 José Victor Pronievicz Barreto1,2†
José Victor Pronievicz Barreto1,2† Elis Lorenzetti1,2,3†Juliana Torres Tomazi Fritzen2Andressa de Melo Jardim1
Elis Lorenzetti1,2,3†Juliana Torres Tomazi Fritzen2Andressa de Melo Jardim1 Thalita Evani Silva Oliveira4
Thalita Evani Silva Oliveira4 Selwyn Arlington Headley1,4,5
Selwyn Arlington Headley1,4,5 Amauri Alcindo Alfieri2,3,5*Luiz Fernando Coelho da Cunha Filho1
Amauri Alcindo Alfieri2,3,5*Luiz Fernando Coelho da Cunha Filho1- 1Post Graduate Program in Animal Health and Production, Department of Agrarian Sciences, Universidade Pitágoras Unopar, Arapongas, Brazil
- 2Laboratory of Animal Virology, Department of Preventive Veterinary Medicine, Universidade Estadual de Londrina, Londrina, Brazil
- 3Multi-User Animal Health Laboratory, Molecular Biology Unit, Department of Veterinary Preventive Medicine, Universidade Estadual de Londrina, Londrina, Brazil
- 4Laboratory of Animal Pathology, Department of Veterinary Preventive Medicine, Universidade Estadual de Londrina, Londrina, Brazil
- 5National Institute of Science and Technology, Dairy Production Chain (INCT-Leite), Universidade Estadual de Londrina, Londrina, Brazil
HoBi-like pestivirus (HoBiPeV) has been reported in several biological samples from cattle worldwide, but there are no descriptions of this virus associated with neurological symptoms. This report described the first occurrence of neurological disease associated with HoBiPeV in a newborn dairy calf. A mixed-breed Holstein calf had severe neurological symptoms at birth and died at 21 days old. The tissue fragments (central nervous system (CNS), myocardium, liver, kidney, lung, intestine, and spleen) were submitted to reverse transcription (RT)–PCR assay for the partial 5'-untranslated region (5'UTR) and N-terminal autoprotease (Npro) gene of the pestivirus genome, and the CNS tissue fragments were submitted to histopathological and immunohistochemical evaluation. The RT–PCR assay indicated that the kidney, CNS, and intestinal tissue fragments were positive for the pestivirus 5'UTR, and the CNS and intestinal tissue fragments were positive for the pestivirus Npro gene. Amplicons with high DNA quantification in the 5'UTR (CNS—cerebral cortex) and Npro (CNS—cerebral cortex and intestine) RT–PCR assays were sequenced. The nucleotide (nt) sequence and phylogenetic analysis of the 5'UTR strain exhibited 93.6 to 99.4%, 85%, 89.4 to 89.9%, 85.1%, and 90.5 to 91.5% nt identity with HoBiPeV strains from clades a, b, c, d, and e, respectively. The Npro amplicons showed 99.7% nt identity to each other and 90.4 to 96.5%, 85.1 to 85.3%, 79.2 to 79.7%, and 85.8 to 86.5% nt identity with HoBiPeV strains from clades a, c, d, and e, respectively. A histopathology revealed neuronal necrosis at the cerebrum, cerebellum, and brain stem. An immunohistochemical assay designed to identify antigens of bovine viral diarrhea virus revealed positive intracytoplasmic immunoreactivity within neurons at the cerebral cortex, cerebrum, cerebellum, and spinal cord. Thus, this report provides information about the first identification of HoBiPeV in tissues of the CNS in a newborn dairy calf with neurological symptoms.
Introduction
The Pestiviruses comprise a genus within the Flaviviridae family and are currently classified into 11 species (Pestivirus A to K) (1). Pestivirus A (bovine viral diarrhea virus 1—BVDV-1), Pestivirus B (BVDV-2), and Pestivirus H (HoBi-like pestivirus—HoBiPeV) are the major pestivirus species infecting cattle (1). HoBiPeV was initially detected in commercial fetal bovine serum from Brazil by Schirrmeier et al. (2), and the viral strain was named D32/00 “HoBi.”
Pestiviruses are spherical enveloped viruses with a single-stranded positive-sense RNA. The genome of 12.5 kb consists of a long and single open reading frame encoding a 3,998 amino acid polyprotein (3) that can be cleaved into 11 to 12 polypeptides: Npro, C, E0/Erns, E1, E2, p7, NS23 (NS2-3), NS4A, NS4B, NS5A, and NS5B (4). The pestivirus 5'-untranslated region (5'UTR) contains 360 to 390 nucleotides (nt) (5) and is used for genetic characterization and phylogeny (6). The N-terminal autoprotease (Npro) coding region is variable in different species and subtypes of pestiviruses (7) and is used for genotyping wild-type virus strains (8, 9). Recently, based on the genomic 5'UTR and Npro gene, the HoBiPeV strains have been classified into clades (a to e) (6, 9–11).
Infections caused by BVDV-1 and BVDV-2 have a worldwide distribution. Like HoBiPeV, they usually result in subclinical infections (12, 13) and may be associated with reproductive, respiratory, gastrointestinal, and/or hemorrhagic disorders (14). The antigenic and genomic differences of these viruses can interfere with diagnosis and vaccination (15, 16). The high mutation rate of HoBiPeV may explain the differences in its biology, virulence, and immunogenicity (17).
HoBi-like pestivirus (HoBiPeV) has been identified by reverse transcription-PCR (RT–PCR) assays performed on different types of biological material and in several countries, such as Bangladesh (18), Thailand (19, 20), India (9), and Italy (12). Occasionally, the genome of HoBiPeV was detected from the central nervous system (CNS) of a persistently infected (PI) calf with mucosal diseases-like in the absence of neurological symptoms (21); however, the association of HoBiPeV with neuropathological lesions was not confirmed by the immunohistochemical (IHC) assay.
Since the early 2000s, several studies have demonstrated the presence of HoBiPeV in biological samples from beef and dairy cattle herds from different geographic regions of Brazil, indicating that this virus may be endemic in this country (6, 13, 15, 22–26).
This case report describes the first occurrence of congenital neurological disease associated with HoBiPeV in a dairy calf.
Materials and Methods
Clinical History
A mixed-breed Holstein one-day-old calf with clinical manifestations of apathy, recumbency, and opisthotonos was examined at the Unopar Veterinary Teaching Hospital (Arapongas, Paraná State, Brazil). Clinical evaluation revealed motor incoordination, ataxia, abnormal menace response, and proprioceptive deficit, suggestive of cerebellar syndrome. However, there was no clinical improvement; the calf died suddenly after 20 days of hospitalization and was submitted for routine postmortem evaluation. There is no report of any previous disease at this herd; furthermore, cattle at this establishment were not vaccinated against infections by BVDV.
Post-mortem Evaluations
A routine postmortem evaluation was performed after death. Tissue sections (CNS, myocardium, liver, kidney, lung, intestine, and spleen) were fixed by immersion in 10% buffered formalin solution and routinely processed for histopathological evaluation with the hematoxylin and eosin staining. Duplicate tissue fragments were collected for the IHC assay, while freshly collected samples were maintained at −80°C until use in molecular assays.
Molecular Investigations
Tissue fragments (cerebral cortex, cerebellum, spinal cord, rete mirabilis, myocardium, liver, kidney, lung, intestine, and spleen) were processed with 0.01 M phosphate-buffered saline (137 mM NaCl, 3 mM KCl, 8 mM Na2HPO4, 14 mM KH2PO4; pH 7.2) in 10% suspension (w/v) and centrifuged at low speed for 5 min. The supernatant (500 μl) was recovered and pretreated at 56°C for 30 min with sodium dodecyl sulfate and proteinase K at final concentrations of 1% (v/v) and 0.2 mg/ml, respectively. The nucleic acid was extracted by combining the phenol/chloroform/isoamyl alcohol (25:24:1) and silica/guanidine isothiocyanate techniques (27). The nucleic acid was eluted in 50 μl of ultrapure diethylpyrocarbonate (DEPC)-treated water (Invitrogen Life Technologies, Carlsbad, CA, USA) and immediately stored at −80°C until use.
The extracted nucleic acid was submitted to RT–PCR assay using the panpestivirus primers (324, forward 5′ATGCCCWTAGTAGGACTAGCA3′ and 326, reverse 5′TCAACTCCATGTGCCATGTAC3′), designed to amplify a 288-base pair (bp) product from the 5′UTR of the pestivirus genome (28). Additionally, the primer pair designed to amplify a 428-bp length amplicon from the Npro gene (BD1, forward 5′TCTCTGCTGTACATGGCACATG3′ and BD3, reverse 5′CCATCTATRCACACATAAATGTGGT3′) (8) was used to confirm the results of the phylogenetic analysis based on the 5′UTR fragment. The RT and PCR assays were performed according to Lunardi et al. (29) and Vilček et al. (8) for the 5′UTR and Npro gene, respectively. Aliquots of ultrapure DEPC-treated water were included as a negative control, and the BVDV-1 cell culture-adapted NADL strain was included as the positive control. The RT–PCR products were analyzed by electrophoresis on 2% agarose gels in Tris-boric acid-ethylenediaminetetraacetic acid (EDTA) buffer, pH 8.4 (89 mM Tris; 89 mM boric acid; 2 mM EDTA), containing 0.5 μg/ml ethidium bromide. After electrophoresis at a constant voltage (100 V) for 40 min, the agarose gel was visualized under ultraviolet light.
Reverse transcription PCR (RT–PCR) amplicons were purified using the Illustra GFX PCR DNA and the Gel Band Purification Kit (GE Healthcare, Little Chalfont, Buckinghamshire, UK), quantified with a Qubit® Fluorometer (Invitrogen Life Technologies, Eugene, OR, USA), and sequenced in an ABI3500 Genetic Analyzer sequencer using the same forward and reverse primers used in the RT–PCR assay with the BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). The sequence quality analysis was carried out using the PHRED software, and contig assembly was obtained using the CAP3 software (http://asparagin.cenargen.embrapa.br/phph/). The nt sequences were compared with sequences deposited in the GenBank using the BLAST software (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Phylogenetic trees based on the nt sequences were constructed using the neighbor-joining method and the Kimura two-parameter model in the MEGA software version 7.0.26. Bootstrapping was statistically supported with 1,000 replicates. The nt sequence identity matrices were performed in the BioEdit software version 7.2.6.1.
Four other etiologic agents that can also cause neurological clinical signs in calves were investigated by PCR assays targeting the glycoprotein C gene of bovine alphaherpesvirus 1 (BoHV-1, 354 bp) and bovine alphaherpesvirus 5 (BoHV-5, 159 bp) (30); the tegument protein gene of ovine gammaherpesvirus 2 (OvHV-2, 422 bp) (31); and the pNC-5 gene of Neospora caninum (337 bp) (32).
Immunohistochemical Identification of BVDV
Formalin-fixed paraffin-embedded (FFPE) tissue fragments of the cerebral cortex, cerebellum, and spinal cord of the calf were used in an IHC assay designed to identify the antigens of BVDV using anti-BVDV #15c-5 antibody, as previously described (33). Positive controls consisted of the FFPE tissue fragments known to contain antigens of BVDV from a previous study (33). Two negative controls were used: the first consisted of substituting the primary antibodies with their respective diluents; the second consisted of utilizing the primary antibodies on FFPE tissues with known negative immunoreactivity to BVDV derived from the study cited above. Positive and negative controls were included in each IHC assay.
Results
Pathological Findings
Significant gross lesions were not observed during routine postmortem evaluations. The histopathologic findings were predominantly neurological and seemed to affect neurons within several anatomic regions of the CNS, resulting in neuronal necrosis at the cerebrum, necrosis and degeneration of Purkinje cells of the cerebellum and brain stem (Figures 1A–C), and mild neuronal necrosis at the spinal cord.
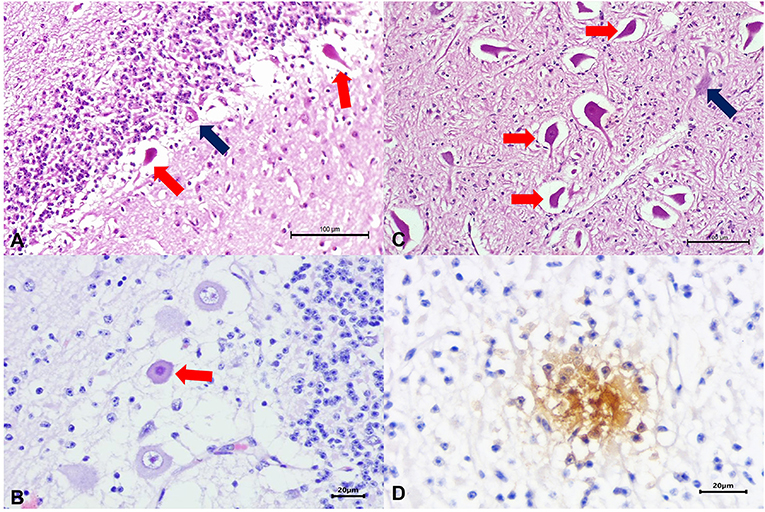
Figure 1. Histopathologic and immunohistochemical findings observed in a newborn calf naturally infected with HoBi-like pestivirus (HoBiPeV). There is necrosis of Purkinje cells at the cerebellum (A,B) and the brain stem (C); compare the severely eosinophilic and shrunken necrotic neurons (red arrows) with the adjacent normal looking neurons (blue arrows). Observed positive intracytoplasmic immunoreactivity to antigens of bovine viral diarrhea virus (BVDV) within necrotic neuron of the cerebellum (D). Hematoxylin and eosin stain (A–C); immunoperoxidase counterstained with hematoxylin (D). Scale bar A and C, 100 μm; B and D, 20 μm.
Molecular Findings
The RT–PCR assay amplified the 5′UTR of pestivirus from fragments of the cerebral cortex, cerebellum, spinal cord, rete mirabilis, kidney, and small intestine. Furthermore, the Npro gene assay was positive in all tissues evaluated in the 5′UTR assay, except fragments from the kidney.
The nucleotide sequence identity analysis of the 5′UTR revealed that the amplicon derived from this study (GenBank accession n° MZ612417) had 93.6 to 99.4%, 85%, 89.4 to 89.9%, 85.1%, and 90.5 to 91.5% nt identity with HoBiPeV strains from clades a, b, c, d, and e, respectively, and 70.6 and 75.6% nt identity with BVDV-1 (NADL strain) and with BVDV-2 (890 strain), respectively.
Additionally, the nt sequence identity analysis of the Npro amplicons (GenBank accession n° MZ612415 and MZ612416) obtained during this study had 99.7% nt identity to each other and 90.4 to 96.5%, 85.1 to 85.3%, 79.2 to 79.7%, and 85.8 to 86.5% nt identity with HoBiPeV strains from clades a, c, d, and e, respectively; and 67.3 to 67.5% and 66.3 to 66.5% nt identity with BVDV-1 (NADL strain) and with BVDV-2 (890 strain), respectively.
The phylogenetic analyses of the 5′UTR (Figure 2A) and Npro gene (Figure 2B) revealed that the wild-type strains identified in this study clustered with the HoBiPeV clade a strains.
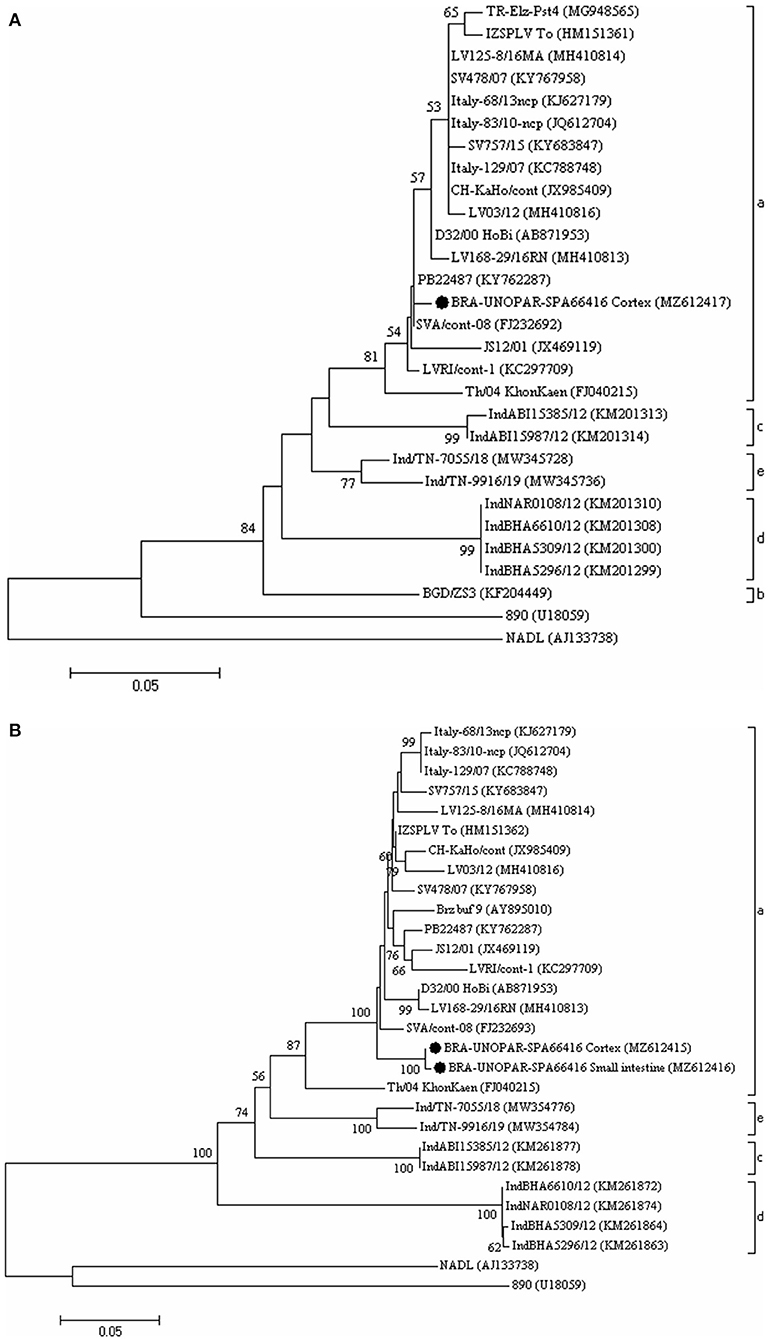
Figure 2. The phylogenetic analysis is based on the partial nucleotide (nt) sequences of the 5'-untranslated region (5'UTR) (A) and of the Npro gene (B) of HoBiPeV strains described in this study. The trees were constructed using the neighbor-joining method and the Kimura two-parameter model for nt substitution. The bootstrap values are shown at the branch nodes (values <50% are not shown). The scale bars at the bottom of the trees represent the number of nt substitutions per site. The GenBank accession numbers of the strains are provided in parentheses. The HoBiPeV strains identified during this study are indicated with a filled circle. The strains used as outgroups were the BVDV-1 prototype strain NADL and BVDV-2 prototype strain 890.
Furthermore, nucleic acids of BoHV-1, BoHV-5, OvHV-2, and N. caninum were not amplified from the tissue fragments evaluated.
Immunohistochemical Findings
Immunohistochemistry (IHC) revealed positive intracytoplasmic immunoreactivity for BVDV antigens within degenerated and necrotic neurons of the cerebral cortex, cerebellum, and spinal cord (Figure 1D).
Discussion
The clinical signs presented by the calf described in this study were strictly neurological, characterizing a congenital cerebellar syndrome, different from those observed in cattle with HoBiPeV infection (9, 11, 12, 21). There are reports of CNS infection by BVDV-1 (34) and BVDV-2 (35, 36) by transplacental transmission with neurologic symptoms in cattle. Passler et al. (37) performed an experiment in which pregnant goats were infected with BVDV-1 and BVDV-2 strains to assess transplacental transmission and the degree of damage to the fetuses. BVDV-2 caused greater damage, and IHC analyses demonstrated the presence of the virus in fetal CNS tissue. The results herein described are similar to those reported by Passler et al. (37), suggesting that HoBiPeV can also have vertical transmission, causing damage to the fetuses, since the one-day-old calf had severe neurological symptoms associated with CNS tissue infection.
HoBi-like pestivirus (HoBiPeV) was first identified in Italy in 2004 from a batch of bovine fetal serum from Brazil (strain D32/00_Hobi) (2). HoBiPeV has been identified in fragments of different organs of cattle with and without clinical signs (12, 21, 22, 24, 25). This study represents the first detection of HoBiPeV causing neurological symptoms. Although Decaro et al. (21) reported the HoBiPeV genome coding the NS2-3 polypeptide by real-time RT-PCR (qPCR) assay in the brain, cerebellum, and brain stem of a PI calf that displayed severe mucosal disease without neurological symptoms, IHC was not performed, so the association of the neuropathological lesions with the viral infection was not confirmed.
Our study detected HoBiPeV RNA in fragment tissues by RT–PCR assay for the 5′UTR and Npro gene. Cruz et al. (25) detected HoBiPeV in the tissue fragments of the spleen, lymph nodes, lung, liver, serum, and kidney using the RT–PCR assay for the 5′UTR and Npro gene from two necropsied calves from an outbreak of the mucosal disease in a Brazilian beef cattle herd. However, they did not report neurological symptoms. Similarly, other studies conducted in Brazil have also reported the presence of HoBiPeV in serum samples and fragments of organs other than the CNS of cattle (6, 13, 15, 22, 23, 25, 26, 38).
This report analyzed the 5′UTR (GenBank accession no. MZ612417) and Npro (GenBank accession no. MZ612415 and MZ612416) sequences from the HoBiPeV strains that were detected in a calf with neurological symptoms belonging to clade a. This clade was previously reported in bovine biological samples, not in CNS, based on previous studies performed in Brazil (6, 10). The HoBiPeV strains from clade b originated from Bangladesh (18), while the other three clades (c, d, and e) were described in India (9, 11). Despite the high frequency of diagnosis of HoBiPeV in Brazil, to date, only the HoBiPeV strains that belong to clade a were identified in this country.
Pestiviruses commonly cause cerebellar hypoplasia, and animals are born showing cerebellar syndrome with dysfunction to regulate and coordinate the motor activity and degeneration of Purkinje cells of the cerebellum as the main histological alteration (39). The calf from this report had no gross evidence of cerebellar hypoplasia, but presented a cerebellar syndrome due to degeneration of Purkinje cells, configuring an atypical manifestation of cerebellar syndrome caused by pestivirus. Histological findings described by Decaro et al. (21) in the brain of a PI calf with HoBiPeV that displayed severe mucosal disease were restricted to neurophagy and spongiosis of the gray cortex. These lesions were less severe than those described in this report, where the calf died shortly after showing severe and irreversible neurological symptoms.
The IHC performed by Cruz et al. (25) using the D89 monoclonal antibody showed positive immunoreactivity for BVDV for the distal limbs, gingiva, tongue, and esophagus of two PI calves by HoBiPeV. On the other hand, this report demonstrated positive immunoreactivity for antigens of BVDV in the cerebral cortex, cerebellum, and spinal cord, confirming the association of neuropathological lesions with a viral infection. Additionally, Marques et al. (24) confirmed the presence of HoBiPeV in the analysis of skin biopsies and ear notches by IHC using the same anti-BVDV #15c-5 antibody applied in this study.
The association of clinical signs, histopathologic and IHC findings, molecular detection and characterization confirmed the participation of HoBiPeV as the etiological agent associated with neurological symptoms observed in this newborn dairy calf. This report describes congenital HoBiPeV infection and contributes to the understanding of the etiopathogenesis of this infection in cattle. To the best of our knowledge, this is the first report of this pestivirus species causing neurological symptoms in a newborn calf.
Data Availability Statement
The sequences determined in this study can be found in the GenBank database under the accession numbers MZ612415 to MZ612417. https://www.ncbi.nlm.nih.gov/genbank/.
Ethics Statement
The animal study was reviewed and approved by Institutional Ethics Committee of Universidade Pitágoras Unopar (Protocol Code 023/17, June 22, 2017). All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Author Contributions
JB, EL, AA, and LC: conceptualization, validation, formal analysis, investigation, visualization, supervision, project administration, and data curation. JB, EL, JF, TO, SH, AA, and LC: methodology. EL: software. AA and LC: resources. JB, EL, JF, SH, AJ, and LC: writing—original draft preparation. JB, EL, SH, AA, and LC: writing—review and editing. AA and LC: funding acquisition. All authors have read, critically analyzed, approved the final draft of this manuscript, and have agreed to be accountable for all aspects of the study in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This research was funded by the following Brazilian institutes: National Foundation for the Development of Private Higher Education (FUNADESP), Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES), National Council of Scientific and Technological Development (CNPq), Araucaria Foundation (FAP/PR), and National Institute of Science and Technology of Dairy Production Chain (INCT-Leite) [Grant Number 465725/2014-7].
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank Dr. Eduardo Flores, from Universidade Federal de Santa Maria, for kindly providing the antibody used in the study. Part of the master's thesis of the first author JB was under the orientation of LFCCF and coorientation of EL in the Post Graduate Program in Animal Health and Production of Universidade Pitágoras Unopar.
References
1. International Committee on Taxonomy of Viruses. Master Species List (MSL36)—Update May 10th, 2021. (2020). Available online at: https://talk.ictvonline.org/files/master-species-lists/m/msl/12314 (accessed November 5, 2021).
2. Schirrmeier H, Strebelow G, Depner K, Hoffmann B, Beer M. Genetic and antigenic characterization of an atypical pestivirus isolate, a putative member of a novel pestivirus species. J Gen Virol. (2004) 85:3647–52. doi: 10.1099/vir.0.80238-0
3. Collett MS, Larson R, Gold C, Strick D, Dennis K, Anderson DK, et al. Molecular cloning and nucleotide sequence of the pestivirus bovine viral diarrhea virus. Virology. (1988) 165:191–9. doi: 10.1016/0042-6822(88)90672-1
4. Tautz N, Tews BA, Meyers G. The molecular biology of pestiviruses. Ad Virus Res. (2015) 93:47–160. doi: 10.1016/bs.aivir.2015.03.002
5. Ridpath JF, Bolin SR, Dubovi EJ. Segregation of bovine viral diarrhea virus into genotypes. Virology. (1994) 205:66–74. doi: 10.1006/viro.1994.1620
6. Monteiro FL, Martins B, Cargnelutti JF, Noll JCG, Weiblen R, Flores EF. Genetic identification of pestiviruses from beef cattle in southern Brazil. Braz J Microbiol. (2019) 50:557–63. doi: 10.1007/s42770-019-00058-6
7. Avalos-Ramirez R, Orlich M, Thiel HJ, Becher P. Evidence for the presence of two novel pestivirus species. Virology. (2001) 286:456–65. doi: 10.1006/viro.2001.1001
8. Vilček Š, Paton DJ, Durkovic B, Strojny L, Ibata G, Moussa A, et al. Bovine viral diarrhea virus genotype 1 can be separated into at least eleven genetic groups. Arch Virol. (2001) 146:99–115. doi: 10.1007/s007050170194
9. Mishra N, Rajukumar K, Pateriya A, Kumar M, Dubey P, Behera SP, et al. Identification and molecular characterization of novel and divergent Hobi-like pestiviruses from naturally infected cattle in India. Vet Microbiol. (2014) 174:239–46. doi: 10.1016/j.vetmic.2014.09.017
10. Silveira S, Cibulski SP, Junqueira DM, Mósena ACS, Weber MN, Mayer FQ, et al. Phylogenetic and evolutionary analysis of Hobi-like pestivirus: insights into origin and dispersal. Transbound Emerg Dis. (2020) 67:1909–17. doi: 10.1111/tbed.13520
11. Kalaiyarasu S, Mishra N, Jayalakshmi K, Selvaraj P, Sudhakar SB, Jhade SK, et al. Molecular characterization of recent HoBi-like pestivirus isolates from cattle showing mucosal disease-like signs in India reveals emergence of a novel genetic lineage. Transbound Emerg Dis. (2021) 21:81. doi: 10.1111/tbed.13981
12. Decaro N, Lucente MS, Mari V, Cirone F, Cordioli P, Camero M, et al. Atypical pestivirus and severe respiratory disease in calves, Europe. Emerg Infect Dis. (2021) 17:1549. doi: 10.3201/eid1708.101447
13. Weber MN, Mósena ACS, Simões SVD, Almeida LL, Pessoa CRM, Budaszewski RF, et al. Clinical presentation resembling mucosal disease associated with ‘HoBi'-like pestivirus in a field outbreak. Transbound Emerg Dis. (2014) 63:12223. doi: 10.1111/tbed.12223
14. Ridpath JF. Bovine viral diarrhea virus: global status. Vet Clin North Am Food Anim Pract. (2010) 20:26. doi: 10.1016/j.cvfa.2009.10.007
15. Cortez A, Heinemann MB, De Castro AMMG, Soares RM, Pinto AMV, Alfieri AA, et al. Genetic characterization of Brazilian bovine viral diarrhea virus isolates by partial nucleotide sequencing of the 5′-UTR region. Braz J Vet Res. (2006) 26:211–216. doi: 10.1590/S0100-736X2006000400005
16. Bauermann FV, Harmon A, Flores EF, Falkenberg SM, Reecy JM, Ridpath JF. In vitro neutralization of HoBi-like viruses by antibodies in serum of cattle immunized with inactivated or modified live vaccines of bovine viral diarrhea viruses 1 and 2. Vet Microbiol. (2013) 166:242–5. doi: 10.1016/j.vetmic.2013.04.032
17. Weber MN, Bauermann FV, Canal CW, Bayles OD, Neill JD, Ridpath JF. Temporal dynamics of ‘HoBi'-like pestivirus quasispecies in persistently infected calves generated under experimental conditions. Virus Res. (2017) 227:18. doi: 10.1016/j.virusres.2016.09.018
18. Haider N, Rahman MS, Khan SU, Mikolon A, Gurley ES, Osmani MG, et al. Identification and epidemiology of a rare HoBi-like pestivirus strain in Bangladesh. Transbound Emerg Dis. (2014) 61:193–8. doi: 10.1111/tbed.12218
19. Ståhl K, Kampa J, Alenius S, Wadman AP, Baule C, Aiumlamai S, et al. Natural infection of cattle with an atypical 'Hobi'-like pestivirus—implications for BVD control and for the safety of biological products. Vet Res. (2007) 38:517–23. doi: 10.1051/vetres:2007012
20. Liu L, Xia H, Baule C, Belák S. Maximum likelihood and bayesian analyses of a combined nucleotide sequence dataset for genetic characterization of a novel pestivirus, SVA/cont-08. Arch Virol. (2009) 154:1111–6. doi: 10.1007/s00705-009-0419-4
21. Decaro N, Lanave G, Lucente MS, Mari V, Varello K, Losurdo M, et al. Mucosal disease-like syndrome in a calf persistently infected by Hobi-like pestivirus. J Clin Microbiol. (2014) 52:2946–54. doi: 10.1128/JCM.00986-14
22. Bianchi E, Martins M, Weiblen R, Flores EF. Genotypic and antigenic profile of bovine viral diarrhea virus isolates from Rio Grande do Sul, Brazil (2000-2010). Bra J Vet Res. (2011) 31:649–55. doi: 10.1590/S0100-736X2011000800003
23. Silveira S, Weber MN, Mósena ACS, Da Silva MS, Streck AF, Pescador CA. Genetic diversity of Brazilian bovine pestiviruses detected between 1995 and 2014. Transbound Emerg Dis. (2015) 64:613–23. doi: 10.1111/tbed.12427
24. Marques ALA, Maia LA, Aguiar GMN, Weber MN, Simões SVD, Azevedo SS. Detection of ‘HoBi'-like (BVDV-3) vírus in a cow in the semiarid of Paraíba, Northeastern Brazil. Braz J Vet Res. (2016) 36:1081–6. doi: 10.1590/s0100-736x2016001100004
25. Cruz RAS, Rodrigues WB, Silveira S, Oliveira VHS, Campos CG, Leite Filho RV, et al. Mucosal disease-like lesions caused by HoBi-like pestivirus in Brazilian calves in 2010-2011: clinical, pathological, immunohistochemical, and virological characterization. Res Vet Sci. (2018) 18:116–21. doi: 10.1016/j.rvsc.2018.06.010
26. Monteiro FL, Cargnelutti JF, Braunig P, Folgueras-Flatschart AV, Santos NC, Pituco EM et al. Detection and genetic identification of pestiviruses in Brazilian lots of fetal bovine serum collected from 2006 to 2014. Braz J Vet Res. (2018) 38:387–92. doi: 10.1590/1678-5150-pvb-5283
27. Alfieri AA, Parazzi ME, Takiuchi E, Médici KC, Alfieri AF. Frequency of group A rotavirus in diarrheic calves in Brazilian cattle herds, 1998-2002. Trop Anim Health Prod. (2006) 38:521–6. doi: 10.1007/s11250-006-4349-9
28. Vilček Š, Herring AJ, Herring JA, Nettleton PF, Lowings JP, Paton DJ. Pestiviruses isolated from pigs, cattle and sheep can be allocated into at least three genogroups using polymerase chain reaction and restriction endonuclease analysis. Arch Virol. (1994) 136:309–23. doi: 10.1007/BF01321060
29. Lunardi M, Headley SA, Lisbôa JAN, Amude AM, Alfieri AA. Outbreak of acute bovine viral diarrhea in Brazilian beef cattle: clinicopathological findings and molecular characterization of a wild-type BVDV strain subtype 1b. Res Vet Sci. (2008) 85:599–604. doi: 10.1016/j.rvsc.2008.01.002
30. Claus PM, Alfieri AF, Folgueras-Flatschart AV, Wosiacki SR, Médici CK, Alfieri AA. Rapid detection and differentiation of bovine herpesvirus 1 and 5 glycoprotein C gene in clinical specimens by multiplex-PCR. J Virol Methods. (2005) 128:183–8. doi: 10.1016/j.jviromet.2005.05.001
31. Baxter SI, Pow I, Bridgen A, Reid HW. PCR detection of the sheep-associated agent of malignant catarrhal fever. Arch Virol. (1993) 132:145–59. doi: 10.1007/BF01309849
32. Müller N, Zimmermann V, Hentrich B, Gottstein B. Diagnosis of Neospora caninum and Toxoplasma gondii infection by PCR and DNA hybridization immunoassay. J Clin Microbiol. (1996) 34:2850–2852. doi: 10.1128/jcm.34.11.2850-2852.1996
33. Oliveira TES, Pelaquim IF, Flores EF, Massi RP, Valdiviezo MJJ, Pretto-Giordano LG, et al. Mycoplasma bovis and viral agents associated with the development of bovine respiratory disease in adult dairy cows. Transbound Emerg Dis. (2020) 67:82–93. doi: 10.1111/tbed.13223
34. Oem JK, Chung JY, Roh IS, Kim HR, Bae YC, Lee KH, et al. Characterization and phylogenetic analysis of bovine viral diarrhea virus in brain tissues from non-ambulatory (downer) cattle in Korea. J Vet Diagn Invest. (2010) 22:518–23. doi: 10.1177/104063871002200404
35. Blas-Machado U, Saliki JT, Duffy JC, Caseltine SL. Bovine viral diarrhea virus type 2-induced meningoencephalitis in a heifer. Vet Pathol. (2004) 41:190–4. doi: 10.1354/vp.41-2-190
36. Liebler-Tenorio EM, Ridpath JF, Neill JD. Distribution of viral antigen and tissue lesions in persistent and acute infection with the homologous strain of noncytopathic bovine viral diarrhea virus. J Vet Diagn Invest. (2004) 16:388–96. doi: 10.1177/104063870401600504
37. Passler T, Riddell KP, Edmondson MA, Chamorro MF, Neill JD, Brodersen BW., et al. Experimental infection of pregnant goats with bovine viral diarrhea virus (BVDV) 1 or 2. Vet Res. (2014) 45:38. doi: 10.1186/1297-9716-45-38
38. Cortez A, Araújo JP, Flores EF, Ribeiro MG, Megid J, Paes AC, et al. Complete genome sequence of a Hobi-like virus isolated from a Nelore cow with gastroenteric disease in the state of São Paulo, Brazil. Genome Announc. (2017) 5:17. doi: 10.1128/genomeA.00767-17
Keywords: dairy cattle, transplacental transmission, neurologic symptoms, Pestivirus H, HoBiPeV
Citation: Barreto JVP, Lorenzetti E, Fritzen JTT, Jardim AdM, Oliveira TES, Headley SA, Alfieri AA and Cunha Filho LFCd (2022) Congenital Neurological Disease Associated With HoBi-like Pestivirus Infection in a Newborn Dairy Calf From Brazil. Front. Vet. Sci. 9:852965. doi: 10.3389/fvets.2022.852965
Received: 11 January 2022; Accepted: 07 February 2022;
Published: 24 March 2022.
Edited by:
Simone Silveira, University of West of Santa Catarina, BrazilReviewed by:
José Manuel Verdes, Universidad de la República, UruguayTereza Cristina Cardoso, Universidade Estadual de São Paulo, Brazil
Copyright © 2022 Barreto, Lorenzetti, Fritzen, Jardim, Oliveira, Headley, Alfieri and Cunha Filho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amauri Alcindo Alfieri, YWxmaWVyaUB1ZWwuYnI=
†These authors have contributed equally to this work