 Jose Denis-Robichaud1†
Jose Denis-Robichaud1† Stephen J. LeBlanc1
Stephen J. LeBlanc1 Andria Jones-Bitton1
Andria Jones-Bitton1 Bruna F. Silper2
Bruna F. Silper2 Ronaldo L. Aoki Cerri2*
Ronaldo L. Aoki Cerri2*- 1Department of Population Medicine, University of Guelph, Guelph, ON, Canada
- 2Applied Animal Biology, Faculty of Land and Food Systems, University of British Columbia, Vancouver, BC, Canada
The ability of automated activity monitoring (AAM) systems to detect estrus is influenced by multiple variables. The luteal phase (LP) preceding estrus has been shown to be important for hormones release, and behavior during estrus in ruminants, but its impact on activity as measured by AAM systems has not been studied. The objective of this study was to investigate the impact of the length of the preceding LP on the intensity and duration of estrus as measured by AAM, and on the expression of estrogen receptor alpha (ERα) in the endometrium. A total of 60 cows between 46 and 53 days in milk were assigned to either a normal or a short length LP. Length of the LP was insured by the use of hormonal synchronization programs. Cows in both treatments were matched on parity, milk production, body condition score (BCS), and lameness score (assessed at enrolment). Expression of ERα receptors in the endometrium were evaluated by nuclear staining by immunohistochemistry of biopsies. Estrus was defined as the day on which the AAM system detected estrus. Cows that were not synchronized by the protocol or that were not detected in estrus by the AAM systems were excluded, which left 21 and 11 cows for analyses in the normal and short length LP, respectively. Peak activity index of estrus, duration of estrus, and expression of ERα were outcomes for multivariable linear regression models. Cows with short length LP tended to have lower peak activity at estrus, but there was no significant effect of treatment when BCS was accounted for. Cows with BCS ≤ 2.5 had less intense and shorter estrus than cows with BCS ≥ 2.75. There was no association between the length of the LP and the expression of ERα in the endometrium.
Introduction
Automated activity monitoring (AAM) systems have been developed and refined as an estrus detection aid in dairy cattle (1–3). They are used on many dairy farms to manage reproduction and decrease the time spent on estrus detection (4, 5). The ability of a system to identify cows in estrus, and the probability of pregnancy for inseminations following an AAM system alert, have been shown to affect the system's economic value in reproduction management programs (6, 7). Previous studies found the sensitivity of a collar-mounted AAM systems (Heatime®, H-Tags, SCR Engineers, Netanya, Israel) varied between 52 and 72%, and its positive predictive value between 64 and 94% (8–11), which is comparable to other systems (11–13). This variability of one system depends on what gold standard was used, but also on the threshold used, and the conditions it is used in (8–13). The ability and accuracy of an AAM system to identify cows in estrus do not depend only on the system itself. Cow factors such as parity, milk production, and body condition score (BCS), as well as concentrations of estradiol and progesterone (P4) at estrus have been shown to be associated with intensity and duration of estrus as detected by AAM systems, and can therefore affect the ability of the system to identify estrus (14–16). Multiparous cows have been shown to have less intense and shorter estrus than primiparous cows, as have thin cows (BCS ≤ 2.5) compared to normal and fat cows (BCS ≥ 2.75), and high-producing cows (≥39 kg milk/days) compared to low-producing cows [ ≤ 31 kg milk/days; (16)]. Parity and BCS are also reported to be associated with estradiol concentration at estrus, which can explain partially these associations (16). High estradiol and low P4 at estrus have been reported to be associated, although weakly, with intensity and duration of estrus as detected by AAM systems (15, 16).
Circulating estradiol concentration at estrus plays a key role in estrous behavior because it activates estrogen receptors in the area of the hypothalamus regulating sexual behavior (17–19). Low concentrations of P4 (because P4 blocks the effect of estradiol on the hypothalamus), and expression of estrogen receptors (ER) in the hypothalamus are necessary for estrus expression (19–21). The expression of ER in the hypothalamus has been shown to vary according to the stage of the estrous cycle and circulating concentrations of estradiol and P4 (18, 19, 22, 23). Moreover, exposure to P4 prior to estrus has been shown to be positively associated with intensity of estrus expression in ewes (24). The expression of ER in the reproductive tract of different species is reported to be regulated by circulating P4 (25, 26). Specifically, ER alpha (ERα) decreased during the luteal phase, likely due to P4 exposure, and reached its inflection point around days 14 of the cycle.
Implementing a controlled trial with binary reproductive outcome variables such as detection of estrus or not requires enrolling a high number of cows (27). Preliminary data is important to evaluate the feasibility of the trial by exploring a similar hypothesis and identifying potential problems and limitations. We hypothesized that the duration of exposure to P4 before estrus (i.e., the length of the LP) is associated with the intensity and duration of estrus, as detected by AAM systems, irrespective of the estradiol concentration at estrus. The objective of this pilot study was to evaluate the association between the length of the LP and the intensity and duration of estrus, as detected by AAM. A secondary objective was to compare the expression of ERα in the endometrium following LP of different length.
Materials and Methods
Animals
This study was evaluated and approved by the University of British Columbia (UBC) Animal Care Committee (Protocol # 14-0019). Cows were enrolled in this controlled trial between 46 and 53 days in milk (DIM), were housed at the UBC Dairy Education and Research Centre (Agassiz, BC, Canada) in a free-stall barn, and stayed in the same pen throughout the study. At enrolment, cows were assessed for lameness using a standardized 5-point score [1 and 2: not lame, 3 and greater: lame; (28)] and for BCS using a 1 (emaciated) to 5 (fat) scoring system; (29). Parity and predicted 305 days milk production at first dairy herd improvement association (DHIA) test were obtained for all enrolled cows from DairyComp 305 (Valley Agricultural Software, Tulare, CA, United States). Cows were assigned to either the normal or short length LP (LP length was controlled by hormonal injections; Figure 1) such that they were matched in the two treatment groups by parity (first, second, or third and greater lactation), milk production quartile, BCS (≤ 2.5, 2.75–3.25, or ≥ 3.5), and lameness status (lame, or not lame).
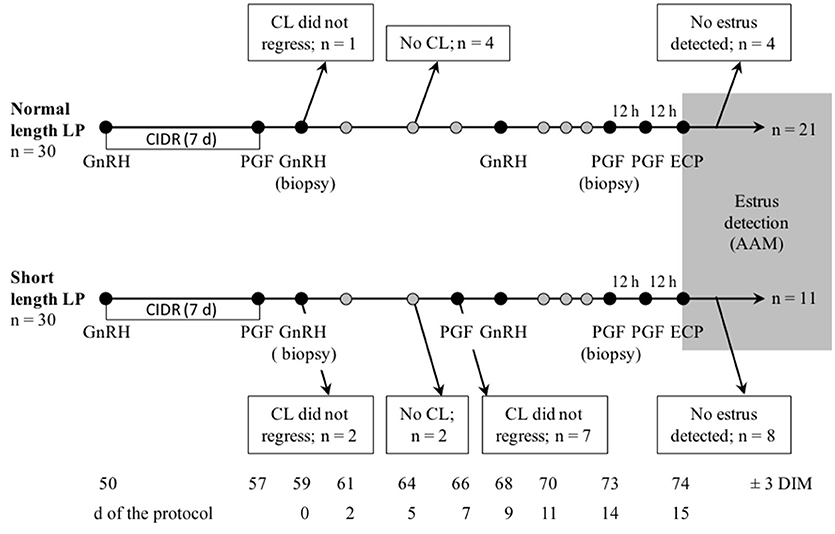
Figure 1. Schematic representation of sampling and treatments, and flow of the number of lactating cows enrolled in a controlled trial to compare estrus detected by AAM systems after a short or normal length luteal phase (LP). A total of 60 cows (30 per treatment) were enrolled at 46–53 DIM in one of two treatments: normal or short length LP. For all cows, a P4 vaginal implant (CIDR) was inserted and 100 μg of gonadorelin (GnRH) was administered. The implant was removed 7 days later and 500 μg of cloprostenol (PGF2α) was administered. Day 0 of the protocol was two d later when the second GnRH was administered. On 7 days of the protocol, cows with short length LP received PGF2α. All cows received GnRH on 9 days of the protocol, then two injections of PGF2α 12 h apart on 14 days, and finally 0.5 mg of estradiol cypionate (ECP) on 15 days (12 h after the last PGF2α). Estrus expression after 14 days (gray area) was recorded using a collar-mounted activity system (Heatime®, H-Tags, SCR Engineers, Netanya, Israel). Samples for serum progesterone were taken on 0, 2, 5, 7, 9, days and daily from days 11 to estrus, as detected by AAM, and ultrasound of the ovaries was done on 2, 5, 9, 12, 14, days and daily from days 16 to estrus. On 0 and 14 days, endometrial biopsies were taken on a subsample of cows (n = 18).
Treatments
To insure the protocol (summarized in Figure 1) started at 59 ± 3 DIM but did not begin with the first cycle of the cow, all cows received a vaginal implant containing 1.38 g of P4 (P4; CIDR, Zoetis, Kirkland, QC, Canada) and an intramuscular (i.m.) injection of 100 μg of gonadorelin (GnRH; Factrel, Zoetis, Kirkland, QC, Canada; 2 ml) at enrolment. After 7 days, the implant was removed, and cows received an i.m. injection of 500 μg of cloprostenol (PGF2α; Estrumate, Merck Animal Health, Kirkland, QC, Canada; 2 ml). The protocol started (0 days) 2 days later, when all cows received a second injection of GnRH (as above). Cows with a short length LP received an injection of PGF2α (as above) on 7 days. All cows then received an injection of GnRH on 9 days, two injections of PGF2α 12 h apart on 14 days, and finally 0.5 mg of estradiol cypionate i.m. (ECP; Estrus, Rafter8, Calgary, AB, Canada; 0.5 ml) on 15 days, 12 h after the last PGF2α. With these protocols, the LP preceding the estrus in the short and normal length LP were 5 and 14 days, respectively.
Examinations, Blood Sampling, and Analyses
Blood samples (10 mL) were taken on 0, 2, 5, 7, 9 days and daily from 11 days to estrus, from the coccygeal vessels into a sterile tube without anticoagulant (Vacutainer, Becton Dickinson, Franklin Lakes, NJ, United States). On 15 days, a blood sample (10 mL) was taken from the coccygeal vessels into a sterile tube with EDTA (Vacutainer, Becton Dickinson, Franklin Lakes, NJ, United States). Samples were kept chilled and allowed to clot. Within 5 h of blood collection, samples were centrifuged to harvest serum or plasma, which was frozen at −80°C. Serum P4 concentrations from 0, 2, 5, 7, 9, and 11 days to estrus were measured using a commercial ELISA kit Ovucheck Plasma; Biovet, St-Hyacinthe, Quebec; (30). This monoclonal antibody kit uses optical density of standards and serum samples read at 405 nm in a microplate absorbance reader. The range of quantification of the test is 0.55–10.45 ng/mL, and the intra-assay coefficient of variation (CV) in the present study was 11.7%. Plasma estradiol concentrations from 15 days were measured using radioimmunoassay [Perkin Elmer NET517250UC; Prairie Diagnostic Services, Saskatoon, SK, Canada; (31)]. This radioimmunoassay kit uses scintillation of charcoal-stripped serum standards and plasma samples after competitive binding with an immunoserum. The range of quantification of the test is 0–200 pg/mL, and the intra-assay CV in the present study was 17.0%.
Ovaries of all cows were scanned with a portable ultrasound (Ibex Pro; E.I. Medical Imaging, Loveland, CO) using a 7.5 MHz linear-array rectal transducer, on 2, 5, 9, 12, 14, days and daily from 16 days to estrus. Follicles and corpus lutea (CL) were examined from several angles and the largest cross-section of ovarian structures ≥5 mm in diameter were recorded. Luteolysis was defined as a P4 ≥ 2 ng/mL on the day of a PGF2α injection, followed by P4 < 2 ng/mL in the next 2 days. The threshold of 2 ng/mL was used as per the instructions in the ELISA manufacturer's insert (Biovet, St-Hyacinthe, Quebec), and because the serum samples were obtained early in the estrous period.
Endometrial Biopsies and Immunohistochemistry
On 0 and 14 days of the protocol, endometrial biopsies were performed on a subset of cows (n = 9 per treatment). Epidural anesthesia was administered using 100 mg of lidocaine (Lidocaine HCl 2%, Vetoquinol, Lavaltrie, QC, Canada). The vulva was cleaned, and a disinfected guarded biopsy instrument (crocodile-type biopsy forceps, made by Aries Surgical, Davis, CA, United States) was introduced via the cervix into the body of the uterus by trans-rectal manipulation. A section of approximately 6 by 4 mm of the endometrium was taken on each d, then put for 24 h in 10% formalin solution (neutral buffered, HT501128, Sigma, Oakville, ON, Canada) at 4°C. Samples were then transferred to a 70% ethanol solution for storage at 4°C until they were dehydrated, embedded in paraffin wax, and sectioned for immunohistochemistry (IHC; Wax-It Histology Services, Vancouver, BC, Canada). Formalin-fixed paraffin-embedded sections of bovine endometrium were de-paraffinised and rehydrated with distilled water. They were retrieved in sodium citrate buffer (pH 6.0) via steamer and blocked with hydrogen peroxide in methanol and protein block to reduce background staining. Sections were then incubated with the primary antibody ERα (sc-7207; Santa Cruz Biotechnology, Dallas, TX, United States) at 1:100 or Rabbit IgG negative isotype control at 4°C overnight. The following day, the sections were incubated with the secondary antibody HRP Labeled Polymer Anti-Rabbit, developed with 3,3′-diaminobenzidine. They were then counterstained, dehydrated, and mounted.
Image acquisition from the biopsy slides was done using Nikon Eclipse e200 microscope. If possible, 3 glands from different areas of the slide were selected. As shown on Figure 2, blue cells were categorized as no staining, bluish-brown, and brown cells were categorized as weak to medium staining, and very dark brown cells were categorized as strong staining (32). Counts of cells in each category were done at 400X magnification for each gland, and the proportion of cells in each category was calculated.
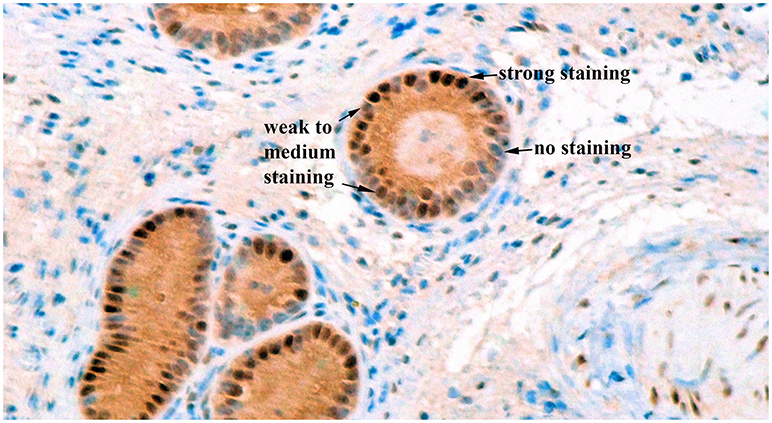
Figure 2. Immunohistochemical staining for estrogen receptor alpha in bovine endometrial glands at a 400X magnification. Blue cells were categorized as no staining, bluish-brown, and brown cells were categorized as weak to medium staining, and very dark brown cells were categorized as strong staining.
Automated Activity Monitoring
Cows were fitted with a collar-mounted AAM system (Heatime®, H-Tags, SCR Engineers, Netanya, Israel), which continuously monitored individual cow activity using an accelerometer detecting movement and recorded average activity for 2 h periods. Data were transmitted every 2 h by a wireless system to the farm computer. The raw activity data were processed into an index value accounting for the baseline values of the previous 14 days. The index represents a value close to the SD of the raw data and ranges from 0 to 100 (larger numbers denoting higher activity). An estrous event was defined as a period when the index was above a threshold of 35 according to the manufacturer recommendation. The duration of estrus was defined as the time (sum of the number of 2 h periods) the activity index was above the threshold, and the peak activity index of estrus was the highest value during this period. Individual cow data (cow identification, date and time of the onset of estrus, peak activity of estrus, and duration of estrus) were extracted from the backup files using a macro in Excel (Microsoft Corporation, Richmond, WA, United States). The day of estrus was defined as the day on which the AAM system signaled an estrus.
Statistical Analysis
The sample size (n = 30 per treatment) was calculated to identify a difference of 10 points (SD = 14) in peak activity index of estrus with 95% confidence and 80% power, accounting for 30% loss to follow up (33). This sample size was also sufficient to identify a difference of 2.5 h (SD = 3.5) in duration of estrus. Cows that did not synchronize through the protocol (CL did not regress following the first PGF2α or the second PGF2α with short length LP, or no CL developed after the removal of the CIDR and the GnRH), and that did not have estrus detected by the AAM system were excluded from the analysis (Figure 1).
All statistical analyses were performed using SAS 9.3 (SAS Institute, Cary, United States). Descriptive statistics (PROC FREQ) for treatment, parity, milk production, BCS, and lameness score were calculated for all enrolled cows, excluded cows, and included cows with normal and short length LP. Peak activity index of estrus and duration of estrus were outcomes for linear regression models (PROC GLM). The proportion of cells in the 3 staining categories for glands were outcomes for linear regression models with repeated measures structure to account for the day of the biopsy (PROC GLIMMIX). In all models, treatment was included as a fixed effect. Even though matching was used to allocate cows to their treatment, the unequal pattern of excluded animals between treatment led us to include parity, milk production, BCS, and lameness score as confounders if the estimate for treatment changed by more than 10% when the covariate was removed (34, 35). Interaction terms between treatment and the other variables in the final model were retained if their P < 0.05. Assumptions of the models (normality and homoscedasticity) were assessed graphically using standardized residuals. Outliers (residuals and Studentized residuals), extreme (leverage), and influential data (Cook's distance and DFFITS) were used to assess the fit of the models (33). Marginal means (least squares means (LSM) ± SE) were obtained for categorical variables.
Results
A total of 60 cows were enrolled in the study. Figure 1 represents the flow of animals throughout the study. Nine and 19 cows in the normal and short length LP, respectively, were excluded during the protocol (Figure 1). A total of 32 (53%) cows were identified in estrus and remained for statistical analyses. Table 1 presents the distribution of parity, milk production, BCS, and lameness score of cows enrolled, excluded, and included in the normal and short length LP.
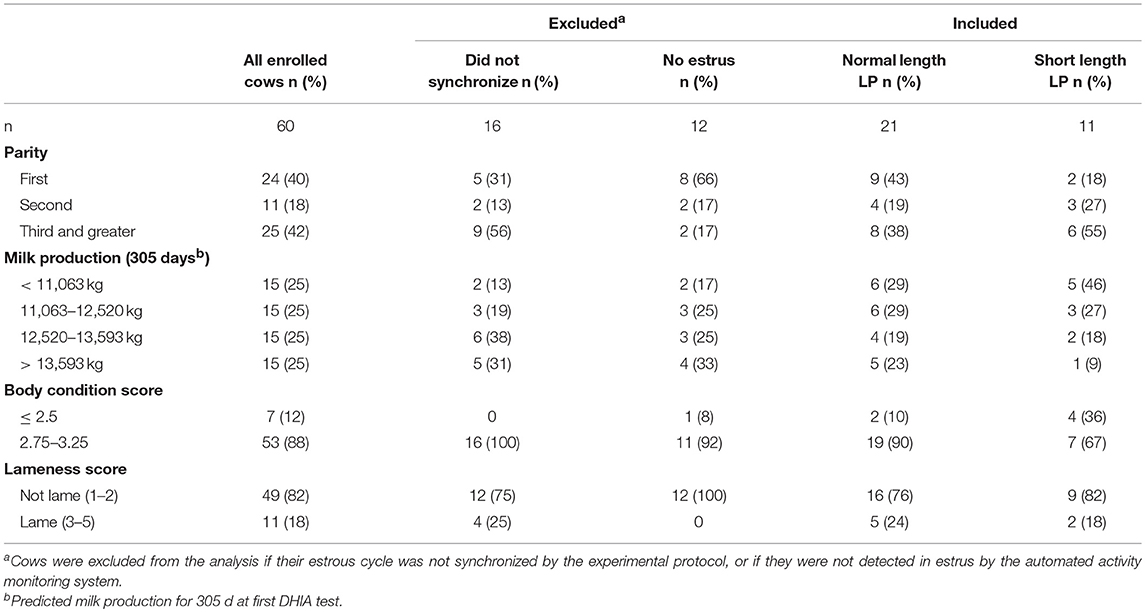
Table 1. Characteristics of cows enrolled in a controlled study of the effects of a normal or short luteal phase (LP) on estrus expression.
Progesterone profiles for the normal and short length LP are presented in Figure 3. By design, the area under the curve was different between treatments (normal length LP: 51.4 ± 2.8; short length LP: 29.0 ± 3.6; P < 0.01). Progesterone at 9 days was 4.7 ± 1.8 ng/mL and 0.9 ± 0.4, in the normal and short length LP, respectively (P < 0.01). Progesterone at estrus (as detected by the AAM system) was 0.7 ± 0.1, and 0.9 ± 0.1 ng/mL in the normal and short length LP, respectively (P = 0.38). The follicular waves for the normal and short length LP are summarized in Figure 4. The size of the dominant follicles did not differ between treatments (P = 0.33). Estradiol at 15 days (before ECP; normal length LP: 3.3 ± 0.4 pg/mL; short length LP: 3.4 ± 0.4 pg/mL; P = 0.88), and dominant follicle diameter at 16 days (normal length LP: 19.5 ± 0.9 mm; short length LP: 18.3 ± 1.1 mm; P = 0.37) did not differ between treatments. Cows ovulated before 16 days (normal length LP = 5; short length LP = 4), from 16 to 17 days (normal length LP = 4; short length LP = 7), from 17 to 18 days (normal length LP = 11; short length LP = 4), or after 18 days (normal length LP = 5; short length LP = 4).
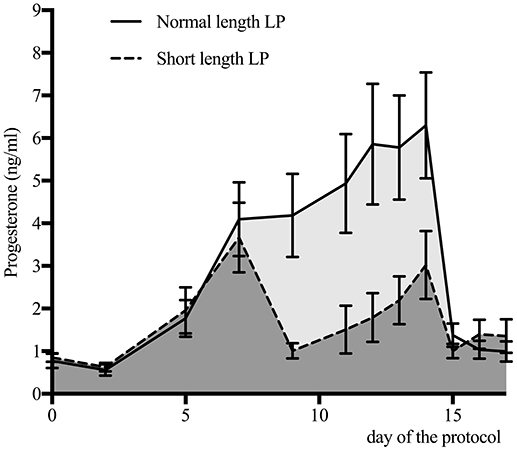
Figure 3. Serum progesterone concentration (mean ± 95% confidence intervals) for cows with a normal (n = 25) or a short length luteal phase (LP; n = 19). The area under the curve was 51.4 for the normal length LP (light gray area), and 29.0 for the short length LP (dark gray area).
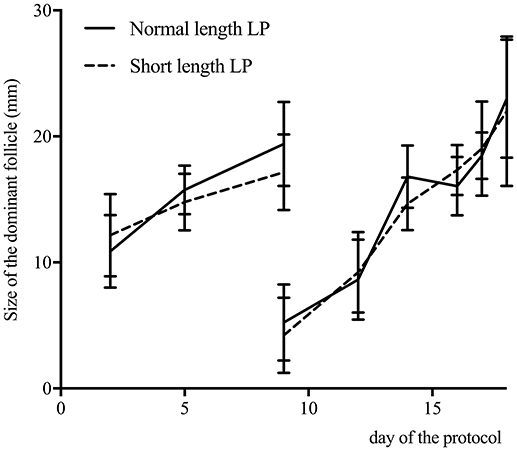
Figure 4. Size of the dominant follicle (mean ± 95% confidence intervals) determined by ultrasound of the ovaries on 2, 5, 9, 12, 14, days and daily from days 16 to estrus, for cows with a normal (n = 25) or a short length luteal phase (LP; n = 19). Cows ovulated before 16 days (normal length LP = 5; short length LP = 4), from 16 to 17 days (normal length LP = 4; short length LP = 7), from 17 to 18 days (normal length LP = 11; short length LP = 4), or after 18 days (normal length LP = 5; short length LP = 4).
Table 2 presents the predicted population marginal means (LSM) of activity in the normal and short length LP. For univariable and multivariable models, the peak of activity and duration of estrus tended (P > 0.05, but < 0.10) to be higher and longer, respectively, with normal length LP than in the short length LP. The one confounder that remained in the multivariable models was BCS, and no variables were statistically significant in the final multivariable models.
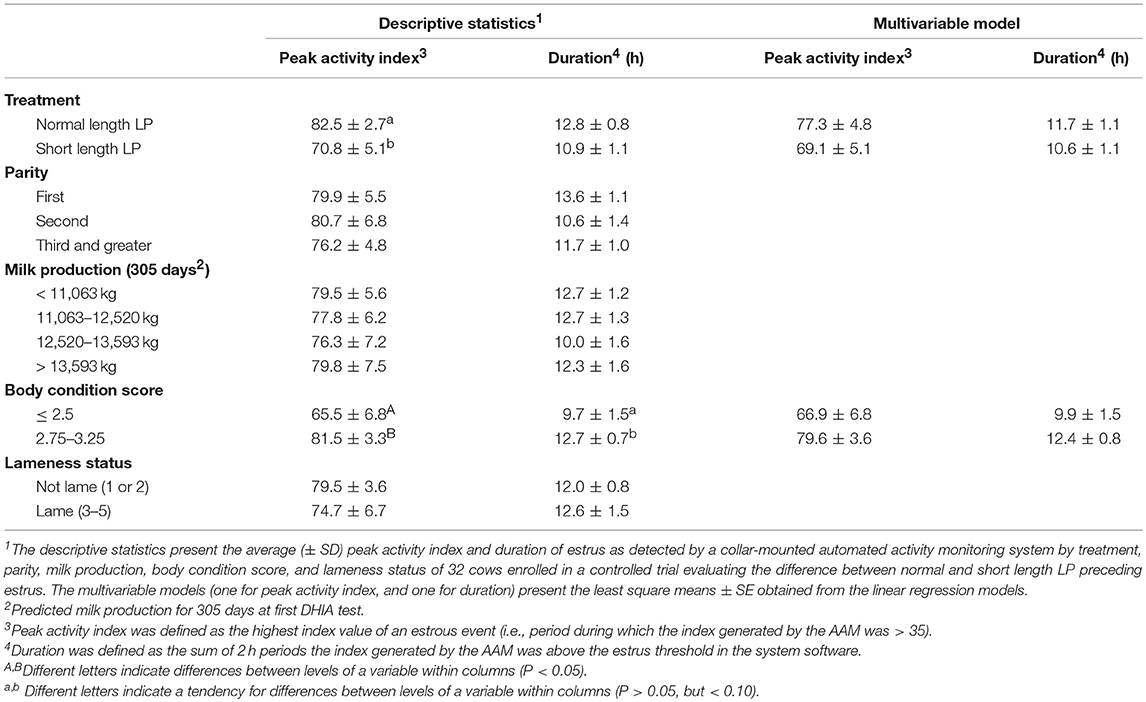
Table 2. Effect of the duration of the preceding luteal phase (LP) on estrus expression.
The expression of ERα did not differ by treatment (P > 0.10), and there was no difference between biopsy 1 and 2 (P > 0.10). Figure 5 presents the predicted population marginal means (LSM) of the proportion of cells in each staining category at biopsy 1 (0 days), and biopsy 2 (14 days), by treatment.
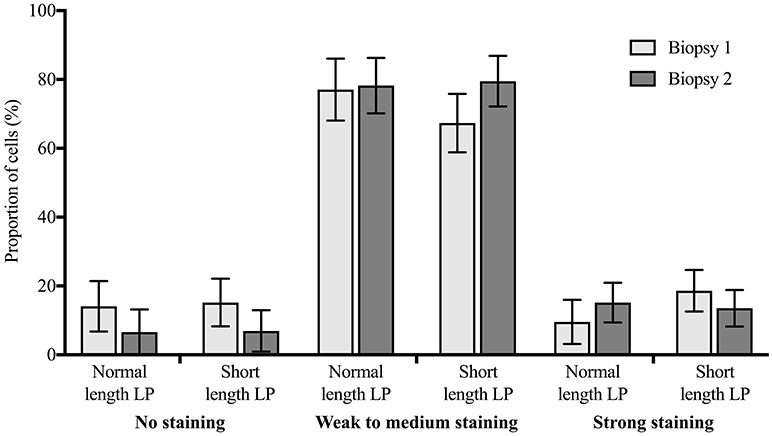
Figure 5. Distribution of immunohistochemical staining for estrogen receptor alpha (proportion ± 95% confidence intervals) in cells from endometrial biopsies 1 (0 days) and 2 (14 days) in 3 staining categories (no staining, weak to medium staining, strong staining), for cows with a short length luteal phase (LP; n = 9) or with a normal length LP (n = 9) preceding estrus. There was no statistically significant difference between biopsy 1 and 2, or between treatments.
Discussion
There was no statistically significant difference in peak activity index or duration of estrus between the normal and short length LP. The proportion of cows that did not synchronize during the protocol was higher than expected in the short length LP (n = 11; 37%), and also higher than in the normal length LP (n = 5; 17%). This could have resulted in a selection bias, as the cows included in the analyses might not be representative of the whole group. For example, cows that did not synchronize could have a common attribute that biases the results by not being represented in the final sample (33). Because of the small numbers, we could not identify a statistical difference between cows that did and did not synchronize, but numerically, cows that did not synchronize were of greater parity (third and greater), normal BCS (2.75–3.25), and had high milk yields (≥12,520 kg). Adding these covariates to the models, despite the matching used for treatment allocation, was used to control for this (35). The higher number of excluded cows with short length LP resulted mainly from cows that did not regress their CL after the PGF2α at 9 days (n = 7; 23% of the short length LP). It was shown that a second PGF2α 24 h apart increased complete CL regression, and could have decreased the exclusions in this case (36, 37). Due to these losses to follow up, the final models did not include as many cows as required according to the sample size calculation, even after accounting for the 30% anticipated loss to follow up. Numerically, the peak activity index and duration of estrus were greater for cows with normal length LP than with short length LP. It is unclear if the difference is not statistically significant because the length of the LP did not have a meaningful effect on estrus intensity and duration as detected by the AAM system, or because the number of cows ended up being insufficient. Post-hoc power calculation is not recommended (33), but intensity and duration had a post-hoc power of 96 and 76%, respectively. That said, the magnitude of the numerical differences (for intensity and duration) between the two treatments is likely too small to have a practical impact on the use of AAM systems. Even though the LP preceding estrus is essential in the estrous cycle for luteinizing hormone release and estradiol sensitization (23, 38), the present results do not support the hypothesis that the length of the LP affects the intensity and duration of estrus as detected by AAM systems.
Only a few studies on a limited number of ovariectomized ruminants support the necessity of exposure to some P4 prior to estrus for estrus expression (24, 39). Fabre-Nys and Martin (24) found a larger proportion of ewes showing estrous behavior, as well as more frequent demonstration of estrous behaviors in ewes that received P4 prior to estradiol, compared to those that did not. Cows that received P4 prior to estradiol were also more likely to show estrous behavior than those that did not, but there was no difference in estrous behavior between cows exposed to P4 for 5 days and those exposed for more than 5 days (39). In both studies, the duration of estrus was not addressed. The present study did not use cows with no previous exposure to P4, but normal or short exposure to P4 resulted in similar intensity and duration of estrus. The study design and sample size did not allow us to compare the difference in the proportion of cows identified in estrus by the AAM. The different length of the LP was not associated with the endometrial expression of ERα either. Circulating estradiol and P4 were previously shown to be associated with the expression of ERα (25, 26), but despite differences in circulating P4 in the week prior to estrus, we found no difference in uterine ERα between the normal and short length LP. As the present study was conducted in a commercial farm, there was no cows with no previous exposure to P4. Cows had a normal or short length LP, that was preceded by P4 exposure (CIDR; Figure 1). This could also have had mitigated the impact of the length of the LP on the intensity and duration of the estrus, as well as the expression of uterine ERα.
Circulating P4 concentrations during follicular growth have previously been shown to affect the diameter of the dominant follicle, and the circulating concentration of estradiol at estrus (40), which could affect estrus intensity and duration. In order to prevent this confounding effect, ECP was used to obtain a comparable peak of circulating estradiol between treatment (41). In the present study, BCS ≤ 2.5 was associated with less intense and shorter duration of estrous activity, as detected by an AAM system. Body condition was also reported to be associated with follicular growth and estradiol concentration at estrus (16). The use of ECP at 15 days of the protocol in the present study should have removed the effect BCS has on estradiol concentration. There was however, a significant association between BCS and estrous activity, which suggests that BCS is associated to estrus expression not only by modifying circulating P4 and estradiol concentrations, but also by mechanisms affecting both BCS and estrus expression such as negative energy balance. The association between BCS and estrus expression has been found before (8, 12, 16), but the exact mechanism involved remains unclear.
In conclusion, we did not detect a difference in intensity or duration of estrus as measured by AAM between a normal and a short length LP in dairy cows, but BCS was associated with intensity and duration of estrus.
Author Contributions
The study design was developed by the collaboration of all the authors. The data collection was done by JD-R, BS, and RA. JD-R performed the statistical analyses and wrote the first draft of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding
This project was funded by the WestGen (Abbotsford, BC, Canada), by the British Columbia Dairy Association (DIREC; Burnaby, BC, Canada), and by the Dairy Research Cluster Initiative [Dairy Farmers of Canada (Ottawa, Canada), Agriculture and Agri-Food Canada (Ottawa, Canada), the Canadian Dairy Network (Guelph, Canada), and the Canadian Dairy Commission (Ottawa, Canada)].
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the employees and the students at University of British Columbia Dairy Education and Research Center for contributing greatly to the on-farm sampling and laboratory work. Part of the results of this project was also published in JD-R's Ph.D. thesis (42).
Abbreviations
AAM, automated activity monitoring; BCS, body condition score; CL, corpus luteum; DIM, days in milk; DHIA, dairy herd improvement association; ERα, estrogen receptor alpha; GnRH, gonadotropin-releasing hormone; IHC, immunohistochemistry; LP, luteal phase; P4, progesterone; PGF2α, prostaglandin F2α.
References
1. Kiddy CA Variation in physical activity as an indication of estrus in dairy cows. J Dairy Sci. (1977) 60:235–43. doi: 10.3168/jds.S0022-0302(77)83859-9
2. At-Taras EE, Spahr SL. Detection and characterization of estrus in dairy cattle with an electronic heatmount detector and an electronic activity tag. J Dairy Sci. (2001) 84:792–8. doi: 10.3168/jds.S0022-0302(01)74535-3
3. Firk R, Stamer E, Junge W, Krieter J. Automation of oestrus detection in dairy cows: a review. Livestock Prod Sci. (2002) 75:219–32. doi: 10.1016/S0301-6226(01)00323-2
4. Neves RC, LeBlanc SJ. Reproductive management practices and performance of Canadian dairy herds using automated activity-monitoring systems. J Dairy Sci. (2015) 98:2801–11. doi: 10.3168/jds.2014-8221
5. Denis-Robichaud J, Cerri RL, Jones-Bitton A, LeBlanc SJ. Survey of reproduction management on Canadian dairy farms. J Dairy Sci. (2016) 99:9339–51 doi: 10.3168/jds.2016-1144
6. Giordano JO, Kalantari AS, Fricke PM, Wiltbank MC, Cabrera VE. A daily herd Markov-chain model to study the reproductive and economic impact of reproductive programs combining timed artificial insemination and estrus detection. J Dairy Sci. (2012) 95:5442–60. doi: 10.3168/jds.2011-4972
7. Galvão KN, Federico P, De Vries A, Schuenemann GM. Economic comparison of reproductive programs for dairy herds using estrus detection, timed artificial insemination, or a combination. J Dairy Sci. (2013) 96:2681–93. doi: 10.3168/jds.2012-5982
8. Aungier SP, Roche JF, Sheehy M, Crowe MA. Effects of management and health on the use of activity monitoring for estrus detection in dairy cows. J Dairy Sci. (2012) 95:2452–66. doi: 10.3168/jds.2011-4653
9. Valenza A, Giordano JO, Lopes Jr G, Vincenti L Amundson MC, Fricke PM. Assessment of an accelerometer system for detection of estrus and treatment with gonadotropin-releasing hormone at the time of insemination in lactating dairy cows. J Dairy Sci. (2012) 95:7115–27. doi: 10.3168/jds.2012-5639
10. Kamphuis C, DelaRue B, Burke CR, Jago J. Field evaluation of 2 collar-mounted activity meters for detecting cows in estrus on a large pasture-grazed dairy farm. J Dairy Sci. (2012) 95:3045–56. doi: 10.3168/jds.2011-4934
11. Holman A, Thompson J, Routly JE, Cameron J, Jones DN, Grove-White D, et al. Comparison of oestrus detection methods in dairy cattle. Vet Rec. (2011) 169:47. doi: 10.1136/vr.d2344
12. Lovendahl P, Chagunda MG. On the use of physical activity monitoring for estrus detection in dairy cows. J Dairy Sci. (2010) 93:249–59. doi: 10.3168/jds.2008-1721
13. Roelofs JB, van Eerdenburg FJCM, Soede NM, Kemp B. Pedometer readings for estrous detection and as predictor for time of ovulation in dairy cattle. Theriogenology. (2005) 64:1690–703. doi: 10.1016/j.theriogenology.2005.04.004
14. Morris MJ, Kaneko K, Walker SL, Jones DN, Routly JE, Smith RF, et al. Influence of lameness on follicular growth, ovulation, reproductive hormone concentrations and estrus behavior in dairy cows. Theriogenology (2011) 76:658–68. doi: 10.1016/j.theriogenology.2011.03.019
15. Aungier SP, Roche JF, Duffy P, Scully S, Crowe MA. The relationship between activity clusters detected by an automatic activity monitor and endocrine changes during the periestrous period in lactating dairy cows. J Dairy Sci. (2015) 98:1666–84. doi: 10.3168/jds.2013-7405
16. Madureira AM, Silper BF, Burnett TA, Polsky L, Cruppe LH, Veira DM, et al. Factors affecting expression of estrus measured by activity monitors and conception risk of lactating dairy cows. J Dairy Sci. (2015) 98:7003–14. doi: 10.3168/jds.2015-9672
17. Blache D, Fabre-Nys CJ, Venier G. Ventromedial hypothalamus as a target for oestradiol action on proceptivity, receptivity and luteinizing hormone surge of the ewe. Brain Res. (1991) 546:241–9. doi: 10.1016/0006-8993(91)91488-M
18. van Eerdenburg FJ, Daemen IA, van der Beek EM, van Leeuwen FW. Changes in estrogen-alpha receptor immunoreactivity during the estrous cycle in lactating dairy cattle. Brain Res. (2000) 880:219–23. doi: 10.1016/S0006-8993(00)02870-5
19. Gillies GE, McArthur S. Estrogen actions in the brain and the basis for differential action in men and women: a case for sex-specific medicines. Pharmacol Rev. (2010) 62:155–98. doi: 10.1124/pr.109.002071
20. Gümen A, Wiltbank MC. An alteration in the hypothalamic action of estradiol due to lack of progesterone exposure can cause follicular cysts in cattle. Biol Reprod. (2002) 66:1689–95. doi: 10.1095/biolreprod66.6.1689
21. Richter TA, Robinson JE, Lozano JM, Evans NP. Progesterone can block the preovulatory gonadotropin-releasing hormone/luteinising hormone surge in the ewe by a direct inhibitory action on oestradiol-responsive cells within the hypothalamus. J Neuroendocrinol. (2005) 17:161–9. doi: 10.1111/j.1365-2826.2005.01287.x
22. Robinson RS, Mann GE, Lamming GE, Wathes DC. Expression of oxytocin, oestrogen and progesterone receptors in uterine biopsy samples throughout the oestrous cycle and early pregnancy in cows. Reproduction (2001) 122:965–79. doi: 10.1530/rep.0.1220965
23. Forde N, Carter F, Fair T, Crowe MA, Evans AC, Spencer TE, et al. Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol Reprod. (2009). 81:784–94. doi: 10.1095/biolreprod.108.074336
24. Fabre-Nys C, Martin GB. Roles of progesterone and oestradiol in determining the temporal sequence and quantitative expression of sexual receptivity and the preovulatory LH surge in the ewe. J Endocrinol. (1991) 130:367–79. doi: 10.1677/joe.0.1300367
25. Martin I, Torres Neto R, Oba E, Buratini J, Binelli M. Laufer-Amorim R, et al. Immunohistochemical detection of receptors for oestrogen and progesterone in endometrial glands and stroma during the oestrous cycle in Nelore (Bos taurus indicus) cows. Reprod Domest Anim. (2008) 43:415–21. doi: 10.1111/j.1439-0531.2007.00928.x
26. Kimmins S, MacLaren LA. Oestrous cycle and pregnancy effects on the distribution of oestrogen and progesterone receptors in bovine endometrium. Placenta (2001) 22:742–8. doi: 10.1053/plac.2001.0708
27. Lean IJ, Lucy MC, McNamara JP, Bradford BJ, Block E, Thomson JM, et al. Invited review: recommendations for reporting intervention studies on reproductive performance in dairy cattle: improving design, analysis, and interpretation of research on reproduction. J Dairy Sci. (2015) 99:1–17. doi: 10.3168/jds.2015-9445
28. Sprecher DJ, Hostetler DE, Kaneene JB. A lameness scoring system that uses posture and gait to predict dairy cattle reproductive performance. Theriogenology (1997) 47:1179–87. doi: 10.1016/S0093-691X(97)00098-8
29. Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster G. A Body condition scoring chart for holstein dairy cows. J Dairy Sci. (1989) 72:68–78. doi: 10.3168/jds.S0022-0302(89)79081-0
30. Broes A, LeBlanc SJ. Comparison of commercial progesterone assays for evaluation of luteal status in dairy cows. Can Vet J. (2014) 55:582–4.
31. Joseph IB, Currie WD, Rawlings NC. Effects of time after ovariectomy, season and oestradiol on luteinizing hormone and follicle-stimulating hormone secretion in ovariectomized ewes. J Reprod Fertil. (1992) 94:511–23. doi: 10.1530/jrf.0.0940511
32. Mylonas I, Jeschke U, Kuhn C, Balle A, et al. Immunohistochemical analysis of estrogen receptor alpha, estrogen receptor beta and progesterone receptor in normal human endometrium. Acta Histo. (2004) 106:245–52. doi: 10.1016/j.acthis.2004.02.005
33. Dohoo IR, Martin SW, Stryhn H. Veterinary Epidemiologic Research. 2nd Edn. Charlottetown, PEI: VER, Inc. 2009. p. 865.
34. Maldonado G, Greenland S. Simulation study of confounder-selection strategies. Am J Epidemiol. (1993) 138:923–36. doi: 10.1093/oxfordjournals.aje.a116813
35. Rothman KJ, Greenland S, Lash TL. Modern Epidemiology. 3rd Edn. Philadelphia, PA Lippincott: Williams and Wilkins (2008).
36. Brusveen DJ, Souza AH, Wiltbank MC. Effects of additional prostaglandin F2alpha and estradiol-17beta during Ovsynch in lactating dairy cows. J Dairy Sci. (2009) 92:1412–22. doi: 10.3168/jds.2008-1289
37. Wiltbank MC, Baez GM, Cochrane F, Barletta RV, Trayford CR, Joseph RT. Effect of a second treatment with prostaglandin F2α during the Ovsynch protocol on luteolysis and pregnancy in dairy cows. J Dairy Sci. (2015) 98:8644–54. doi: 10.3168/jds.2015-9353
38. Gümen A, Sartori R, Costa FM, Wiltbank MC. A GnRH/LH surge without subsequent progesterone exposure can induce development of follicular cysts. J Dairy Sci. (2002) 85:43–50. doi: 10.3168/jds.S0022-0302(02)74051-4
39. Carrick MJ, Shelton JN. Oestrogen-progesterone relationships in the induction of oestrus in spayed heifers. J Endocrinol. (1969) 45:99–109. doi: 10.1677/joe.0.0450099
40. Cerri RL, Rutigliano HM, Chebel RC, Santos JE. Period of dominance of the ovulatory follicle influences embryo quality in lactating dairy cows. Reproduction (2009) 137:813–23. doi: 10.1530/REP-08-0242
41. Santos JEP, Galvão KN, Cerri RLA, Chebel R, Juchem SO. Controlled breeding programs for reproductive management. Adv Dairy Technol. (2003) 15:49–68.
42. Denis-Robichaud J. (2016). Assessment of Automated Activity Monitors for Managing Reproduction in Dairy Cows. Ph.D. thesis, Guelph (ON), Canada: University of Guelph. Available online at: https://atrium.lib.uoguelph.ca/xmlui/handle/10214/10123
Keywords: automated activity monitoring, dairy cows, estrogen receptor, estrus expression, progesterone
Citation: Denis-Robichaud J, LeBlanc SJ, Jones-Bitton A, Silper BF and Aoki Cerri RL (2018) Pilot Study to Evaluate the Association Between the Length of the Luteal Phase and Estrous Activity Detected by Automated Activity Monitoring in Dairy Cows. Front. Vet. Sci. 5:210. doi: 10.3389/fvets.2018.00210
Received: 21 June 2018; Accepted: 13 August 2018;
Published: 31 August 2018.
Edited by:
Heriberto Rodriguez-Martinez, Linköping University, SwedenReviewed by:
Martin G. Maquivar, Washington State University, United StatesDivakar Justus Ambrose, University of Alberta, Canada
Copyright © 2018 Denis-Robichaud, LeBlanc, Jones-Bitton, Silper and Aoki Cerri. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ronaldo L. Aoki Cerri, cm9uYWxkby5jZXJyaUB1YmMuY2E=
† Present Address: Jose Denis-Robichaud, Independent Researcher, Vector Consulting, Amqui, QC, Canada