 Mughees Aizaz Alvi1
Mughees Aizaz Alvi1 John Asekhaen Ohiolei1
John Asekhaen Ohiolei1 Muhammad Saqib2
Muhammad Saqib2 Muhammad Haleem Tayyab2
Muhammad Haleem Tayyab2 Muhammad Umar Zafar Khan3
Muhammad Umar Zafar Khan3 Li Li1
Li Li1 Amjad Islam Aqib4Ali Hassan2Anum Aizaz Alvi5Warda Qamar6Bao-Quan Fu1
Amjad Islam Aqib4Ali Hassan2Anum Aizaz Alvi5Warda Qamar6Bao-Quan Fu1 Hong-Bin Yan1*
Hong-Bin Yan1* Wan-Zhong Jia1*
Wan-Zhong Jia1*- 1State Key Laboratory of Veterinary Etiological Biology, National Professional Laboratory for Animal Echinococcosis, Key Laboratory of Veterinary Parasitology of Gansu Province, Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Lanzhou, China
- 2Department of Clinical Medicine and Surgery, University of Agriculture, Faisalabad, Pakistan
- 3Depeartment of Veterinary Medicine, University of Veterinary and Animal Sciences, Lahore, Pakistan
- 4Department of Medicine, Cholistan University of Veterinary and Animal Sciences, Bahawalpur, Pakistan
- 5Institute of Pharmacy, Physiology and Pharmacology, University of Agriculture, Faisalabad, Pakistan
- 6Department of Pathobiology, University of Veterinary and Animal Sciences, Lahore (Jhang Campus), Jhang, Pakistan
Coenurus cerebralis is the larval stage of Taenia multiceps commonly found in the brain (cerebral form), intramuscular and subcutaneous tissues (non-cerebral form) of ungulates. Globally, few reports exist on the molecular characterization and genetic diversity of C. cerebralis with none available for Pakistan. The current study molecularly characterized 12 C. cerebralis isolates surgically recovered from sheep (n = 4) and goats (n = 8) from a total of 3,040 small ruminants using a portion of the cytochrome c oxidase subunit 1 (cox1) mitochondrial (mt) gene. NCBI BLAST search confirmed the identity of each isolate. A high haplotype and a low nucleotide diversity with three haplotypes from the 12 isolates were observed. The findings suggest the existence of unique haplotypes of C. cerebralis in Pakistan. The negative value of Tajima's D and the positive value of Fu's Fs were inconsistent with population expansion, however, the sample size was small. Bayesian phylogeny revealed that all Pakistani isolates alongside the Chinese sequences (obtained from GenBank) constituted a cluster while sequences from other regions constituted another cluster. This is the first molecular study to determine the genetic diversity of C. cerebralis in Pakistan and serves as a foundation for prospective studies on the prevalence and population structure of C. cerebralis in the country. Furthermore, in this study, we amplified only a partial segment of the cox1 gene from a limited sample size. This could have implications on the interpretation of the actual population structure in reality. Thus, we recommend future studies to consider a larger sample size in a massive epidemiological survey for further insights.
Introduction
The genus Taenia is a diverse group of helminths with more than 40 known species that infect livestock and humans causing cysticercosis and/or taeniasis (1, 2). Coenurus cerebralis refers to the larval stage of T. multiceps (an obligate intestinal tapeworm of dogs) infecting sheep and goat population across the world especially in tropical countries in Asia, Africa, and the Middle East (3). Sheep and goats are the common intermediate hosts but infection has also been reported in cattle and humans (4). Intermediate hosts including sheep and goats often become infected following feeding on vegetation and water contaminated by eggs from dogs (5, 6). This is followed by the development of the larval stage in different tissues including the brain, muscles, and skin (7, 8). The parasite completes its life cycle when the definitive hosts consume poorly discarded infected carcasses, (9). Predominantly, Coenurus cysts in the brain (10) present as either acute or chronic form. Acute clinical manifestations include fever, muscle tremors, hemorrhagic retinal lesions, ataxia, blindness, nystagmus, dysmetria, and scoliosis (11, 12). In chronic stages, in addition to acute signs, painful response to pressure over the cystic area, paralysis, blindness, lack of coordination, lethargy, and lack of response to stimuli are evident (13–15). Cyst formation is also frequently seen in subcutaneous and intramuscular tissues (16–21) where they possibly impair normal organ functioning and induce muscular pain (4).
Coenurosis is more prevalent in developing countries (22) with ovine coenurosis varying between 1 and 40% across countries in Africa, Asia, and the Middle East. In Asia and the Middle East, the prevalence in Bangladesh, India, and Jordan has been reported to be 2.5, 2.88, and 3% (13, 23, 24) while in Iran, prevalence in sheep and goat sits between 1.7 and 18.7% (16, 25–28). In countries like Ethiopia, Mozambique, Tanzania, Kenya, and Egypt, the prevalence may range from 1.5 to 42.1% (29–33). In the developed world like the United Kingdom, the infection is uncommon. However, abattoir-based surveys conducted during the 1980s put the prevalence between 0.5 and 5.58% (9). Low prevalence has also been reported in countries like France and Italy (34). Although the infection is cosmopolitan, to the best of our knowledge, only two necropsy-based case reports are available for Pakistan (35, 36).
Genetic variability among many species found within the family taeniidae has been well-documented especially for the genus Echinococcus. Nonetheless, the importance and degree of intraspecific variations within some species of Taenia remain to be fully understood. However, a good understanding of the genetic diversity and population structure of cestodes remains an important aspect that is essential for the implementation of control and prevention programs as it provides insights related to host specificity or geographical peculiarities. For T. multiceps, the genetic variability is poorly studied and there are arguments whether genetic variation may exist between isolates from different hosts, geographical location, or cerebral or non-cerebral forms. In a recent study, the phylogenetic analysis of nad1, cox1, and 12S rRNA mtDNA sequences showed that no monophyletic groups were based on geographical origin, organ location in the intermediate host (cerebral or non-cerebral), or species of the intermediate host (37). Meanwhile, from Sardinia sheep in Italy, Varcasia et al. (38) reported three variants of C. cerebralis using the cox1 and nad1 genes. In Iran, a high scale genetic diversity was reported in C. cerebralis isolates obtained from sheep and goats (12, 39) with the cox1 gene demonstrating 11 segregation sites resulting in seven haplotypes in sheep (39). Five distinct haplotypes of C. cerebralis cysts recovered from sheep, goats and cattle based on partial cox1 gene have also been documented in Greece (40). In China, the cox1, cytb, nad1, and nad4 genes have been employed in investigating the genetic diversity of C. cerebralis in Sichuan, Gansu, and Inner Mongolia (41–44) and the evidence suggests very low sequence variation 0–0.8%. Li et al. (41) further demonstrated that the cox1 mitochondrial gene of C. cerebralis obtained from sheep and goats from Gansu province of China were highly conserved.
On the other hand, small ruminant farming is one of the major determinants of advancement in developing countries. However, infectious diseases including parasitic infections have been known to constitute major obstacles to livestock production. Parasitic diseases cause enormous financial losses to owners of sheep and goat population as a result of poor growth, morbidity, poor carcasses value and mortality (34, 45). Mortality in infected sheep and goats reaches about 5% in Ethiopia (32), 25% in Istanbul, Turkey (46) while in Iran losses amounting to 10.2 million rials (27) due to condemnation of infected carcasses highlights the economic setbacks by tapeworm metacestodes. While Pakistan hosts a very large population of small ruminants, little is known on the prevalence, molecular and phylogenetic characteristics of C. cerebralis infecting small ruminants. Therefore, the current study was designed to molecularly identify and determine the genetic variability of C. cerebralis isolates surgically removed from sheep and goats using the mt cytochrome oxidase subunit 1 (cox1) gene to provide preliminary data for Pakistan besides contributing to our understanding of the global molecular epidemiology and population structure of C. cerebralis.
Materials and Methods
Study Area
Faisalabad is one of the three most populous cities in Pakistan. Geographically, the city is located 186 meters above sea level lying between 73° and 74° East (longitude), and 30° and 31.5° North (latitude). An extensive canal system and the presence of Ravi and Chenab rivers in the outskirts make the agricultural land highly fertile, fetching the attention of peasants toward agriculture farming and livestock rearing to earn their livelihoods.
Parasite Material
A total of 3,040 sheep and goats presented to the outdoor patient department of Clinical Medicine and Surgery, University of Agriculture, Faisalabad, Pakistan (irrespective of their disease or purpose of visiting) during May-December, 2019 were palpated thoroughly using digital palpation. Sheep and goats having subcutaneous or intramuscular palpable (cystic) swelling or bulge giving asymmetry to the infested organ were subjected to ultrasound scan procedure for the diagnosis of non-cerebral coenurosis. Hair from the bulging area was clipped and shaved and then animals (sheep and goats) were anesthetized with a combination of xylazine and diazepam at a dose rate of 1 mg/kg (47) and 0.25 mg/kg (48), respectively. Further, local infiltration of 3 ml of 2% lignocaine was also administered subcutaneously. Sterile Aquasonic® 100 Ultrasound Gel (Parker Laboratories) was applied and scanning was performed using 2–5 MHz convex probe (transducer) that was directly placed in the transverse plane over the cystic area using Convex scanner HS-1500 (HONDA® Electronics) and then was spun/moved in cranial and caudal planes for more enhanced visualization of the coenurus cysts. Transcutaneous ultrasonographic confirmation of cysts using convex array was carried out in <1 min followed by ultrasound-guided aseptic aspiration of the cystic fluid using fine needle aspiration technique (FNA). Each cyst was surgically removed and placed carefully in sterile falcon tubes and transferred to the In-House Preventive Veterinary Medicine Public Health Laboratory of the Department of Clinical Medicine and Surgery, University of Agriculture, Faisalabad, Pakistan. Gross morphological identification of the cysts was carried out as described previously by Soulsby (49), Faruk et al. (50), and Oryan et al. (45). The walls of the cysts were translucent and thin filled with a fluid having water like consistency. Protoscolices were firmly adhered to the germinal layer (45).
DNA Extraction, Amplification, and Sequencing
Genomic DNA from each cyst was extracted using Qiagen Blood and Tissue Kit (Qiagen, Hilden, Germany). A reaction mixture of 25 μl consisting of Ex Taq™ (12.5 μl) from Takara Bio, Kusatsu, Japan, forward (10 pmol) and reverse primers 1(10 pmol), extracted genomic DNA (0.5 μl), and RNAse free water up to the final volume of 25 μl was used to carry amplification of the portion of the cox1 gene (1,073 bp) using the forward (5′- CTTTGAGTGCGTGGTTGTTG-3′) and reverse (5′- AGAACCTACAGTGCACACAAT-3′) primers. In place of DNA, RNAse free water was added as a negative control. The PCR program was as follows: an initial denaturation at 95°C for 5 min followed by 35 cycles of denaturation (95°C for 30 sec), annealing (55°C for 40 sec), and extension (72°C for 1 min) and a final extension step at 72°C for 10 min. The PCR products were observed under UV light transilluminator using an agarose gel (1.5% w/v) stained with GelRed™. The sizes of the amplicons were estimated by running a 2,000-bp ladder in each gel. The PCR products were sent to Tsingke Biotechnology Company, Beijing for sequencing.
Molecular Analysis
DNA sequences were corrected manually for any misread nucleotides using Unipro UGENE v1.32.0 software. Using the same software, multiple sequence alignment was carried out. The identities of isolates were confirmed by NCBI BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Population Genetics and Phylogenetic Analyses
DnaSP 4.5 software was used to calculate nucleotide diversity (π), number of haplotypes (h), and haplotype diversity (Hd). Fu's Fs (51) and Tajima's D (52) were estimated by Arlequin software (53). A median-joining network (54) was constructed based on the partial mt cox1 sequences to examine intraspecific variation between haplotypes.
A dataset consisting of cox1 gene (639 bp) sequences of the representative haplotypes and other Taenia spp. (T. asiatica, T. crassiceps, T. hydatigena, T. ovis, T. pisiformis, T. saginata, T. solium, and T. taeniaeformis) retrieved from GenBank were used to construct a phylogenetic tree using the Bayesian method in MrBayes v.3.1.1 software. Echinococcus granulosus whose larvae can infect sheep and goats was used as an out-group.
Results
Ultrasonographic Findings
During the study period (May–December, 2019), 4 sheep and 8 goats were found infected with T. multiceps. The cysts were observed in the face, neck, trunk, and groin regions (Figure 1a). All cysts showed characteristics morphology containing clear fluid of varying volume with many protoscolices. Many scolices were attached to the internal wall of the cyst (Figure 1b). Ultrasound examination showed a hypoechoic cyst wall (Figure 1c, blue arrow) with anechoic cystic fluid (Figure 1c, white arrow) and protoscolices (Figure 1c, red arrow). The cysts appeared as hypoechoic structures with acoustic enhancement at the distal edge.
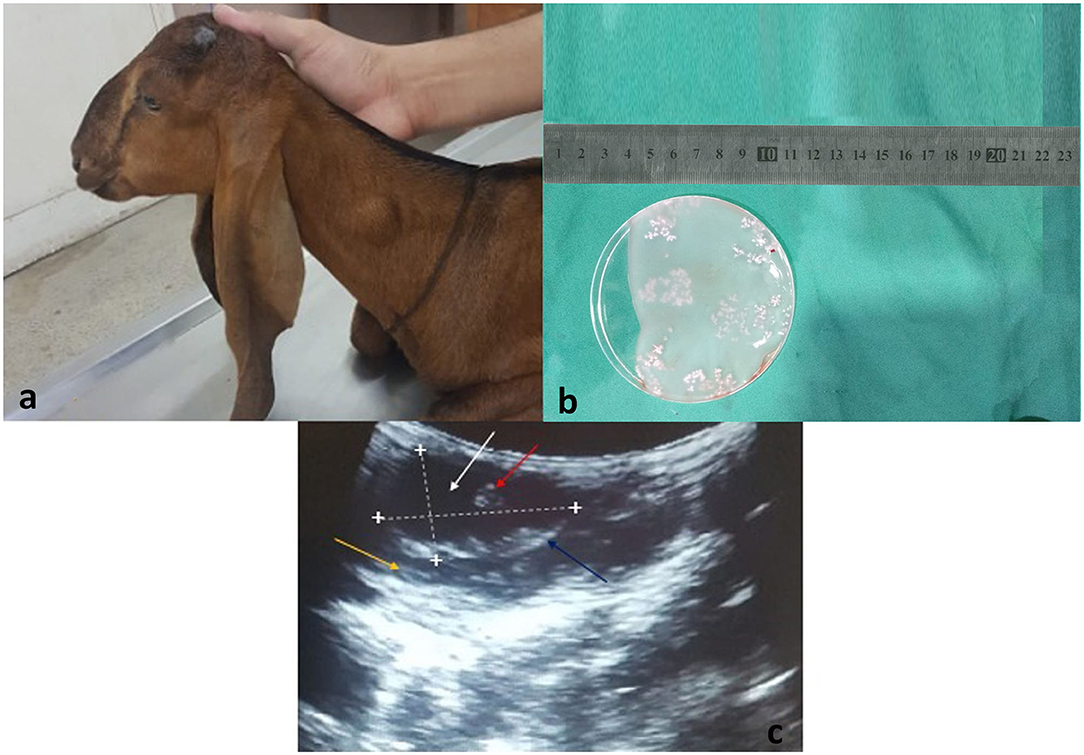
Figure 1. (a) A goat presented to the CMS Department of University of Agriculture, Faisalabad, Pakistan with a swollen mass around the neck region. (b) Surgically isolated Coenurus cerebralis cyst from the neck region of a goat. Scolices are attached to the internal layer of the cyst (white clusters). (c) Ultrasound scan of T. multiceps cyst in goat. Lengthwise and widthwise boundaries of the cyst are demarcated with dotted lines and plus (+) signs. The yellow arrow indicates the host's muscular layer, blue arrow indicates hypoechoic cyst wall, white arrow indicates anechoic cystic fluid, and red arrow indicates protoscolices.
Nucleotide Polymorphism and Population Indices
A PCR product of ~1,073 bp was obtained. After multiple sequence alignment, polymorphism analysis resulted in three parsimony informative sites from six mutation sites producing three haplotypes. According to the median-joining network of the partial cox1 gene sequences, PAK-Hap3 haplotype comprised only goat isolates, PAK-Hap2 of sheep origin, while PAK-Hap-1 from both hosts (Figure 2A). The examined nucleotide polymorphism resulted in amino acid changes (PAK-Hap2: 103L-103S; PAK-Hap1: 145N-145D).
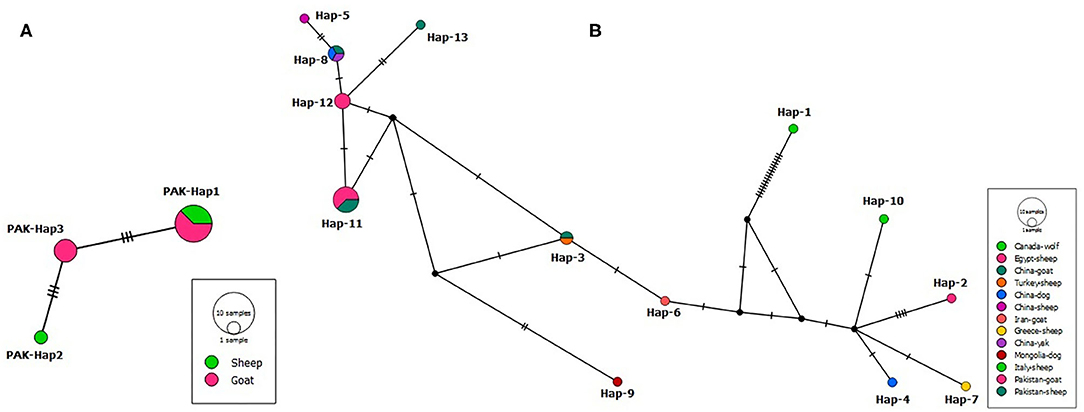
Figure 2. (A) Median-joining network of Coenurus cerebralis isolates from Pakistani sheep and goats based on cox1 gene (639 bp) sequences. Circle sizes are proportional to the haplotype frequencies. Hatch marks represent the number of mutations. Small black spheres are median vectors i.e., unsampled or hypothetical haplotypes. (B) Median-joining network of Taenia multiceps isolates from different hosts and geographical locations based on the mitochondrial cox1 gene (339 bp) sequences. Circle sizes are proportional to the haplotype frequencies. Hatch marks represent the number of mutations. Small black circles are median vectors (i.e., hypothetical haplotypes).
A nucleotide diversity of 0.00297 and haplotype diversity of 0.718 were observed for the overall T. multiceps population. The overall values of Tajima's D (−0.06198) and Fu's Fs (2.248) were not significant (p > 0.05). A similar observation was also found for T. multiceps from the separate populations (sheep and goats) (Table 1).
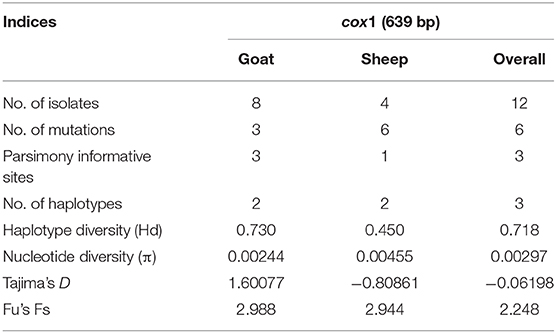
Table 1. Diversity and neutrality indices for Taenia multiceps populations from Pakistan.
To evaluate the geographical relatedness of the 12 Pakistani isolates from this study with those from different parts of the globe, a dataset of 25 cox1 sequences (339 bp) was used to draw a median-joining network (Figure 2B). The analysis showed 13 haplotypes, with a high haplotype diversity (0.883), and nucleotide diversity (0.23691). Non-significant Tajima's D = 2.24407 and Fu's Fs = 14.002 were obtained.
Haplotype Data Availability
Sequences of the representative haplotypes from this study have been deposited in the GenBank database under the accession numbers MT863708–MT863710.
Phylogenetic Analysis
The phylogenetic tree was inferred based on the mt cox1 gene sequences of the representative haplotypes along with those retrieved from GenBank. The Pakistani and Chinese isolates formed a cluster and were found at a reasonable distance to another cluster comprising isolates from other countries (Figure 3). In the phylogenetic tree, other Taenia species (T. asiatica, T. crassiceps, T. hydatigena, T. ovis, T. pisiformis, T. saginata, T. solium, and T. taeniaeformis) were represented while Echinococcus granulosus sensu stricto (genotype 1,3) were used as outgroups.
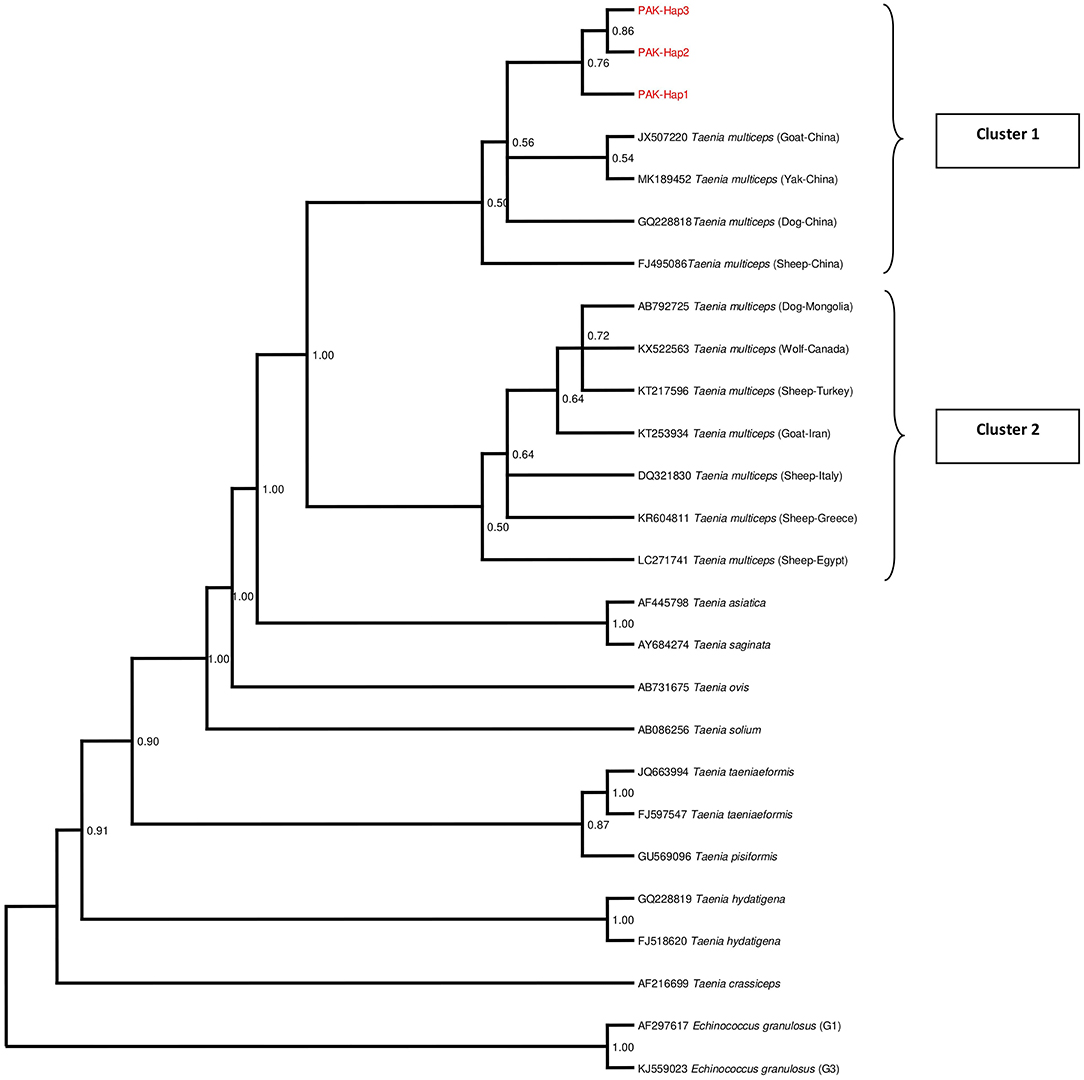
Figure 3. Bayesian phylogeny of Pakistani Coenurus cerebralis isolates inferred from the cox1 gene. Echinococcus granulosus is used as an outgroup. Red = Coenurus cerebralis haplotypes representing isolates from this study. Posterior probability values are depicted at the nodes.
Discussion
Progress in the caprine and ovine rearing sector is thought to be an important indicator of protracted advancement of developing countries. Parasitic diseases pose a serious threat to livestock production all over the world leading to hampered economic progress (55, 56). Coenurosis is one of the production limiting diseases of ungulates and the exact prevalence is almost impossible to evaluate since the animals are sent for slaughter without prior knowledge of infection by the farmers. Meanwhile, molecular analyses have demonstrated huge importance in the identification and characterization of taeniids cysts (57), as well as the epidemiology, population genetics and phylogeny (58, 59).
Cerebral and non-cerebral coenurosis are terms used to describe infection caused by T. multiceps based on the location of the coenuri. T. multiceps is regarded as the causative agent for cerebral coenurosis while the larval stages of Multiceps gaigeri and M. skrjabini were previously believed to be responsible for non-cerebral coenurosis in goats and sheep until recently when modern studies have adopted T. multiceps to refer to causative agents for both cerebral and non-cerebral coenurosis (37). Information related to non-cerebral coenurosis is scanty and mainly available in countries like Bangladesh (60), India (4, 61, 62), and Iran (63). It has also been reported in Sudan, Oman and Namibia (17).
Here, we detected non-cerebral coenurosis in 12 small ruminants (eight goats and four sheep) out of 3,040 sheep and goats examined from May–December, 2019 which demonstrate a prevalence of 0.39%. Similar low prevalence has been observed in Iran where the prevalence recorded were 0.48 and 1.79% in sheep and goats, respectively (45). A study conducted in Greece to determine the prevalence of non-cerebral coenurosis in sheep also demonstrated a low prevalence of 0.008% (7/90,415) (18).
In this study, a molecular description of C. cerebralis isolates from sheep and goats was reported for the first time in Pakistan based on mt cox1 gene as mtDNA remains an important marker in exploring intraspecific variation due to maternal inheritance, conserved structure, higher evolution rate, high genetic divergence, and absence of recombination (64–68). The result demonstrates a high haplotype and low nucleotide diversity comparable to the population indices reported in Italy (Hd = 0.664, π = 0.004) (69) as well as in Egypt, where a considerable number of SNP in the cox1 gene, mostly non-parsimony informative with low genetic diversity and fewer haplotypes were reported (70). Although this observation is mostly characteristic of an expanding population, our study revealed inconsistency with population expansion as demonstrated by the population indices: an insignificant negative Tajima's D and a positive Fu's Fs, unlike the Italian study where negative but insignificant values were observed for both Tajima's D and a positive Fu's Fs (69).
To date, the importance of the nucleotide substitutions and corresponding amino acid changes within C. cerebralis is unclear (44) though genetic variability leading to phenotypic differences in many other cestodes has been documented (57).
The phylogenetic analysis of the cox1 sequences from these isolates revealed a separate cluster suggesting the existence of unique C. cerebralis haplotypes in Pakistan. This pattern was also observed in the population network and it is also similar to the cluster pattern in a study conducted in Greece (40). All the sequences obtained from GenBank were found to be in one cluster irrespective of the country of origin, predilection site and host while Pakistani isolates occupied a second cluster with the Chinese sequences. This network pattern was also observed by Rostami et al. (22, 39) and Al-Riyami et al. (40) unlike Varcasia et al. (69). Nonetheless, this clustering pattern may indicate a genetic distinction between cerebral and non-cerebral coenurosis agents or intermediate host-specific specificity. Howbeit, further investigation is still warranted. The phylogenetic tree also confirmed T. asiatica and T. saginata as the closest to T. multiceps as reported previously (41).
Conclusion
In this study, C. cerebralis surgically isolated from sheep and goats in Faisalabad, Pakistan were characterized in what we believe is the first attempt to molecularly identify T. multiceps in Pakistan. Despite the sample size limitation, this study constitutes a significant preliminary data for Pakistan and also a contribution to understanding the global molecular epidemiology and population structure of C. cerebralis.
Data Availability Statement
Representative haplotypes sequences of cox1 gene from this study have been deposited in the GenBank database under the accession numbers MT863708–MT863710.
Ethics Statement
The animal study was reviewed and approved by the Laboratory of Veterinary Preventive Medicine and Public Health, University of Agriculture, Faisalabad, Pakistan. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author Contributions
MA, H-BY, MS, and W-ZJ conceived and designed the experiments. MT, AAq, AH, and MZ collected the samples. MA, AAl, WQ, and LL carried out the experiments as well as the data analyses. The initial draft of the manuscript was written by MA. JO, AAq, and H-BY. B-QF and W-ZJ gave constructive suggestions for revisions. All authors read and approved the final manuscript.
Funding
We are thankful to the National Key Research and Development Program of China (2018YFC1602504; 2017YFD0501301), and the Central Public-interest Scientific Institution Basal Research Fund (1610312020016; 161032017001) for funding this study. This study was part of a PhD research supported by Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences, the People's Republic of China.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Haukisalmi V, Konyaev S, Lavikainen A, Isomursu M, Nakao M. Description and life-cycle of Taenia lynciscapreoli sp. n. (Cestoda, Cyclophyllidea). ZooKeys. (2016)584:1–23. doi: 10.3897/zookeys.584.8171
2. Hoberg EP. Phylogeny of Taenia: species definitions and origins of human parasites. Parasitol Int. (2006) 55:23–30. doi: 10.1016/j.parint.2005.11.049
3. Amrabadi O, Oryan A, Moazeni M, Shari-Fiyazdi H, Akbari M. Histopathological and molecular evaluation of the experimentally infected goats by the larval forms of Taenia multiceps. Iran J Parasitol. (2019) 14:95. doi: 10.18502/ijpa.v14i1.722
4. Sharma DK, Chauhan PPS. Coenurosis status in Afro-Asian region: a review. Small Ruminant Res. (2006) 64:197–202. doi: 10.1016/j.smallrumres.2005.05.021
5. Sami M, Mirzaei M, Kheirandish R. Intramuscular and subcutaneous coenurosis in goats and sheep in south-east Iran. Comp Clin Path. (2012) 23:141–4. doi: 10.1007/s00580-012-1585-4
6. Tanowitz HB, Weiss LM, Wittner M. Tapeworms. Curr Infect Dis Rep. (2001) 3:77–84. doi: 10.1007/s11908-001-0062-z
7. Avcioglu H, Yildirim A, Duzlu O, Inci A, Terim KA, Balkaya I. Prevalence and molecular characterization of bovine coenurosis from Eastern Anatolian region of Turkey. Vet Parasitol. (2011) 176:59–64. doi: 10.1016/j.vetpar.2010.10.033
8. El-Neweshy MS, Khalafalla RE, Ahmed MMS, Al Mawly JH, El-Manakhly EM. First report of an outbreak of cerebral coenurosis in Dhofari goats in Oman. Rev Bras Parasitol Vet. (2019) 28:479–88. doi: 10.1590/s1984-29612019054
9. Scott PR. Diagnosis and treatment of coenurosis in sheep. Vet Parasitol. (2012) 189:75–8. doi: 10.1016/j.vetpar.2012.03.034
10. Christodoulopoulos G, Theodoropoulos G, Petrakos G. Epidemiological survey of cestode-larva disease in Greek sheep flocks. Vet Parasitol. (2008) 153:368–73. doi: 10.1016/j.vetpar.2008.02.002
11. Giadinis ND, Psychas V, Polizopoulou Z, Papadopoulos E, Papaioannou N, Komnenou AT, et al. Acute coenurosis of dairy sheep from 11 flocks in Greece. N Z Vet J. (2012) 60:247–53. doi: 10.1080/00480169.2012.665343
12. Akbari M, Moazeni M, Oryan A, Sharifiyazdi H, Amrabadi O. Experimental cerebral and non-cerebral coenurosis in goats: a comparative study on the morphological and molecular characteristics of the parasite. Vet Parasitol. (2015) 211:201–7. doi: 10.1016/j.vetpar.2015.06.013
13. Abo-Shehada MN, Jebreen E, Arab B, Mukbel R, Torgerson PR. Prevalence of Taenia multiceps in sheep in northern Jordan. Prev Vet Med. (2002) 55:201–7. doi: 10.1016/S0167-5877(02)00056-9
14. Giadinis N, Brellou G, Pourliotis K, Papazahariadou M, Sofianidis G, Poutahidis T, et al. Coenurosis in a beef cattle herd in Greece. Vet Rec. (2007) 161:697. doi: 10.1136/vr.161.20.697
15. Komnenou A, Dessiris A, Giadinis N. Surgical treatment of coenurosis (gid) in sheep. Vet Rec. (2000) 147:242–4. doi: 10.1136/vr.147.9.242
16. Oryan A, Nazifi S, Sharifiyazdi H, Ahmadnia S. Pathological, molecular, and biochemical characterization of Coenurus gaigeri in Iranian native goats. J Parasitol. (2010) 96:961–7. doi: 10.1645/GE-2399.1
17. Schuster RK, Sivakumar S, Wieckowsky T. Non-cerebral coenurosis in goats. Parasitol Res. (2010) 107:721–6. doi: 10.1007/s00436-010-1919-6
18. Christodoulopoulos G, Kassab A, Theodoropoulos G. Occurrence of non-cerebral coenurosis in sheep. J Helminthol. (2013) 87:125–7. doi: 10.1017/S0022149X1100085X
19. Christodoulopoulos G, Kassab A, Theodoropoulos G. Characteristics of non-cerebral coenurosis in tropical goats. Vet Parasitol. (2015) 211:216–22. doi: 10.1016/j.vetpar.2015.05.020
22. Rostami S, Beech RN, Salavati R, Baneshi MR, Kamyabi H, Harandi MF. Morphometric analysis of larval rostellar hooks in Taenia multiceps of sheep in Iran and its association with mitochondrial gene variability. Iranian J Parasitol. (2013) 8:579. Available online at: https://ijpa.tums.ac.ir/index.php/ijpa/article/view/452
23. Islam AW, Rahman MS. A report on incidence of gid of calves of Bangladesh. Indian J Anim Health. (1997) 36:187–8.
24. Varma TK, Malviya HC. Prevalence of coenuriosis in sheep, goat and pigs in Bareilly, Utar Pradesh. J Vet Parasitol. (1989) 3:69–71.
25. Kheirandish R, Sami M, Azizi S, Mirzaei M. Prevalence, predilection sites and pathological findings of Taenia multiceps coenuri in slaughtered goats from south-east Iran. Onderstepoort J Vet Res. (2012) 79:436. doi: 10.4102/ojvr.v79i1.436
26. Tavassoli M, Malekifard F, Soleimanzadeh A, Tajik H. Prevalence of Coenurus cerebralis in sheep in Northwest of Iran. Vet Res Forum. (2011) 2:274–6. Available online at: https://scholar.google.com.pk/scholar?q=Prevalence+of+Coenurus+cerebralis+in+sheep+in+Northwest+of+Iran.&hl=en&as_sdt=0&as_vis=1&oi=scholart
27. Oryan A, Moghaddar N, Gaur SNS. Metacestodes of sheep with special reference to their epidemiological status, pathogenesis and economic implications in Fars Province Iran. Vet Parasitol. (1994) 51:231–40. doi: 10.1016/0304-4017(94)90160-0
28. Javadi A, Hashemzadeh H, Shahbaz P. Study of Cerebral coenurosis rate in slaughtered sheep and goat brains in slaughterhouse in Tabriz. Iran Vet J. (2010) 6:59–63. Available online at: https://www.sid.ir/en/journal/ViewPaper.aspx?ID=176325
29. Asmare K, Sibhat B, Abera M, Haile A, Degefu H, Fentie T, et al. Systematic review and meta-analysis of metacestodes prevalence in small ruminants in Ethiopia. Prev Vet Med. (2016) 129:99–107. doi: 10.1016/j.prevetmed.2016.05.006
30. Afonso SMS, Mukaratirwa S, Hajovska K, Capece BPS, Cristofol C, Arboix M, et al. Prevalence and morphological characteristics of Taenia multiceps Cysts (Coenurus cerebralis) from Abattoir-Slaughtered and experimentally infected goats. J Neuroparasitol. (2011) 2:1–5. doi: 10.4303/jnp/235532
31. Miran MB, Nzalawahe J, Kassuku AA, Swai ES. Prevalence of coenurosis in sheep and goats at three slaughter slabs in Ngorongoro District, Tanzania. Trop Anim Health Prod. (2015) 47:1591–7. doi: 10.1007/s11250-015-0903-7
32. Achenef M, Markos T, Feescha G, Hibret A, Tembley S. Coenurus cerebralis infection in Ethiopian highland sheep: incidence and observations on pathogenesis and clinical signs. Trop Anim Health Prod. (1999) 31:15–24. doi: 10.1023/A:1005125316275
33. Abedl-Maogood SZ. Studies on metacestode of sheep with reference to serodiagnosis of Coenurus cerebralis (PhD dissertation). Faculty of Veterinary Medicine, Cairo University, Cario, Egypt (2005).
34. Scala A, Cancedda GM, Varcasia A, Ligios C, Garippa G, Genchi C. A survey of Taenia multiceps coenurosis in Sardinian sheep. Vet Parasitol. (2007) 143:294–8. doi: 10.1016/j.vetpar.2006.08.020
35. Abbas G, Saqib M, Muhammad G, Mughal MN, Rashid I, Ullah Q. Two unusual cases of generalized Coenurus gaigeri cyst infection in Beetal goats. J Parasit Dis. (2017) 41:859–61. doi: 10.1007/s12639-017-0906-6
36. Sikandar A, Adil M, Zaneb H, Arshad M, Ali HM, Khan MA, et al. Concurrent cerebral and extra-cerebral caprine coenurosis: a case report. Pak J Life Soc Sci. (2018) 16:55–8. Available online at: http://www.pjlss.edu.pk/pdf_files/2018_1/55-58.pdf
37. Christodoulopoulos G, Dinkel A, Romig T, Ebi D, Mackenstedt U, Loos-Frank B. Cerebral and non-cerebral coenurosis: on the genotypic and phenotypic diversity of Taenia multiceps. Parasitol Res. (2016) 115:4543–58. doi: 10.1007/s00436-016-5246-4
38. Varcasia A, Lightowlers MW, Cattoli G, Cancedda GM, Canu S, Garippa G, et al. Genetic variation within Taenia multiceps in Sardinia, Western Mediterranean (Italy). Parasitol Res. (2006) 99:622–6. doi: 10.1007/s00436-006-0179-y
39. Rostami S, Salavati R, Beech RN, Sharbatkhori M, Babaei Z, Saedi S, et al. Cytochrome c oxidase subunit 1 and 12S ribosomal RNA characterization of Coenurus cerebralis from sheep in Iran. Vet Parasitol. (2013) 197:141–51. doi: 10.1016/j.vetpar.2013.07.001
40. Al-Riyami S, Ioannidou E, Koehler AV, Hussain MH, Al-Rawahi AH, Giadinis ND, et al. Genetic characterisation of Taenia multiceps cysts from ruminants in Greece. Infect Genet Evol. (2016) 38:110–6. doi: 10.1016/j.meegid.2015.12.008
41. Li WH, Jia WZ, Qu ZG, Xie ZZ, Luo JX, Yin H, et al. Molecular characterization of Taenia multiceps isolates from Gansu Province, China by sequencing of mitochondrial cytochrome C oxidase subunit 1. Korean J Parasitol. (2013) 51:197–201. doi: 10.3347/kjp.2013.51.2.197
42. Gui-Ying H, Ying-Dong Y, Ying-Zi Z, Guang-You Y. Population genetic diversity of Taenia multiceps isolated from Sichuan based on mitochondrial cox1 and cytb gene. Chinese J Anim Vet Sci. (2014) 4:631–8. Available online at: https://oversea.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFD&dbname=CJFD2014&filename=XMSY201404018&v=37rfZo%25mmd2FStUOlDbaeb42j0dHuol%25mmd2F7PvZLog8h8qxtQsaAg4NiDBGOicixQpAD0GPv
43. Gui-Ying H, Ying-Dong Y, Ying-Zi Z, Guang-You Y. Population genetic diversity of Taenia multiceps isolated from Sichuan Province based on mitochondrial nad1 and nad4 genes. Chinese Vet Sci. (2014) 3:251–7. Available online at: https://oversea.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFD&dbname=CJFD2014&filename=ZGSY201403006&v=%25mmd2BW8wDVgW9hGikjfBpH1qx6ys2OvwY2GDzGIwqWxgC%25mmd2Btn9NnCmyEP2K3kY0pRs6rP
44. Zhang Y, Zhao W, Yang D, Tian Y, Zhang W, Liu A. Genetic characterization of three mitochondrial gene sequences of goat/sheep-derived Coenurus cerebralis and cysticercus tenuicollis isolates in Inner Mongolia, China. Parasite. (2018) 25:1. doi: 10.1051/parasite/2018002
45. Oryan A, Goorgipour S, Moazeni M, Shirian S. Abattoir prevalence, organ distribution, public health and economic importance of major metacestodes in sheep, goats and cattle in Fars, southern Iran. Trop Biomed. (2012) 29:349–59. Available online at: https://pubmed.ncbi.nlm.nih.gov/23018497/
46. Biyikoglu G, Bagci O, Oncel T. A coenurosis outbreak of sheep in Istanbul, Turkey. Pendik Veteriner-Mikrobiyoloji-Dergisi. (2001) 32:27–30. Available online at: https://www.researchgate.net/publication/284676366_A_coenurosis_outbreak_of_sheep_in_Istanbul_Turkey
47. Garcia-Villar R, Toutain PL, Alvinerie M, Ruckebusch Y. The pharmacokinetics of xylazine hydrochloride: an interspecific study. J Vet Pharmacol Therap. (1981) 4:87–92. doi: 10.1111/j.1365-2885.1981.tb00715.x
48. Abrahamsen EJ. Ruminant field anaesthesia. Vet Clin Food Anim. (2008) 24:429–41. doi: 10.1016/j.cvfa.2008.07.001
49. Soulsby EJL. Helminths, Arthropods and Protozoa of Doemsticated Animals. 7th ed. London: Bailliére Tindall (1982). p. 117.
50. Faruk MO, Siddiki AMAMZ, Islam MS, Rekha A, Chowdhury S, Masuduzzaman M, et al. Identification of non-cerebral cyst: zoonotic Taenia multiceps in domestic goat in Bangladesh. Vet World. (2017) 10:1156–60. doi: 10.14202/vetworld.2017.1156-1160
51. Fu YX. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics. (1997) 147:915–25.
52. Tajima F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics. (1989) 123:585–95.
53. Excoffier L, Laval G, Schneider S. Arlequin ver. 3.0: an integrated software package for population genetics data analysis. Evol Bioinform Online. (2007) 1:47–50. doi: 10.1177/117693430500100003
54. Bandelt H, Forster P, Röhl A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. (1999) 16:37–48. doi: 10.1093/oxfordjournals.molbev.a026036
55. Vercruysse J, Claerebout E. Treatment vs non-treatment of helminth infections in cattle: defining the threshold. Vet Parasitol. (2001) 98:195–214. doi: 10.1016/S0304-4017(01)00431-9
56. Espinoza JR, Terashima A, Herrera-Velit P, Marcos LA. Fasciolosis humana y animal en el Peru: impacto en la economía de las zonas endémicas. Rev Peru Med Exp Salud Pública. (2010) 27:604–12. doi: 10.1590/S1726-46342010000400018
57. McManus DP. Molecular discrimination of taeniid cestodes. Parasitol Int. (2006) 55:31–7. doi: 10.1016/j.parint.2005.11.004
58. Zhang L, Hu M, Jones A, Allsopp BA, Beveridge I, Schindler AR, et al. Characterization of Taenia madoquae and Taenia regis from carnivores in Kenya using genetic markers in nuclear and mitochondrial DNA, and their relationships with other selected taeniids. Mol Cell Probes. (2007) 21:379–85. doi: 10.1016/j.mcp.2007.05.003
59. Yan H, Lou Z, Li L, Ni X, Guo A, Li H, et al. The nuclear 18S ribosomal RNA gene as a source of phylogenetic information in the genus Taenia. Parasitol Res. (2013) 112:1343–7. doi: 10.1007/s00436-012-3199-9
60. Alim MA, Islam MK, Rahman MM, Khan MA, Mondan MM. Further observation on metacestodiosis in black Bengal goat in Bangladesh. Pak J Sci Ind Res. (2002) 45:330–3. Available online at: https://www.researchgate.net/publication/233991952_Further_observations_on_metacestodiosis_in_Black_Bengal_goat_in_Bangladesh
61. Gosh RC, Dubey S, Mandal SC, Sharma N. Occurrence of Coenurus gaigeri cyst in a goat. Indian Vet J. (2005) 82:90–1. Available online at: https://www.researchgate.net/publication/291859616_Occurrence_of_coenurus_gaigeri_cyst_in_a_goat
62. Islam S, Kalita D, Bhuyan D, Rahman T, Saleque A. Ocular coenurosis in goat. J Vet Parasitol. (2006) 20:53–5. Available online at: https://www.indianjournals.com/ijor.aspx?target=ijor:jvp&volume=20&issue=1&article=012
63. Moghaddar N. Occurrence of Coenurus gaigeri in goats in Shirz, Iran. J Appl Anim Res. (2007) 31:83–4. doi: 10.1080/09712119.2007.9706635
64. Brown WM, George M, Wilson AC. Rapid evolution of animal mitochondrial DNA. Proc Natl Acad Sci USA. (1979) 76:1967–71. doi: 10.1073/pnas.76.4.1967
65. Mueller RL, Macey JR, Jaekel M, Wake DB, Boore JL. Morphological homoplasy, life history evolution, and historical biogeography of plethodontid salamanders inferred from complete mitochondrial genomes. Proc Natl Acad Sci USA. (2004) 101:13820–5. doi: 10.1073/pnas.0405785101
66. Shen X, Wang H, Ren J, Tian M, Wang M. The mitochondrial genome of Euphausia superba (Prydz Bay) (Crustacea: Malacostraca: Euphausiacea) reveals a novel gene arrangement and potential molecular markers. Mol Biol Rep. (2010) 37:771–84. doi: 10.1007/s11033-009-9602-7
67. Wei SJ, Tang P, Zheng LH, Shi M, Chen XX. The complete mitochondrial genome of Evania appendigaster (Hymenoptera: Evaniidae) has low A+T content and a long intergenic spacer between atp8 and atp6. Mol Biol Rep. (2010) 37:1931–42. doi: 10.1007/s11033-009-9640-1
68. Singh BB, Sharma R, Gill JP, Sharma JK. Prevalence and morphological characterisation of Cysticercus tenuicollis (Taenia hydatigena cysts) in sheep and goat from north India. J Parasit Dis. (2015) 39:80–4. doi: 10.1007/s12639-013-0284-7
69. Varcasia A, Pipia AP, Dessi G, Zidda A, Tamponi C, Pau M, et al. Morphology and genetic variability within Taenia multiceps in ruminants from Italy. Vet Parasitol. (2016) 223:181–5. doi: 10.1016/j.vetpar.2016.04.039
Keywords: Coenurus cerebralis, cox1, Taenia multiceps, genetic diversity, phylogeny, Pakistan
Citation: Alvi MA, Ohiolei JA, Saqib M, Tayyab MH, Zafar Khan MU, Li L, Aqib AI, Hassan A, Alvi AA, Qamar W, Fu B-Q, Yan H-B and Jia W-Z (2020) First Report on Molecular Characterization of Taenia multiceps Isolates From Sheep and Goats in Faisalabad, Pakistan. Front. Vet. Sci. 7:594599. doi: 10.3389/fvets.2020.594599
Received: 13 August 2020; Accepted: 19 October 2020;
Published: 10 November 2020.
Edited by:
Donato Traversa, University of Teramo, ItalyReviewed by:
Antonio Varcasia, University of Sassari, ItalyYue Xie, Sichuan Agricultural University, China
Copyright © 2020 Alvi, Ohiolei, Saqib, Tayyab, Zafar Khan, Li, Aqib, Hassan, Alvi, Qamar, Fu, Yan and Jia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wan-Zhong Jia, amlhd2FuemhvbmdAY2Fhcy5jbg==; Hong-Bin Yan, eWFuaG9uZ2JpbkBjYWFzLmNu