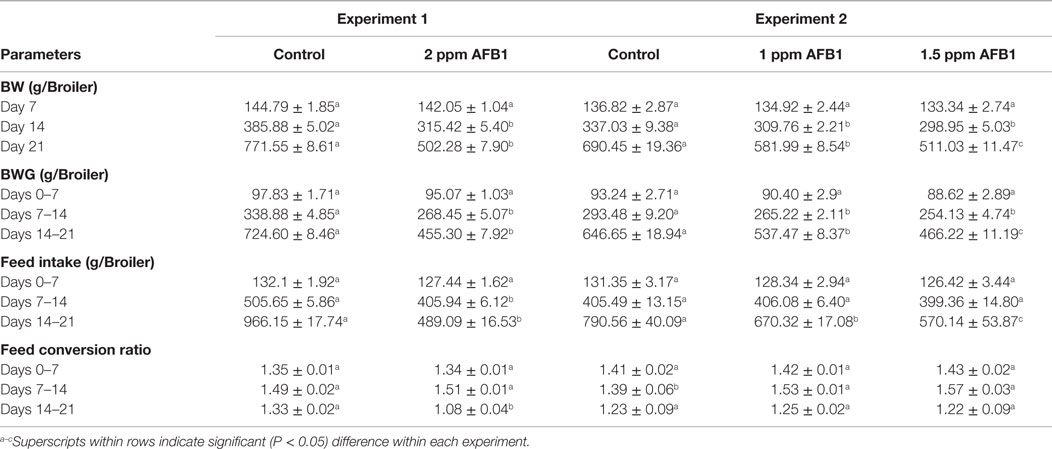
Table 1. Effect of dietary administration of 2, 1.5, and 1 ppm of aflatoxin B1 on body weight (BW), body weight gain (BWG), feed intake, and feed conversion ratio at 7, 14, and 21 days in broiler chickens or experiments 1 and 2.
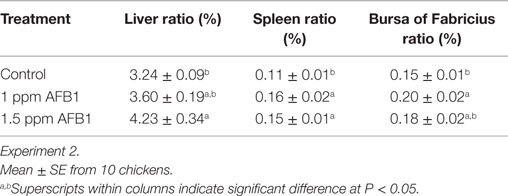
Table 2. Effect of 1 and 1.5 ppm of aflatoxin B1 on body weight ratios for liver, spleen, and bursa of Fabricius in 21-day-old broiler chickens.
Total Bacterial Counts in Cecum
In experiment 1, chicks receiving 2 ppm of AFB1 had an increase in the number of total Gram-negative bacteria and total LAB, but the total numbers of aerobes were similar between chickens that received 2 ppm of AFB1 and Control chickens (Table 3). In experiment 2, the total number of aerobic bacteria and total Gram negatives were higher in 1.5 ppm AFB1 group. Conversely, the number of total LAB was reduced in chickens fed with 1 ppm AFB1. No difference was observed in total yeast count between groups in neither of both experiments (Table 3).
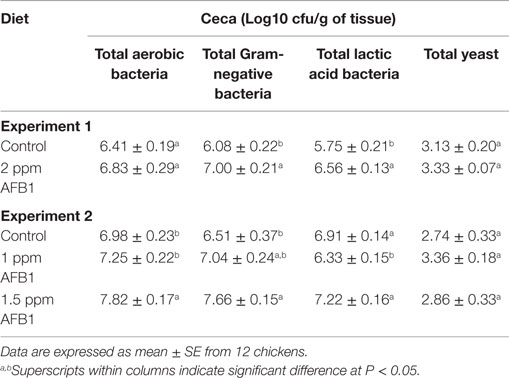
Table 3. Effect of 2 ppm of aflatoxin B1 (experiment 1) or 1 and 1.5 ppm of aflatoxin B1 (experiment 2) on total bacterial and yeast counts from cecum samples in broiler chickens at 21 days.
Hematology
In experiment 2, a significant heterophilia with a marked lymphopenia was observed in both groups that received AFB1 (Table 4). Consequently, an increase in the heterophils-to-lymphocyte ratio was also observed in those groups when compared with Controls. No significant differences were found in the numbers of monocytes, eosinophils, or basophils (data not shown). Hemoglobin, MVC, and MCH were significantly decreased in chickens that consumed 1.5 ppm of AFB1 when compared with Controls. These values were not affected in chickens that received 1 ppm when compared with Controls (Table 4).
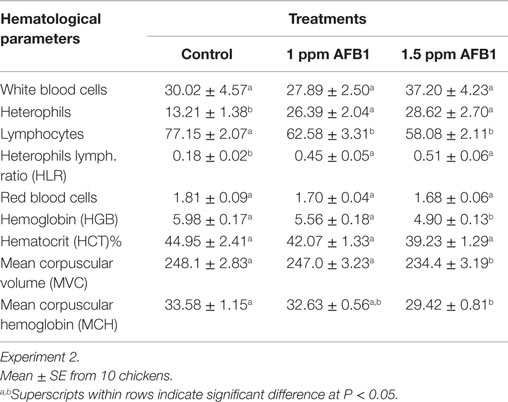
Table 4. Effect of 1 and 1.5 ppm of aflatoxin B1 on blood parameters and serum chemistry in broiler chickens at 21 days.
Bacterial Translocation and FITC-d Leakage
Chickens receiving a diet with 2 ppm of AFB1 had a significant reduction in BT to the liver when compared to Control chickens (Table 5). Interestingly, there were no differences in serum levels of FITC-d levels between Control and treated chickens. On the other hand, in experiment 2, chicks fed 1.5 ppm AFB1 did not show significant differences in BT when compared with Control chickens, but no bacteria recovery was observed from livers of chickens fed with 1 ppm AFB1. Nevertheless, similar to experiment 1, no significant differences were observed in the levels of serum FITC-d between chicks that received 1 or 1.5 ppm of AFB1 and Control chickens (Table 5).
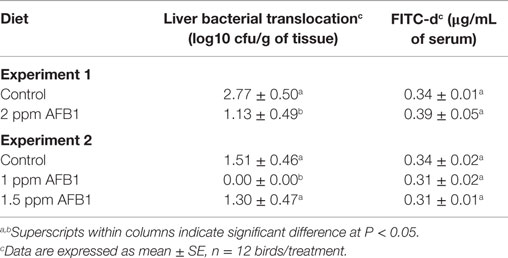
Table 5. Effect of 2 ppm of aflatoxin B1 (experiment 1) or 1 and 1.5 ppm of aflatoxin B1 (experiment 2) on liver bacterial translocation and serum FITC-d levels in broiler chickens at 21 days.
Morphometric Analysis
Villus length in both duodenum and ileum sections was significantly increased in a dose-related fashion in chickens that received 1 and 1.5 ppm of AFB1 when compared with Control chickens (Table 6). However, a significant reduction in duodenum crypt depth was observed in chickens that received 1 and 1.5 ppm of AFB1 when compared with Control chickens. On the other hand, similar changes in ileum crypt depth were found in chickens that received 1.5 ppm of AFB1 when compared with Control or 1 ppm chickens. Changes in duodenum villus height/crypt depth ratio were inconsistent between doses of AFB1 in this study.
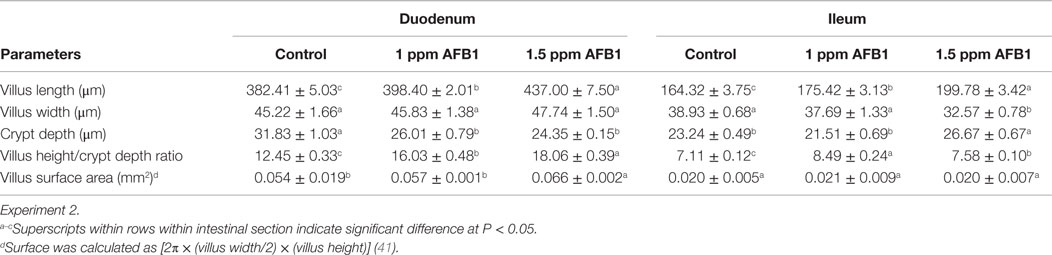
Table 6. Morphometric analysis of duodenum and ileum samples from broiler chickens at 21 days.
In the ileum, this relationship was increased in chickens that received 1 ppm, followed by chicks that received 1.5 ppm of AFB1 and Control chickens had the lower villus height/crypt depth ratio. The surface area of the duodenum was significantly higher in chicks that received 1.5 ppm of AFB1, but no changes in ileum surface area were observed between the three groups (Table 6).
Discussion
Aflatoxins have several effects in poultry, including poor performance, liver pathology, immunosuppression, and changes in relative organ weights (30, 31, 43, 44). Our results were consistent with these previous studies demonstrating dose-related effects on reduction of BW, BWG, FI, and feed conversion as well as increase relative weights of liver, spleen, and bursa of Fabricius.
In spite of the indicated antimicrobial potential of AFB1, we found few reports regarding the effects of the toxin on gut microbial populations. Kubena et al. (44) reported a significant increase in total volatile fatty acids at 5 days of age in chickens that received 2.5 and 7.5 ppm of AFB1, suggesting changes in LAB populations (45, 46). In other studies, Lactobacillus spp. have been noted to change under the influence of AFB1; however, these changes did not warrant any beneficial effects of AFB1 on intestinal microbial population (47).
In the present study, AFB1 significantly increased the total number of Gram-negative bacteria in chickens fed with 2 and 1.5 ppm and numerically in chickens fed with 1 ppm, and a similar trend was observed in the total number of LAB for chickens receiving 2 and 1.5 ppm of AFB1. However, chickens that received 1 ppm showed a significant reduction of total LAB but higher total number of aerobic bacteria when compared with Control chickens. Interestingly, little information about the outcomes of AFB1 on gut microbiome is available. In one study, Kubena et al. (44) reported that 2.5 ppm of AFB1 increased the production of total volatile fatty acids in broilers, which suggest higher number of total LAB populations. In the present study, no differences were observed in total yeast counts between groups in neither of both experiments; nevertheless, we could not find any other report to compare our results. Perhaps, such inconsistent results may be a reason of the lack of publications reporting yeast evaluation. Interestingly, it has been showed that fermentation patterns of Saccharomyces cerevisiae also change under the influence of AFB1 (48). AFB1 has also been reported to change fermentation patterns with increase gas production, due to fermentation of other carbohydrates of LAB, that affects negatively the cheese industry (46, 47, 49). Several investigators have reported the effects that aflatoxins cause to heterophilia lymphopenia and hemolytic anemia in poultry (6, 10, 30, 50, 51). In experiment 2, a dramatic increase in the heterophils occurred while the lymphocytes were reduced. Consequently, an increase in the heterophils-to-lymphocyte ratio was also observed in those groups when compared with Control chickens. A similar response of circulating leukocytes was also found when a physiological stress was applied to chickens (38). In aflatoxicosis, the spleen is enlarged due to the hemolytic anemia (52) and some reports indicate that the spleen of chickens is almost doubled in size (53). In experiment 2, spleens of chickens that received 1 and 1.5 ppm were significantly larger when compared with Control. The elevated WBC counts caused by both doses of AFB1 also support the clinical presentation of hemolytic anemia. Additionally, hemoglobin, MVC, and MCH were significantly increased in chickens that consumed 1.5 ppm of AFB1 when compared with Control chickens, confirming that aflatoxicosis causes a hemolytic anemia in chickens as has been previously reported (30, 51, 52, 54, 55).
We have previously shown that intestinal inflammation can be induced by diet ingredients or stress, affecting intestinal permeability (33–36). As the largest barrier in the body, IEC are responsible for absorption of water and nutrients, but they also prevent the entry of antigens into the blood (56–58).
Contrary to our initial hypothesis, 2 ppm of AFB1 did not increase intestinal permeability, as was evidenced by a significant reduction in BT or similar levels of serum FITC-d when compared with Control chickens. It is possible that the inflammation of the liver that is characterized by infiltration of heterophils and other inflammatory cells may handle cleaning any bacterial leakage that arrives from the porta system to the liver. Those results encouraged us to repeat and extend the experiment with lower doses of AFB1 and by comparing the morphometric changes between Control and treated groups. Our findings from experiment 2 showed that chickens fed 1.5 ppm AFB1 showed a numerical reduction in BT when compared with Control chickens, but no bacteria were recovered from livers of chickens fed with 1 ppm AFB1. Also, similar to experiment 1, no significant differences were observed in the levels of serum FITC-d between chicks that received 1 or 1.5 ppm of AFB1 and Control chickens.
Increased intestinal leakage is also associated with BT in the portal circulation (59, 60). Likewise, FITC-d is a bulky molecule (3–5 kDa) which is not observed under normal conditions. Nevertheless, if TJs between epithelial cells are altered, FITC-d can be detected in serum, indicating damage to the TJs following FITC-d gavage administration (61). It has been reported that AFB1 does not destroy TJs (32), it has only minor effects on the gut-associated lymphoid tissue (GALT) (62), confirming that AFB1 does not induce inflammation in the GIT.
Literature reports on the effects of AFB1 on histology of GIT are limited and not conclusive (10, 11, 18, 27, 63). However, it is important to mention that the few studies that have evaluated the effect of AFB1 on intestinal histology are reports using different concentrations of AFB1, different avian species, different ages, as well as time of AFB1 administration. Interpretation of our morphometric results was also inconclusive. Nevertheless, the GIT is highlighted as a dynamic organ that is able to adapt to a chronic AFB1 as has been demonstrated by several scientists (18, 20–23, 26, 27). In summary, the results of the present study suggest that AFB1 does not increase gut leakage as is evidenced by the lack of increase permeability of FITC-d in the serum. On the other hand, further studies are needed to clarify the BT and morphometric results with AFB1.
Author Contributions
RS: contributions to conception and design, acquisition of data, and analysis and interpretation of data; JL: contributions to conception and design, acquisition of data, and/or analysis and interpretation of data; LB: contributions to conception and design, and/or analysis and interpretation of data; VK: contributions to conception and design and acquisition of data; AW: acquisition of data; XV: final approval of the version to be submitted, drafting the article, or revising it critically for important intellectual content; RG: acquisition of data; JV: acquisition of data; AD: final approval of the version to be submitted; DC: acquisition of data; BH: final approval of the version to be submitted; and GT: contributions to conception and design, acquisition of data, and analysis and interpretation of data.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors gratefully acknowledge the excellent technical assistance of Sonia Tsai (Poultry Production and Product Safety Research Unit, USDA, Agricultural Research Service, Poultry Science Center, University of Arkansas, Fayetteville) and all the professional staff, who work at the Interlibrary Loan Service of the University of Arkansas, for their unconditional help and support.
References
2. Siller W, Ostler D. The histopathology of an enterohepatic syndrome of turkey poults. Vet Rec (1961) 73:134–8.
3. Nesbitt BF, O’kelly J, Sargeant K, Sheridan A. Aspergillus flavus and turkey X disease: toxic metabolites of Aspergillus flavus. Nature (1962) 195:1062–3. doi: 10.1038/1951062a0
4. Spensley PC. Aflatoxin, the active principle in turkey ‘X’ disease. Endeavour (1963) 22:75–9. doi:10.1016/0160-9327(63)90097-8
5. Cole RJ. Etiology of turkey “X” disease in retrospect: a case for the involvement of cyclopiazonic acid. Mycotoxin Res (1986) 2:3–7. doi:10.1007/BF03191956
6. McLean M, Dutton MF. Cellular interactions and metabolism of aflatoxin: an update. Pharmacol Ther (1995) 65:163–92. doi:10.1016/0163-7258(94)00054-7
7. Fox JG, Feng Y, Theve EJ, Raczynski AR, Fiala JLA, Doernte AL, et al. Gut microbes define liver cancer risk in mice exposed to chemical and viral transgenic hepatocarcinogens. Gut (2010) 59:88–97. doi:10.1136/gut.2009.183749
8. Rawal S, Kim JE, Coulombe R. Aflatoxin B1 in poultry: toxicology, metabolism and prevention. Res Vet Sci (2010) 89:325–31. doi:10.1016/j.rvsc.2010.04.011
9. Rawal S, Coulombe RA. Metabolism of aflatoxin B1 in turkey liver microsomes: the relative roles of cytochromes P450 1A5 and 3A37. Toxicol Appl Pharmacol (2011) 254:349–54. doi:10.1016/j.taap.2011.05.010
10. Yunus AW, Razzazi-Fazeli E, Bohm J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: a review of history and contemporary issues. Toxins (Basel) (2011) 3:566–90. doi:10.3390/toxins3060566
11. Zhang S, Peng X, Fang J, Cui H, Zuo Z, Chen Z. Effects of aflatoxin B1 exposure and sodium selenite supplementation on the histology, cell proliferation, and cell cycle of jejunum in broilers. Biol Trace Elem Res (2014) 160:32–40. doi:10.1007/s12011-014-0009-5
12. Ledoux DR, Rottinghaus GE, Bermudez AJ, Alonso-Debolt M. Efficacy of a hydrated sodium calcium aluminosilicate to ameliorate the toxic effects of aflatoxin in broiler chicks. Poult Sci (1999) 78:204–10. doi:10.1093/ps/78.2.204
13. Andrade PD, Silva J, Caldas ED. Simultaneous analysis of aflatoxins B1, B2, G1, G2, M1 and ochratoxin A in breast milk by high-performance liquid chromatography/fluorescence after liquid-liquid extraction with low temperature purification (LLE-LTP). J Chromatogr A (2013) 1304:61–8. doi:10.1016/j.chroma.2013.06.049
14. Greco MV, Franchi ML, Rico Golba SL, Pardo AG, Pose GN. Mycotoxins and mycotoxigenic fungi in poultry feed for food-producing animals. ScientificWorldJournal (2014) 2014:968215. doi:10.1155/2014/968215
15. Warren MF, Hamilton PB. Intestinl fragility during ochratoxicosis and aflatoxicosis in broiler chickens. Appl Environ Microbiol (1980) 40:641–5.
16. Xu Z, Hu C, Xia M, Zhan X, Wang M. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult Sci (2003) 82:1030–6. doi:10.1093/ps/82.6.1030
17. Diaz GJ, Calabrese E, Blain R. Aflatoxicosis in chickens (Gallus gallus): an example of hormesis? Poult Sci (2008) 87:727–32. doi:10.3382/ps.2007-00403
18. Applegate TJ, Schatzmayr G, Prickel K, Pricket K, Troche C, Jiang Z. Effect of aflatoxin culture on intestinal function and nutrient loss in laying hens. Poult Sci (2009) 88:1235–41. doi:10.3382/ps.2008-00494
19. Yunus AW, Ghareeb K, Abd-El-Fattah AAM, Twaruzek M, Böhm J. Gross intestinal adaptations in relation to broiler performance during chronic aflatoxin exposure. Poult Sci (2011) 90:1683–9. doi:10.3382/ps.2011-01448
20. Smith LE, Stoltzfus RJ, Prendergast A. Food chain mycotoxin exposure, gut health, and impaired growth: a conceptual framework. Adv Nutr (2012) 3:526–31. doi:10.3945/an.112.002188
21. Ruff M, Wyatt R. Intestinal absorption of L-methionine and glucose in chickens with aflatoxicosis. Toxicol Appl Pharmacol (1976) 37:257–62. doi:10.1016/0041-008X(76)90088-0
22. Fan Y, Croom J, Christensen V, Black B, Bird A, Daniel L, et al. Jejunal glucose uptake and oxygen consumption in turkey poults selected for rapid growth. Poult Sci (1997) 76:1738–45. doi:10.1093/ps/76.12.1738
23. Nelson TS, Johnson ZB, Kirby LK, Beasley JN. Digestion of dry matter and amino acids and energy utilization by chicks fed molded corn containing mycotoxins. Poult Sci (1982) 61:584–5. doi:10.3382/ps.0610584
24. Verma J, Swain BK, Johri TS. Effect of various levels of aflatoxin and ochratoxin A and combinations thereof on protein and energy utilisation in broilers. J Sci Food Agric (2002) 82:1412–7. doi:10.1002/jsfa.1203
25. Verma J, Johri TS, Swain BK. Effect of aflatoxin, ochratoxin and their combination on protein and energy utilisation in white leghorn laying hens. J Sci Food Agric (2007) 87:760–4. doi:10.1002/jsfa.2655
26. Kermanshahi H, Reza AM, Afzali N. Effect of low-level administration of aflatoxin B1 into diet on performance and activity of some blood enzymes in broiler chickens. J Sci Technol Agric Nat Resour (2007) 11:443–50.
27. Yunus AW, Awad WA, Kröger S, Zentek J, Böhm J. In vitro aflatoxin B(1) exposure decreases response to carbamylcholine in the jejunal epithelium of broilers. Poult Sci (2010) 89:1372–8. doi:10.3382/ps.2009-00617
28. Ramos A, Hernandez E. In situ absorption of aflatoxins in rat small intestine. Mycopathologia (1996) 134:27–30. doi:10.1007/BF00437049
29. Ortatatli M, Oguz H. Ameliorative effects of dietary clinoptilolite on pathological changes in broiler chickens during aflatoxicosis. Res Vet Sci (2001) 71:59–66. doi:10.1053/rvsc.2001.0487
30. Huff W, Kubena L, Harvey R, Corrier D, Mollenhauer H. Progression of aflatoxicosis in broiler chickens. Poult Sci (1986) 65:1891–9. doi:10.3382/ps.0651891
31. Kubena L, Harvey R, Huff W, Elissalde M, Yersin A, Phillips T, et al. Efficacy of a hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and diacetoxyscirpenol. Poult Sci (1993) 72:51–9. doi:10.3382/ps.0720051
32. Caloni F, Cortinovis C, Pizzo F, De Angelis I. Transport of aflatoxin M(1) in human intestinal Caco-2/TC7 cells. Front Pharmacol (2012) 3:111. doi:10.3389/fphar.2012.00111
33. Tellez G, Latorre JD, Kuttappan VA, Kogut MH, Wolfenden A, Hernandez-Velasco X, et al. Utilization of rye as energy source affects bacterial translocation, intestinal viscosity, microbiota composition, and bone mineralization in broiler chickens. Front Genet (2014) 5:339. doi:10.3389/fgene.2014.00339
34. Tellez G, Latorre JD, Kuttappan VA, Hargis BM, Hernandez-Velasco X. Rye affects bacterial translocation, intestinal viscosity, microbiota composition and bone mineralization in turkey poults. PLoS One (2015) 10:e0122390. doi:10.1371/journal.pone.0122390
35. Vicuña E, Kuttappan V, Galarza-Seeber R, Latorre J, Faulkner O, Hargis B, et al. Effect of dexamethasone in feed on intestinal permeability, differential white blood cell counts, and immune organs in broiler chicks. Poult Sci (2015) 94:2075–80. doi:10.3382/ps/pev211
36. Vicuña E, Kuttappan V, Tellez G, Hernandez-Velasco X, Seeber-Galarza R, Latorre J, et al. Dose titration of FITC-D for optimal measurement of enteric inflammation in broiler chicks. Poult Sci (2015) 94:1353–9. doi:10.3382/ps/pev111
37. Cobb-Vantress Inc. Broiler Management Supplement (2012). Available from: http://cobb-vantress.com/docs/default-source/guides/cobb-broiler-management-guide---english.pdf
38. Gross WB, Siegel HS. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Dis (1983) 27:972–9. doi:10.2307/1590198
39. Latorre JD, Hernandez-Velasco X, Kogut MH, Vicente JL, Wolfenden R, Wolfenden A, et al. Role of a Bacillus subtilis direct-fed microbial on digesta viscosity, bacterial translocation, and bone mineralization in turkey poults fed with a rye-based diet. Front Vet Sci (2014) 1:26. doi:10.3389/fvets.2014.00026
40. Aptekmann K, Artoni S, Stefanini M, Orsi M. Morphometric analysis of the intestine of domestic quails (Coturnix coturnix japonica) treated with different levels of dietary calcium. Anat Histol Embryol (2001) 30:277–80. doi:10.1046/j.1439-0264.2001.00331.x
41. Sakamoto K, Hirose H, Onizuka A, Hayashi M, Futamura N, Kawamura Y, et al. Quantitative study of changes in intestinal morphology and mucus gel on total parenteral nutrition in rats. J Surg Res (2000) 94:99–106. doi:10.1006/jsre.2000.5937
43. Kubena LF, Harvey RB, Buckley SA, Edrington TS, Rottinghaus GE. Individual and combined effects of moniliformin present in Fusarium fujikuroi culture material and aflatoxin in broiler chicks. Poult Sci (1997) 76:265–70. doi:10.1093/ps/76.2.256
44. Kubena L, Bailey R, Byrd J, Young C, Corrier D, Stanker L, et al. Cecal volatile fatty acids and broiler chick susceptibility to Salmonella typhimurium colonization as affected by aflatoxins and T-2 toxin. Poult Sci (2001) 80:411–7. doi:10.1093/ps/80.4.411
45. Mohran MA, Megalla SE, Said MR. Effect of aflatoxin B-1 on the proteolytic activity of some lactic-acid bacteria. Mycopathologia (1984) 86:99–101. doi:10.1007/BF00436494
46. Suti’c M, Banina A. Influence of aflatoxin B1 on gas production by lactic acid bacteria. J Environ Pathol Toxicol Oncol (1989) 10:149–53.
47. Peltonen K, El-Nezami H, Haskard C, Ahokas J, Salminen S. Aflatoxin B 1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J Dairy Sci (2001) 84:2152–6. doi:10.3168/jds.S0022-0302(01)74660-7
48. Reiss J. Influence of the mycotoxins aflatoxin B1, rubratoxin B, patulin and diacetoxyscirpenol on the fermentation activity of baker’s yeast. Mycopathol Mycol Appl (1973) 51:337–45. doi:10.1007/BF02057804
49. Georgianna DR, Payne GA. Genetic regulation of aflatoxin biosynthesis: from gene to genome. Fungal Genet Biol (2009) 46:113–25. doi:10.1016/j.fgb.2008.10.011
50. Oğuz H, Keçeci T, Birdane YO, Onder F, Kurtoğlu V. Effect of clinoptilolite on serum biochemical and haematological characters of broiler chickens during aflatoxicosis. Res Vet Sci (2000) 69:89–93. doi:10.1053/rvsc.2000.0395
51. Yousef M, Salem M, Kamel K, Hassan G, El-Nouty F. Influence of ascorbic acid supplementation on the haematological and clinical biochemistry parameters of male rabbits exposed to aflatoxin B1. J Environ Sci Health (2003) 38:193–209. doi:10.1081/PFC-120018449
52. Tung HT, Cook F, Wyatt R, Hamilton P. The anemia caused by aflatoxin. Poult Sci (1975) 54:1962–9. doi:10.3382/ps.0541962
53. Smith J, Hamilton P. Aflatoxicosis in the broiler chicken. Poult Sci (1970) 49:207–15. doi:10.3382/ps.0490207
54. Smith R, Griffin JM, Hamilton PB. Survey of aflatoxicosis in farm animals. Appl Environ Microbiol (1976) 31:385–8.
55. Tessari EN, Kobashigawa E, Cardoso ALS, Ledoux DR, Rottinghaus GE, Oliveira CA. Effects of aflatoxin B1 and fumonisin B1 on blood biochemical parameters in broilers. Toxins (Basel) (2010) 2:453–60. doi:10.3390/toxins2040453
56. Salminen S, Isolauri E. Intestinal colonization, microbiota, and probiotics. J Pediatr (2006) 149:S115–20. doi:10.1016/j.jpeds.2006.06.062
57. Salzman NH. Microbiota-immune system interaction: an uneasy alliance. Curr Opin Microbiol (2011) 14:99–105. doi:10.1016/j.mib.2010.09.018
58. Elson CO, Cong Y. Host-microbiota interactions in inflammatory bowel disease. Gut Microbes (2012) 3:332–44. doi:10.4161/gmic.20228
59. Ilan Y. Leaky gut and the liver: a role for bacterial translocation in nonalcoholic steatohepatitis. World J Gastroenterol (2012) 18:2609–18. doi:10.3748/wjg.v18.i21.2609
60. Seki E, Schnabl B. Role of innate immunity and the microbiota in liver fibrosis: crosstalk between the liver and gut. J Physiol (2012) 590:447–58. doi:10.1113/jphysiol.2011.219691
61. Yan Y, Kolachala V, Dalmasso G, Nguyen H, Laroui H, Sitaraman SV, et al. Temporal and spatial analysis of clinical and molecular parameters in dextran sodium sulfate induced colitis. PLoS One (2009) 4:e6073. doi:10.1371/journal.pone.0006073
62. Watzl B, Neudecker C, Hänsch GM, Rechkemmer G, Pool-Zobel BL. Short-term moderate aflatoxin B1 exposure has only minor effects on the gut-associated lymphoid tissue of Brown Norway rats. Toxicology (1999) 138:93–102. doi:10.1016/S0300-483X(99)00088-8
Keywords: aflatoxin B1, bacterial translocation, broilers, gut leakage, performance
Citation: Galarza-Seeber R, Latorre JD, Bielke LR, Kuttappan VA, Wolfenden AD, Hernandez-Velasco X, Merino-Guzman R, Vicente JL, Donoghue A, Cross D, Hargis BM and Tellez G (2016) Leaky Gut and Mycotoxins: Aflatoxin B1 Does Not Increase Gut Permeability in Broiler Chickens. Front. Vet. Sci. 3:10. doi: 10.3389/fvets.2016.00010
Received: 10 October 2015; Accepted: 01 February 2016;
Published: 15 February 2016
Edited by:
Massimo Amadori, Istituto Zooprofilattico Sperimentale della Lombardia e dell’Emilia-Romagna, ItalyReviewed by:
Justin C. Fowler, University of Georgia, USAAdebukunola Olufunmilayo Lala, Federal University of Agriculture, Nigeria
Copyright: © 2016 Galarza-Seeber, Latorre, Bielke, Kuttappan, Wolfenden, Hernandez-Velasco, Merino-Guzman, Vicente, Donoghue, Cross, Hargis and Tellez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guillermo Tellez, Z3RlbGxlekB1YXJrLmVkdQ==