 Tomoyuki Nakano1
Tomoyuki Nakano1 Hirooki Matsui1
Hirooki Matsui1 Toshiaki Tanaka1Yasukazu Hozumi1Ken Iseki2Kaneyuki Kawamae3
Toshiaki Tanaka1Yasukazu Hozumi1Ken Iseki2Kaneyuki Kawamae3 Kaoru Goto1*
Kaoru Goto1*- 1Department of Anatomy and Cell Biology, Yamagata University School of Medicine, Yamagata, Japan
- 2Department of Emergency and Critical Care Medicine, Fukushima Medical University School of Medicine, Fukushima, Japan
- 3Department of Anesthesiology, Yamagata University School of Medicine, Yamagata, Japan
The endoplasmic reticulum (ER) comprises an interconnected membrane network, which is made up of lipid bilayer and associated proteins. This organelle plays a central role in the protein synthesis and sorting. In addition, it represents the synthetic machinery of phospholipids, the major constituents of the biological membrane. In this process, phosphatidic acid (PA) serves as a precursor of all phospholipids, suggesting that PA synthetic activity is closely associated with the ER function. One enzyme responsible for PA synthesis is diacylglycerol kinase (DGK) that phosphorylates diacylglycerol (DG) to PA. DGK is composed of a family of enzymes with distinct features assigned to each isozyme in terms of structure, enzymology, and subcellular localization. Of DGKs, DGKε uniquely exhibits substrate specificity toward arachidonate-containing DG and is shown to reside in the ER. Arachidonic acid, a precursor of bioactive eicosanoids, is usually acylated at the sn-2 position of phospholipids, being especially enriched in phosphoinositide. In this review, we focus on arachidonoyl-specific DGKε with respect to the historical context, molecular basis of the substrate specificity and ER-targeting, and functional implications in the ER.
Introduction
The endoplasmic reticulum (ER), which comprises a tubular and planar network of lipid bilayer membranes (Croze and Morré, 1984), represents a specialized site of protein synthesis and subsequent folding machinery. In addition, the ER plays a central role in the synthesis and transport of major membrane phospholipids such as phosphatidylcholine (PC), phosphatidylserine (PS), and phosphatidylinositol (PI; Gaspar et al., 2007). In response to cellular requirements, this tubular and planar ER network extends to all regions of cell interfaces at membrane contact sites with the plasma membrane, mitochondria, and Golgi apparatus for lipid transfer, integration of metabolic pathways, and calcium homeostasis (Lagace and Ridgway, 2013). In terms of energy homeostasis, fatty acids supply a major source of energy for organisms, but they can also be toxic. When exposed to excess fatty acids, cells esterify fatty acids into neutral lipids and package them into lipid droplets (LDs). Actually, an LD is an ER-derived organelle that is necessary for the storage and mobilization of neutral lipids in a specialized cell type: adipocyte (Martin and Parton, 2006; Brasaemle and Wolins, 2012). Under pathological conditions including nutrient and oxygen starvation, calcium depletion and altered redox status, protein folding, and lipid biosynthesis are impaired, thereby producing ER stress. Therefore, the ER integrates cellular activities of protein and lipid synthesis as well as pathological responses such as unfolding protein response (UPR; Berridge, 2002; Ron and Walter, 2007; Sano and Reed, 2013).
During the process of lipid synthesis, phosphatidic acid (PA) serves as an intermediate molecule for all phospholipids. It is therefore conceivable that PA synthetic activity is intimately involved in the ER function, and that one enzyme responsible for this activity is diacylglycerol kinase (DGK; Kanoh et al., 1990). Actually, DGK comprises a family of enzymes. Each of the isozymes exhibits a characteristic feature in terms of structural, enzymological, and morphological aspects (Goto et al., 2007; Sakane et al., 2007; Mérida et al., 2008; Topham and Epand, 2009; Table 1). Each member of the DGK family presents a unique subcellular localization in transfected cells and presumably plays a specific role at each site (Kobayashi et al., 2007). Of the DGKs, DGKε is unique in its substrate specificity toward arachidonate-containing DG and resides in the ER (Matsui et al., 2014). In this review, we specifically examine the functional role of DGKε in this organelle.
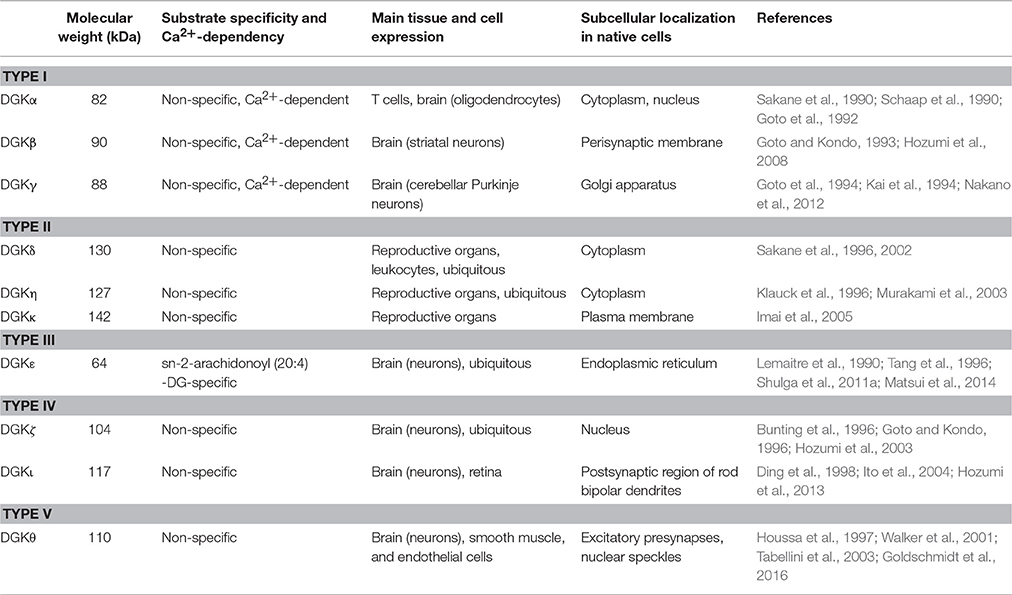
Table 1. Characteristic features of mammalian DGK isozymes.
Identification of Arachidonoyl DGK
Since the first discovery of DGK activity in a brain microsome fraction (Hokin and Hokin, 1959), it has been reported as distributed widely in animal tissues (Hokin and Hokin, 1963; Sastry and Hokin, 1966; Prottey and Hawthorne, 1967; Lapetina and Hawthorne, 1971; Farese et al., 1981). The DGK activity was associated with various fractions of cells, including soluble, membranous, and cytoskeletal fractions (Call and Rubert, 1973; Daleo et al., 1974). These features suggest the heterogeneity of DGK in animal tissues and cells. In this respect, Glomset group reported that cytosolic and membrane-bound DGKs in Swiss 3T3 cells show different substrate selectivity (MacDonald et al., 1988). Intriguingly, the membrane-bound DGK is unique in that it selectively catalyzes DG containing arachidonate at the sn-2 position (Lemaitre et al., 1990). Moreover, it is rapidly inactivated by preincubation with its preferred substrate. Generally, DGK activity is determined using several assay systems with different detergents, including octylglucoside mixed micelle assay, deoxycholate assay, and Triton X-100 assay (Walsh et al., 1994). It is particularly noteworthy that the detection of arachidonoyl-specificity depends on the assay system that is used. The substrate selectivity toward arachidonate-containing DG is most sensitive in the octylglucoside assay, but is not detected in the deoxycholate assay. The sensitive assay system together with presumed thermal lability made it difficult to purify the enzyme. Biochemical purification of this “arachidonoyl DGK” from bovine testis estimated the molecular mass as 58,000 (Walsh et al., 1994), although PCR cloning using degenerate primers succeeded in isolating the cDNA clone encoding arachidonoyl DGK, designated as DGKε (Tang et al., 1996).
Molecular Basis for Arachidonoyl Specificity of DGKε
DGKε is the only isozyme that shows substrate specificity toward arachidonate (20:4)-containing DG. As a substrate for DGKε, sn-1-stearoyl-2-arachidonoyl-DG (18:0/20:4-DG) is preferred over saturated DG (sn-1,2-didecanoyl-DG, 10:0/10:0-DG) or monounsaturared DG (sn-1,2-dioleoyl-DG, 18:1/18:1-DG). It should be mentioned that DGKε prefers 18:0/20:4-DG to sn-1-stearoyl-2-linoleoyl-DG (18:0/18:2-DG) and sn-1-stearoyl-2-docosahexaenoyl-DG (18:0/22:6-DG) (Lemaitre et al., 1990; Tang et al., 1996; Shulga et al., 2011a). Therefore, it is concluded that DGKε prefers arachidonate at the sn-2 position. Arachidonic acid, an essential polyunsaturated fatty acid, contains four double bonds. Arachidonate is not only a major component of membrane phospholipid; it is also the precursor of bioactive molecules designated as eicosanoids, such as prostaglandins and leukotrienes that are catalyzed, respectively, by cyclooxygenase (COX) and lipoxygenase (LOX; Funk, 2001; Buczynski et al., 2009). Because these arachidonate-derivatives serve as key mediators of several pathophysiological events, free arachidonate itself should be maintained within a restricted concentration (Peters-Golden and Henderson, 2007). Under physiological conditions, arachidonate is incorporated into the sn-2 position of phospholipids by the enzymes arachidonoyl-CoA synthetase and lysophospholipid acyltransferases (Pérez-Chacón et al., 2009). These enzymes, together with DGKε, specifically recognize arachidonate moiety. However, how is the arachidonoyl specificity achieved?
To investigate the molecular basis of the substrate specificity of DGKε toward arachidonate, the Epand group compared amino acid sequences of the enzymes that specifically recognize this fatty acid (Shulga et al., 2011b). They identified in the catalytic domain of DGKε (aa. 421–453 in human sequence) the motif L-X(3-4)-R-X(2)-L-X(4)-G, in which -X(n)- is n residues of any amino acid. This domain, which is contained in DGKε of various species as well as phosphatidylinositol-4-phosphate-5-kinase type Iα, resembles a polyunsaturated fatty acid-recognizing domain identified in lipoxygenases (Neau et al., 2009). Mutations of the essential residues in this motif, L431I and L438I, significantly reduce arachidonoyl specificity. Furthermore, the group found a sequence similar to this LOX-like motif in non-specific isozyme DGKα, with a V656 residue instead of Leu in DGKε. They confirm that V656L mutation introduces some specificity for arachidonate-containing DG to DGKα.
Targeting of DGKε to the ER
The DGK family is localized to distinct subcellular compartments in cDNA-transfected cells, including the cytoplasm, ER, Golgi complex, actin-cytoskeleton, and nucleus (Kobayashi et al., 2007). In an early fractionation study using Swiss 3T3 cells, “arachidonoyl DGK activity” comigrated with that of the ER marker enzymes, together with other PI-metabolizing enzyme PI synthase (Glomset, 1996). In agreement with the biochemical data, DGKε is targeted to the ER. Because DGKε is highly insoluble, the hydrophobic region was presumed to play a key role in the ER targeting. We investigated the sequence responsible for ER targeting of DGKε (Matsui et al., 2014). Various deletion and substitution mutations of rat DGKε tagged with GFP were transfected in cells and were compared with ER markers. Results show clearly that a stretch of hydrophobic amino acid sequence 20–40 (DGKε 20–40) in the N-terminus is a determinant sequence in controlling the ER targeting of DGKε. This hydrophobic region adopts an α-helical structure of the transmembrane segment (Glukhov et al., 2007).
In this regard, a detailed modeling study suggests the possibility that this sequence structure can take two representative models of low-energy conformations, such as a long straight helix and a U-bend helix (Decaffmeyer et al., 2008). Two interchangeable structures of monotopic and bitopic nature might confer on DGKε a unique feature in relation to the ER. Changing conditions such as a redox state and pH can regulate the conformation of DGKε between these two structures, thereby affecting the relation of DGKε and the ER membrane.
The α-helical structure of the hydrophobic sequence 20–40 creates a “hydrophobic patch” composed of L22, L25, and L29 (according to the rat sequence; Figure 1). To test whether the hydrophobicity is critical in the ER targeting, we produced two substitution mutants: one containing less hydrophobic Ala (hydrophobic score 1.8) and the other with hydrophilic Gln (hydrophobic score -3.5), instead of wild-type Leu (hydrophobic score 3.8; (Matsui et al., 2014)). Ala substitution fragment DGKε (20–40/L22A, L25A, L29A) is targeted to the ER membrane. It is recovered in the membrane fraction, along with wild-type fragment. However, Gln substitution fragment DGKε (20–40/L22G, L25G, L29G) containing a “hydrophilic patch” is distributed diffusely in the cytoplasm and is recovered in the soluble fraction. Furthermore, full-length Ala mutant DGKε (L22A, L25A, L29A) is shown to reside in the ER whereas Gln mutant DGKε (L22G, L25G, L29G) abolishes it. These findings suggest that the hydrophobic patch composed of L22, L25, and L29 is crucially important for ER targeting of DGKε.
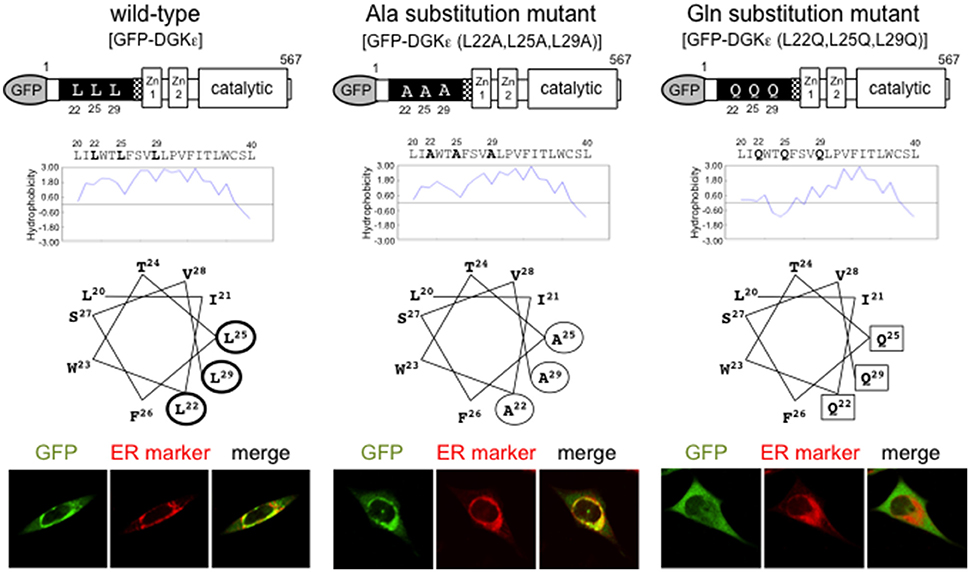
Figure 1. Features of wild-type DGKε and Ala and Gln substitution mutants. Hydropathy plot analysis and helical wheel projections of the hydrophobic region of DGKε are shown. Wild-type DGKε contains highly hydrophobic residues L22, L25, L29 (thick circles), which represent a prominent hydrophobic patch. Alanine substitution mutant DGKε (L22A, L25A, L29A) have a reduced hydrophobic patch (thin circles), whereas glutamine substitution mutant DGKε (L22Q, L25Q, L29Q) includes a hydrophilic patch (squares). Immunofluorescence images of GFP for full-length wild-type and substitution mutants of DGKε together with ER marker staining in transfected HeLa cells (lower panels). Wild-type DGKε and alanine mutant DGKε (L22A, L25A, L29A) clearly colocalize with ER marker, whereas glutamine mutant DGKε (L22Q, L25Q, L29Q) shows diffuse cytoplasmic pattern. Modified from Matsui et al. (2014) with permission from Elsevier.
No general consensus sequence for ER localization, such as the ER retention signal, is found in this hydrophobic or in other regions of DGKε (Matsui et al., 2014). Therefore, details of the ER targeting mechanism of DGKε remain unclear. How does this hydrophobic patch specifically lead DGKε to the ER, instead of the other membranes such as mitochondrial membrane? The ER consists of phospholipid bilayer containing a plethora of proteins. Is the membrane or the protein of the ER recognized by the hydrophobic patch? Does the hydrophobic patch bind to some microdomain of the membrane? Because DGKε-kinase dead mutant also resides in the ER, the substrate DG and the product PA are not involved in subcellular localization of DGKε. Therefore, the current data can be summarized as follows: ER targeting is mediated through the N-terminal hydrophobic patch composed of L22, L25, and L29. Subsequent recognition of the arachidonoyl acyl chain of DG is achieved by a LOX-like motif in the catalytic domain of DGKε (aa. 421–453). Additional studies must be conducted to elucidate the ER targeting mechanism of DGKε.
ER Stress
ER homeostasis is crucially important for cellular activity and survival (Ellgaard and Helenius, 2003). Stress in the ER induces the UPR, which represents a complex signaling system that controls translation and transcription in response to increased demands on the protein folding capacity of the ER for cell survival (Rutkowski and Kaufman, 2004; Koumenis and Wouters, 2006; Malhotra and Kaufman, 2007; Hetz, 2012). To meet this demand, the UPR coordinates membrane growth and phospholipid metabolism, thereby leading to ER membrane expansion and enhanced protein folding capacity (Sriburi et al., 2004). In addition to misfolding or incomplete assembly of proteins, alteration of the ER lipid composition also is shown to initiate ER stress (Devries-Seimon et al., 2005), indicating that disruption of membrane lipid homeostasis triggers directly or indirectly a mechanism to reestablish ER lipid composition (Fagone and Jackowski, 2009).
Under ER stress conditions, the glucose-regulated protein GRP78 plays a key role in UPR (Bertolotti et al., 2000). GRP78, a member of the heat shock protein 70 superfamily, serves as a major ER chaperone protein with ATPase activity. It is a key regulator of the transmembrane ER stress sensors comprised of inositol requiring enzyme 1 (IRE1), protein kinase RNA-activated (PKR)-like ER kinase (PERK), and activating transcription factor-6 (ATF6) (Lee, 2014). IRE1 is a transmembrane ribonuclease that splices and activates X-box-binding protein (XBP-1) mRNA. Spliced form of XBP-1 [XBP-1(S)], together with cleaved ATF6 and ATF4, translocates to the nucleus where they induce the expression of ER stress-responsive genes (Hetz, 2012). In this regard, XBP-1(S) serves as a regulator of PC synthesis and ER membrane development (Fagone and Jackowski, 2009). PERK mediates activation of the pro-apoptotic factor C/EBP homologous protein (CHOP), thereby leading to apoptosis if the response is insufficient to reestablish homeostasis (Xu et al., 2005; Shore et al., 2011).
Therefore, the ER membrane expansion is supported by phospholipid synthesis, in which PA serves as an intermediate product. The initial step in the PA synthesis is catalyzed using a family of glycerol 3-phosphate acyltransferases located in the ER and the outer mitochondrial membrane, followed by acyl-CoA-dependent acylation of lyso-PA to form PA (Lagace and Ridgway, 2013).
Another intermediate product DG is a precursor for PA, which is catalyzed by DGK. Therefore, DGK is intimately involved in phospholipid synthesis in the ER and presumably in the UPR. Earlier, we examined whether ER-resident DGKε participates in this process and assessed the ER stress pathways in DGKε knockdown cells under experimental ER stress conditions using tunicamycin and thapsigargin (Matsui et al., 2014). From DGKε deletion experiments conducted under ER stress conditions, we found the following: (1) The major protein chaperone GRP78 is induced to the same extent in both wild-type and DGKε-deficient cells. (2) Eukaryotic initiation factor 2α (eIF2α) is slightly, although not significantly, downregulated at the total and phosphorylated protein levels. (3) CHOP is significantly suppressed at the protein level. Analysis of cellular vulnerability, however, clearly shows that DGKε deletion reduces cell viability under ER stress conditions to some degree. Therefore, DGKε deletion seems to exert conflicting effects on apoptosis in terms of CHOP expression. In this regard, recent studies suggest that although CHOP is identified originally as a repressive member of the C/EBP family of transcription factors (Ron and Habener, 1992), it is capable of either transcriptional repression or activation, depending on the context (Oyadomari and Mori, 2004). Further studies need to be done to elucidate this point.
Concluding Remarks
Gene duplication contributes to the evolution of living creatures by expanding DNA information. Escherichia coli is equipped with two forms of DGK (Van Horn and Sanders, 2012; Jennings et al., 2015) whereas mammalian cells contain at least 10 isozymes. In the course of evolution, one branch of the diversified DGKs might have gained substrate specificity toward arachidonate-containing DG. Of DGs, arachidonoyl DG is phosphorylated selectively by arachidonoyl DGK to produce corresponding PA, which is further incorporated into inositol phospholipids. Multiple steps of this process are expected to enrich arachidonate in PI (Glomset, 1996). Because PIP2 is a major substrate for PLC, its enzymatic action results in the production of arachidonoyl DG. Functional implication of arachidonoyl DGK is suggested by an experimental seizure model at the organismal level. It reveals that DGKε-KO mice show lower degradation of brain PIP2 and lower accumulation of arachidonoyl-DG and free arachidonate although resting levels of PIP and PIP2 are similar between wild-type and DGKε-KO mice brains (Rodriguez de Turco et al., 2001). These findings suggest that DGKε is clearly involved in recycling PI metabolism presumably through the enrichment of arachidonate moiety. Therefore, DGKε downregulation may lead to a slowdown of PI turnover, thereby downregulating various membrane functions. In addition, it is particularly noteworthy that arachidonoyl DG also represents a substrate for another enzyme DG lipase, which cleaves sn-1-acyl chain to produce 2-arachidonoyl glycerol (Maejima et al., 2005). Because 2-arachidonoyl glycerol serves as an endocannabinoid for retrograde synaptic transmission, arachidonoyl DG is located at the crossroad of the two signaling cascades: DG-PA and DG-2AG pathways directed respectively by DGK and DG lipase. The mechanisms for how these signaling pathways are coordinated in parallel are just beginning to be explored.
Author Contributions
TN, HM, TT, YH, KI, and KK did the experiments in the original papers and summarized the results for the mini review. TN and KG constituted and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study is dedicated to Dr. John A. Glomset, University of Washington. Dr. Glomset reported for the first time the enzyme that preferentially phosphorylates arachidonoyl-DG and investigated the functional implication of arachidonate at the sn-2 position of phosphoinositide. This work was supported by Grants-in-Aid from The Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan [grant numbers 24390044, 26670782 (KG), 26460265 (TN), 15K20178 (HM), and 15K10530 (KK)].
References
Berridge, M. J. (2002). The endoplasmic reticulum: a multifunctional signaling organelle. Cell Calcium 32, 235–249. doi: 10.1016/S0143416002001823
Bertolotti, A., Zhang, Y., Hendershot, L. M., Harding, H. P., and Ron, D. (2000). Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2, 326–332. doi: 10.1038/35014014
Brasaemle, D. L., and Wolins, N. E. (2012). Packaging of fat: an evolving model of lipid droplet assembly and expansion. J. Biol. Chem. 287, 2273–2279. doi: 10.1074/jbc.R111.309088
Buczynski, M. W., Dumlao, D. S., and Dennis, E. A. (2009). Thematic review series: proteomics. An integrated omics analysis of eicosanoid biology. J. Lipid Res. 50, 1015–1038. doi: 10.1194/jlr.R900004-JLR200
Bunting, M., Tang, W., Zimmerman, G. A., McIntyre, T. M., and Prescott, S. M. (1996). Molecular cloning and characterization of a novel human diacylglycerol kinase zeta. J. Biol. Chem. 271, 10230–10236. doi: 10.1074/jbc.271.17.10237
Call, F. L. II, Rubert, M. (1973). Diglyceride kinase in human platelets. J. Lipid Res. 14, 466–474.
Croze, E. M., and Morré, D. J. (1984). Isolation of plasma membrane, golgi apparatus, and endoplasmic reticulum fractions from single homogenates of mouse liver. J. Cell. Physiol. 119, 46–57. doi: 10.1002/jcp.1041190109
Daleo, G. R., Piras, M. M., and Piras, R. (1974). The presence of phospholipids and diglyceride kinase activity in microtubules from different tissues. Biochem. Biophys. Res. Commun. 61, 1043–1050. doi: 10.1016/0006-291X(74)90260-5
Decaffmeyer, M., Shulga, Y. V., Dicu, A. O., Thomas, A., Truant, R., Topham, M. K., et al. (2008). Determination of the topology of the hydrophobic segment of mammalian diacylglycerol kinase epsilon in a cell membrane and its relationship to predictions from modeling. J. Mol. Biol. 383, 797–809. doi: 10.1016/j.jmb.2008.08.076
Devries-Seimon, T., Li, Y., Yao, P. M., Stone, E., Wang, Y., Davis, R. J., et al. (2005). Cholesterol-induced macrophage apoptosis requires ER stress pathways and engagement of the type A scavenger receptor. J. Cell Biol. 171, 61–73. doi: 10.1083/jcb.200502078
Ding, L., Traer, E., McIntyre, T. M., Zimmerman, G. A., and Prescott, S. M. (1998). The cloning and characterization of a novel human diacylglycerol kinase, DGKiota. J. Biol. Chem. 273, 32746–32752. doi: 10.1074/jbc.273.49.32746
Ellgaard, L., and Helenius, A. (2003). Quality control in the endoplasmic reticulum. Nat. Rev. Mol. Cell Biol. 4, 181–191. doi: 10.1038/nrm1052
Fagone, P., and Jackowski, S. (2009). Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 50, S311–S316. doi: 10.1194/jlr.R800049-JLR200
Farese, R. V., Sabir, M. A., and Larson, R. E. (1981). Effects of adrenocorticotropin and cycloheximide on adrenal diglyceride kinase. Biochemistry 20, 6047–6051. doi: 10.1021/bi00524a020
Funk, C. D. (2001). Prostaglandins and leukotrienes: advances in eicosanoid biology. Science 294, 1871–1875. doi: 10.1126/science.294.5548.1871
Gaspar, M. L., Aregullin, M. A., Jesch, S. A., Nunez, L. R., Villa-García, M., and Henry, S. A. (2007). The emergence of yeast lipidomics. Biochim. Biophys. Acta 1771, 241–254. doi: 10.1016/j.bbalip.2006.06.011
Glomset, J. A. (1996). “A branched metabolic pathway in animal cells converts 2-monoacylglycerol into sn-1-stearoyl-2-arachdonoyl phosphatidylinositol and other phosphoglycerides,” in Advances in Lipobiology, ed R. W. Cross (Greemwich, CT: JAI Press), 61–100.
Glukhov, E., Shulga, Y. V., Epand, R. F., Dicu, A. O., Topham, M. K., Deber, C. M., et al. (2007). Membrane interactions of the hydrophobic segment of diacylglycerol kinase epsilon. Biochim. Biophys. Acta 1768, 2549–2558. doi: 10.1016/j.bbamem.2007.06.012
Goldschmidt, H. L., Tu-Sekine, B., Volk, L., Anggono, V., Huganir, R. L., and Raben, D. M. (2016). DGKtheta catalytic activity is required for efficient recycling of presynaptic vesicles at excitatory synapses. Cell Rep. 14, 200–207. doi: 10.1016/j.celrep.2015.12.022
Goto, K., Funayama, M., and Kondo, H. (1994). Cloning and expression of a cytoskeleton-associated diacylglycerol kinase that is dominantly expressed in cerebellum. Proc. Natl. Acad. Sci. U.S.A. 91, 13042–13046. doi: 10.1073/pnas.91.26.13042
Goto, K., Hozumi, Y., Nakano, T., Saino, S. S., and Kondo, H. (2007). Cell biology and pathophysiology of the diacylglycerol kinase family: morphological aspects in tissues and organs. Int. Rev. Cytol. 264, 25–63. doi: 10.1016/S0074-7696(07)64002-9
Goto, K., and Kondo, H. (1993). Molecular cloning and expression of a 90-kDa diacylglycerol kinase that predominantly localizes in neurons. Proc. Natl. Acad. Sci. U.S.A. 90, 7598–7602. doi: 10.1073/pnas.90.16.7598
Goto, K., and Kondo, H. (1996). A 104-kDa diacylglycerol kinase containing ankylin-like repeats localizes in the cell nucleus. Proc. Natl. Acad. Sci. U.S.A. 93, 11196–11201. doi: 10.1073/pnas.93.20.11196
Goto, K., Watanabe, M., Kondo, H., Yuasa, H., Sakane, F., and Kanoh, H. (1992). Gene cloning, sequence, expression and in situ localization of 80kDa diacylglycerol kinase specific to oligodendrocyte of rat brain. Mol. Brain Res. 16, 75–87.
Hetz, C. (2012). The unfolded protein response: controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 13, 89–102. doi: 10.1038/nrm3270
Hokin, L. E., and Hokin, M. R. (1963). Diglyceride kinase and other pathways for phosphatidic acid synthesis in the erythrocyte membrane. Biochim. Biophys. Acta 67, 470–484. doi: 10.1016/0926-6569(63)90263-3
Hokin, M. R., and Hokin, L. E. (1959). The synthesis of phosphatidic acid from diglyceride and adenosine triphosphate in extracts of brain microsomes. J. Biol. Chem. 234, 1381–1386.
Houssa, B., Schaap, D., van der Val, J., Goto, K., Kondo, H., Yamakawa, A., et al. (1997). Cloning of a novel human diacylglycerol kinase (DGKtheta) containing three cysteine-rich domains, a proline-rich region, and a pleckstrin homology domain with an overlapping Ras-associating domain. J. Biol. Chem. 272, 10422–10428. doi: 10.1074/jbc.272.16.10422
Hozumi, Y., Fukaya, M., Adachi, N., Saito, N., Otani, K., Kondo, H., et al. (2008). Diacylglycerol kinase beta accumulates on the perisynaptic site of medium spiny neurons in the striatum. Eur. J. Neurosci. 28, 2409–2422. doi: 10.1111/j.1460-9568.2008.06547.x
Hozumi, Y., Ito, T., Nakano, T., Nakagawa, T., Aoyagi, M., Kondo, H., et al. (2003). Nuclear localization of diacylglycerol kinase ζ in neurons. Eur. J. Neurosci. 18, 1448–1457. doi: 10.1046/j.1460-9568.2003.02871.x
Hozumi, Y., Matsui, H., Sakane, F., Watanabe, M., and Goto, K. (2013). Distinct Expression and Localization of Diacylglycerol Kinase Isozymes in Rat Retina. J. Histochem. Cytochem. 61, 462–476. doi: 10.1369/0022155413483574
Imai, S., Kai, M., Yasuda, S., Kanoh, H., and Sakane, F. (2005). Identification and characterization of a novel human type II diacylglycerol kinase, DGKκ. J. Biol. Chem. 280, 39870–39881. doi: 10.1074/jbc.M500669200
Ito, T., Hozumi, Y., Sakane, F., Saino-Saito, S., Kanoh, H., Aoyagi, M., et al. (2004). Cloning and characterization of diacylglycerol kinase iota splice variants in rat brain. J. Biol. Chem. 279, 23317–23326. doi: 10.1074/jbc.M312976200
Jennings, W., Doshi, S., D'souza, K., and Epand, R. M. (2015). Molecular properties of diacylglycerol kinase-epsilon in relation to function. Chem. Phys. Lipids 192, 100–108. doi: 10.1016/j.chemphyslip.2015.06.003
Kai, M., Sakane, F., Imai, S., Wada, I., and Kanoh, H. (1994). Molecular cloning of a diacylglycerol kinase isozyme predominantly expressed in human retina with a truncated and inactive enzyme expression in most other human cells. J. Biol. Chem. 269, 18492–18498.
Kanoh, H., Yamada, K., and Sakane, F. (1990). Diacylglycerol kinase: a key modulator of signal transduction? Trends Biochem. Sci. 15, 47–50. doi: 10.1016/0968-0004(90)90172-8
Klauck, T. M., Xu, X., Mousseau, B., and Jaken, S. (1996). Cloning and characterization of a glucocorticoid-induced diacylglycerol kinase. J. Biol. Chem. 271, 19781–19788. doi: 10.1074/jbc.271.33.19781
Kobayashi, N., Hozumi, Y., Ito, T., Hosoya, T., Kondo, H., and Goto, K. (2007). Differential subcellular targeting and activity-dependent subcellular localization of diacylglycerol kinase isozymes in transfected cells. Eur. J. Cell Biol. 86, 433–444. doi: 10.1016/j.ejcb.2007.05.002
Koumenis, C., and Wouters, B. G. (2006). “Translating” tumor hypoxia: unfolded protein response (UPR)-dependent and UPR-independent pathways. Mol. Cancer Res. 4, 423–436. doi: 10.1158/1541-7786.MCR-06-0150
Lagace, T. A., and Ridgway, N. D. (2013). The role of phospholipids in the biological activity and structure of the endoplasmic reticulum. Biochim. Biophys. Acta 1833, 2499–2510. doi: 10.1016/j.bbamcr.2013.05.018
Lapetina, E. G., and Hawthorne, J. N. (1971). The diglyceride kinase of rat cerebral cortex. Biochem. J. 122, 171–179. doi: 10.1042/bj1220171
Lee, A. S. (2014). Glucose-regulated proteins in cancer: molecular mechanisms and therapeutic potential. Nat. Rev. Cancer 14, 263–276. doi: 10.1038/nrc3701
Lemaitre, R. N., King, W. C., MacDonald, M. L., and Glomset, J. A. (1990). Distribution of distinct arachidonoyl-specific and non-specific isoenzymes of diacylglycerol kinase in baboon (Papio cynocephalus) tissues. Biochem. J. 266, 291–299. doi: 10.1042/bj2660291
MacDonald, M. L., Mack, K. F., Williams, B. W., King, W. C., and Glomset, J. A. (1988). A membrane-bound diacylglycerol kinase that selectively phosphorylates arachidonoyl-diacylglycerol. Distinction from cytosolic diacylglycerol kinase and comparison with the membrane-bound enzyme from Escherichia coli. J. Biol. Chem. 263, 1584–1592.
Maejima, T., Oka, S., Hashimotodani, Y., Ohno-Shosaku, T., Aiba, A., Wu, D., et al. (2005). Synaptically driven endocannabinoid release requires Ca2+-assisted metabotropic glutamate receptor subtype 1 to phospholipase Cbeta4 signaling cascade in the cerebellum. J. Neurosci. 25, 6826–6835. doi: 10.1523/JNEUROSCI.0945-05.2005
Malhotra, J. D., and Kaufman, R. J. (2007). The endoplasmic reticulum and the unfolded protein response. Semin. Cell Dev. Biol. 18, 716–731. doi: 10.1016/j.semcdb.2007.09.003
Martin, S., and Parton, R. G. (2006). Lipid droplets: a unified view of a dynamic organelle. Nat. Rev. Mol. Cell Biol. 7, 373–378. doi: 10.1038/nrm1912
Matsui, H., Hozumi, Y., Tanaka, T., Okada, M., Nakano, T., Suzuki, Y., et al. (2014). Role of the N-terminal hydrophobic residues of DGKepsilon in targeting the endoplasmic reticulum. Biochim. Biophys. Acta 1842, 1440–1450. doi: 10.1016/j.bbalip.2014.07.007
Mérida, I., Avila-Flores, A., and Merino, E. (2008). Diacylglycerol kinases: at the hub of cell signalling. Biochem. J. 409, 1–18. doi: 10.1042/BJ20071040
Murakami, T., Sakane, F., Imai, S., Houkin, K., and Kanoh, H. (2003). Identification and characterization of two splice variants of human diacylglycerol kinase eta. J. Biol. Chem. 278, 34364–34372. doi: 10.1074/jbc.M301542200
Nakano, T., Hozumi, Y., Goto, K., and Wakabayashi, I. (2012). Involvement of diacylglycerol kinase gamma in modulation of iNOS synthesis in Golgi apparatus of vascular endothelial cells. Naunyn Schmiedebergs Arch. Pharmacol. 385, 787–795. doi: 10.1007/s00210-012-0760-0
Neau, D. B., Gilbert, N. C., Bartlett, S. G., Boeglin, W., Brash, A. R., and Newcomer, M. E. (2009). The 1.85 Å structure of an 8R-lipoxygenase suggests a general model for lipoxygenase product specificity. Biochemistry 48, 7906–7915. doi: 10.1021/bi900084m
Oyadomari, S., and Mori, M. (2004). Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 11, 381–389. doi: 10.1038/sj.cdd.4401373
Pérez-Chacón, G., Astudillo, A. M., Balgoma, D., Balboa, M. A., and Balsinde, J. (2009). Control of free arachidonic acid levels by phospholipases A2 and lysophospholipid acyltransferases. Biochim. Biophys. Acta 1791, 1103–1113. doi: 10.1016/j.bbalip.2009.08.007
Peters-Golden, M., and Henderson, W. R. Jr. (2007). Leukotrienes. N. Engl. J Med. 357, 1841–1854. doi: 10.1056/NEJMra071371
Prottey, C., and Hawthorne, J. N. (1967). The biosynthesis of phosphatidic acid and phosphatidylinositol in mammalian pancreas. Biochem. J. 105, 379–392. doi: 10.1042/bj1050379
Rodriguez de Turco, E. B., Tang, W., Topham, M. K., Sakane, F., Marcheselli, V. L., Chen, C., et al. (2001). Diacylglycerol kinase epsilon regulates seizure susceptibility and long-term potentiation through arachidonoyl- inositol lipid signaling. Proc. Natl. Acad. Sci. U.S.A. 98, 4740–4745. doi: 10.1073/pnas.081536298
Ron, D., and Habener, J. F. (1992). CHOP, a novel developmentally regulated nuclear protein that dimerizes with transcription factors C/EBP and LAP and functions as a dominant-negative inhibitor of gene transcription. Genes Dev. 6, 439–453. doi: 10.1101/gad.6.3.439
Ron, D., and Walter, P. (2007). Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 8, 519–529. doi: 10.1038/nrm2199
Rutkowski, D. T., and Kaufman, R. J. (2004). A trip to the ER: coping with stress. Trends Cell Biol. 14, 20–28. doi: 10.1016/j.tcb.2003.11.001
Sakane, F., Imai, S., Kai, M., Wada, I., and Kanoh, H. (1996). Molecular cloning of a novel diacylglycerol kinase isozyme with a pleckstrin homology domain and a C-terminal tail similar to those of the EPH family of protein tyrosine kinase. J. Biol. Chem. 271, 8394–8401. doi: 10.1074/jbc.271.14.8394
Sakane, F., Imai, S., Kai, M., Yasuda, S., and Kanoh, H. (2007). Diacylglycerol kinases: why so many of them? Biochim. Biophys. Acta 1771, 793–806. doi: 10.1016/j.bbalip.2007.04.006
Sakane, F., Imai, S., Yamada, K., Murakami, T., Tsushima, S., and Kanoh, H. (2002). Alternative splicing of the human diacylglycerol kinase delta gene generates two isoforms differing in their expression patterns and in regulatory functions. J. Biol. Chem. 277, 43519–43526. doi: 10.1074/jbc.M206895200
Sakane, F., Yamada, K., Kanoh, H., Yokoyama, C., and Tanabe, T. (1990). Porcine diacylglycerol kinase sequence has zinc finger and E–F handmotifs. Nature 344, 345–348. doi: 10.1038/344345a0
Sano, R., and Reed, J. C. (2013). ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 1833, 3460–3470. doi: 10.1016/j.bbamcr.2013.06.028
Sastry, P. S., and Hokin, L. E. (1966). Studies on the role of phospholipids in phagocytosis. J. Biol. Chem. 241, 3354–3361.
Schaap, D., de Widt, J., van der Wal, J., Vandekerckhove, J., van Damme, J., Gussow, D., et al. (1990). Purification, cDNA-cloning and expression of human diacylglycerol kinase. FEBS Lett. 275, 151–158. doi: 10.1016/0014-5793(90)81461-V
Shore, G. C., Papa, F. R., and Oakes, S. A. (2011). Signaling cell death from the endoplasmic reticulum stress response. Curr. Opin. Cell Biol. 23, 143–149. doi: 10.1016/j.ceb.2010.11.003
Shulga, Y. V., Topham, M. K., and Epand, R. M. (2011a). Substrate specificity of diacylglycerol kinase-epsilon and the phosphatidylinositol cycle. FEBS Lett. 585, 4025–4028. doi: 10.1016/j.febslet.2011.11.016
Shulga, Y. V., Topham, M. K., and Epand, R. M. (2011b). Study of arachidonoyl specificity in two enzymes of the PI cycle. J. Mol. Biol. 409, 101–112. doi: 10.1016/j.jmb.2011.03.071
Sriburi, R., Jackowski, S., Mori, K., and Brewer, J. W. (2004). XBP1: a link between the unfolded protein response, lipid biosynthesis, and biogenesis of the endoplasmic reticulum. J. Cell Biol. 167, 35–41. doi: 10.1083/jcb.200406136
Tabellini, G., Bortul, R., Santi, S., Riccio, M., Baldini, G., Cappellini, A., et al. (2003). Diacylglycerol kinase-θ is localized in the speckle domains of the nucleus. Exp. Cell Res. 287, 143–154. doi: 10.1016/S0014-4827(03)00115-0
Tang, W., Bunting, M., Zimmerman, G. A., McIntyre, T. M., and Prescott, S. M. (1996). Molecular cloning of a novel human diacylglycerol kinase highly selective for arachidonate-containing substrates. J. Biol. Chem. 271, 10237–10241. doi: 10.1074/jbc.271.17.10230
Topham, M. K., and Epand, R. M. (2009). Mammalian diacylglycerol kinases: molecular interactions and biological functions of selected isoforms. Biochim. Biophys. Acta 1790, 416–424. doi: 10.1016/j.bbagen.2009.01.010
Van Horn, W. D., and Sanders, C. R. (2012). Prokaryotic diacylglycerol kinase and undecaprenol kinase. Annu. Rev. Biophys. 41, 81–101. doi: 10.1146/annurev-biophys-050511-102330
Walker, A. J., Draeger, A., Houssa, B., van Blitterswijk, W. J., Ohanian, V., and Ohanian, J. (2001). Diacylglycerol kinase theta is translocated and phosphoinositide 3-kinase-dependently activated by noradrenaline but not angiotensin II in intact small arteries. Biochem. J. 353, 129–137. doi: 10.1042/bj3530129
Walsh, J. P., Suen, R., Lemaitre, R. N., and Glomset, J. A. (1994). Arachidonoyl-diacylglycerol kinase from bovine testis. Purification and properties. J. Biol. Chem. 269, 21155–21164.
Keywords: diacylglycerol kinase, arachidonate, substrate specificity, endoplasmic reticulum, phosphoinositide, ER stress
Citation: Nakano T, Matsui H, Tanaka T, Hozumi Y, Iseki K, Kawamae K and Goto K (2016) Arachidonoyl-Specific Diacylglycerol Kinase ε and the Endoplasmic Reticulum. Front. Cell Dev. Biol. 4:132. doi: 10.3389/fcell.2016.00132
Received: 03 August 2016; Accepted: 31 October 2016;
Published: 18 November 2016.
Edited by:
Isabel Merida, Spanish National Research Council, SpainReviewed by:
Uwe Lendeckel, University of Greifswald, GermanyFrank Thévenod, Witten/Herdecke University, Germany
Copyright © 2016 Nakano, Matsui, Tanaka, Hozumi, Iseki, Kawamae and Goto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaoru Goto, a2dvdG9AbWVkLmlkLnlhbWFnYXRhLXUuYWMuanA=