 Michael Kosoy
Michael Kosoy Irina Goodrich
Irina Goodrich- Centers for Disease Control and Prevention, Fort Collins, CO, United States
Phylogenetic sister clades Bartonella and Brucella within the order Rhizobiales present some common biological characteristics as well as evident differences in adaptations to their mammalian reservoirs. We reviewed published data on Bartonella and Brucella infections in wild carnivores to compare the ecology of these bacteria in relatively similar host environments. Arthropod vectors are the main mechanism for Bartonella species transmission between mammalian hosts. The role of arthropods in transmission of Brucella remains disputed, however experimental studies and reported detection of Brucella in arthropods indicate potential vector transmission. More commonly, transmission of Brucella occurs via contact exposure to infected animals or the environment contaminated with their discharges. Of 26 species of carnivores tested for both Bartonella and Brucella, 58% harbored either. Among them were bobcats, African lions, golden jackals, coyotes, wolves, foxes, striped skunks, sea otters, raccoons, and harbor seals. The most common species of Bartonella in wild carnivores was B. henselae, found in 23 species, followed by B. rochalimae in 12, B. clarridgeiae in ten, and B. vinsonii subsp. berkhoffii in seven. Among Brucella species, Br. abortus was reported in over 30 terrestrial carnivore species, followed by Br. canis in seven. Marine carnivores, such as seals and sea lions, can host Br. pinnipedialis. In contrast, there is no evidence of a Bartonella strain specific for marine mammals. Bartonella species are present practically in every sampled species of wild felids, but of 14 Brucella studies of felids, only five reported Brucella and those were limited to detection of antibodies. We found no reports of Bartonella in bears while Brucella was detected in these animals. There is evident host-specificity of Bartonella species in wild carnivores (e.g., B. henselae in felids and B. vinsonii subsp. berkhoffii in canids). A co-adaptation of Brucella with terrestrial wild carnivore hosts is not as straightforward as in domestic animals. Wild carnivores often carry the same pathogens as their domesticated relatives (cats and dogs), but the risk of exposure varies widely because of differences in biology, distribution, and historical interactions.
Introduction
Sixty percent of emerging infectious diseases are zoonoses and majority of these (71.8%) originate from wildlife (1). Among pathogens, Bartonella species might represent an underappreciated danger for human and animal health (2) and human brucellosis remains one of the most common zoonotic diseases worldwide, with more than 500,000 new cases every year (3).
Bartonella and Brucella are phylogenetic sister clades in the order Rhizobiales (4, 5). The genus Brucella is composed of 12 recognized species defined to their preferential hosts (6, 7). The more diverse genus Bartonella includes over 33 validated species exhibiting extremely high genetic diversity (8). Genome analyses of representative species of these bacterial genera have confirmed their shared ancestry. Alsmark et al. (9) identified 760 Bartonella henselae genes, for which homologs are present in one of chromosomes of Brucella suis. In addition to their genetic proximity, the Bartonella and Brucella genera present analogies in their life history and ecology that are even more important for our analysis. Whereas, most closely related species of the order Rhizobiales are symbiotic on plant roots, both Bartonella and Brucella are adapted to diverse mammalian hosts. Each Bartonella and Brucella species has one or a few closely related mammal reservoir hosts (5).
Investigations of wild animals, including predators, for Brucella infections started much earlier and were more intensive compared to studies of Bartonella infections. Research of Brucella infections in animals has been dominated by studies of domestic animals and, to a lesser degree, of wild ruminants. Although Brucella canis was identified in domestic dogs more than 50 years ago and is well known to veterinary community as a causative agent of canine abortion (10), investigations of Brucella in dogs are much fewer than those of Brucella in cattle, sheep, goats, and pigs. This is mainly due to lack of good tests for rough Brucella species, not because of lack of interest. Publications on the distribution of Brucella species among wild canids, as well as among other wild carnivores, are even more limited in the western literature. At the same time, a good number of reports on this topic are scattered across Russian literature. Most of these were published during Soviet times, sometimes in classified proceedings, and are not easily available (11–13). Identification of novel species and genotypes of Brucella in rodents, bats, marine mammals, and amphibians stimulated epidemiological research of diverse animal species, including wildlife (7).
Extensive investigations of animals for Bartonella species started in the early 1990s after the discovery that one or more Bartonella species could cause cat scratch disease in people. For this reason, most studies targeted domestic cats and dogs with limited investigations of stray dogs and feral cats (14, 15). Studies on detection, identification, and characterization of Bartonella species in wild animals usually targeted small mammals: rodents (8) and bats (16). Chomel et al. (17) pioneered Bartonella research in wild carnivores and ruminants. Since then, numerous wildlife studies have been conducted in various parts of the world. However, a comprehensive analysis of the available data on prevalence and diversity of Bartonella and Brucella infections in wild carnivores has yet to be published. Such an analysis would allow comparisons to be made between the ecologies of these two bacterial groups living in similar host environments and identify possible directions for future research.
In this review we undertook such an analysis through an extensive literature review. We examined the similarities and differences in the Bartonella and Brucella ecology and, more importantly, analyzed biological features that may reveal ways of these phylogenetically close bacterial genera exhibit evolutionary adaptations to the same or related mammalian hosts, presumably during the long periods over which they have co-occurred. Considering the differences in the genera's life history, we paid special attention to possible arthropod-mediated transmission of these bacteria between mammalian hosts.
For this review, we followed the more accepted taxonomic division of the order Carnivora into suborders Feliformia (“cat-like”) and Caniformia (“dog-like”), with pinnipeds included as a separate superfamily level clade (Pinnipedia). We chose these divisions not for preference for a specific taxonomic scheme, but as a convenient basis for analysis of available data on Bartonella and Brucella infections in 12 families: suborder Feliformia (Felidae, Herpestidae, Hyaenidae, and Viverridae), suborder Caniformia (Canidae, Mephitidae, Mustelidae, Procyonidae, and Ursidae), and clade Pinnipedia (Odobenidae, Otariidae, and Phocidae).
We conducted a thorough literature search by using PubMed, Scopus, OVID Medline, BioOne, Crossref, WorldCat, Web of Science, Google Scholar, and other databases. In the search we used keywords: “Bartonella ecology,” “Brucella ecology,” “Bartonella AND wild animals,” “Brucella AND wild animals,” “Bartonella AND carnivores,” “Brucella AND carnivores,” “Bartonella AND predators,” “Brucella AND predators,” “Bartonella AND marine mammals,” “Brucella AND marine mammals,” “Bacteria AND wild carnivores,” “Bartonella AND fleas AND mammals,” “Brucella AND arthropods,” and their variations. We realized that all these search engines had missed numerous reports on detection of Brucella in wild animals in the Russian language literature and we conducted our own search of such sources in the Russian Internet and search engines as well as by working through the references in related articles and reviews in the Russian language.
We used the word “wild” in the meaning of “free-ranging” and apart from a few publications of particular interest, we excluded reports of Bartonella and Brucella in captive and zoo animals as the composition of bacterial communities in such animals could have been modified by separation from the natural environment or via acquisition of bacterial infections from the surrounding environment (e.g., from urban rats). The literature on Bartonella and Brucella infections in domestic carnivores (cats and dogs) is abundant, so we limited inclusion for comparative purposes only.
We collected data from serological, bacteriological, and molecular investigations of Bartonella and Brucella infections in all families of wild terrestrial and marine carnivores worldwide. Providing data from various techniques, we need to acknowledge that discrimination power of characterization of pure cultures and sequence analyses for identification of Bartonella and Brucella species is greater than that of serological procedures. However, serological methods remain an important tool in detection and identification of these infections in animals and should be taken in consideration with full awareness of their limitations.
Then we collated the obtained information in Bartonella, Brucella and combined tables by carnivore species divided into their respective families listed in alphabetical order of their Latin names. We listed information on location where the samples were collected, investigation method, prevalence and bacterial species, and reference to the study. Both positive and negative results of investigations were included. The combined table shows only references listed by carnivore species in their respective families.
Features of Bartonella and Brucella Bacteria Related to Their Ecology in Wild Animals
Biological Characteristics
Bartonella and Brucella bacteria share some biological characteristics, yet there are evident differences in their adaptations to their animal reservoirs. Infections caused by bacteria of both taxonomic groups can lead to a long-lasting bacteremia with ability to invade specific mammalian cells and survive inside them. Via analogous mechanisms, the specialized secretion system (T4SS) works as a molecular syringe to inject effector molecules into their target cells (18, 19). Bartonella and Brucella modulate their gene expression to adapt to the different environments during the infectious process (20, 21). The VirB systems of Bartonella and Brucella are associated with distinct groups of effector proteins that collectively mediate interactions with host cells (19).
Bartonella bacteria infect endothelial cells and seed into the bloodstream, colonizing erythrocytes which provide a persistence niche for the bacteria. The ability of these bacteria to exploit their reservoir hosts with diminished morbidity and to cause a high level of bacteremia justifies the definition of “elegant hemotrophic parasites” given by Birtles (22) to bartonellae. In incidental hosts, Bartonella infections can cause various clinical manifestations commonly without high-level bacteremias (21). In contrast to Bartonella, Brucella bacteria invade and multiply within mammalian host's macrophages and placental trophoblasts (18, 19, 23). Although bacteremia is common during brucellosis, data on duration and mechanisms of Brucella persistence in animal blood are limited.
Transmission of Bartonella and Brucella Bacteria
The persistence of Bartonella bacteria in red blood cells optimizes transmission of these bacteria by blood-sucking arthropods. High prevalence and long-term bacteremia in reservoir mammals and adaptation to specific vectors seem to be the common strategy of bartonellae for transmission and host diversity (24). Many described Bartonella species are vector-borne bacteria transmitted by fleas, sand flies, lice, and biting flies depending on the bacteria species involved and their vertebrate reservoirs (24, 25). Experimental studies demonstrated louse and flea transmission of B. henselae and B. quintana (26–28). Some investigators provided evidence of potential role of ticks and mites in transmission of Bartonella species, but debates continue on their role of as vectors (25, 29). A 2008 study by Cotté et al. (30) showed that Ixodes spp. ticks are capable of transmitting B. henselae via salivary contents, but Telford and Wormser (31) found no convincing evidence that ticks were vectors of Bartonella species. Molecular detection of Bartonella spp. in terrestrial leeches (Haemadipsa rjukjuana) by Kang et al. (32) opens up a discussion of the pathogen transmission by land leeches.
Transcutaneous transmission of Bartonella via animal bites and scratches during hunting, as well as through butchering or handling wild meat is another possibility (33). Cat scratch disease, caused by B. henselae is the best-documented example of direct animal-to-human transmission of a Bartonella species by scratch or bite inoculation (14). Finkelstein et al. (27) showed that B. henselae can remain viable in flea feces for over 72 hours. Therefore, transmission potentially can occur via inoculation of B. henselae from infected flea feces into the skin via open wounds. Suspected Bartonella alsatica transmission from wild rabbits to humans, presumably occurring during hunting and butchering, was reported in patients with endocarditis or lymphadenitis in France (34, 35). Suspected dog bite transmission of B. vinsonii to a human was reported based upon serological evidence (36). There is little information on possible vertical transmission of bartonellae in animals. However, Bartonella species were isolated from the embryos and neonates of naturally infected cotton rats (Sigmodon hispidus) and white-footed mice (Peromyscus leucopus) (37). Experimental inoculation of B. henselae to adult female cats was accompanied by decreased conception or failure to maintain pregnancy (38). Considering the extensive animal reservoirs and the large number of insects that have been implicated in the transmission of Bartonella species, animal exposure to these organisms may be more substantial than is currently believed.
Transmission of Brucella occurs mainly via close contact with placenta, aborted fetuses, fetal fluids, reproductive tract discharges, and secretions (7). Infected dogs intermittently shed low concentrations of bacteria in seminal fluids and nonestrus vaginal secretions. Postabortion vaginal fluids contain a high level of bacteria and are a source of infection for other dogs (39). In addition, dogs can shed the bacteria in the saliva, nasal secretions, and urine (40). Studies suggest that the concentration of Br. canis in urine is higher in male than female dogs; this difference is attributed to urine contamination with seminal fluid (41). However, the role of urine as a source of infection is not fully understood (39).
There is a widely accepted perception that absence of transmission of Brucella species via arthropod vectors is the most essential difference in ecology of these bacteria compared to the Bartonella species. We found a number of rarely cited publications on either detection of Brucella species in arthropods or experimental studies designed to verify a possibility of vector transmission of these bacteria. Although detection of Brucella in arthropods collected from different sources does not often directly relate to carnivores, such information can help interpret potential mechanisms of bacterial transmission. The invasion of Brucella into erythrocytes and its persistence in blood suggest a possibility for transmission by bloodsucking arthropods in nature (42). Although Brucella may be found in erythrocytes, these bacteria exhibit strong tissue tropism and replicate within vacuoles in macrophages, dendritic cells, and placental trophoblasts. Evidence that Brucella species can be spread among animals by arthropods is very limited. Some Russian authors argued that parasitic arthropods, especially ticks, could preserve Brucella in nature and transmit them within a population from one animal to another (43, 44). Rementsova (43) listed 20 observations of Brucella detection in ticks. Experiments in Russia reported that both ixodid and argasid ticks were infected with Brucella at different phases of their development and could transmit the pathogen to uninfected animals during bloodsucking (43). Brucella in ticks retained their virulence even after 2 years (43). More recently, Neglia et al. (45) detected Br. abortus DNA and RNA in different stages of development of the sucking louse (Haematopinus tuberculatus).
Alimentary transmission is important for Brucella as proved by experimental studies in wild carnivores. Scanlan et al. (46) infected gray foxes with Br. abortus in dog food. Seven of eight foxes became seropositive. Neiland and Miller (47) infected six beagle dogs, two wolves (Canis lupus), one black bear (Ursus americanus), and two grizzly bears (Ursus arctos horribilis) with a strain of Br. suis biovar 4 isolated from a sled dog from Alaska. Their experiments demonstrated that canids and ursids are susceptible to the infection via intraperitoneal inoculation and through oral mucous membranes. During acute stages of the infection, Brucella congregated in these species in high numbers in lymph nodes and distributed throughout the body. Importantly, Brucella invaded salivary glands and probably also mammary glands and kidney, thus providing conditions for shedding the bacteria in saliva, milk, and urine. The authors reported reproductive failure during infection in wolves, but were not confident that the failure was a consequence of the infection (47). Morton (48) experimentally infected foxes with Br. suis biovar 4 and observed that the incidence of positive titers, positive cultures, and shedding of bacteria was related to the number of Brucella organisms experimentally fed to the animals. Lowest doses did not produce infection. Highest doses produced positive titers and cultures.
Tests on rats showed transmission of Br. abortus biovar 1 from infected male to uninfected female rats resulted from sexual intercourse (49). Vertical transmission of Br. abortus caused sterility in pregnant mice (50); Wang et al. (51) documented vertical transmission of Br. melitensis on a pregnant mouse model. Guzman-Verri et al. (52) cited the more likely modes of transmission of Br. ceti to be through sexual intercourse, maternal feeding, aborted fetuses, placental tissues, vertical transmission from mother to the fetus or through fish or helminth reservoirs.
Brucellae have high viability and can survive in the environment for 3–21 days in spring-summer and for 151–233 days in winter-fall seasons. Brucellae maintain viability in carcasses (muscles, internal organs, and lymph nodes) at −7.2° to 38.4°C for 1–12 months (53). Long-term survival of Br. microti in soil was described and, thus, soil might act as a reservoir of infection (54).
Prevalence of Bartonella Infections in Wild Carnivores
General Prevalence Pattern
Overall, prevalence of Bartonella infections in carnivores was higher compared to Brucella infections. In the studies of over two Feliformia animals, the highest prevalence was registered by culture in bobcats [37%, 7/19, (55)], by IFA in bobcats again [74%, (56)], and by PCR of blood in Iberian lynx [33.3%, 10/30, (57)]. In studies of over two Caniformia animals, the highest Bartonella prevalence was registered by culture in gray foxes [49%, 26/53, (58)], by IFA in coyotes [89%, 48/53, (58)], and by PCR of blood in raccoons [43%, 16/37, (59)]. A very high overall prevalence of antibodies to B. henselae (95%) was detected among Brazilian free-ranging felids (60) (Table 1). As expected, the number of seropositive animals was usually higher than the numbers of culture or PCR positive individuals from the same study. Thus, of the 54 lions from South Africa, 5.2% were positive by culture, 3.7% were PCR positive for Bartonella DNA, and 17% had Bartonella antibodies (62). A study on golden jackals in Iraq found 14.5% of animals positive by PCR and 40.4% (23/57) by IFA (86).
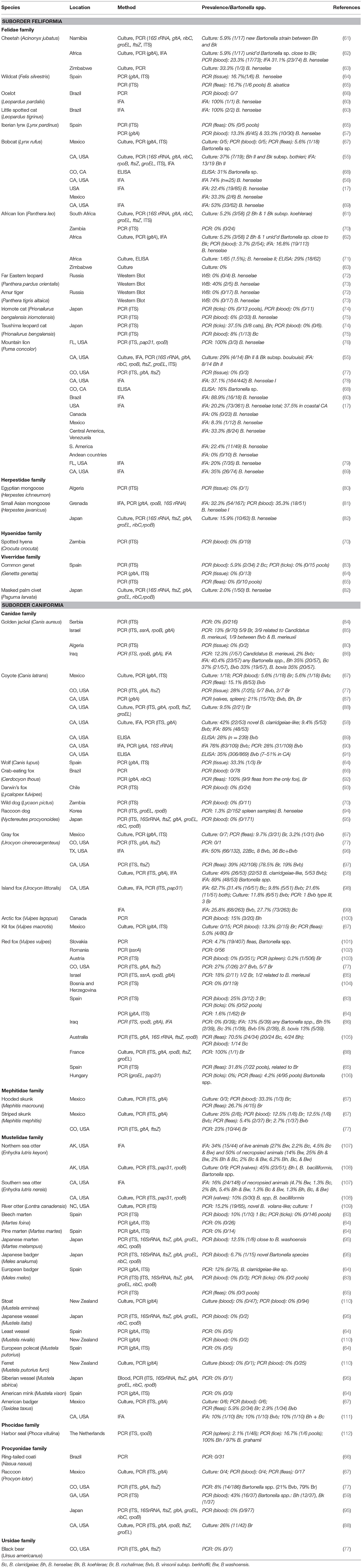
Table 1. Bartonella studies in wild carnivores by species.
Age and Gender Pattern
Prior studies usually show no statistical difference in prevalence by age or gender in felines (61, 69, 79). However, Chomel et al. (17) found antibody prevalence for B. henselae to increase with age in pumas in California. In contrast, Rotstein et al. (79) found antibody prevalence higher in Florida panthers under 2 years of age (40%) compared to panthers over 2 years of age (13%).
Geographic Pattern
Prevalence of B. henselae antibodies in mountain lions and bobcats varied significantly between different states of the U.S. (17). Mountain lions from Arizona, California, and Texas were more likely to be seropositive for B. henselae (26.7–40.0%) than pumas from the Northwest and Mountain states (0–11.8%) (17). In California, the highest prevalence in bobcats was from the coastal range (37.5%), while the highest prevalence in pumas was from Southern California and Sierra Nevada (17). The reported pattern was similar to the geographic distribution of Bartonella infection in domestic cats. It has been demonstrated that in cat populations (stray or pets), prevalence of infection was demonstrated to vary considerably with an increasing gradient from cold climates (0% in Norway) to warm and humid climates (68% in the Philippines) (14). In the U.S., prevalence of B. henselae antibodies in pet cats varied significantly with the highest average prevalence in the southeastern United States, Hawaii, coastal California, the Pacific Northwest, and low prevalence in Alaska, the Rocky Mountain-Great Plains region, and the Midwest (113). Comparing wild felids at four sites in California and Colorado, Bevins et al. (68) noted that seroprevalence varied considerably, but in almost all cases, it was higher in warmer and more humid California than in Colorado. For mountain lions, suburban land use predicted increased exposure to Bartonella species in southern California (114).
Seasonal Pattern
Studies have yielded conflicting evidence about the seasonality of B. vinsonii subsp. berkhoffii infection in coyotes. First, Chang et al. (91) reported that the prevalence of Bartonella antibodies was highest in summer (42%) and lowest in spring (29%), whereas a geographically more restricted study conducted in coastal central California, U.S., by the same authors found the highest seroprevalence in winter (100%) and the lowest in summer (62%) (90). Investigating antibody prevalence in 239 coyotes from northern California, Beldomenico et al. (89) identified some environmental factors associated with the seropositivity. In that study, prevalence of antibodies against B. vinsinii subsp. berkhoffii was 44% in the summer, 40% in the spring, 27% in the winter, and 19% in the fall. The authors noticed that Bartonella seropositivity was associated with higher precipitation and proximity to the coast. In addition, coyotes seropositive for B. vinsonii subsp. berkhoffii were more likely to be seropositive for tick-borne agents Anaplasma phagocytophilum and mosquito-vectored Dirofilaria immitis (89). Interestingly, California Zoo felids of the genus Felis were found almost three times more likely to be seropositive for B. henselae than animals belonging to the genera Panthera and Acinonyx (69).
Prevalence of Brucella Infections in Wild Carnivores
General Pattern
Eighty-nine percent of Brucella studies of wild carnivores were conducted by serological and bacteriological methods, but no reports were found on culturing Brucella from representatives of suborder Feliformia. Only a few wild felid species (lion, jaguar, and bobcat), mongooses, and spotted hyena were serologically positive. Apart from one bobcat that had antibodies against Br. canis (115), the rest of seropositive Feliformia animals had antibodies against Br. abortus. We have to be cautious with the claim about presence of specific antibodies in this paper, as well in many other reports, because Br. abortus suspensions can also detect Br. melitensis. The highest seroprevalence was registered in white-tailed mongoose [33.3%, 1/3, (116)]. In evident contrast to Feliformia animals, prevalence of Brucella in various Caniformia species varied greatly, with many reporting high prevalences of positive antibody titeres. Antibodies to Brucella species were recorded in 40% of coyotes (117), 42% of wolves (118), 43% of black-backed jackals (116), 50% of Arctic foxes and 40% of red foxes (48), 64% of grizzly bears (119), 28% of Asian sea otters (120), 23% of California sea lions (121), and 74% of Australian seals (122). Brucella was cultured from 30.8% of wolves (123) (Table 2).
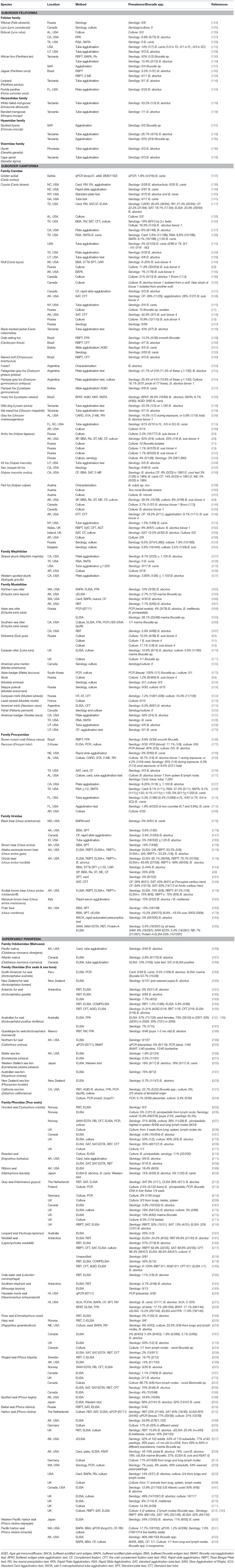
Table 2. Brucella studies in wild Carnivores by species.
Age Pattern
We could find information on age dependence only in marine Brucella. In the 2018 study on gray and harbor seals, Kroese et al. (212) noted remarkable age-dependent prevalence of Br. pinnipedialis in both serology and in the investigation of the tissues from stranded animals. The PCR positivity was 84% (26/31) in juveniles compared to 57% (4/7) in adults and Br. pinnipedialis was cultured only from juveniles and not from adults in that study. Similar age dependence was shown in harbor seals by Miller et al. (169) and Ewalt et al. (229). Nymo et al. (206) noted the age-dependent prevalence of anti-Brucella antibodies in hooded seals. Pups (<1 mo old) had a substantially lower probability of being seropositive (4/159, 2.5%) than yearlings (6/17, 35.3%), suggesting that exposure may occur post-weaning, during the first year of life. For seals over 1 year old, the mean probability of being seropositive decreased with age, with no seropositives older than 5 years, indicating loss of antibody titer with either chronicity or clearance of infection (206).
Bartonella Species Identified in Wild Carnivores
Bartonella Species in Wild Feliformia Animals
Wild Feliformia animals mostly carry the same Bartonella species as domestic cats, namely B. henselae (types I and II), B. koehlerae, and B. clarridgeiae (234). The same species were detected in feral cats from Georgia, U.S. (59). In Africa, free-ranging lions were found infected with B. henselae type II and B. koehlerae subsp. koehlerae and Namibian cheetah with a strain that clustered between B. henselae and B. koehlerae and was considered a new subspecies of B. koehlerae (61, 63). In Japan, B. henselae was found in Iriomote leopard cats and B. clarridgeiae DNA was detected in Tsushima leopard cats (74, 82).
In a study on free-ranging mountain lions and bobcats from California, U.S., Chomel et al. (55) described new Bartonella strains, which were similar to but different from B. henselae and B. koehlerae, and named them B. koehlerae subsp. boulouisii and B. koehlerae subsp. bothieri. Phylogenetic analysis based on comparison of four genetic markers revealed two clusters: one with five strains obtained from bobcats and another with three strains obtained from mountain lions indicating a degree of host-speciation of these strains (55). In Brazil, sequencing analysis revealed a Bartonella strain close to but different from B. henselae and B. koehlerae in wild-born captive margay (Leopardus wiedii) (235).
Other Bartonella species were detected in fleas collected from wild felids. For example, Bartonella alsatica was found in one of six rabbit fleas Spilopsyllus cuniculi collected from a European wildcat (F. silvestris) in Spain (65). This Bartonella species is usually associated with rabbits and possibly fleas were infected or they contained blood meal from infected rabbits, as S. cuniculi is normally found on European rabbits (Oryctolagus cuniculus). A different situation has been reported by López-Pérez et al. (67) regarding a genetic variant obtained from a flea (Pulex simulans) collected from a bobcat (L. rufus) in northwestern Mexico. This variant had ITS sequence 99.1% similar to a strain previously isolated from another bobcat from California, U.S., but distant from all other Bartonella genotypes.
Bartonella Species in Wild Caniformia Animals
In the studies, Caniformia animals were found to carry B. henselae, B. clarridgeiae, B. vinsonii subsp. berkhoffii, B.rochalimae, B. washoensis, and B. bacilliformis. In an investigation of wild carnivores from Colorado, U.S., Bai et al. (77) identified two Bartonella species, B. vinsonii subsp. berkhoffii and B. rochalimae. Striped skunks exclusively carried B. rochalimae, while coyotes, red foxes, and raccoons were infected with either or both Bartonella species. Bartonella rochalimae DNA was found in a wolf (C. lupus) in northern Spain (64). Investigating wild canids along with stray dogs throughout Iraq, Chomel et al. (86) identified a novel strain of Bartonella, which was named Candidatus B. merieuxii, in six jackals (Canis aureus). By three genetic markers, the “jackal” strain was aligned most closely with B. bovis and the other ruminant Bartonella species. Sequences closely related to Candidatus Bartonella merieuxii later were found in three jackals and one red fox (V. vulpes) in Israel (85). Besides this strain, B. rochalimae and B. rochalimae-like were found in five jackals and one fox, and one jackal harbored B. vinsonii subsp. berkhoffii (85).
Kehoe et al. (87) documented the presence of three Bartonella species in heart valves and/or spleen of free-ranging coyotes from northern California, U.S. Partial DNA sequencing showed that aortic valves from 8 (53%) of 15 coyotes were B. vinsonii subsp. berkhoffii positive, B. rochalimae DNA was amplified from the spleen of one coyote, and B. henselae DNA was amplified from the mitral valve of another coyote. By sequence analyses, four coyotes were infected with B. vinsonii subsp. berkhoffii genotype I, three with genotype II, and one with genotype III (87).
Two species of Bartonella, a novel Bartonella clarridgeiae-like bacterium and B. vinsonii subsp. berkhoffii, were isolated from rural dogs and gray foxes in northern California (58). Two B. henselae sequences detected in the spleen of raccoon dogs in Korea matched the strain Houston-1 and by ITS sequences were 99.8% similar to a strain found in dogs in China (94). Northern and Southern sea otters were found IFA positive for B. washoensis (107, 108). The authors also detected B. bacilliformis by PCR in heart valves of both species. A strain close to B. washoensis was detected by PCR in Japanese marten (95). Chinnadurai et al. (109) detected a novel strain in river otters by PCR with a sequence matched a strain previously described in Southern flying squirrel. In the Netherlands, harbor seals were found to carry a strain 97% similar to B. grahamii (112).
Brucella Species Identified in Wild Carnivores
There are no reports on identification of Brucella species by culture or by sequence analysis in animals belonging to Felidae, Herpestidae, and Hyaenidae families. Except for one bobcat that had antibodies against Br. canis (115), the other few seropositive Feliformia animals had antibodies against Br. abortus (116, 132). Since the authors did not use specific tests that identify rough Brucella species, they were not able to find antibodies.
In contrast, multiple Brucella species can infect Caniformia animals. Brucella species identified by culture or PCR/sequencing in terrestrial carnivores included Br. canis in coyotes (137); Br. abortus in wolves, red foxes, gray foxes, pampas gray foxes, and raccoons (123, 126, 154, 177); Br. suis biovar 4 in wolves, arctic foxes, and red foxes (11, 157, 164); Br. microti and Br. vulpis in red foxes (159, 161). One red fox species, V. vulpes, can carry four different Brucella species (Br. abortus, Br. vulpis, Br. microti, and Br. canis) (115, 125, 159–161). All isolates obtained from arctic foxes were identified as Br. suis biovar 4 (11, 13, 48). This is not surprising as reindeer are common hosts of Br. suis biovar 4, and arctic foxes often scavenge dead reindeer.
Various aquatic carnivores carry a different species, Brucella pinnipedialis. It was identified in the harbor seal (Phoca vitulina), the ringed seal (P. hispida), the harp seal (Pagophilus groenlandicus), the gray seal (Halichoerus grypus), the hooded seal (Cystophora cristata), Asian sea otter (Enhydra lutris), and European river otter (Lutra lutra) (168, 171, 207, 208, 214). Characterization of the isolates belonging to this species indicated that Br. pinipedialis may contain different biovars (208).
Bartonella and Brucella Infections in Wild Carnivores by Family
Family Felidae
Genus Panthera
Bartonella infection was reported in three big cats species: African lion (P. leo), jaguar (P. onca), and Far Eastern leopard (P. pardus orientalis). B. henselae and B. koehlerae subsp. koehlerae were cultured from the blood of three (5.2%) of 58 lions from Kruger National Park in South Africa (61, 62). The level of bacteremia in the culture-positive lions varied from 35 to 2,000 bacteria per 1 ml of blood. Bartonella culture- and antibody-positive lions were found among semi-captive lions from three ranches in South Africa (71). Interestingly, all studied lions from Zambia and Zimbabwe were negative for Bartonella by culture and PCR (63, 70). A wild-caught jaguar in Brazil, which was maintained in captivity for only a week, was found B. henselae positive (236). This finding led the authors to believe that the animal had been infected in the wild. In the Russian Far East, wild Amur tigers (P. tigris altaica) tested negative for antibodies to B. henselae (72, 73), but two of five Far Eastern leopards from that area had antibodies against B. henselae (73).
Limited information exists about Brucella in the wild cats of the genus Panthera. During the investigation of Brucella infection in the human, livestock and wildlife interface in the Katavi-Rukwa ecosystem in Tanzania, Assenga et al. (130) found one of the two tested lions serologically positive for Brucella at a titer 1:200 by three different tests (RBPT, BAPA, and Riv.T). In a 1968 study in Tanzania, Sachs et al. (116) found two of 13 lions had antibodies to Brucella species by tube agglutination test. De Vos and Van Niekerk (131) were not able to detect Brucella antibodies in four lions from the Kruger National Park, South Africa. Furtado et al. (132) tested serum samples from 31 free-ranging jaguars (P. onca) from Brazil using Br. abortus as antigen and reported antibodies in one jaguar.
Genus Puma
Two Bartonella species were cultured from mountain lions (P. concolor) (55). Bartonella antibodies were found in mountain lions from Arizona, California, Idaho, Oregon, Texas, Wyoming, and Florida in the U.S. (17, 68, 69, 78, 79). No Bartonella DNA was detected in spleen samples of three mountain lions from Colorado, U.S. (77). B. henselae antibodies were found in pumas from Bolivia, Peru, and Venezuela (17). Filoni et al. (60) reported 16 out of 18 pumas serologically positive to B. henselae in Brazil. B. henselae DNA was detected in lung tissues of three Florida pumas with the first and only up to date reported association of B. henselae infection with a fatal disease syndrome of necrotizing interstitial pneumonia and suppurative myocarditis in pumas (76).
All 24 free-ranging Florida panthers (P. c. coryi) were seronegative for Brucella (134). Reports of Brucella in populations of pumas from elsewhere in the Americas were unavailable.
Genus Acinonyx
The cheetah (A. jubatus) is only member of its genus. Kelly et al. (63) reported isolation of B. henselae genotype II from an African pet cheetah from Zimbabwe. In 2016, Molia and colleagues (61) isolated Bartonella bacteria from blood of 5.9% (1/17) Namibian cheetahs, and the cheetah was infected with a previously unidentified Bartonella strain. The Namibian cheetah strain was close but distinct from isolates from North American wild felids and clustered between B. henselae and B. koehlerae; it was claimed to be a new subspecies of B. koehlerae (61). The same study documented that 23% of the 73 animals were positive for Bartonella DNA by PCR and 31% (23/74) of cheetahs had antibodies to B. henselae. No reports on Brucella infections in the cheetah were found.
Genus Lynx
Those are medium-sized cats represented by four species: Canadian lynx (Lynx canadensis), Eurasian lynx (L. lynx), Iberian lynx (L. pardinus), and bobcat (L. rufus). Chomel et al. (55) isolated two Bartonella species (B. henselae and B. koehlerae subsp. bothieri) from bobcats in California, U.S. A high prevalence of Bartonella antibodies (22.4–74.0%) was reported in bobcats from California, Colorado, Florida, Nevada, and Oregon in the U.S. and from Mexico (17, 56, 68, 69). In northwestern Mexico, a Bartonella genotype was found in a flea P. simulans collected from a bobcat, but not in the blood of that animal (67). B. henselae DNA was found in 16 of 75 (21.3%) blood samples of Iberian lynx from southern Spain (57).
Antibodies against Brucella species in bobcats were reported in two studies: Br. abortus at 6.6% (5/75) in California (127) and Br. canis at 33% (1/3) in Texas (115). Serological investigations of bobcats from Alabama, Texas, and Utah in the U.S. did not result in identification of antibodies to Brucella (126, 128, 129). Tessaro (125) reported the absence of Brucella bacteria and anibodies in Canadian lynx.
Genus Leopardus
These are small spotted cats mostly native to Middle and South America. Representatives are the ocelot (L. pardalis), the little spotted cat (L. tigrinus), Geoffroy's cat (L. geoffroyi), and the margay (L. wiedii). Antibodies to B. henselae were reported in the ocelot (1/1) and the little spotted cat (2/2) in Brazil (60). A Bartonella sequence similar to B. koehlerae and B. henselae was detected in the captive margay in Brazil (235). The animal was born in the wild and lived in captivity prior to sampling, thus it is not possible to ascertain if the infection was acquired in the wild or in captivity. The authors claimed this animal exhibited clinical signs of bartonellosis: episodes of accentuated weight loss, dullness, dehydration, and anemia (235). The main reason why we have included the case of captive margay into our review is that the identified strain was different from all strains described in domestic and wild felines.
Genus Prionailurus
This is a genus of small spotted wild cats native to Asia. The genus includes the Iriomote cat (P. iriomotensis) and the Tsushima leopard cat (P. bengalensis euptilura), both endangered in Japan. A molecular epidemiologic survey in Japan resulted in identification of B. henselae in 6% (2/33) of Iriomote leopard cats and B. clarridgeiae in 8% (1/13) of Tsushima leopard cats (75). In the following study, four ixodid ticks collected from Tsushima leopard cats were PCR positive for B. henselae (74).
Genus Felis
The European wildcat (F. silvestris silvestris) is a subspecies of the same species that includes domestic cats (F. s. catus). This species is found in forest habitats of Europe. There are two reports of the presence of Bartonella in wildcats from Spain. First, Márquez et al. (65) identified B. alsatica, strain associated with rabbits, in a flea Spilopsyllus cuniculi collected from a wildcat in Spain. Then, Gerrikagoitia et al. (64) detected B. henselae DNA in a carcass of a wildcat. A study of feral cats in the U.S. state of Georgia by Hwang and Gottdenker (59) also reported that 48% of feral cats were PCR positive for three Bartonella species: B. henselae, B. koehlerae, and B. clarridgeiae.
Family Viverridae
The most common species are civets and genets widely distributed in South and Southeast Asia, Africa, and Southern Europe. The first evidence suggesting that civets can host Bartonella came from a description of a human cat scratch disease case reported in 2001 in Japan. In the case, a patient scratched by a masked palm civet (Paguma larvata) developed fever and inguinal lymphadenopathy with a high antibody titer (1:1,024) to B. henselae (237). Later, Sato et al. (82) cultured B. henselae from blood of one of 50 masked palm civets collected in Chiba Prefecture of Japan. The level of bacteremia was high (7,000 bacteria per 1 mL of blood). Importantly, the multi-locus sequence type detected from the isolated strain revealed a unique genotype. Though the prevalence of Bartonella in cats in Chiba prefecture was 5%, the same genotype had never been found in any B. henselae strains from cats from the same and other prefectures (82). Bartonella DNA was detected in another Viverridae species, the common genet (Genetta genetta). Conducting molecular detection of vector-borne pathogens in wild carnivores in natural parks and adjacent residential areas in Barcelona, Spain, Millán et al. (83) identified B. clarridgeiae in tissues of two of 34 (6%) common genets, but ticks collected from genets were free of Bartonella DNA. In another study conducted in Northern Spain (Basque County), Gerrikagoitia et al. (64) did not detect Bartonella DNA in 13 common genets tested. Márquez et al. (65) also found no Bartonella DNA in 18 fleas S. cuniculi collected from 10 common genets in Andalusia, Spain.
Reports of Brucella testing among viverrids are nearly nonexistent. No Brucella antibodies were found in two common genets (G. genetta) and three Cape genets (G. tigrina) from eastern Africa tested by tube agglutination test (116, 136).
Family Herpestidae
Mongooses is the common name for the weasel-like small carnivores that live in southern Asia, Africa, and southern Europe, and are introduced to some other areas. We have information about Bartonella in one species of this genus—the small Asian mongoose (Herpestes javanicus). Sato et al. (82) isolated B. henselae from 15.9% (10/63) of small Asian mongooses from Okinawa prefecture, Japan. Based on multi-locus sequence analysis, they identified four types of B. henselae strains cultured from mongooses (82). Jaffe et al. (81) tested small Asian mongooses in Grenada and found 32% (54/167) of the animals IFA positive and 35% (18/51) PCR positive for B. henselae. The only additional report of investigation of mongooses was from testing a single Egyptian mongoose (Herpestes ichneumon) in Algeria and the PCR test was negative (80).
There is a report of antibodies against Br. abortus in one white-tailed mongoose (Ichneumia albicauda) (33%, 1/3) and one banded mongoose (Mungos mungo) (100%, 1/1) in Tanzania (116).
Family Hyaenidae
The family contains four species of hyenas and phylogenetically belongs to the suborder Feliformia despite the dog-like appearance of these animals. The only available report on testing hyenas for Bartonella is from a molecular survey of 19 spotted hyenas (Crocuta crocuta) from two sites in Zambia with no positive results (70).
Serological observation of 15 spotted hyenas from Tanzania resulted in detection of antibodies against Brucella in four out of 15 (27%) animals (116). In the prior study, Sachs and Staak (135) found Brucella species exposure in two out of four hyenas in Tanzania. Another serological study did not detect Brucella antibodies in two spotted hyenas from the Kruger National Park in South Africa (131).
Family Canidae
Genus Canis
Multiple wild species, including coyotes, jackals, and wolves belong to this genus. The golden jackal (C. aureus) is a species experiencing rapid geographic expansion with significant public health impacts (238). Of 57 golden jackals sampled from four sites in Iraq, seven (12.3%) were PCR positive for Candidatus B. merieuxii and one (2%) for B. vinsonii subsp. berkhoffii (86). In Israel, Marciano et al. (85) found nine out of 70 (13%) golden jackals PCR positive for Bartonella species: 5/9 B. rochalimae, 3/9 close to Candidatus B. merieuxii, and 1/9 between B. vinsonii subsp. berkhoffii and Candidatus B. merieuxii. A search for Bartonella in coyotes (C. latrans) from California and Colorado in the U.S. and from Mexico demonstrated a high prevalence of up to 89% by IFA, 42% by culture, and 28% by PCR (58, 67, 77, 87–91). There is one report of PCR detection of Bartonella DNA in a wolf (C. lupus) from northern Spain (64).
Most reports of Brucella infections in canids are based on detection of antibodies. Serologically positive coyotes were identified from California and Texas, U.S. (115, 117, 127, 128, 142). In wolves, evidence of Brucella infections also included Brucella isolations in Canada and Russia (13, 125). Brucella was found in two jackal species: 1.9% (4/216) of golden jackals (C. aureus) in Serbia were positive for Br. canis by PCR (137) and 43% (3/7) of black-backed jackals (C. mesomelas) in Tanzania were seropositive for Br. abortus by tube agglutination test (116).
Genus Vulpes
There are more reports on detection of Bartonella in red foxes (V. vulpes) than in any other species of wild carnivores. Bartonella DNA was identified in red fox tissues from Australia, Austria, France, Israel, Spain, and U.S. (64, 77, 83, 85, 88, 103, 105). Most sequences were identified as B. vinsonii subsp. berkhoffii and B. rochalimae. Out of two red foxes positive for Bartonella DNA in Israel, one harbored DNA sequences that were 100% identical to B. rochalimae and the other was positive for Candidatus B. merieuxii (85). Hodžić et al. (104) did not detect Bartonella DNA in 119 fox spleen samples from Bosnia and Herzegovina. Blood samples from 39 red foxes from Iraq were also negative for Bartonella by PCR; however, 12.8% of these foxes were serologically positive for Bartonella antibodies (86). Mascarelli et al. (100) detected B. henselae DNA in three out of 20 tested arctic foxes (V. lagopus) from Canada and López-Pérez et al. (67) identified B. rochalimae DNA in two out of 15 kit foxes (V. macrotis) tested.
There are several reports about screening of ectoparasites from red foxes. DNA of a Bartonella strain, closely related to B. rochalimae, was found in fleas (Pulex irritans) from red foxes in Andalusia, Spain (65). PCR tests detected B. clarridgeiae and B. henselae in 20/34 and 4/34 fleas (Ctenocephalides felis), respectively, from red foxes in Australia, where it is an introduced species (105). Sréter-Lancz et al. (106) found Bartonella DNA in 4.2% (4/95) pools of fleas (P. irritans) from red foxes in Hungary, but all ticks from foxes were negative.
Similarly, there are numerous reports of Brucella infections in red foxes in Austria, Canada, Ireland, Russia, the U.S., and the UK (115, 125, 159–161). Tessaro (125) cultured Br. abortus from red foxes in Canada. Morton (48) cultured Br. suis biovar 4 from three out of 38 red foxes from Alaska. Scholtz et al. (161) cultured Br. microti and the proposed novel species Br. vulpis from red foxes in Austria in 2016. Br. suis biovar 4 cultures were obtained from arctic foxes from Alaska and Russia (11, 13, 48). McCue and O'Farrell (158) conducted a serological survey of San Joaquin kit foxes in California, U.S. and reported antibodies to Br. abortus in 8% in 1981–1982 and 3% in 1984 and to Br. canis in 14% in 1981–1982 and none in 1984.
Genus Cerdocyon
Investigators tested another fox species, the crab-eating fox (Cerdocyon thous), in Brazil and found B. rochalimae DNA in all nine P. irritans fleas collected from one animal (92). In another study by De Sousa et al. (66), none of the 78 sampled crab-eating foxes showed presence of Bartonella DNA in blood samples by qPCR.
Genus Urocyon
This genus contains two species of Western Hemisphere foxes: the gray fox (U. cinereoargenteus) and closely related island fox (U. littoralis), which is a dwarf cousin of the gray fox (239). There is a comprehensive study of Bartonella in gray foxes in northern California, U.S., conducted by Henn et al. (58). A novel B. clarridgeiae-like bacterium was isolated from 22 (42%) of 53 gray foxes and B. vinsonii subsp. berkhoffii from five gray foxes (9.4%). Serology showed that 48 gray foxes (89%) had detectable antibodies against Bartonella. The authors made the conclusion that the high prevalence of bacteremia and seroreactivity in gray foxes suggests that they may act as a reservoir species for the B. clarridgeiae-like species in this region. In another study of gray foxes in northern California, 14 (64%) of 22 foxes were infected with Bartonella species at one or more of the capture dates (97). Fleas collected from gray foxes in the study were identified as P. simulans, and 39% of the fleas were PCR positive for Bartonella, with B. rochalimae and B. vinsonii subsp. berkhoffii identified in 81% and 19% of the PCR positive fleas, respectively.
A serological survey of 132 gray foxes from Texas, U.S., demonstrated an antibody prevalence of 50% (66/132), with 22 (33.3%) individuals seropositive for B. clarridgeiae, eight (12.2%) for B. vinsonii subsp. berkhoffii, and 36 (54.5%) for both B. clarridgeiae and B. vinsonii subsp. berkhoffii (96). In gray foxes from Colorado, U.S., and northern Mexico Bartonella DNA was not detected (67, 77). Serological survey of the endangered island foxes (U. littoralis) conducted on several islands near the Californian coast by Namekata et al. (99) demonstrated a wide range of seroprevalence for B. clarridgeiae and B. vinsonii subsp. berkhoffii from 0% on San Nicolas Island to 86% on Santa Cruz Island. The following serological survey of 51 island foxes on Santa Rosa Island identified the overall antibody prevalence of 62.7% with 16 (31.4%) foxes seropositive for B. clarridgeiae only, five (9.8%) for B. vinsonii subsp. berkhoffii only, and 11 (21.6%) for both antigens (98). Importantly, B. vinsonii subsp. berkhoffii was isolated from six (11.8%) foxes using blood culture medium. All of the isolated B. vinsonii subsp. berkhoffii belonged to type III, the same type found in mainland gray foxes (98).
A culture of Br. abortus was obtained from one gray fox (U. cinereoargenteus) from Alabama, U.S. (126) while there were no positive results in foxes of this species in Arkansas, Florida, and South Carolina, U.S. (115, 156).
Genus Lycalopex
Several investigations of the South American foxes for Brucella infection have been published, including those investigating the pampas gray fox (L. gymnocercus) and Patagonian gray fox (L. griseus). Szyfres and González Tomé (154) found evidence of Brucella in both species from Argentina and isolated B. abortus biovar 1 from a pampas gray fox.
Genus Nyctereutes
The DNA identified as B. henselae was detected in spleens of two out of 142 raccoon dogs (N. procyonoides) in Korea, but not in any of 51 blood samples tested (94).
Family Ursidae
We found research on Bartonella and Brucella in three bear species, namely black bear (U. americanus), brown bear (U. arctos), and polar bear (U. maritimus). Bartonella DNA was not detected in seven black bears from Colorado, U.S. (77). All other research was focused on Brucella in bears (119, 147, 148, 179, 180, 182–185). Despite high seroprevalence levels for Br. abortus antibodies in all investigated bear species, we could not find any report on successful isolation of Brucella from these animals. Serological tests of 61 black bears for Br. canis by Bronson et al. (178) were negative.
Family Mephitidae
Twelve skunks of two species, the hooded skunk (Mephitis macroura) and the striped skunk (M. mephitis), from Colorado, U.S., and Mexico were found infected with B. rochalimae and one skunk from Mexico was infected with B. vinsonii subsp. berkhoffii (67, 77).
Antibodies against B. abortus were found in 8.7% of striped skunks and 3.9% of western spotted skunks (Spilogale gracilis) from California, U.S. (127).
Family Procyonidae
The common raccoon (Procyon lotor) has a natural range from southern Canada to Panama. Of 37 raccoons trapped on St. Simon Island in Georgia, U.S., 12 were positive for B. henselae and one for B. koehlerae (59). Interestingly, raccoons from the western regions of the U.S. carried different species of Bartonella. Henn et al. (88) isolated B. rochalimae from 11 of 42 raccoons from California, and Bai et al. (77) found 11 of 186 raccoons from Colorado PCR positive for B. rochalimae and three for B. vinsinii subsp. berkhoffii. All 977 raccoons from Japan, where it is an introduced species, were PCR negative for Bartonella (95).
Two Brucella strains cultured from raccoons from Alabama were identified as Br. abortus biovar 1 (126, 177). Raccoons seropositive to Brucella species were found in California, Alabama, Florida, and Texas in the U.S. (115, 126–128). None of 63 raccoons from Nebraska, U.S., had antibodies to Br. canis (139). In South Korea, Brucella DNA was found in blood (1/9) and tissues (2/5) of introduced raccoons (172). Three (8.8%) of 34 brown-nosed coatis (Nasua nasua), which also belong to family Procyonidae, were serologically positive for Brucella in the Brazilian Pantanal (149).
Family Mustelidae
Mustelidae is the largest family in the order Carnivora. Many terrestrial species of this genus were tested for Bartonella, including the beech marten (Martes foina), pine marten (M. martes), Japanese marten (M. melampus), American badger (Taxidea taxus), stoat (Mustela erminea), Japanese weasel (M. itatsi), least weasel (M. nivalis), Siberian weasel (M. sibirica), American mink (M. vison), European polecat (M. putorius), and ferret (M. putorius furo) (Table 1). However, out of 16 mustelid species tested for Bartonella DNA, only two cultures were obtained: one from a Japanese badger (Meles anakuma) and another from a Japanese marten (95). The isolate from the marten was close to Bartonella washoensis, a species typically found in squirrels, suggesting that it could have potentially “jumped” from a squirrel to its natural predator. The isolate from the Japanese badger was unique, with the closest match being to B. clarridgeiae and B. rochalimae (95). Bartonella clarridgeiae or related sequences were also detected in a beech marten and in European badgers, all from Spain (64, 83).
In North Carolina, U.S., Chinnadurai et al. (109) revealed a novel Bartonella species in 19 (29%) of 65 tested river otters (Lontra canadensis). Bartonella infection was detected in 45% (23/51) and 10% (3/30) of heart valves of northern and southern sea otters (Enhydra lutris kenyoni and E. l. nereis), respectively, by PCR (108). Analysis of the Bartonella ITS region identified two Bartonella species in those animals: a novel species closely related to Bartonella washoensis and Candidatus B. volans, whereas another genotype was molecularly identical to B. henselae. Sera from 50% of necropsied and 34% of presumed healthy, live-captured northern sea otters and in 16% of necropsied southern sea otters contained antibodies against Bartonella species (107).
Antibodies against Brucella species were detected in the American badger (Taxidea taxus), American mink (Neovison vison), European mink (Mustela lutreola), Eurasian otter (Lutra lutra), wolverine (Gulo gulo), and northern, southern and Asian sea otters (Enhydra lutris keyoni, lutris, nereis) from Europe, Asia, North and South Americas (11, 124, 125, 127, 174). We found only one report of successful culturing of Brucella (Br. abortus) from terrestrial mustelids (farmed European mink) and only one report of PCR detection of Brucella DNA in tissues of Asian badger (Meles leucurus) (172, 240). Similar to Bartonella, sea otters carry different species of Brucella than terrestrial mustelids. Investigating rectal swab samples of Asian sea otters (E. l. lutris) from Russia (168) found DNA of three Brucella species (Br. abortus, Br. melitensis, and Br. pinnipedialis). Miller et al. (169) isolated marine Brucella from a southern sea otter (E. l. nereis) with osteolytic lesions that was stranded on the central California coast. Antibodies to Brucella were detected in Northern sea otters (E. l. keyoni) from Alaska in the U.S. and Russia (120).
Families Phocidae, Otariidae, and Odobenodae
There is only one report on the identification of Bartonella in any of the pinnipeds, including walruses, eared seals, and true seals. Morick et al. (112) tested spleen samples and seal lice (Echinophtirius horridus) collected from seven harbor seals (Phoca vitulina). One spleen of 48 tissue samples and one of six lice pools were positive. The Bartonella species identified in the spleen and lice were found to be identical to each other by two genetic loci. One genetic marker identified the genotype as B. henselae, while another marker indicated 97% sequence similarity with B. grahamii.
In contrast to Bartonella, there is abundant evidence of Brucella infections in various species of the clade Pinnipedia. In family Phocidae (true seals), cultures of Brucella species were obtained from hooded seals (Cystophora cristata), gray seals (Halichoerus grypus), ringed seal (Phoca hispida), harp seal (Pagophilus groenlandicus), and harbor seal (Ph. vitulina) (170, 171, 206, 208, 214, 223, 224, 226, 231, 233). All identified cultures from true seals were Br. pinnipedialis. Serological evidence of Brucella was reported from investigation of even more species of true seals, including the bearded seal (Erignathus barbatus), ribbon seal (Histriophoca fasciata), leopard seal (Hydrurga leptonyx), Weddell seal (Leptonychotes weddellii), crab-eater seal (Lobodon carcinophaga), southern elephant seal (Mirounga leonina), Hawaiian monk seal (Neomonachus schauinslandi), Ross seal (Ommatophoca rossii), and several species of the genus Phoca.
In the family Odobenidae (walruses), Nielsen et al. reported serological prevalence of 12% (7/59) in 1996 and 3% (5/170) in 2001 in Atlantic walrus (Odobenus rosmarus rosmarus) from Canada; however serological tests of 40 Pacific walruses (O. r. divergens) from Alaska by Calle et al. (211) showed no antibodies to Brucella species
There are multiple reports of Brucella antibodies in fur seals and sea lions of the family Otariidae—nine species of the genera (Arctocephalus, Callorhinus, Eumetopias, Neophoca, Phocarctos, and Zalophus) (Table 2).
Differences in Distribution of Bartonella and Brucella Species in Wild Carnivores
Carnivores have regular exposure to both Bartonella and Brucella bacteria through predation on pathogen hosts, scavenging, and arthropod vectors. As with plague caused by Yersinia pestis (241), testing one carnivore for Bartonella and Brucella species could be equivalent to sampling a large number of its prey animals and give an idea of the epidemiological situation in the local environment. Overall, both Bartonella and Brucella are common in wildlife. Our review demonstrated numerous reports of infections caused by bacteria of both taxa in wild carnivores. We analyzed over 170 Bartonella and Brucella studies covering 109 species and subspecies of carnivores (Table 3). Eighty-four species of carnivores were tested for Brucella and 79% of these species were found positive by serological, bacteriological, or molecular methods. Out of 51 species examined in Bartonella studies, 71% tested positive.
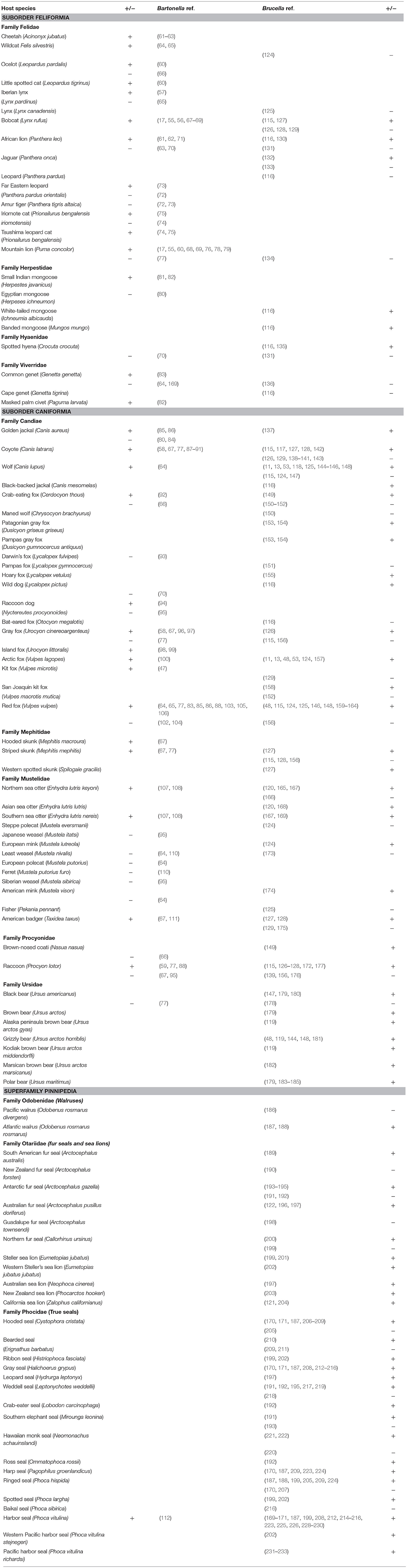
Table 3. Bartonella and Brucella studies in wild carnivores by species.
Although no species of wild carnivores were tested for both pathogens in a single study, 26 species were tested for both pathogens in different studies. Of those, 15 (58%) species were positive for both Bartonella and Brucella (among them bobcat, African lion, golden jackal, coyote, wolf, foxes, striped skunk, sea otters, raccoon, and harbor seal), meaning these carnivores can harbor either pathogen or potentially both. We know that other mammalian groups [bats for example, (242)] can be co-infected with Bartonella and Brucella species, and we speculate that this is also possible in carnivores, a hypothesis that definitely needs more investigation.
The most commonly identified Bartonella species was B. henselae, which was found in at least 23 species of wild carnivores, followed by B. rochalimae in 12, B. clarridgeiae in ten, and B. vinsinii subsp. berkhofii in seven species. Similarly, Br. abortus led the list of Brucella species, being identified in 36 terrestrial carnivore species, followed by Br. canis in eight. However, most of the reports are based on serology that cannot reliably discriminate these species until there are bacteriological data or sequences of PCR amplicons. Br. pinnipedialis is prevalent in marine carnivores, and some of the early reports of antibodies to Br. abortus in marine animals probably can be attributed to Br. pinnipedialis as well.
The analysis revealed some striking differences in distributions of these infectious agents in wild populations belonging to different carnivore families. One of the evident differences is abundance of several species of Bartonella practically in every explored species of wild felids. In contrast, very few reports of Brucella in the same species are available and those are limited to detection of antibodies that may indicate an exposure to the agent rather than direct involvement of these animals in the circulation of Brucella. At the same time, we could not find any report of Bartonella in bears while the presence of Brucella in these animals was well documented. An even more evident difference was found in marine carnivores, such as seals and sea lions, with practically every species reported infected with a specific species of Brucella (Br. pinnipedialis). In contrast, there is only one report of detection of Bartonella DNA in one tissue sample of a seal and there is no evidence of a Bartonella strain specific to marine mammals. A comparison with other marine mammals, such as dolphins, porpoises, and whales, which were not the subjects of our paper, also indicated a presence of specific Brucella species in blood of these animals, known as Brucella ceti. Whereas, the cat pathogen B. henselae was found in cetaceans, albeit less commonly than species of Brucella (243, 244).
Prevalence and the spectrum of bacterial species present depends on a potential exchange of bacteria between domestic and wild terrestrial carnivores. Wild carnivores are often infected with the same pathogens as their domesticated relatives (cats and dogs) though the risk of exposure varies widely because of differences in biology, distribution, and historical interactions. Confirmation of the identity of the bacterial species, however, remains critical for making such a statement regarding host specificity. Using a rapid test for differentiation of Bartonella species without sequencing amplicons, Carver et al. (114) came to the conclusion that free-ranging felids (pumas and bobcats) could be infected with Bartonella species that are generally considered to cross felid species barriers from domestic cats. Sequence analysis of some cultures and PCR amplicons has challenged such a conclusion. For example, in Californian mountain lions and bobcats Chomel et al. (55) found Bartonella species, typical for domestic cats (B. henselae and B. koehlerae); however, their detailed analysis demonstrated that these strains were sufficiently different for them to propose new subspecies of B. koehlerae (55). The authors who described the novel strains noted that these strains appear highly adapted to their particular species of wild cats and likely originated from a common ancestor.
There are some limitations in the analysis provided herein on the distribution of Bartonella and Brucella species in wild carnivores. The timing of samples collection for the animals listed in our review varied among studies and this factor could influence prevalence of infections. Differences in diagnostic methods used for identification can significantly affect comparison of the results. For a number of reasons, the number of Brucella studies relying on detection of antibodies in wild carnivores was much higher compared to the number of Bartonella studies in the same species that included either culturing or molecular detection. Several species of Brucella (Br. suis, Br. abortus, and Br. melitensis) are select agents and culturing of these species requires BSL-3 level capacity. Investigations of Brucella in wildlife started much earlier than similar investigations of Bartonella when DNA amplification techniques were not available. We should be carefull with interpretation of Brucella antibodies since available serological tests cannot identify all species of Brucella. There are separate tests for rough Brucella species (Br. canis) and for smooth Brucella species (Br. abortus, Br. melitensis, and Br. suis), and reported serology depends on the used tests.There are more described species of Bartonella (>35) and multiple diverse strains exist within this genus than for Brucella species. For many decades, the genus Brucella included six species, with some experts arguing that this genus is monospecific. In the past decade, new and more diverse Brucella species have been described (7). Recognition of the ubiquitous presence of Brucella in the environment will most likely continue (6). Nevertheless, reports of Brucella in wildlife without discrimination between species and biovars are still common, whereas future studies of Bartonella infections are more likely to be accompanied by proper identification down to species or subspecies level. Clearly, serological investigations are less informative for identification of bacterial species because of possible cross-reactivity between different antigens. The analysis presented in this review demonstrates the need for more information on genetic polymorphism of bacterial pathogens for the purposes of making comparison of strains from domestic and wild carnivores.
Evolutionary Aspects
Another issue that may influence the choice of methods for discriminating among Bartonella species is the effective level of association between these bacteria and their mammalian hosts, ranging from host species to host genus (245). Presumably, such a close bacteria-host association relates to the long-history of co-adaptation between Bartonella and their mammalian hosts and possibly arthropod vectors (245). An association of these bacteria with rodents, bats, and ruminants is described elsewhere, but analysis of the literature on Bartonella in wild carnivores also supports some degree of host-specificity (e.g., B. henselae in felids and B. vinsonii subsp. berkhoffii in canids).
A co-adaptation of Brucella with terrestrial wild carnivore hosts is not as straightforward as in domestic animals. A clear exception to this observation is Br. pinnipedialis, a species found in true seals only. Typical for domestic dogs, Br. canis may be expected to be commonly shared with wild canids, such as wolves and coyotes. However, this bacterial species has not been cultured from these predators and only few serological findings are available (115, 128, 139). Noticing the absence of Br. canis in wolves and coyotes, Moreno (7) proposed that this bacterial species evolved in the dog's ancestor after its predation on Br. suis biovar 4 infected animals (e.g., caribou/raindeer). This can be also explained by lack of specific serological tests available and low yield of culture.
Recent phylogenetic reconstructions and diversification analyses of prokaryotes have led to a better understanding of patterns of bacterial macroevolution. According to the analysis of prokaryote evolution based on the 16S rRNA gene (246), the common ancestor between the Brucella and the Bartonella genera split from the common ancestor with Phyllobacteriaceae in the order Rhizobiales about 567 million years ago and diverged about 507.4 million years ago (247) around the time of the Cambrian explosion and diversification of life during the Paleozoic Era, still on the giant supercontinent Pangea. As the species of the order Rhizobiales most closely related to Bartonella and Brucella are symbionts on plant roots, we can speculate that the ancestor of the two genera may have been a plant symbiont as well.
Bartonella evolved around 134 million years ago during Early Cretaceous Period around the time the flowering plants appeared in the middle of the dinosaur era (247). Segers et al. (248) suggest that the last common ancestor of the Bartonella was a gut symbiont of insects that produced its own amino acids and vitamins and that the adaptation to blood-feeding insects facilitated colonization of the mammalian bloodstream. Indeed, Bartonellaceae species were identified in honeybees (248, 249) and ants (250) filling the gap between the pathogenic Bartonella clade and more ancient bacterial symbionts. The honeybee strains of B. apis form a clade basal to species of the genus Bartonella (249). However, the B. apis genomes are almost twice as large (2.6 to 2.9 Mb) as the ant symbionts, suggesting that the association with the bee is more recent or that the association is less intimate (251). The phylogenetic trees show that the ant-related bacterial clade is a sister group to bee-related clade and other mammal-related Bartonella species (249, 252). Ants predate bees by some 35 million years in the order Hymenoptera which is 325 million years old itself (246). We can only speculate how the Bartonella ancestor adapted from a plant symbiont to gut symbiont through possible consumption routes and suggest looking into other “ancient” insect orders, like Archaeognata, or the orders that have maintained connection with water in their metamorphosis, like mayflies or dragonflies; and the ones that include sap-sucking insects.
Genomic and functional similarities between Br. suis and organisms from the Rhizobium Agrobacterium group suggest that the Brucella may have evolved from a soil plant-associated ancestral bacteria and speculatively, it may be metabolically active outside of a mammalian host (253). According to the analysis of prokaryote evolution based on the ribosomal gene, the genus Brucella is much younger than Bartonella and diverged about 230 thousand years ago (247) during Middle Pleistocene epoch. Previously it was hypothesized that Brucella species diverged roughly 20 million years ago following the divergence of their bovine and goat hosts (254). However, whole-genome-based phylogeny (255) supports the ribosomal gene analysis suggesting a much younger age for Brucella than previously estimated. Their rooted phylogeny suggests that brucellosis in various mammalian species emerged from infected sheep roughly in the past 86,000 to 296,000 years. This analysis has also suggested that transmittal of Brucella from pigs to canids likely happened within the past 22,500 years from infection of wolves or other canids feeding on pigs that were themselves infected (255). So, while possible paleo-brucellosis cases in the Bronze Age and later (256) fit perfectly within the timeframe, the possibility of brucellosis in a 2.5-million-year old hominid (257) brings an exciting prospect of an ancestral Brucella-like strain that either became extinct or has not been detected yet.
Conclusion
We can only speculate that a longer period of evolution of Bartonella has resulted in higher diversity and better co-adaptation to specific mammalian hosts compared to Brucella. Asymptomatic persistence of Bartonella bacteria in their natural reservoir animals contrasts with the well-documented pathological manifestations of Brucella in host animals. The only until the present time association of Bartonella infection with fatal cases of clinical disease in wild carnivores was reported in Florida pumas (76). Three diseased pumas had spent time in captivity prior to being released in the wild and were found later exhibiting respiratory signs and reluctance to move. Autopsy findings included necrotizing interstitial pneumonia and suppurative myocarditis associated with B. henselae infection (76). There is much more information on pathology caused by Brucella in domestic animals than in wildlife in general and even less in wild carnivores. Describing a range of pathologies caused by Brucella in sea mammals, Foster et al. (208) listed sub-blubber abscesses, hepatic and splenic necrosis, macrophage infiltration in liver and spleen, possible abortion, epididymitis, and meningitis.
In spite of shared mammalian reservoirs, the difference in transmission cycles presents distinct ecological traits. While Bartonella species use arthropod vectors as a main mechanism for transmission between mammalian hosts, the role of arthropod vectors in transmission of Brucella remains disputed. In our review, we provided some data, mostly from Russian sources, which support a potential role of ticks and other arthropods in transmission of Brucella. Nevertheless, it is hard to argue that such means of transmission are significant, let alone dominant, in transmission of these bacteria. Commonly, wild terrestrial predators contract brucellosis through consumption of infective tissues during predation and scavenging (258). Considering potential modes of Brucella transmission between marine mammals, Foster et al. (208) also included social interactions, sexual activity, maternal transmission, physical trauma, ingestion during feeding, and carriage by parasites.
We realize that our analyses create more questions than answers; the current review brought up significant parallels and differences in Bartonella and Brucella ecologies in wild carnivores and we hope it will prove to be useful for a wide range of specialists and can stimulate interest in comparing the ecologies of Bartonella and Brucella in wildlife and, at a larger scale, in investigating ecological trends of phylogenetically related zoonotic agents; benefitting epidemiological research and wildlife conservation.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Drs. Paul Mead, Kenneth Gage, Maria Negron, and John Goodrich for careful revision of the manuscript, thoughtful comments, and constructive suggestions. Special thanks should be given to the staff of the Steven B. Thacker CDC Library for their invaluable assistance in obtaining scientific materials.
References
1. Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature (2008) 451:990–3. doi: 10.1038/nature06536
2. Regier Y, O'Rourke F, Kempf VA. Bartonella spp. - a chance to establish One Health concepts in veterinary and human medicine. Parasit Vectors (2016) 9:261. doi: 10.1186/s13071-016-1546-x
3. Godfroid J, Garin-Bastuji B, Saegerman C, Blasco JM. Brucellosis in terrestrial wildlife. Rev Sci Tech. (2013) 32:27–42. doi: 10.20506/rst.32.1.2180
4. Moreno E, Stackebrandt E, Dorsch M, Wolters J, Busch M, Mayer H. Brucella abortus 16S rRNA and lipid A reveal a phylogenetic relationship with members of the alpha-2 subdivision of the class Proteobacteria. J Bacteriol. (1990) 172:3569–76. doi: 10.1128/jb.172.7.3569-3576.1990
5. Ben-Tekaya H, Gorvel JP, Dehio C. Bartonella and Brucella–weapons and strategies for stealth attack. Cold Spring Harb Perspect Med. (2013) 1:a010231. doi: 10.1101/cshperspect.a010231
6. Pappas G. The changing Brucella ecology: novel reservoirs, new threats. Int J Antimicrob Agents (2010) 36(Suppl. 1):S8–11. doi: 10.1016/j.ijantimicag.2010.06.013
7. Moreno E. Retrospective and prospective perspectives on zoonotic brucellosis. Front Microbiol. (2014) 5:213. doi: 10.3389/fmicb.2014.00213
8. Buffet JP, Kosoy M, Vayssier-Taussat M. Natural history of Bartonella-infecting rodents in light of new knowledge on genomics, diversity and evolution. Future Microbiol. (2013) 8:1117–28. doi: 10.2217/fmb.13.77
9. Alsmark CM, Frank AC, Karlberg EO, Legault BA, Ardell DH, Canbäck B, et al. The louse-borne human pathogen Bartonella quintana is a genomic derivative of the zoonotic agent Bartonella henselae. Proc. Natl. Acad. Sci. USA. (2004) 101:9716–21. doi: 10.1073/pnas.0305659101
10. Carmichael LE, Kenney RM. Canine abortion caused by Brucella canis. J Am Vet Med Assoc. (1968) 152:605–16.
11. Petukhova OS, Pinigin AF, Zabrodin VA, Vagina LA, Zabrodina EF. Isolation of Brucella from wild animals. Veterinariia (1971) 4:41–2.
12. Pavlov P. Teaching of E. N. Pavlovskii on natural foci of diseases and development of this teaching in Bulgaria. Zh Mikrobiol Epidemiol Immunobiol. (1960) 31:80–4.
13. Pinigin AF, Zabrodin VA. Natural foci of brucellosis. Vestnik sel' skokhoz nauki (1970) 7:96–99.
14. Chomel BB, Boulouis HJ, Maruyama S, Breitschwerdt EB. Bartonella spp. in pets and effect on human health. Emerg Infect Dis. (2006) 12:389–94. doi: 10.3201/eid1203.050931
15. Breitschwerdt EB, Maggi RG, Chomel BB, Lappin MR. Bartonellosis: an emerging infectious disease of zoonotic importance to animals and human beings. J Vet Emerg Critic Care (2010) 20:8–30. doi: 10.1111/j.1476-4431.2009.00496.x
16. Stuckey MJ, Chomel BB, de Fleurieu EC, Aguilar-Setién A, Boulouis HJ, Chang CC. Bartonella, bats and bugs: a review. Comp Immunol Microbiol Infect Dis. (2017) 55:20–9. doi: 10.1016/j.cimid.2017.09.001
17. Chomel BB, Kikuchi Y, Martenson JS, Roelke-Parker ME, Chang CC, Kasten RW, et al. Seroprevailance of Bartonella infection in American free-ranging and captive pumas (Felis concolor) and bobcats (Lynx rufus). Vet Res. (2004) 35:233–41. doi: 10.1051/vetres:2004001
18. Batut J, Andersson SG, O'Callaghan D. The evolution of chronic infection strategies in the alpha-proteobacteria. Nat Rev Microbiol. (2004) 12:933–45. doi: 10.1038/nrmicro1044
19. Dehio C, Tsolis RM. Type IV effector secretion and subversion of host functions by Bartonella and Brucella species. Curr Top Microbiol Immunol. (2017) 413:269–95. doi: 10.1007/978-3-319-75241-9_11
20. Boschiroli ML, Foulongne V, O'Callaghan D. Brucellosis: a worldwide zoonosis. Curr Opin Microbiol. (2001) 4:58–64. doi: 10.1016/S1369-5274(00)00165-X
21. Harms A, Dehio C. (2012). Intruders below the radar: molecular pathogenesis of Bartonella spp. Clin Microbiol Rev. 25, 42–78. doi: 10.1128/CMR.05009-11
22. Birtles RJ. Bartonellae as elegant hemotropic parasites. Ann NY Acad Sci. (2005) 1063:270–9. doi: 10.1196/annals.1355.044
23. Cutler SJ, Whatmore AM, Commander NJ. Brucellosis-new aspects of an old disease. J Appl Microbiol. (2005) 98:1270–81. doi: 10.1111/j.1365-2672.2005.02622.x
24. Chomel BB, Boulouis HJ, Breitschwerdt EB, Kasten RW, Vayssier-Taussat M, Birtles RJ, et al. Ecological fitness and strategies of adaptation of Bartonella species to their hosts and vectors. Vet Res. (2009) 40:29. doi: 10.1051/vetres/2009011
25. Billeter SA, Levy MG, Chomel BB, Breitschwerdt EB. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med Vet Entomol. (2008) 22:1–15. doi: 10.1111/j.1365-2915.2008.00713.x
26. Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, et al. Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. (1996) 34:1952–6.
27. Finkelstein JL, Brown TP, O'Reilly KL, Wedincamp JJr, Foil LD. Studies on the growth of Bartonella henselae in the cat flea (Siphonaptera: Pulicidae). J Med Entomol. (2002) 39:915–9. doi: 10.1603/0022-2585-39.6.915
28. Seki N, Kasai S, Saito N, Komagata O, Mihara M, Sasaki T, et al. Quantitative analysis of proliferation and excretion of Bartonella quintana in body lice, Pediculus humanus L. Am J Trop Med Hyg. (2007) 77:562–6. doi: 10.4269/ajtmh.2007.77.562
29. Reis C, Cote M, Le Rhun D, Lecuelle B, Levin ML, Vayssier-Taussat M, et al. Vector competence of the tick Ixodes ricinus for transmission of Bartonella birtlesii. PLoS Negl Trop Dis. (2011) 5:e1186. doi: 10.1371/journal.pntd.0001186
30. Cotté V, Bonnet S, Le Rhun D, Le Naour E, Chauvin A, Boulouis HJ, et al. Transmission of Bartonella henselae by Ixodes ricinus. Emerg Infect Dis. (2008) 14:1074–80. doi: 10.3201/eid1407.071110
31. Telford SR, Wormser GP. Bartonella spp. transmission by ticks not established. Emerg Infect Dis. (2010) 16:379–84. doi: 10.3201/eid1603.090443
32. Kang JG, Won S, Kim HW, Kim BJ, Park BK, Park TS, et al. Molecular detection of Bartonella spp. in terrestrial leeches (Haemadipsa rjukjuana) feeding on human and animal blood in Gageo-do, Republic of Korea. Parasit Vectors (2016) 9:326. doi: 10.1186/s13071-016-1613-3
33. Cantlay JC, Ingram DJ, Meredith AL. A review of zoonotic infection risks associated with the wild meat trade in Malaysia. Ecohealth (2017) 14:361–88. doi: 10.1007/s10393-017-1229-x
34. Angelakis E, Lepidi H, Canel A, Rispal P, Perraudeau F, Barre I, et al. Human case of Bartonella alsatica lymphadenitis. Emerging Infect Dis. (2008) 14:1951–3. doi: 10.3201/eid1412.080757
35. Raoult D, Roblot F, Rolain JM, Besnier JM, Loulergue J, Bastides F, et al. First isolation of Bartonella alsatica from a valve of a patient with endocarditis. J Clin Microbiol. (2006) 44:278–9. doi: 10.1128/JCM.44.1.278-279.2006
36. Rolain JM, Boureau-Voultoury A, Raoult D. Serological evidence of Bartonella vinsonii lymphadenopathies in a child bitten by a dog. Clin Microbiol Infect. (2009) 15 Suppl 2:122–3. doi: 10.1111/j.1469-0691.2008.02197.x
37. Kosoy MY, Regnery RL, Kosaya OI, Jones DC, Marston EL, Childs JE. Isolation of Bartonella spp. from embryos and neonates of naturally infected rodents. J Wildl Dis. (1998) 34:305–9. doi: 10.7589/0090-3558-34.2.305
38. Guptill L, Slater LN, Wu CC, Lin TL, Glickman LT, Welch DF, et al. Evidence of reproductive failure and lack of perinatal transmission of Bartonella henselae in experimentally infected cats. Vet Immunol Immunopathol. (1998) 65:177–89. doi: 10.1016/S0165-2427(98)00153-6
39. Hensel ME, Negron M, Arenas-Gamboa AM. Brucellosis in dogs and public health risk. Emerging Infect Dis. (2018) 24:1401–6. doi: 10.3201/eid2408.171171
41. Carmichael LE, Joubert JC. Transmission of Brucella canis by contact exposure. Cornell Vet. (1988) 78:63–73.
42. Vitry MA, Hanot Mambres D, Deghelt M, Hack K, Machelart A, Lhomme F, et al. Brucella melitensis invades murine erythrocytes during infection. Infect. Immun. (2014) 82:3927–38. doi: 10.1128/IAI.01779-14
43. Rementsova MM. Brucellosis in Wild Animals. In: Galuzo and Gvozdev, editors. New Delhi: Oxonian Press (1987).
44. Zheludkov MM, Tsilerson LE. Reservoirs of Brucella infection in nature. Biol Bull Russ Acad Sci. (2010) 37:709–15. doi: 10.1134/S106235901007006X
45. Neglia G, Veneziano V, De Carlo E, Galiero G, Borriello G, Francillo M, et al. Detection of Brucella abortus DNA and RNA in different stages of development of the sucking louse Haematopinus tuberculatus. BMC Vet Res. (2013) 9:236. doi: 10.1186/1746-6148-9-236
46. Scanlan CM, Pidgeon GL, Swango LJ, Hannon SS, Galik PA. Experimental infection of gray foxes (Urocyon cinereoargenteus) with Brucella abortus. J Wildl Dis. (1984) 20:27–30. doi: 10.7589/0090-3558-20.1.27
47. Neiland KA, Miller LG. Experimental Brucella suis type 4 infections in domestic and wild Alaskan carnivores. J Wildl Dis. (1981) 17:183–9. doi: 10.7589/0090-3558-17.2.183
48. Morton JK. Brucella suis Type 4 in Foxes and Their Role as Reservoirs/Vectors Among Reindeer. Ph.D. thesis, University of Alaska, Fairbanks, AK (1989).
49. Islam MA, Khatun MM, Baek BK. Male rats transmit Brucella abortus biotype 1 through sexual intercourse. Vet Microbiol. (2013) 165:475–7. doi: 10.1016/j.vetmic.2013.04.016
50. Hashino M, Kim S, Tachibana M, Shimizu T, Watarai M. Vertical transmission of Brucella abortus causes sterility in pregnant mice. J Vet Med Sci. (2012) 74:1075–7. doi: 10.1292/jvms.11-0566
51. Wang Z, Wang SS, Wang GL, Wu TL, Lv YL, Wu QM. A pregnant mouse model for the vertical transmission of Brucella melitensis. Vet J. (2014) 200:116–21. doi: 10.1016/j.tvjl.2013.12.021
52. Guzmán-Verri C, González-Barrientos R, Hernández-Mora G, Morales JA, Baquero-Calvo E, Chaves-Olarte E, et al. Brucella ceti and brucellosis in cetaceans. Front Cell Infect Microbiol. (2012) 2:3. doi: 10.3389/fcimb.2012.00003
53. Egorov IY, Kalinovskii AI, Maramovich AS, Chernyavskii VF. Problems of epidemiological surveillance of brucellosis under conditions of deer breeding in the North. Epidemiol Infekts Bolezni. (1997) 3:18–21.
54. Scholz HC, Hubalek Z, Nesvadbova J, Tomaso H, Vergnaud G, Le Flèche P, et al. Isolation of Brucella microti from soil. Emerging Infect Dis. (2008) 14:1316–7. doi: 10.3201/eid1408.080286
55. Chomel BB, Molia S, Kasten RW, Borgo GM, Stuckey MJ, Maruyama S, et al. Isolation of Bartonella henselae and two new Bartonella subspecies, Bartonella koehlerae subspecies boulouisii subsp. nov and Bartonella koehlerae subspecies bothieri subsp nov from free-ranging Californian mountain lions and bobcats. PLoS ONE (2016) 11:e0148299. doi: 10.1371/journal.pone.0148299
56. Riley SP, Foley J, Chomel B. Exposure to feline and canine pathogens in bobcats and gray foxes in urban and rural zones of a national park in California. J Wildl Dis. (2004) 40:11–22. doi: 10.7589/0090-3558-40.1.11
57. Meli ML, Cattori V, Martínez F, López G, Vargas A, Simón MA, et al. Feline leukemia virus and other pathogens as important threats to the survival of the critically endangered Iberian lynx (Lynx pardinus). PLoS ONE (2009) 4:e4744. doi: 10.1371/journal.pone.0004744
58. Henn JB, Gabriel MW, Kasten RW, Brown RN, Theis JH, Foley JE, et al. Gray foxes (Urocyon cinereoargenteus) as a potential reservoir of a Bartonella clarridgeiae-like bacterium and domestic dogs as part of a sentinel system for surveillance of zoonotic arthropod-borne pathogens in northern California. J Clin Microbiol. (2007) 45:2411–8. doi: 10.1128/JCM.02539-06
59. Hwang J, Gottdenker NL. Bartonella species in raccoons and feral cats, Georgia, USA. Emerging Infect Dis. (2013) 19:1167–8. doi: 10.3201/eid1907.130010
60. Filoni C, Catão-Dias JL, Bay G, Durigon EL, Jorge RS, Lutz H, et al. First evidence of feline herpesvirus, calicivirus, parvovirus, and Ehrlichia exposure in Brazilian free-ranging felids. J Wildl Dis. (2006) 42:470–7. doi: 10.7589/0090-3558-42.2.470
61. Molia S, Kasten RW, Stuckey MJ, Boulouis HJ, Allen J, Borgo GM, et al. Isolation of Bartonella henselae, Bartonella koehlerae subsp. koehlerae, Bartonella koehlerae subsp bothieri and a new subspecies of B koehlerae from free-ranging lions (Panthera leo) from South Africa, cheetahs (Acinonyx jubatus) from Namibia and captive cheetahs from California. Epidemiol Infect. (2016) 144:3237–43. doi: 10.1017/S0950268816001394
62. Molia S, Chomel BB, Kasten RW, Leutenegger CM, Steele BR, Marker L, et al. Prevalence of Bartonella infection in wild African lions (Panthera leo) and cheetahs (Acinonyx jubatus). Vet Microbiol. (2004) 100:31–41. doi: 10.1016/j.vetmic.2004.01.007
63. Kelly PJ, Rooney JJ, Marston EL, Jones DC, Regnery RL. Bartonella henselae isolated from cats in Zimbabwe. Lancet (1998) 351:1706. doi: 10.1016/S0140-6736(05)77744-8
64. Gerrikagoitia X, Gil H, García-Esteban C, Anda P, Juste RA, Barral M. Presence of Bartonella species in wild carnivores of northern Spain. Appl Environ Microbiol. (2012) 78:885–8. doi: 10.1128/AEM.05938-11
65. Márquez FJ, Millán J, Rodríguez-Liébana JJ, García-Egea I, Muniain MA. Detection and identification of Bartonella sp. in fleas from carnivorous mammals in Andalusia, Spain. Med Vet Entomol. (2009) 23:393–8. doi: 10.1111/j.1365-2915.2009.00830.x
66. De Sousa KCM, do Amaral RB, Herrera HM, Santos FM, Macedo GC, de Andrade Pinto PCE, et al. Genetic diversity of Bartonella spp. in wild mammals and ectoparasites in Brazilian Pantanal. Microb Ecol. (2018) 76:544–54. doi: 10.1007/s00248-017-1138-0
67. López-Pérez AM, Osikowicz L, Bai Y, Montenieri J, Rubio A, Moreno K, et al. Prevalence and phylogenetic analysis of Bartonella species of wild carnivores and their fleas in Northwestern Mexico. Ecohealth (2017) 14:116–29. doi: 10.1007/s10393-017-1216-2
68. Bevins SN, Carver S, Boydston EE, Lyren LM, Alldredge M, Logan KA, et al. Three pathogens in sympatric populations of pumas, bobcats, and domestic cats: implications for infectious disease transmission. PLoS ONE (2012) 7:e31403. doi: 10.1371/journal.pone.0031403
69. Yamamoto K, Chomel BB, Lowenstine LJ, Kikuchi Y, Phillips LG, Barr BC, et al. Bartonella henselae antibody prevalence in free-ranging and captive wild felids from California. J Wildl Dis. (1998) 34:56–63. doi: 10.7589/0090-3558-34.1.56
70. Williams BM, Berentsen A, Shock BC, Teixiera M, Dunbar MR, Becker MS, et al. Prevalence and diversity of Babesia, Hepatozoon, Ehrlichia, and Bartonella in wild and domestic carnivores from Zambia, Africa. Parasitol Res. (2014) 113:911–8. doi: 10.1007/s00436-013-3722-7
71. Pretorius AM, Kuyl JM, Isherwood DR, Birtles RJ. Bartonella henselae in African lion, South Africa. Emerging Infect Dis. (2004) 10:2257–8. doi: 10.3201/eid1012.031054
72. Goodrich JM, Lewis JCM, Quigley KS, Roelke M, Astafiev AA, Slabii EV, et al. Infectious diseases of Amur tigers and Far Eastern leopards. In: Seryodkin IV, and Miquelle DG, editors. Diseases and Parasites of Wildlife in Siberia and the Russian Far East: Monograph. Vladivostok: Dalnauka (2012). p. 19–26.
73. Quigley KS, Armstrong DL, Miquelle DG, Goodrich JM, Quigley HB. Health evaluation of wild Siberian tigers (Panthera tigris altaica) and Amur leopards (Panthera pardus orientalis) in the Russian Far East. In: Proceedings AAZV, AAWV, ARAV, NAZWV Joint Conference (Orlando, FL) (2001).
74. Tateno M, Sunahara A, Nakanishi N, Izawa M, Matsuo T, Setoguchi A, et al. Molecular survey of arthropod-borne pathogens in ticks obtained from Japanese wildcats. Ticks Tick Borne Dis. (2015) 6:281–9. doi: 10.1016/j.ttbdis.2015.01.009
75. Tateno M, Nishio T, Sakuma M, Nakanishi N, Izawa M, Asari Y, et al. Molecular epidemiologic survey of Bartonella, Ehlichia, and Anaplasma infections in Japanese Iriomote and Tsushima leopard cats. J Wildl Dis. (2013) 49:646–52. doi: 10.7589/2012-07-194
76. Elsmo EJ, Fenton H, Cleveland CA, Shock B, Cunningham M, Howerth EW, et al. Necrotizing interstitial pneumonia and suppurative myocarditis associated with Bartonella henselae infection in three Florida pumas. J Vet Diagnos Invest. (2018) 30:728–732. doi: 10.1177/1040638718789226
77. Bai Y, Gilbert A, Fox K, Osikowicz L, Kosoy M. Bartonella rochalimae and B. vinsonii subsp berkhoffii in wild carnivores from Colorado, USA. J Wildl Dis. (2016) 52:844–9. doi: 10.7589/2016-01-015
78. Girard YA, Swift P, Chomel BB, Kasten RW, Fleer K, Foley JE, et al. Zoonotic vector-borne bacterial pathogens in California mountain lions (Puma concolor), 1987-2010. Vector Borne Zoonotic Dis. (2012) 12:913–21. doi: 10.1089/vbz.2011.0858
79. Rotstein DS, Taylor SK, Bradley J, Brieitschwerdt EB. Prevalence of Bartonella henselae antibody in Florida panthers. J Wildl Dis. (2000) 36:157–60. doi: 10.7589/0090-3558-36.1.157
80. Leulmi H, Aouadi A, Bitam I, Bessas A, Benakhla A, Raoult D, et al. Detection of Bartonella tamiae, Coxiella burnetii and rickettsiae in arthropods and tissues from wild and domestic animals in northeastern Algeria. Parasit Vectors (2016) 9:27. doi: 10.1186/s13071-016-1316-9
81. Jaffe DA, Chomel BB, Kasten RW, Breitschwerdt EB, Maggi RG, McLeish A, et al. Bartonella henselae in small Indian mongooses (Herpestes auropunctatus) from Grenada, West Indies. Vet Microbiol. (2018) 216:119–22. doi: 10.1016/j.vetmic.2018.02.009
82. Sato S, Kabeya H, Shigematsu Y, Sentsui H, Une Y, Minami M, et al. Small Indian mongooses and masked palm civets serve as new reservoirs of Bartonella henselae and potential sources of infection for humans. Clin Microbiol Infect. (2013) 19:1181–7. doi: 10.1111/1469-0691.12164
83. Millán J, Proboste T, Fernández de Mera IG, Chirife AD, de la Fuente J, Altet L. Molecular detection of vector-borne pathogens in wild and domestic carnivores and their ticks at the human-wildlife interface. Ticks Tick Borne Dis. (2016) 7:284–90. doi: 10.1016/j.ttbdis.2015.11.003
84. Sukara R, Chochlakis D, Ćirović D, Penezić A, Mihaljica D, Ćakić S, et al. Golden jackals (Canis aureus) as hosts for ticks and tick-borne pathogens in Serbia. Ticks Tick Borne Dis. (2018) 9:1090–7. doi: 10.1016/j.ttbdis.2018.04.003
85. Marciano O, Gutiérrez R, Morick D, King R, Nachum-Biala Y, Baneth G, et al. Detection of Bartonella spp. in wild carnivores, hyraxes, hedgehog and rodents from Israel. Parasitology (2016) 143:1232–42. doi: 10.1017/S0031182016000603
86. Chomel BB, McMillan-Cole AC, Kasten RW, Stuckey MJ, Sato S, Maruyama S, et al. Candidatus Bartonella merieuxii, a potential new zoonotic Bartonella species in canids from Iraq. PLoS Negl Trop Dis. (2012) 6:e1843. doi: 10.1371/journal.pntd.0001843
87. Kehoe SP, Chomel BB, Stuckey MJ, Kasten RW, Balakrishnan N, Sacks BN, et al. Zoonotic Bartonella species in cardiac valves of healthy coyotes, California, USA. Emerging Infect Dis. (2014) 20:2133–6. doi: 10.3201/eid2012.140578
88. Henn JB, Chomel BB, Boulouis HJ, Kasten RW, Murray WJ, Bar-Gal GK, et al. Bartonella rochalimae in raccoons, coyotes, and red foxes. Emerging Infect Dis. (2009) 15:1984–7. doi: 10.3201/eid1512.081692
89. Beldomenico PM, Chomel BB, Foley JE, Sacks BN, Baldi CJ, Kasten RW, et al. Environmental factors associated with Bartonella vinsonii subsp. berkhoffii seropositivity in free-ranging coyotes from northern California. Vector Borne Zoonotic Dis. (2005) 5:110–9. doi: 10.1089/vbz.2005.5.110
90. Chang CC, Kasten RW, Chomel BB, Simpson DC, Hew CM, Kordick DL, et al. Coyotes (Canis latrans) as the reservoir for a human pathogenic Bartonella sp: molecular epidemiology of Bartonella vinsonii subsp berkhoffii infection in coyotes from central coastal California. J Clin Microbiol. (2000) 38:4193–200.
91. Chang C, Yamamoto K, Chomel BB, Kasten RW, Simpson DC, Smith CR, et al. Seroepidemiology of Bartonella vinsonii subsp. berkhoffii infection in California coyotes, 1994–1998. Emerg Infect Dis. (1998) 5:711–5. doi: 10.3201/eid0505.990514
92. Fontalvo MC, Favacho ARM, Araujo AC, Santos NMD, Oliveira GMB, Aguiar DM, et al. Bartonella species pathogenic for humans infect pets, free-ranging wild mammals and their ectoparasites in the Caatinga biome, Northeastern Brazil: a serological and molecular study. Braz J Infect Dis. (2017) 21:290–6. doi: 10.1016/j.bjid.2017.02.002
93. Cabello J, Altet L, Napolitano C, Sastre N, Hidalgo E, Dávila JA, et al. Survey of infectious agents in the endangered Darwin's fox (Lycalopex fulvipes): high prevalence and diversity of hemotrophic mycoplasmas. Vet Microbiol. (2013) 167:448–54. doi: 10.1016/j.vetmic.2013.09.034
94. Kang JG, Chae JB, Cho YK, Jo YS, Shin NS, Lee H, et al. Molecular detection of Anaplasma, Bartonella, and Borrelia theileri in raccoon dogs (Nyctereutes procyonoides) in Korea. Am J Trop Med Hyg. (2018) 98:1061–8. doi: 10.4269/ajtmh.17-0380
95. Sato S, Kabeya H, Miura T, Suzuki K, Bai Y, Kosoy M, et al. Isolation and phylogenetic analysis of Bartonella species from wild carnivores of the suborder Caniformia in Japan. Vet Microbiol. (2012) 161:130–6. doi: 10.1016/j.vetmic.2012.07.012
96. Schaefer JD, Moore GM, Namekata MS, Kasten RW, Chomel BB. Seroepidemiology of Bartonella infection in gray foxes from Texas. Vector Borne Zoonotic Dis. (2012) 12:428–30. doi: 10.1089/vbz.2011.0805
97. Gabriel MW, Henn J, Foley JE, Brown RN, Kasten RW, Foley P, et al. Zoonotic Bartonella species in fleas collected on gray foxes (Urocyon cinereoargenteus). Vector Borne Zoonotic Dis. (2009) 9:597–602. doi: 10.1089/vbz.2008.0134
98. Schaefer JD, Kasten RW, Coonan TJ, Clifford DL, Chomel BB. Isolation or detection of Bartonella vinsonii subspecies berkhoffii and Bartonella rochalimae in the endangered island foxes (Urocyon littoralis). Vet Microbiol. (2011) 154:135–9. doi: 10.1016/j.vetmic.2011.06.031
99. Namekata MS, Clifford DL, Kasten RW, Henn JB, Garcelon DK, Coonan TJ, et al. Seroprevalence of Bartonella spp. in the endangered island fox (Urocyon littoralis). Vet Microbiol. (2009) 136:184–7. doi: 10.1016/j.vetmic.2008.10.017
100. Mascarelli PE, Elmore SA, Jenkins EJ, Alisauskas RT, Walsh M, Breitschwerdt EB, et al. Vector-borne pathogens in arctic foxes, Vulpes lagopus, from Canada. Res Vet Sci. (2015) 99:58–9. doi: 10.1016/j.rvsc.2014.12.011
101. Víchová B, Bona M, Miterpáková M, Kraljik J, Čabanová V, Nemcíková G, et al. Fleas and ticks of red foxes as vectors of canine bacterial and parasitic pathogens, in Slovakia, Central Europe. Vec Borne Zoonot Dis. (2018) 18:611–9. doi: 10.1089/vbz.2018.2314
102. Andersson MO, Tolf C, Tamba P, Stefanache M, Radbea G, Frangoulidis D, et al. Molecular survey of neglected bacterial pathogens reveals an abundant diversity of species and genotypes in ticks collected from animal hosts across Romania. Parasit Vectors (2018) 11:144. doi: 10.1186/s13071-018-2756-1
103. Hodžić A, Mrowietz N, Cézanne R, Bruckschwaiger P, Punz S, Habler VE, et al. Occurrence and diversity of arthropod-transmitted pathogens in red foxes (Vulpes vulpes) in western Austria, and possible vertical (transplacental) transmission of Hepatozoon canis. Parasitology (2017) 24:1–10. doi: 10.1017/S0031182017001536
104. Hodžić A, Alić A, Fuehrer HP, Harl J, Wille-Piazzai W, Duscher GG. A molecular survey of vector-borne pathogens in red foxes (Vulpes vulpes) from Bosnia and Herzegovina. Parasit Vectors (2015) 8:88. doi: 10.1186/s13071-015-0692-x
105. Kaewmongkol G, Kaewmongkol S, Fleming PA, Adams PJ, Ryan U, Irwin PJ, et al. Zoonotic Bartonella species in fleas and blood from red foxes in Australia. Vector Borne Zoonotic Dis. (2011) 11:1549–53. doi: 10.1089/vbz.2011.0646
106. Sréter-Lancz Z, Tornyai K, Széll Z, Sréter T, Márialigeti K. Bartonella infections in fleas (Siphonaptera: Pulicidae) and lack of bartonellae in ticks (Acari: Ixodidae) from Hungary. Folia Parasitol. (2006) 53:313–6. doi: 10.14411/fp.2006.039
107. Carrasco SE, Chomel BB, Gill VA, Doroff AM, Miller MA, Burek-Huntington KA, et al. Bartonella spp. exposure in northern and southern sea otters in Alaska and California. Vector Borne Zoonotic Dis. (2014) 14:831–7. doi: 10.1089/vbz.2014.1612
108. Carrasco SE, Chomel BB, Gill VA, Kasten RW, Maggi RG, Breitschwerdt EB, et al. Novel Bartonella infection in northern and southern sea otters (Enhydra lutris kenyoni and Enhydra lutris nereis). Vet Microbiol. (2014) 170:325–34. doi: 10.1016/j.vetmic.2014.02.021
109. Chinnadurai SK, Birkenheuer AJ, Blanton HL, Maggi RG, Belfiore N, Marr HS, et al. Prevalence of selected vector-borne organisms and identification of Bartonella species DNA in North American river otters (Lontra canadensis). J Wildl Dis. (2010) 46:947–50. doi: 10.7589/0090-3558-46.3.947
110. McDonald RA, Day MJ, Birtles RJ. Diseases and pathogens of stoats and other wildlife in New Zealand. Dep. Conserv. Sci. Inter. Series (2004) 171:1–23.
111. Quinn JH, Girard YA, Gilardi K, Hernandez Y, Poppenga R, Chomel BB, et al. Pathogen and rodenticide exposure in American badgers (Taxidea taxus) in California. J Wildl Dis. (2012) 48:467–72. doi: 10.7589/0090-3558-48.2.467
112. Morick D, Osinga N, Gruys E, Harrus S. Identification of a Bartonella species in the harbor seal (Phoca vitulina) and in seal lice (Echinophtirius horridus). Vector Borne Zoonotic Dis. (2009) 9:751–3. doi: 10.1089/vbz.2008.0202
113. Jameson P, Greene C, Regnery R, Dryden M, Marks A, Brown J, et al. Prevalence of Bartonella henselae antibodies in pet cats throughout regions of North America. J. Infect. Dis. (1995) 172:1145–9. doi: 10.1093/infdis/172.4.1145
114. Carver S, Bevins SN, Lappin MR, Boydston EE, Lyren LM, Alldredge M, et al. Pathogen exposure varies widely among sympatric populations of wild and domestic felids across the United States. Ecol Appl. (2016) 26:367–81. doi: 10.1890/15-0445
115. Hoff GL, Bigler WJ, Trainer DO, Debbie JG, Brown GM, Winkler WG, et al. Survey of selected carnivore and opossum serums for agglutinins to Brucella canis. J Am Vet Med Assoc. (1974) 165:830–1.
116. Sachs R, Staak C, Groocock CM. Serological investigation of brucellosis in game animals in Tanzania. Bull Epizoot Dis Afr. (1968) 16:93–100.
117. Williams JD, Heck FC, Davis DS, Adams LG. Comparison of results from five serologic methods used for detecting Brucella abortus antibody activity in coyote sera. Vet Immunol Immunopathol. (1991) 29:79–87. doi: 10.1016/0165-2427(91)90054-G
118. Neiland KA. Rangiferine brucellosis in Alaskan canids. J. Wildl. Dis. (1970) 6:136–19. doi: 10.7589/0090-3558-6.3.136
119. Godfroid J, Beckmen K, Helena Nymo I. Removal of lipid from serum increases coherence between brucellosis rapid agglutination test and enzyme-linked immunosorbent assay in bears in Alaska, USA. J Wildl Dis. (2016) 52:912–5. doi: 10.7589/2015-11-298
120. Goldstein T, Gill VA, Tuomi P, Monson D, Burdin A, Conrad PA, et al. Assessment of clinical pathology and pathogen exposure in sea otters (Enhydra lutris) bordering the threatened population in Alaska. J Wildl Dis. (2011) 47:579–92. doi: 10.7589/0090-3558-47.3.579
121. Avalos-Téllez R, Ramírez-Pfeiffer C, Hernández-Castro R, Díaz-Aparicio E, Sánchez-Domínguez C, Zavala-Norzagaray A, et al. Infection of California sea lions (Zalophus californianus) with terrestrial Brucella spp. Vet J. (2014) 202:198–200. doi: 10.1016/j.tvjl.2014.06.021
122. Lynch M, Nielsen O, Duignan PJ, Kirkwood R, Hoskins A, Arnould JP. Serologic survey for potential pathogens and assessment of disease risk in Australian fur seals. J Wildl Dis. (2011b) 47:555–65. doi: 10.7589/0090-3558-47.3.555
123. Johnson MR. The disease ecology of brucellosis and tuberculosis in potential relationship to Yellowstone wolf populations. In: Wolves for Yellowstone? A Report to the United States Congress, Volume IV Research and Analysis (Yellowstone National Park, WY) (1992). p. 5–72.
125. Tessaro SV. The existing and potential importance of brucellosis and tuberculosis in Canadian wildlife: a review. Can Vet J. (1986) 27:119–24.
126. Schnurrenberger PR, Brown RR, Hill EP, Scanlan CM, Altiere JA, Wykoff JT. Brucella abortus in wildlife on selected cattle farms in Alabama. J Wildl Dis. (1985) 21:132–6. doi: 10.7589/0090-3558-21.2.132
127. Hoq A. A serologic survey of Brucella agglutinins in wildlife and sheep. Calif Vet. (1978) 32:15–17.
128. Randhawa AS, Kelly VP, Baker EF Jr. Agglutinins to Coxiella burnetii and Brucella spp, with particular reference to Brucella canis, in wild animals of southern Texas. J. Am. Vet. Med. Assoc. (1977) 171:939–942.
129. Vest ED, Lundgren DL, Parker DD, Johnson DE, Morse EL, Bushman JB, et al. Results of a five-year survey for certain enzootic diseases in the fauna of western Utah. Am J Trop Med Hyg. (1965) 14:124–35. doi: 10.4269/ajtmh.1965.14.124
130. Assenga JA, Matemba LE, Muller SK, Malakalinga JJ, Kazwala RR. Epidemiology of Brucella infection in the human, livestock and wildlife interface in the Katavi-Rukwa ecosystem, Tanzania. BMC Vet Res. (2015) 11:189. doi: 10.1186/s12917-015-0504-8
131. De Vos V, Van Niekerk CAWJ. Brucellosis in the Kruger National Park. J S Afr Vet Med Assoc. (1969) 40:331–4.
132. Furtado MM, Gennari SM, Ikuta CY, Jácomo AT, de Morais ZM, Pena HF, et al. Serosurvey of smooth Brucella, Leptospira spp. and Toxoplasma gondii in free-ranging jaguars (Panthera onca) and domestic animals from Brazil. PLoS ONE (2015) 10:e0143816. doi: 10.1371/journal.pone.0143816
133. Onuma SSM, Kantek DLZ, Crawshaw PG Jr, Morato RG, May-Junior JA, Morais ZMD, et al. Detection of Leptospira spp. and Brucella abortus antibodies in free-living jaguars (Panthera onca) in two protected areas of Northern Pantanal, Brazil. Rev Instit Med Trop São Paulo (2015) 57:177–80. doi: 10.1590/S0036-46652015000200014
134. Roelke ME, Forrester DJ, Jacobson ER, Kollias GV, Scott FW, Barr MC, et al. Seroprevalence of infectious disease agents in free-ranging Florida panthers (Felis concolor coryi). J Wildl Dis. (1993) 29:36–49. doi: 10.7589/0090-3558-29.1.36
135. Sachs R, Staak C. Evidence of brucellosis in antelopes of the Serengeti. Vet Rec. (1966) 79:857–8. doi: 10.1136/vr.79.26.857
137. Cirović D, Chochlakis D, Tomanović S, Sukara R, Penezić A, Tselentis Y, et al. Presence of Leishmania and Brucella species in the golden jackal Canis aureus in Serbia. Biomed Res Int. (2014) 2014:728516. doi: 10.1155/2014/728516
138. Chitwood MC, Swingen MB, Lashley MA, Flowers JR, Palamar MB, Apperson CS, et al. Parasitology and serology of free-ranging coyotes (Canis latrans) in North Carolina, USA. J Wildl Dis. (2015) 51:664–9. doi: 10.7589/2015-01-002
139. Bischof R, Rogers DG. Serologic survey of select infectious diseases in coyotes and raccoons in Nebraska. J Wildl Dis. (2005) 41:787–91. doi: 10.7589/0090-3558-41.4.787
140. Gese EM, Schultz RD, Johnson MR, Williams ES, Crabtree RL, Ruff RL. Serological survey for diseases in free-ranging coyotes (Canis latrans) in Yellowstone National Park, Wyoming. J Wildl Dis. (1997) 33:47–56. doi: 10.7589/0090-3558-33.1.47
141. Holzman S, Conroy MJ, Davidson WR. Diseases, parasites and survival of coyotes in south-central Georgia. J Wildl Dis. (1992) 28:572–80. doi: 10.7589/0090-3558-28.4.572
142. Davis DS, Boeer WJ, Mims JP, Heck FC, Adams LG. Brucella abortus in coyotes. I A serologic and bacteriologic survey in eastern Texas. J Wildl Dis. (1979) 15:367–72. doi: 10.7589/0090-3558-15.3.367
143. Trainer DO, Knowlton FF. Serologic evidence of diseases in Texas coyotes. J Wildl Manage. (1968) 32:981–3. doi: 10.2307/3799581
144. Zarnke RL, Ver Hoef JM, DeLong RA. Geographic pattern of serum antibody prevalence for Brucella spp. in caribou, grizzly bears, and wolves from Alaska, 1975-1998. J Wildl Dis. (2006) 42:570–7. doi: 10.7589/0090-3558-42.3.570
145. Zarnke RL, Ballard WB. Serologic survey for selected microbial pathogens of wolves in Alaska, 1975-1982. J Wildl Dis. (1987) 23:77–85. doi: 10.7589/0090-3558-23.1.77
146. Tessaro SV. (1987). A Descriptive and Epizootiologic Study of Brucellosis and Tuberculosis in Bison in Northern Canada. Ph.D. thesis, Univ. Saskatchewan, Saskatoon.
147. Zarnke RL, Yuill TM. Serologic survey for selected microbial agents in mammals from Alberta, 1976. J Wildl Dis. (1981) 17:453–61. doi: 10.7589/0090-3558-17.3.453
148. Neiland KA. Further observations on rangiferine brucellosis in Alaskan carnivores. J Wildl Dis. (1975) 11:45–53. doi: 10.7589/0090-3558-11.1.45
149. Seles Dorneles EM, Pellegrin AO, Péres IAFS, Mathias LA, Mourão G, Bianchi RC, et al. Serology for brucellosis in free-ranging crab-eating foxes (Cerdocyon thous) and brown-nosed coatis (Nasua nasua) from Brazilian Pantanal. Ciência Rural. (2014) 44:2193–6. doi: 10.1590/0103-8478cr20131167
150. Proença LM, Silva JC, Galera PD, Lion MB, Marinho-Filho JS, Ragozo AM, et al. Serologic survey of infectious diseases in populations of maned wolf (Chrysocyon brachyurus) and crab-eating fox (Cerdocyon thous) from Aguas Emendadas Ecological Station, Brazil. J Zoo Wildl Med. (2013) 44:152–5. doi: 10.1638/1042-7260-44.1.152
151. Fiorello CV, Noss AJ, Deem SL, Maffei L, Dubovi EJ. Serosurvey of small carnivores in the Bolivian Chaco. J Wildl Dis. (2007) 43:551–7. doi: 10.7589/0090-3558-43.3.551
152. Standley WG, McCue PM. Prevalence of antibodies against selected diseases in San Joaquin kit foxes at Camp Roberts, California. Calif Fish Game (1997) 83:30–7.
153. Parnas J, Zalichta S, Sidor-Wójtowicz A. The taxonomic properties of Brucella from foxes (Chifres-Argentina), isolated in natural foci. Zentralblatt Veterinarmedizin Reihe B. (1969) 16:183–92. doi: 10.1111/j.1439-0450.1969.tb00103.x
154. Szyfres B, González Tomé J. Natural Brucella infection in Argentine wild foxes. Bull World Health Organ. (1966) 34:919–23.
155. Azevedo SSD, Silva MLCR, Batista CDSA, Gomes AADB, Vasconcellos SA, Alves CJ. Detection of anti Brucella abortus, anti Brucella canis and anti Leptospira spp. antibodies in hoary foxes (Pseudalopex vetulus) from semi-arid of Paraiba state, Northeastern region of Brazil. Ciência Rural (2010) 40:190–2. doi: 10.1590/S0103-84782009005000232
156. Corey RR, Paulissen LJ, Schwartz D. Prevalence of brucellae in the wildlife of Arkansas. J. Wildl. Dis. (1964) 39:WD-63–2.
157. Pinigin AF, Zabrodin VA, Nikulina VI. [Brucellosis in arctic foxes, Alopex lagopus]. Krolikovodstvo i zverovodstvo. (1970) 5:39–40.
158. McCue PM, O'Farrell TP. Serological survey for selected diseases in the endangered San Joaquin kit fox (Vulpes macrotis mutica). J Wildl Dis. (1988) 24:274–81. doi: 10.7589/0090-3558-24.2.274
159. Scholz HC, Revilla-Fernández S, Al Dahouk S, Hammerl JA, Zygmunt MS, Cloeckaert A, et al. Brucella vulpis sp. nov, isolated from mandibular lymph nodes of red foxes (Vulpes vulpes). Int J Syst Evol Microbiol. (2016) 66:2090–8. doi: 10.1099/ijsem.0.000998
160. Hofer E, Revilla-Fernández S, Al Dahouk S, Riehm JM, Nöckler K, Zygmunt MS, et al. A potential novel Brucella species isolated from mandibular lymph nodes of red foxes in Austria. Vet Microbiol. (2012) 155:93–9. doi: 10.1016/j.vetmic.2011.08.009
161. Scholz HC, Hofer E, Vergnaud G, Le Fleche P, Whatmore AM, Al Dahouk S, et al. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector Borne Zoonotic Dis. (2009) 9:153–6. doi: 10.1089/vbz.2008.0036
162. Davies G, Ockey JH, Lloyd HG. Isolation of Brucella abortus from a fox (Vulpes vulpes). State Vet. J. (1973) 28:250–2.
163. McCaughey WJ, Fairley JS. Serological reactions to Brucella and Leptospira in foxes. Vet. Rec. (1969) 84:21.
164. Pavlov P, Tehiley D, Matev M, Milanov M, Tatarov B, Krastev V. Recherches sur des reservoirs de Brucella chez le porc vivant en liberte. Bull. Off. Int. Epizoot. (1960) 53:1511–26.
165. White CL, Schuler KL, Thomas NJ, Webb JL, Saliki JT, Ip HS, et al. Pathogen exposure and blood chemistry in the Washington, USA population of northern sea otters (Enhydra lutris kenyoni). J Wildl Dis. (2013) 49:887–99. doi: 10.7589/2013-03-053
166. Brancato MS, Milonas L, Bowlby CE, Jameson R, Davis JW. Chemical Contaminants, Pathogen Exposure and General Health Status of Live and Beach-Cast Washington Sea Otters (Enhydra lutris kenyoni). Marine Sancturaries Conservation Series ONMS 08-08. US Department of Commerce, National Oceanic and Atmospheric Administration, Office of National Marine Sancturaies, Silver Springs, MD, (2009). 181 p.
167. Hanni KD, Mazet JA, Gulland FM, Estes J, Staedler M, Murray MJ, et al. Clinical pathology and assessment of pathogen exposure in southern and Alaskan sea otters. J Wildl Dis. (2003) 39:837–50. doi: 10.7589/0090-3558-39.4.837
168. Burgess TL, Johnson CK, Burdin A, Gill VA, Doroff AM, Tuomi P, et al. Brucella infection in Asian sea otters (Enhydra lutris lutris) on Bering Island, Russia. J Wildl Dis. (2017) 53:864–8. doi: 10.7589/2016-09-220
169. Miller MA, Burgess TL, Dodd EM, Rhyan JC, Jang SS, Byrne BA, et al. Isolation and characterization of marine Brucella from southern sea otter (Enhydra lutris lutris), California, USA. J Wildl Dis. (2017) 53:215–27. doi: 10.7589/2015-12-326
170. Patterson IAP, Howie FE, Reid RJ, Ross HM, MacMillan A, Foster G, et al. Brucella infections in marine mammals from Scottish waters. In: International Association for Aquatic Animal Medicine: archival CD for Proceedings (New Orleans, LA) (2000). p. 396–398.
171. Foster G, Jahans KL, Reid RJ, Ross HM. Isolation of Brucella species from cetaceans, seals and an otter. Vet Rec. (1996) 138:583–6. doi: 10.1136/vr.138.24.583
172. Truong LQ, Kim JT, Yoon BI, Her M, Jung SC, Hahn TW. Epidemiological survey for Brucella in wildlife and stray dogs, a cat and rodents captured on farms. J Vet Med Sci. (2011) 73:1597–601. doi: 10.1292/jvms.11-0222
173. Verger JM, Dufrenoy J. Isolement de Brucella abortus biotype 1 chez le mulot (Apodemus sylvaticus). Ouest Med. (1972) 25:989–91.
174. Martino PE, Samartino LE, Stanchi NO, Radman NE, Parrado EJ. Serology and protein electrophoresis for evidence of exposure to 12 mink pathogens in free-ranging American mink (Neovison vison) in Argentina. Vet Q. (2017) 37:207–11. doi: 10.1080/01652176.2017.1336810
175. Stoenner HG, Holdenried R, Lackman D, Orsborn JS Jr. The occurrence of Coxiella burnetii, Brucella, and other pathogens among fauna of the Great Salt Lake desert in Utah. Am J Trop Med Hyg. (1959) 8:590–6. doi: 10.4269/ajtmh.1959.8.590
176. Boeer WJ, Crawford RP, Hidalgo RJ, Robinson RM. Small mammals and white-tailed deer as possible reservoir hosts of Brucella abortus in Texas. J Wildl Dis. (1980) 16:19–24. doi: 10.7589/0090-3558-16.1.19
177. Swann AI, Schnurrenberger PR, Brown RR, Garby CL. Brucella abortus isolations from wild animals. Vet Rec. (1980) 106:57. doi: 10.1136/vr.106.3.57-a
178. Bronson E, Spiker H, Driscoll CP. Serosurvey for selected pathogens in free-ranging American black bears (Ursus americanus) in Maryland, USA. J Wildl Dis. (2014) 50:829–36. doi: 10.7589/2013-07-155
179. O'Hara TM, Holcomb D, Elzer P, Estepp J, Perry Q, Hagius S, et al. Brucella species survey in polar bears (Ursus maritimus) of northern Alaska. J Wildl Dis. (2010) 46:687–94. doi: 10.7589/0090-3558-46.3.687
180. Binninger CE, Beecham JJ, Thomas LA, Winward LD. A serologic survey for selected infectious diseases of black bears in Idaho. J Wildl Dis. (1980) 16:423–30. doi: 10.7589/0090-3558-16.3.423
181. Zarnke RL. Serologic survey for selected microbial pathogens in Alaskan wildlife. J Wildl Dis. (1983) 19:324–9. doi: 10.7589/0090-3558-19.4.324
182. Di Francesco CE, Gentile L, Di Pirro V, Ladiana L, Tagliabue S, Marsilio F. Serologic evidence for selected infectious diseases in Marsican brown bears (Ursus arctos marsicanus) in Italy (2004-09). J Wildl Dis. (2015) 51:209–13. doi: 10.7589/2014-01-021
183. Atwood TC, Duncan C, Patyk KA, Nol P, Rhyan J, McCollum M, et al. Environmental and behavioral changes may influence the exposure of an Arctic apex predator to pathogens and contaminants. Sci Rep. (2017) 7:13193. doi: 10.1038/s41598-017-13496-9
184. Rah H, Chomel BB, Follmann EH, Kasten RW, Hew CH, Farver TB, et al. Serosurvey of selected zoonotic agents in polar bears (Ursus maritimus). Vet Rec. (2005) 156:7–13. doi: 10.1136/vr.156.1.7
185. Tryland M, Derocher AE, Wiig Y, Godfroid J. Brucella sp. antibodies in polar bears from Svalbard and the Barents Sea. J Wildl Dis. (2001) 37:523–31. doi: 10.7589/0090-3558-37.3.523
186. Calle PP, Seagars DJ, McClave C, Senne D., House C, House JA. Viral and bacterial serology of free-ranging Pacific walrus. J Wildlife Dis. (2002) 38:93–100. doi: 10.7589/0090-3558-38.1.93
187. Nielsen O, Stewart RE, Nielsen K, Measures L, Duignan P. Serologic survey of Brucella spp. antibodies in some marine mammals of North America. J Wildl Dis. (2001) 37:89–100. doi: 10.7589/0090-3558-37.1.89
188. Nielsen O, Nielsen K, Stewart REA. Serologic evidence of Brucella spp. exposure in Atlantic walruses (Odobenus rosmarus rosmarus) and ringed seals (Phoca hispida) of Arctic. Canada Arctic (1996) 49:383–6. doi: 10.14430/arctic1214
189. Jankowski G, Adkesson MJ, Saliki JT, Cárdenas-Alayza S, Majluf P. Survey for infectious disease in the South American fur seal (Arctocephalus australis) population at punta San Juan, Peru. J Zoo Wildl Med. (2015) 46:246–54. doi: 10.1638/2014-0120.1
190. Mackereth GF, Webb KM, O'Keefe JS, Duignan PJ, Kittelberger R. Serological survey of pre-weaned New Zealand fur seals (Arctocephalus forsteri) for brucellosis and leptospirosis. N Z Vet J. (2005) 53:428–32. doi: 10.1080/00480169.2005.36588
191. Jensen SK, Nymo IH, Forcada J, Hall A, Godfroid J. Brucella antibody seroprevalence in Antarctic seals (Arctocephalus gazella, Leptonychotes weddellii and Mirounga leonina). Dis Aquat Org. (2013) 105:175–81. doi: 10.3354/dao02633
192. Tryland M, Nymo IH, Nielsen O, Nordøy ES, Kovacs KM, Krafft BA, et al. Serum chemistry and antibodies against pathogens in Antarctic fur seals, Weddell seals, crabeater seals, and Ross seals. J Wildl Dis. (2012) 48:632–45. doi: 10.7589/0090-3558-48.3.632
193. Abalos Pineda P, Blank Hidber O, Torres Navarro D, Torres Castillo D, Valdenegro Vega V, Retamal Merino P. Brucella infection in marine mammals in Antarctica. Vet Rec. (2009) 164:250. doi: 10.1136/vr.164.8.250
194. Blank O, Retamal P, Abalos P, Torres D. Additional data on anti-Brucella antibodies in Arctocephalus gazella from Cape Shirreff, Livingston Island, Antarctica. CCAMLR Sci. (2001) 8:147–54.
195. Retamal P, Blank O, Abalos P, Torres D. Detection of anti-Brucella antibodies in pinnipeds from the Antarctic territory. Vet Rec. (2000) 146:166–7. doi: 10.1136/vr.146.6.166
196. Lynch M, Duignan PJ, Taylor T, Nielsen O, Kirkwood R, Gibbens J, et al. Epizootiology of Brucella infection in Australian fur seals. J Wildl Dis. (2011a) 47:352–63. doi: 10.7589/0090-3558-47.2.352
198. Ziehl-Quirós EC, García-Aguilar MC, Mellink E. Colony-level assessment of Brucella and Leptospira in the Guadalupe fur seal, Isla Guadalupe, Mexico. Dis Aquat Org. (2017) 122:185–93. doi: 10.3354/dao03073
199. Nymo IH, Rødven R, Beckmen K, Larsen AK, Tryland M, Quakenbush L, et al. Brucella antibodies in Alaskan True seals and eared seals—Two different stories. Front Vet Sci. (2018) 5:8. doi: 10.3389/fvets.2018.00008
200. Duncan CG, Tiller R, Mathis D, Stoddard R, Kersh GJ, Dickerson B, et al. Brucella placentitis and seroprevalence in northern fur seals (Callorhinus ursinus) of the Pribilof Islands, Alaska. J Vet Diagn Invest. (2014) 26:507–12. doi: 10.1177/1040638714532647
201. Burek KA, Gulland FM, Sheffield G, Beckmen KB, Keyes E, Spraker TR, et al. Infectious disease and the decline of Steller sea lions (Eumetopias jubatus) in Alaska, USA: insights from serologic data. J Wildl Dis. (2005) 41:512–24. doi: 10.7589/0090-3558-41.3.512
202. Abe E, Ohishi K, Ishinazaka T, Fujii K, Maruyama T. Serologic evidence of Brucella infection in pinnipeds along the coast of Hokkaido, the northernmost main island of Japan. Microbiol. Immunol. (2017) 61:114–22. doi: 10.1111/1348-0421.12474
203. Roe WD, Rogers LE, Gartrell BD, Chilvers BL, Duignan PJ. Serologic evaluation of New Zealand sea lions for exposure to Brucella and Leptospira spp. J Wildl Dis. (2010) 46:1295–9. doi: 10.7589/0090-3558-46.4.1295
204. Goldstein T, Zabka TS, Delong RL, Wheeler EA, Ylitalo G, Bargu S, et al. The role of domoic acid in abortion and premature parturition of California sea lions (Zalophus californianus) on San Miguel Island, California. J Wildl Dis. (2009) 45:91–108. doi: 10.7589/0090-3558-45.1.91
205. Sonne C, Andersen-Ranberg E, Rajala EL, Agerholm JS, Bonefeld-Jørgensen E, Desforges JP, et al. Seroprevalence for Brucella spp. in Baltic ringed seals (Phoca hispida) and East Greenland harp (Phagophilus groenlandicus) and hooded (Cystophora cristata) seals. Vet Immunol Immunopathol. (2018) 198:14–8. doi: 10.1016/j.vetimm.2018.02.005
206. Nymo IH, Tryland M, Frie AK, Haug T, Foster G, Rødven R, et al. Age-dependent prevalence of anti-Brucella antibodies in hooded seals Cystophora cristata. Dis Aquat Org. (2013) 106:187–96. doi: 10.3354/dao02659
207. Tryland M, Sørensen KK, Godfroid J. Prevalence of Brucella pinnipediae in healthy hooded seals (Cystophora cristata) from the North Atlantic Ocean and ringed seals (Phoca hispida) from Svalbard. Vet Microbiol. (2005) 105:103–11. doi: 10.1016/j.vetmic.2004.11.001
208. Foster G, MacMillan AP, Godfroid J, Howie F, Ross HM, Cloeckaert A, et al. A review of Brucella sp. infection of sea mammals with particular emphasis on isolates from Scotland. Vet Microbiol. (2002) 90:563–80. doi: 10.1016/S0378-1135(02)00236-5
209. Tryland M, Kleivane L, Alfredsson A, Kjeld M, Arnason A, Stuen S, et al. Evidence of Brucella infection in marine mammals in the North Atlantic Ocean. Vet Rec. (1999) 144:588–92. doi: 10.1136/vr.144.21.588
210. Foster G, Nymo IH, Kovacs KM, Beckmen KB, Brownlow AC, Baily JL, et al. First isolation of Brucella pinnipedialis and detection of Brucella antibodies from bearded seals Erignathus barbatus. Dis Aquat Org. (2018) 128:13–20. doi: 10.3354/dao03211
211. Calle PP, Seagars DJ, McClave C, Senne D, House C, House JA. Viral and bacterial serology of six free-ranging bearded seals Erignathus barbatus. Dis Aquat Organ. (2008) 81:77–80. doi: 10.3354/dao01927
212. Kroese MV, Beckers L, Bisselink YJWM, Brasseur S, van Tulden PW, Koene MGJ, et al. Brucella pinnipedialis in grey seals (Halichoerus grypus) and harbor seals (Phoca vitulina) in the Netherlands. J. Wildl. Dis. (2018) 54:439–49. doi: 10.7589/2017-05-097
213. Hirvelä-Koski V, Nylund M, Skrzypczak T, Heikkinen P, Kauhala K, Jay M, et al. Isolation of Brucella pinnipedialis from grey seals (Halichoerus grypus) in the Baltic Sea. J Wildl Dis. (2017) 53:850–3. doi: 10.7589/2016-06-144
214. Prenger-Berninghoff E, Siebert U, Stede M, König A, Weiss R, Baljer G. Incidence of Brucella species in marine mammals of the German North Sea. Dis Aquat Org. (2008) 81:65–71. doi: 10.3354/dao01920
215. Jepson PD, Brew S, MacMillan AP, Baker JR, Barnett J, Kirkwood JK, et al. Antibodies to Brucella in marine mammals around the coast of England and Wales. Vet Rec. (1997) 141:513–5. doi: 10.1136/vr.141.20.513
216. Ross HM, Jahans KL, MacMillan AP, Reid RJ, Thompson PM, Foster G. Brucella species infection in North Sea seal and cetacean populations. Vet Rec. (1996) 138:647–8. doi: 10.1136/vr.138.26.647
217. McFarlane RA. Health assessment and diseases of the Weddell seal, Leptonochotes weddelli, in Vestfold Hills, East Antarctica. In: Kerry KR, and Riddle MJ, editors. Health of Antarctic Wildlife. Berlin: Springer-Verlag (2009). p. 139–66.
218. Yochem PK, Stewart BS, Gelatt TS, Siniff DB. Health assessment of Weddell seals, Leptonychotes weddellii, in McMurdo Sound, Antarctica. In: Kerry KR, and Riddle MJ, editors. Health of Antarctic Wildlife. Berlin: Springer-Verlag (2009) 123–38.
219. Blank O, Retamal P, Abalos P, Torres D. Detection of anti-Brucella antibodies in Weddell seals (Leptonychotes weddellii) from Cape Shirreff, Antarctica. Arch Med Vet. (2002) 34:117–22. doi: 10.4067/S0301-732X2002000100013
220. Barbieri M, Duncan C, Harting AL, Pabilonia KL, Johanos TC, Goldstein T, et al. Survey for placental disease and reproductive pathogens in the endangered Hawaiian monk seal (Neomonachus schauinslandi). J. Wildl. Dis. (2018) 54:564–8. doi: 10.7589/2017-07-164
221. Aguirre AA, Keefe TJ, Reif JS, Kashinsky L, Yochem PK, Saliki JT, et al. Infectious disease monitoring of the endangered Hawaiian monk seal. J Wildl Dis. (2007) 43:229–41. doi: 10.7589/0090-3558-43.2.229
222. Nielsen O, Nielsen K, Braun R, Kelly L. A comparison of four serologic assays in screening for Brucella exposure in Hawaiian monk seals. J Wildl Dis. (2005) 41:126–33. doi: 10.7589/0090-3558-41.1.126
223. Maratea J, Ewalt DR, Frasca S Jr, Dunn JL, De Guise S, Szkudlarek L, et al. Evidence of Brucella sp. infection in marine mammals stranded along the coast of southern New England. J Zoo Wildl Med. (2003) 34:256–61. doi: 10.1638/02-053
224. Forbes LB, Nielsen O, Measures L, Ewalt DR. Brucellosis in ringed seals and harp seals from Canada. J Wildl Dis. (2000) 36:595–8. doi: 10.7589/0090-3558-36.3.595
225. Siebert U, Rademaker M, Ulrich SA, Wohlsein P, Ronnenberg K, Prenger-Berninghoff E. Bacterial microbiota in harbor seals (Phoca vitulina) from the North Sea of Schleswig-Holstein, Germany, around the time of morbillivirus and influenza epidemics. J Wildl Dis. (2017) 53:201–14. doi: 10.7589/2015-11-320
226. Kershaw JL, Stubberfield EJ, Foster G, Brownlow A, Hall AJ, Perrett LL. Exposure of harbour seals Phoca vitulina to Brucella in declining populations across Scotland. Dis Aquat Org. (2017) 126:13–23. doi: 10.3354/dao03163
227. Hoover-Miller A, Dunn JL, Field CL, Blundell G, Atkinson S. Seroprevalence of Brucella antibodies in harbor seals in Alaska, USA, with age, regional, and reproductive comparisons. Dis. Aquat. Org. (2017) 126:1–12. doi: 10.3354/dao03153
228. Hueffer K, Gende SM, O'Hara TM. Assay dependence of Brucella antibody prevalence in a declining Alaskan harbor seal (Phoca vitulina) population. Acta Vet Scand. (2013) 55:52. doi: 10.1186/1751-0147-55-2
229. Ewalt D, Lambourn D, Sidor I, Gaydos J, Garner M, Lockwood S, et al. Endemic marine Brucella sp. infection in harbor seals: Is there human risk? In: Proc. 45th Annual Meeting of the Infectious Diseases Society of America, 4–7 October. San Diego, CA (2007).
230. Ross HM, Foster G, Reid RJ, Jahans KL, MacMillan AP. Brucella species infection in sea-mammals. Vet Rec. (1994) 134:359. doi: 10.1136/vr.134.14.359-b
231. Lambourn DM, Garner M, Ewalt D, Raverty S, Sidor I, Jeffries SJ, et al. Brucella pinnipedialis infections in Pacific harbor seals (Phoca vitulina richardsi) from Washington State, USA. J Wildl Dis. (2013) 49:802–15. doi: 10.7589/2012-05-137
232. Zarnke RL, Saliki JT, Macmillan AP, Brew SD, Dawson CE, Ver Hoef JM, et al. Serologic survey for Brucella spp., phocid herpesvirus-1, phocid herpesvirus-2, and phocine distemper virus in harbor seals from Alaska, 1976-1999. J. Wildl. Dis. (2006) 42:290–300. doi: 10.7589/0090-3558-42.2.290
233. Garner MM, Lambourn DM, Jeffries SJ, Hall PB, Rhyan JC, Ewalt DR, et al. Evidence of Brucella infection in Parafilaroides lungworms in a Pacific harbor seal (Phoca vitulina richardsi). J Vet Diagn Invest. (1997) 9:298–303. doi: 10.1177/104063879700900311
234. Lappin MR, Hawley J. Presence of Bartonella species and Rickettsia species DNA in the blood, oral cavity, skin and claw beds of cats in the United States. Vet Dermatol. (2009) 20:509–14. doi: 10.1111/j.1365-3164.2009.00800.x
235. Filoni C, Catão-Dias JL, Cattori V, Willi B, Meli ML, Corrêa SH, et al. Surveillance using serological and molecular methods for the detection of infectious agents in captive Brazilian neotropic and exotic felids. J Vet Diagn Invest. (2012) 24:166–73. doi: 10.1177/1040638711407684
236. Guimaraes AM, Brandão PE, Moraes W, Kiihl S, Santos LC, Filoni C, et al. Detection of Bartonella spp. in neotropical felids and evaluation of risk factors and hematological abnormalities associated with infection. Vet Microbiol. (2010) 142:346–51. doi: 10.1016/j.vetmic.2009.10.002
237. Miyazaki S, Ishii T, Matoba S, Awatani T, Toda I. A case of cat-scratch disease from a masked palm civet in Japan. Monthly Community Med. (2001) 15:564–6.
238. Gherman CM, Mihalca AD. A synoptic overview of golden jackal parasites reveals high diversity of species. Parasit Vectors (2017) 10:419. doi: 10.1186/s13071-017-2329-8
239. Wozencraft WC. Order Carnivora. In: Wilson DE, and Reeder, DM. Mammal Species of the World: A Taxonomic and Geographic Reference. Baltimore, MD: Johns Hopkins University Press (2005) 532–628.
240. Allegri L, Socci A, Di Natale M. (1969). [Isolation of Brucella abortus from the organs of mink]. G Mal Infett Parassit. 21:708–10.
241. Gage KL. (1999). Plague surveillance. In: Plague Manual: Epidemiology, Distribution, Surveillance and Control. on World Health Organization. Available online at: http://www.who.int/csr/resources/publications/plague/WHO_CDS_CSR_EDC_99_2_EN/en/
242. Bai Y, Urushadze L, Osikowicz L, McKee C, Kuzmin I, Kandaurov A, et al. Molecular survey of bacterial zoonotic agents in bats from the country of Georgia (Caucasus). PLoS ONE (2017) 12:e0171175. doi: 10.1371/journal.pone.0171175
243. Harms CA, Maggi RG, Breitschwerdt EB, Clemons-Chevis CL, Solangi M, Rotstein DS, et al. Bartonella species detection in captive, stranded and free-ranging cetaceans. Vet Res. (2008) 39:59. doi: 10.1051/vetres:2008036
244. Maggi RG, Harms CA, Hohn AA, Pabst DA, McLellan WA, Walton WJ, et al. Bartonella henselae in porpoise blood. Emerging Infect Dis. (2005) 11:1894–8. doi: 10.3201/eid1112.050969
245. Kosoy M, Hayman DT, Chan KS. Bartonella bacteria in nature: where does population variability end and a species start? Infect. Genet. Evol. (2012) 12:894–904. doi: 10.1016/j.meegid.2012.03.005
246. Kumar S, Stecher G, Suleski M, Hedges SB. TimeTree: a resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. (2017) 34:12–1819. doi: 10.1093/molbev/msx116
247. Marin J, Battistuzzi FU, Brown AC, Hedges SB. The timetree of prokaryotes: new insights into their evolution and speciation. Mol. Biol. Evol. (2016) 34:437–46. doi: 10.1093/molbev/msw245
248. Segers FH, Kešnerov áL, Kosoy M, Engel P. Genomic changes associated with the evolutionary transition of an insect gut symbiont into a blood-borne pathogen. ISME J. (2017) 11:1232. doi: 10.1038/ismej.2016.201
249. Kešnerová L, Moritz R, Engel P. Bartonella apis sp. nov, a honey bee gut symbiont of the class Alphaproteobacteria. Int J Syst Evol Microbiol. (2016) 66:414–21. doi: 10.1099/ijsem.0.000736
250. Russell JA, Moreau CS, Goldman-Huertas B, Fujiwara M, Lohman DJ, Pierce NE. Bacterial gut symbionts are tightly linked with the evolution of herbivory in ants. Proc Natl Acad Sci USA. (2009) 106:21236–41. doi: 10.1073/pnas.0907926106
251. Bisch G, Neuvonen MM, Pierce NE, Russell JA, Koga R, Sanders JG, et al. Genome evolution of Bartonellaceae symbionts of ants at the opposite ends of the trophic scale. Genome Biol. Evol. (2018) 10:1687–704. doi: 10.1093/gbe/evy126
252. Neuvonen MM, Tamarit D, Näslund K, Liebig J, Feldhaar H, Moran NA, et al. The genome of Rhizobiales bacteria in predatory ants reveals urease gene functions but no genes for nitrogen fixation. Sci Rep. (2016) 6:39197. doi: 10.1038/srep39197
253. Paulsen IT, Seshadri R, Nelson KE, Eisen JA, Heidelberg JF, Read TD, et al. The Brucella suis genome reveals fundamental similarities between animal and plant pathogens and symbionts. Proc Natl Acad Sci USA. (2002) 99:13148–53. doi: 10.1073/pnas.192319099
254. Moreno E, Cloeckaert A, Moriyón I. Brucella evolution and taxonomy. Vet Microbiol. (2002) 90:209–27. doi: 10.1016/S0378-1135(02)00210-9
255. Foster JT, Beckstrom-Sternberg SM, Pearson T, Beckstrom-Sternberg JS, Chain PS, Roberto FF, et al. Whole-genome-based phylogeny and divergence of the genus Brucella. J Bacteriol. (2009) 191:2864–70. doi: 10.1128/JB.01581-08
256. D'Anastasio R, Staniscia T, Milia ML, Manzoli L, Capasso L. Origin, evolution and paleoepidemiology of brucellosis. Epidemiol Infect. (2011) 139:149–56. doi: 10.1017/S095026881000097X
257. D'Anastasio R, Zipfel B, Moggi-Cecchi J, Stanyon R, Capasso L. Possible brucellosis in an early hominin skeleton from Sterkfontein, South Africa. PLoS ONE (2009) 4:e6439. doi: 10.1371/journal.pone.0006439
Keywords: Bartonella, Brucella, carnivores, disease ecology, wildlife disease
Citation: Kosoy M and Goodrich I (2019) Comparative Ecology of Bartonella and Brucella Infections in Wild Carnivores. Front. Vet. Sci. 5:322. doi: 10.3389/fvets.2018.00322
Received: 23 October 2018; Accepted: 03 December 2018;
Published: 04 January 2019.
Edited by:
David Modrý, University of Veterinary and Pharmaceutical Sciences Brno, CzechiaReviewed by:
Shimon Harrus, Hebrew University of Jerusalem, IsraelAndrei Daniel Mihalca, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, Romania
Copyright © 2019 Kosoy and Goodrich. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Kosoy, bWNrM0BjZGMuZ292