 Shunhong Zhang1
Shunhong Zhang1 Jun Jiang1Yi Luo1Guangyue Liu2
Jun Jiang1Yi Luo1Guangyue Liu2 Saidi Hu3Siran Wan4Chenchen Luo5Hong Li6Nian Li7Jailson da Graça Espírito Santo Vasconcelos8Leonilde Lavres Ceita de Carvalho8Monazeri Lima Bragança da Costa9José Etchu Takounjou8
Saidi Hu3Siran Wan4Chenchen Luo5Hong Li6Nian Li7Jailson da Graça Espírito Santo Vasconcelos8Leonilde Lavres Ceita de Carvalho8Monazeri Lima Bragança da Costa9José Etchu Takounjou8 Karem Maimite Das Neves8Luzimery dos Ramos da Conceição8Marinela da Costa Encarnação8
Karem Maimite Das Neves8Luzimery dos Ramos da Conceição8Marinela da Costa Encarnação8 Lin-Yong Zhao10*†
Lin-Yong Zhao10*†
- 1Department of Cardiology, Pangang Group General Hospital, Panzhihua, China
- 2Department of Anesthesiology, West China Hospital, Sichuan University, Chengdu, China
- 3Department of Stomatology, Yaan People’s Hospital, Yaan, China
- 4Department of Gynaecology and Obstetrics, Yaan People’s Hospital, Yaan, China
- 5Department of Outpatient Chengbei, The Affiliated Stomatological Hospital, Southwest Medical University, Luzhou, China
- 6Department of Ultrasound Medicine, Panzhihua Women & Enfants Healthcare Hospital, Panzhihua, China
- 7Department of Traditional Chinese Medicine, Panzhihua Central Hospital, Panzhihua, China
- 8Department of General Surgery, Hospital Dr. Ayres de Menezes, Sao Tome, Sao Tome and Principe
- 9Department of Stomatology, Delegation of Health of Sao Tome and Principe, Sao Tome, Sao Tome and Principe
- 10Department of General Surgery & Laboratory of Gastric Cancer, State Key Laboratory of Biotherapy/Collaborative Innovation Center of Biotherapy and Cancer Center, West China Hospital, Sichuan University, Chengdu, China
The escalating recognition of perivascular adipose tissue (PVAT) as a molecular nexus in cardiovascular disease (CVD) pathogenesis necessitates a comprehensive synthesis of its spatiotemporal dynamics and therapeutic potential. This review synthesizes PVAT's roles in vascular inflammation, metabolic dysregulation, and emerging diagnostic strategies, emphasizing molecular cross-talk and spatial heterogeneity. We explore PVAT's molecular interactions in obesity, diabetes, and hypertension, elucidating its contribution to inflammation, oxidative stress, and endothelial dysfunction. Advanced imaging techniques, notably the perivascular fat attenuation index (FAI) and circulating biomarkers, are highlighted for early CVD detection. Novel therapeutic strategies, including lifestyle modifications, pharmacological interventions, and gut microbiota modulation, are discussed. Finally, we emphasize multi-omics approaches and propose a roadmap bridging basic and clinical research to advance PVAT-based CVD management.
1 Introduction
Perivascular adipose tissue (PVAT) is of paramount importance as it closely surrounds blood vessels and releases a variety of active substances through paracrine and autocrine mechanisms (1, 2). PVAT is essential in the complex process of regulating vascular tension, which is vital for the proper functioning of the cardiovascular system. By maintaining the elasticity of blood vessels, PVAT ensures that they can efficiently accommodate the flow of blood, adapting to changes in pressure and volume (3). Furthermore, PVAT is involved in the intricate processes of lipid and energy metabolism, highlighting its importance in the body's overall metabolic functions (4). In addition to these roles, PVAT also plays a significant part in regulating inflammation and immune responses, demonstrating its multifaceted contributions to both vascular health and systemic metabolism (5).
Conversely, under pathological conditions such as obesity and diabetes, PVAT function becomes impaired. The secretion of active substances becomes unbalanced, leading to excessive release of inflammatory factors and disordered immune regulation (6, 7). This dysfunction contributes to impaired vascular endothelial function, accelerated atherosclerosis, and a series of vascular pathologies, thereby becoming a significant driving factor in the onset and progression of cardiovascular diseases (CVDs), which profoundly impacts overall health and disease processes (8–10). The recently discovered role of PVAT as a modulator of circadian rhythms—via Bmal1-dependent secretion of heme-binding protein—links nocturnal blood pressure dipping to adipose redox cycles (11), thereby offering mechanistic insights into cardiovascular events related to morning surges.
Although the significance of PVAT in both cardiovascular physiological and pathological processes has gained increasing recognition; however, current research in this area continues to encounter several limitations. This review challenges three prevailing assumptions: (1) Homogeneity Fallacy: PVAT is not a monolithic entity; rather, it represents a mosaic of adipocyte subtypes and immune niches characterized by distinct spatial metabolic programming. (2) Paracrine-Centric View: Mechanotransduction through the extracellular matrix-integrin signaling of PVAT may be as significant as soluble factors in contributing to vascular stiffening. (3) Therapeutic Neutrality: The PVAT-specific effects of current anti-diabetic drugs remain unquantified, which poses a risk of off-target consequences. By integrating single-cell omics, advanced imaging techniques, and data from interventional trials, we propose a novel framework for PVAT-centric cardiovascular therapeutics—one that emphasizes depot-specific targeting, optimization of circadian rhythms, and modulation of the microbiome-adipose axis.
2 Paracrine effects and bi-directional cross-talk of PVAT in cardiovascular function
2.1 Anatomical location and cellular composition
Anatomically, PVAT surrounds most large blood vessels, excluding the pulmonary and brain vasculature and the microcirculation (12, 13). PVAT is specifically designated as adipose tissue that resides within a radial distance equivalent to the vessel's own width, measured from the outer wall of a blood vessel. When dealing with vessels exceeding a diameter of 2 cm, notably the aorta, PVAT can expand up to a maximum of 2 cm away from the vessel wall (1). This tissue is intimately associated with blood vessels, forming a distinct layer encircling the adventitia. Its distribution patterns vary considerably, contingent upon the vessel's location and size. Notably, in larger arteries like the aorta, PVAT can constitute a substantial depot, whereas in smaller vessels, it may manifest as a thinner, more delicate layer. In rodents, the PVAT surrounding the thoracic aorta consists of brown adipose tissue (BAT), whereas the PVAT around the abdominal aorta is made up of a combination of white adipose tissue (WAT) and a portion of BAT (14, 15). In humans, the phenotype of thoracic aortic PVAT is characterized as brown. In the case of neonates and during the early developmental stages, human epicardial adipose tissue (EAT) exhibits both morphological and functional characteristics that closely resemble those of BAT. This similarity is particularly significant in the context of early life. However, under typical physiological conditions, the brown fat-like attributes of EAT diminish considerably as a person ages, transitioning from childhood into adulthood. This decline highlights the dynamic nature of PVAT and its changing roles throughout the life course (16, 17).
Cellularly, PVAT harbors adipocytes, macrophages, neutrophils, dendritic cells, mast cells, eosinophils, T cells, B cells, and various other cell types. These cells interact to maintain the functionality and microenvironmental homeostasis of PVAT (18) (Figure 1A). The adipocytes can be white, brown, or beige, each with unique characteristics and functions, BAT and beige fat function differently than WAT (Table 1). While WAT mainly acts as an energy reservoir, accumulating surplus chemical energy in the form of triacylglycerol (TAG), both BAT and beige fat are noted for their elevated metabolic activity, employing chemical energy for the generation of heat (24). The thermogenic capacity of BAT and beige fat plays an essential role in sustaining body temperature regulation, achieving energy equilibrium, and managing body weight (25). PVAT's role transcends its anatomical proximity to vasculature, functioning as a dynamic signaling interface with depot-specific molecular signatures.
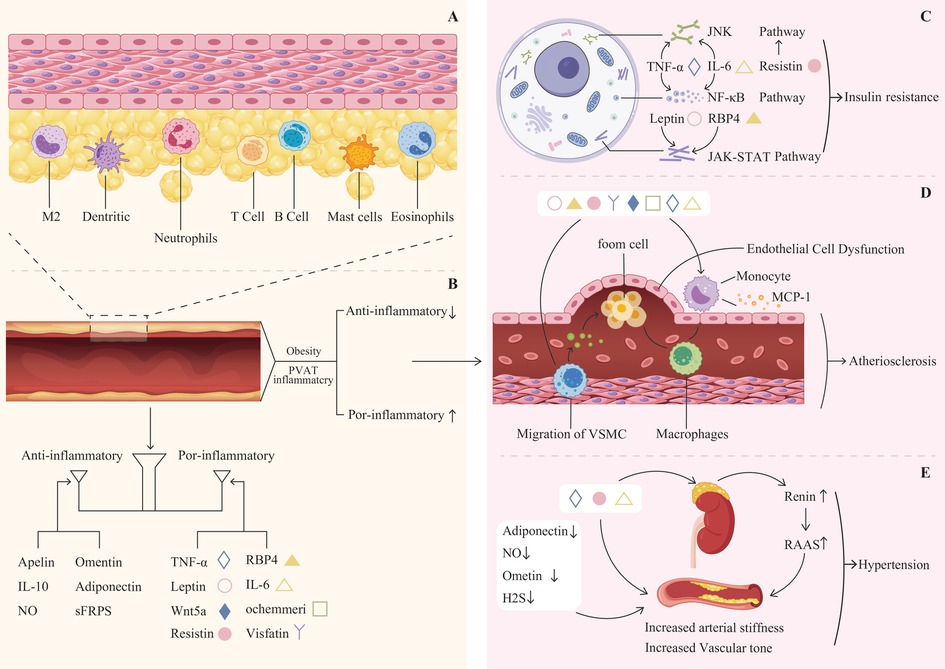
Figure 1. (A) The cellular composition of perivascular adipose tissue (PVAT) includes adipocytes, macrophages(M2), neutrophils, dendritic cells, mast cells, eosinophils, T cells, B cells, and various other cell types. (B) Balance of anti-inflammatory and pro-inflammatory adipokines. (C) Leptin, resistin, RBP4, TNF-αand IL-6 can activate the JNK, NF-κB, and JAK-STAT pathways, leading to insulin resistance. (D) Pro-inflammatory factors contribute to endothelial cell dysfunction, as well as the migration of vascular smooth muscle cells and monocytes, ultimately leading to atherosclerosis. (E) The reduction of adiponectin, Ometin, and H2S, alongside the increase in TNF-α, IL-6, and resistin, contributes to increased arterial stiffness and vascular tone. This biochemical imbalance promotes the secretion of renin and activates the RAS system, ultimately resulting in hypertension.
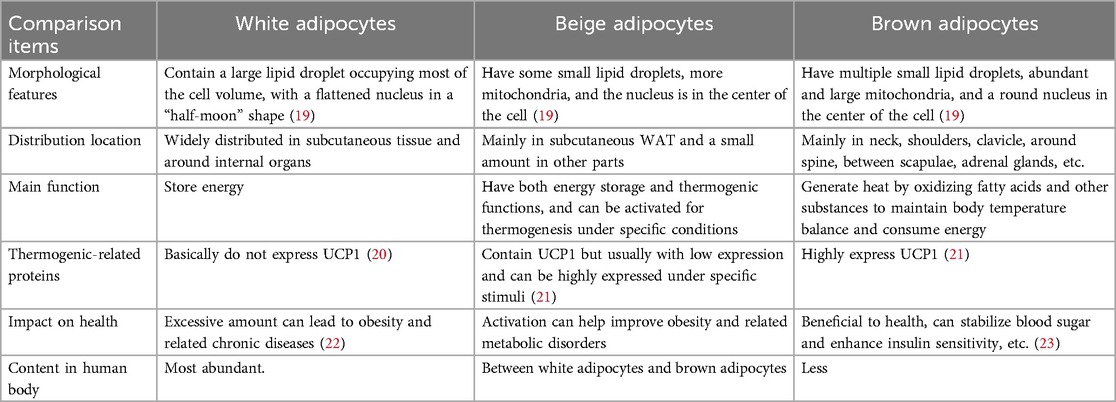
Table 1. Distribution and functionality of white, beige, and brown adipocytes.
Recent single-nucleus RNA sequencing (snRNA-seq) analyses reveal three distinct adipocyte subpopulations in human thoracic PVAT: uncoupling protein 1(UCP1) thermogenic, ADIPOQ endocrine, and fibro-inflammatory adipocytes—each exhibiting unique vascular crosstalk patterns (24, 26–30). Notably, In murine models, periaortic PVAT demonstrates higher mitochondrial density compared to mesenteric depots, correlating with enhanced fatty acid oxidation and reactive oxygen species (ROS) buffering capacity (31). This spatial metabolic specialization challenges the traditional dichotomy between white and brown adipocytes, suggesting that PVAT exists as a continuum of plastic phenotypes. Furthermore, it contests the conventional viewpoint of the “Homogeneity Fallacy” regarding PVAT.
2.2 PVAT in normal vascular physiology: paracrine and endocrine effects
PVAT releases a variety of bioactive molecules, including adipokines (32), cytokines (33), and growth factors (34). Studies conducted in animal models have demonstrated that these molecules can influence vascular function through endocrine or paracrine mechanisms. Under normal physiological circumstances, PVAT has a net effect of vasodilation, antioxidation, and anti-inflammation on the blood vessels. The vasodilatory function is influenced by the secretion of various factors from adipocytes, which are specialized cells in adipose tissue. Among these factors are adiponectin (35), apelin (36), leptin (37), and omentin (38), each playing a significant role in the regulation of vascular tone. These bioactive compounds contribute to vasodilation through two primary mechanisms. Secondly, they may indirectly promote the production of endothelium-derived vasodilators, including nitric oxide (NO) and the endothelium-derived hyperpolarizing factor. This results in enhanced blood circulation and improved vascular performance. Animal experiments have confirmed this effect (39). Interestingly, differences have been observed among various ethnic groups (40). This intricate interplay underscores the importance of adipocyte-derived factors in maintaining vascular health and regulating blood pressure. The paracrine functions of adipocytokines released from PVAT exhibit significant antioxidant and anti-inflammatory properties. These effects may be mediated by activating the 5′adenosine monophosphate-activated protein kinase (AMPK) and NO signaling pathways in nearby blood vessels. In mouse experiments, the downregulation of AMPK inhibits the synthesis of NO, which lead to a reduction in the anti-contraction activity PVAT and a decrease in the BAT phenotype (31). This interaction suggests that the release of adipokines from PVAT not only influences the local vascular environment but also plays a crucial role in modulating vascular health. By engaging these signaling mechanisms, adipokines contribute to the maintenance of vascular homeostasis, potentially offering protective benefits against inflammatory processes and oxidative stress in the surrounding tissues (41).
PVAT is crucial in the intricate two-way communication with the vascular wall (Table 2). Through this interaction, PVAT can react to paracrine signals released by the vascular cells, resulting in phenotypic alterations in the adipocytes located within the PVAT. As a result of these alterations, the composition of the secretory products released by PVAT is modified. These secretory products subsequently exert paracrine effects on the vascular wall itself, influencing its function and behavior (18, 60). It plays a crucial role in maintaining vascular health by secreting a variety of vasoactive factors. These factors encompass a diverse range of substances, including (61–63). Through the release of these biologically active molecules, PVAT contributes to the modulation of vascular functions, which can influence processes such as blood flow and vascular tone (18, 64). Under conditions of heightened oxidative stress in the vascular system, substances resulting from lipid peroxidation, such as 4-hydroxynonenal (4-HNE), are emitted by the walls of blood vessels (65). The substances infiltrate the adjacent PVAT, subsequently activating signaling pathways associated with peroxisome proliferator-activated receptor (PPAR)-γ in the perivascular adipocytes. This activation leads to elevated levels of adiponectin expression and release from PVAT, an essential factor for diminishing oxidative stress in blood vessels, acting as a protective mechanism against oxidative damage in the vascular environment (66, 67).
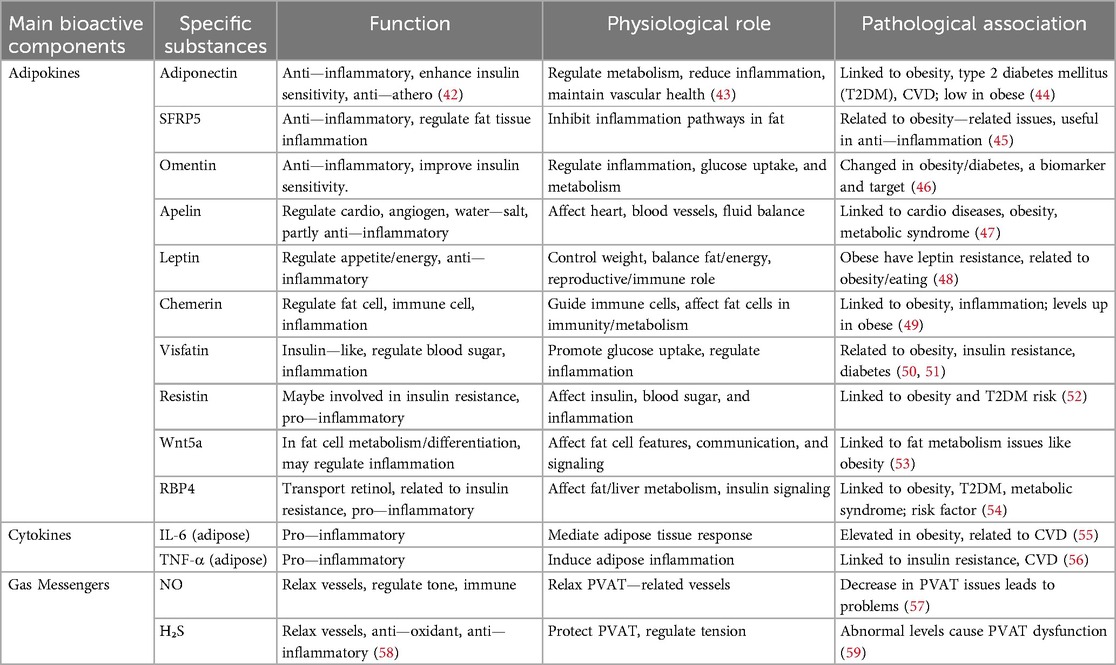
Table 2. Main bioactive components in PVAT: biological functions in the vascular bed.
The balance between adipokines may be crucial in determining the overall impact of PVAT on cardiovascular health (Figure 1B). Understanding the complex interactions among these factors could yield new insights into the pathophysiology of obesity-related CVD and inform the development of novel treatment strategies. Adiponectin, a vasodilatory adipokine primarily secreted by adipocytes, plays an important role in the anti-contractile function of PVAT and is significant for regulating insulin sensitivity and glucose metabolism (68). In instances of obesity, the levels of plasma adiponectin decrease, which aids in the progression of hypertension (69), atherosclerosis (70), and diabetes (71). Leptin is another prominent adipokine that plays a crucial role in regulating appetite and body weight (72). In instances of obesity, there is an increase in leptin secretion; nonetheless, it is common for individuals to develop resistance to leptin (73). Leptin is capable of promoting vasodilation via mechanisms that rely on the endothelium as well as those that do not. In contrast, consistently elevated leptin levels may contribute to vasoconstriction (74, 75). Additionally, high levels of leptin have been associated with obesity-related conditions, including myocardial infarction (76, 77) and stroke (78). Apelin expression increases in obesity and interacts with the abnormal function of PVAT during the onset and progression of CVD, such as hypertension (79) and atherosclerosis (80). Visfatin is highly secreted by VAT and PVAT, and it appears to play a role in atherosclerosis while being associated with a vasopressor effect (81). Omentin is an adipokine that possesses both anti-contractile and anti-inflammatory characteristics. It enhances the bioavailability of NO, exhibits antioxidant properties, and provides protective effects against ischemic stroke by improving atherosclerosis (82, 83).
In addition to adipokines, PVAT also secretes various other factors, including cytokines, growth factors, and gaseous messengers. These factors can influence vascular function and play a role in the pathophysiology of CVDs. For instance, interleukin-6 (IL-6) (84) and tumor necrosis factor-alpha (TNF-α) (85) are pro-inflammatory cytokines released from PVAT in conditions such as obesity. Vascular inflammation, oxidative stress, and dysfunction of the endothelium can be initiated by these cytokines, which, in turn, play a role in the progression of CVDs (86). Additionally, hydrogen sulfide (H₂S), a gaseous messenger released from PVAT, exhibits antioxidant and vasodilatory effects under physiological conditions (87). However, in obesity and other pathological states, the production of H₂S may be altered, resulting in modifications to vascular function (88, 89). NO is a well-recognized vasodilator that plays a significant role in mediating the anti-contractile effect in PVAT (90). Initially, obesity may induce an adaptive increase in NO levels; however, chronic obesity ultimately results in diminished NO bioavailability (91). NO-dependent endothelial dysfunction is a critical initial step in the development of CVDs, particularly in the pathogenesis of atherosclerosis and hypertension (92). Furthermore, NO dysfunction also impacts myocardial remodeling (93). The above conclusions are mainly based on the results of a large number of animal experiments.
Recent studies reveal that PVAT transfers functional mitochondria to endothelial cells via extracellular vesicles (EVs), primarily exosomes, thereby enhancing vascular bioenergetics and redox balance (94–96). This intercellular mitochondrial transfer is mediated through exosomal packaging mechanisms regulated by the Endosomal Sorting Complex Required for Transport (ESCRT) and Rab GTPase proteins, which facilitate the formation of multivesicular bodies (MVBs) and subsequent vesicle docking to recipient cells (97). Exosomes selectively encapsulate mitochondria via “mitochondrial sorting” signals involving syntaxin-17 and HSP90, ensuring functional organelle delivery (98). However, the depot-specific efficiency, with thoracic PVAT exhibiting greater efficacy than abdominal PVAT, may arise from higher exosomal yield and enhanced mitochondrial quality control (e.g., PINK1/Parkin-mediated mitophagy) in thoracic depots (31). Notably, obesity disrupts this process by inducing mitochondrial dysfunction (e.g., reduced oxidative phosphorylation and mtDNA mutations) in transferred organelles, while concurrently suppressing protective adipokines such as Neuregulin-4 (Nrg4) (99). Nrg4, secreted by adipocytes, attenuates metabolic inflammation by repressing pro-inflammatory macrophage polarization and restoring T cell function, thereby preserving PVAT homeostasis and exosomal mitochondrial integrity (100). These findings challenge the “Paracrine-Centric View” associated with PVAT and highlight Nrg4 agonism as a potential strategy to counteract obesity-induced mitochondrial transfer deficits.
2.3 Immune and inflammatory changes of PVAT in pathological states
2.3.1 Obesity-induced PVAT remodeling and immune dysregulation
Obesity profoundly alters the phenotype of PVAT, characterized by white adipocyte hypertrophy and hyperplasia within this depot (101–103). Excess body weight may also promote the conversion of aortic BAT into a WAT-like phenotype (104). These adipocyte changes are metabolically significant, as hypertrophy is linked to increased inflammation and insulin resistance (105). Critically, obesity drives remarkable shifts in the immune cell landscape of PVAT. Populations of macrophages, neutrophils, dendritic cells, eosinophils, natural killer cells, B cells, and T cells exhibit altered abundance and function (2). Macrophages, the most extensively studied immune component, increase in number and activation state (polarization) during obesity, with a distinct shift towards pro-inflammatory M1 over anti-inflammatory M2 polarization (106). This M1 dominance is associated with insulin resistance and hypertension (103). Additional immune cells also play a role in the inflammatory response within PVAT, though they do so via different mechanisms. For example, Neutrophils contribute to early macrophage polarization through the release of CCL2 and TNF in response to high-fat diets (107). Dendritic cells are found in minimal quantities within the lean phenotype but are elevated in specific adipose regions when exposed to a high-fat diet (107, 108). Additionally, mast cells in adipose tissue demonstrate an increase during obesity, although not as prominently as macrophages (109, 110). In contrast, eosinophils show a decline in obesity, and their emerging function as potential key regulators of metabolic stability is significant (111, 112). The combined populations of B (113, 114) and T cells (115, 116) make up the second largest group of immune cells located in adipose tissue, following the macrophages found within this tissue. In the context of obesity, there is a greater overall presence of T cells, with their expression varying by depot (117). This dysfunctional state of PVAT contributes to increased vascular tone, vessel stiffness, activation of the renin-angiotensin-aldosterone system (RAAS), and diminished anti-contractile effects, promoting peripheral resistance and hypertension (118, 119).
2.3.2 Reciprocal interactions between diabetes and PVAT
A complex bidirectional relationship exists between diabetes and PVAT dysfunction. Hyperglycemia induces systemic oxidative stress and the overproduction of ROS (120, 121), which activate inflammatory pathways such as NF-κB. This activation triggers the infiltration of inflammatory cells into PVAT and the release of cytokines, including IL-6 and TNF-α (121, 122), establishing a chronic inflammatory state. Concurrently, dysfunction in PVAT adipocytes manifests as increased lipolysis and the release of free fatty acids, which induce lipotoxicity (123). While also reducing the secretion of the beneficial adipokine adiponectin. This deficiency in adiponectin impairs vascular function by decreasing NO bioavailability and increasing endothelin-1, thereby disrupting the balance of vascular tone (124). Conversely, dysfunctional PVAT actively contributes to the pathogenesis of diabetes. Disrupted adipokine secretion (e.g., decreased adiponectin and elevated levels of leptin, resistin, retinol binding protein 4, TNF-α, and IL-6) impairs insulin sensitivity in peripheral tissues. Inflammatory factors such as TNF-α and IL-6 interfere with insulin signaling by activating serine kinases (e.g., via JNK, NF-κB, and JAK-STAT pathways) (Figure 1C), leading to the phosphorylation of insulin receptor substrates and the blockade of signal transduction (125–127). Furthermore, vascular lesions driven by PVAT can compromise tissue perfusion, hindering the systemic delivery of insulin and glucose.
2.3.3 The role of PVAT in atherosclerosis development
PVAT has a pivotal role in the development of atherosclerosis, a condition characterized by the thickening and hardening of arterial walls. This tissue functions by secreting various adipokines and inflammatory mediators, including IL-6 and TNF-α. These substances activate inflammatory signaling pathways within both vascular endothelial cells and vascular smooth muscle cells (128). For instance, upon stimulation by tumor necrosis factor-α, endothelial cells activate the nuclear factor-κB signaling pathway, leading to increased expression of intercellular adhesion molecule-1 and vascular cell adhesion molecule-1. This enhances the adhesion of inflammatory cells to the endothelial surface of blood vessels (129). Concurrently, inflammatory mediators induce vascular endothelial cells to secrete chemokines, including monocyte chemoattractant protein-1. Guided by the gradient of these chemokines, monocytes and other inflammatory cells traverse the gaps between endothelial cells and migrate into the subintima (130, 131). Additionally, PVAT serves as a significant source of ROS. The substantial production of ROS can oxidatively modify low-density lipoprotein, resulting in the formation of oxidized low-density lipoprotein, which impairs the relaxation function of endothelial cells (132). Ultimately resulting in thickening of the blood vessel wall and narrowing of the lumen, thereby effectively promoting the development of atherosclerosis (Figure 1D).
2.3.4 Relationship between hypertension and PVAT function
Hypertension exerts significant structural and functional impacts on PVAT. Chronically elevated pressure stimulates the differentiation of precursor cells into adipocytes and promotes lipid accumulation in existing adipocytes (133). Additionally, hypertension remodels the PVAT extracellular matrix by increasing collagen and fibronectin deposition, which disrupts local substance exchange and signaling. Additionally, hypertension alters the extracellular matrix components of PVAT, leading to increased deposition of collagen and fibronectin. These structural modifications disrupt the exchange of substances and signal transduction within PVAT, thereby impairing its normal physiological function (134, 135). Functionally, the hemodynamic alterations and heightened oxidative stress induced by hypertension contribute to PVAT dysfunction. Oxidative stress generates substantial amounts of ROS, which can damage cellular components in PVAT. Furthermore, hypertension disrupts the balance of vasoactive substances and inflammatory factors secreted by PVAT, transitioning it from a normal physiological regulatory state to a pathological state that promotes vasoconstriction and inflammation, further aggravating hypertension (136, 137). Reciprocally, PVAT critically influences blood pressure regulation and becomes a key site for immune cell infiltration in hypertension (138). Infiltrating immune cells, such as T cells, release effector cytokines—including interferon-gamma (IFN-γ), interleukin-17 (IL-17), TNF-α, and IL-6 which initiate biological cascades that contribute to vascular dysfunction, heightened oxidative stress, and increased vascular stiffness. These PVAT-driven inflammatory and vascular responses significantly contribute to the pathogenesis of hypertension and its complications (139, 140) (Figure 1E).
3 PVAT as a source of biomarkers
3.1 Advancements in imaging techniques for assessing PVAT inflammation
Computed Tomography (CT) imaging has established itself as the benchmark for effectively visualizing and characterizing PVAT (141). This is largely attributed to its exceptional spatial resolution, which allows for detailed imaging of structures within the body. Moreover, the distinct attenuation signals generated by adipose tissue enhance the accuracy and clarity of the images obtained. As a result, CT Imaging proves to be an invaluable tool in the study and analysis of PVAT, facilitating better understanding and insights into its role in various physiological and pathological conditions (142). The fat attenuation index (FAI) serves as a measure aimed at quantifying the three-dimensional attenuation gradients identified in coronary computed tomography angiography (CCTA) imaging of PVAT (141). FAI has received both biological and clinical validation, and it is now acknowledged as the main imaging biomarker for coronary inflammation sourced from CCTA. Standardized FAI evaluations are capable of accounting for side branches, thus yielding a more precise evaluation of PVAT inflammation (143, 144). In addition, the artificial intelligence (AI)-driven imaging biomarker (FRP) significantly improves the prediction of cardiac risk (143).
However, various imaging modalities, including magnetic resonance imaging (MRI) (145), positron emission tomography (PET), and ultrasound, offer promising applications for evaluating PVAT. MRI allows for the assessment of PVAT volume around major arteries, including the aorta, and has demonstrated an independent association with markers of subclinical atherosclerosis (146). PET provides functional insights into the metabolic activity of adipose tissue; however, its low spatial resolution limits its effectiveness in evaluating PVAT around coronary arteries (142). Although ultrasound may yield some surrogate measurements of EAT (147–149), its limited ability to distinguish adipose tissue from other structures, combined with its operator-dependent nature, reduces its utility in assessing PVAT in the coronary region.
3.2 Circulating biomarkers in evaluating PVAT function
In addition to imaging biomarkers, circulating biomarkers, including adipokines and cytokines, have potential applications in evaluating PVAT function and predicting CVDs (150). For instance, plasma adiponectin levels are diminished in individuals with obesity and diabetes, and lower adiponectin levels have been linked to an elevated risk of CVDs (151). Similarly, plasma levels of IL-6 and TNF-α are increased in obesity and other pathological conditions, with high levels of these cytokines associated with a greater risk of CVDs (152). However, it is important to note that circulating biomarkers are not specific to PVAT function and may be influenced by other factors, such as systemic inflammation and tissue damage.
4 PVAT as a therapeutic target
4.1 Lifestyle modifications for enhancing PVAT function and reducing cardiovascular risk
Lifestyle modifications, including diet and exercise, hold significant promise for enhancing PVAT function and mitigating the risk of CVDs (153–155). In humans, exercise-induced weight loss is more effective than dieting alone in improving circulating adipokine profiles and insulin resistance (156, 157). High-intensity interval training has been particularly effective in reducing total adipose tissue and visceral adipose tissue (VAT) mass in adults (158, 159), as well as enhancing cardiorespiratory fitness in obese children (158, 160).
4.2 The role of novel pharmacological strategies in restoring PVAT function in CVD
PVAT has a well-established significance in the development and progression of vascular diseases, making it a promising target for innovative therapeutic strategies in cardiovascular medicine. Research has demonstrated that current anti-diabetic medications can partly mediate their positive cardiovascular effects through their interactions with adipose tissues, including PVAT (161). For example, glucagon-like peptide-1 (GLP-1) receptor agonists facilitate adipocyte differentiation and enhance overall adipocyte health (162, 163). GLP-1 and its agonists exert various direct effects on PVAT biology, including the reduction of lipid accumulation (164) and the promotion of lipogenesis (165). Furthermore, GLP-1 receptor agonists upregulate the expression of adipokines (166) and promote M2 macrophage polarization (167), clinical studies have demonstrated that administering the GLP-1 analogue liraglutide to individuals with obesity results in favorable modifications in plasma lipid profiles and a reduction in apolipoprotein B levels, which could significantly lower the risk of CVDs (168–171). Additionally, sodium-glucose cotransporter 2 inhibitors (SGLT2-i) have been found to exert beneficial effects on human PVAT, improving mitochondrial efficiency, reducing oxidative stress and inflammation, and enhancing tissue function (172). The SGLT2 inhibitor empagliflozin is notable for its ability to enhance the browning of WAT and activate resident M2 macrophages, which helps decrease inflammation and insulin resistance linked to obesity, as demonstrated in animal research (173, 174). In human subjects, treatment with empagliflozin is linked to a decrease in the volume of EAT, which is critically associated with cardiometabolic risks (175, 176). Moreover, statins are known to decrease vascular inflammation, and their administration correlates with significant changes in the phenotype of PVAT following instances of acute coronary syndrome (ACS) and the commencement of statin treatment (177, 178). Other potential methods for restoring the health of PVAT include the use of PPAR-γ agonists such as rosiglitazone (179, 180). It is essential to acknowledge the recognized negative impacts on individuals suffering from congestive heart failure. Furthermore, agonists of the cannabinoid CB1 receptor might facilitate the release of H2S from PVAT (181, 182). Mineralocorticoid receptor blockers have been shown to reduce adipose tissue inflammation and increase adiponectin levels (183, 184). However, the specific effects of current drugs on the PVAT remain unquantified, which poses a risk of off-target consequences. Therefore, it is essential to examine this issue from a more neutral perspective and to conduct further research.
4.3 Effect of surgical intervention on PVAT function
In patients with type 2 diabetes and obesity, undergoing metabolic surgery (defined as procedures that influence metabolism by inducing weight loss and altering gastrointestinal physiology) has been associated with a significantly reduced risk of major adverse cardiovascular events (MACE) compared to traditional nonsurgical management approaches (185). This finding highlights the potential of metabolic surgical interventions to enhance cardiovascular outcomes for this patient population, underscoring the necessity for a shift in treatment strategies. The deterioration of the anticontractile properties of PVAT observed in obesity is partly attributable to a reduction in adiponectin levels within the PVAT. This hormone is crucial for maintaining vascular health. Importantly, the weight loss achieved through bariatric surgery has the potential to restore adiponectin levels, which may lead to improvements in PVAT function and, consequently, cardiovascular health (186). In contrast, procedures such as abdominal liposuction have not shown significant benefits in correcting the metabolic abnormalities associated with obesity, thereby highlighting the limitations of these methods in addressing the complex nature of obesity-related health issues (187).
4.4 Targeting gut microbiota as a therapeutic approach to enhance PVAT function and vascular health
The gut microbiota plays a substantial role in influencing PVAT through various mechanisms, particularly by producing metabolites and regulating immune responses (188). Among these metabolites, short-chain fatty acids produced by gut microbiota have been found to enhance the metabolic activity of adipocytes within the PVAT, which can lead to a reduction in inflammation levels. On the other hand, the presence of trimethylamine-N-oxide can trigger oxidative stress and inflammation in PVAT, negatively affecting its function. Additionally, emerging research in the field of cancer therapy shows that gut microbiota can regulate immune responses and produce metabolites that impact cancer development and treatment responses, suggesting its role in PVAT may have implications for broader health aspects related to cancer and vascular health (189, 190). Maintaining a healthy balance of gut microbiota is crucial, as it regulates immune cells that are essential for preserving PVAT homeostasis. When dysbiosis occurs, this balance is disrupted, potentially compromising both the structure and the functional integrity of PVAT. This finding highlights the promising potential of targeting gut microbiota as a therapeutic approach to enhance PVAT function and overall vascular health (191).
4.5 Other potential therapeutic targets
The activation of β3-adrenergic receptors (β3-AR) by mirabegron presents another fascinating avenue for bolstering vascular health. Research has indicated that this pharmacological agent can prevent serious vascular conditions such as aortic dissection and aneurysm by promoting lymphangiogenesis within PVAT (192). Furthermore, colchicine has shown promise in inhibiting inflammatory responses in PVAT, as well as curbing abnormal cellular behaviors such as proliferation and migration that can lead to vascular complications (193–195). The role of perivascular relaxing factors, including methyl palmitate, is equally critical in managing PVAT and enhancing vascular functionality. These factors facilitate several important processes, including the relaxation of vascular smooth muscle, reduction of vascular resistance, and improvement of blood perfusion. Additionally, they work to inhibit inflammatory responses and decrease both inflammatory cell infiltration and mediator production (196, 197). Collectively, these multifaceted effects play a vital role in addressing CVDs associated with PVAT dysfunction. Despite the promising implications of these interventions, it is essential to conduct further research to ascertain their specific effects and ensure their safety for clinical applications.
5 Discussion and outlook
5.1 Discussion
The evolving recognition of PVAT as a pivotal regulator of vascular homeostasis and a driver of CVD underscores its potential as a therapeutic target. However, translating mechanistic insights into clinical applications remains fraught with challenges, necessitating a critical reevaluation of current paradigms and the integration of cutting-edge methodologies. While recent advancements in single-cell omics, imaging biomarkers, and therapeutic strategies have illuminated PVAT's spatiotemporal complexity, significant gaps still persist in reconciling its depot-specific heterogeneity with systemic vascular pathology. Although animal experiments have shown that β3-AR, methyl palmitate, and cannabinoid CB1 receptor agonists can alleviate the inflammatory response of PVAT (181, 182, 192), it is crucial to note that due to differences in PVAT composition and function across species, the applicability of preclinical findings (primarily from rodent models) to human pathophysiology requires rigorous validation. Furthermore, gender and age profoundly influence PVAT biology and CVD risk, though mechanistic insights into these effects remain limited (198, 199). For instance, postmenopausal women exhibit accelerated PVAT inflammation linked to estrogen loss (200, 201), and aging-associated PVAT fibrosis exacerbates endothelial dysfunction (122, 202).
To address these complexities, in this review, we systematically synthesized the molecular crosstalk, spatial heterogeneity, and therapeutic potential of PVAT in the pathogenesis of CVD. Concurrently, we highlighted three limitations of previous studies: (1) the oversimplified assumption of PVAT homogeneity, (2) the paracrine-centric view that overshadows mechanotransduction pathways, and (3) the unquantified depot-specific effects of existing therapies.
Given this landscape, the emerging role of PVAT in cardiovascular pathophysiology necessitates a paradigm shift from observational research to precision therapeutics. While single-cell omics elucidate its cellular heterogeneity—such as UCP1 + thermogenic, ADIPOQ + endocrine, and COL1A1 + fibro-inflammatory adipocytes—and depot-specific metabolic programming, clinical translation is hindered by insufficient human models and an overreliance on rodent data. Although advanced imaging biomarkers, such as fat attenuation index (FAI), and AI-driven radiomics enhance the detection of coronary inflammation (141, 142); standardization gaps combined with systemic confounding factors limit their practical utility. Consequently, therapeutic innovation should prioritize PVAT-centric strategies, including macrophage-targeted PPAR-γ delivery and CRISPR-mediated ADIPOQ activation. Despite the preclinical promise shown by agents like β3-adrenergic receptor agonists (192) and colchicine (194), human trials remain limited. Hence, global consortia are essential to standardize protocols, address disparities, and ethically integrate AI-driven platforms. Ultimately, through bridging mechanistic depth with clinical pragmatism, PVAT research has the potential to redefine precision cardiology and mitigate residual cardiovascular risk.
5.2 Future research directions
To address the specific gaps and broader translational challenges, such as species differences, inadequacies in human models, biomarker standardization, and the necessity for human trials, we propose a multidisciplinary roadmap that integrates molecular insights with translational applications. Firstly, single-cell and spatial multi-omics techniques (e.g., snRNA-seq and single-cell ATAC sequencing) will elucidate PVAT's cellular diversity and epigenetic mechanisms, complemented by dynamic three-dimensional models to validate mechanosensitive pathways. Additionally, the development of clinical biomarkers requires cross-population validation of PVAT-specific signatures, while also addressing obesity-related fibrotic suppression. Moreover, therapeutic innovation depends on nanotechnology-enabled delivery systems [e.g., macrophage-targeted lipid nanoparticles (203)] and CRISPR activation-mediated ADIPOQ activation (204), combined with circadian optimization (e.g., timed melatonin administration). Finally, global consortia must standardize protocols and utilize AI-driven data harmonization to accelerate discovery, while adaptive trials address sex and racial disparities and ensure gene-editing safety (Figure 2).
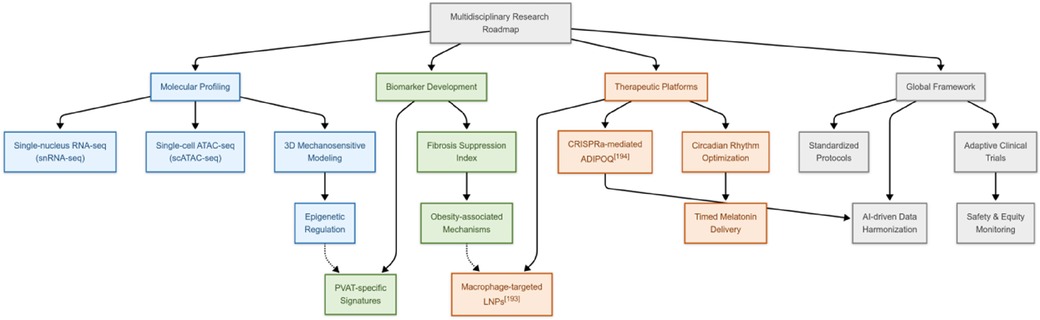
Figure 2. Multidisciplinary research roadmap for PVAT.
Moving forward, precision therapies will concentrate on cell-specific PVAT targets by employing systems biology and network pharmacology. Personalized lifestyle modifications and surgical interventions will be refined based on long-term outcome data, while mechanistic studies of gut microbiota and PVAT will inform targeted interventions. By emphasizing mechano-metabolic pathways, microbiota interactions, and ethical considerations, this roadmap seeks to transform PVAT biology into stratified cardiovascular therapies, thereby reducing residual risk and enhancing disease management through molecular precision.
6 Conclusion
PVAT serves as a critical regulator of vascular homeostasis, with its dysfunction contributing to cardiovascular pathology through paracrine imbalance, metabolic dysregulation, and extracellular matrix remodeling. Advances in imaging techniques and circulating biomarkers now facilitate the early detection of PVAT abnormalities, while emerging therapies, such as SGLT2 inhibitors and circadian rhythm modulation, show promise in restoring PVAT function. However, challenges persist in standardizing diagnostic protocols, optimizing tissue-specific drug delivery, and evaluating long-term safety. Future research ought to prioritize translational studies and multidisciplinary collaboration to integrate PVAT-targeted strategies into precision cardiovascular medicine, ultimately aiming to reduce residual disease risks and enhance clinical outcomes.
Author contributions
SZ: Data curation, Investigation, Project administration, Writing – original draft, Writing – review & editing. JJ: Writing – original draft, Writing – review & editing. YL: Writing – original draft, Writing – review & editing. GL: Writing – original draft, Writing – review & editing. SH: Writing – original draft, Writing – review & editing. SW: Writing – original draft, Writing – review & editing. CL: Writing – original draft, Writing – review & editing. HL: Writing – original draft, Writing – review & editing. NL: Writing – original draft, Writing – review & editing. JG: Writing – original draft, Writing – review & editing. LL: Writing – original draft, Writing – review & editing. ML: Writing – original draft, Writing – review & editing. JT: Writing – original draft, Writing – review & editing. KM: Writing – original draft, Writing – review & editing. LR: Writing – original draft, Writing – review & editing. MC: Writing – original draft, Writing – review & editing. L-YZ: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
PVAT, perivascular adipose tissue; CVDs, cardiovascular diseases; BAT, brown adipose tissue; WAT, white adipose tissue; EAT, epicardial adipose tissue; TAG, triacylglycerol; snRNA-seq, single-nucleus RNA sequencing; UCP1, uncoupling protein 1; ROS, reactive oxygen species; AMPK, adenosine monophosphate-activated protein kinase; 4-HNE, 4-hydroxynonenal; PPAR, peroxisome proliferator-activated receptor; T2DM, type 2 diabetes mellitus; IL-6, interleukin-6; TNF-α, tumor necrosis factor-alpha; EVs, extracellular vesicles; ESCRT, endosomal sorting complex required for transport; MVBs, multivesicular bodies; Nrg4, neuregulin-4; RAAS, renin-angiotensin-aldosterone system; IFN-γ, interferon-gamma; IL-17, interleukin-17; CT, computed tomography; FAI, fat attenuation index; CCTA, coronary computed tomography angiography; AI, artificial intelligence; MRI, magnetic resonance imaging; PET, positron emission tomography; VAT, visceral adipose tissue; GLP-1, glucagon-like peptide-1; SGLT2-i, sodium-glucose cotransporter 2 inhibitors; ACS, acute coronary syndrome; MACE, major adverse cardiovascular events; β3-AR, β3-adrenergic receptors.
References
1. Antoniades C, Tousoulis D, Vavlukis M, Fleming I, Duncker DJ, Eringa E, et al. Perivascular adipose tissue as a source of therapeutic targets and clinical biomarkers. Eur Heart J. (2023) 44(38):3827–44. doi: 10.1093/eurheartj/ehad484
2. Saxton SN, Clark BJ, Withers SB, Eringa EC, Heagerty AM. Mechanistic links between obesity, diabetes, and blood pressure: role of perivascular adipose tissue. Physiol Rev. (2019) 99(4):1701–63. doi: 10.1152/physrev.00034.2018
3. Meijer RI, Hoevenaars FPM, Serné EH, Yudkin JS, Kokhuis TJA, Weijers EM, et al. JNK2 in myeloid cells impairs insulin’s vasodilator effects in muscle during early obesity development through perivascular adipose tissue dysfunction. Am J Physiol Heart Circ Physiol. (2019) 317(2):H364–74. doi: 10.1152/ajpheart.00663.2018
4. Chen JY, Wu YP, Li CY, Jheng HF, Kao LZ, Yang CC, et al. PPARγ activation improves the microenvironment of perivascular adipose tissue and attenuates aortic stiffening in obesity. J Biomed Sci. (2021) 28(1):22. doi: 10.1186/s12929-021-00720-y
5. Wang Y, Wang X, Chen Y, Zhang Y, Zhen X, Tao S, et al. Perivascular fat tissue and vascular aging: a sword and a shield. Pharmacol Res. (2024) 203:107140. doi: 10.1016/j.phrs.2024.107140
6. Azul L, Leandro A, Boroumand P, Klip A, Seiça R, Sena CM. Increased inflammation, oxidative stress and a reduction in antioxidant defense enzymes in perivascular adipose tissue contribute to vascular dysfunction in type 2 diabetes. Free Radic Biol Med. (2020) 146:264–74. doi: 10.1016/j.freeradbiomed.2019.11.002
7. Stanek A, Brożyna-Tkaczyk K, Myśliński W. The role of obesity-induced perivascular adipose tissue (PVAT) dysfunction in vascular homeostasis. Nutrients. (2021) 13(11):3843. doi: 10.3390/nu13113843
8. Rafeh R, Viveiros A, Oudit GY, El-Yazbi AF. Targeting perivascular and epicardial adipose tissue inflammation: therapeutic opportunities for cardiovascular disease. Clin Sci (Lond). (2020) 134(7):827–51. doi: 10.1042/CS20190227
9. Liu C-L, Ren J, Wang Y, Zhang X, Sukhova GK, Liao M, et al. Adipocytes promote interleukin-18 binding to its receptors during abdominal aortic aneurysm formation in mice. Eur Heart J. (2020) 41(26):2456–68. doi: 10.1093/eurheartj/ehz856
10. Chang L, Villacorta L, Li R, Hamblin M, Xu W, Dou C, et al. Loss of perivascular adipose tissue on peroxisome proliferator-activated receptor-γ deletion in smooth muscle cells impairs intravascular thermoregulation and enhances atherosclerosis. Circulation. (2012) 126(9):1067–78. doi: 10.1161/CIRCULATIONAHA.112.104489
11. Chang L, Xiong W, Zhao X, Fan Y, Guo Y, Garcia-Barrio M, et al. Bmal1 in perivascular adipose tissue regulates resting-phase blood pressure through transcriptional regulation of angiotensinogen. Circulation. (2018) 138(1):67–79. doi: 10.1161/CIRCULATIONAHA.117.029972
12. Dorothea S-A, Häring HU. Perivascular adipose tissue: an unique fat compartment relevant for the cardiometabolic syndrome. Rev Endocr Metab Disord. (2016) 17(1):51–60. doi: 10.1007/s11154-016-9346-3
13. Gu P, Hui X, Zheng Q, Gao Y, Jin L, Jiang W, et al. Mitochondrial uncoupling protein 1 antagonizes atherosclerosis by blocking NLRP3 inflammasome-dependent interleukin-1β production. Sci Adv. (2021) 7(50):eabl4024. doi: 10.1126/sciadv.abl4024
14. Padilla J, Jenkins NT, Vieira-Potter VJ, Laughlin MH. Divergent phenotype of rat thoracic and abdominal perivascular adipose tissues. Am J Physiol Regul Integr Comp Physiol. (2013) 304(7):R543–52. doi: 10.1152/ajpregu.00567.2012
15. Mestres-Arenas A, Villarroya J, Giralt M, Villarroya F, Peyrou M. A differential pattern of batokine expression in perivascular adipose tissue depots from mice. Front Physiol. (2021) 12:714530. doi: 10.3389/fphys.2021.714530
16. Iacobellis G. Epicardial adipose tissue in contemporary cardiology. Nat Rev Cardiol. (2022) 19(9):593–606. doi: 10.1038/s41569-022-00679-9
17. Iacobellis G, Bianco AC. Epicardial adipose tissue: emerging physiological, pathophysiological and clinical features. Trends Endocrinol Metab. (2011) 22(11):450–7. doi: 10.1016/j.tem.2011.07.003
18. Ahmadieh S, Kim HW, Weintraub NL. Potential role of perivascular adipose tissue in modulating atherosclerosis. Clin Sci (Lond). (2020) 134(1):3–13. doi: 10.1042/CS20190577
19. Cinti S. Adipocyte differentiation and transdifferentiation: plasticity of the adipose organ. J Endocrinol Invest. (2002) 25(10):823–35. doi: 10.1007/BF03344046
20. Himms-Hagen J, Melnyk A, Zingaretti MC, Ceresi E, Barbatelli G, Cinti S. Multilocular fat cells in WAT of CL-316243-treated rats derive directly from white adipocytes. Am J Physiol Cell Physiol. (2000) 279(3):C670–81. doi: 10.1152/ajpcell.2000.279.3.C670
21. Rahbani JF, Bunk J, Lagarde D, Samborska B, Roesler A, Xiao H, et al. Parallel control of cold-triggered adipocyte thermogenesis by UCP1 and CKB. Cell Metab. (2024) 36(3):526–40.e7. doi: 10.1016/j.cmet.2024.01.001
22. Giroud M, Jodeleit H, Prentice KJ, Bartelt A. Adipocyte function and the development of cardiometabolic disease. J Physiol. (2022) 600(5):1189–208. doi: 10.1113/JP281979
23. Xiao L, De Jesus DF, Ju CW, Wei JB, Hu J, Distefano-Forti A, et al. M(6)A mRNA methylation in brown fat regulates systemic insulin sensitivity via an inter-organ prostaglandin signaling axis independent of UCP1. Cell Metab. (2024) 36(10):2207–27.e9. doi: 10.1016/j.cmet.2024.08.006
24. Angueira AR, Sakers AP, Holman CD, Cheng L, Arbocco MN, Shamsi F, et al. Defining the lineage of thermogenic perivascular adipose tissue. Nat Metab. (2021) 3(4):469–84. doi: 10.1038/s42255-021-00380-0
25. van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, et al. Cold-activated brown adipose tissue in healthy men. N Engl J Med. (2009) 360(15):1500–8. doi: 10.1056/NEJMoa0808718
26. Ye M, Ruan CC, Fu M, Xu L, Chen D, Zhu M, et al. Developmental and functional characteristics of the thoracic aorta perivascular adipocyte. Cell Mol Life Sci. (2018) 76(4):777–89. doi: 10.1007/s00018-018-2970-1
27. Long JZ, Svensson KJ, Tsai L, Zeng X, Roh HC, Kong X, et al. A smooth muscle-like origin for beige adipocytes. Cell Metab. (2014) 19(5):810–20. doi: 10.1016/j.cmet.2014.03.025
28. Vijay J, Gauthier M-F, Biswell RL, Louiselle DA, Johnston JJ, Cheung WA, et al. Single-cell analysis of human adipose tissue identifies depot and disease specific cell types. Nat Metab. (2020) 2(1):97–109. doi: 10.1038/s42255-019-0152-6
29. Li X, Ma Z, Zhu YZ. Regional heterogeneity of perivascular adipose tissue: morphology, origin, and secretome. Front Pharmacol. (2021) 12:697720. doi: 10.3389/fphar.2021.697720
30. Wang Z, Lu H, Garcia-Barrio M, Guo Y, Zhang J, Chen YE, et al. RNA sequencing reveals perivascular adipose tissue plasticity in response to angiotensin II. Pharmacol Res. (2022) 178:106183. doi: 10.1016/j.phrs.2022.106183
31. Hwej A, Al-Ferjani A, Alshuweishi Y, Naji A, Kennedy S, Salt IP. Lack of AMP-activated protein kinase-α1 reduces nitric oxide synthesis in thoracic aorta perivascular adipose tissue. Vasc Pharmacol. (2024) 157:107437. doi: 10.1016/j.vph.2024.107437
32. Wilcox CS, Herbert C, Wang C, Ma Y, Sun P, Li T, et al. Signals from inflamed perivascular adipose tissue contribute to small-vessel dysfunction in women with the human immunodeficiency virus. J Infect Dis. (2024) 230(1):67–77. doi: 10.1093/infdis/jiae094
33. Krawczyńska A, Herman AP, Antushevich H, Bochenek J, Wojtulewicz K, Zięba DA. The influence of photoperiod on the action of exogenous leptin on gene expression of proinflammatory cytokines and their receptors in the thoracic perivascular adipose tissue (PVAT) in ewes. Mediators Inflamm. (2019) 2019:7129476. doi: 10.1155/2019/7129476
34. Zhang ZB, Ruan CC, Lin JR, Xu L, Chen XH, Du YN, et al. Perivascular adipose tissue-derived PDGF-D contributes to aortic aneurysm formation during obesity. Diabetes. (2018) 67(8):1549–60. doi: 10.2337/db18-0098
35. Jüttner AA, Ataei Ataabadi E, Golshiri K, de Vries R, Garrelds IM, Danser AHJ, et al. Adiponectin secretion by perivascular adipose tissue supports impaired vasodilation in a mouse model of accelerated vascular smooth muscle cell and adipose tissue aging. Vascul Pharmacol. (2024) 154:107281. doi: 10.1016/j.vph.2024.107281
36. Kagota S, Maruyama-Fumoto K, Iwata S, Shimari M, Koyanagi S, Shiokawa Y, et al. Perivascular adipose tissue-enhanced vasodilation in metabolic syndrome rats by apelin and N -acetyl−l-cysteine-sensitive factor(s). Int J Mol Sci. (2018) 20(1):106. doi: 10.3390/ijms20010106
37. Nel S, Strijdom H, Genis A, Everson F, Van Wijk R, Kotzé SH. A histomorphometric study on the effects of antiretroviral therapy (ART) combined with a high-calorie diet (HCD) on aortic perivascular adipose tissue (PVAT). Acta Histochem. (2017) 119(5):555–62. doi: 10.1016/j.acthis.2017.05.009
38. Berti L, Hartwig S, Irmler M, Rädle B, Siegel-Axel D, Beckers J, et al. Impact of fibroblast growth factor 21 on the secretome of human perivascular preadipocytes and adipocytes: a targeted proteomics approach. Arch Physiol Biochem. (2016) 122(5):281–8. doi: 10.1080/13813455.2016.1212898
39. Kudryavtseva O, Lyngsø KS, Jensen BL, Dimke H. Nitric oxide, endothelium-derived hyperpolarizing factor, and smooth muscle-dependent mechanisms contribute to magnesium-dependent vascular relaxation in mouse arteries. Acta Physiol (Oxf). (2024) 240(3):e14096. doi: 10.1111/apha.14096
40. Ozkor MA, Rahman AM, Murrow JR, Kavtaradze N, Lin J, Manatunga A, et al. Differences in vascular nitric oxide and endothelium-derived hyperpolarizing factor bioavailability in blacks and whites. Arterioscler Thromb Vasc Biol. (2014) 34(6):1320–7. doi: 10.1161/ATVBAHA.113.303136
41. Adachi Y, Ueda K, Nomura S, Ito K, Katoh M, Katagiri M, et al. Beiging of perivascular adipose tissue regulates its inflammation and vascular remodeling. Nat Commun. (2022) 13(1):5117. doi: 10.1038/s41467-022-32658-6
42. Marek G, Pannu V, Shanmugham P, Pancione B, Mascia D, Crosson S, et al. Adiponectin resistance and proinflammatory changes in the visceral adipose tissue induced by fructose consumption via ketohexokinase-dependent pathway. Diabetes. (2015) 64(2):508–18. doi: 10.2337/db14-0411
43. Hotta K, Funahashi T, Bodkin NL, Ortmeyer HK, Arita Y, Hansen BC, et al. Circulating concentrations of the adipocyte protein adiponectin are decreased in parallel with reduced insulin sensitivity during the progression to type 2 diabetes in rhesus monkeys. Diabetes. (2001) 50(5):1126–33. doi: 10.2337/diabetes.50.5.1126
44. Kumada M, Kihara S, Sumitsuji S, Kawamoto T, Matsumoto S, Ouchi N, et al. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler Thromb Vasc Biol. (2003) 23(1):85–9. doi: 10.1161/01.ATV.0000048856.22331.50
45. Wang B, Pan Y, Yang G, Cui Z, Yu W, Liu H, et al. Sfrp5/Wnt5a and leptin/adiponectin levels in the serum and the periarterial adipose tissue of patients with peripheral arterial occlusive disease. Clin Biochem. (2021) 87:46–51. doi: 10.1016/j.clinbiochem.2020.11.002
46. Yamawaki H, Kuramoto J, Kameshima S, Usui T, Okada M, Hara Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem Biophys Res Commun. (2011) 408(2):339–43. doi: 10.1016/j.bbrc.2011.04.039
47. Yu S, Zhang Y, Li MZ, Xu H, Wang Q, Song J, et al. Chemerin and apelin are positively correlated with inflammation in obese type 2 diabetic patients. Chin Med J (Engl). (2012) 125(19):3440–4.23044303
48. Gálvez-Prieto B, Somoza B, Gil-Ortega M, García-Prieto CF, De Las Heras AI, González MC, et al. Anticontractile effect of perivascular adipose tissue and leptin are reduced in hypertension. Front Pharmacol. (2012) 3:103. doi: 10.3389/fphar.2012.00103
49. Stejskal D, Karpisek M, Hanulova Z, Svestak M. Chemerin is an independent marker of the metabolic syndrome in a Caucasian population–a pilot study. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. (2008) 152(2):217–21. doi: 10.5507/bp.2008.033
50. Atasoy A, Çakır E, Ahbab S, Erdoğan Döventaş Y, Koldaş M, Ataoğlu E, et al. Visfatin levels in hormonally inactive adrenal adenoma and their association with metabolic parameters. Turk J Med Sci. (2018) 48(3):548–53. doi: 10.3906/sag-1709-74
51. Eyileten T, Sonmez A, Saglam M, Cakir E, Caglar K, Oguz Y, et al. Effect of renin-angiotensin-aldosterone system (RAAS) blockade on visfatin levels in diabetic nephropathy. Nephrology (Carlton). (2010) 15(2):225–9. doi: 10.1111/j.1440-1797.2009.01173.x
52. Pagano C, Marin O, Calcagno A, Schiappelli P, Pilon C, Milan G, et al. Increased serum resistin in adults with prader-willi syndrome is related to obesity and not to insulin resistance. J Clin Endocrinol Metab. (2005) 90(7):4335–40. doi: 10.1210/jc.2005-0293
53. Tong S, Du Y, Ji Q, Dong R, Cao J, Wang Z, et al. Expression of Sfrp5/Wnt5a in human epicardial adipose tissue and their relationship with coronary artery disease. Life Sci. (2020) 245:117338. doi: 10.1016/j.lfs.2020.117338
54. Ghorbanian B, Wong A, Iranpour A. The effect of dietary carbohydrate restriction and aerobic exercise on retinol binding protein 4 (RBP4) and fatty acid binding protein 5 (FABP5) in middle-aged men with metabolic syndrome. Br J Nutr. (2023) 130(4):553–63. doi: 10.1017/S0007114522003580
55. Gruzdeva OV, Dyleva YA, Belik EV, Sinitsky MY, Stasev AN, Kokov AN, et al. Relationship between epicardial and coronary adipose tissue and the expression of adiponectin, leptin, and interleukin 6 in patients with coronary artery disease. J Pers Med. (2022) 12(2):129. doi: 10.3390/jpm12020129
56. Virdis A, Colucci R, Bernardini N, Blandizzi C, Taddei S, Masi S. Microvascular endothelial dysfunction in human obesity: role of TNF-α. J Clin Endocrinol Metab. (2019) 104(2):341–8. doi: 10.1210/jc.2018-00512
57. Carlström M, Larsen FJ, Nyström T, Hezel M, Borniquel S, Weitzberg E, et al. Dietary inorganic nitrate reverses features of metabolic syndrome in endothelial nitric oxide synthase-deficient mice. Proc Natl Acad Sci U S A. (2010) 107(41):17716–20. doi: 10.1073/pnas.1008872107
58. Szabo C. Gaseotransmitters: new frontiers for translational science. Sci Transl Med. (2010) 2(59):59ps4. doi: 10.1126/scitranslmed.3000721
59. Szabó C. Hydrogen sulphide and its therapeutic potential. Nat Rev Drug Discov. (2007) 6(11):917–35. doi: 10.1038/nrd2425
60. Szasz T, Webb RC. Perivascular adipose tissue: more than just structural support. Clin Sci. (2011) 122(1):1–12. doi: 10.1042/CS20110151
61. Hancock JT, Veal D. Nitric oxide, other reactive signalling compounds, redox, and reductive stress. J Exp Bot. (2021) 72(3):819–29. doi: 10.1093/jxb/eraa331
62. Nakladal D, Sijbesma JWA, Visser LM, Tietge UJF, Slart R, Deelman LE, et al. Perivascular adipose tissue-derived nitric oxide compensates endothelial dysfunction in aged pre-atherosclerotic apolipoprotein E-deficient rats. Vascul Pharmacol. (2021) 142:106945. doi: 10.1016/j.vph.2021.106945
63. Lan A, Xu W, Zhang H, Hua X, Zheng D, Guo R, et al. Inhibition of ROS-activated p38MAPK pathway is involved in the protective effect of H2S against chemical hypoxia-induced inflammation in PC12 cells. Neurochem Res. (2013) 38(7):1454–66. doi: 10.1007/s11064-013-1044-x
64. Xia N, Li H. The role of perivascular adipose tissue in obesity-induced vascular dysfunction. Br J Pharmacol. (2017) 174(20):3425–42. doi: 10.1111/bph.13650
65. Hsu CG, Chávez CL, Zhang C, Sowden M, Yan C, Berk BC. The lipid peroxidation product 4-hydroxynonenal inhibits NLRP3 inflammasome activation and macrophage pyroptosis. Cell Death Differ. (2022) 29(9):1790–803. doi: 10.1038/s41418-022-00966-5
66. Zahr T, Liu L, Chan M, Zhou Q, Cai B, He Y, et al. PPARγ (peroxisome proliferator-activated receptor γ) deacetylation suppresses aging-associated atherosclerosis and hypercholesterolemia. Arterioscler Thromb Vasc Biol. (2023) 43(1):30–44. doi: 10.1161/ATVBAHA.122.318061
67. Żółkiewicz J, Stochmal A, Zaremba M, Rudnicka L, Czuwara J. Circulating peroxisome proliferator-activated receptor γ is elevated in systemic sclerosis. Postepy Dermatol Alergol. (2020) 37(6):921–6. doi: 10.5114/ada.2019.84746
68. Almabrouk TAM, White AD, Ugusman AB, Skiba DS, Katwan OJ, Alganga H, et al. High fat diet attenuates the anticontractile activity of aortic PVAT via a mechanism involving AMPK and reduced adiponectin secretion. Front Physiol. (2018) 9:51. doi: 10.3389/fphys.2018.00051
69. Li Y, Zheng H, Yang J, Zhang B, Xing X, Zhang Z, et al. Association of genetic variants in leptin, leptin receptor and adiponectin with hypertension risk and circulating leptin/adiponectin changes. Gene. (2023) 853:147080. doi: 10.1016/j.gene.2022.147080
70. Katsiki N, Mantzoros C, Mikhailidis DP. Adiponectin, lipids and atherosclerosis. Curr Opin Lipidol. (2017) 28(4):347–54. doi: 10.1097/MOL.0000000000000431
71. Baker JF, England BR, Wysham KD, Sauer B, Joseph AM, Lenert A, et al. Associations between adiponectin and the development of diabetes in rheumatoid arthritis. J Clin Endocrinol Metab. (2024) 109(10):e1839–46. doi: 10.1210/clinem/dgae010
72. Gündüz B, Okimoto DK. Methyl donor supplementation alters serum leptin levels and increases appetite but not body weight in cross-fostered male Syrian hamster offspring (mesocricetus auratus). J Anim Physiol Anim Nutr (Berl). (2022) 106(5):1130–8. doi: 10.1111/jpn.13665
73. Simonds SE, Pryor JT, Ravussin E, Greenway FL, Dileone R, Allen AM, et al. Leptin mediates the increase in blood pressure associated with obesity. Cell. (2014) 159(6):1404–16. doi: 10.1016/j.cell.2014.10.058
74. Rodríguez A, Frühbeck G, Gómez-Ambrosi J, Catalán V, Sáinz N, Díez J, et al. The inhibitory effect of leptin on angiotensin II-induced vasoconstriction is blunted in spontaneously hypertensive rats. J Hypertens. (2006) 24(8):1589–97. doi: 10.1097/01.hjh.0000239295.17636.6e
75. Fortuño A, Rodríguez A, Gómez-Ambrosi J, Muñiz P, Salvador J, Díez J, et al. Leptin inhibits angiotensin II-induced intracellular calcium increase and vasoconstriction in the rat aorta. Endocrinology. (2002) 143(9):3555–60. doi: 10.1210/en.2002-220075
76. Syed AH, Lohana S, Aung NH, Memon MK, Shaikh A, Memon S, et al. Correlation of leptin with acute myocardial infarction: a case control study. Cureus. (2020) 12(12):e12190. doi: 10.7759/cureus.12190
77. Baig M, Alghalayini KW, Gazzaz ZJ, Atta H. Association of serum omentin-1, chemerin, and leptin with acute myocardial infarction and its risk factors. Pak J Med Sci. (2020) 36(6):1183–8. doi: 10.12669/pjms.36.6.2372
78. Saber H, Himali JJ, Shoamanesh A, Beiser A, Pikula A, Harris TB, et al. Serum leptin levels and the risk of stroke: the Framingham study. Stroke. (2015) 46(10):2881–5. doi: 10.1161/STROKEAHA.115.009463
79. Zhang Q, Shen Y, Niloy SI, O’rourke ST, Sun C. Chronic effects of apelin on cardiovascular regulation and angiotensin II-induced hypertension. Pharmaceuticals (Basel. (2023) 16(4):600. doi: 10.3390/ph16040600
80. Liu LQ, Zhang P, Qi YZ, Li H, Jiang YH, Yang CH. Quercetin attenuates atherosclerosis via modulating apelin signaling pathway based on plasma metabolomics. Chin J Integr Med. (2023) 29(12):1121–32. doi: 10.1007/s11655-023-3645-9
81. Kärberg K, Forbes A, Lember M. Visfatin and subclinical atherosclerosis in type 2 diabetes: impact of cardiovascular drugs. Medicina (Kaunas). (2023) 59(7):1324. doi: 10.3390/medicina59071324
82. Du Y, Ji Q, Cai L, Huang F, Lai Y, Liu Y, et al. Association between omentin-1 expression in human epicardial adipose tissue and coronary atherosclerosis. Cardiovasc Diabetol. (2016) 15:90. doi: 10.1186/s12933-016-0406-5
83. Nishimura M, Morioka T, Hayashi M, Kakutani Y, Yamazaki Y, Kurajoh M, et al. Plasma omentin levels are inversely associated with atherosclerosis in type 2 diabetes patients with increased plasma adiponectin levels: a cross-sectional study. Cardiovasc Diabetol. (2019) 18(1):167. doi: 10.1186/s12933-019-0973-3
84. Zhang Y, Lv P, Li Y, Zhang Y, Cheng C, Hao H, et al. Inflammatory cytokine interleukin-6 (IL-6) promotes the proangiogenic ability of adipose stem cells from obese subjects via the IL-6 signaling pathway. Curr Stem Cell Res Ther. (2023) 18(1):93–104. doi: 10.2174/1574888X17666220429103935
85. Eswar S, Rajagopalan B, Ete K, Nageswara Rao Gattem S. Serum tumor necrosis factor alpha (TNF-α) levels in obese and overweight adults: correlations with metabolic syndrome and inflammatory markers. Cureus. (2024) 16(7):e64619. doi: 10.7759/cureus.64619
86. Ait-Oufella H, Libby P, Tedgui A. Anticytokine immune therapy and atherothrombotic cardiovascular risk. Arterioscler Thromb Vasc Biol. (2019) 39(8):1510–9. doi: 10.1161/ATVBAHA.119.311998
87. Geddo F, Querio G, Asteggiano A, Antoniotti S, Porcu A, Occhipinti A, et al. Improving endothelial health with food-derived H(2)S donors: an in vitro study with S-allyl cysteine and with a black-garlic extract enriched in sulfur-containing compounds. Food Funct. (2023) 14(9):4163–72. doi: 10.1039/D3FO00412K
88. Wang C, Shu L, Cheng R, Yan M, Liang W, Zhou J, et al. Exercise enhances anti-contractile effects of PVAT through endogenous H2S in high-fat diet-induced obesity hypertension. Cardiovasc Drugs Ther. (2024). doi: 10.1007/s10557-024-07612-x
89. Oliveira PB, Zochio GP, Caetano ESP, Da Silva MLS, Dias-Junior CA. Vasodilator responses of perivascular adipose tissue-derived hydrogen sulfide stimulated with L-cysteine in pregnancy hypertension-induced endothelial dysfunction in rats. Antioxidants (Basel). (2023) 12(11):1919. doi: 10.3390/antiox12111919
90. Lee RM, Lu C, Su LY, Gao YJ. Endothelium-dependent relaxation factor released by perivascular adipose tissue. J Hypertens. (2009) 27(4):782–90. doi: 10.1097/HJH.0b013e328324ed86
91. Xia N, Horke S, Habermeier A, Closs EI, Reifenberg G, Gericke A, et al. Uncoupling of endothelial nitric oxide synthase in perivascular adipose tissue of diet-induced obese mice. Arterioscler Thromb Vasc Biol. (2016) 36(1):78–85. doi: 10.1161/ATVBAHA.115.306263
92. Virdis A, Duranti E, Rossi C, Dell’agnello U, Santini E, Anselmino M, et al. Tumour necrosis factor-alpha participates on the endothelin-1/nitric oxide imbalance in small arteries from obese patients: role of perivascular adipose tissue. Eur Heart J. (2015) 36(13):784–94. doi: 10.1093/eurheartj/ehu072
93. Chen Y, Chen C, Feng C, Tang A, Ma Y, He X, et al. AVE 3085, a novel endothelial nitric oxide synthase enhancer, attenuates cardiac remodeling in mice through the smad signaling pathway. Arch Biochem Biophys. (2015) 570:8–13. doi: 10.1016/j.abb.2015.02.020
94. Li X, Ballantyne LL, Yu Y, Funk CD. Perivascular adipose tissue–derived extracellular vesicle miR-221-3p mediates vascular remodeling. FASEB J. (2019) 33(11):12704–22. doi: 10.1096/fj.201901548R
95. Ballasy N, Jadli A, Edalat P, Gomes K, Wijesuriya T, Belke D, et al. Paracrine effects of perivascular adipose tissue on atherogenesis: role of extracellular vesicles-mediated intercellular communications. FASEB J. (2021) 35(Suppl 1):01712. doi: 10.1096/fasebj.2021.35.S1.01712
96. Sigdel S, Udoh G, Albalawy R, Wang J. Perivascular adipose tissue and perivascular adipose tissue-derived extracellular vesicles: new insights in vascular disease. Cells. (2024) 13(16):1309. doi: 10.3390/cells13161309
97. Chen HH, Li HF, Tseng TL, Lin H. Perivascular adipose tissue and adipocyte-derived exosomal miRNAs maintain vascular homeostasis. Heliyon. (2023) 9(12):e22607. doi: 10.1016/j.heliyon.2023.e22607
98. Liu J, Chen Y. Cell-cell crosstalk between fat cells and immune cells. Am J Physiol Endocrinol Metab. (2024) 327(3):E371–83. doi: 10.1152/ajpendo.00024.2024
99. Tutunchi H, Ostadrahimi A, Hosseinzadeh-Attar MJ, Miryan M, Mobasseri M, Ebrahimi-Mameghani M. A systematic review of the association of neuregulin 4, a brown fat-enriched secreted factor, with obesity and related metabolic disturbances. Obes Rev. (2020) 21(2):e12952. doi: 10.1111/obr.12952
100. Liu Y, Chen M. Neuregulin 4 as a novel adipokine in energy metabolism. Front Physiol. (2022) 13:1106380. doi: 10.3389/fphys.2022.1106380
101. Jeffery E, Wing A, Holtrup B, Sebo Z, Kaplan JL, Saavedra-Peña R, et al. The adipose tissue microenvironment regulates depot-specific adipogenesis in obesity. Cell Metab. (2016) 24(1):142–50. doi: 10.1016/j.cmet.2016.05.012
102. Siang DTC, Lim YC, Kyaw AMM, Win KN, Chia SY, Degirmenci U, et al. The RNA-binding protein HuR is a negative regulator in adipogenesis. Nat Commun. (2020) 11(1):213. doi: 10.1038/s41467-019-14001-8
103. Kumar RK, Yang Y, Contreras AG, Garver H, Bhattacharya S, Fink GD, et al. Phenotypic changes in T cell and macrophage subtypes in perivascular adipose tissues precede high-fat diet-induced hypertension. Front Physiol. (2021) 12:616055. doi: 10.3389/fphys.2021.616055
104. Wang S, Cao S, Arhatte M, Li D, Shi Y, Kurz S, et al. Adipocyte Piezo1 mediates obesogenic adipogenesis through the FGF1/FGFR1 signaling pathway in mice. Nat Commun. (2020) 11(1):2303. doi: 10.1038/s41467-020-16026-w
105. Veerapaneni P, Goo B, Ahmadieh S, Shi H, Kim DS, Ogbi M, et al. Transgenic overexpression of HDAC9 promotes adipocyte hypertrophy, insulin resistance and hepatic steatosis in aging mice. Biomolecules. (2024) 14(4):494. doi: 10.3390/biom14040494
106. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest. (2003) 112(12):1821–30. doi: 10.1172/JCI200319451
107. Talukdar S, Oh DY, Bandyopadhyay G, Li D, Xu J, Mcnelis J, et al. Neutrophils mediate insulin resistance in mice fed a high-fat diet through secreted elastase. Nat Med. (2012) 18(9):1407–12. doi: 10.1038/nm.2885
108. Soedono S, Cho KW. Adipose tissue dendritic cells: critical regulators of obesity-induced inflammation and insulin resistance. Int J Mol Sci. (2021) 22(16):8666. doi: 10.3390/ijms22168666
109. Hirai S, Ohyane C, Kim Y-I, Lin S, Goto T, Takahashi N, et al. Involvement of mast cells in adipose tissue fibrosis. Am J Physiol Endocrinol Metab. (2014) 306(3):E247–55. doi: 10.1152/ajpendo.00056.2013
110. Żelechowska P, Agier J, Kozłowska E, Brzezińska-Błaszczyk E. Mast cells participate in chronic low-grade inflammation within adipose tissue. Obes Rev. (2018) 19(5):686–97. doi: 10.1111/obr.12670
111. Moussa K, Gurung P, Adams-Huet B, Devaraj S, Jialal I. Increased eosinophils in adipose tissue of metabolic syndrome. J Diabetes Complicat. (2019) 33(8):535–8. doi: 10.1016/j.jdiacomp.2019.05.010
112. Vohralik EJ, Psaila AM, Knights AJ, Quinlan KGR. EoTHINophils: eosinophils as key players in adipose tissue homeostasis. Clin Exp Pharmacol Physiol. (2020) 47(8):1495–505. doi: 10.1111/1440-1681.13304
113. Nishimura S, Manabe I, Takaki S, Nagasaki M, Otsu M, Yamashita H, et al. Adipose natural regulatory B cells negatively control adipose tissue inflammation. Cell Metab. (2013) 18(5):759–66. doi: 10.1016/j.cmet.2013.09.017
114. Srikakulapu P, Upadhye A, Rosenfeld SM, Marshall MA, Mcskimming C, Hickman AW, et al. Perivascular adipose tissue harbors atheroprotective IgM-producing B cells. Front Physiol. (2017) 8:719. doi: 10.3389/fphys.2017.00719
115. Yu Y, Bai H, Wu F, Chen J, Li B, Li Y. Tissue adaptation of regulatory T cells in adipose tissue. Eur J Immunol. (2022) 52(12):1898–908. doi: 10.1002/eji.202149527
116. Jacks RD, Lumeng CN. Macrophage and T cell networks in adipose tissue. Nat Rev Endocrinol. (2024) 20(1):50–61. doi: 10.1038/s41574-023-00908-2
117. Nishimura S, Manabe I, Nagasaki M, Eto K, Yamashita H, Ohsugi M, et al. CD8 + effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat Med. (2009) 15(8):914–20. doi: 10.1038/nm.1964
118. Dos Reis Costa DEF, de Araújo NF, Nóbrega NRC, de Assis Rabelo Ribeiro N, de Oliveira ACC, Dos Santos Aggum Capettini L, et al. Contribution of RAS, ROS and COX-1-derived prostanoids to the contractile profile of perivascular adipose tissue in cafeteria diet-induced obesity. Life Sci. (2022) 309:120994. doi: 10.1016/j.lfs.2022.120994
119. Das E, Moon JH, Lee JH, Thakkar N, Pausova Z, Sung HK. Adipose tissue and modulation of hypertension. Curr Hypertens Rep. (2018) 20(11):96. doi: 10.1007/s11906-018-0894-7
120. Pritchard KA, Groszek L, Smalley DM, Sessa WC, Wu M, Villalon P, et al. Native low-density lipoprotein increases endothelial cell nitric oxide synthase generation of superoxide anion. Circ Res. (1995) 77(3):510–8. doi: 10.1161/01.RES.77.3.510
121. Ma Y, Li L, Shao Y, Bai X, Bai T, Huang X. Methotrexate improves perivascular adipose tissue/endothelial dysfunction via activation of AMPK/eNOS pathway. Mol Med Rep. (2017) 15(4):2353–9. doi: 10.3892/mmr.2017.6225
122. Tai GJ, Ma YJ, Feng JL, Li JP, Qiu S, Yu QQ, et al. NLRP3 inflammasome-mediated premature immunosenescence drives diabetic vascular aging dependent on the induction of perivascular adipose tissue dysfunction. Cardiovasc Res. (2024) 121(1):77–96. doi: 10.1093/cvr/cvae07
123. Mitri J, Tomah S, Furtado J, Tasabehji MW, Hamdy O. Plasma free fatty acids and metabolic effect in type 2 diabetes, an ancillary study from a randomized clinical trial. Nutrients. (2021) 13(4):1145. doi: 10.3390/nu13041145
124. Tang YH, Wang YH, Chen CC, Chan CJ, Tsai FJ, Chen SY. Genetic and functional effects of adiponectin in type 2 diabetes mellitus development. Int J Mol Sci. (2022) 23(21):13544. doi: 10.3390/ijms232113544
125. Bako HY, Ibrahim MA, Isah MS, Ibrahim S. Inhibition of JAK-STAT and NF-κB signalling systems could be a novel therapeutic target against insulin resistance and type 2 diabetes. Life Sci. (2019) 239:117045. doi: 10.1016/j.lfs.2019.117045
126. Seo JB, Riopel M, Cabrales P, Huh JY, Bandyopadhyay GK, Andreyev AY, et al. Knockdown of Ant2 reduces adipocyte hypoxia and improves insulin resistance in obesity. Nat Metab. (2019) 1(1):86–97. doi: 10.1038/s42255-018-0003-x
127. Yalçın T, Oğuz SH, Bayraktar M, Rakıcıoğlu N. Anthropometric measurements and serum TNF-α, IL-6 and adiponectin in type 2 diabetes. Diabetol Int. (2022) 13(2):396–406. doi: 10.1007/s13340-021-00553-y
128. Sprague AH, Khalil RA. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem Pharmacol. (2009) 78(6):539–52. doi: 10.1016/j.bcp.2009.04.029
129. Hattori Y, Suzuki K, Hattori S, Kasai K. Metformin inhibits cytokine-induced nuclear factor kappaB activation via AMP-activated protein kinase activation in vascular endothelial cells. Hypertension. (2006) 47(6):1183–8. doi: 10.1161/01.HYP.0000221429.94591.72
130. Li M, Van Esch B, Henricks PAJ, Folkerts G, Garssen J. The anti-inflammatory effects of short chain fatty acids on lipopolysaccharide- or tumor necrosis factor α-stimulated endothelial cells via activation of GPR41/43 and inhibition of HDACs. Front Pharmacol. (2018) 9:533. doi: 10.3389/fphar.2018.00533
131. Miao CY, Li ZY. The role of perivascular adipose tissue in vascular smooth muscle cell growth. Br J Pharmacol. (2012) 165(3):643–58. doi: 10.1111/j.1476-5381.2011.01404.x
132. Haghikia A, Landmesser U. Lipoproteins and cardiovascular redox signaling: role in atherosclerosis and coronary disease. Antioxid Redox Signal. (2018) 29(3):337–52. doi: 10.1089/ars.2017.7052
133. McCarthy CG, Martinez-Quinones P, Klee NS, Watts SW, Calmasini FB, Komic A, et al. Hypertension induced morphological and physiological changes in cells of the arterial wall. Am J Hypertens. (2018) 31(10):1067–78. doi: 10.1093/ajh/hpy083
134. Song Y, Jia H, Hua Y, Wu C, Li S, Li K, et al. The molecular mechanism of aerobic exercise improving vascular remodeling in hypertension. Front Physiol. (2022) 13:792292. doi: 10.3389/fphys.2022.792292
135. Persson P, Marchetti M, Friederich-Persson M. Browning of perivascular adipose tissue prevents vascular dysfunction and reduces hypertension in angiotensin II-infused mice. Am J Physiol Regul Integr Comp Physiol. (2023) 325(3):R290–8. doi: 10.1152/ajpregu.00043.2023
136. Cerecedo D, Martínez-Vieyra I, Hernández-Rojo I, Hernández-Cruz A, Rincón-Heredia R, Millán-Aldaco D, et al. Reactive oxygen species downregulate dystroglycans in the megakaryocytes of rats with arterial hypertension. Exp Cell Res. (2023) 433(2):113847. doi: 10.1016/j.yexcr.2023.113847
137. Abais-Battad JM, Lund H, Dasinger JH, Fehrenbach DJ, Cowley AW, Mattson DL. NOX2-derived reactive oxygen species in immune cells exacerbates salt-sensitive hypertension. Free Radic Biol Med. (2020) 146:333–9. doi: 10.1016/j.freeradbiomed.2019.11.014
138. Mikolajczyk TP, Nosalski R, Skiba DS, Koziol J, Mazur M, Justo-Junior AS, et al. 1,2,3,4,6-Penta-O-galloyl-β-d-glucose modulates perivascular inflammation and prevents vascular dysfunction in angiotensin II-induced hypertension. Br J Pharmacol. (2019) 176(12):1951–65. doi: 10.1111/bph.14583
139. Sena CM, Pereira A, Fernandes R, Letra L, Seiça RM. Adiponectin improves endothelial function in mesenteric arteries of rats fed a high-fat diet: role of perivascular adipose tissue. Br J Pharmacol. (2017) 174(20):3514–26. doi: 10.1111/bph.13756
140. Itani HA, Mcmaster WG Jr, Saleh MA, Nazarewicz RR, Mikolajczyk TP, Kaszuba AM, et al. Activation of human T cells in hypertension: studies of humanized mice and hypertensive humans. Hypertension. (2016) 68(1):123–32. doi: 10.1161/HYPERTENSIONAHA.116.07237
141. Oikonomou EK, Marwan M, Desai MY, Mancio J, Alashi A, Hutt Centeno E, et al. Non-invasive detection of coronary inflammation using computed tomography and prediction of residual cardiovascular risk (the CRISP CT study): a post-hoc analysis of prospective outcome data. Lancet. (2018) 392(10151):929–39. doi: 10.1016/S0140-6736(18)31114-0
142. Antoniades C, Antonopoulos AS, Deanfield J. Imaging residual inflammatory cardiovascular risk. Eur Heart J. (2020) 41(6):748–58. doi: 10.1093/eurheartj/ehz474
143. Oikonomou EK, Williams MC, Kotanidis CP, Desai MY, Marwan M, Antonopoulos AS, et al. A novel machine learning-derived radiotranscriptomic signature of perivascular fat improves cardiac risk prediction using coronary CT angiography. Eur Heart J. (2019) 40(43):3529–43. doi: 10.1093/eurheartj/ehz592
144. Chan K, Wahome E, Tsiachristas A, Antonopoulos AS, Patel P, Lyasheva M, et al. Inflammatory risk and cardiovascular events in patients without obstructive coronary artery disease: the ORFAN multicentre, longitudinal cohort study. Lancet. (2024) 403(10444):2606–18. doi: 10.1016/S0140-6736(24)00596-8
145. Ardissino M, Mccracken C, Bard A, Antoniades C, Neubauer S, Harvey NC, et al. Pericardial adiposity is independently linked to adverse cardiovascular phenotypes: a CMR study of 42 598 UK biobank participants. Eur Heart J Cardiovasc Imaging. (2022) 23(11):1471–81. doi: 10.1093/ehjci/jeac101
146. Antonopoulos AS, Angelopoulos A, Papanikolaou P, Simantiris S, Oikonomou EK, Vamvakaris K, et al. Biomarkers of vascular inflammation for cardiovascular risk prognostication: a meta-analysis. JACC Cardiovasc Imaging. (2022) 15(3):460–71. doi: 10.1016/j.jcmg.2021.09.014
147. Vasques ACJ, Souza JR, Yamanaka A, De Oliveira Mda S, Novaes FS, Pareja JC, et al. Sagittal abdominal diameter as a marker for epicardial adipose tissue in premenopausal women. Metab Clin Exp. (2013) 62(7):1032–6. doi: 10.1016/j.metabol.2013.01.022
148. Gaborit B, Julla JB, Fournel J, Ancel P, Soghomonian A, Deprade C, et al. Fully automated epicardial adipose tissue volume quantification with deep learning and relationship with CAC score and micro/macrovascular complications in people living with type 2 diabetes: the multicenter EPIDIAB study. Cardiovasc Diabetol. (2024) 23(1):328. doi: 10.1186/s12933-024-02411-y
149. Li Y, Zhang W, Hu Y, Xu Z, Huo Q, Qi H, et al. Pericoronary adipose tissue radiomics features as imaging markers for coronary artery disease risk assessment: insights from gene expression analysis. Cardiovasc Diabetol. (2024) 23(1):444. doi: 10.1186/s12933-024-02530-6
150. Su X, Peng D. Emerging functions of adipokines in linking the development of obesity and cardiovascular diseases. Mol Biol Rep. (2020) 47(10):7991–8006. doi: 10.1007/s11033-020-05732-9
151. Qi L, Doria A, Manson JAE, Meigs JB, Hunter D, Mantzoros CS, et al. Adiponectin genetic variability, plasma adiponectin, and cardiovascular risk in patients with type 2 diabetes. Diabetes. (2006) 55(5):1512–6. doi: 10.2337/db05-1520
152. Guzik TJ, Nosalski R, Maffia P, Drummond GR. Immune and inflammatory mechanisms in hypertension. Nat Rev Cardiol. (2024) 21(6):396–416. doi: 10.1038/s41569-023-00964-1
153. Lingvay I, Cohen RV, Roux CWL, Sumithran P. Obesity in adults. Lancet. (2024) 404(10456):972–87. doi: 10.1016/S0140-6736(24)01210-8
154. Davis CL, Lorig K. Lifestyle interventions for obesity. JAMA. (2024) 332(17):1488. doi: 10.1001/jama.2024.17533
155. Elmaleh-Sachs A, Schwartz JL, Bramante CT, Nicklas JM, Gudzune KA, Jay M. Obesity management in adults: a review. JAMA. (2023) 330(20):2000–15. doi: 10.1001/jama.2023.19897
156. Khoo J, Dhamodaran S, Chen DD, Yap SY, Chen RY, Tian RH. Exercise-induced weight loss is more effective than dieting for improving adipokine profile, insulin resistance, and inflammation in obese men. Int J Sport Nutr Exerc Metab. (2015) 25(6):566–75. doi: 10.1123/ijsnem.2015-0025
157. Saxton SN, Toms LK, Aldous RG, Withers SB, Ohanian J, Heagerty AM. Restoring perivascular adipose tissue function in obesity using exercise. Cardiovasc Drugs Ther. (2021) 35(6):1291–304. doi: 10.1007/s10557-020-07136-0
158. Guo P, Zhou Y, Zhu Y. Effects of a school-based lifestyle intervention on ideal cardiovascular health in Chinese children and adolescents: a national, multicentre, cluster-randomised controlled trial. Lancet Glob Health. (2023) 11(Suppl 1):S14. doi: 10.1016/S2214-109X(23)00097-9
159. Li M, Li J, Xu Y, Gao J, Cao Q, Ding Y, et al. Effect of 5:2 regimens: energy-restricted diet or low-volume high-intensity interval training combined with resistance exercise on glycemic control and cardiometabolic health in adults with overweight/obesity and type 2 diabetes: a three-arm randomized controlled trial. Diabetes Care. (2024) 47(6):1074–83. doi: 10.2337/dc24-0241
160. Woo KS, Chook P, Yu CW, Sung RYT, Qiao M, Leung SSF, et al. Effects of diet and exercise on obesity-related vascular dysfunction in children. Circulation. (2004) 109(16):1981–6. doi: 10.1161/01.CIR.0000126599.47470.BE
161. Kusminski CM, Bickel PE, Scherer PE. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat Rev Drug Discov. (2016) 15(9):639–60. doi: 10.1038/nrd.2016.75
162. Ussher JR, Drucker DJ. Glucagon-like peptide 1 receptor agonists: cardiovascular benefits and mechanisms of action. Nat Rev Cardiol. (2023) 20(7):463–74. doi: 10.1038/s41569-023-00849-3
163. Szekeres Z, Nagy A, Jahner K, Szabados E. Impact of selected glucagon-like peptide-1 receptor agonists on Serum lipids, adipose tissue, and muscle metabolism-a narrative review. Int J Mol Sci. (2024) 25(15):8214. doi: 10.3390/ijms25158214
164. Meier JJ. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat Rev Endocrinol. (2012) 8(12):728–42. doi: 10.1038/nrendo.2012.140
165. Delessa Challa T, Beaton N, Arnold M, Rudofsky G, Langhans W, Wolfrum C. Regulation of adipocyte formation by GLP-1/GLP-1R signaling. J Biol Chem. (2012) 287(9):6421–30. doi: 10.1074/jbc.M111.310342
166. Le Kim Chung T, Yoshida M, Hosaka T, Sakaue H, Harada N, Sakai T, et al. Exendin-4, a GLP-1 receptor agonist, directly induces adiponectin expression through protein kinase A pathway and prevents inflammatory adipokine expression. Biochem Biophys Res Commun. (2009) 390(3):613–8. doi: 10.1016/j.bbrc.2009.10.015
167. Fujiwara Y, Shiraishi D, Mizuta H, Komohara Y, Takeya M. Glucagon-like peptide-1 (GLP-1) induces M2 polarization of human macrophages via STAT3 activation. Biochem Biophys Res Commun. (2012) 425(2):304–8. doi: 10.1016/j.bbrc.2012.07.086
168. Marso SP, Daniels GH, Brown-Frandsen K, Kristensen P, Mann JF, Nauck MA, et al. Liraglutide and cardiovascular outcomes in type 2 diabetes. N Engl J Med. (2016) 375(4):311–22. doi: 10.1056/NEJMoa1603827
169. Santilli F, Simeone PG, Guagnano MT, Leo M, Maccarone MT, Di Castelnuovo A, et al. Effects of liraglutide on weight loss, fat distribution, and β-cell function in obese subjects with prediabetes or early type 2 diabetes. Diabetes Care. (2017) 40(11):1556–64. doi: 10.2337/dc17-0589
170. Verma S, Bhatt DL, Bain SC, Buse JB, Mann JFE, Marso SP, et al. Effect of liraglutide on cardiovascular events in patients with type 2 diabetes mellitus and polyvascular disease: results of the LEADER trial. Circulation. (2018) 137(20):2179–83. doi: 10.1161/CIRCULATIONAHA.118.033898
171. Que Y, Shi J, Zhang Z, Sun L, Li H, Qin X, et al. Ion cocktail therapy for myocardial infarction by synergistic regulation of both structural and electrical remodeling. Exploration (Beijing). (2024) 4(3):20230067. doi: 10.1002/EXP.20230067
172. Zelniker TA, Wiviott SD, Raz I, Im K, Goodrich EL, Bonaca MP, et al. SGLT2 inhibitors for primary and secondary prevention of cardiovascular and renal outcomes in type 2 diabetes: a systematic review and meta-analysis of cardiovascular outcome trials. Lancet. (2019) 393(10166):31–9. doi: 10.1016/S0140-6736(18)32590-X
173. Xu L, Ota T. Emerging roles of SGLT2 inhibitors in obesity and insulin resistance: focus on fat browning and macrophage polarization. Adipocyte. (2018) 7(2):121–8. doi: 10.1080/21623945.2017.1413516
174. Xu L, Nagata N, Nagashimada M, Zhuge F, Ni Y, Chen G, et al. SGLT2 inhibition by empagliflozin promotes fat utilization and browning and attenuates inflammation and insulin resistance by polarizing M2 macrophages in diet-induced obese mice. EBioMedicine. (2017) 20:137–49. doi: 10.1016/j.ebiom.2017.05.028
175. Zinman B, Wanner C, Lachin JM, Fitchett D, Bluhmki E, Hantel S, et al. Empagliflozin, cardiovascular outcomes, and mortality in type 2 diabetes. N Engl J Med. (2015) 373(22):2117–28. doi: 10.1056/NEJMoa1504720
176. Wanner C, Lachin JM, Inzucchi SE, Fitchett D, Mattheus M, George J, et al. Empagliflozin and clinical outcomes in patients with type 2 diabetes mellitus, established cardiovascular disease, and chronic kidney disease. Circulation. (2018) 137(2):119–29. doi: 10.1161/CIRCULATIONAHA.117.028268
177. Šatný M, Jaroslav H, Vrablík M. Statins and inflammation. Current atherosclerosis reports. (2021).
178. Sanna F, Margaritis M, Antoniades C. Perivascular adipose tissue as an endocrine organ: the role of statins. Curr Pharm Des. (2017). doi: 10.2174/1381612823666170926133843
179. Guri AJ, Hontecillas R, Bassaganya-Riera J. Abscisic acid synergizes with rosiglitazone to improve glucose tolerance and down-modulate macrophage accumulation in adipose tissue: possible action of the cAMP/PKA/PPAR γ axis. Clin Nutr. (2010) 29(5):646–53. doi: 10.1016/j.clnu.2010.02.003
180. Derosa G, Maffioli P. Peroxisome proliferator-activated receptor-γ (PPAR-γ) agonists on glycemic control, lipid profile and cardiovascular risk. Curr Mol Pharmacol. (2012) 5(2):272–81. doi: 10.2174/1874467211205020272
181. Bajzer M, Olivieri M, Haas MK, Pfluger PT, Magrisso IJ, Foster MT, et al. Cannabinoid receptor 1 (CB1) antagonism enhances glucose utilisation and activates brown adipose tissue in diet-induced obese mice. Diabetologia. (2011) 54(12):3121–31. doi: 10.1007/s00125-011-2302-6
182. Han JH, Shin H, Rho JG, Kim JE, Son DH, Yoon J, et al. Peripheral cannabinoid 1 receptor blockade mitigates adipose tissue inflammation via NLRP3 inflammasome in mouse models of obesity. Diabetes Obes Metab. (2018) 20(9):2179–89. doi: 10.1111/dom.13350
183. Epstein M. Reduction of cardiovascular risk in chronic kidney disease by mineralocorticoid receptor antagonism. Lancet Diabetes Endocrinol. (2015) 3(12):993–1003. doi: 10.1016/S2213-8587(15)00289-2
184. Guo C, Ricchiuti V, Lian BQ, Yao TM, Coutinho P, Romero JR, et al. Mineralocorticoid receptor blockade reverses obesity-related changes in expression of adiponectin, peroxisome proliferator-activated receptor-gamma, and proinflammatory adipokines. Circulation. (2008) 117(17):2253–61. doi: 10.1161/CIRCULATIONAHA.107.748640
185. Aminian A, Zajichek A, Arterburn DE, Wolski KE, Brethauer SA, Schauer PR, et al. Association of metabolic surgery with major adverse cardiovascular outcomes in patients with type 2 diabetes and obesity. JAMA. (2019) 322(13):1271–82. doi: 10.1001/jama.2019.14231
186. Aghamohammadzadeh R, Greenstein AS, Yadav R, Jeziorska M, Hama S, Soltani F, et al. Effects of bariatric surgery on human small artery function: evidence for reduction in perivascular adipocyte inflammation, and the restoration of normal anticontractile activity despite persistent obesity. J Am Coll Cardiol. (2013) 62(2):128–35. doi: 10.1016/j.jacc.2013.04.027
187. Klein S, Fontana L, Young VL, Coggan AR, Kilo C, Patterson BW, et al. Absence of an effect of liposuction on insulin action and risk factors for coronary heart disease. N Engl J Med. (2004) 350(25):2549–57. doi: 10.1056/NEJMoa033179
188. Shen J, Obin MS, Zhao L. The gut microbiota, obesity and insulin resistance. Mol Aspects Med. (2013) 34(1):39–58. doi: 10.1016/j.mam.2012.11.001
189. Zhao LY, Mei JX, Yu G, Lei L, Zhang WH, Liu K, et al. Role of the gut microbiota in anticancer therapy: from molecular mechanisms to clinical applications. Signal Transduct Target Ther. (2023) 8(1):201. doi: 10.1038/s41392-023-01406-7
190. Li Z, Xiong W, Liang Z, Wang J, Zeng Z, Kołat D, et al. Critical role of the gut microbiota in immune responses and cancer immunotherapy. J Hematol Oncol. (2024) 17(1):33. doi: 10.1186/s13045-024-01541-w
191. Chen L, Ishigami T, Nakashima-Sasaki R, Kino T, Doi H, Minegishi S, et al. Commensal microbe-specific activation of B2 cell subsets contributes to atherosclerosis development independently of lipid metabolism. EBioMedicine. (2016) 13:237–47. doi: 10.1016/j.ebiom.2016.10.030
192. Zhang ZB, Cheng YW, Xu L, Li JQ, Pan X, Zhu M, et al. Activation of β3-adrenergic receptor by mirabegron prevents aortic dissection/aneurysm by promoting lymphangiogenesis in perivascular adipose tissue. Cardiovasc Res. (2024) 120(17):2307–19. doi: 10.1093/cvr/cvae213
193. Chiabrando F, Lanzillotta M, Palumbo D, Pedica F, Caruso M, Capurso G, et al. Treating type 2 autoimmune pancreatitis with colchicine: a case series. Ann Intern Med. (2021) 174(12):1775–6. doi: 10.7326/L21-0281
194. Deftereos SG, Beerkens FJ, Shah B, Giannopoulos G, Vrachatis DA, Giotaki SG, et al. Colchicine in cardiovascular disease: in-depth review. Circulation. (2022) 145(1):61–78. doi: 10.1161/CIRCULATIONAHA.121.056171
195. Nidorf SM, Ben-Chetrit E, Ridker PM. Low-dose colchicine for atherosclerosis: long-term safety. Eur Heart J. (2024) 45(18):1596–601. doi: 10.1093/eurheartj/ehae208
196. Gollasch M. Adipose-vascular coupling and potential therapeutics. Annu Rev Pharmacol Toxicol. (2017) 57:417–36. doi: 10.1146/annurev-pharmtox-010716-104542
197. Lee YC, Chang HH, Chiang CL, Liu CH, Yeh JI, Chen MF, et al. Role of perivascular adipose tissue-derived methyl palmitate in vascular tone regulation and pathogenesis of hypertension. Circulation. (2011) 124(10):1160–71. doi: 10.1161/CIRCULATIONAHA.111.027375
198. Rendon CJ, Sempere L, Lauver A, Watts SW, Contreras GA. Anatomical location, sex, and age modulate adipocyte progenitor populations in perivascular adipose tissues. Front Physiol. (2024) 15:1411218. doi: 10.3389/fphys.2024.1411218
199. Victorio JA, Da Costa RM, Tostes RC, Davel AP. Modulation of vascular function by perivascular adipose tissue: sex differences. Curr Pharm Des. (2020) 26(30):3768–77. doi: 10.2174/1381612826666200701211912
200. Somani YB, Pawelczyk JA, De Souza MJ, Kris-Etherton PM, Proctor DN. Aging women and their endothelium: probing the relative role of estrogen on vasodilator function. Am J Physiol Heart Circ Physiol. (2019) 317(2):H395–404. doi: 10.1152/ajpheart.00430.2018
201. Tankó LB, Christiansen C. Adipose tissue, insulin resistance and low-grade inflammation: implications for atherogenesis and the cardiovascular harm of estrogen plus progestogen therapy. Climacteric. (2006) 9(3):169–80. doi: 10.1080/13697130600738765
202. Chen Y, Qin Z, Wang Y, Li X, Zheng Y, Liu Y. Role of inflammation in vascular disease-related perivascular adipose tissue dysfunction. Front Endocrinol (Lausanne). (2021) 12:710842. doi: 10.3389/fendo.2021.710842
203. Luo YL, Xu CF, Li HJ, Cao ZT, Liu J, Wang JL, et al. Macrophage-specific in vivo gene editing using cationic lipid-assisted polymeric nanoparticles. ACS Nano. (2018) 12(2):994–1005. doi: 10.1021/acsnano.7b07874
Keywords: perivascular adipose tissue, biomarkers, vascular inflammation, atherosclerosis, cardiovascular disease
Citation: Zhang S, Jiang J, Luo Y, Liu G, Hu S, Wan S, Luo C, Li H, Li N, da Graça Espírito Santo Vasconcelos J, Lavres Ceita de Carvalho L, Lima Bragança da Costa M, Takounjou JE, Maimite Das Neves K, dos Ramos da Conceição L, da Costa Encarnação M and Zhao L-Y (2025) Molecular crosstalk in perivascular adipose tissue: mechanisms of inflammation, metabolic dysregulation, and therapeutic opportunities in cardiovascular disease. Front. Cardiovasc. Med. 12:1613900. doi: 10.3389/fcvm.2025.1613900
Received: 28 April 2025; Accepted: 11 June 2025;
Published: 26 June 2025.
Edited by:
Hong Jin, Karolinska Institutet (KI), SwedenReviewed by:
Genmao Cao, Second Hospital of Shanxi Medical University, ChinaXinyi Li, Karolinska Institutet (KI), Sweden
Copyright: © 2025 Zhang, Jiang, Luo, Liu, Hu, Wan, Luo, Li, Li, da Graça Espírito Santo Vasconcelos, Lavres Ceita de Carvalho, Lima Bragança da Costa, Takounjou, Maimite Das Neves, dos Ramos da Conceição, da Costa Encarnação and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lin-Yong Zhao, MTUzNzk1MzUyQHNjdS5lZHUuY24=
†ORCID:
Lin-Yong Zhao
orcid.org/0000-0003-0884-4657