 Elizabeth Santin1*
Elizabeth Santin1* Ricardo Mitsuo Hayashi1
Ricardo Mitsuo Hayashi1 Jessica Caroline Wammes1
Jessica Caroline Wammes1 Ricardo Gonzalez-Esquerra2
Ricardo Gonzalez-Esquerra2 Marcelo Falsarella Carazzolle3
Marcelo Falsarella Carazzolle3 Caio César de Melo Freire4
Caio César de Melo Freire4 Paulo Sérgio Monzani5
Paulo Sérgio Monzani5 Anderson Ferreira da Cunha4
Anderson Ferreira da Cunha4
- 1Laboratório de Microbiologia e Ornitopatologia, Universidade Federal do Paraná, Curitiba, Brazil
- 2Novus International, Inc., Indaiatuba, Brazil
- 3Laboratório de Genômica e Expressão, Universidade Estadual de Campinas, Campinas, Brazil
- 4Laboratório de Bioquímica e Genética Aplicada, Departamento de Genética e Evolução, Centro de Ciências Biológicas e da Saúde, Universidade Federal de São Carlos, São Carlos, Brazil
- 5Faculdade de Zootecnia e Engenharia de Alimentos, Departamento de Medicina Veterinária, Universidade de São Paulo, Pirassununga, Brazil
Salmonella enterica serovar Heidelberg is a human pathogen also found in broilers. A strain (UFPR1) has been associated with field reports of resistance to short-chain organic acids (SCOA) in broilers in the South of Brazil, but was susceptible to a Bacillus subtilis-based probiotic added in feed in a related study. This work aimed to (i) report clinical symptoms caused by SH UFPR1 in broilers, (ii) study its susceptibility to some antibiotics in vitro, and (iii) SCOA in vivo; and (iv) relate these phenotypic observations with its genome characteristics. Two in vivo trials used 1-day-old chicks housed for 21 days in 8 sterilized isolated negative pressure rooms with 4 battery cages of 12 birds each. Birds were challenged or not with 107 CFU/bird of SH UFPR1 orally and exposed or not to SCOA in a 2 × 2 factorial design. Zootechnical parameters were unaffected (P > 0.05), no clinical signs were observed, and few cecal and hepatic histologic and immune-related alterations were seen, in birds challenged with SH. Formic and propionic acids added together in drinking water, fumaric and benzoic acid in feed (Trial 1), and coated calcium butyrate in feed (Trial 2) did not reduce the SH isolation frequencies seen in cecum and liver in broilers after SH challenge (P > 0.05). SH UFPR1 was susceptible to amikacin, amoxicillin + clavulanate, ceftiofur, cephalexin, doxycycline and oxytetracycline; and mildly susceptible to ampicillin + sulbactam, cephalothin, ciprofloxacin, enrofloxacin, and gentamycin in an in vitro minimum inhibitory concentration model using Mueller–Hinton agar. The whole genome of SH UFPR1 was sequenced and consisted of a circular chromosome, spanning 4,760,321 bp with 52.18% of GC-content encoding 84 tRNA, 22 rRNA, and 4,427 protein-coding genes. The comparison between SH UFPR1 genome and a multidrug-resistant SL476 strain revealed 11 missing genomic fragments and 5 insertions related to bgt, bgr, and rpoS genes. The deleted genes codify proteins associated with cell cycle regulation, virulence, drug resistance, cellular adhesion, and salt efflux which collectively reveal key aspects of the evolution and adaptation of SH strains such as organic acids resistance and antibiotic sensitivity and provide information relevant to the control of SH in poultry.
Introduction
Despite recent advances in the treatment of infectious diseases, pathogenic microorganisms, including Salmonella, remain an important threat to human and animal health worldwide (1). Non-typhoid Salmonella serovars are well-known pathogens but they also silently infect animals, particularly poultry, as transient members of the intestinal microbial population without causing disease (2, 3). Intestinal colonization of several Salmonella enterica serovars often have no effect on poultry zootechnical performance (3). Kogut et al. (4) described that the infection in chickens caused by Salmonella Enteritidis leads to immune tolerance beginning around 3–4 days post-primary infection. This decreases the host immune responsiveness resulting in the establishment of Salmonella and persistent colonization. This asymptomatic infection could increase the probability of transmission to humans via contaminated food (5).
Therefore, reducing Salmonella colonization and fecal shedding in live chickens, and its subsequent chicken meat contamination, can reduce the burden of salmonellosis in humans. Although many aspects related to their mechanisms of action are unknown, short-chain organic acids (SCOA) have been added to chicken feed, drinking water, and other matrices, as part of several strategies to prevent Salmonella colonization in animal tissues and transmission through the food chain with many positives results (6).
In that context, Salmonella enterica serovar Heidelberg is one of the most prevalent serovars and is commonly isolated from patients with salmonellosis in North America. That region has the greatest prevalence comparing to other continents (7). More invasive human infections, such as myocarditis and bacteremia, are produced by SH compared to other non-typhoid Salmonella (8). Since 1962, SH has been isolated and reported from poultry and their products worldwide (9) including Brazil (10). Voss-Rech et al. (11) reported 20 different Salmonella serovars in samples from broilers and SH was prevalent in 7.31% of them. SH UFPR1 strain was isolated from a chicken carcass in the South of Brazil. This strain showed susceptibility and resistance to antibiotics and organic acids, respectively. In a previous trial, our group showed reduction in SH counts in the cecum and liver of broilers fed with a probiotic composed of three strains of Bacillus subtillis previously challenged orally with SH UFPR1 strain. No studies comparing the genome of Brazilian UFPR1 to other SH strains have been reported.
Whole-genome sequencing is a tool that allows to investigate the genomic features of any organism. Several genomes from Salmonella strains have been decoded using this technique, aiming at improving the understanding of some aspects of their evolutionary biology, distinguishing outbreak-related strains of sporadic infections (12) and comparing genomes of strains with different clinical history and resistance profile (1, 13, 14). Whole-genome sequencing was recently used to study the differences among SH serotypes (15, 16) and resistance to different antibiotics (17, 18).
The objective of this work was to report possible clinical symptoms caused by SH UFPR1 strain in broilers, test the efficacy of SCOA to reduce SH UFPR1 strain proliferation in broilers, study the susceptibility of this strain to some antibiotics in vitro, and relate these phenotypic observations with its genome characteristics through comparative genomics.
Materials and Methods
In Vivo Experiments
The SH UFPR1 strain used herein was isolated from commercial broiler carcasses in the South of Brazil according to the Brazilian Ministry of Agriculture’s procedures [MAPA (19)] and sent to Fiocruz Institute (protocol number 6830/2012) for further serological identification.
Two experiments were conducted at CERIA (Center of Immune Response in Poultry) at the Federal University of Parana, Curitiba, Brazil, to evaluate the effectiveness of several SCOA to control UFPR1 in broilers, and report possible clinical symptoms in orally challenged birds. In Trial 1, an SCOA blend (30% of formic acid and 18% of propionic acid) was offered in drinking water at 0.05% from 1 to 7 and from 15 to 21 days of age, along with 3 kg/ton of feed of a SCOA blend composed of fumaric and benzoic acids at 92% and fed from 1 to 21 days of age. In Trial 2, the effect of adding a product containing 89% of coated calcium butyrate when added at 2 kg/ton in feed from 1 to 21 days was studied.
All experimental procedures were approved by the Ethical Committee of Agricultural Sector of Federal University of Parana under approval number 037/2016 and 014/2016, respectively. All other methods were equal for both trials as described below.
Chicken Housing and Sample Collection
Eight previously disinfected negative-pressured isolation rooms were used. Rooms were equipped with automatic temperature and lighting controls, and each contained four battery cages (replications) stacked vertically with sterilized litter and nipple drinkers. Before the start of each trial, swabs from all walls and cages within the rooms were collected to verify the absence of Salmonella by qualitative analysis. One day-old male chicks (Cobb® 500; n = 192) were kept from 1 to 21 days of age and distributed using a completely randomized design of four treatments (n = 48 birds per treatment, with four replicates per treatment, 12 birds per replication) detailed as follows: T1: non-challenged plus control diet, T2: non-challenged plus SCOA treatment according to Trial, T3: challenged with SH plus control diet, T4: challenged with SH plus SCOA according to Trial (Table 1).
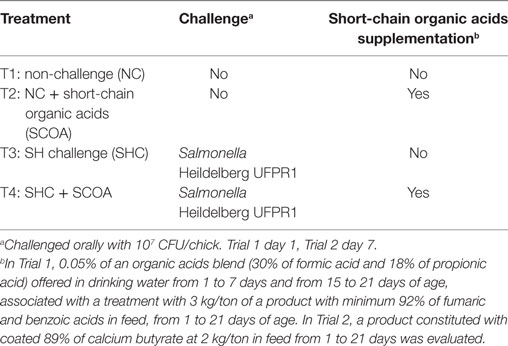
Table 1. Experimental treatment design.
At day 1, 10 chicks were necropsied, and liver and cecum samples were collected to confirm the negativity in both in vivo experiments by qualitative analysis and at day 21, feed intake (FI), body weight gain (BWG), and feed conversion (FC) were calculated.
At days 7 and 21 in Trial 1 and days 14 and 21 in Trial 2, 12 birds from each treatment were euthanized by cervical dislocation, necropsied and liver and cecum were collected for Salmonella sp. counting procedure. In Trial 2, liver and cecum of five birds per treatment were collected for histology at day 14. At that age, mRNA expression of IL-10 and IL-12 was also evaluated in liver.
Diets
The nutritional value of experimental diets was formulated to supply nutrients at requirements (20). Diets were corn and soybean meal based and were offered in mash form ad libitum at all times. Rations were formulated without coccidiostatics or antibiotics and were designed for a unique feeding phase (Starter) from 1 to 21 days of age for all treatments.
A basal diet with all ingredients except amino acids, vitamin, and mineral premix was autoclaved at 120°C/15 min. After this process, SCOA, amino acids, vitamin, and mineral premix were added according to each treatment. All dietary components were mixed for 10 min in a 50 kg blender. Batches were blended in such an order as to avoid interference among treatments.
Salmonella Heidelberg Challenge and Quantification
At 3 or 7 days of age for Trials 1 or 2, respectively, chicks from T3 and T4 were challenged orally with 107 CFU/chick of SH UFPR1.
The quantification of typical colonies of Salmonella sp. (quantitative analysis) was performed in liver and cecum samples processed according to the modified methodology by Pickler et al. (21). The organs were weighed, mashed and homogenized in 2% buffered peptone water (1:9). Further dilution was conducted by successively placing 1 mL of the solution in a test tube with 9 mL 0.1% peptone water until a 10−3 dilution was achieved. Then, 100 µL aliquots of each dilution were transferred to duplicate plates in Brilliant Green Agar (BGA) medium and uniformly spread with a sterile Drigalsky loop. Plates were incubated at 35°C for 24 h before typical colonies were counted. For all samples, pre-enrichment was performed with 2% buffered peptone water at 35°C for 24 h. Samples that did not show typical Salmonella colonies during BGA counting, were enriched with 10 mL Rappaport-Vassiliadis broth and incubated at 42°C for 24 h. Thereafter, a drop of the enriched broth was placed on BGA medium. The samples that were negative after direct BGA plate counting, but positive after enrichment were assumed to have 101 CFU/g. Samples that were negative after enrichment were assumed to have 0 CFU/g. To verify the Salmonella serotype, isolated Salmonella samples were sent to the Sector of Enterobacteria of the Oswaldo Cruz Institute, Brazil for serotyping.
Cecal and Hepatic Histologic Evaluation
For trial 2 only, samples of cecum and liver were processed according to Kraieski et al. (22). Tissues embedded in paraffin were cut in 5 µm sections and both later stained with hematoxylin and eosin, and with Alcian Blue for cecum also. Liver samples were examined using 5 fields per bird with a 10× objective and 100× of magnification. Congestion, hydropic degeneration, cell vacuolation, bile-duct proliferation, immune cells infiltration, pericholangitis, and lymphocytic aggregate were observed. The “I See Inside” (ISI) methodology applied herein was described by Santin et al. (23) where an impact factor (IF) was assigned to each type of alteration according to its potential to reduce organ functionality. The basis for these criteria was previous literature review on the relationship of organ functionality and type of lesion, and background research. The IF ranges from 1 to 3, where 3 defines an alteration that impacts organ functionality the most, and 1 the least. For instance, necrosis has the highest IF because the functional capacity of affected cells is totally lost. In addition, an observer score (OS) is assigned to each lesion based on its observed intensity or frequency compared to non-affected organs, during histologic inspection. This evaluation is performed in each organ/tissue per animal and OS values range from score 0 (absence of lesion or frequency), score 1 (alteration up to 25% of the area or observed frequency), score 2 (alteration ranges from 25 to 50% of the area or observed frequency), and score 3 (alteration extent more than 50% of the area or observed frequency). In order to calculate the final ISI Index, the IF of each alteration is multiplied by the respective score number, and the results of all alterations are summed [ISI = Σ (IF × OS)]. The scale ranges from 0 to 39 for the liver.
For cecum samples, 5 fields per bird in 40× objective and 400× of magnification were evaluated, and villus height, villus thickness, presence of erythrocytes, and infiltration of immune cell on lamina propria were measured.
Cytokines mRNA Expression in Liver (Trial 2)
Six birds per treatment were euthanized, their livers removed and immediately stored at −20°C until further analysis. Total RNA from that tissue was isolated using Trizol reagent (15596-018, Invitrogen, Carlsbad, CA, USA) following the manufacturer’s procedures. Turbo-DNAse kit (AM1907, Applied Biosystems, Foster City, CA, USA) was used for the collected samples. RNA concentrations were quantified by NanoDrop Spectrophotometer (ND1000, Thermo Scientific, Bonn, Germany) and RNA integrity determined by Experion Automated Electrophoresis System (700-7000, Bio-Rad, Hercules, CA, USA). RNA samples were reverse transcribed and RT-qPCR analyses performed with a MyiQ System (170-9740, Bio-Rad). One microgram of RNA was converted to cDNA in a 20 µL reaction volume using the iScript™ Reverse Transcription Supermix kit (170-8841, Bio-rad) at 25°C for 1 h, 42°C for 30 min, and then 85°C for 5 min.
The genes analyzed by RT-qPCR were: IL-10 (5′-cgggagctgagggtgaa-3′ and 5′-gtgaagaagcggtgacagc-3′), IL-12 (5′-agactccaatgggcaaatga-3′ and 5′-ctcttcggcaaatggacagt-3′), and GAPDH (5′-ggtggtgctaagcgtgttat-3′ and 5′-acctctgtcatctctccaca-3′). The final 20 µL PCR contained 2 µL reverse transcription product, 2 µL of the forward and reverse gene, and 10 µL of iTAq® Universal SYBR Green Supermix (172-5122, Bio-Rad). PCR cycle conditions of all primer pairs used an initial 60 s denaturation step at 95°C followed by 40 cycles of denaturation (15 s at 95°C), annealing and extension (30 s at 60°C). The melting profile of each sample was analyzed after every PCR run to confirm PCR product specificity; and later determined by heating samples at 65°C for 30 s and then increasing the temperature at a linear rate of 20°C/s to 95°C while continuously monitoring fluorescence. Sample PCR amplification efficiencies were determined in the log-linear phase with the LinRegPCR program (24). Additionally, the delta–delta equation subtracts sample and reference Ct values from an endogenous control. However, the endogenous control (GAPDH) Ct was affected by treatments in this study (P < 0.05) and, therefore, was removed from the equation. All data were normalized to the mRNA level of the control group (group non-challenged and without SCOA) and reported as the fold-change from the reference, which was calculated as ES(40 − Ct Sample)/ER(40 − Ct Reference), where ES and ER are the sample and reference PCR amplification efficiencies, respectively (25).
Statistical Analysis of In Vivo Studies
Data were analyzed using the statistical software Statistix 9. For microbiological analysis, data were evaluated by the Shapiro–Wilk normality test. Parametric data were subjected to analysis of variance (ANOVA) and Tukey’s test (P < 0.05), while the Kruskal–Wallis test (P < 0.05) was used for non-parametric data (quantitative microbiological data and histology data). The chi-square test was used in microbiological results of presence/absence (qualitative) of Salmonella in liver for Trial 2. For zootechnical performance, and immunohistochemistry analysis, data were subjected to ANOVA using a factorial 2 × 2 design.
In Vitro Antimicrobial Susceptibility Tests
The susceptibility of SH UFPR1 against a panel of 12 antimicrobials commonly used in human and veterinary clinics in Brazil was determined by the dilution antimicrobial method using Mueller–Hinton agar after incubation at 37°C for 18–24 h. The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) results were interpreted in agreement with the interpretative criteria provided by Clinical and Laboratory Standards Institute (26). The 12 antimicrobials tested included amikacin (250 mg/mL), amoxicillin + clavulanate (14 g + 3.5 g/100 mL), ampicillin + sulbactam (1 g + 0.5 g/10 mL), ceftiofur (50 mg/mL), cephalexin (250 mg/5 mL), cephalothin (1 g/10 mL), ciprofloxacin (2 mg/mL), doxycycline (4.6 g/100 mL), enrofloxacin (10 g/100 mL), gentamycin (40 mg/mL), penicillin (6,000,000 UI/15 mL), and tetracycline (20 g/100 mL). The Escherichia coli ATCC 25,922 was used as reference strain. The MIC breakpoints were set based on CLSI (26) and FDA (9).
Genomic Analysis and Comparative Analysis
The isolated SH UFPR1 strain was cultured overnight in liquid LB medium, its genomic DNA extracted using the QIAamp DNA Mini Kit (Qiagen) and quantified using the NanoVue spectrophotomer (GE Healthcare). A total of 70 µg of DNA was sent to the High-Throughput Sequencing Facility at University of North Carolina. The library was prepared using the PacBio’s 20 Kb template prep protocol (PN_100-286-000-06) and it was size-selected by using a range setting of 8,000 bp to 50,000 bp. De Novo assembly was performed using PacBio native pipeline (27). Comparative genomic analysis was independently performed with MAUVE v.20150225 (28) and Mummer v.3.23 (29) programs, using the annotated genome of SH strain SL476 as reference (GenBank assembly accession: GCA_000020705.1) (30). The shared genomic fragments between UFPR1 and SL476 were identified with Mummer while the regions with no match between them were identified with a Perl script (available in https://github.com/CaioFreire/Scripts). PROKKA v.1.12 software was used for genome annotation (31) and the circular map was drawn using DNAPlotter v.10.2 (32). The fully sequenced SH UFPR1 genome was deposited at the NCBI genome database under the number CP020101. In addition, missing fragments between each other were found using Megablast (https://blast.ncbi.nlm.nih.gov) and verifying if the gene sequences in these missing fragments were present in other parts of the genome.
Results
At the in vivo trails, SCOA were offered to the birds during the early days of life to allow the gut to adapt to the treatment before the SH challenge. No effect in zootechnical parameters was observed at any time in both trials (P > 0.05) either from the adding SCOA in drinking water and/or feed, from challenging birds with SH UFPR1 strain, or from the interaction between these factors (Table 2). It should be mentioned that the primary objective of this work was not measuring performance and that more replicates would be needed to appropriately test possible effects on these parameters.
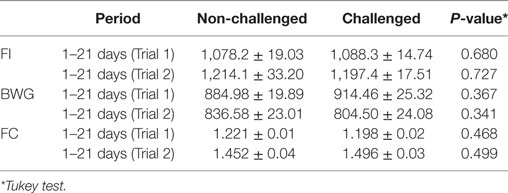
Table 2. Mean ± SD of feed intake (FI), body weight gain (BWG), and feed conversion (FC) during the periods of 1–21 days of age of broilers on Trials 1 and 2 in non-challenged and challenged birds.
As expected, all non-challenged groups tested negative for Salmonella while all challenged groups tested positive. Therefore, data on Salmonella counts were statistically evaluated in a completely randomized design using only challenged groups. Still, the use of SCOA did not influence the percentage of SH positive in cecum or liver (Figure 1; Trial 1). Similar findings were observed in Trial 2 in cecum (Figure 2) where SH counts were only performed in that organ. In that experiment, liver microbiological results were qualitative only and showed that, at 14 days of age, 100 and 42% of the samples were SH positive in challenged and challenged + SCOA birds, respectively, while, at 21 days of age, 25 and 58% tested SH positive for those two groups of broilers (P < 0.05). When comparing results from both trials, it seems that challenging birds with SH UFPR1 later in life (7 days in Trial 2 vs. 3 days in Trial 1) reduced the recovery of SH in liver.
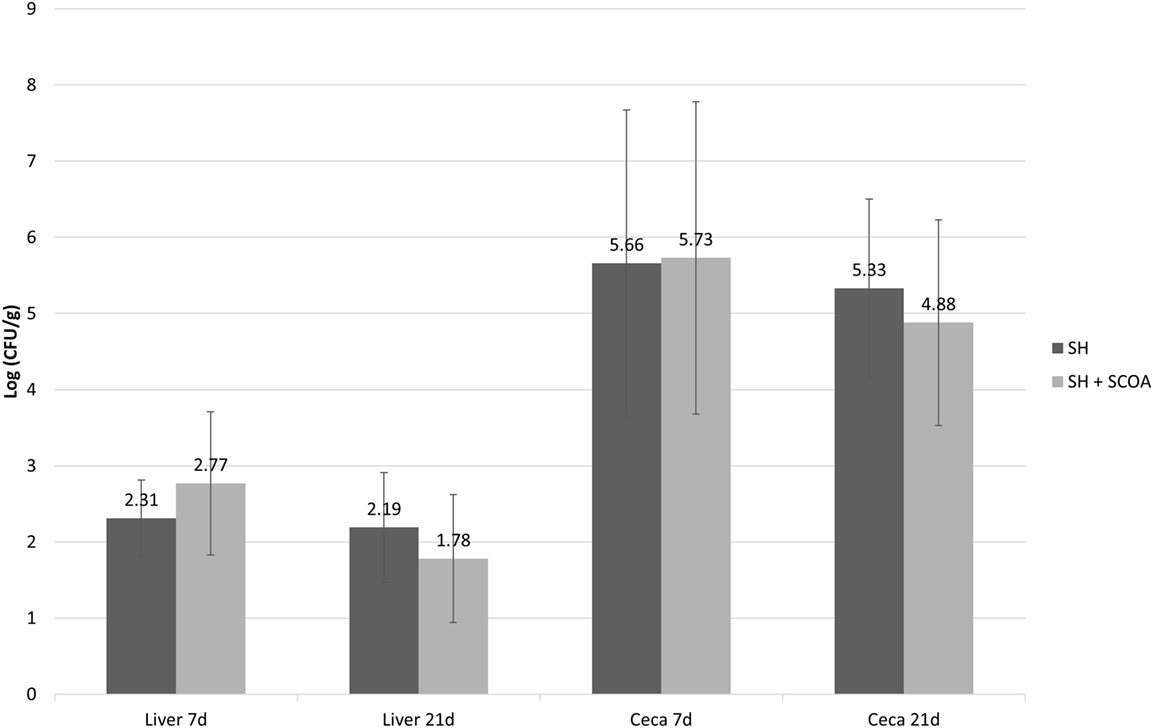
Figure 1. Trial 1. Salmonella sp. quantification (Log CFU/g) in liver and cecum at 7 and 21 days of age in treatments challenged with Salmonella Heildelberg (SH) or Salmonella Heidelberg challenged + Short-Chain Organic Acids (SH + SCOA).
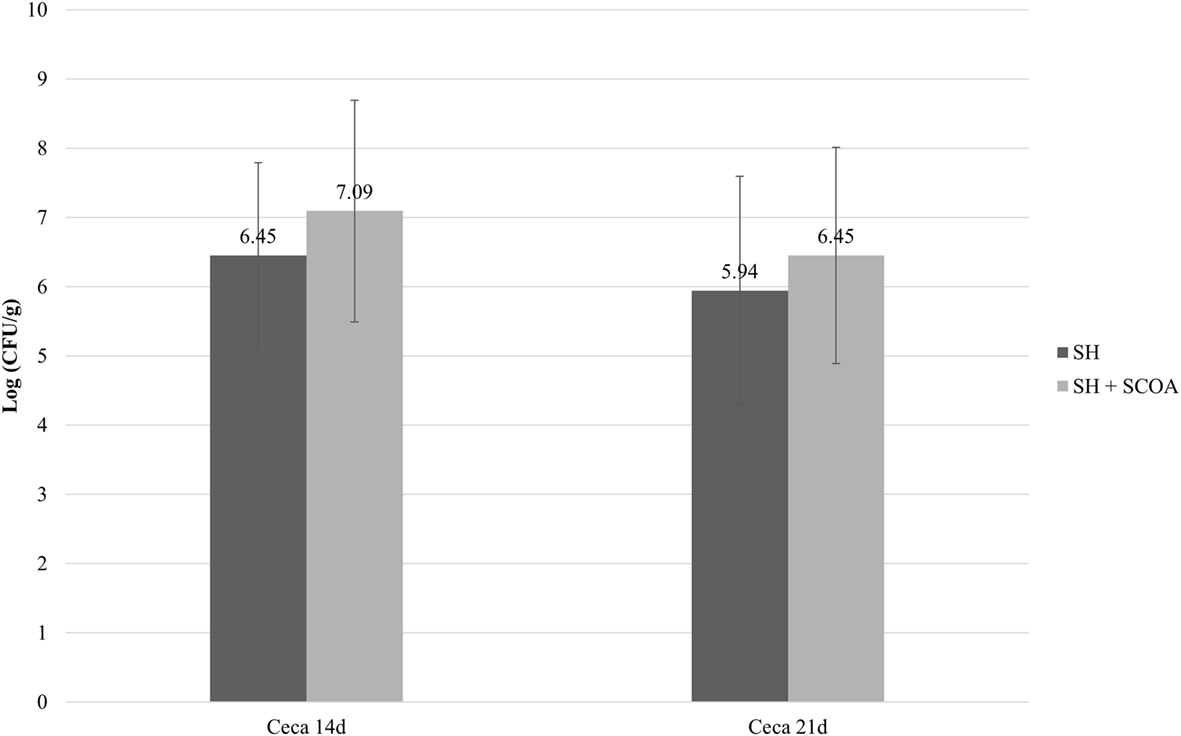
Figure 2. Trial 2. Salmonella sp. quantification (Log CFU/g) in cecum at 14 and 21 days of age in treatments challenged with Salmonella Heidelberg (SH) or Salmonella Heidelberg challenged + Short-Chain Organic Acids (SH + SCOA).
Liver histology revealed that challenging birds with SH increased the ISI score at 14 days indicating greater histological alterations (Table 3). The main alterations found were congestion, vacuolation, and immune cell infiltration as presented in Figure 3. The SCOA treatment did not influence liver histology measures.
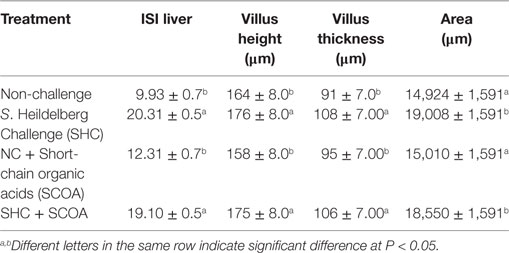
Table 3. Mean ± SE of histological alterations by I See Inside (ISI) methodology in liver and cecum at 14 days of age, evaluating the challenge and the use of feed additives interaction.
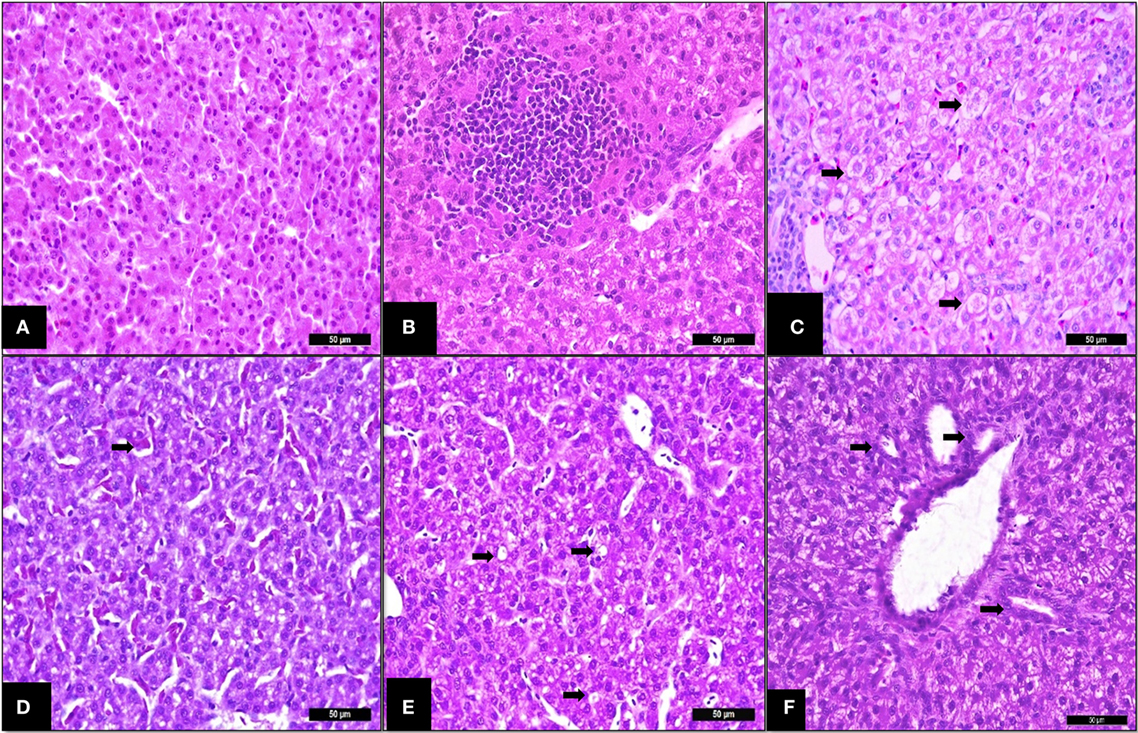
Figure 3. Trial 2. Liver of broilers (14 days). (A) Non-challenged group—normal tissue (parenchyma), I See Inside (ISI) score 23. (B) SH-Challenged group—ISI score 25, cell infiltrate in parenchyma grade II. (C) SH-Challenged group—hydropic degeneration grade III. (D) SH-Challenged group—congestion grade II. (E) SH-Challenged group—Vacuolization grade II. (F) SH-Challenged group—bile-duct proliferation grade II. Hematoxylin and eosin, 400×.
At 14 days of age, SH-challenged birds showed increased cecal villi height, villi thickness and villi surface area compared to the non-challenged group (P < 0.05; Table 3; Figure 4). Supplementing SCOA treatment did not affect the histological parameters in cecum having no interaction between SH challenge and SCOA supplementation (P > 0.05).
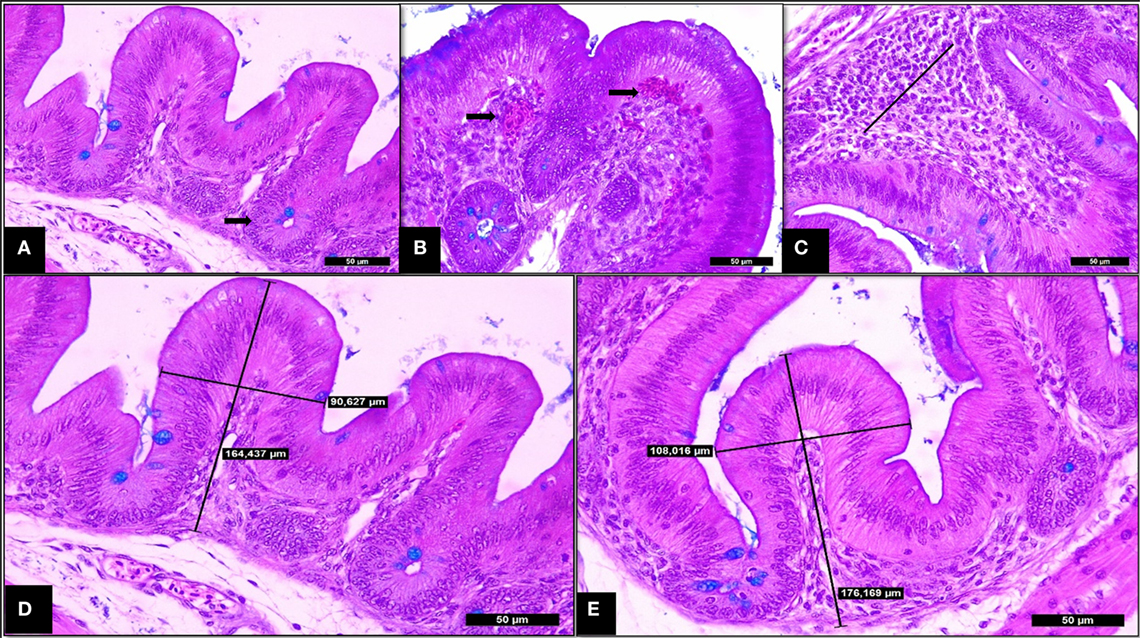
Figure 4. Trial 2. Cecum of broilers (14 days). (A) Non-challenged group—normal villi and crypts of Lieberkhün. (B) SH-Challenged group—congestion grade III. (C) SH-challenged group—cell infiltrate in lamina propria grade II. (D) Non-challenged group—villus height and thickness axes of measurement. (E) SH-challenged group—villus height and thickness axes of measurement. Hematoxylin and eosin plus Alcian Blue, 400×.
The results of mRNA expression of cytokines on liver at 14 days (Figure 5) showed higher IL-10 (P > 0.05) cytokines in SH-challenged birds compared to non-challenged, while IL-12 mRNA expression remained unaffected in all treatments.
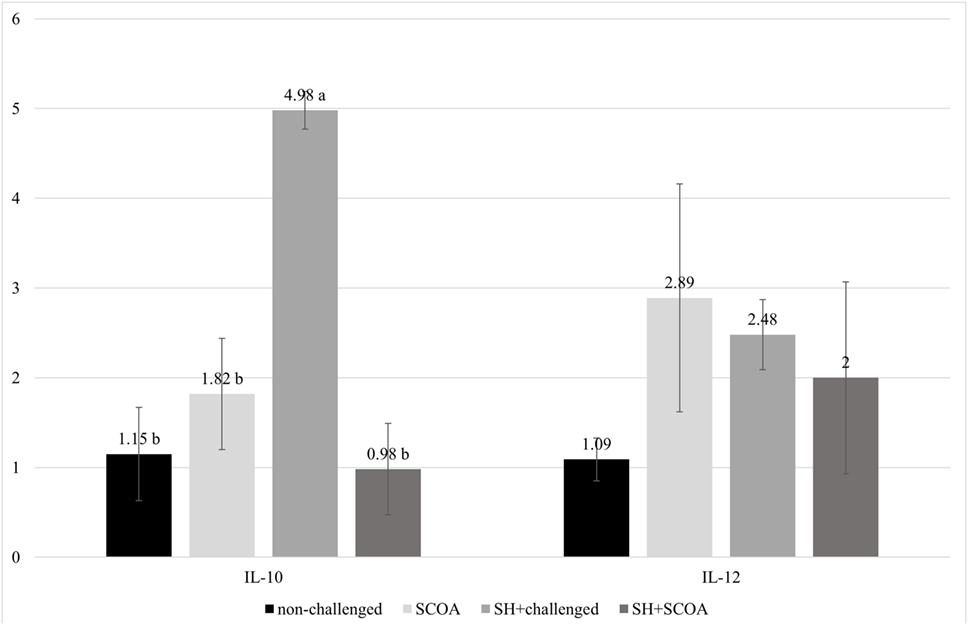
Figure 5. Trial 2. mRNA expression of IL-10 and IL-12 at 14 days of age in non-challenged group, SH-challenged group, short-chain organic acids (SCOA) and SCOA + SH-challenged group. a,bDifferent letters indicate significant differences at P < 0.05 at Kruskal–Wallis test.
The in vitro work suggests that SH UFPR1 strain is susceptible to amikacin, amoxicillin + clavulanate, ceftiofur, cephalexin, doxycycline and oxytetracycline and presents intermediary resistance to ampicillin + sulbactam, cephalothin, ciprofloxacin, enrofloxacin, and gentamycin (Table 4).
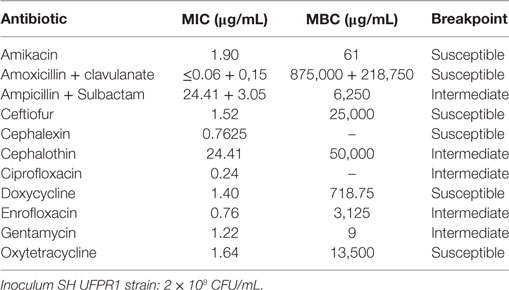
Table 4. Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of Brazilian Salmonella enterica serovar Heidelberg (UFPR1 strain).
The whole genome of UFPR1 was sequenced to better understand the genotypic particularities of this strain and compared to the genomic sequence of a multidrug-resistant SH SL476 strain. As shown in Figure 6, the assembled genomic sequence from UFPR1 strain was 128 kb smaller than SH SL476 sequence, with important deletions of 11 chromosomal fragments in the Brazilian strain. Three of them were greater than 30, 40, and 50 kb, encompassing several important genes (Datasheet S1 in Supplementary Material). Genomic regions without similar sequences in the compared genome can be observed by red dashes in Figures 6A,B. Nevertheless, the comparison between the genomes of these strains revealed high similarity with few translocation events and conserved synteny (Figure 7). Moreover, no plasmid-sequences were found in the assembled sequences from reads of the UFPR1 strain (BioProject NCBI number PRJNA378710), using Canu software v1.3 (33) to correct all input data.
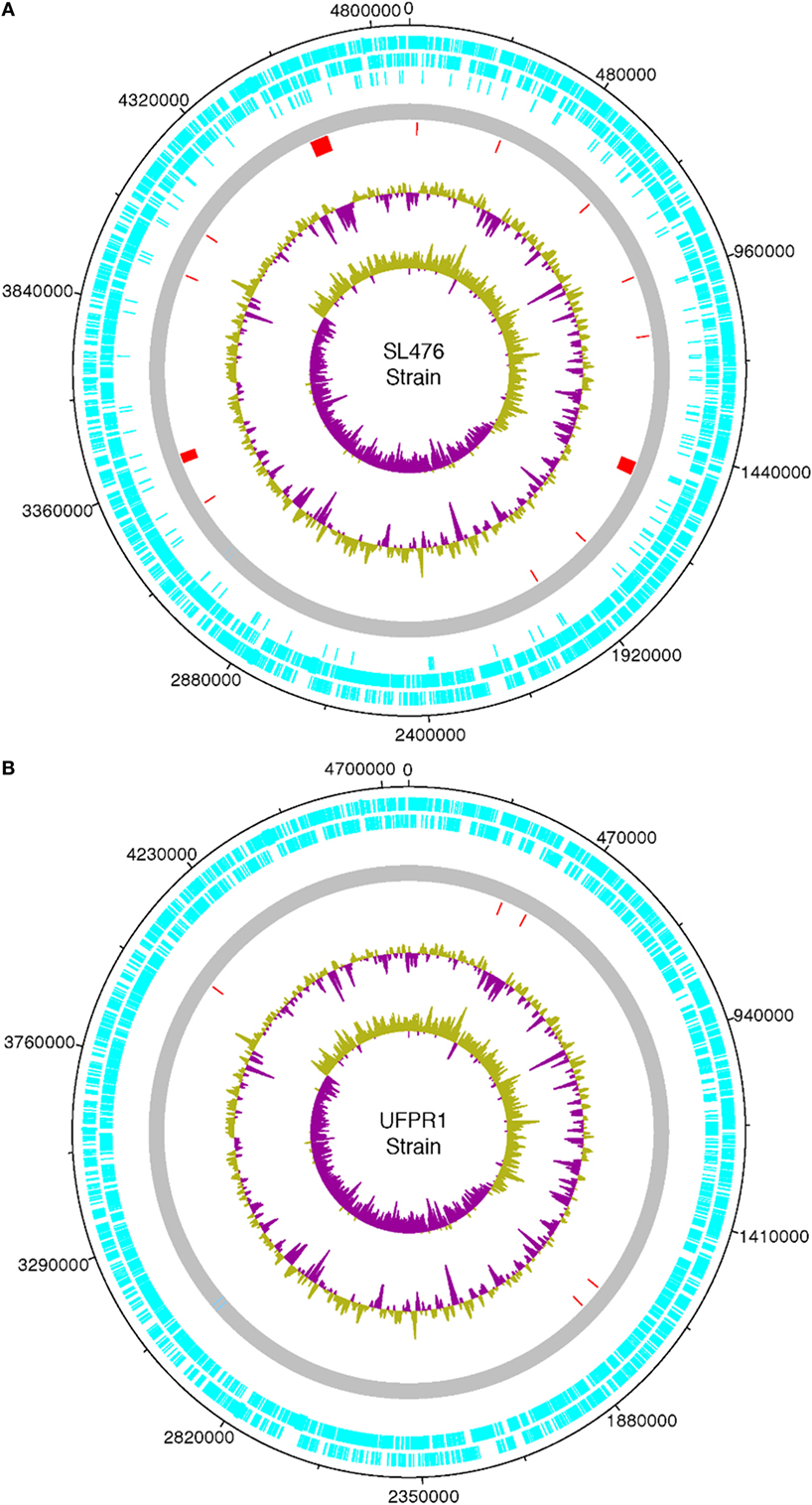
Figure 6. Chromosome features of a Brazilian UFPR1 strain (B) compared to SL476 strain (A) isolate. The circular map was drawn using DNA Plotter. Different features are shown in different colored bars. The coding sequences are shown in light blue (forward and reverse). The complete genome is shown in gray, the red dashes represents unique chromosome regions that have no homologous sequence in the genome of the other strain, green and purple in the major circle represent the GC content, while in the central circle show the GC skew [(G − C)/G + C]. Regions with GC content below the average are shown in purple and those with content above the average are shown in green.
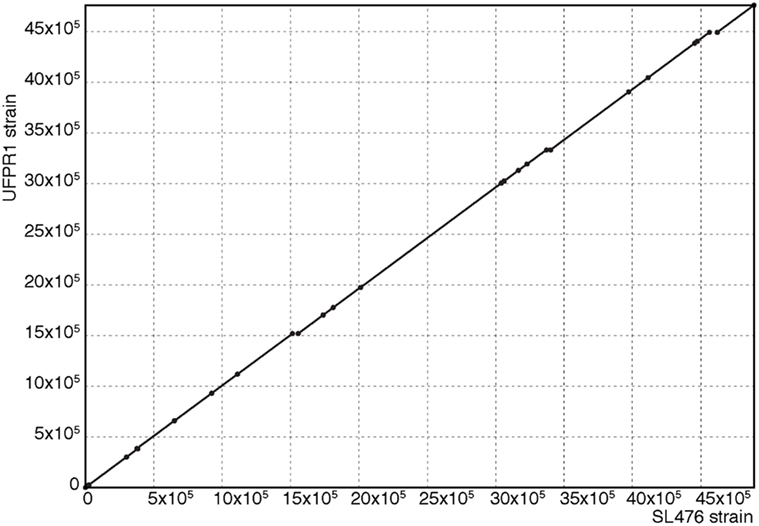
Figure 7. Alignment between genomic sequences from SL476 and UFPR1 strains. This dot-plot was generated with Mummer Software. The exact matches between genomic sequences are represented on the diagonal, showing the high conservation between the genomes with few missing fragments, as shown in Figure 6.
The genomic analysis of deleted regions revealed that 171 genes were present in SH SL476 strain but absent in UFPR1 strain (Datasheet S1 in Supplementary Material). The analyses of the 171 genes of SL476 showed that: (i) 46% were related to the codification of hypothetical proteins; (ii) 28% were mRNA sequences use to produce proteins with known functions involved in cell cycle regulation, DNA replication, virulence, drug resistance, and salt efflux; (iii) 16% correspond to encoded proteins related to the DNA recombination process (transposases and invertases genes), and (iv) 10% are encoded viral proteins (conjugal transfer, integrase, capsid, and tail proteins). Some of the genes in the absent regions were duplicated in SL476 genome or were located in another genomic region in UFPR1. Some genes were completely absent in UFPR1 strain (Datasheet S1 in Supplementary Material) such as (i) aph3 and aph6 genes that codify two isoforms of aminoglycoside O-phosphotransferase proteins; (ii) tem-1 gene that codifies a protein associated with an antibiotic resistance mechanism in bacteria; (iii) qacEΔ1 gene involved in resistance to a large spectrum of quaternary ammonium compounds (QAC); (iv) sul1 gene involved in the sulfonamide resistance; (v) tetB gene linked to the efflux of tetracycline, and (vi) lysR gene that codifies the transcriptional activator of lysA gene, which encodes the diaminopimelate decarboxylase involved in a lysine synthesis pathway. LysR gene belongs to the LYSR-type family transcriptional regulator, which regulates a varied set of genes involved in virulence, metabolism, quorum sensing, and motility (34). In the alignment of the genomes, the presence of five insertions was observed in the UFPR1 strain coding some genes like bgt, bgr, and rpoS. These genes are also present in the SL476 strain in other genomic regions and are correlated with important phenotypes found in UFPR1, such as virulence and organic acids resistance. Only five chromosomal fragments were found in UFPR1 compared to SL476 (Datasheet S2 in Supplementary Material); however, these fragments have been identified in several other strains demonstrating that they are not exclusive to UFPR1.
Discussion
The results from the in vivo studies showed that SH UFPR1 strain do not seem to affect the zootechnical performance of broilers although these results should be taken with care given the fact that our experimental layout in terms of number of replicates was not designed to be highly sensible to changes in performance, but rather to be appropriate for all the other measurements described in this report.
No clinical signs were observed after SH UFPR1 oral challenge. Other studies have shown that non-typhoid Salmonella infection in chickens does not result in morbidity or severe clinical signs in spite of intestinal colonization, and liver and spleen bacterial infiltration (2, 3, 35). The SH UFPR1 strain infection produced mild histologic alterations in liver and cecum compared to non-challenged birds, mainly associated with inflammatory processes. No differences on IL-12 mRNA expression in liver on SH-challenged vs. non-challenged birds were observed as opposed to IL-10 mRNA liver expression which increased in the former group.
Shanmugasundaram et al. (36) showed that CD4+CD25+ (Treg) cells increase in number in the cecum of chickens infected with Salmonella Enteritidis. These cells collected from cecal tonsils of S. Enteritidis-infected birds and re-stimulated in vitro with Salmonella antigen had higher IL-10 mRNA content compared to those in the control group (P < 0.05). The CD4+CD25+ cells were associated with suppressing the immune response and maintaining the Salmonella infection in the host (4). The Treg cell was not marked in this study, but the absence of clinical signs, the apparent lack of effect on zootechnical parameters, along with the increase of IL-10 mRNA found in the present work suggest that this mechanism could be similar in the UFPR1 strain.
Associating phenotypic observations with genomic sequence information of an organism can contribute to understand some of its key biologic aspects, such as mechanisms for genetic information storage, genome organization, the effect of deletion, insertion, inversion, and translocation on the genome function. To that aim, the maximum number of possible genome sequences is necessary.
Whole-chromosome alignments made in the present genomic study showed that, besides phenotypic differences, the UFPR1 strain has a genome very similar to that of the multidrug resistance SH SL476 strain overall (Figure 6). However, several chromosomal fragments that harbor various important genes were lost in UFPR1 (Datasheet S1 in Supplementary Material). The aph3 and aph6 genes that were deleted in UFPR1, encode two isoforms of aminoglycoside O-phosphotransferase that participate in the primary mechanism of resistance to aminoglycosides, such as kanamycin, gentamycin, streptomycin, and neomycin. Both genes are frequently found together with transposable elements (37). The tem-1 gene, which codifies the β-lactamase protein in bacteria, is associated with β-lactam antibiotic resistance and was also deleted in UFPR1. The protein produced by the translation of this mRNA fragment is able to hydrolyze penicillin and first-generation cephalosporin (38). Fragments of chromosome in which genes related to the production of proteins involved in DNA replication, such as DNA polymerase, DNA helicase, DNA resolvase, and DNA topoisomerase, were also found to be deleted in UFPR1 with copies of these genes present in other genomic DNA regions, indicating that their deletion did not affect UFPR1’s replication. In agreement with this observation, UFPR1 presented normal growth when cultured in vivo in the present report.
Likewise, deletion of qacEΔ1, sul1, and tetB genes were found in SH UFPR1 strain. The former gene has been associated to resistance against a large spectrum of cationic compounds such as intercalating dyes, diamidines, and biguanides (39), sul1 to tolerance to sulfonamide (40) and tetB to tetracycline efflux (41). Although other genes linked to tetracycline tolerance were found in UFPR1 strain, such as tetA class B and tetA class C genes, the deletion of tetB gene could explain the intermediate tetracycline resistance observed in this report. The deletion of LysR gene in UFPR1 may be linked to its low pathogenicity since microorganisms lacking it have been found to be less virulent (34, 42).
Gene deletion is used as an evolutionary process in bacteria in which small genomes have evolved from large genomes, with natural selection acting as a significant driver of gene loss and reductive genome evolution (43). However, bacteria genome could be increased by the acquisition of genetic fragments transferred horizontally (44).
Interestingly, no sequence from plasmids among those assembled was observed. Genome stability is threatened by transposons, which are able to create repetitive sequence islands that can initiate ectopic recombination (45). The present results showed that UFPR1 has several deletions of genes of various transposases suggesting that DNA transposition could be decreased in this strain. The UFPR1 genome presented five different fragments that are absent in SL476 but present in other Salmonella genomes evidencing the fact that they are not exclusively of UFPR1. In those five fragments, important genes related to some of the phenotypic characteristics reported herein in UFPR1 were found, such as the Bgt and Bgr genes that are related to serotype transformation; and the rpoS gene, linked to sensitivity to lower temperatures. The proteins produced by the expression of Bgt and Bgr genes are related to the glucosylation of the O-antigen repeated units of lipopolysaccharide (LPS) and are correlated to serotype conversion in Shigella flexneri and also in Salmonella (46). The presence of this gene is also correlated to the increase of virulence and resistance to oxidative stress (47).
Activation of rpoS gene is involved in cold sensitivity of Salmonella enterica serovar Typhimurium (48). The rpoS gene codifies for an alternative sigma factor that regulates many cellular responses to environmental conditions, such as heat, alkaline, and acid stress. Mutations of this gene have been detected in pathogenic bacteria (49). Bacteria are subjected to acid stress situations such as to the extreme low pH of the stomach and to the organic acids that are produced in large quantities in the gut including acetic, propionic, and butyric acids. In both situations, bacteria activates mechanisms for acid tolerance response (ATR), for which rpoS is a key regulator, thus minimizing the lethal effects of the acid stress. However, Salmonella rpoS mutant fails to provide the same level of protection when compared to a wild-type strain. Therefore, rpoS mutant is ineffective to sustain the ATR resulting in rapid cell death when exposed to pH 3.0 (50, 51). The product of the rpoS gene regulates the virulence gene expression in Salmonella Typhimurium in response to conditions encountered in the host tissue. Mutations in the rpoS gene yield that bacteria unable to develop a complete ATR significantly reducing its virulence potential (51). The presence of rpoS gene in the UFPR1 strain could be involved to the resistance to SCOA found. It has been observed that the alternative sigma factor clearly plays an important role in protecting Salmonella enterica serovar Typhimurium against weak acids (52).
In a recent study, Dhanani et al. (15) demonstrated in an in vivo experiment that the resistant genes found in SL476 may explain its pathogenicity, colonization ability and persistence in chickens. The absence of several genes involved in tolerance to antibiotics and in the efflux of salt reported in this paper, could explain why UFPR1 was in general susceptible to antibiotics, but resistant to SCOA likely due to the presence of rpoS gene.
The use of SCOA can be effective for the control of Salmonella in broilers being an important tool for the poultry industry (6, 21). However, testing for the presence of the ATR gene in Salmonella strains could help avoiding the misuse of these substances.
In this report, we used comparative genomics to study some of the genotypic peculiarities of UFPR1 linking those findings to phenotypic observations such as its tolerance to SCOA and sensitivity to various antibiotics. The comparison of several genome sequences could reveal important aspects of the evolution of the different Salmonella strains, and in a more accurate analysis, help identifying single nucleotide polymorphisms involved in potentially unknown pathways that could be relevant to the study of the metabolism of Salmonella and its control. Our findings can also help developing effective strategies to control this agent in broilers, thus preventing food-borne disease in humans.
Salmonellosis causes great economic damage to the poultry industry worldwide from strains that are either pathogenic and non-pathogenic to humans. Developing methodologies that reliably and promptly differentiate pathogenic from non-pathogenic strains could ameliorate that economic loss.
Conclusion
The infection of Salmonella enterica serovar Heildelberg UFPR1 in broilers did not affect zootechnical performance and promoted a mild inflammatory reaction on cecum and liver.
The use of different SCOA in drinking water or feed was ineffective against Salmonella enterica serovar Heildelberg UFPR1 strain in the present layout.
The genomic findings showed several differences between SH UFPR1 strain and the pathogenic SL476 strain.
The absence of several genes involved in antibiotics resistance and in salt efflux; along with the presence of rpoS gene, could explain the overall high susceptibility of UFPR1 to the antibiotics tested, and its resistance to SCOA. This information can help the poultry industry on designing SH control programs targeted against this specific strain.
Understanding the phenotypic and genotypic differences among Salmonella strains could help improving our knowledge on their metabolism, which could ultimately lead to their effective control.
Ethics Statement
The experiments were approved by the Ethical Committee of Agricultural Sector of Federal University of Parana under approval number: 037/2016 and 014/2016.
Author Contributions
ES—isolation of Salmonella Heildelber strain UFPR1, design in vivo and in vitro trial, analysis and interpretation of data, drafting and revising it critically, and final approval of the version. RH—design in vivo and in vitro trial, analysis of data, drafting the work and revising it critically, and final approval of the version. JW—carry on the in vivo and in vitro trial, analysis of data, drafting the work and revising it critically, and final approval of the version. RG-E—design in vivo and in vitro trial, analysis, interpretation of data for the work, drafting the work, and final approval of the version. MC, CF, PM, AC—analysis, interpretation of genomic data and revising it critically, and final approval of the final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor and reviewer KG declared their shared affiliation and involvement as co-editors in the Research Topic.
Funding
Novus International, Inc., Indaiatuba, SP, Brazil.
Supplementary Material
The Supplementary Material for this article can be found online at http://www.frontiersin.org/article/10.3389/fvets.2017.00184/full#supplementary-material.
References
1. Abd El Ghany M, Shi X, Li Y, Ansari HR, Hill-Cawthorne GA, Ho YS, et al. Genomic and phenotypic analyses reveal the emergence of an atypical Salmonella enterica serovar Senftenberg variant in China. J Clin Microbiol (2016) 54:2014–22. doi:10.1128/JCM.00052-16
2. Barrow PA, Simpson JM, Lovell MA, Binns MM. Contribution of Salmonella Gallinarum large plasmid toward virulence in fowl typhoid. Infect Immun (1987) 55(2):388–92.
3. Muniz E, Pickler L, Lourenço MC, Kraieski LA, Fiaga DAM, Westphal P, et al. Avaliação da resposta imunológica da mucosa intestinal de frangos de corte desafiados com diferentes sorovares de Salmonella. Pesqui Vet Bras (2015) 35:241–8. doi:10.1590/S0100-736X2015000300006
4. Kogut MH, Swaggerty CL, Byrd JA, Selvaraj R, Arsenault RJ. Chicken-specific kinome array reveals that Salmonella enterica serovar Enteritidis modulates host immune signaling pathways in the cecum to establish a persistence infection. Int J Mol Sci (2016) 17:1207–27. doi:10.3390/ijms17081207
5. Carter AJ, Adams MR, Woodward MJ, La Ragione RM. Control strategies for Salmonella colonization of poultry: the probiotic perspective. Food Sci Technol Bull Funct Foods (2009) 5(9):103–15. doi:10.1616/1476-2137.15682
6. Van Immerseel F, Russell JB, Flythe MD, Gantois I, Tim-bermont L, Pasmans F, et al. The use of organic acids to combat Salmonella in poultry: a mechanistic explanation of the efficacy. Avian Pathol (2006) 35:182–8. doi:10.1080/03079450600711045
7. CDC (Centers for Disease Control and Prevention). National Salmonella Surveillance Overview. (2013). Available from: https://www.cdc.gov/nationalsurveillance/pdfs/salmonella-annual-report-2013-508c.pdf
8. Hoffmann M, Zhao S, Pettengill J, Luo Y, Monday SR, Abbott J, et al. Comparative genomic analysis and virulence differences in closely related Salmonella enterica serotype Heidelberg isolates from humans, retail meats, and animals. Genome Biol Evol (2014) 6:1046–68. doi:10.1093/gbe/evu079
9. FDA (Food and Drug Administration). National Antimicrobial Resistance Monitoring System–Enteric Bacteria (NARMS). NARMS Integrated Report. (2013). Available from: https://www.fda.gov/downloads/AnimalVeterinary/SafetyHealth/AntimicrobialResistance/NationalAntimicrobialResistanceMonitoringSystem/UCM453398.pdf
10. Hofer E, Silva Filho SJ, Reis EMF. Prevalência de sorovares de Salmonella isolados de aves no Brasil. Pesqui Vet Bras (1997) 17(2):55–62. doi:10.1590/S0100-736X1997000200003
11. Voss-Rech D, Vaz CSL, Alves L, Coldebella A, Leão JA, Rodrigues DP, et al. A temporal study of Salmonella enterica serotypes from broiler farms in Brazil. Poult Sci (2015) 94:433–41. doi:10.3382/ps/peu081
12. Allard MW, Luo Y, Strain E, Li C, Keys CE, Son I, et al. High resolution clustering of Salmonella enterica serovar Montevideo strains using a next-generation sequencing approach. BMC Genomics (2012) 13:32. doi:10.1186/1471-2164-13-32
13. Ogunremi D, Blais B, Huang H, Wang L, Elmufti M, Allain R, et al. Draft genome sequences of two strains of Salmonella enterica serovar Typhimurium displaying different virulence in an experimental chicken model. Genome Announc (2017) 5(6):e01526–16. doi:10.1128/genomeA.01526-16
14. Reimschuessel R, Grabenstein M, Guag J, Nemser SM, Song K, Qiu J, et al. Multi-laboratory survey to evaluate Salmonella prevalence in diarrheic and non-diarrheic dogs and cats-in the USA between 2012 and 2014. J Clin Microbiol (2017) 55(5):1350–68. doi:10.1128/JCM.02137-16
15. Dhanani AS, Block G, Dewar K, Forgetta V, Topp E, Beiko RG, et al. Genomic comparison of non-typhoidal Salmonella enterica serovars Typhimurium, Enteritidis, Heidelberg, Hadar and Kentucky isolates from broiler chickens. PLoS One (2015) 10:e0128773. doi:10.1371/journal.pone.0128773
16. Labbe G, Ziebell K, Bekal S, Macdonald KA, Parmley EJ, Agunos A, et al. Complete genome sequences of 17 Canadian isolates of Salmonella enterica subsp. enterica serovar Heidelberg from human, animal, and food sources. Genome Announc (2016) 4(5):e00990–16. doi:10.1128/genomeA.00990-16
17. Donado-Godoy P, Bernal JF, Rodriguez F, Gomez Y, Agarwala R, Landsman D, et al. Genome sequences of multidrug-resistant Salmonella enterica serovar Paratyphi B (dT+) and Heidelberg strains from the Colombian poultry chain. Genome Announc (2015) 3(5):e01265–15. doi:10.1128/genomeA.01265-15
18. Fricke WF, Mammel MK, McDermott PF, Tartera C, White DG, Leclerc JE, et al. Comparative genomics of 28 Salmonella enterica isolates: evidence for CRISPR-mediated adaptive sublineage evolution. J Bacteriol (2011) 193:3556–68. doi:10.1128/JB.00297-11
19. Brasil MAPA (Ministério da Agricultura, Pecuária e Abastecimento). Instrução Normativa n° 6 publicada em 26 de agosto de 2003. (2003). Available from: http://extranet.agricultura.gov.br/sislegis-consulta/consultarLegislacao.do?operacao=visualizar&id=2851
20. Rostagno HS. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. Viçosa: UFV University Press (2011). p. 3–252.
21. Pickler L, Hayashi RM, Lourenço MC, Miglino LB, Caron LF, Beirão BCB, et al. Avaliação microbiológica, histológica e imunológica de frangos de corte desafiados com Salmonella Enteritidis e Minnesota e tratados com ácidos orgânicos. Pesqui Vet Bras (2012) 32:27–36. doi:10.1590/S0100-736X2012000100006
22. Kraieski AL, Hayashi RM, Sanches A, Almeida GC, Santin E. Effect of aflatoxin experimental ingestion and Eimeira vaccine challenges on intestinal histopathology and immune cellular dynamic of broilers: applying an intestinal health index. Poult Sci (2017) 96(5):1078–87. doi:10.3382/ps/pew397
23. Santin E, Kraieski AL, Hayashi RM, Inventors; Universidade Federal do Paraná, Assignee. ISI – I see Inside: Sistema de avaliação econômica de saúde em aves produzidas em sistema intensivos. Brazil Patent INPI BR 10 2015 003601 9 (2015).
24. Ramakers C, Ruiter JM, Deprez RH, Moorman AF. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett (2003) 339:62–6. doi:10.1016/S0304-3940(02)01423-4
25. Humphrey T. Salmonella, stress responses and food safety. Nat Rev Microbiol (2004) 2(6):504–9. doi:10.1038/nrmicro907
26. CLSI (Clinical and Laboratory Standards Institute). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standards. CLSI/NCCLS document M31-A2. (2002). Available from: http://www.dbt.univr.it/documenti/OccorrenzaIns/matdid/matdid485539.pdf
27. Berlin K, Koren S, Chin CS, Drake JP, Landolin JM, Phillippy AM. Assembling large genomes with single-molecule sequencing and locality-sensitive hashing. Nat Biotechnol (2015) 33:623–30. doi:10.1038/nbt.3238
28. Darling AE, Mau B, Perna NT. Progressive mauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One (2010) 5(6):e11147. doi:10.1371/journal.pone.0011147
29. Edirmanasinghe R, Finley R, Parmley EJ, Avery BP, Carson C, Bekal S, et al. A whole genome sequencing approach to study cefoxitin-resistant Salmonella enterica serovar Heidelberg from various sources. Antimicrob Agents Chemother (2017) 61(4):e01919–16. doi:10.1128/AAC.01919-16
30. Kurtz S, Phillippy A, Delcher AL, Smoot M, Shumway M, Antonescu C, et al. Versatile and open software for comparing large genomes. Genome Biol (2004) 5:R12. doi:10.1186/gb-2004-5-2-r12
31. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics (2014) 30:2068–9. doi:10.1093/bioinformatics/btu153
32. Carver T, Thomson N, Bleasby A, Berriman M, Parkhill J. DNAPlotter: circular and linear interactive genome visualization. Bioinformatics (2009) 25:119–20. doi:10.1093/bioinformatics/btn578
33. Koren S, Walenz BP, Berlin K, Miller JR, Bergman NH, Phillippy AM. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res (2017) 27(5):722–36. doi:10.1101/gr.215087.116
34. Maddocks SE, Oyston PC. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology (2008) 154:3609–23. doi:10.1099/mic.0.2008/022772-0
35. Berndt A, Wilhelm A, Jugert C, Pieper J, Sachse K, Methner U. Chicken cecum immune response to Salmonella enterica serovars of different levels of invasiveness. Infect Immun (2007) 75(12):5993–6007. doi:10.1128/IAI.00695-07
36. Shanmugasundaram R, Kogut MH, Arsenault RJ, Swaggerty CL, Cole K, Reddish MJ. Effect of Salmonella infection on cecal tonsil T regulatory T cell properties in the chicken. Poult Sci (2015) 94(8):1828–35. doi:10.3382/ps/pev161
37. Wright GD, Thompson PR. Aminoglycoside phosphotransferases: proteins, structure, and mechanism. Front Biosci (1999) 4:D9–21. doi:10.2741/Wright
38. Shaikh S, Fatima J, Shakil S, Rizvi SM, Kamal MA. Antibiotic resistance and extended spectrum beta-lactamases: types, epidemiology and treatment. Saudi J Biol Sci (2015) 22:90–101. doi:10.1016/j.sjbs.2014.08.002
39. Jaglic Z, Cervinkova D. Genetic basis of resistance to quaternary ammonium compounds – the qac genes and their role: a review. Vet Med (2012) 6:275–81.
40. Byrne-Bailey KG, Gaze WH, Kay P, Boxall AB, Hawkey PM, Wellington EM. Prevalence of sulfonamide resistance genes in bacterial isolates from manured agricultural soils and pig slurry in the United Kingdom. Antimicrob Agents Chemother (2009) 53:696–702. doi:10.1128/AAC.00652-07
41. Li XZ, Nikaido H. Efflux-mediated drug resistance in bacteria: an update. Drugs (2009) 69:1555–623. doi:10.2165/11317030-000000000-00000
42. Sheehan LM, Budnick JA, Blanchard C, Dunman PM, Caswell CC. A LysR-family transcriptional regulator required for virulence in Brucella abortus is highly conserved among the alpha-proteobacteria. Mol Microbiol (2015) 98:318–28. doi:10.1111/mmi.13123
43. Koskiniemi S, Sun S, Berg OG, Andersson DI. Selection-driven gene loss in bacteria. PLoS Genet (2012) 8(6):e1002787. doi:10.1371/journal.pgen.1002787
44. Davison J. Genetic exchange between bacteria in the environment. Plasmid (1999) 42:73–91. doi:10.1006/plas.1999.1421
45. Kazazian HHJ. Mobile elements: drivers of genome evolution. Science (2004) 303:1626–32. doi:10.1126/science.1089670
46. Mavris M, Manning PA, Morona R. Mechanism of bacteriophage SfII-mediated serotype conversion in Shigella flexneri. Mol Microbiol (1997) 26:939–50. doi:10.1046/j.1365-2958.1997.6301997.x
47. Szemes T, Vlkova B, Minarik G, Drahovska H, Turna J, Celec P. Does phage P22 contribute to resistance of Salmonella to oxidative stress? Med Hypotheses (2012) 79:484–6. doi:10.1016/j.mehy.2012.06.030
48. Knudsen GM, Nielsen MB, Thomsen LE, Aabo S, Rychlik I, Olsen JE. The role of ClpP, RpoS and CsrA in growth and filament formation of Salmonella enterica serovar Typhimurium at low temperature. BMC Microbiol (2014) 14:208. doi:10.1186/s12866-014-0208-4
49. Bhagwat AA, Tan J, Sharma M, Kothary M, Low S, Tall BD, et al. Functional heterogeneity of RpoS in stress tolerance of enterohemorrhagic Escherichia coli strains. Appl Environ Microbiol (2006) 72:4978–86. doi:10.1128/AEM.02842-05
50. Foster JW. The acid tolerance response of Salmonella Typhimurium involves transient synthesis of key acid shock proteins. J Bacteriol (1993) 175:1981–7. doi:10.1128/jb.175.7.1981-1987.1993
51. Lee IS, Lin J, Hall HK, Bearson B, Foster JW. The stationary-phase sigma factor sigma S (RpoS) is required for a sustained acid tolerance response in virulent Salmonella Typhimurium. Mol Microbiol (1995) 17:155–67. doi:10.1111/j.1365-2958.1995.mmi_17010155.x
Keywords: antibiotic, comparative genomics, organic acids, resistance, UFPR1 strain
Citation: Santin E, Hayashi RM, Wammes JCS, Gonzalez-Esquerra R, Carazzolle MF, Freire CCM, Monzani PS and Cunha AF (2017) Phenotypic and Genotypic Features of a Salmonella Heidelberg Strain Isolated in Broilers in Brazil and Their Possible Association to Antibiotics and Short-Chain Organic Acids Resistance and Susceptibility. Front. Vet. Sci. 4:184. doi: 10.3389/fvets.2017.00184
Received: 20 May 2017; Accepted: 11 October 2017;
Published: 01 November 2017
Edited by:
Michael Kogut, Agricultural Research Service (USDA), United StatesReviewed by:
Lisa Bielke, The Ohio State University Columbus, United StatesKenneth James Genovese, Agricultural Research Service (USDA), United States
Copyright: © 2017 Santin, Hayashi, Wammes, Gonzalez-Esquerra, Carazzolle, Freire, Monzani and Cunha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elizabeth Santin, QmVzYW50aW5AaG90bWFpbC5jb20=