 Israel Barbosa Guedes1
Israel Barbosa Guedes1 Gisele Oliveira de Souza1Juliana Fernandes de Paula Castro1
Gisele Oliveira de Souza1Juliana Fernandes de Paula Castro1 Antônio Francisco de Souza Filho1Matheus Burilli Cavalini1Sueli Akemi Taniwaki1Anderson Luiz Pinheiro Maia2Isaías Corrêa Pereira3Marcos Bryan Heinemann1*
Antônio Francisco de Souza Filho1Matheus Burilli Cavalini1Sueli Akemi Taniwaki1Anderson Luiz Pinheiro Maia2Isaías Corrêa Pereira3Marcos Bryan Heinemann1*- 1Departamento de Medicina Veterinária Preventiva e Saúde Animal, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, Brazil
- 2Médico Veterinário, Auditor Fiscal Agropecuário, Agência de Defesa e Inspeção Agropecuária Do Estado Do Amapá, Santana, Brazil
- 3Secretaria de Estado da Saúde Do Amapá, Macapá, Brazil
In the current context of deforestation and fire in the Amazon, buffaloes could be a cost-effective and sustainable alternative for cattle production in the region, as they can convert low-quality foods and be raised in floodplain areas. However, little is known about the reproductive diseases that affect these animals; thus, the purpose of this study was to perform the molecular characterization of Leptospira spp. in the urogenital tract of water buffaloes (Bubalus bubalis) raised in the Amazon River Delta region in Brazil. Samples were collected from 114 kidneys, 204 ovaries, and 160 uterine swabs of slaughtered buffaloes in the Macapá microregion of Amapá State (Brazil) and were subjected to PCR to detect bacterial DNA. Positive amplicons were sequenced to identify Leptospira species. Among the total samples, 11/473 were PCR positive (2.3%), including 10 kidney samples and one uterine swab sample. DNA sequencing identified two pathogenic species from the kidney samples: L. interrogans, accounting for 60.0% (6/10) of these samples, and L. borgpetersenii, accounting for 20.0% (2/10), while 20.0% (2/10) were identified only at the genus level. The bacterium in the uterine swab sample was identified as L. interrogans with genetic proximity to strains belonging to the serovar Hardjo. This is the first report of leptospires species identified in buffaloes from the Amazon River Delta region and revealed that these animals may be carriers of different pathogenic Leptospira species, similar to bovines, including showing genital colonization.
Introduction
Leptospirosis is a bacterial disease that affects humans and several species of domestic and wild animals and is considered a zoonosis (1). The genus Leptospira can be considered to be dynamic and diversified in relation to the species it includes; with the advancement of molecular methodologies, it has been possible to define 64 species, which are now divided into two major clades, one of which contains pathogenic species, while the other contains saprophytes, leading to a new proposal for the systematic classification of the genus (2). Nevertheless, the serological classification of leptospires is still accepted and is the basis of serological assays such as the microscopic agglutination test (MAT), in which a limited number of serovars that represent prevalent serogroups for a specific region are used (1, 3).
Similar to the disease in cattle, buffalo leptospirosis is mainly characterized by reproductive disorders, especially abortions, in which Leptospira spp. have been detected (4, 5). In Brazil, there has been only one report of Leptospira isolation from the urine of a healthy buffalo from the southeastern region of the country, and the isolate was classified as L. santarosai serovar Guaricura (6). Due to the difficulties in isolating leptospires from biological samples, direct DNA sequencing from PCR products is used, which allows the identification of leptospires at the species level, providing a new epidemiological analysis of the disease (7–9).
In 2018, the Brazilian buffalo herd consisted of 1,390,066 buffaloes, and ~37% of this herd was concentrated in the Amazon River Delta region (considering the east coast of Amapá state and Marajó Island) (10). Buffaloes are part of the Amazonian culture since it is used as a work animal and it plays a role in tourism, so these animals are in close proximity to humans (11). This close contact may represent a public health problem since leptospirosis is a zoonosis, and it has been reported that buffaloes can be directly involved in the transmission of leptospirosis to humans (12). In this region, serological studies have revealed the presence of anti-Leptospira spp. antibodies in buffaloes, with a prevalence ranging from 34.37 to 80.0% and predominance of the Sejroe serogroup (13–15), similar to what is observed in cattle. Nevertheless, there have been few studies on leptospirosis in buffaloes; thus, the aim of this study was to detect and perform the molecular characterization of Leptospira spp. in the urogenital tract of buffaloes raised in the Amazon River Delta region, providing new knowledge about leptospirosis in these animals.
Materials and Methods
This work was approved by the Ethics Committee on Animal Use of the School of Veterinary Medicine and Animal Science (Universidade de São Paulo)—CEUA/FMVZ n° 5613211118. For this study, 114 kidney fragments (~5 g), 204 ovaries and 160 uterine swabs were collected from buffaloes slaughtered in a slaughterhouse in the Macapá microregion of Amapá State, Brazil. Sampling was carried out by convenience, and for logistical reasons during slaughter, the samples were collected in groups on different days according to the type of tissue (kidneys—day 1, ovaries—day 2, uterine swabs—day 3); thus, each sample represented a single animal, totaling 478 animals.
The animals came from different farms located in the Amazon River Delta region in northern Brazil, which is characterized as a geographical area formed by hundreds of islands and islets between the states of Pará and Amapá, encompassing Marajó Island (Figure 1). The animals predominantly belonged to the River Buffalo group breeds (Murrah, Mediterranean, and Jafarabadi) and included males and females that were at least 12 months old, showed meat production capacity, had not been vaccinated against leptospirosis and had an unknown reproductive history.
Figure 1. Amazon River Delta region in Brazil (circle).
During the slaughter of the animals, fragments of the kidneys, and ovaries were collected with the aid of sterile forceps and scissors and stored individually in sterile plastic bags for homogenization. At the slaughterhouse immediately after collection, the organs were macerated and diluted 1:10 in phosphate-buffered saline [PBS; 0.137 M NaCl; 0.0027 M KCl; 0.01 M Na2HPO4; and 0.0018 M KH2PO4 (pH 7.4)]. The uterine swab samples were obtained with the aid of a disposable cervical brush fixed to a swab, which was vigorously rubbed against the mucosa of the uterine horns and body, and were also diluted in phosphate-buffered saline. A 1 ml aliquot of the diluted samples was frozen and sent to the laboratory for tests.
The extraction and purification of DNA from the samples were performed using the PureLink® Genomic DNA Mini Kit (Invitrogen™) according to the manufacturer's protocol. PCR targeting the Bubalus bubalis cytochrome b (cytb) gene, as described by Bottero et al. (16), was used as an internal control to verify the validity of the extracted DNA. The detection of Leptospira spp. was carried out by PCR with the Lep1 and Lep2 primer pair, which amplifies a 330 bp region of the 16S rRNA gene (rrs) (17), using Go Taq™ Green Master Mix (Promega, Brazil). The positive samples were subjected to another round of PCR for typing using primer pairs that amplify a 549 bp region of the secY gene (18). L. interrogans serovar Hardjo-prajitno and ultrapure water were used as positive and negative controls, respectively. A negative control sample was inserted between every five test samples to assess the presence of contamination in the extracted DNA.
Positive amplicons were sequenced by the Sanger method with BigDye Terminator v.3.1 chemistry (Applied Biosystems) and an ABI-3500 (Applied Biosystems) automatic sequencer according to the manufacturer's instructions. The sequences were assembled with the BioEdit Sequence Alignment Editor (19). The phylogenetic trees were built using homologous sequences retrieved from the GenBank database (accession numbers in Figures 2, 3) with the neighbor-joining method, the Tamura-3-parameter model and 1,000 bootstrap replicates in MEGA 7 software (20).
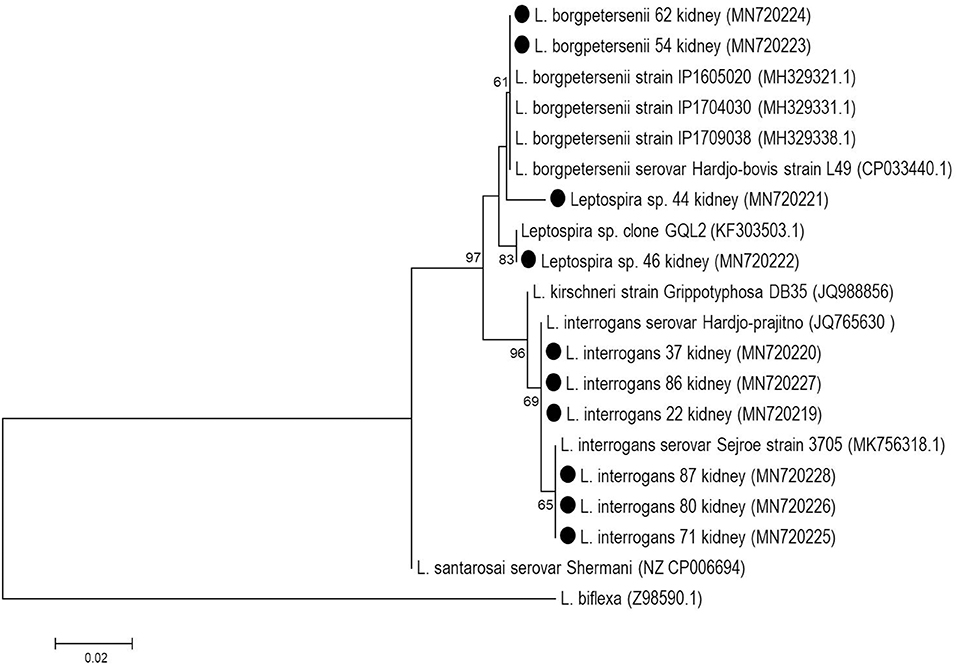
Figure 2. Phylogenetic reconstruction based on partial rrs gene of Leptospira spp. from kidney samples of buffaloes (black circles). The tree was elaborated with Neighbor-Joining method, Tamura-3 parameter model with bootstrap test of 1,000 replicates.
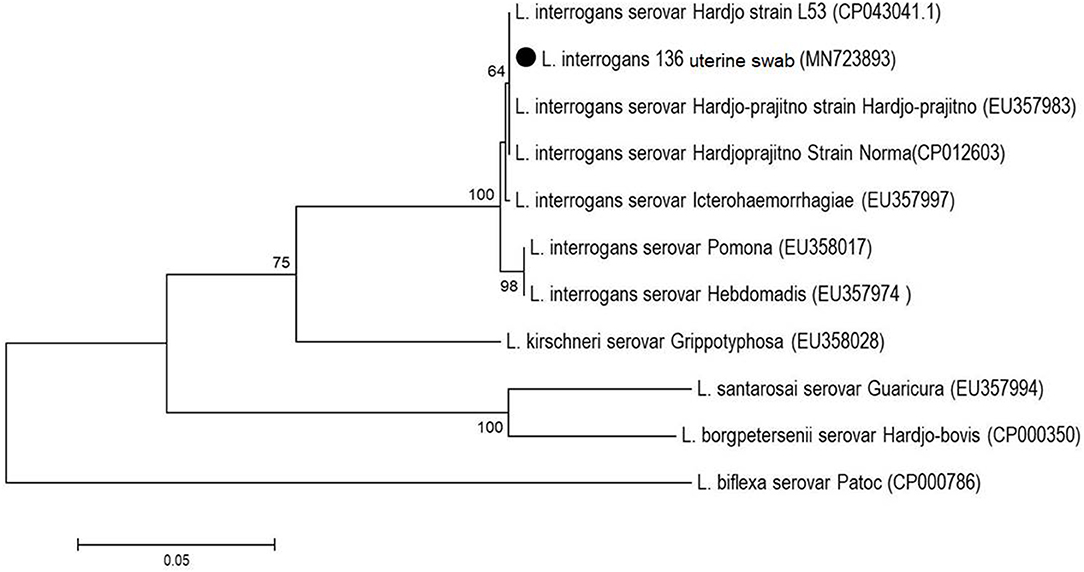
Figure 3. Phylogenetic analysis based on partial secY gene of Leptospira spp. from the uterine swab sample (black circle). The tree was elaborated with Neighbor-Joining method and Tamura-3 parameter model with bootstrap test of 1,000 replicates.
Results
Among the total samples (478) subjected to the internal control PCR assay, only five ovarian samples were not amplified and were disregarded, resulting in a total of 473 samples that were analyzed. In the rrs PCR analysis, 11/473 samples were positive (2.3%), 10 of which were kidney samples, and one was a uterine swab. All ovarian samples were negative. None of these positive kidney samples showed amplification of the secY gene. However, the rrs-positive amplicons were sequenced (GenBank accession numbers from MN720219 to MN720228), and it was possible to identify two pathogenic species: L. interrogans, accounting for 60.0% (6/10) of these samples, and L. borgpetersenii, accounting for 20.0% (2/10), while 20.0% (2/10) of the samples were identified only to the genus level and also corresponded to pathogenic species (Figure 2).
The positive uterine swab sample was identified as L. interrogans by the partial sequencing of the secY gene (GenBank accession number: MN723893) and showed genetic proximity to strains belonging to the serovar Hardjo (Figure 3). As each sample represented a single animal, it was not possible to correlate the samples with each other.
Discussion
Although buffaloes are considered rustic, as reflected in their natural resistance to disease in the environment, these animals can be affected by infectious agents, including those related to reproductive disorders (21). In this work, the identification of bacterial DNA in animals showed that buffaloes may be as sensitive to Leptospira spp. as cattle, including showing renal colonization and, consequently, the elimination of bacteria through the urine, which becomes a source of environmental contamination (22).
In the Amazon, climatic conditions may favor the occurrence of leptospirosis in cattle and buffalo, since the risk of animals contracting the disease is higher in regions affected by seasonal flooding (23). In bovines, the control of leptospirosis is difficult because different strains of leptospires may be adaptable to cattle (24) and the disease is not yet well-understood in buffalo, as few studies have provided information about the species of Leptospira circulating in these animals.
The direct sequencing of PCR products from kidney samples showed that the pathogenic species L. interrogans was most prevalent (6/10) in these samples, whereas the L. borgpetersenii is usually reported to be the most prevalent species in cattle (7, 9). In other animal species such as ewes and horses, L. interrogans is most commonly found in the genital tract (25, 26); in association with such infections, recurrent uveitis, a classic leptospirosis injury in horses, seems to be directly related to strains of L. interrogans (27). Two sequences (MN720221 and MN720222) formed two independent clades and could not be identified at the species level, but is suggested that this two samples are L. borgpetersenii-like species, as these two samples showed >98.9% identity with L. borgpetersenii; in particular the MN720222 sample was homologous to L. borgpetersenii isolated from a human patient in Guayaquil, Ecuador (28).
A difficulty found in the present study was that the kidney samples positive for the rrs gene could not be amplified by PCR targeting the secY gene. This situation has also been observed in other studies, and the discrepancy between PCR assays could arise due to the amount of original bacterial DNA in the material as well as the quality of the sample tested (7, 9). In addition, not all PCR products can always be sequenced (29) or generate interpretable data (30).
Despite the limitations of the use of the rrs gene to separate Leptospira species (2), this target and secY gene, which are used for the molecular characterization of leptospires, have many characterized samples deposited on GenBank which facilitates sample typing (8, 31–33). Increasingly, different target genes have been described and proposed to assist in the molecular characterization of leptospires (lipL32, flaB, gyrB, rpoB, among others) which had success both in characterizing isolates and directly from PCR products, although this last option is less sensitive. These targets can generate “barcodes” which have variable discriminatory power, and can be combined to increase the differentiation of Leptospira species (34).
An interesting result was that L. santarosai was not detected in the buffalo samples, differing from what is reported for cattle in other regions of the country, such as the southeast, where this species is one of the most prevalent (8, 24, 35). The dimensions and heterogeneity of the with climatic conditions in the Amazon make this region unique and different from other parts of the country, which is reflected in the etiological agent-host relationships and may account for the situation found in cattle from the Amazon region, where L. santarosai may not be involved in leptospirosis in these animals (9, 36). Regarding buffalo, more work is needed to evaluate the participation of L. santarosai and others species in buffalo leptospirosis.
No ovarian specimens were PCR positive, suggesting that this organ is not be a preferred site for the colonization of leptospires in buffalos, although there has been one report of leptospiral DNA detected in the ovaries of slaughtered sheep (37). The isolation of L. interrogans from ovaries has been reported, but only in experimentally infected hamsters in the acute phase of the disease (38). The potential adaptability of leptospires to their hosts, especially in cattle, leads to a chronic character of these infections, where extrarenal colonization by leptospires has been described focusing on the structures of the female reproductive system, such as the vagina, uteri, and oviducts (39–41).
The positive uterine swab sample (MN723893), belonging to L. interrogans, showed homology to sequences characterized as belonging to serovar Hardjo strains, including an autochthonous sample (CP043041.1) isolated from the urine of dairy cattle in Paraná, in the southern region of Brazil (42). This finding may suggest that sexual transmission may be occurring among these animals. The presence of Leptospira in the reproductive tract of animals (25, 40, 41) demonstrates the tissue tropism of the bacterium, causing reproductive disorders, especially the strains of the Sejroe serogroup, which are considered to be adapted to cattle, promoting chronic infection of the genital tract and subtly compromising the reproductive performance of these animals for long periods. This condition has recently been described as a syndrome, referred to as bovine genital leptospirosis (BGL) (43).
Conclusion
This is the first report of leptospires species identified in buffaloes from the Brazilian Amazon and revealed that these animals may be carriers of different pathogenic Leptospira species, similar to bovines. Genital colonization was found, showing that the reproductive system of the buffaloes may be affected.
Data Availability Statement
The datasets generated for this study can be found in the Genbank NCBI. Accession numbers from MN720219 to MN720228 and Genbank accession number: MN723893.
Ethics Statement
The animal study was reviewed and approved by Ethics Committee on Animal Use of the School of Veterinary Medicine and Animal Science (University of São Paulo).
Author Contributions
IG, AM, and IP carried out the collection of samples at the slaughterhouse. IG, GO, JF, AF, and MC performed all the laboratory tests. IG and ST performed the interpretation of DNA sequencing results. IG wrote the manuscript and ST did the translation. MH accurately reviewed the manuscript. All authors have read and approved the final version of the manuscript.
Funding
This study was financed in part by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), Brasil – Finance Code 001 and by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico)–(420110/2018-6).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
MH thanked CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) the fellowship. IG thanked CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for the scholarship.
References
1. Faine S, Adler B, Bolin C, Perolat P. Leptospira and Leptospirosis. 2nd ed. Melbourne: Medisci. (1999). 272 p.
2. Vincent AT, Schiettekatte O, Goarant C, Neela VK, Bernet E, Thibeaux R, et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl Trop Dis. (2019) 13:e0007270. doi: 10.1371/journal.pntd.0007270
3. Levett PN. Leptospirosis. Clin Microbiol Rev. (2001) 13:296–326. doi: 10.1128/CMR.14.2.296-326.2001
4. Galiero G. Causes of infectious abortion in the mediterranean buffalo. Ital J Anim Sci. (2007) 6:194–9. doi: 10.4081/ijas.2007.s2.194
5. Marianelli C, Tarantino M, Astarita S, Martucciello A, Capuano F, Galiero G. Molecular detection of Leptospira species in aborted fetuses of water buffalo. Vet Rec. (2007) 161:310–11. doi: 10.1136/vr.161.9.310
6. Vasconcellos SA, Oliveira JCF, Morais ZM, Baruselli PS, Amaral R, Pinheiro SR, et al. Isolation of Leptospira santarosai, serovar guaricura from buffaloes (Bubalus bubalis) in vale do Ribeira, São Paulo, Brazil. Braz J Microbiol. (2001) 32:298–300. doi: 10.1590/S1517-83822001000400008
7. Hamond C, Pestana CP, Medeiros MA, Lilenbaum W. Genotyping of Leptospira directly in urine samples of cattle demonstrates a diversity of species and strains in Brazil. Epidemiol Infect. (2015) 144:72–5. doi: 10.1017/S0950268815001363
8. Loureiro AP, Hamond C, Pinto P, Bremont S, Bourhy P, Lilenbaum W. Molecular analysis of leptospires from serogroup Sejroe obtained from asymptomatic cattle in Rio de Janeiro – Brazil reveals genetic proximity to serovar guaricura. Res Vet Sci. (2016) 105:249–53. doi: 10.1016/j.rvsc.2016.02.012
9. Guedes IB, Araújo SAA, Souza GO, Silva SOS, Taniwaki SA, Cortez A, et al. Circulating Leptospira species identified in cattle of the Brazilian Amazon. Acta Trop. (2019) 191:212–16. doi: 10.1016/j.actatropica.2019.01.011
10. IBGE. Pesquisa pecuária municipal. Instituto Brasileiro de Geografia e Estatística. (2018). Available online at: https://sidra.ibge.gov.br/pesquisa/ppm/quadros/brasil/2018 (accessed April 20, 2020).
11. Barreto EO, Lobato AS, Pereira PVV, Serra DRO. Caracterização do turismo de base comunitária em polos turísticos do estado do pará. Revista Brasileira de Ecoturismo. (2017) 10:113–27. doi: 10.34024/rbecotur.2017.v10.6620
12. Agampodi SB, Peacock SJ, Thevanesam V, Nugegoda DB, Smythe L, Thaipadungpanit J, et al. Leptospirosis outbreak in Sri Lanka in 2008: lessons for assessing the global burden of disease. Am J Trop Med Hyg. (2011) 85:471–8. doi: 10.4269/ajtmh.2011.11-0276
13. Viana RB, Del Fava C, Moura ACB, Cardoso EC, De Araújo CV, Monteiro BM, et al. Ocorrência de anticorpos anti- Neospora caninum, Brucella sp. e leptospira spp. em búfalos (bubalus bubalis) criados na amazônia. Arq Inst Biol. (2009) 76:453–57. doi: 10.13140/2.1.1966.1446
14. Oliveira GC, Silva DB, Pinheiro VLC, Langoni H. Pesquisa de anticorpos anti-leptospíricos em bubalino. Ars Vet. (2013) 29:33. doi: 10.15361/2175-0106.2013v29n4p33
15. Rocha KS, Lima MS, Paz GS, Langoni H, Moraes CCG. Detecção de anticorpo anti-Brucella sp. e anti- Leptospira spp. em búfalos (Bubalus bubalis) abatidos em matadouro na cidade de Belém, Pará. Revista de Ciências Agrárias. (2019) 62:4. doi: 10.22491/rca.2019.3046
16. Bottero MT, Civera T, Anastacio A, Turi RM, Rosati S. Identification of cow's milk in buffalo cheese by duplex polymerase chain reaction. J Food Prot. (2002) 65:362–6. doi: 10.4315/0362-028X-65.2.362
17. Mérien F, Amouriaux P, Perolat P, Baranton G, Saint-girons I. Polymerase chain reaction goes detection of Leptospira spp. in clinical samples. J Clin Microbiol. (1992) 30:2219–24.
18. Ahmed N, Devi SM, Valverde MDLÁ, Vijayachari P, Machang RS, Ellis W, et al. Multilocus sequence typing method for identification and genotypic classification of pathogenic Leptospira species. Ann Clin Microbiol Antimicrob. (2006) 5:28. doi: 10.1186/1476-0711-5-28
19. Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Sympo Ser. (1999) 41:95–8.
20. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. (2016) 33:1870–74. doi: 10.1093/molbev/msw054
21. Fujii TU, Kasai N, Vasconcellos SA, Richtzenhain LJ, Cortez A, Souza SLP, et al. Anticorpos anti- Neospora caninum e contra outros agentes de abortamentos em búfalas do vale do Ribeira, São Paulo, Brasil. Arq Inst Biol. (2001) 68:5–9.
22. Denipitiya DTH, Chandrasekharan NV, Abeyewickreme W, Hartskeerl RA, Hapugoda MD. Identification of cattle, buffaloes and rodents as reservoir animals of Leptospira in the district of Gampaha, Sri Lanka. BMC Research Notes. (2017) 10:134. doi: 10.1186/s13104-017-2457-4
23. Chadsuthi S, Chalvet-Monfray K, Wiratsudakul A, Suwancharoen D, Cappelle J. A remotely sensed flooding indicator associated with cattle and buffalo leptospirosis cases in Thailand 2011–2013. BMC Infect Dis. (2018) 18:602. doi: 10.1186/s12879-018-3537-3
24. Pinto PS, Pestana C, Medeiros MA, Lilenbaum W. Plurality of Leptospira strains on slaughtered animals suggest a broader concept of adaptability of leptospires to cattle. Acta Trop. (2017) 172:156–9. doi: 10.1016/j.actatropica.2017.04.032
25. Director A, Penna B, Hamond C, Loureiro AP, Martins G, Medeiros MA, et al. Isolation of Leptospira interrogans hardjoprajitno from vaginal fluid of a clinically healthy ewe suggests potential for venereal transmission. J Med Microbiol. (2014) 63:1234–6. doi: 10.1099/jmm.0.065466-0
26. Hamond C, Martins G, Bremont S, Medeiros MA, Bourhy P, Lilenbaum W. Predominance of Leptospira interrogans serovar bratislava DNA in vaginal fluid of mares suggests sexual transmission of leptospirosis. Anim. Reprod Sci. (2014) 151:275–9. doi: 10.1016/j.anireprosci.2014.10.019
27. Lucchesi PM, Parma AE, Arroyo GH. Serovar distribution of a DNA sequence involved in the antigenic relationship between Leptospira and equine cornea. BMC Microbiol. (2002) 2:3. doi: 10.1186/1471-2180-2-3
28. Chiriboga J, Barragan V, Arroyo G, Sosa A, Birdsell DN, España K, et al. High prevalence of intermediate Leptospira spp. DNA in febrile humans from Urban and rural ecuador. Emerg Infect Dis. (2015) 21:2141–7. doi: 10.3201/eid2112.140659
29. Agampodi SB, Dahanayaka NJ, Bandaranayaka AK, Perera M, Priyankara S, Weerawansa P, et al. Regional differences of leptospirosis in Sri Lanka: observations from a flood-associated outbreak in 2011. PLoS Negl Trop Dis. (2014) 8:e2626. doi: 10.1371/journal.pntd.0002626
30. Agampodi SB, Moreno AC, Vinetz JM, Matthias MA. Utility and limitations of direct multi-locus sequence typing on qPCR-positive blood to determine infecting Leptospira strain. Am J Trop Med Hyg. (2013) 88:184–5. doi: 10.4269/ajtmh.2012.12-0526
31. Victoria B, Ahmed A, Zuerner RL, Ahmed N, Bulach DM, Quinteiro J, et al. Conservation of the S10-SPC-A locus within otherwise highly plastic genomes provides phylogenetic insight into the genus Leptospira. PLoS ONE. (2008) 3:e2752. doi: 10.1371/journal.pone.0002752
32. Ahmed A, Engelberts MF, Boer KR, Ahmed N, Hartskeerl RA. Development and validation of a real-time PCR for detection of pathogenic Leptospira species in clinical materials. PLoS ONE. (2009) 4:e7093. doi: 10.1371/journal.pone.0007093
33. Miotto BA, Moreno LZ, Guilloux AG, Sousa GO, Loureiro AP, Moreno AM, et al. Molecular and serological characterization of the first Leptospira santarosai strain isolated from a dog. Acta Trop. (2016) 162:1–4. doi: 10.1016/j.actatropica.2016.06.007
34. Guernier V, Allan KJ, Goarant C. Advances and challenges in barcoding pathogenic and environmental Leptospira. Parasitology. (2018) 145:595–607. doi: 10.1017/S0031182017001147
35. Barbosa C, Martins G, Lilenbaum W. Infectivity and virulence of leptospiral strains of serogroup sejroe other than hardjo on experimentally infected hamsters. Braz J Microbiol. (2019) 50:1129–32. doi: 10.1007/s42770-019-00122-1
36. Guedes IB, Souza GO, Castro JFP, Souza-Filho AF, Rocha KS, Gomes MET, et al. Development of a pooled antigen for use in the macroscopic slide agglutination test (MSAT) to detect Sejroe serogroup exposure in cattle. J Microbiol Methods. (2019) 166:105737. doi: 10.1016/j.mimet.2019.105737
37. Silva AF, Farias PJA, Silva MLCR, Araújo júnior JP, Malossi CD, Ullmann LS, et al. High frequency of genital carriers of Leptospira sp. in sheep slaughtered in the semi-arid region of northeastern Brazil. Trop Anim Health Prod. (2019) 51:43–7. doi: 10.1007/s11250-018-1657-9
38. Camargo CRA, Vasconcellos SA, Nümberger R Jr, Passos EC, Morais ZM, Visintin AJ. Investigação sobre a presença de leptospiras nos ovários de hamsters experimentalmente infectados com Leptospira interrogans sorotipo pomona. Braz J Vet Res Ann Sci. (1993) 30:129–135.
39. Ellis WA, Thiermann AB. Isolation of leptospires from the genital tracts of Iowa cows. Am J Vet Res. (1986) 47:1694–6.
40. Loureiro AP, Pestana C, Medeiros MA, Lilenbaum W. High frequency of leptospiral vaginal carriers among slaughtered cows. Anim Reprod Sci. (2017) 178:50–54. doi: 10.1016/j.anireprosci.2017.01.008
41. Pires BC, Grapiglia JB, Moreira L, Jaeger LH, Carvalho-Costa A, Lilenbaum W. Ocurrence of uterine carriers for Leptospira interrogans on slaughtered cows. Microb Pathog. (2018) 114:163–5. doi: 10.1016/j.micpath.2017.11.056
42. Chideroli RT, Gonçalves DD, Suphoronski SA, Alfieri AF, Alfieri AA, Oliveira AG, et al. Culture strategies for isolation of fastidious Leptospira serovar hardjo and molecular differentiation of genotypes hardjobovis and hardjoprajitno. Front. Microbiol. (2017) 8:2155. doi: 10.3389/fmicb.2017.02155
Keywords: Leptospira, buffalo, PCR, DNA sequencing, Amazon
Citation: Guedes IB, Oliveira de Souza G, Fernandes de Paula Castro J, de Souza Filho AF, Cavalini MB, Taniwaki SA, Maia ALP, Pereira IC and Heinemann MB (2020) Identification of Pathogenic Leptospira Species in the Urogenital Tract of Water Buffaloes (Bubalus bubalis) From the Amazon River Delta Region, Brazil. Front. Vet. Sci. 7:269. doi: 10.3389/fvets.2020.00269
Received: 28 November 2019; Accepted: 21 April 2020;
Published: 14 May 2020.
Edited by:
Antonio Minervino, Federal University of Western Pará, BrazilReviewed by:
Pollyanna Mafra Soares, Federal University of Uberlandia, BrazilCyrille Goarant, Institut Pasteur de Nouvelle Calédonie, New Caledonia
Copyright © 2020 Guedes, Oliveira de Souza, Fernandes de Paula Castro, de Souza Filho, Cavalini, Taniwaki, Maia, Pereira and Heinemann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcos Bryan Heinemann, bWFyY29zYnJ5YW5AdXNwLmJy