 Maykel Andrés Galloso-Hernández1*
Maykel Andrés Galloso-Hernández1* Vicente Rodríguez-Estévez1Carlos Armando Alvarez-Díaz2Mildrey Soca-Perez3
Vicente Rodríguez-Estévez1Carlos Armando Alvarez-Díaz2Mildrey Soca-Perez3 Devon Ronald Dublin4Jesús Iglesias-Gómez3Leonel Simon Guelmes3†
Devon Ronald Dublin4Jesús Iglesias-Gómez3Leonel Simon Guelmes3†- 1Department of Animal Production, Universidad de Córdoba, Córdoba, Spain
- 2Department of Basic Science, Universidad Técnica de Machala, Machala, Ecuador
- 3Department of Research, Sustainable Systems of Animal Production, Experimental Station of Pastures and Forrage: Indio Hatuey, Matanzas, Cuba
- 4Language Department, Hokkaido University of Education, Sapporo, Japan
Water buffaloes (Bubalus bubalis) manifest different levels of selectivity for different pastures and forages. Knowledge of feed selectivity is important to facilitate the design of efficient production systems that take into account optimal animal welfare. In this study, the selectivity of nine 18-month old female water buffaloes for Leucaena leucocephala, Albizia lebbeck, Gliricidia sepium, and Moringa oleifera was evaluated. After 12 h of grazing Megathyrsus maximum, the animals were housed in individual shelters and 1.2 kg of leaves from each of the four tree species were offered to the animals simultaneously. The selectivity, measured as the intake of dry matter (DM), was highest for A. lebbeck (mean = 0.34 kgDM, SD = 0.05 kg), followed by L. leucocephala (mean = 0.30 kgDM, SD = 0.03 kg), M. oleifera (mean = 0.11 kgDM, SD = 0.05 kg), and G. sepium (mean = 0.10 kgDM, SD = 0.02 kg) (P < 0.01). The crude protein intake was highest for A. lebbeck and L. leucocephala. Notably, the less selected leaves were those of G. sepium and M. oleifera. This study suggests that the inclusion of A. lebbeck and L. leucocephala in silvopastoral systems may increase both the consumption and well-being of water buffaloes in the tropics.
Introduction
The importance of buffaloes (Bubalus bubalis) as a productive species has increased worldwide by 2% in recent years with a total population of 202 million (1). For example, Cuba has a population of 6 × 104 buffaloes (2, 3) with an annual increase of 13.7% and comparatively less than Brazil with 3 × 106 buffaloes (4). In Argentina, the population amounts to 8.5 × 104 with an annual growth of 13.1% (5).
The advantages of using buffaloes in tropical environments are the resistance to heat stress under shade (6, 7), and the fact that it allows this species to maintain birth rates above 80%, which is higher than cattle in similar conditions (8). However, the milk production per hectare (ha) is lower than cattle due to the low stocking rates (0.6–0.8 animals/ha) currently used to breed buffaloes (8, 9). The capacity of buffaloes to digest highly fibrous diets (10, 11) makes the use of trees an attractive strategy for the incorporation of proteins in the diet of these animals (12–15).
Tropical tree species are included in pastures in the form of silvopastoral systems. The incorporation of Leucaena leucocephala (13), Albizia lebbeck (16), Gliricidia sepium, and Moringa oleifera (17, 18) in silvopastoral systems improves the quality of feed for ruminants and decreases the impact of environmental stressors on the animals (6). The use of trees as a source of feed for ruminants has been ignored due to the limited understanding of their positive impact on production systems (19). Leguminous trees are a source of feed in sustainable livestock farming in the dry season (19–22). A good selection of trees maximizes the benefits of agroforestry systems (13, 21, 24). Leguminous trees (L. leucocephala, A. lebbeck, G. sepium, and M. oleifera) have perennial leaves and fruits with high nutritional value. Ruminal evaluations showed that diets comprised of pasture and tree leaves have a higher rate of degradation (i.e., organic material, dry matter, crude protein, gross energy). Leucaena trees, for example, have the highest rumen by-pass protein supply. This digestibility of the protein is 63% in L. leucocephala and around 60% in other tree species (25–28), which decreases the amount of commercial concentrates being used (21, 29, 30). According to Ku Vera et al. (26), we need to deepen our understanding through the manipulation of different diets in ruminants including the use of leguminous trees and grasses in tropical conditions. The branches of leguminous trees also contain condensed tannins with antiparasitic properties (31) that could reduce enteric CH4 emissions (30).
The productivity of buffaloes has been evaluated in silvopastoral systems in tropical countries including Cuba (8, 22, 33), and Brazil (34). Silvopastoral systems, combined with rusticity and high productive indexes (12, 21), improve the productive performance of buffaloes per ha by increasing the daily weight gain and milk production when compared to conventional systems (35). Previous studies have tested the effect of individual leguminous trees on ruminant metabolism (12, 25, 36). However, in natural conditions, animals select a mixture of foliage from different trees. The aim of this study was to evaluate the selectivity of water buffaloes for four tree species (L. leucocephala, A. lebbeck, G. sepium, M. oleifera) that are commonly used in silvopastoral systems as demonstrated in the aforementioned studies.
Materials and Methods
Ethics Statement
The experiment received the approval of the Scientific Council and the Ethics Committee of the “Indio Hatuey” Grass and Forage Experimental Station, Matanzas, Cuba. This was an observational study that did not involve any harm or cruelty to animals.
Animals and Study Site
Clinically healthy 18-month old female water buffaloes (n = 9), with an average body weight of 373.77 kg, and daily weight gain of 362 g were used in the study. Before being included in the experiment, the animals were treated with ivermectin (1 μg/kg, Labiomec, Labiofam, Habana, Cuba) to ensure that their consumption behavior is not affected by intestinal parasites.
The study was conducted between August and September of 2008 in the municipality of Périco, Matanzas, Cuba, located at 22°48′7″ of latitude north and 81°1′ of longitude west and 19.01 m above sea level. The experimental phase was carried out on hydrated red ferralitic soil with medium fertility, moderately acidic (pH 5.60), low in phosphorus content (2.43 mg/100 g), and containing 0.18% total nitrogen, 3.2 % organic matter, calcium predominates (11.84 mEq/100 g) among the exchangeable cations and the cation exchange capacity was slightly low (19.21 mEq/100 g) as previously described (37). The climate of the region is tropical, seasonally humid, with an annual average temperature between 24.3°C and 33.4°C, and relative humidity of 80%. The annual rainfall is 1,331 mm, where 79.8% of it occurs between May and October (38).
Twelve paddocks, each measuring 0.54 ha, were used through a rotation of 3 days of occupation and 42 days of rest. In the morning, the animals were moved to the paddocks, where the availability of feed was 2.45 Ton/ha of dry matter (DM), and from 18:00 h the animals had access to the leaves of the four species in the feeders (Figure 1). Grass availability was determined by the visual estimation method and the botanical composition was measured as previously described (6, 39). During the experiment, we collected five grass samples and one sample per tree leaf type. The feeding regime included the grazing of Megathyrsus maximum in paddocks and nocturnal supplementation with Saccharum officinarum (sugar cane) (0.52 kg DM/animal) and the PCN002 commercial feed for growing animals (MINAG, Habana, Cuba) (Table 1) (0.44 Kg DM/animal). In the grazing area (Figure 1A), free access to water for wallowing and natural shade by Dichrostachys cinerea (Marabú) was available in the wallowing areas. Drinking water and mineral salts were provided ad libitum.
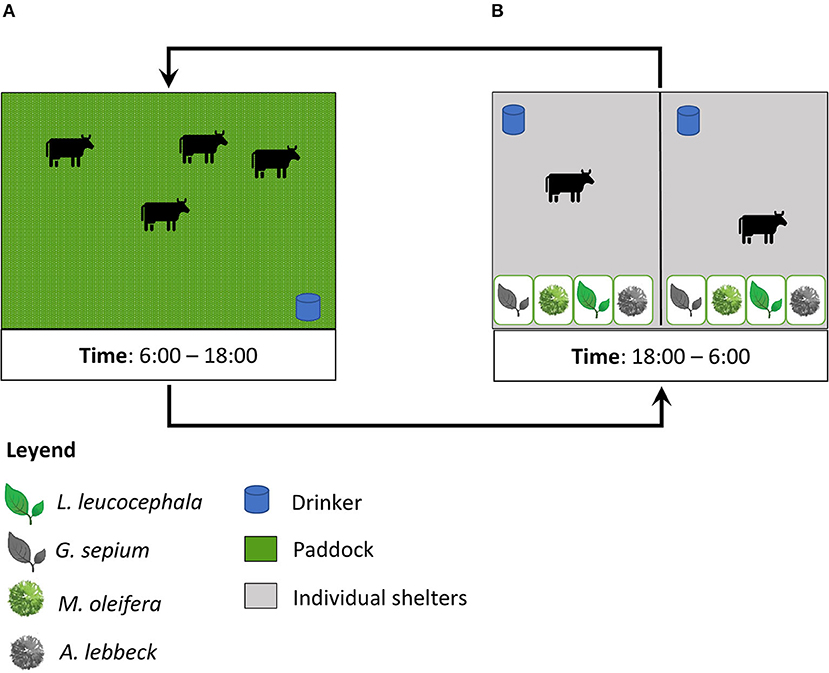
Figure 1. Description of management and experimental design. The figure shows the time animals spent in the pasture (A) and in the shelter (B).
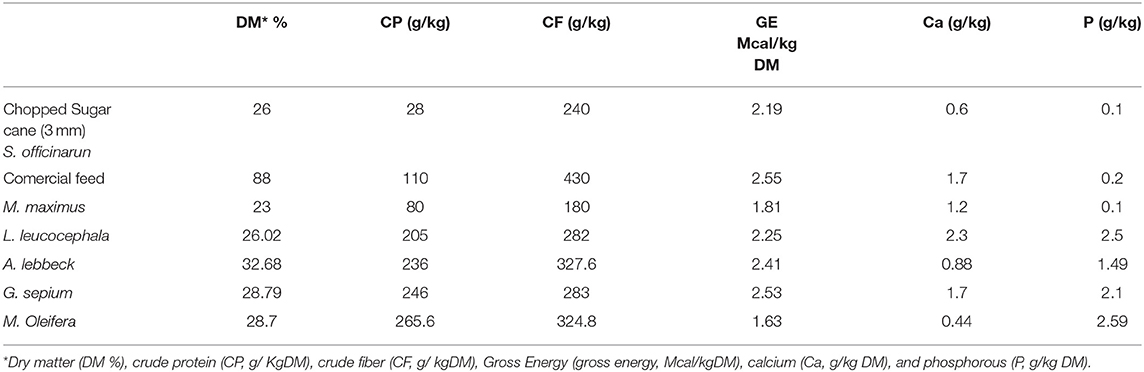
Table 1. Chemical composition of ingredients in the diet.
Experimental Design
In this study, the preference and consumption of fresh tree leaves were determined in a group of water buffaloes. Leucaena leucocephala, Albizia lebbeck, Gliricidia sepium, and Moringa oleifera were offered simultaneously (1.2 kg/animal of each fresh leaf type) to the animals. The buffaloes had no previous consumption experiences of the leguminous tree species studied. The trees were pruned and only the leaves that sprouted after 60 days were used in the experiment. Leaves of the trees were cut at a height of 1.7 meters, and the pods and stems were excluded. Fresh leaves were collected daily between the hours of 9:00–11:00. The trees were not irrigated nor treated with agrochemicals.
The animals were fed for 15 days with tree leaves and a base diet (Table 1). Experimental measurements were made daily from day 16 to day 32. After returning from the paddock, the animals were separated into individual shelters and each type of leaf was randomly placed separately in one of four feeders (each feeder measured 50 × 90 × 30 cm).
We used the repeated measured model shown in equation (1) below:
Yijklm = m + Ti + Pj + Aj + eijklm
where:
Yijklm = represented the m-n measurement made in the l-n square the j-n period in the i-n treatment in the k-n sampling within the j-n period.
m = population means or general intercept.
Ti = fixed effect of the i-n treatment (i = 1: Ll, 2: Al, 3: Gs, 4: Mo).
Pj = fixed effect of the i-n day (i = 1,…. 16).
Ai = fixed effect of the i-n animal (i = 1, 2, 3.9)
eijklm = random residual associated with the m- n measurement. ~ N (0, s2).
Analysis of Consumption and Chemical Composition of Fodder
The consumption of each leaf type was measured by subtracting the amount left in the feeders from the initial amount of leaves placed. For the weighing of fresh leaves, a digital scale with a sensitivity of 0.01 kg was used. Laboratory analysis was carried out at the beginning of the experiment to determine the DM, crude protein (CP), crude fiber (CF), and gross energy (GE). All procedures of feed analysis were performed as described in the manual “Official methods of analysis of AOAC international” (40).
The DM content of the leaves was determined individually in the laboratory of the Indio Hatuey Grass and Forage Experimental Station as described previously (41). To estimate the DM content, 500 g of homogenized grass samples were dried in a forced-air oven at 60°C for 48 h. The weight of each leaf sample was determined before and after the dehydration process (40) using an analytical balance of sensitivity 0.001 kg. After dehydration, the leaves were ground to a size of 2 mm and stored in amber glass jars.
The CP was calculated by the Kjeldahl method (42). CF was estimated by the Van Soest method (43, 44) briefly described by Garcia et al. (17). The GE was calculated with calorimetric methods using benzoic acid as the internal standard (45). This analytical procedure was performed with an adiabatic calorimetric pump (Model 1341EB, Parr instrument, Illinois, United States).
Statistical Analysis
The results were analyzed using the SPSS program version 22 (IBM Corp, New York, United States). An analysis of variance (ANOVA) was applied to determine whether there were any statistically significant differences between the measurements. Each buffalo was considered as an experimental unit (9) during the experimental period of 16 days. The weight of the feed (leaves and other supplements) before and after its placement in the feeders was evaluated individually. A Pearson correlation test was performed for the consumption of dry matter for the four leaf types.
Results
Water Buffaloes Leaf Type Preferences
There was a daily variation in the consumption of the four types of leaves studied (Figure 2A). The leaves of A. lebbeck (0.34 kgDM) and L. leucocephala (0.30 kgDM) were the most consumed, respectively, while the leaves of M. oleifera (0.11 kgDM) and G. sepium (0.10 kgDM) were the least consumed (Figures 2A, 3A). Each animal consumed 0.85 kg of DM of leaves daily (P < 0.01). The consumption of A lebbeck and L. leucocephala was always the most consumed, but it was observed that this amount reduced when the buffaloes consumed higher quantities of G. sepium and M. oleifera.
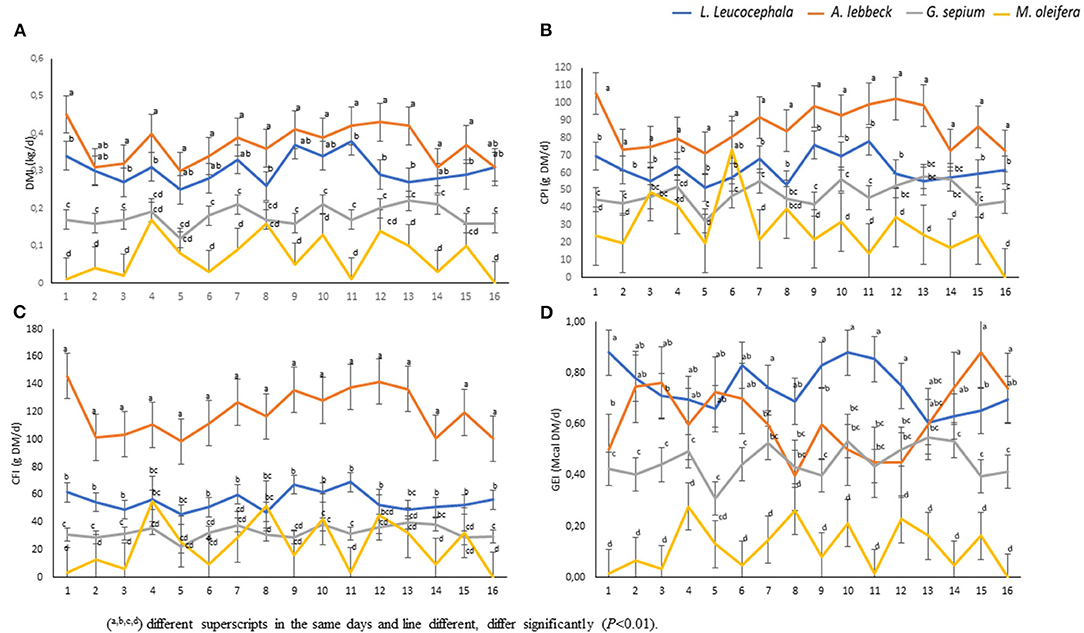
Figure 2. Consumption and overall nutritional contribution of the leaves. The statistical differences in the consumption of dry matter of the leaves are shown, as dry matter intake (DMI) (A); crude protein intake (CPI) (B); crude fiber intake (CFI) (C); and gross energy intake (GEI) (D). (Duncan, P < 0.01). N = 9 during 16 days of evaluation (P < 0.01).
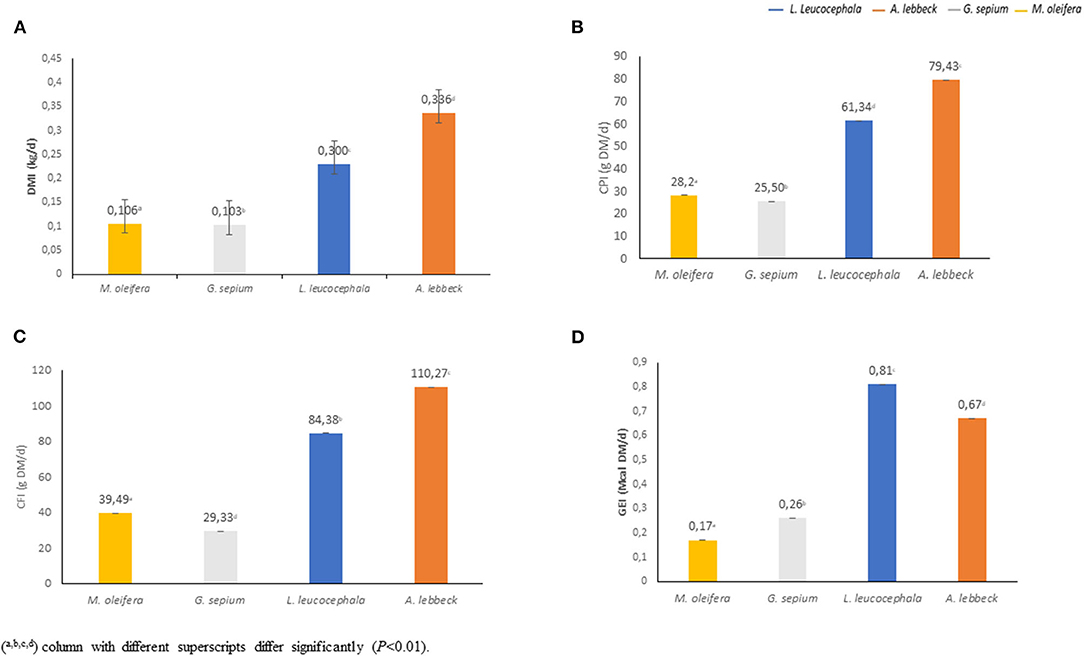
Figure 3. Nutritional contribution of each leaf type. The figure represents the average contribution (per animal/day) of dry matter intake (DMI) (A), crude protein intake (CPI) (B), crude fiber intake (CFI) (C) and gross energy intake (GEI) (D) (P < 0.01).
Contribution of Legumes to the Buffalo Diet
The analysis of the chemical composition revealed differences between the evaluated species (Table 1). The DM content was lower in M. oleifera and G. sepium. Notably, M. oleifera had the lowest GE values. L. leucocephala and M. oleifera were the species with the lowest and highest CP, respectively. CF content was lower in G. sepium and L. leucocephala but higher in M. oleífera and A. lebbeck (Table 1). The contribution of each of the species in terms of feed intake was different (Table 2 and Figures 2A–D). Of the leaves consumed, DM content differed between the four tree species during the period evaluated (Figure 2A). The CPI was highest in A. lebbeck and L. leucocephala followed by M. oleifera and G. sepium (P < 0.01; Figures 2B, 3B). The CFI was different between treatments. A. lebbeck had the highest CF content followed by L. leucocephala, M. oleifera, and G. sepium, respectively (P < 0.01; Figures 2C, 3C). However, the consumption of GE was highest in A. lebbeck followed by L. leucocephala, G. sepium, and M. oleifera, respectively (P < 0.01; Figures 2D, 3D). No differences were found in the consumption of calcium, phosphorous, and organic material present in the four types of leaves.
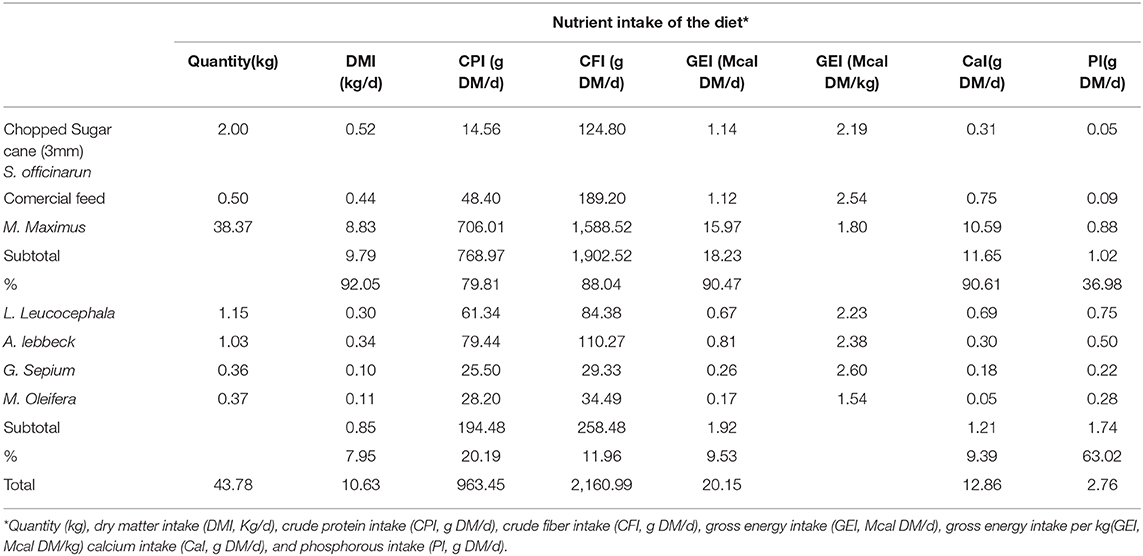
Table 2. Principal nutrient composition in the diet.
The average intake of fresh leaves was 2.91 kg/day and represents a daily DM consumption of 0.85 kg in this study (Table 2). The CPI of leaves was 194.48 g /animal/d; CFI was 258.48 g/ animal/d and GEI was 1.92 Mcal/animal/d.
The total DM Intake (leaves + M. maximum + S. officinarum + commercial feed) in this study was 10.62 KgDM/animal corresponding to 2.83 % body weight, and 936.45 g of CPI/animal, CFI = 2,160.99 g/animal; GEI 20.15 Mcal /animal, Ca = 12,86 g/animal and phosphorous = 2,76 g/animal (Table 2).
The DMI between L. leucocephala and A. lebbeck showed significant correlation (Pearson r = 0.54, P < 0.01); DM consumption of G. sepium and M. oleifera showed a positive correlation (Pearson r = 0.26; P < 0.01). Notably, the correlation of DMI between L. leucocephala and M. oleifera was negative (Pearson r = −0.28; P < 0.01); A. lebbeck and G. sepium had a value of r = 0.32. DMI of the most preferred and least preferred legume leaves (L. leucocephala + A. lebbeck vs. G. sepium + M. oleifera) showed a negative correlation (Pearson r = −0.43; P < 0.01).
Discussion
Consumption of the leguminous trees was higher in L. leucocephala but provided less DM, CP and CF when compared to A. lebbeck. In this study, A. lebbeck was the leaf most consumed on the basis of DM, except for GE, where L. leucocephala was the most consumed species in the diet.
Previously, an evaluation in the paddock was conducted on bovine browsing behavior in an arboretum of 60 tree species. The findings showed a preference for L. leucocephala and A. lebbeck in cattle (46). This evaluation was repeated with buffaloes resulting in a similar preference for L. leucocephala (24). According to García et al. (18), M. oleifera was a moderately-consumed plant. A. lebbeck and L leucocephala was the most consumed species, probably due to the ease of collecting these branches in the feeders. L. leucocephala has very small leaves which are easily licked from the bottom of the feeders while A. lebbeck and G. sepium are more resistant to this type of action (47). However, it was observed that the rate of wilting of M. oleifera, prevents its consumption by the animals. We could attribute the preference for L. leucocephala and A. lebbeck to the “feeding memory” (i.e., the tendency of animals to remember the information of feed for up to 3 years) effect, reported in sheep (48) and buffaloes (49). The difference in the consumption and acceptability of the tree leaves could be suggestive of the satisfaction of the needs of the appetite and not that of “hunger” (50, 51). Additionally, ruminants prefer leaf types as a supplement that rapidly provides the highest satiety level of nutrients (52, 53).
The DM content of the feed in ruminants is partially responsible for limiting its consumption and ingestion through short-term regulation along with the fast fermentation of the carbohydrate contents (10, 50). These results are indicative of the satisfaction of the animal's needs and coincide with what was reported by Mendez and Lima (54) who indicated that the voluntary consumption (expressed in the percentage of body weight) is 2.59–3.09%) in buffalos. Similar results were previously published by Paul and Lai (55).
Tree leaves (L. leucocephala, A. lebbeck, G. sepium, and M. oleifera) contributed 194.48 g DM to the daily CPI. This represents 20.19% of the protein even though the trees only provided 7.95% of the DM in the diet. The rest of the diet provided 768.97 g CPI/animal/day (79.81% of diet). Paul and Lai (55) reported a daily CPI of 298–310 g in the diet of female buffaloes with a similar body weight of 300–350 kg and daily weight gain of 250 g/day, while being fed with different proportions of feed in India. The value of the proportion of CPI due to the ingestion of tree leaves in this study was similar to a study in bovines in Cuba (21, 45, 56). Of the four types of leaves, A. lebbeck was the one that contributed the most to the protein needs of buffaloes and was sufficient to cover all the requirements of the animals used. Valenciaga et al. (25, 57) estimated in Cuba that a DM intake of 9.95 and 10.25 kg DM in buffaloes with 600 kg of BW was needed.
Delgado et al. (12) reported an apparent digestibility above 60% for trees such as G. sepium and L. leucocephala in similar conditions in Cuba. In this sense, Albores-Moreno et al. (58) reported high voluntary consumption in cattle (10.26–12.48 kg of D/animal/day) of legumes and found that the secondary metabolite content of plants did not interfere with the voluntary intake. Additionally, Barros-Rodríguez et al. (28) report that up to 50% of L. leucocephala can be included in the diet. These criteria could explain the favorable consumption in favor of L. leucocephala and A. lebbeck, in the same way, that it can be attributed to the digestibility as reported by Delgado et al. (12). However, they do not explain why G. sepium and M. olifera were scarcely consumed.
Notably, the lower intake of M. oleifera could be related to the effects of this plant on the ruminal flora, volatile fatty acids, and ruminal ammonium concentration (59). This legume is low in fiber with higher degradability, gas production, short chain fatty acids production, and lower methane emissions as compared to wheat straw.
An in vitro study with an M. oleifera leaf extract as an alternative to monensin in sheep diets, found similar effects on the ruminal parameters (ruminal degradability, ammonia concentrations, and gas production) (60). The lower consumption of M. oleifera could also depend on the concentration of anti-nutritional factors (condensed tannin, tannins which are precipitants of proteins, terpenoids, and total sterols) (18, 31) and the ability of this leaf type to wilt, rather than its nutritional value. Previous studies attributed the texture, anti-nutritional factors and the apparent digestibility (17, 18, 32, 48) to the speed with which the branches lose qualities after being cut (61). It could also be attributed to the vegetative state or age (60- days-old) in which the leaves of the plants were collected (62, 63), climatic conditions (13, 56), the animal category (48), the amount and nutritional composition of the tree resource (64), DM content and apparent digestibility (32, 65–67). However, there does not seem to be a direct relationship between the nutritional composition and the palatability of the leaves of leguminous trees, and the leaves of L. leucocephala and A. lebbeck may generate similar stimuli in animals, meaning they are consumed more.
Pearson's correlation was significantly negative between the two most consumed species (L. leucocephala and A. lebbeck) and the two least consumed (G. sepium and M. oleífera). In this sense, there was no correspondence between what was reported by Santana-Perez et al. (47) regarding the low correlations of consumption of A. lebbeck (stems with irregular barks) compared to L. leucocephala, G. sepium, and M. oleifera (smooth stems), attributing it to the physical characteristics of the stems.
Analysis showed a negative correlation between L. leucocephala and M. oleifera in the DM intake, similar results were obtained previously (17, 18) where during grazing they observed M. olifera being rejected in comparison with L. leucocephala and other leguminous trees. Santana-Perez et al. (47) found differences in voluntary consumption in sheep and cattle, attributing it to the differences between the maximum diameters of the stems consumed. With G. sepium, L. leucocephala, M. oleifera, and A. lebbeck being the most consumed, respectively, the sheep did not consume the stems of G. sepium, attributing this phenomenon to the aversive stimulus generated by this plant.
Conclusions
There were significant differences in the consumption and acceptability of the leaves of the tree species. The highest consumption of leaves was that of L. leucocephala and A. lebbeck, while M. oleifera and G. sepium species were consumed in lower quantities. The leaves of A. lebbeck contributed more nutritionally to the animals, although the amounts consumed were equal to L. leucocephala. This study helps to define which tree species could be used in production systems with buffaloes in the tropics by providing insights into buffaloe preferences for different leaves. We suggest that future studies explore the effects of secondary metabolites on feed intake.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by the ethics committee of the Experimental Station: Indio Hatuey, Cuba.
Author Contributions
All authors also participated equally in the design, preparation, and review of this research.
Funding
BIOMAS-CUBA PROJECT. COSUDE. For grant and support of the execution of these experiments and Experimental Station: “Indio Hatuey” (Financial support of this research in to the topic's of sustainable Livestock production).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. FAOSTAT. FAOSTAT. Agriculture Datebase. (2010). Available online at: http://apps.fao.org/page/collections?.Subs~ety=agriculture (accessed October 24, 2020).
2. MINAG. Conservación de los Recursos zoogenéticos (rzg) en Animales de Granja de la República de Cuba. (2014). Available online at: http://www.fao.org/3/i4787e/i4787s103b.pdf consulted (accessed July 7, 2018).
3. López JR. Perspectivas de La crianza del búfalo de agua (Bubalus bubalis) en la amazonía ecuatoriana. Rev Amazón Ciencia Tecnol. (2013) 2:19–29.
4. Brcko CC, Silva JAR, Martorano LG, Vilela RA, Nahúm BS, Silva AGM, et al. Infrared thermography to assess thermoregulatory reactions of female buffaloes in a humid tropical environment. Front Vet Sci. (2020) 7:180. doi: 10.3389/fvets.2020.00180
6. Galloso-Hernández MA, Rodríguez-Estévez V, Alvarez-Díaz CA, Soca-Pérez M, Dublin D, Iglesias-Gómez J, et al. Effect of silvopastoral systems in the thermoregulatory and feeding behaviors of water buffaloes under different conditions of heat stress. Front Vet Sci. (2020) 7:393. doi: 10.3389/fvets.2020.00393
7. Galloso Hernández MA, Rodríguez-Estevez V, Simon-Guelmes L, Soca-Perez M, Alvarez-Diaz CA, Dublin D, et al. Can trees replace the need for wallowing in river buffalo (Bubalus bubalis) in the tropics? In: Dupraz C, Gosme M, Lawson G, editors. Preliminary Results. (2019). pp. 696.
8. Simón L, Galloso M. Presencia y perspectivas de los búfalos en Cuba Presence and perspective of buffaloes in Cuban. Pastos Forrajes. (2011) 34:3–20.
9. Fundora O, Quintana FO, María GE. Comportamiento y composición de la canal de búfalos de río alimentados con una mezcla de pasto estrella, pastos naturales y leguminosas nativas. Rev Cubana Cienc Agric. (2004) 38:43–6.
10. Wanapat M, Phesatcha K. Rumen adaptation of swamp buffaloes (Bubalus bubalis) by high level of urea supplementation when fed on rice straw-based diet. Trop Animal Health Product. (2016) 48:1135–1140. doi: 10.1007/s11250-016-1064-z
11. Patra AK, Pal K, Lalhriatpuii M. Prediction of nitrogen excretion in buffalo production systems using dietary and animal variables. Agricult Syst.(2020) 182:102845. doi: 10.1016/j.agsy.2020.102845
12. Delgado D, Cairo J, Moreira O. Fermentación ruminal en búfalos de río y cebú comerciales, alimentados con forraje integral de caña de azúcar. Rev Cubana Ciencia Agrícola. (2005) 39:53–7.
13. Simón L. Del monocultivo de pastos al silvopastoreo: La experiencia de la EEPF Indio Hatuey. In: Simón L, editor. Silvopastoreo Un Nuevo Concepto de Pastizal. Matanzas: EEPF (2012). p. 11–24.
14. Fundora O. Performance of river buffaloes (Bubalus bubalis) from Buffalypso breed in feeding systems based on grazing: fifteen years of research in the Instituto de ciencia animal. Cuban J Agric Sci. (2015) 49:2015. Available online at: http://cjascience.com/index.php/CJAS/issue/view/35
15. FAO. (2011). Perspectivas Alimentarias, Análisis del Mercado de los Mercados Mundiales. Available online at: http://www.fao.org/giews/ (accessed October 24, 2020).
16. Palma JM, Flores R. Aproximación al estudio de la vegetación arbórea del estado de Colima, México. In: Centro Nacional de Investigaciones Agropecuarias, INIA-CENIAP, editor. Memorias: X Reunión de Avances en Investigación Agropecuaria Trópico'97. Maracay: Estado Aragua (1997). p. 88–90.
17. García DE, Medina MG. Composición química, metabolitos secundarios, valor nutritivo y aceptabilidad relativa de diez árboles forrajeros. Zoo Trop. (2006) 24:233.
18. García DE, Medina MG, Cova LJ, Torres A, Soca M, Pizzani P, et al. Preferencia de vacunos por el follaje de doce especies con potencial para sistemas agrosilvopastoriles en el Estado de Trujillo, Venezuela. Pastos Forrajes. (2008) 31:255.
19. Ørskov EB. Conferencia silvopastoral systems : technical, environmental and socio-economic challenges. Pastos Forrajes. (2005) 28:5–9.
20. Domínguez FA. Árboles y arbustos forrajeros: usos y perspectivas. In: Memoria: XII Reunión Anual COTECOCA. Boca del Río: Experimental Station of Pastures an Forrages: “Indio Hatuey” (1996). p. 141–56.
21. Milera MC. Recursos Forrajeros Herbáceos y Arbóreos. In: Estación Experimental de Pastos y 379. Experimental Station of Grass and Forages: “Indio Hatuey” (2016).
22. Iglesias-Gómez JM, Galloso-Hernández M, Toral-Pérez CO, Aguilar-Hernández A. Comportamiento Productivo y Conducta de Búfalos de Río y Toros Cebú En Silvopastoreo. Pastos y Forrajes. (2019) 42:223–9.
23. Vasquez F, Pezo D, Mora-Delgado J, Skarpe CH. Selectividad de especies forrajeras por bovinos en pastizales seminaturales del trópico centroamericano: un estudio basado en la observación sistemática del pastoreo. Zoo Trop. (2012) 30:63–80. Available at:http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0798-72692012000100008&lng=es&nrm=iso&tlng=es
24. Toral-Pérez OC, Iglesias-Gomez JM, Aguilar A. Selectividad de especies arbóreas por búfalos en pastoreo. Avan Investi Agropec. (2019) 23:41–54.
25. Valenciaga D, López JR, Chongo B, Saliba E, La OO, Cairo JG. Consumo y digestibilidad aparente de nutrientes en búfalas de río (Bubalus bubalis) alimentadas con Pennisetum purpureum vc. Cuba CT-115. Rev Cubana Ciencia Agrícola. (2007) 41:249–52.
26. Ku Vera JC, Briceño EG, Ruiz A, Mayo R, Ayala AJ, Aguilar CF, et al. Manipulación del metabolismo energético de los rumiantes en los trópicos: opciones para mejorar la producción y la calidad de la carne y leche. Rev Cubana Ciencia Agrícola. (2014) 48:43–53.
27. Delgado, et al. Degradabilidad ruminal del follaje de Gliricidia sepium y Leucaenaleucocephala en búfalos de río y toros cebú Revista Cubana de Ciencia Agrícola. (2008) 42:375–88.
28. Barros-Rodríguez M, Sandoval-Castro CA, Solorio-Sánchez J, Sarmiento-Franco L, Rojas-Herrera R, Klieve AV. Leucaena leucocephala IN RUMINANT NUTRITION. Tropical and Subtropical Agroecosystems. (2014) 17.
29. Baumer M. Trees as browse and to support animal production. In: Speedy A and Pugliese PL, editors. Legume Trees and Other Fodder Trees as Protein Sources for Livestock. Roma: FAO, Animal Production and Health Paper (1992). p. 1–10.
30. López-Vigoa O. Silvopastoral systems as alternative for sustainable animal production in the current context of tropical livestock production. Pastos Forrajes. (2017) 40:83–95.
31. Thi M, Van Binh D, Ørskov ER. “Effect of foliages containing condensed tannins and on gastrointestinal parasites. Anim Feed Sci Technol. (2005) 121:77–87. doi: 10.1016/j.anifeedsci.2005.02.013
32. Molina-Botero IC, Arroyave-Jaramillo J, Valencia-Salazar S, Barahona-Rosales R, Aguilar-Pérez CF, Ayala A, et al. Effects of tannins and saponins contained in foliage of Gliricidia Sepium and pods of Enterolobium cyclocarpum on fermentation, methane emissions and rumen microbial population in crossbred heifers. Anim Feed Sci Technol. (2019) 251:1–11. doi: 10.1016/j.anifeedsci.2019.01.011
33. Galloso-Hernández M, Rodríguez-Estevez V, Simon L, Alvarez-Diaz CA. Comportamiento alimentario y aptitud de búfalos en sistemas arborizados (SSP) y sin arborización (SSA). In: Chica AF, Julieta P, Garcia M, editors. VI Congreso de Jóvenes Investigadores. Creando Redes Doctorales: La Generación Del Conocimiento. Córdoba: UCO-Press, Editorial Universidad de Córdoba (2018).
34. Castro AC, Lourenço Júnior JDB, Santos NDFAD, Monteiro EMM, Aviz MABD, Garcia AR. Silvopastoral system in the Amazon region: tool to increase the productive performance of buffaloes. Cienc Rural. (2008) 38:2395–402. doi: 10.1590/S0103-84782008000800050
35. Simón L, Galloso MA. Efecto de la arborización en la crianza de búfalos de rio. Zoo Trop. (2008) 26:399–401.
36. La O, Chongo B, Delgado D, Valenciaga D, Pratt A, Elías A, et al. Caracterización de la degradabilidad ruminal in situ de nutrientes y de la digestibilidad intestinal in vitro de nitrógeno en leucaena leucocephala Cv. CIAT-7929. Rev Cubana Ciencia Agrícola. (2003) 37:387–93.
37. Hernández-Jiménez A, Pérez-Jiménez JM, Bosch-Infante D, Castro-Speck N. Clasificación De Los Suelos De Cuba 2015. San José de las Lajas: Ediciones INCA (2015).
38. INSMET. El Clima de Cuba. Características Generales. La Habana: Instituto de Meteorología. (2016). Available online at: http://www.insmet.cu/asp/genesis.asp?TB0=PLANTILLAS&TB1=CLIMAC&TB2=/clima/ClimaCuba.htm
39. Pérez Infante F. Nueva Ganadería en Pastos. La Habana: Asociación Cubana de Producción Animal (2013).
40. AOAC. International Official Methods of Analysis of AOAC International. 18th ed. Washintong, DC: AOAC International (2005).
41. Cáceres O, Ojeda F, González E, Arece J, Simón L, Lamela L, et al. Valor nutritivo de los principales recursos forrajeros en el trópico. In: Milagros Milera (comp.), Recursos Forrajeros Herbáceos y Arbóreos. Matanzas: EEPF: “Indio Hatuey” (2010). p. 174–98.
42. Kjendhal J. Official Methods of Analysis, 14th ed. Association of Official Agricultural Chemists. Washington, DC (1984).
43. Goering HK, Van Soest PJ. Forage fiber analyses (apparatus, reagents, procedures and some applications). In: AOAC International, editor. Agricultural Handbook. US Department of Agriculture (1970). p. 379
44. Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds.S0022-0302(91)78551-2
45. Caceres O, González-García E. Valor nutritivo de árboles, arbustos y otras plantas forrajeras para los rumiantes. Pastos y Forrajes. (2002) 25:15–20. Available online at: https://hal.archives-ouvertes.fr/hal-01190060 (accessed September 1, 2015).
46. Toral OC, Iglesias JM. Selectividad de especies arbóreas potencialmente útiles para sistemas de producción ganaderos. Zoo Trop. (2008) 26:197–200.
47. Santana-Pérez AA, Borrás-Sandoval LM, Cheng L, Verdecia-Acosta DM, Iglesias-Gómez JM, Vega-Albi AM, et al. Influencia de las dimensiones de las ramas de árboles forrajeros en el aprovechamiento por rumiantes. Ciencia y Agricultura. (2019) 16:25–38. doi: 10.19053/01228420.v16.n2.2019.9118
48. Forbes JM, Kyriazakis I. Feed preferences in farm animals: why don't they choose wisely? Proc Nutr Soc. (1995) 54:429–40. doi: 10.1079/PNS19950012
49. Alcantara PB, Alcantara VBG. Evaluation of “feeding intelligence” in Heitor Penteado, editor. Murrah Water Buffaloes Grazing Three Different Tropical Grasses. Sao Pablo: Instituto de Zootecnia–SAA (1996).
50. Forbes JM. Voluntary Feed Intake and Diet Selection of Farm Animals. Wallingford, GB: CAB International (2007).
51. Alvarez Díaz CA, Pérez H, De la Cruz Martín T, Quincosa J, Sánchez A. Fisiología Animal Aplicada. Antoquia: Editorial Universidad de Antioquia (2009). 380 p.
52. Baumont R. Palatability and feeding behaviour in ruminants. Ann Zoo. (1996) 45:385–400. doi: 10.1051/animres:19960501
53. Forbes J. M. (2000). Physiological and metabolic aspects of feed intake control. Farm Anim Metab Nutr. 1:319–34. doi: 10.1079/9780851993782.0319
55. Paul SS, Lai LD. Nutrient Requirements of Buffaloes. Hisar: Satish Serial Publishing House, CIRB (2010).
56. López-Vigoa O, Lamela-López L, Sánchez-Santana T, Olivera-Castro Y, García-López R, González-Ronquillo M. Influencia de la época del año sobre el valor nutricional de los forrajes, en un sistema silvopastoril. Pastos Forrajes. (2019) 42:57–67.
57. Gutiérrez O, Cairo J, Ramírez B, Vasallo G, Varela M. Comparison between two techniques for estimating voluntary intake of female buffaloes under grazing conditions. Technical note. Rev Cubana Ciencia Agrícola. (2015) 49:317–20.
58. Albores-Moreno S, Alayón-Gamboa JA, Morón-Ríos A, Ortiz-Colin PN, Ventura-Cordero J, González-Pech PG, et al. Influence of the composition and diversity of tree fodder grazed on the selection and voluntary intake by cattle in a tropical forest. Agroforest Syst. (2020) 94:1651–64. doi: 10.1007/s10457-020-00483-9
59. Dey A, Paul SS, Pandey P, Rathore R. Potential of moringa oleifera leaves in modulating in vitro methanogenesis and fermentation of wheat straw in buffalo potential of Moringa oleifera leaves in modulating in vitro methanogenesis and fermentation of wheat straw in buffalo. Ind J Anim Sci. (2014) 84:533–8.
60. Soltan YA, Morsy AS, Hashem NM, Sallam SMA. Impact of supplementary Moringa Oleifera leaf extract on ruminal nutrient degradation and mitigation methane formation in vitro Egyptian. J Nutr Feeds. (2019) 22:55–62. doi: 10.21608/ejnf.2019.75840
61. Montejo-Sierra I, Lamela-López L, López-Vigoa O. Deshidratación del follaje, al sol y a la sombra, de tres plantas forrajeras proteicas. Pastos Forrajes. (2018) 41:21–9.
62. Wencomo HB, Ortiz R. Phenological Performance of 23 Leucaena spp. Accession. Pastos Forrajes. (2010) 33:1–12.
63. Toral OC, Iglesias JM. Evaluation of forage tree and shrub accessions during the establishment period. Pastos Forrajes. (2012) 35:17–28.
64. Palma JM, Román L, Morales A, Aguirre MA. Comportamiento productivo y composición químico-nutricional de cuatro especies arbóreas. In: Memorias: III Taller Internacional Silvopastoril. Los árboles y Arbustos en la Ganadería. Matanzas: EEPF “Indio Hatuey” (1998). p. 45–7.
65. González E., and Cáceres, O. (2002). Valor nutritivo de arboles, arbustos y otras plantas forrajeras para los rumiantes. Pastos Forrajes 25:1–6.
66. González N, Galindo J, Abdalla AL. Effect of four inclusion levels of Morus alba L. cv cubana on microbial populations and fermentative products in river buffalo (Bubalus bubalis) rumen liquid Cuban. J Agricult Sci. (2015) 49:349–56.
Keywords: selectivity, feeding behavior, buffaloes (Bubalus bubalis), consumption, trees leaves, semi-intensive system
Citation: Galloso-Hernández MA, Rodríguez-Estévez V, Alvarez-Díaz CA, Soca-Perez M, Dublin DR, Iglesias-Gómez J and Guelmes LS (2020) Selectivity of Leguminous Trees by Water Buffaloes in Semi-intensive Systems. Front. Vet. Sci. 7:542338. doi: 10.3389/fvets.2020.542338
Received: 12 March 2020; Accepted: 16 September 2020;
Published: 24 November 2020.
Edited by:
André Mendes Jorge, São Paulo State University, BrazilReviewed by:
Octavio Alonso Castelán-Ortega, Universidad Autónoma del Estado de México, MexicoYosra Ahmed Soltan, Alexandria University, Egypt
Copyright © 2020 Galloso-Hernández, Rodríguez-Estévez, Alvarez-Díaz, Soca-Perez, Dublin, Iglesias-Gómez and Guelmes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maykel Andrés Galloso-Hernández, ejYyZ2FoZWFAdWNvLmVz; orcid.org/0000-0002-4247-4840
†Deceased, 6 August 2016. Contribution: Research Design