 Danilo Fernández Ríos1,2*
Danilo Fernández Ríos1,2* Nidia Benítez Candia1
Nidia Benítez Candia1 Silverio Andrés Quintana1
Silverio Andrés Quintana1 María Florencia Goberna3
María Florencia Goberna3 Eva Nara Pereira4
Eva Nara Pereira4 Andrea Alejandra Arrúa1,5
Andrea Alejandra Arrúa1,5 Andrés Castro Alegría2*
Andrés Castro Alegría2*- 1Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Asunción, San Lorenzo, Paraguay
- 2Doctorado en Ciencias Agrarias, Universidad San Carlos, Asunción, Paraguay
- 3Coordinación de Innovación y Biotecnología, Dirección Nacional de Bioeconomía, Subsecretaría de Producción Agropecuaria y Forestal, Secretaría de Agricultura, Ganadería y Pesca, Buenos Aires, Argentina
- 4Departamento de Biología Molecular y Biotecnología, Instituto de Investigaciones en Ciencias de la Salud, Universidad Nacional de Asunción, San Lorenzo, Paraguay
- 5Mycology Investigation and Safety Team, Centro Multidisciplinario de Investigaciones Tecnológicas, Universidad Nacional de Asunción, San Lorenzo, Paraguay
1 Introduction
Horizontal gene transfer (HGT) is a widespread phenomenon across all domains of life, and has been a driving force of evolution (Keeling and Palmer, 2008; Boto, 2010; Wickell and Li, 2020). Viral sequences have been found in all eukaryotic (Liu et al., 2011; Gilbert and Cordaux, 2013; Takemura, 2020) and prokaryotic kingdoms (Schleper et al., 1992; Rambo et al., 2022), and HGT has been found to occur in all directions between kingdoms of the same domain (Nelson et al., 1999; Keeling, 2009; Fuchsman et al., 2017).
Plant species have stably integrated foreign sequences into their genomes. This natural transgenesis has occurred repeatedly in the evolution of plants, affecting their biology and genetic diversification (Ma et al., 2022). Some of the mechanisms of natural HGT have been characterized to sufficient extent to be used for genetic engineering applications, and the list of mechanisms of gene transfer mastered and applied to engineering might expand with the advancement of scientific knowledge. In order to make the case that natural HGT must be taken into account when designing regulatory frameworks for transgenic organisms, and in particular of transgenic crops, we will address the particular case of HGT from bacteria to plants.
The term “transgenic” usually refers in the literature to DNA constructs resulting from the process of gene transfer between species through genetic engineering (Gordon and Ruddle, 1981; Horsch et al., 1985). Agrobacterium1-mediated transformation has established itself as the most widely used method for this purpose. In this procedure, a modified Agrobacterium plasmid transfers the desired DNA into the recipient cell, integrating it into its genome and allowing its hereditary transmission (Gelvin, 2009).
2 Horizontal gene transfer in plants
For stable incorporation of a sequence into a host organism and its transmission to offspring, certain conditions must be met. First, the foreign sequence must be integrated into the host genome. Then, the incorporated sequence must not be lost in the genomic rearrangements during cell divisions. In addition, the transformed cell must be part of the germline, to ensure inheritance. Finally, the integrated sequence must persist throughout evolution (Lacroix and Citovsky, 2016).
HGT is a process by which genes are transferred between unrelated organisms, as opposed to inheritance from parents. A clear example of gene acquisition by HGT is nitrogen fixation, a metabolic process present in certain bacteria of the genus Paenibacillus and regulated by the nif (nitrogen fixation) operon. These metabolic pathways are not specific to Paenibacillus, but have been acquired from phylogenetically distant organisms, including some of the Archaea domain and closely related bacterial phyla. HGT plays a key role in these changes, which has resulted in great diversity in the sequence and structure of nitrogen fixation regulatory elements, reflecting the multiplicity of such events from different donor organisms (Fuchsman et al., 2017). Although this phenomenon has been widely documented in Bacteria and Archaea, it has also been observed in eukaryotes, including plants (Keeling, 2024). In the latter, one of the most studied examples of HGT is the transfer of DNA from bacteria of the genus Agrobacterium to various plant species (Matveeva, 2021b).
Agrobacterium can transfer part of its DNA (T-DNA) to plant cells. Once incorporated, this T-DNA is integrated into the recipient genome, resulting in naturally occurring transgenic plants, or naturally occurring genetically modified plants (nGMs) (Matveeva and Otten, 2019). These plants have sequences in their genomes called cellular T-DNA (cT-DNA), homologous to Agrobacterium T-DNAs (White et al., 1983).
Most cT-DNAs identified to date appear to originate from Agrobacterium rhizogenes. However, cT-DNAs have also been found with previously unknown T-DNA sequences or unusual combinations thereof (Matveeva and Otten, 2019).
T-DNA sequences naturally transferred by various Agrobacterium species contain two types of genes, both regulated by promoters compatible with expression in eukaryotic cells. The first group of genes, called “oncogenes”, encodes proteins that regulate the biosynthesis or response of plant cells to phytohormones, particularly auxins and cytokinins. Their expression causes uncontrolled cell division, leading to tissue proliferation and the formation of neoplastic growths, known as crown galls (De Cleene and De Ley, 1976; Lacroix and Citovsky, 2016). The second group of genes encodes enzymes involved in the synthesis of opines that can be used by Agrobacterium cells as a source of carbon and nitrogen (Lacroix and Citovsky, 2016). It has been proposed that for the emergence of a natural transgenic plant, two conditions must be met: the naturally infected plant must be able to regenerate from tissues transformed upon infection; and the structure of the incorporated T-DNA must allow or favor such regeneration (Otten, 2016).
3 Evidence of natural transgenesis in plants
HGT in plants was initially identified in species of the genus Nicotiana, in whose genomes the presence of Agrobacterium T-DNA was detected (White et al., 1983). Studies in N. glauca and N. sylvestris showed that bacterial DNA insertion was not an isolated event (Khafizova and Matveeva, 2022).
The identification of new cT-DNA sequences in several plant species has been possible thanks to whole genome sequencing databases. The evidence suggests that HGT from bacteria to plants is a more common phenomenon than previously thought and that it has occurred in multiple plant lineages (Bogomaz et al., 2024) (Table 1).
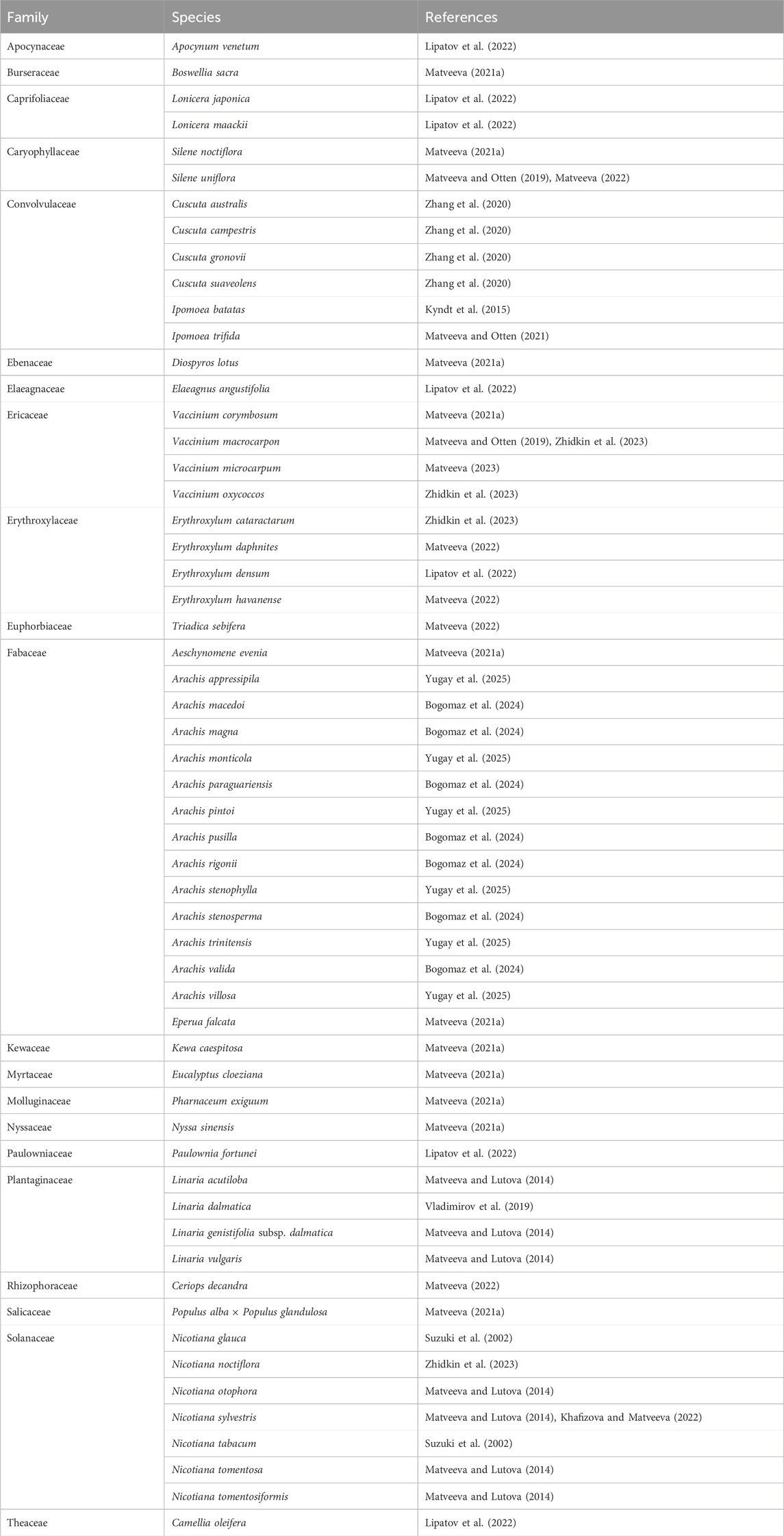
Table 1. Examples of nGM plants reported in the literature.
Genes acquired by HGT can retain their functionality in recipient plants and influence their traits. An example of this is Ipomoea batatas, where a cT-DNA has been identified with functional Agrobacterium genes which have remained stable over time (Kyndt et al., 2015). In addition, such genes can affect certain phenotypic traits, such as the rol genes, associated with root development (Quispe-Huamanquispe et al., 2017).
Unlike transgenics obtained through genetic engineering, in which genes are inserted in a targeted manner in the laboratory, nGMs acquired foreign DNA through natural infections (Chen and Otten, 2017). Between 5%–10% of dicotyledonous species are estimated to contain cT-DNAs (Matveeva, 2021b). With approximately 200 million species in this class, about 10,000 species would be nGM plants (Folta and Otten, 2021). The existence of nGM plants challenges the separation between “natural” and “artificial” made by regulatory triggers when determining which types of plants should be subjected to biosafety assessments, by showing that transgenics are not only the result of human manipulation, but also a naturally occurring phenomenon.
4 Discussion
The natural presence of Agrobacterium sequences in plant organisms questions the logic of strictly regulating transgenics obtained through genetic engineering, while exempting organisms that are similar, but obtained through conventional methods (Ammann, 2014; McHughen, 2016). cT-DNA evidence suggests that regulation focused on the method of production may be inadequate (Gould et al., 2022). In many regulatory frameworks, a transgenic organism is one that contains deliberately altered genetic material which does not occur “naturally” through breeding or selection (EFSA, 2024). This inconsistency becomes more evident when considering that the same trait can be obtained both by genetic engineering techniques and by conventional breeding, creating different regulatory thresholds for products with the same traits.
These inconsistencies also extend to relevant aspects of risk assessment, given that HGT represents an important topic in the evaluation of GM plants. In regulatory practice, HGT is typically evaluated using a pathway-to-harm approach (OECD, 2023). However, to date, no empirical evidence supports HGT from GM plants to soil bacteria under field conditions (Badosa et al., 2004; Demanèche et al., 2011; Ma et al., 2011). Similarly, while humans and animals routinely ingest DNA from multiple biological sources, the likelihood of HGT from GM plant-derived DNA to gut microbiota or host tissues remains extremely low (Jennings et al., 2003; Netherwood et al., 2004; Sieradzki et al., 2013; Korwin-Kossakowska et al., 2016). A detailed assessment of the potential for HGT from GM plants to microorganisms is beyond the scope of this work. For further information, readers are encouraged to consult Philips et al. (2022) for a detailed review.
Given these complexities, the existence of nGM plants highlights the need for a product-based regulatory trigger in which biosafety assessment focuses on the traits and phenotype of the final organism rather than the process by which it was obtained (McHughen, 2016). This approach offers several important advantages over traditional process-based frameworks, particularly in the context of emerging breeding techniques. Focusing on the characteristics and potential risks of the final product ensures regulatory coherence and risk assessment proportionality, avoiding inconsistencies where crops with similar traits are subject to different oversight (Caccamo, 2023; Brookes and Smyth, 2024). Not all GMOs pose the same level of risk; some have well-characterized, low-risk profiles; just as not all conventionally bred crops are inherently safe. Conventional methods such as wide crosses, mutagenesis, or spontaneous mutations can also result in traits with biosafety implications, including increased toxicity, allergenicity, or invasiveness (McHughen, 2016). While these products are generally not subject to a complete risk assessment, they are often regulated at various stages of the production chain (registration for the crop, safety assessment for the byproducts).
A product-based approach enables regulators to focus their efforts on the actual risk presented by a crop rather than presuming risk based on the technique employed (Sprink et al., 2016). This logic has already been adopted in the case of NBTs by countries such as Argentina, Brazil and Canada, that exclude certain products developed through NBTs from GMO regulations when no novel combination of genetic material is present in the final product (Goberna et al., 2023; Fernandes et al., 2024; Lubieniechi et al., 2025). This aligns with risk assessment principles that prioritize the traits of the crop. It also allows for the inclusion of conventionally bred crops in biosafety assessments when they present novel or potentially hazardous traits, which process-based systems tend to overlook (Gould et al., 2022). Altogether, adopting a product-based perspective would contribute to building a coherent, adaptable, and science-driven regulatory framework for novel organisms.
Author contributions
DF: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. NB: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing – review and editing. SQ: Formal Analysis, Investigation, Methodology, Writing – original draft. MG: Data curation, Methodology, Validation, Writing – review and editing. EN: Data curation, Methodology, Validation, Writing – review and editing. AA: Data curation, Methodology, Validation, Writing – review and editing. AC: Data curation, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the projects “Regulatory sciences in agricultural biotechnology” (PIC-45–2023), and “Innovation in Regulatory Science” (PIC-01–2024) from the Faculty of Exact and Natural Sciences, National University of Asunción.
Acknowledgments
The authors express their sincere appreciation to Rocío Riveros, Marcos Florentín, Samuel Gabaglio, and Vitor Pinoti for their kind contributions of scientific references.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1The collective term Agrobacterium is used in recognition of tradition and due to the impossibility of precisely identifying the bacterium responsible for the plant transformation that occurred millions of years ago. The T-DNA fragments present in plant genomes are insufficient for this determination (Matveeva, 2021a). The classification of the genus is still evolving, as the taxonomic affiliation of five Agrobacterium genomospecies has not yet been determined, suggesting a possible increase in the number of species in the future (Flores-Félix et al., 2020).
References
Ammann, K. (2014). Genomic Misconception: a fresh look at the biosafety of transgenic and conventional crops. A plea for a process agnostic regulation. New Biotechnol. 31, 1–17. doi:10.1016/j.nbt.2013.04.008
Badosa, E., Moreno, C., and Montesinos, E. (2004). Lack of detection of ampicillin resistance gene transfer from Bt176 transgenic corn to culturable bacteria under field conditions. FEMS Microbiol. Ecol. 48, 169–178. doi:10.1016/j.femsec.2004.01.005
Bogomaz, O. D., Bemova, V. D., Mirgorodskii, N. A., and Matveeva, T. (2024). Evolutionary fate of the opine synthesis genes in the Arachis L. Genomes. Biology 13, 601. doi:10.3390/biology13080601
Boto, L. (2010). Horizontal gene transfer in evolution: facts and challenges. Proc. R. Soc. B Biol. Sci. 277, 819–827. doi:10.1098/rspb.2009.1679
Brookes, G., and Smyth, S. J. (2024). Risk-appropriate regulations for gene-editing technologies. Gm. Crops Food 15, 1–14. doi:10.1080/21645698.2023.2293510
Caccamo, M. (2023). New precision-breeding law unlocks gene editing in England. Nat. Biotechnol. 41, 752–753. doi:10.1038/s41587-023-01795-8
Chen, K., and Otten, L. (2017). Natural Agrobacterium transformants: recent results and some theoretical considerations. Front. Plant Sci. 8, 1600. doi:10.3389/fpls.2017.01600
De Cleene, M., and De Ley, J. (1976). The host range of crown gall. Bot. Rev. 42, 389–466. doi:10.1007/BF02860827
Demanèche, S., Monier, J.-M., Dugat-Bony, E., and Simonet, P. (2011). Exploration of horizontal gene transfer between transplastomic tobacco and plant-associated bacteria: HGT from transgenic plants to bacteria. FEMS Microbiol. Ecol. 78, 129–136. doi:10.1111/j.1574-6941.2011.01126.x
EFSA (2024). GMO. Glossary. Available online at: https://www.efsa.europa.eu/en/glossary/gmo (Accessed February 26, 2025).
Fernandes, P. M. B., Favaratto, L., Merchán-Gaitán, J. B., Pagliarini, R. F., Zerbini, F. M., and Nepomuceno, A. L. (2024). “Regulation of CRISPR-edited plants in Latin America,” in Global regulatory outlook for CRISPRized plants (Elsevier), 197–212. doi:10.1016/B978-0-443-18444-4.00012-0
Flores-Félix, J. D., Menéndez, E., Peix, A., García-Fraile, P., and Velázquez, E. (2020). History and current taxonomic status of genus Agrobacterium. Syst. Appl. Microbiol. 43, 126046. doi:10.1016/j.syapm.2019.126046
Folta, K., and Otten, L. (2021). Widespread GMOs in nature. Colabra. Available online at: https://www.colabra.ai/podcasts/talking-biotech/294-widespread-gmos-in-nature/(Accessed March 13, 2025).
Fuchsman, C. A., Collins, R. E., Rocap, G., and Brazelton, W. J. (2017). Effect of the environment on horizontal gene transfer between bacteria and archaea. PeerJ 5, e3865. doi:10.7717/peerj.3865
Gelvin, S. B. (2009). Agrobacterium in the genomics age. Plant Physiol. 150, 1665–1676. doi:10.1104/pp.109.139873
Gilbert, C., and Cordaux, R. (2013). Horizontal transfer and evolution of prokaryote transposable elements in eukaryotes. Genome Biol. Evol. 5, 822–832. doi:10.1093/gbe/evt057
Goberna, M. F., Lewi, D. M., Godoy, P., and Hopp, E. (2023). “Regulatory framework of CRISPR-edited crops in Argentina,” in Global regulatory outlook for CRISPRized plants. Editors K. A. Abd-Elsalam, and A. Ahmad (Cambridge, Massachusetts: Elsevier Academic Press).
Gordon, J. W., and Ruddle, F. H. (1981). Integration and stable germ line transmission of genes injected into mouse pronuclei. Science 214, 1244–1246. doi:10.1126/science.6272397
Gould, F., Amasino, R. M., Brossard, D., Buell, C. R., Dixon, R. A., Falck-Zepeda, J. B., et al. (2022). Toward product-based regulation of crops. Science 377, 1051–1053. doi:10.1126/science.abo3034
Horsch, R. B., Fry, J. E., Hoffmann, N. L., Wallroth, M., Eichholtz, D., Rogers, S. G., et al. (1985). A simple and general method for transferring genes into plants. Science 227, 1229–1231. doi:10.1126/science.227.4691.1229
Jennings, J. C., Kolwyck, D. C., Kays, S. B., Whetsell, A. J., Surber, J. B., Cromwell, G. L., et al. (2003). Determining whether transgenic and endogenous plant DNA and transgenic protein are detectable in muscle from swine fed Roundup Ready soybean meal1,2,3. J. Anim. Sci. 81, 1447–1455. doi:10.2527/2003.8161447x
Keeling, P. J. (2009). Functional and ecological impacts of horizontal gene transfer in eukaryotes. Curr. Opin. Genet. Dev. 19, 613–619. doi:10.1016/j.gde.2009.10.001
Keeling, P. J. (2024). Horizontal gene transfer in eukaryotes: aligning theory with data. Nat. Rev. Genet. 25, 416–430. doi:10.1038/s41576-023-00688-5
Keeling, P. J., and Palmer, J. D. (2008). Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 9, 605–618. doi:10.1038/nrg2386
Khafizova, G. V., and Matveeva, T. (2022). Agrobacterium-mediated transformation of Nicotiana glauca and Nicotiana sylvestris. Vavilov J. Genet. Breed. 26, 697–703. doi:10.18699/VJGB-22-84
Korwin-Kossakowska, A., Sartowska, K., Tomczyk, G., Prusak, B., and Sender, G. (2016). Health status and potential uptake of transgenic DNA by Japanese quail fed diets containing genetically modified plant ingredients over 10 generations. Br. Poult. Sci. 57, 415–423. doi:10.1080/00071668.2016.1162281
Kyndt, T., Quispe, D., Zhai, H., Jarret, R., Ghislain, M., Liu, Q., et al. (2015). The genome of cultivated sweet potato contains Agrobacterium T-DNAs with expressed genes: an example of a naturally transgenic food crop. Proc. Natl. Acad. Sci. 112, 5844–5849. doi:10.1073/pnas.1419685112
Lacroix, B., and Citovsky, V. (2016). Transfer of DNA from bacteria to eukaryotes. mBio 7, e008633–e916. doi:10.1128/mBio.00863-16
Lipatov, P.Yu., Bogomaz, F. D., Gosudarev, K. D., Kondrashova, S. A., Kuchevsky, M. V., Malyuga, N. L., et al. (2022). New cellular T-DNAs in naturally transgenic plants. Ecol. Genet. 20, 40–41. doi:10.17816/ecogen112352
Liu, H., Fu, Y., Li, B., Yu, X., Xie, J., Cheng, J., et al. (2011). Widespread horizontal gene transfer from circular single-stranded DNA viruses to eukaryotic genomes. BMC Evol. Biol. 11, 276. doi:10.1186/1471-2148-11-276
Lubieniechi, S. A., Van Eenennaam, A. L., and Smyth, S. J. (2025). Regulation of animal and plant agricultural biotechnology. Trends Biotechnol. 43, 511–521. doi:10.1016/j.tibtech.2024.11.003
Ma, B. L., Blackshaw, R. E., Roy, J., and He, T. (2011). Investigation on gene transfer from genetically modified corn (Zea mays L.) plants to soil bacteria. J. Environ. Sci. Health Part B 46, 590–599. doi:10.1080/03601234.2011.586598
Ma, J., Wang, S., Zhu, X., Sun, G., Chang, G., Li, L., et al. (2022). Major episodes of horizontal gene transfer drove the evolution of land plants. Mol. Plant 15, 857–871. doi:10.1016/j.molp.2022.02.001
Matveeva, T. (2021a). New naturally transgenic plants: 2020 update. Biol. Commun. 66. doi:10.21638/spbu03.2021.105
Matveeva, T. (2021b). Why do plants need agrobacterial genes? Ecol. Genet. 19, 365–375. doi:10.17816/ecogen89905
Matveeva, T. (2022). Natural GMOs: a history of research. Ecol. Genet. 20, 7–8. doi:10.17816/ecogen112371
Matveeva, T. (2023). Prospects for the study of natural GMOs. Ecol. Genet. 21, 56. doi:10.17816/ecogen487646
Matveeva, T., and Lutova, L. A. (2014). Horizontal gene transfer from Agrobacterium to plants. Front. Plant Sci. 5, 326. doi:10.3389/fpls.2014.00326
Matveeva, T., and Otten, L. (2019). Widespread occurrence of natural genetic transformation of plants by Agrobacterium. Plant Mol. Biol. 101, 415–437. doi:10.1007/s11103-019-00913-y
Matveeva, T., and Otten, L. (2021). Opine biosynthesis in naturally transgenic plants: genes and products. Phytochemistry 189, 112813. doi:10.1016/j.phytochem.2021.112813
McHughen, A. (2016). A critical assessment of regulatory triggers for products of biotechnology: product vs. process. Gm. Crops Food 7, 125–158. doi:10.1080/21645698.2016.1228516
Nelson, K. E., Clayton, R. A., Gill, S. R., Gwinn, M. L., Dodson, R. J., Haft, D. H., et al. (1999). Evidence for lateral gene transfer between Archaea and Bacteria from genome sequence of Thermotoga maritima. Nature 399, 323–329. doi:10.1038/20601
Netherwood, T., Martín-Orúe, S. M., O’Donnell, A. G., Gockling, S., Graham, J., Mathers, J. C., et al. (2004). Assessing the survival of transgenic plant DNA in the human gastrointestinal tract. Nat. Biotechnol. 22, 204–209. doi:10.1038/nbt934
OECD (2023). “Safety assessment of transgenic organisms in the environment, volume 10,” in OECD consensus document on environmental considerations for the release of transgenic plants (Paris: OECD). doi:10.1787/62ed0e04-en
Otten, L. (2016). Functional analysis of cT-DNAs in naturally transformed plants, recent findings and general considerations. Ecol. Genet. 14, 26–31. doi:10.17816/ecogen14426-31
Philips, J. G., Martin-Avila, E., and Robold, A. V. (2022). Horizontal gene transfer from genetically modified plants - regulatory considerations. Front. Bioeng. Biotechnol. 10, 971402. doi:10.3389/fbioe.2022.971402
Quispe-Huamanquispe, D. G., Gheysen, G., and Kreuze, J. F. (2017). Horizontal gene transfer contributes to plant evolution: the case of Agrobacterium T-DNAs. Front. Plant Sci. 8, 2015. doi:10.3389/fpls.2017.02015
Rambo, I. M., Langwig, M. V., Leão, P., De Anda, V., and Baker, B. J. (2022). Genomes of six viruses that infect Asgard archaea from deep-sea sediments. Nat. Microbiol. 7, 953–961. doi:10.1038/s41564-022-01150-8
Schleper, C., Kubo, K., and Zillig, W. (1992). The particle SSV1 from the extremely thermophilic archaeon Sulfolobus is a virus: demonstration of infectivity and of transfection with viral DNA. Proc. Natl. Acad. Sci. 89, 7645–7649. doi:10.1073/pnas.89.16.7645
Sieradzki, Z., Mazur, M., Kwiatek, K., Świątkiewicz, S., Świątkiewicz, M., Koreleski, J., et al. (2013). Assessing the possibility of genetically modified DNA transfer from GM feed to broiler, laying hen, pig and calf tissues. Pol. J. Vet. Sci. 16, 435–441. doi:10.2478/pjvs-2013-0061
Sprink, T., Eriksson, D., Schiemann, J., and Hartung, F. (2016). Regulatory hurdles for genome editing: process-vs. product-based approaches in different regulatory contexts. Plant Cell Rep. 35, 1493–1506. doi:10.1007/s00299-016-1990-2
Suzuki, K., Yamashita, I., and Tanaka, N. (2002). Tobacco plants were transformed by Agrobacterium rhizogenes infection during their evolution. Plant J. 32, 775–787. doi:10.1046/j.1365-313X.2002.01468.x
Takemura, M. (2020). Medusavirus ancestor in a proto-eukaryotic cell: updating the hypothesis for the viral origin of the nucleus. Front. Microbiol. 11, 571831. doi:10.3389/fmicb.2020.571831
Vladimirov, I. A., Pavlova, O. A., Polev, D. E., and Bogomaz, D. I. (2019). cT-DNA in Linaria vulgaris L. is multicopy, inverted and homogenized. doi:10.1101/615328
White, F. F., Garfinkel, D. J., Huffman, G. A., Gordon, M. P., and Nester, E. W. (1983). Sequences homologous to Agrobacterium rhizogenes T-DNA in the genomes of uninfected plants. Nature 301, 348–350. doi:10.1038/301348a0
Wickell, D. A., and Li, F. (2020). On the evolutionary significance of horizontal gene transfers in plants. New Phytol. 225, 113–117. doi:10.1111/nph.16022
Yugay, Y., Rusapetova, T., Vasyutkina, E., Sorokina, M., Grigorchuk, V., Degtyareva, V., et al. (2025). A naturally integrated RolD-like gene in sweet potato mediates stress-responsive pathways. Plant Physiol. biochem. 223, 109875. doi:10.1016/j.plaphy.2025.109875
Zhang, Y., Wang, D., Wang, Y., Dong, H., Yuan, Y., Yang, W., et al. (2020). Parasitic plant dodder (Cuscuta spp.): a new natural Agrobacterium-to-plant horizontal gene transfer species. Sci. China Life Sci. 63, 312–316. doi:10.1007/s11427-019-1588-x
Keywords: naturally transgenic plants, horizontal gene transfer (HGT), cellular T-DNA (cT-DNA), Agrobacterium, regulatory triggers
Citation: Fernández Ríos D, Benítez Candia N, Quintana SA, Goberna MF, Nara Pereira E, Arrúa AA and Castro Alegría A (2025) Naturally transgenic plants and the need to rethink regulatory triggers in biotechnology. Front. Bioeng. Biotechnol. 13:1600610. doi: 10.3389/fbioe.2025.1600610
Received: 26 March 2025; Accepted: 16 May 2025;
Published: 30 June 2025.
Edited by:
Monica Garcia-Alonso, Estel Consult Ltd., United KingdomReviewed by:
Paul Keese, University of Ghana, GhanaCopyright © 2025 Fernández Ríos, Benítez Candia, Quintana, Goberna, Nara Pereira, Arrúa and Castro Alegría. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Danilo Fernández Ríos, ZGZlcm5hbmRlekBmYWNlbi51bmEucHk=; Andrés Castro Alegría, YW5kcmVzLmNhc3Ryb0B1c2MuZWR1LnB5