1
The Center for Innovative Visual Rehabilitation, Boston VA Medical Center, Boston, MA, USA
2
Department of Neurosurgery, Massachusetts General Hospital, Harvard Medical School, Boston, MA, USA
3
Department of Neurophthalmology, Massachusetts Eye and Ear Infirmary, Boston, MA, USA
We are investigating the use of novel stimulus waveforms in neural prostheses to determine whether they can provide more precise control over the temporal and spatial pattern of elicited activity as compared to conventional pulsatile stimulation. To study this, we measured the response of retinal ganglion cells to both sinusoidal and white noise waveforms. The use of cell-attached and whole cell patch clamp recordings allowed the responses to be observed without significant obstruction from the stimulus artifact. Electric stimulation with sinusoids elicited robust responses. White noise analysis was used to derive the linear kernel for the ganglion cell’s spiking response as well as for the underlying excitatory currents. These results suggest that in response to electric stimulation, presynaptic retinal neurons exhibit bandpass filtering characteristics with a peak response that occurs 25 ms after onset. The experimental approach demonstrated here may be useful for studying the temporal response properties of other neurons in the CNS.
Neural prostheses generally use electric stimulation with pulse trains to modulate neural activity. However, pulses do not allow fine control over the pattern of elicited activity. For example, in retinal prostheses the incidental stimulation of axons on the retinal surface (Jensen et al., 2003
) diminishes the fidelity over the spatial pattern of activation. In addition, the temporal resolution of elicited spike trains through activation of the synaptic network with pulsatile stimulation has been quite limited (Jensen and Rizzo III, 2007
). The use of alternative stimulus waveforms to improve control over neural activation has not been well studied, possibly because the electrical artifact associated with such waveforms obscures the recorded spikes. As a result, only those spikes that appear after the electric stimulus is turned off can be studied. Here, we demonstrate that patch clamp recordings minimize the stimulus artifact, allowing the neuronal response during stimulation to be studied. We were able to observe both the spiking response and synaptic currents that were elicited by sinusoidal and white noise stimulation.
The choice of stimulus waveform for use in neural prostheses should be made with some understanding of the temporal response properties of the neurons being activated. For example, if the goal is to use sinusoidal stimulation to excite a group of neurons, it is first necessary to determine the timescale over which these neurons respond in order to guide the choice of stimulus frequency. While this could be estimated by varying the frequency of sinusoidal stimulation until the optimal frequency range is found, such an approach is time consuming, and these recordings are often difficult to maintain for extended periods of time. We present a more efficient method using white noise analysis to estimate the temporal response properties of the target neuron. Such analysis is commonly applied to study neuronal responses in the visual (Chichilnisky, 2001
) and auditory (van Dijk et al., 1997
) systems. Using electric stimulation with white noise, the linear kernel can be quickly measured, giving an estimate of the time course of the response to electric stimulation. This approach does not take into account the many nonlinear features of neuronal activation (e.g., nonlinear ion channels, rectification, response saturation). However, by minimizing the complexity of the model, the basic temporal relationship between the input and output can be quickly determined. Estimating the linear kernel with white noise is ideal for this purpose.
The use of novel stimulus waveforms has the potential to improve control over the patterns of elicited neural activation, both in terms of the temporal structure of elicited spike trains, and in the types of neurons or neuronal substructures being activated. Such techniques may have widespread application in neural prostheses, including the retina and other CNS targets.
Methods for extracting the retina and studying the response to electric stimulation have been described previously (Fried et al., 2006
). Briefly, retinas from 2.5 kg New Zealand White Rabbits were isolated and placed photoreceptor-side down in a small recording chamber. An Axopatch 200B amplifier (Axon Instruments) was used to record ganglion cell spikes from loose, cell-attached patch electrodes (4–8 MΩ) filled with Ames solution. For whole-cell recordings, the patch electrode was filled with (in mM): 113 CsMeSO4, 1 MgSO4, 7.8 × 10−3 CaCl2, 0.1 BABTA, 10 HEPES, 4 AATP-Na2, 0.5 GTP-Na3, 5 lidocaine N-ethyl bromide (QX314-BR), 7.5 neurobiotin chloride, pH 7.2. Excitatory currents were isolated by adjusting the voltage clamp to −60 mV (ECl). The retina was continuously perfused at 4 ml/min with Ames (pH 7.4) at 36°C, equilibrated with 95% O2 and 5% CO2. Electric stimulation was delivered via a 10-kΩ Platinum-Iridium electrode (MicroProbes); the exposed area was conical with an approximate height of 100 μm and base diameter of 35 μm. The height of the stimulating electrode remained fixed at 25 μm above the inner limiting membrane, which is a thin transparent membrane that comprises the inner surface of the retina. The current injected through the stimulating electrode was either a sinusoid of 5–100 Hz, or a zero-mean, pseudo-random binary noise sequence (peak-to-peak, 3.0 μA, unless noted otherwise), where the stimulus level was randomly fluctuated between −1.5 and 1.5 μA at a time resolution of 1 ms.
We used cell-attached patch clamp recordings to measure the spiking response of retinal ganglion cells to electric stimulation. We found that a relatively tight seal (∼40 MΩ) allowed large amplitude spikes to be recorded without obstruction from the stimulus artifact. Typical responses to sinusoidal stimulation and to stimulation with white noise are shown in Figures 1
A,B, respectively. Robust spiking was observed at all sinusoidal stimulus frequencies tested (5–100 Hz), however cells were most sensitive to low frequencies. In fact, some cells responded to 5–10 Hz stimulation at the lowest stimulus levels we could deliver (1 μA).
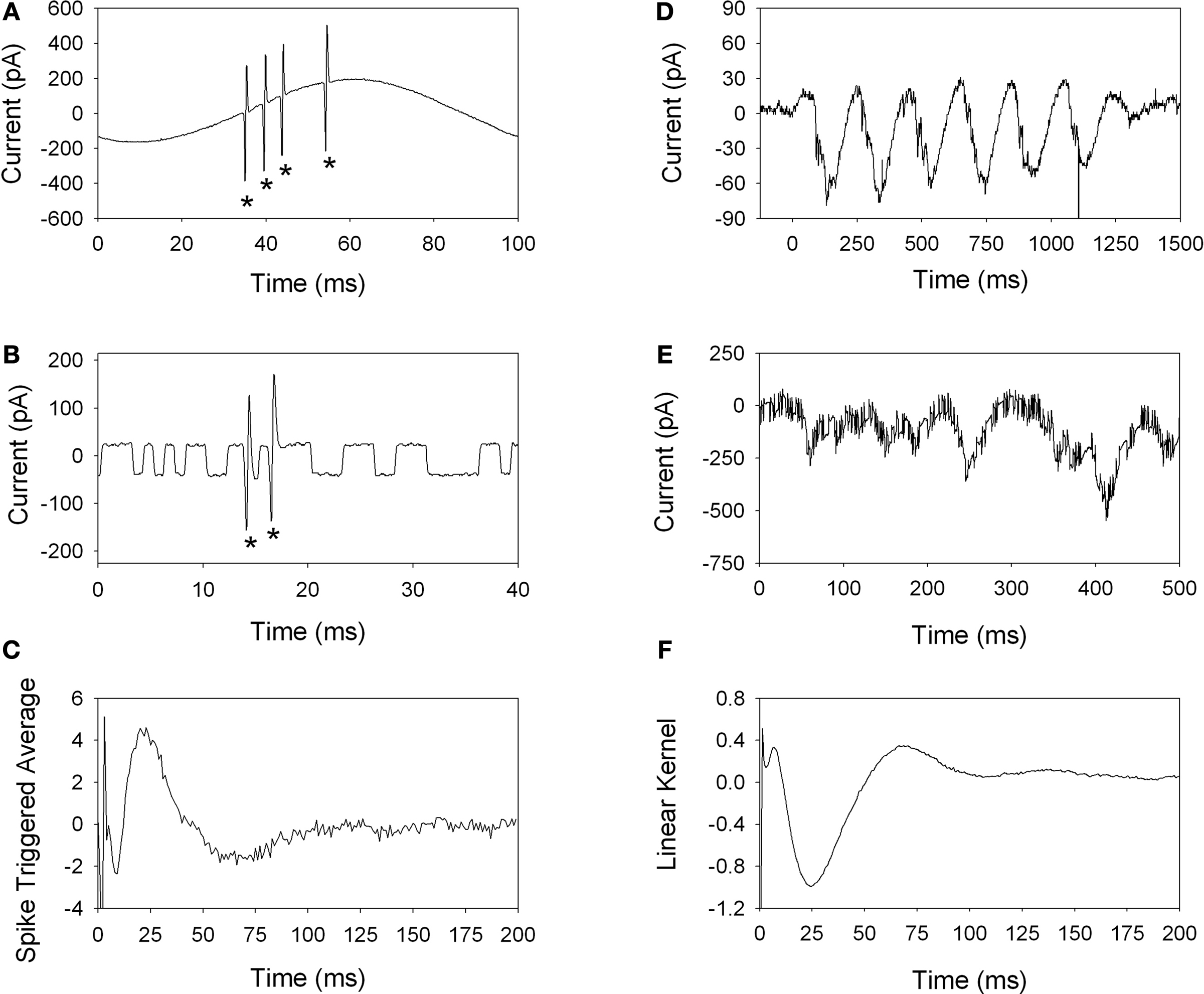
Figure 1. Spike recordings using cell-attached patch clamping allows visualization of the spikes (*) through the stimulus artifact, as shown for sinusoidal (A) and binary white noise (B) stimulation. Both stimulus waveforms elicited robust spiking responses. The spike-triggered average was estimated using 60 s of white noise stimulation (C), which is biphasic in shape with a peak near 25 ms. Excitatory currents were recorded in response to electric stimulation using sinusoids (D) and binary white noise (E). Both stimulus waveforms were highly effective at modulating the synaptic current to the ganglion cell. For stimulation with noise, the linear kernel (F) is estimated by cross-correlation of the stimulus and response.
Electric stimulation with white noise was highly effective at activating some cells, while other cells did not elicit a response even at the highest stimulus amplitude tested (peak-to-peak 4.4 μA). Following stimulation for 60 s with white noise, the spike times were extracted and the spike-triggered average (STA) was computed by averaging the stimulus waveform that preceded each spike (Rieke et al., 1997
). The STA represents the best linear description of the neuronal response to electric stimulation. A typical STA from an ON-ganglion cell is shown in Figure 1
C. The STA has two peaks (Figure 1
C): an early peak (<5 ms) and a late phase peak (∼25 ms). The late phase and overshoot of the STA were abolished following application of the synaptic blocker cadmium chloride (not shown), suggesting that the early peak of the STA is mediated by direct excitation, while the late phase is mediated by activation of presynaptic neurons.
Retinal ganglion cells receive synaptic inputs from excitatory bipolar cells and inhibitory amacrine cells. By voltage clamping the ganglion cell membrane potential at −60 mV, we isolated the excitatory currents in response to electric stimulation. Examples are shown for stimulation with sinusoidal stimulation (Figure 1
D) and stimulation with noise (Figure 1
E). By convention, inward, excitatory currents are depicted as negative, downward deflections. Consistent with the spiking response, both sinusoidal and white noise stimulation elicited robust excitatory currents in the ganglion cell.
White noise analysis was used to estimate the temporal relationship between the electric stimulus and the resulting current. The stimulus artifact is very small compared to the excitatory currents, as seen in Figure 1
E where the rapid fluctuations in the response (∼50 pA peak-to-peak) do not obstruct the input currents, which are hundreds of picoamps. After collecting 60 s of white noise data, the raw current response was cross-correlated with the stimulus waveform, giving an estimate of the linear kernel (Figure 1
F). As with the STA in Figure 1
C, the peak of the kernel occurs at about 25 ms, followed by an overshoot. The early peak (<1 ms) of the kernel likely results from a capacitive transient, which is present for whole-cell recordings since the cell membrane now separates the stimulating electrode from the recording electrode.
Here, we show that both sinusoidal and white noise stimulation elicits robust activity in retinal neurons and that these responses can be studied via use of patch clamp recordings. Our results, although preliminary, raise the possibility that each class of retinal neuron may respond optimally to different frequencies of stimulation. If so, this would be highly advantageous as it would allow very specific patterns of neural activity to be elicited. Specifically, sinusoids may have the ability to be more selective in activating certain types of neurons because the energy in a sinusoid is confined to a single frequency. Conversely, pulses have energy distributed across a wide range of frequencies, thus activating all neurons within the vicinity of the stimulating electrode regardless of their temporal response properties.
The results presented here are consistent with previous studies that revealed two potential means of eliciting a response in ganglion cells with electric stimulation: (1) direct activation of the ganglion cell, (2) activation of neurons that are presynaptic to the ganglion cells (Jensen et al., 2005
). Using white noise analysis, we showed the STA contained both a short-latency response (<1 ms), as well as a long-latency response (tens of milliseconds) for activation of neurons presynaptic to the ganglion cell. The long-latency component of the response was abolished with synaptic blockers, further confirming that this component of the STA is mediated by the activation of presynaptic neurons. The presynaptic component of the STA, as well as the linear kernel estimated with excitatory currents, can be approximated as a bandpass filter with peak response near 25 ms.
These results suggest that white noise analysis techniques can be used in conjunction with patch clamp recordings to quickly estimate the temporal relationship between the electric stimulus and the neural response. Estimating the linear approximation to the system using this method has been commonly applied to retinal ganglion cell responses using stimulation with light (Chichilnisky, 2001
; Kim and Rieke, 2001
), but has yet to be used with electric stimulation. While this linear approximation does not fully account for the many nonlinear features in the system (e.g., rectification or response saturation), the filter provides a general description of the time course of the response. Furthermore, white-noise analysis is more suited to characterize complex nonlinear systems than are pulse responses (Marmarelis and Naka, 1972
). Using white noise to measure the system dynamics is also more efficient than measuring the response to a wide range of sinusoidal frequencies and amplitudes. While much work remains in elucidating the underlying neural mechanisms responsive to pulsatile versus non-pulsatile stimulus waveforms, the methods described here provide a means to further explore these questions.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Donald Eddington for helpful discussions during the data analysis, and Neal Desai for technical assistance.