 Margarida Beatriz1,2†
Margarida Beatriz1,2† Rita Vilaça
Rita Vilaça Carla Lopes
Carla Lopes- 1CNC-Center for Neuroscience and Cell Biology, University of Coimbra, Coimbra, Portugal
- 2IIIUC-Institute for Interdisciplinary Research, University of Coimbra, Coimbra, Portugal
Extracellular vesicles (EVs) are nano-sized membrane-enclosed particles released by cells that participate in intercellular communication through the transfer of biologic material. EVs include exosomes that are small vesicles that were initially associated with the disposal of cellular garbage; however, recent findings point toward a function as natural carriers of a wide variety of genetic material and proteins. Indeed, exosomes are vesicle mediators of intercellular communication and maintenance of cellular homeostasis. The role of exosomes in health and age-associated diseases is far from being understood, but recent evidence implicates exosomes as causative players in the spread of neurodegenerative diseases. Cells from the central nervous system (CNS) use exosomes as a strategy not only to eliminate membranes, toxic proteins, and RNA species but also to mediate short and long cell-to-cell communication as carriers of important messengers and signals. The accumulation of protein aggregates is a common pathological hallmark in many neurodegenerative diseases, including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, amyotrophic lateral sclerosis, and prion diseases. Protein aggregates can be removed and delivered to degradation by the endo-lysosomal pathway or can be incorporated in multivesicular bodies (MVBs) that are further released to the extracellular space as exosomes. Because exosome transport damaged cellular material, this eventually contributes to the spread of pathological misfolded proteins within the brain, thus promoting the neurodegeneration process. In this review, we focus on the role of exosomes in CNS homeostasis, their possible contribution to the development of neurodegenerative diseases, the usefulness of exosome cargo as biomarkers of disease, and the potential benefits of plasma circulating CNS-derived exosomes.
Introduction
Extracellular vesicles (EVs) are secreted by all types of cells, including neuronal cells, and can be found in almost all body fluids, including urine, saliva, blood, and cerebrospinal fluid (CSF) (Faure et al., 2006; Yanez-Mo et al., 2015). EVs can be divided according to their size and mechanism of release. In 2018, the International Society for Extracellular Vesicles (ISEV) endorsed the use of a new standardized nomenclature considering terms for subtypes that refer to physical properties such as size (small < 200 nm) or biochemical composition, among others (Théry et al., 2018). Apoptotic bodies and microvesicles (>100 nm in diameter) are released directly from the outward budding of the plasma membrane, while exosomes (30–150 nm in diameter, most common size described in the literature) are released after fusion of endosome-derived vesicles with the plasma membrane (Figure 1). Thus, different pools of EVs are characterized based on distinct intracellular origins and biogenesis (Kowal et al., 2014). Exosome biogenesis and secretion are modulated by different cellular stress and/or pathological conditions (Raposo and Stoorvogel, 2013; Record et al., 2014; Becker et al., 2016). These vesicles act as selective transporters of proteins, lipids, and genetic material, thus constituting an important intercellular communication system, capable of influencing distinct cellular activities in the recipient cells (Mathivanan et al., 2010; Yanez-Mo et al., 2015). The growing interest in EVs has been reflected in the creation of distinct databases that compile data on exosome content such as Exocarta (Mathivanan and Simpson, 2009), miRandola (Russo et al., 2018), EVpedia (Kim et al., 2013), and Vesiclepedia (Kalra et al., 2012), which are constantly updated with released studies. For the scope of this review, we will focus on exosomes and their biological function in central nervous system (CNS) homeostasis and discuss the role, as biomarkers, in the context of neurological disorders, particularly CNS-derived exosomes that can be isolated from the blood.
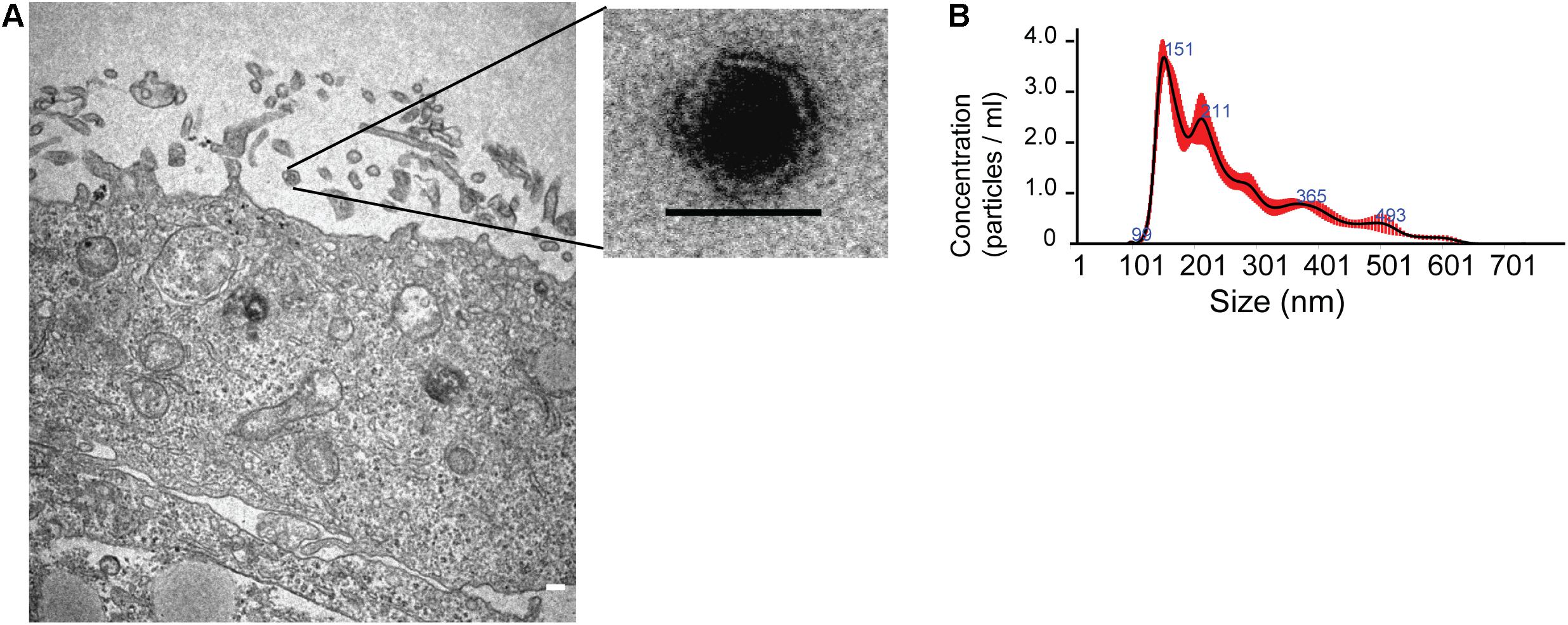
Figure 1. Characterization of exosomes released from human neural stem cells differentiated from Huntington’s Disease iPS cells. (A) Left: Electron micrographs of exosomes released from neural stem cell membrane, scale 200 nm. Right: Exosome with a lipid bilayer membrane sizing 50 nm. (B) Nanoparticle Tracking Analysis (NTA) of exosomes with sizes ranging from 67.7 to 152.1 nm (10th; 90th percentile) cited from Lopes et al. (2020).
Exosome Biogenesis and Cargo Loading
Exosomes originate during the maturation of early endosomes through the inward budding of the endosomal membrane that generates intraluminal vesicles (ILVs), resulting in the formation of late endosomes (LEs) with multiple small vesicles termed MVB (Huotari and Helenius, 2011). During this process, cytosolic portions, proteins, and nucleic acids are enveloped into the vesicles, and the resulting MVBs can be either targeted to lysosomal degradation or fused with the plasma membrane, releasing the ILVs to the extracellular space as exosomes.
Exosome biogenesis can occur through an Endosomal Sorting Complex Required for Transport (ESCRT)-dependent mechanism or by an ESCRT-independent pathway. The first relies on multiple proteins that assemble into four ESCRT complexes (ESCRT-0, I, II, and III) and accessory proteins, such as an AAA-ATPase, VPS4 complex, and ALIX that are involved in MVB budding and loading (Babst et al., 2002; Henne et al., 2011; Colombo et al., 2014).
The exosome biogenesis also occurs through an ESCRT-independent pathway mediated by tetraspanins and ceramide-enriched lipid rafts (Trajkovic et al., 2008; Van Niel et al., 2011). Tetraspanins are recruited at early steps to endosome membranes, before ILV formation (Escola et al., 1998; Pols and Klumperman, 2009) and at least CD63, CD9, CD81, and CD82 are found in endosome and exosome membranes (Andreu and Yanez-Mo, 2014).
Ceramide and its derived metabolites are organized in raft-based microdomains that interact with proteins, such as flotillins. These lipid-enriched structures are involved not only in endosomal membrane invagination for ILV formation (Trajkovic et al., 2008) but also in cargo loading. The selective cargo loading occurs during exosome biogenesis through tetraspanin-dependent and/or ESCRT-dependent mechanisms. The composition and quantity of exosome released are dictated by specific molecules present in the membrane and lumen of ILVs and in the membrane of MVB, which is determinant for their selective trafficking toward plasma membrane for exosome release (Figure 2A).
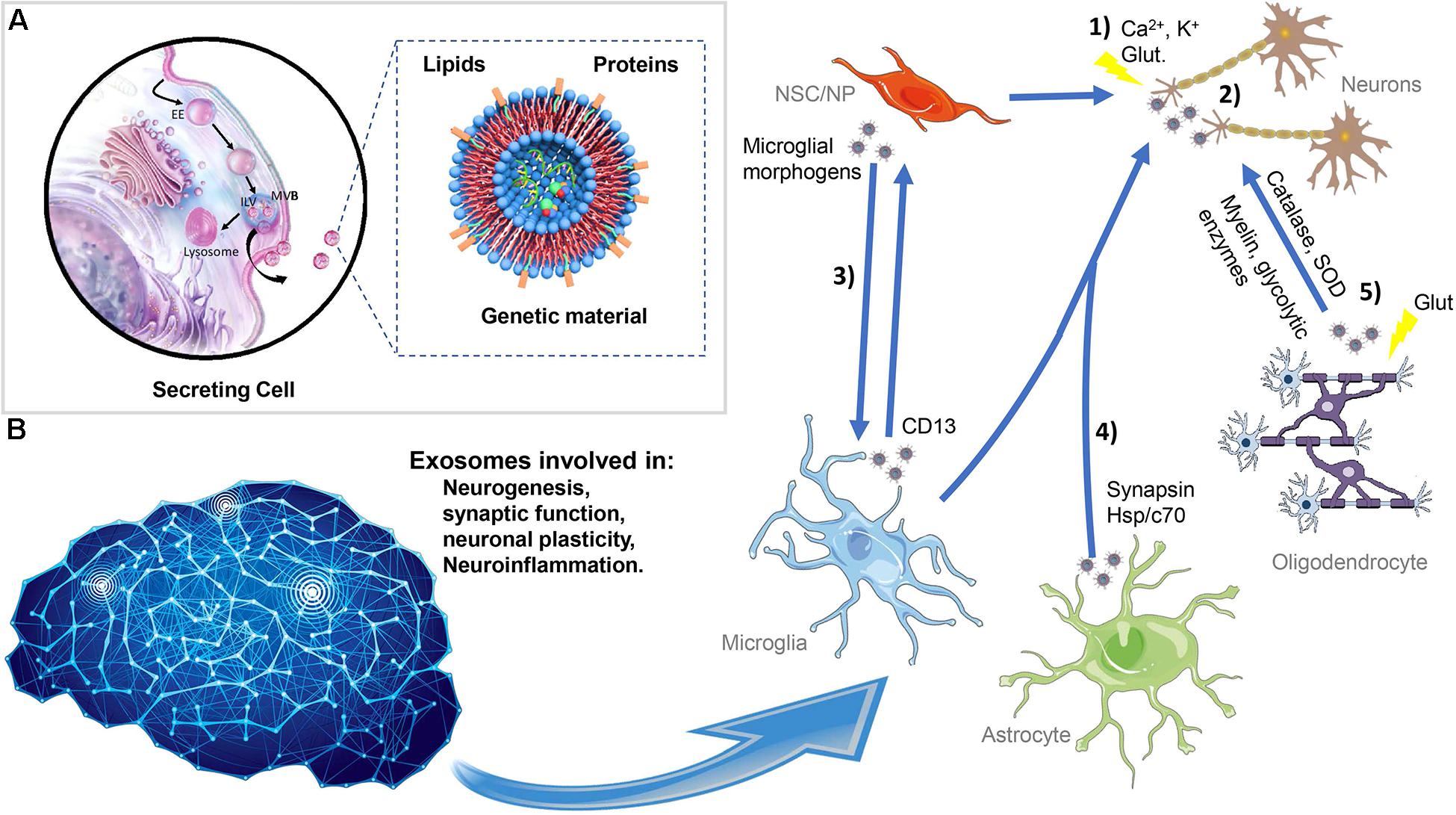
Figure 2. Exosome biogenesis, secretion, and interaction in the central nervous system. (A) Exosomes originated in the endocytic pathway as ILVs resulting in the formation of LE with multiple small vesicles termed MVBs. During this process, exosomes are loaded with proteins, lipids, and genetic material. Afterward, the MVBs can be destined for lysosomal degradation or fused with the plasma membrane and the exosomes released into the extracellular space. The exosomes have a complex composition of proteins such as ALIX, TSG101 and flotillin, Rab family, tetraspanins (e.g., CD63, CD81, and CD9), mitochondrial proteins, and others; lipids including cholesterol, sphingomyelin, glycosphingolipids, and ceramide; and nucleic acids especially RNA and also nuclear and mitochondrial DNA. (B) Exosomes can be released from NSC/NP, neurons, astrocytes, oligodendrocytes, and microglia acting in proximal or distal cells. Exosomes can be shuttled between the different neural cells modulating distinct neuronal processes such as neurogenesis, synaptic function, neuronal plasticity, and neuroinflammation. (1) In neurons, exosomes are released in response to stimulus, such as calcium influx, glutamatergic synaptic activity, and potassium-induced depolarization. (2) The exosome signaling pathways include neuron-to-neuron communication. (3) NSC can influence the microglia function, and exosomes released from glial cells exert effects on NSC. (4) Additionally, exosomes originated in astrocytes can be fundamental for the trophic support of neurons. (5) Oligodendrocytes can also shuttle to neuron exosomes containing myelin proteins and glycolytic enzymes. ILVs, intraluminal vesicles; EEs, early endosomes; MVBs, multivesicular bodies, NSC/NP, neural stem/progenitor cells.
Proteins
Proteins that are commonly found in exosomes are the ones involved in their biogenesis and are located in the endosomal membrane. The proteins detected in exosome fractions comprise components of the ESCRT machinery such as ALIX, TSG101, and flotillins (Kowal et al., 2016), proteins responsible for transport and fusion of MVB with the plasma membrane (RAB27A, RAB11B) and tetraspanins (e.g., CD63, CD81, and CD9) (Ostrowski et al., 2010; Kowal et al., 2016). Proteins associated with lipid rafts are found in exosomes, as they originate primarily from early endosomes formed by invagination of the plasma membrane (de Gassart et al., 2003). Membrane-associated and signaling proteins were found in exosomes released from stem cells after stimulation of their trafficking to lipid rafts, thus supporting a raft-mediated protein sorting mechanism (Lin et al., 2017).
Ubiquitinated proteins were detected in exosomes (Buschow et al., 2005) and might be sorted into ILVs by an ESCRT-dependent mechanism (MacDonald et al., 2012). ESCRT-0 can recognize and bind ubiquitin-like molecules, such as small ubiquitin-like modifiers (SUMO), thus recruiting SUMOylated proteins like RNA-binding proteins (RBPs) (Villarroya-Beltri et al., 2013). NEDD4, an E3 ubiquitin-protein ligase that recognizes PPXY motifs in cargo proteins, might also be involved in exosome loading with ubiquitinated proteins (Putz et al., 2008). Recently, it has been described that about 10% of exosome protein content are mitochondria-associated proteins (Choi et al., 2013). Despite the fact that initial studies have shown that mitochondrial proteins are absent in tumor-derived exosomes (Mears et al., 2004), further developments of the methodologies for exosome proteomic analysis revealed the presence of mitochondrial proteins in exosomes (Burke et al., 2014; Wang et al., 2020). A recent study showed that exosomes released from Huntington’s disease (HD) patient-derived iPSC also carried mitochondrial proteins (Lopes et al., 2020). A preliminary report also identified the presence of functional mitochondria in neural stem cell-derived exosomes (Peruzzotti-Jametti et al., 2020). The generation of mitochondrial-derived vesicles (MDVs) containing malfunctioning parts (proteins or peptides) seems to constitute a novel mitochondrial quality control mechanism. The MDVs can enter the secretory pathway by fusing with LE/MVB for further degradation into lysosomes (Soubannier et al., 2012), but, alternatively, they can be sorted to the plasma membrane, thus releasing the mitochondria content through exosomes.
Most of the proteins used as exosome markers are found in exosomes from different sources and are not exclusive to neuronal cells. Despite the enrichment of exosomes with some of these proteins, they can also be found in other types of EVs, as they are membrane-associated proteins (Lötvall et al., 2014). This raises concerns on the definition of exosome-specific cargo and on the characterization of cell-specific exosome fractions. Still, some cell-specific proteins can be used to distinguish the source of exosomes from different neuronal cells. For instance, neural secreted exosomes present cell adhesion molecule L1 (L1CAM) and glutamate receptors (Faure et al., 2006; Lachenal et al., 2011), oligodendrocyte exosomes contain specific myelin proteins (Kramer-Albers et al., 2007), and microglial exosomes contain CD13 and monocarboxylate transporter 1, which are cell specific (Potolicchio et al., 2005). Therefore, it is expected that exosomes might have a specific signature that correlates with their cellular origin and further research will bring light on their physiological roles.
Lipids
Lipids in exosomes share similarities with plasma membrane composition, particularly lipids that are normally found in lipid rafts such as plasma membrane composition cholesterol, sphingomyelin, glycosphingolipids, and ceramide (de Gassart et al., 2003; Llorente et al., 2013; Skotland et al., 2017). The raft-like regions are important platforms for protein stabilization in the outer left of the ILV: cholesterol is important for the assembly of ESCRT protein complexes (Boura et al., 2012) while lysophosphatidic acid (LBPA), a glycerophospholipid mainly found in MVB, stabilizes ALIX (Kufareva et al., 2014). Modulating the lipid content of exosomes affects their release. Treatment of oligodendroglial cells with an inhibitor of cholesterol egress from lysosomes (U18666A) or with a soluble cholesterol analog compromises exosome release (Strauss et al., 2010). On the other hand, cholesterol depletion induced by methyl-β-cyclodextrin treatment led to an increase of exosome release (Llorente et al., 2007). Exosomes are uniquely enriched in phosphatidylserine in the outer face of the membrane, which may help in the internalization process by the recipient cells (Morelli et al., 2004). Also, ceramide and glycosphingolipids are found in exosomes as they are involved in their biogenesis and release. Inhibition of neutral sphingomyelinase 2 (nSMase2), an enzyme that metabolizes sphingomyelin into ceramide, altered the loading of CD63, flotillin, and proteolipid proteins into exosomes and also impaired exosome secretion (Trajkovic et al., 2008; Yuyama et al., 2012; Menck et al., 2017; Miranda et al., 2018). Inhibition of ceramide conversion into sphingomyelin by siRNA or chemical inhibition of sphingomyelin synthase 2 (SMS2) promotes exosome secretion (Yuyama et al., 2012; Dinkins et al., 2014).
Genetic Material
Exosomes are natural carriers of genetic material, with almost any type of RNA being found inside, and to some extent also nuclear and mitochondrial DNA (mtDNA) (Valadi et al., 2007; Waldenstrom et al., 2012; Ridder et al., 2014). Exosomes are particularly enriched in small RNAs around 200 nucleotides in length, and only a few have more than 3 Kb, which supports the notion that most of the mRNA and long non-coding RNA detected are fragmented (Crescitelli et al., 2013). RNA loading into ILVs is mediated by RBPs that assist in the transport of RNAs to raft-like regions by a selective mechanism that involves specific RNA sequences and RNA hydrophobic modifications (Janas et al., 2004; Batagov et al., 2011).
One of the most important exosome cargoes are miRNAs, which are non-coding RNAs with 17–21 nucleotides, that regulate protein expression by binding to the untranslated regions (UTR) of mRNAs thus inhibiting their translation. Several studies have shown enrichment of selective miRNAs in exosomes, which suggests a selective mechanism of miRNA loading during the vesicle biogenesis (Guduric-Fuchs et al., 2012).
The packaging of miRNAs into MVBs is favored by the presence of specific exosome-sorting motifs in the 3′ portion of miRNA that is recognized by RBPs such as the chaperone hnRNPA2B1 (Heterogeneous Nuclear Ribonucleoprotein A2/B1), by a ceramide-dependent mechanism (Villarroya-Beltri et al., 2013). Also, 3′end uridylation of miRNA is involved with its sorting into EVs, demonstrating that posttranslational modifications regulate selective sorting of miRNA (Wyman et al., 2011; Koppers-Lalic et al., 2014). Mature miRNAs can interact with proteins forming the miRNA-induced silencing complex (miRISC). Argonaute 2 (Ago2) is a component of miRISC that localizes in MVBs, thus promoting miRNA loading (Gibbings et al., 2009; Guduric-Fuchs et al., 2012; Cha et al., 2015). Some studies have shown that Ago2 is present in exosomes (Goldie et al., 2014) while others reported its localization only at MVBs inside cells (Gibbings et al., 2009). More recent studies showed that the impairment of Ago2 localization in MVB affected its secretion in exosomes (McKenzie et al., 2016). In agreement, knockout of Ago2 impaired the export of selective miRNAs in HEK293T-derived exosomes (Guduric-Fuchs et al., 2012).
The miRNAs are also players involved in the loading mechanism of mRNA into exosomes. Specific sequences in 3′UTR of mRNA, with approximately 25 nucleotides and a CTGCC motif within a stem-loop structure, act as a sorting sequence to exosomes through a mechanism mediated by miR-1289 (Bolukbasi et al., 2012).
Exosomes also contain mtDNA (Guescini et al., 2010; Sansone et al., 2017) and single- and double-stranded DNA (Balaj et al., 2011; Yang et al., 2017). The secretion of DNA by exosomes may constitute an alternative process to maintain intracellular homeostasis by eliminating harmful cytoplasmic DNA (Takahashi et al., 2017). Moreover, part of the exosome DNA was bound to histones, which have been profiled in distinct exosome proteomic analysis (Simpson et al., 2008). So, the authors speculate that DNA loading into exosomes might depend on histones (Takahashi et al., 2017).
In a recent report, the authors employed a novel approach for exosome isolation with a combined methodology of high-resolution density gradient fractionation followed by a direct immunoaffinity capture to selectively isolate exosomes (Jeppesen et al., 2019). The study showed contradicting results, such as the presence of miRNA in non-vesicular fractions rather than in exosome fractions. This was supported by the absence of Ago2 or any miRNA machinery and RBPs in exosomes, in contrast with previous studies (Melo et al., 2014). Also, this study showed that isolated exosomes lacked dsDNA. Instead, this molecule is co-purified with these vesicles when standard isolation protocols are used. The authors proposed an EV-independent mechanism for dsDNA extracellular release, contradicting previous findings (Thakur et al., 2014). Most of the initial studies reporting dsDNA within exosomes were performed in cell conditional media, which might lead to premature conclusions on exosomes circulating on body fluids without validated protocols and standardized nomenclature (Ludwig et al., 2019). The Jeppesen study among others has pointed out that the isolation and purification methods have an important outcome in the final exosome content characterization and it is urgent to establish unified criteria for vesicle standard definition (size and density) and characterization of their cargo (Théry et al., 2018; Jeppesen et al., 2019; Witwer and Thery, 2019). Researchers have to be mindful of setting a subset characterization of exosome fractions before taking general conclusions from distinct studies. The vast majority of the current knowledge on the exosome field was obtained from cell line studies using conditional media. This remains the best approach for examining exosome content derived from a single or well-defined cell-type population and their function on other cells (Abramowicz et al., 2018).
The Transport of MVB to the Plasma Membrane and Exosome Release
The interaction of MVBs with actin and microtubules is essential for their transport to the plasma membrane (Villarroya-Beltri et al., 2014). The translocation of MVB toward the plasma membrane depends on several molecules via the cytoskeleton (Mittelbrunn et al., 2015). Kifc2 is a neuronal protein that moves directionally toward the plus end of microtubules and was found to be associated with MVB, suggesting a possible role in MVB transport to the neuronal plasma membrane (Saito et al., 1997). Rab GTPases such as RAB11, RAB27A/B, and RAB35 are mediators of selective sorting of MVB to the plasma membrane and exosome release (Hsu et al., 2010; Ostrowski et al., 2010; Escudero et al., 2014). RAB11 and p75 neurotrophin receptors are involved in directing MBV to the plasma membrane and exosome release in neuronal cells (Escudero et al., 2014). Also, Rab35 is involved in the mechanism of exosome release by oligodendrocytes (Fruhbeis et al., 2013). The MVBs are decorated with tethering protein complexes, such as HOPS and SNAREs, that mediate the fusion of these vesicles with the plasma membrane (Itakura et al., 2012; Jiang et al., 2014). The presence of tetraspanins (e.g., CD63) and lysosomal-associated membrane proteins LAMP1 and LAMP2 in late endosomes also facilitate the fusion of MVB with the plasma membrane (Jaiswal et al., 2002).
Uptake of Exosomes by Recipient Cells
After secretion, the exosomes will dock into the membrane of the target cells and activate signaling events or will be internalized through specific receptor–ligand interactions (Mulcahy et al., 2014). The transmembrane proteins present in the surface of exosomes (tetraspanins) can be recognized by signaling receptors in the target cells (Munich et al., 2012), resulting in activation of transducing pathways and modulation of intracellular processes without entering the target cell. Exosomes can merge with the plasma membrane of target cells releasing its cargo directly into the cytosol by a low pH-dependent mechanism (Parolini et al., 2009). However, the main route for exosome uptake seems to be mediated by endocytic events. Exosome uptake can occur by clathrin-mediated (Escrevente et al., 2011) or caveolin-dependent endocytosis (Nanbo et al., 2013), and the presence of lipid rafts in the membrane facilitates the process (Izquierdo-Useros et al., 2009; Svensson et al., 2013). After internalization, exosomes are sorted into LE/MVB with two possible fates: to be released again to neighboring cells or to be degraded after fusion of LE/MVB with lysosomes (Tian et al., 2013).
The uptake of exosomes by brain cells seems to be cell-type dependent. For instance, neurons and glial cells seem to uptake exosomes by clathrin-mediated endocytosis (Alvarez-Erviti et al., 2011b; Fruhbeis et al., 2013). Some neurons can also use specific receptors from the SNARE family, such as SNAP25, for exosome uptake (Zhang et al., 2017a). Curiously, the uptake of exosomes seems to be a selective pathway; exosomes derived from cortical neurons were primarily internalized by hippocampal neurons whereas exosomes released by neuroblastoma cell line N2A were taken up by astrocytes and oligodendrocytes (Chivet et al., 2014). Also, exosomes derived from oligodendrocytes were mainly internalized by microglia but not by neurons or astrocytes (Fitzner et al., 2011). Thus, the mechanisms that regulate the selective targeting of exosomes to recipient cells can determine their biological effect. Moreover, the uptake of exosomes was also more active in pre-synaptic regions, which might indicate that these vesicles use constitutive endocytosis processes at these regions for neuronal cell entrance (Granseth et al., 2006; Chivet et al., 2014). Nevertheless, the exact mechanism involved in exosome entrance into neuronal cells is still uncharacterized.
Exosomes and the CNS
If originally exosomes were considered to be disposable vehicles for the elimination of cellular components, now multiple studies demonstrated that exosomes play multiple physiological roles in the nervous system. Exosomes can be released from all brain cells, including neural stem/progenitor cells (Ma et al., 2019), neurons (Faure et al., 2006), astrocytes (Taylor et al., 2007; Fitzner et al., 2011), and microglia (Xia et al., 2019) acting in proximal or distal cells. Importantly, exosomes can be shuttled between the different neural cells modulating distinct neuronal processes such as neurogenesis (Luarte et al., 2017), synaptic function (Lachenal et al., 2011), neuronal plasticity (Lafourcade et al., 2016), and neuroinflammation (Dalvi et al., 2017; Figure 2B).
Neural stem/neural progenitor cells (NSCs/NPCs), found in neurogenic niches, can secrete or uptake exosomes, thus playing key roles in neuronal development (Ma et al., 2019). Exosomes derived from neural stem cells were able to promote neuronal growth and differentiation as their protein cargo affects neuronal development and function (Sharma et al., 2019). The exosomes derived from NSCs carry specific miRNA that targets neural regeneration, neuroprotection, and neural plasticity (Stevanato et al., 2016). Moreover, miRNA present in exosomes released from NSCs at the hypothalamic subventricular zone might play a beneficial role in the control of aging (Zhang et al., 2017b).
Exosomes are also important mediators of neuron-to-neuron communication. The secreted exosomes enter the recipient cells and can be re-secreted along with the recipient neuron endogenously formed exosomes, which facilitates specialized long-distance communication within a cell type (Polanco et al., 2018). Neuronal exosomes are mostly released from a somato-dendritic compartment in response to calcium influx and glutamatergic synaptic activity (Lachenal et al., 2011; Men et al., 2019). Moreover, potassium-induced depolarization stimulates the release of exosomes at synaptic terminals containing subunits of glutamate receptors (AMPA), GPI-anchored prion proteins, and L1 cell adhesion molecule (L1CAM) (Faure et al., 2006). The presence of proteins involved in neuronal function, such as amyloid precursor protein (APP) (Vingtdeux et al., 2007) or adaptor protein Ndfip1 (Putz et al., 2008) in exosomes further supported their modulatory role in synaptic function. Neuronal-derived exosomes (NDEs) containing microtubule-associated protein 1B (MAP1B) and a specific subset of miRNA related to synaptic plasticity were also described (Goldie et al., 2014). The release of exosomes by neurons is an important event in neuron-to-neuron communication. The Synaptotagmin 4 (Syt4)-containing exosomes were implicated in retrograde postsynaptic signaling by mediating activity-dependent presynaptic growth and neurotransmitter release (Korkut et al., 2013). Neuronal exosomes also carry Wnt proteins thus modulating the Wnt signaling pathway in the recipient cells (Korkut et al., 2009). Exosomes secreted by neurons contain a subset of miRNA that is distinct from the donor cells (Men et al., 2019), which support a selective packaging mechanism.
Trophic support of neurons is assisted by exosomes released from oligodendrocytes and astrocytes (Fruhbeis et al., 2012). Glutamate release by neurons triggers the secretion of exosomes enriched in specific proteins, and RNA from oligodendrocytes was shown to improve the cellular viability of neuronal cells under stress conditions (Fruhbeis et al., 2013). Moreover, the presence of catalase and superoxide dismutase in oligodendroglial exosomes might help neuronal survival under stress conditions of oxygen/glucose deprivation (Frohlich et al., 2014). Exosomes can mediate communication between oligodendrocytes and axons by supplying neurons with myelin proteins and glycolytic enzymes and substrates (Kramer-Albers et al., 2007; Kramer-Albers and White, 2011). During CNS tissue regeneration, oligodendrocytes communicate with microglia via exosomes to induce microglia-mediated elimination of oligodendroglial membranes without activation of the immune response (Fitzner et al., 2011).
Exosome-mediated signaling by astrocytes seems to be involved in neurite growth and survival (Wang et al., 2011; Xin et al., 2012). Synapsin I, a vesicle protein involved in neural development, was detected in exosomes released from astrocytes and, when added to hippocampal and cortical neurons under stress conditions, exerts a protective effect via promotion of neurite outgrowth and neuronal survival, respectively (Wang et al., 2011). The transport of neuroglobin, a hypoxic neuroprotective protein, from astrocytes to neurons was also reported by an exosome-dependent mechanism (Venturini et al., 2019). The exosome-mediated transfer of prion protein, an important receptor protein that protects against oxidative stress, from astrocytes to neurons improves their protection to oxidative and ischemic stress (Guitart et al., 2016). Also, the exosome-dependent release of Hsp/c70 from astrocytes might be fundamental to locally support neighboring neurons (Taylor et al., 2007). The neuron-to-astrocyte communication is also involved in the modulation of synaptic activation. For instance, exosomes from cortical neurons modulate the activation of GLT1 protein expression in astrocytes through miR-124a found in its cargo (Morel et al., 2013).
Microglia, which constitutes the first line of defense during brain injury or infection, releases exosomes enriched in aminopeptidase CD13 involved in neuropeptide catabolism (Potolicchio et al., 2005). Exosomes released from microglia can modulate neuronal activity through the activation of sphingolipid metabolism (Antonucci et al., 2012). Serotonin stimulates the release of exosomes from microglia (Glebov et al., 2015). Moreover, neuron-to-microglia communication via exosomes enhanced the removal of degenerative neurites, thus promoting proper neuronal function and synaptic plasticity (Bahrini et al., 2015). Exosomes released from neural stem cells target microglia and act as microglial morphogens by activating transcriptional signaling pathways primarily associated with the immune system in the post-natal nervous system (Morton et al., 2018). Exosomes released from NSC seem to control the state, organization, and morphology of microglia, which in turn forms a negative feedback loop to NSC, by interfering in cell proliferation.
Exosomes are also released by the peripheral nervous system, namely, Schwann cells, which were demonstrated to further support local axonal maintenance and regeneration (Lopez-Verrilli et al., 2013). The presence of miRNA from Schwann cells was also observed in axon terminals, in a process mediated through exosome, that has an impact on gene expression and neurite growth (Chang et al., 2013b; Ching et al., 2018).
Critical Roles of Exosomes in Neurological Disorders
EVs have been a recent object of research interest, resulting until now in several studies that search for connections between their physiological and/or pathological role and the development of neurodegenerative diseases. Several neurodegenerative disorders share other mechanistic features besides the accumulation of insoluble proteins. Exosomes are known to participate in these processes. Finding how the structure, biogenesis, cargo, communication, and final target of exosomes are altered in disease will give insights into their role in disease modulation and importantly enlighten their potential as a source of disease biomarkers. In this section, we describe the latest advances in exosome research in neurodegenerative disorders, as summarized in Table 1. Given the need for biomarkers of a large array of neurological diseases, understanding the mechanism underlying exosome regulation is of great interest for future research.
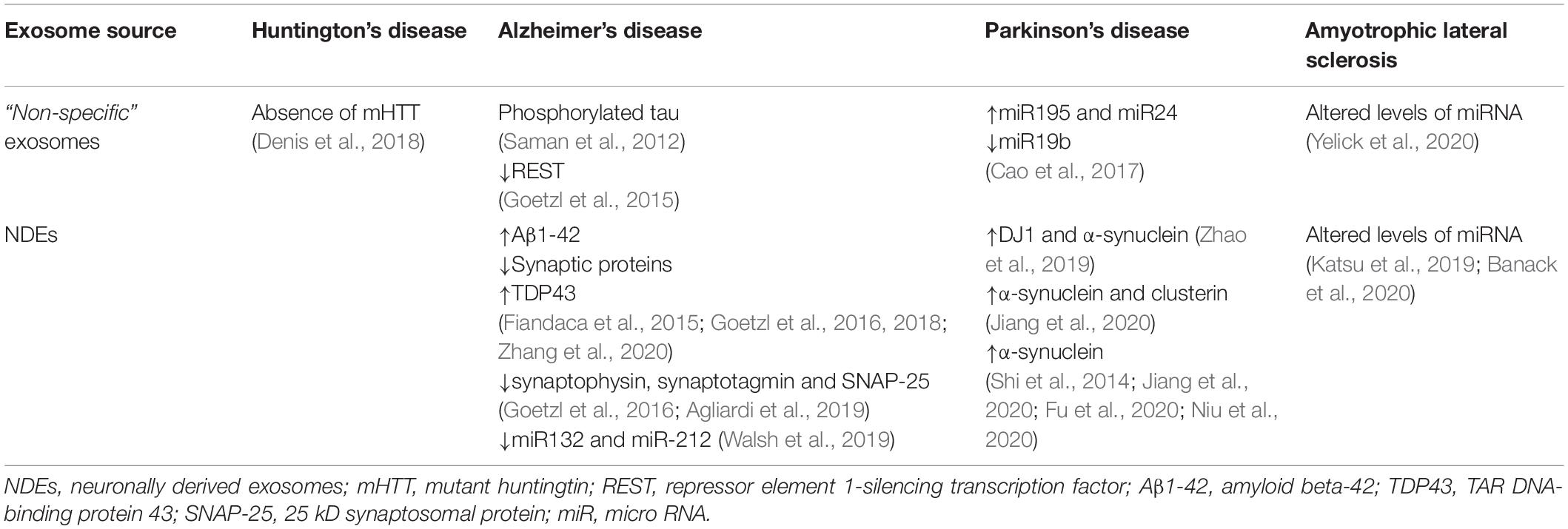
Table 1. Candidate biomarkers for neurodegenerative disorders transported in exosomes from different human biofluids.
Huntington’s Disease
The pathology of HD is caused by a CAG expansion in the HTT gene (Huntington’s Disease Collaborative Research Group, 1993), which encodes a protein with a polyglutamine expansion at the N-terminal, mutant huntingtin (mHTT). Accumulation of the mutated protein results in cognitive deficits and involuntary movements, correlated with a selective loss of striatal medium spiny neurons and cortical atrophy (Roos, 2010). The implication of the possible transfer of pathological biomolecules through a non-cell-autonomous pathway in HD has aroused attention after Cicchetti and colleagues showed that healthy fetal neural tissue engrafted in HD patients displayed mHTT aggregates several years after transplantation (Cicchetti et al., 2014). The unconventional mHTT spread throughout exosome transport was then suggested as a novel mechanism involved in HD pathology and opened new possibilities to find therapeutic targets aiming to mitigate this neurodegenerative condition.
Several studies suggested that exosomes can transport the expanded polyglutamine tract of HTT RNA and protein, as well as mHTT aggregates (Jeon et al., 2016; Zhang et al., 2016). Zhang and colleagues showed that after infecting human HEK293T cells with a lentivirus encoding mutant exon 1 huntingtin fragments, these cells secreted exosomes containing transcripts of the mutant polyglutamine tail and HTT peptides (Zhang et al., 2016). The addition of these exosomes to mouse striatal cells carrying both normal and mutant CAG repeats (Q7/7, Q111/111, respectively) led to the uptake of RNA but curiously not the protein (Zhang et al., 2016). However, others have acknowledged the uptake of exosomes containing mHTT by SH-SY5Y cells when exposed to conditioned media from HEK293 cells overexpressing GFP-mHtt-Q19 and -Q103 (Jeon et al., 2016). This is a topic with variable outcomes since the presence of mHTT is not always observed in exosomes, as described for astrocytes of an HD knock-in mouse model (HD140Q KI) (Hong et al., 2017). Huntingtin is a large protein with 350 kDa, which makes it difficult to be fully packed into exosomes. mHTT spreading through the exosome pathway is a complex and not yet defined mechanism that surely has an associated variability between HD models. However, strong evidence supports the idea that mHTT can be carried by exosomes reinforcing the need to better clarify how the protein is loaded into exosomes. The acknowledgment that mHTT is present in exosomes was shown when murine embryonic fibroblasts (MEFs) overexpressing the exon 1 of the Htt gene showed constitutive interaction of mHTT with exosome structural proteins Alix and TSG101 mediated by Transglutaminase 2 (Diaz-Hidalgo et al., 2016). In HD astrocytes, mHTT was shown to impair the exosome secretion by decreasing αB-crystallin levels, a glial protein involved in exosome secretion. The overexpression of αB-crystallin could revert the decrease in exosome release and simultaneously decreased the mHTT aggregates in the striatum of HD140Q KI mice (Hong et al., 2017). On the other hand, in the same study, the injection of exosomes from WT astrocytes was described to prevent aggregation of mHTT (Hong et al., 2017). The exosome transport of mHTT was confirmed when neurons differentiated from WT mice NSC displayed mHTT aggregates upon co-culture with HD fibroblasts (143 CAG repeats); moreover, mice injected with HD-derived exosomes also revealed that mHTT aggregates specifically in the DARPP-32 + medium spiny neurons of the host’s striatum. Eight weeks post-incubation, the animals exhibited motor impairment and cognitive deficits (Jeon et al., 2016). In contrast, exosomes isolated from platelets of HD patients failed to show any presence of mHTT (Table 1; Denis et al., 2018). Altogether, this data awareness on the importance of exosomes in the progression and pathophysiology of HD supports the need for deeper research in exosome-mediated mechanisms.
Alzheimer’s Disease
Alzheimer’s disease (AD), the most common form of dementia, has, as a causative mechanism, the accumulation of aggregated β-amyloid (Aβ) in the brain and phosphorylated tau in neurofibrillary tangles (Palmqvist et al., 2017). The accumulation of Aβ fibrils can precede symptoms for decades, which has long intrigued researchers looking for the mechanisms by which neurodegeneration occurs (Sperling et al., 2014). The discovery of multivesicular bodies (MVBs) as a site for the accumulation of Aβ42 in pre- and postsynaptic compartments preceded the later acknowledgment that the cleavage of β-amyloid peptides occurs in early endosomes before routing to MVB and the resulting products released from cells through exosomes (Takahashi et al., 2002; Rajendran et al., 2006). The presence of Alix associated with amyloid plaques in post-mortem AD patient’s brains, a fact not observed in healthy brains, reinforced the existence of a role for Aβ-associated exosomes in amyloid plaque formation (Rajendran et al., 2006). The presence of secretases (γ-, α-, and β-secretases), enzymes involved in the proteolytic cleavage of APP, in isolated exosomes from an AD animal model raised the possibility for these vesicles to be a potential site of APP cleavage (Sharples et al., 2008). Indeed, it was observed that APP goes through cleavage inside exosomes, with existing evidence that products resulting from APP processing are present in exosomes from AD human and mouse AD model brains, evidencing the exosome intervention in the pathological process (Vingtdeux et al., 2007; Sharples et al., 2008; Perez-Gonzalez et al., 2012; Guix et al., 2017; Laulagnier et al., 2018).
After evidence of existing Aβ in association with exosomes, the concern was to understand whether these vesicles contribute, as a vehicle, to spreading the pathology or can be protective by clearing the toxic elements associated with AD pathophysiology. Exosomes derived from neuroblastoma cells injected into the brain of the hAPP-J20 AD mouse model were able to trap Aβ bound to the glycosphingolipid surface and later transport to microglia for degradation. These findings supporting the exosome mentioned “clearance role” through the decrease in the levels of Aβ and Aβ-mediated synaptotoxicity (Yuyama et al., 2014). Altogether, these data reinforce the capacity of exosomes to carry Aβ and “deliver” their cargo to microglia, reducing the levels of synaptic toxicity and inflammatory response.
The levels of certain enzymes and structural components in exosomes promote Aβ elimination by microglial and lysosomal processes, including the insulin-degrading enzyme, ceramide, or the ganglioside GM1 (Tamboli et al., 2010; Wang et al., 2012; Yuyama et al., 2012).
Thereby, there is evidence that microglial phagocytic action on Aβ is promoted by exosomes being mediated by sphingolipids integrating their structure. Moreover, β-amyloid oligomers (Aβo) incubated with exosomes were sequestered to their superficial structure by interacting with surface proteins including the prion protein (PrPC) (An et al., 2013). The sequestration of Aβo by exogenous exosomes can exert a protective effect against the synaptic disruption induced by Aβ (An et al., 2013). Indeed, PrPC plays a dual role: a protective one, by capturing Aβ in exosomes, through superficial recognition, thus facilitating the formation of amyloidogenic fibrils and preventing the derivative neurotoxic complications mediated by Aβo and a neurotoxic effect when expressed in the neuronal surface as a receptor for Aβ (Falker et al., 2016). Other exosome components from the outer membrane were reported to be associated with Aβ, including glycans of glycosphingolipids and GM1 ganglioside, which was seen to induce exosome release and promote Aβ fibril formation (Yuyama et al., 2008, 2014). Hypoxia can accelerate the production of Aβ by increasing the activity of β and PS1/γ-secretase. In SH-SY5Y cells (expressing human WT APP695), maintained in hypoxic conditions, was observed an increase in Aβ40 and Aβ42 content in exosomes (Xie et al., 2018).
Although the abovementioned studies reveal an eliminatory role for exosomes to discard Aβ, C-terminal fragments, and full-length APP, recent data suggest that these vesicles can also transport Aβo, further contributing to the neurotoxicity seen in AD. The use of biological material/samples from AD patients can more accurately help to unravel what specifically happens in the development of the disease other than cellular or animal models. Recently, a study in post-mortem tissue from the temporal neocortex of AD patients showed that flotilin-1 and Aβo co-labeled and there were increased levels of oligomers in exosomes compared to controls (Sinha et al., 2018). Incubation of human-derived induced-pluripotent stem (hiPSC)-differentiated neurons and SH-SY5Y cells with labeled exosomes isolated from brain tissue of AD patients revealed the incorporation of vesicles carrying Aβo, leading to subsequent cytotoxicity when comparing to healthy brains (Sinha et al., 2018). The ablation of Aβo spreading was achieved by using siRNA to inhibit transcription of TSG101 and VPS4A, proteins involved in exosome formation and secretion (Sinha et al., 2018). In sum, these studies confirmed the transport of Aβo, APP, and its cleaved products in exosomes and their further uptake and internalization by neuronal cells. Moreover, the neurotoxic effects of adding exosomes derived from AD human plasma, cell, and animal models or previously exposed to toxicity-inducing Aβ-protofibrils has also been described (Eitan et al., 2016; Beretta et al., 2020).
Tau has also been found in exosomes isolated from CSF and brain of AD patients (Saman et al., 2012; Wang et al., 2017; Guix et al., 2018; Crotti et al., 2019; Ruan et al., 2021). In accordance, the injection into WT mice of exosomes from iPSC-derived neurons and post-mortem brain of AD patients led to the increase in tau phosphorylation and formation of inclusions (Aulston et al., 2019; Ruan et al., 2021).
Generally, the diagnosis of AD patients is based on the expression of tau and Aβ; however, the observation that exosomes from AD patients have differences in cargo has sparked curiosity in the field of biomarkers (Table 1). In accordance, exosomes isolated from plasma of AD patients exhibited decreased levels of transcription factors involved in neuronal protection from external damages like the repressor element 1-silencing transcription factor (REST), as already observed in individuals at pre-clinical stages without cognitive impairments (Goetzl et al., 2015).
The recent developments in the methods to isolate exosomes of neuronal origin from plasma have gained interest as they may include more sensitive and accurate neuronal biomarkers than total plasma exosomes (Table 1). Some reports have shown the presence of higher levels of Aβ1–42 and TAR DNA-binding protein 43 (TDP43) and decreased levels of synaptic proteins, as synaptophysin, synaptotagmin, and 25-kDa synaptosomal-associated protein (SNAP-25), in NDEs from AD patients (Fiandaca et al., 2015; Goetzl et al., 2016, 2018). Importantly, these first two presynaptic proteins, in particular, were already diminished at the pre-clinical stage and were correlated with the cognitive decline, as evaluated by the mini-mental state examination (Goetzl et al., 2016). In accordance, the exosome-associated growth-associated protein (GAP43) neurogranin, SNAP25, and synaptotagmin 1 proteins can be detected in AD up to several years before clinical onset (Jia et al., 2020). Following the same procedure, Zhao and colleagues showed that NDE Aβ1–42 levels allow discriminating between cognitive controls, mild cognitive impairment (MCI) patients, and AD dementia patients and predicting the risk of MCI progressing to AD dementia (Zhao et al., 2020). The levels of miR132 and miR-212 were also shown to be decreased in NDEs from AD patients (Walsh et al., 2019). Another comprehensive study involving EV isolated from post-mortem brain tissue from AD subjects has provided more insights into differential RNA biotype exosome content, namely, expression patterns of AD-associated miRNA compared with brain homogenates, adding to the pallet of possible targets for studying AD (Cheng et al., 2020). The NDEs’ novel approach, considered a liquid biopsy of the brain, resorts to exosomes released from neuronal cells found in the peripheral source, to discover more accurate biomarkers for neurodegenerative disorders. This technique has mainly been used in AD studies; however, strong proof of concept has also been assessed in other neurodegenerative diseases.
Parkinson’s Disease
Parkinson’s disease (PD) is the most common neurodegenerative movement disorder affecting 0.3% of the general population. The pathological hallmark is the loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc) of the striatum and the accumulation of α-synuclein in cytoplasmic inclusions called Lewy Bodies (Balestrino and Schapira, 2020). The formation of Lewy Bodies in healthy transplanted areas of PD patients years after grafting raised curiosity for the study of possible non-autonomous spreading mechanisms including related EVs (Kordower et al., 2008).
The first evidence of exosome involvement in PD was the presence of α-synuclein in Alix- and flotillin-positive vesicles derived from α-synuclein-expressing cells (Emmanouilidou et al., 2010). In 2012, the oligomeric form of α-synuclein was shown to be transported via exosomes, and additionally that exosomes may favor the formation of α-synuclein aggregates due to its lipid and/or protein composition thus facilitating the uptake of α-synuclein by cells (Danzer et al., 2012; Grey et al., 2015; Delenclos et al., 2017). Moreover, oligomerized α-synuclein seemed to be preferentially uptaken by cells when associated with exosomes rather than isolated, reinforcing the role of exosomes to transfer and deliver toxic oligomeric proteins to cells (Delenclos et al., 2017). Similar results were obtained when exosomes obtained from CSF of PD patients were added to H4 glioma cells, resulting in the elevated formation of α-synuclein oligomers (Stuendl et al., 2016). Surprisingly, this study reported reduced levels of α-synuclein in exosomes from CSF of early stage PD patients when compared to controls. On the other hand, others claim that the presence of α-synuclein seemed to be elevated in exosomes from plasma of PD patients, suggesting a positive correlation between exosome-associated α-synuclein levels and disease severity studies (Shi et al., 2014). In fact, the presence of α-synuclein in L1CAM-positive NDEs validated the protein as a biomarker for PD with origins in the nervous system (Shi et al., 2014). Henceforward, investigators have searched for mechanisms behind the release of exosomes aiming to manipulate it and find new outcomes to treat neurodegeneration in PD. α-synuclein was also shown to induce the release of exosomes by microglia cells in mice, thus promoting apoptosis, as well as lysosomal dysfunction in α-synuclein overexpressing SH-SY5Y cells, leading to an increase in the release of exosome-associated α-synuclein (Alvarez-Erviti et al., 2011a; Chang et al., 2013a). Additionally, when exosomes from serum of PD patients, highly enriched in α-synuclein, were injected in mice, it induced protein aggregation along with a neuroinflammatory response, reinforcing the neurotoxic role of exosome transfer in PD (Han et al., 2019). Alterations in genes’ expression like ATPase cation transporting 13A2 (ATP13A2) and β-glucocerebrosidase have also been associated with differences in the exosome-associated release of α-synuclein (Kong et al., 2014; Papadopoulos et al., 2018). While the overexpression of ATP13A2 (overexpressed in neurons of PD patients) in SH-SY5Y differentiated cells caused a decrease in intracellular α-synuclein with enhanced levels in exosomes, inhibition of β-glucocerebrosidase enzymatic activity in vivo caused the enhancement of α-synuclein release in association with exosomes and the levels of α-synuclein oligomers (Kong et al., 2014; Papadopoulos et al., 2018).
Altogether, these data suggest a transmission role for exosomes, as carriers of pathogenic/toxicity-inducing elements. The same applies to the loading of nucleic/proteic fragments that could rescue neurotoxic processes happening in PD contexts. Strikingly, a study showed that PD plasma-derived exosomes, when added to rat cortical neurons, were capable of diminishing apoptosis and increase metabolic activity when compared to the addition of exosomes from healthy individuals (Tomlinson et al., 2015). A possible explanation for the different outcomes of this study could be the incapacity to detect α-synuclein in PD-derived serum microvesicles. In line with these results, another study showed that a mouse model of PD equally benefited from injection with exosomes from blood of healthy controls, by experiencing diminished cell loss and movement alterations, demonstrating the therapeutic role of exosomes as potential rescuers in neurodegeneration (Sun et al., 2020).
Access to plasma samples from PD patients also allowed the isolation of plasma NDEs and the detection of more sensitive disease biomarkers, such as DJ1 and α-synuclein (Table 1; Zhao et al., 2019). The same increased levels of α-synuclein were, however, not confirmed in exosomes isolated from the urine of PD patients (Ho et al., 2014). More recently, in plasma L1CAM-positive vesicles, an increase of α-synuclein levels in combination with clusterin, a stress-induced chaperone, allowed the differentiation of patients with PD from those with atypical parkinsonism (Jiang et al., 2020). The α-synuclein levels in NDEs from plasma and serum of PD patients were shown to be augmented but with some divergences between studies on how this aspect correlates with disease progression (Fu et al., 2020). The number of NDEs was described to be higher in plasma of PD patients and correlated with either early or very advanced stages of the disease (Ohmichi et al., 2019). NDEs positive for L1CAM were also used as a tool to assess the concentration of proteins of interest in the serum of PD patients who participated in a clinical trial for the study of possible beneficial effects of a diabetes type 2 treatment drug (Athauda et al., 2019). The study of the mechanisms linking PD and exosome is of major interest to understand the neurotoxic events in PD contexts and brings hope for new therapeutic approaches focusing on nanovesicles as key interlocutors.
Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) affects both upper motor neurons and lower motor neurons leading to motor and extra-motor symptoms. The majority of cases is related to alterations in four genes, namely, C9orf72, TARDBP (encoding TAR DNA-binding protein 43, TDP43), SOD1 (encoding superoxide dismutase), and FUS (encoding RNA binding protein FUS), which results in aggregation and accumulation of ubiquitinated protein inclusions in motor neurons (Mejzini et al., 2019).
ALS was first associated with exosome trafficking in 2007 when a study showed the transport of both WT and mutant Cu/Zn superoxide dismutase (SOD1) by exosomes in a cell model for ALS (mouse neuron-like NSC-34 overexpressing human mutant SOD1) (Gomes et al., 2007). Further ahead, Grad and colleagues demonstrated that the same ALS cellular model was capable of incorporating misfolded SOD1, pointing that this protein was localized in the external part of exosomes isolated from the supernatant of cells transfected with human WT and mutant GFP-SOD1 (Grad et al., 2014). Astrocytes from primary cultures overexpressing mutant SOD1 released more exosomes than controls and more efficiently transferred these proteins, resulting in the loss of motor neurons as shown by the decrease in the SMI32/NeuN ratio in cultures where exosomes were added (Basso et al., 2013). Indeed, the release of mutant SOD1 by EV has been observed in both astrocytes and neurons from an ALS animal model (transgenic SOD1G93A mouse model) (Basso et al., 2013; Silverman et al., 2019). Moreover, mutant SOD1 was also present in EV from the brain and spinal cord of ALS patients, evidencing the spreading role of these vesicles (Silverman et al., 2019). In accordance, an EV-associated miRNA (miR-494-3p) released by astrocytes obtained through direct conversion of fibroblasts of ALS patients contributed to the loss of motor neurons in mice (Varcianna et al., 2019). Additionally, alteration of miR-124 levels associated with inflammatory processes was observed in exosomes from NSC-34 (motor neuron-like cultures) transfected with mutant SOD1 (Pinto et al., 2017). These exosomes were preferentially incorporated by microglial cells (co-culture of microglia and NSC) and lead to the decrease of microglia phagocytic ability.
The continuous search for the role of exosomes in ALS motivated concerns about the plausible spreading of other pathological agents in the disease than the mutant protein mainly implied, SOD1. Further analysis showed that TDP-43 protein was transported through exosomes isolated from the CSF of ALS patients (also suffering from frontotemporal dementia) to U251 human glioma cells (Ding et al., 2015). The uptake of these exosomes was accompanied by an increase in apoptosis- and autophagy-related proteins, which demonstrated that the transport of toxic cargo by nanovesicles can spread and promote negative effects throughout cells.
The identification of miRNA (e.g., miR-124-3p, hsa-miR-4736, and miR-146a-5p) with altered levels in both neuronal-derived and total exosomes from ALS patients (CSF and plasma) gave some insight into novel targets to overcome the pathology (Table 1; Banack et al., 2020). Nonetheless, the research regarding the role of exosome vesicles in ALS has still a long way to go to allow a better understanding of how we can use the already acquired knowledge to mitigate the progression of these neurodegenerative processes.
Future Perspectives: Diagnostic and Therapeutic Potentials of Exosomes
In the last decade, the research into the exosome’s biological characteristics has seen exponential growth. Exosomes are secreted from virtually all types of cells acting as mediators of intercellular communication physiologically. Interestingly, evidence has been reported that exosomes are involved in the development of human neural circuits by regulating signaling pathways. The maturation and function of neurogenic niches are highly regulated by miRNA (e.g., miR-let7b and miR-9 regulates NSC proliferation and differentiation; miR-34a promotes NSC differentiation and neuron maturation among others) and most of them were found in exosome cargo from different types of cells (Bátiz et al., 2015). Moreover, exosomes have shown the potential to restore brain cellular function in the NSC model of Rett syndrome incubated with isogenic control iPSC-derived exosomes, resulting in amelioration of the developmental deficit associated with the disease (Sharma et al., 2019). Exosomes from NSCs were also able to reduce the lesion extension in a spinal cord injury mouse model, improve functional recovery, and mitigate neuroinflammation by inducing autophagy (Rong et al., 2019). These data are feasible such that exosome content can target NSC/NPC to modulate the neurogenic process and influence neuronal plasticity. Importantly, exosomes from NSCs exert anti-inflammatory, neurogenic, and neurotrophic effects that are likely to be useful to treat conditions such as dementia, AD, HD, PD, ALS, stroke, traumatic brain injury, multiple sclerosis, and major depressive disorder.
In contrast, exosomes have been suggested to be important players in the propagation of neurodegenerative diseases by providing a mechanism for the spreading of toxic content such as misfolded, aggregated forms of proteins. In the last few years, a causal connection between exosomes and the development and progression of CNS diseases such as AD, PD, prion disease, multiple sclerosis, traumatic encephalopathy, and stroke, among others, has been explored (Liu et al., 2019). These diseases have in common the activation of the autophagic–lysosomal pathways for the elimination of misfolded proteins. Exosomes can act as an alternative pathway to the overload of these protective cellular routes by loading and secreting the deleterious proteins (ExoCarta), although the mechanism is not yet fully comprehended. Our understanding of the cellular and molecular mechanisms involved in the biogenesis, release, uptake, and function of EV remains elusive, and further studies are needed to help clarify these aspects. Whereas the importance of exosomes in cell communication is undisputed, the knowledge of how they can influence and regulate cell responses is critical to deciphering their potential physiological and pathological impacts. While the current studies offer contradictory data on the favorable or adverse effects exerted by exosomes, they are not certainly bystanders in any step of these biological processes.
A major pitfall in the exosome field is the technical difficulties in the isolation and characterization of “pure” exosomes. The exosome cargo is enriched in nucleic acids, proteins, lipids, amino acids, and metabolites that vary according to the cell of origin. The heterogeneity among preparations and the possible contamination of EVs/exosome fraction with body fluid contaminants might interfere with the characterization of vesicular cargo. Our knowledge about exosome research is challenged due to the lack of reliable and accurate isolation protocols, the lack of selective biomarkers, and the lack of high-resolution characterization techniques. Routine isolation methods include ultracentrifugation, density-gradient centrifugation, and ultrafiltration with all having their own advantages and disadvantages, and a universally accepted methodology for exosome isolation is still an intense debate. Identifying and separating different subpopulations of exosomes from specific cellular types will provide tremendous advantages in defining an exosome biosignature to act as a diagnostic biomarker. Isolation of neural-derived exosomes from human plasma is a particularly attractive method to improve the specificity of disease fingerprints. The immunoprecipitation of neuronal exosomes by antibodies targeting the marker proteins NCAM or L1CAM has proven to significantly increase the accuracy of exosome content as a disease biomarker. NCAM and L1CAM are not exclusive from neurons, despite their enrichment, impelling the search for more precise markers that will allow to obtain purer exosome populations from different brain cell types and eventually compare exosome biomarkers in the same sample. Importantly, there is an urge for more robust biomarkers obtained through a minimally invasive method for diagnostic purposes, preferably in the early stages of the disease, to monitor disease progression and to evaluate treatment response.
Exosomes are a promising tool with potential clinical application both as a diagnostic biomarker and as a therapeutic agent, particularly as drug/gene delivery systems. For a long time, mesenchymal stem cell-derived exosomes have been used in a myriad of pathological conditions with success (Xin et al., 2012; Ibrahim et al., 2014; Hao et al., 2017; Mendt et al., 2019). Undifferentiated iPSCs were also reported to release an elevated number of exosomes that exert multiple protective effects in different situations such as in cardiovascular diseases or by reducing senescence and stimulating proliferation of human dermal fibroblasts (Wang et al., 2015; Oh et al., 2018). Exosomes derived from human iPSC-neural stem cells led to significant improvements at the tissue and functional levels in different animal models of stroke, strongly supporting exosomes as a biological treatment (Webb et al., 2018a,b). These studies emphasize the important role of exosomes derived from neural cells in maintaining neural functions.
Exosomes have natural cargo transportation properties, turning them into an excellent vehicle for transferring bioactive molecules such as drugs, proteins, non-coding miRNA, and gene therapeutic agents to specific cells involved in CNS diseases. Exosomes combine a myriad of advantages as nanocarriers including stability, biocompatibility, low immunogenicity, ability to cross the blood–brain barrier, and efficient cellular uptake that can be modulated to target specific cells according to appropriate membrane proteins. The field of nanotherapy centered on nanoparticle-based exosome mimetics has evolved rapidly and can be a promising tool for drug delivery. Of major interest is the incorporation of miRNA into exosomes to be delivered into recipient cells for protective effects. However, despite the advances stressed, there are many challenging points to consider. One is related to the low yield of exosomes obtained from cells since large-scale production of exosomes for clinical use must be achieved. Furthermore, the use of exosomes for therapeutic purposes should clarify the dose–response output and the distribution and most importantly increase the specificity for the target cells. The lack of an efficient method for cargo loading into the exosomes is also a limitation. The current strategies include conjugation of harvested exosomes with drugs by different methods (e.g., electroporation, sonication, extrusion, and freeze/thaw), incubating cells with compounds to be incorporated into the exosomes, or transfection of donor cells with active molecules before the collection of the exosomes. So far, none of the strategies have been validated for clinical use. The short run is expected to have exosome-based therapeutics treating a wide range of maladies. Meanwhile, fully exploiting the exosomes’ potential is critical to define the safest and more efficient modes of delivery into the target tissue, either by systemic administration or by local delivery. Exosomes in circulation are rapidly sequestered by organ or cleared, remaining in the serum less than 5% at 5 min upon intravenous injection (Takahashi et al., 2013).
Importantly, the examples discussed above have demonstrated that exosomes comprise a great potential for the diagnosis and treatment of neurodegenerative disorders. Currently, the efforts in the field are focused on not only obtaining exosomes from specific brain cell types but also exploring the possibility of targeting exosomes to a specific neuronal type in the brain. These findings are expected to open up new avenues for pharmacological treatments with minor side effects. Overall, we have come a long way in exosome research, although several obstacles remain to be overcome before transitioning from innovation to widespread clinical use.
Author Contributions
MB, RV, and CL conceived, wrote, and participated in the revising of the manuscript. All authors have approved the final version of the manuscript.
Funding
This work was financed by the European Regional Development Fund (ERDF), through the Centro 2020 Regional Operational Programme under project CENTRO-01-0145-FEDER- 000012-HealthyAging2020, and through the COMPETE 2020–Operational Programme for Competitiveness and Internationalization and Portuguese national funds via FCT-Fundação para a Ciência e a Tecnologia, under project POCI-01-0145-FEDER-029621 and UIDB/04539/2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to acknowledge Dr. Felipe Serrano for his critical review of the manuscript.
References
Abramowicz, A., Marczak, L., Wojakowska, A., Zapotoczny, S., Whiteside, T. L., Widlak, P., et al. (2018). Harmonization of exosome isolation from culture supernatants for optimized proteomics analysis. PLoS One 13:e0205496. doi: 10.1371/journal.pone.0205496
Agliardi, C., Guerini, F. R., Zanzottera, M., Bianchi, A., Nemni, R., and Clerici, M. (2019). SNAP-25 in serum is carried by exosomes of neuronal origin and is a potential biomarker of Alzheimer’s disease. Mol. Neurobiol. 56, 5792–5798. doi: 10.1007/s12035-019-1501-x
Alvarez-Erviti, L., Seow, Y., Schapira, A. H., Gardiner, C., Sargent, I. L., Wood, M. J. A., et al. (2011a). Lysosomal dysfunction increases exosome-mediated alpha-synuclein release and transmission. Neurobiol. Dis. 42, 360–367. doi: 10.1016/j.nbd.2011.01.029
Alvarez-Erviti, L., Seow, Y., Yin, H., Betts, C., Lakhal, S., and Wood, M. J. (2011b). Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 29, 341–345. doi: 10.1038/nbt.1807
An, K., Klyubin, I., Kim, Y., Jung, J. H., Mably, A. J., O’Dowd, S. T., et al. (2013). Exosomes neutralize synaptic-plasticity-disrupting activity of Aβ assemblies in vivo. Mol. Brain 6:47. doi: 10.1186/1756-6606-6-47
Andreu, Z., and Yanez-Mo, M. (2014). Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 5:442. doi: 10.3389/fimmu.2014.00442
Antonucci, F., Turola, E., Riganti, L., Caleo, M., Gabrielli, M., Perrotta, C., et al. (2012). Microvesicles released from microglia stimulate synaptic activity via enhanced sphingolipid metabolism. EMBO J. 31, 1231–1240. doi: 10.1038/emboj.2011.489
Athauda, D., Gulyani, S., Kumar Karnati, H., Li, Y., Tweedie, D., Mustapic, M., et al. (2019). Utility of neuronal-derived exosomes to examine molecular mechanisms that affect motor function in patients with parkinson disease: a secondary analysis of the exenatide-PD trial. JAMA Neurol. 76, 420–429. doi: 10.1001/jamaneurol.2018.4304
Aulston, B., Liu, Q., Mante, M., Florio, J., Rissman, R. A., and Yuan, S. H. (2019). Extracellular vesicles isolated from familial alzheimer’s disease neuronal cultures induce aberrant tau phosphorylation in the wild-type mouse brain. J. Alzheimer’s Dis. 72, 575–585. doi: 10.3233/JAD-190656
Babst, M., Katzmann, D. J., Estepa-Sabal, E. J., Meerloo, T., and Emr, S. D. (2002). Escrt-III: an endosome-associated heterooligomeric protein complex required for mvb sorting. Dev. Cell 3, 271–282. doi: 10.1016/s1534-5807(02)00220-224
Bahrini, I., Song, J. H., Diez, D., and Hanayama, R. (2015). Neuronal exosomes facilitate synaptic pruning by up-regulating complement factors in microglia. Sci. Rep. 5:7989. doi: 10.1038/srep07989
Balaj, L., Lessard, R., Dai, L., Cho, Y. J., Pomeroy, S. L., Breakefield, X. O., et al. (2011). Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat. Commun. 2:180. doi: 10.1038/ncomms1180
Balestrino, R., and Schapira, A. (2020). Parkinson disease. Eur. J. Neurol. 27, 27–42. doi: 10.1111/ene.14108
Banack, S. A., Dunlop, R. A., and Cox, P. A. (2020). An miRNA fingerprint using neural-enriched extracellular vesicles from blood plasma: towards a biomarker for amyotrophic lateral sclerosis/motor neuron disease. Open Biol. 10:200116. doi: 10.1098/rsob.200116
Basso, M., Pozzi, S., Tortarolo, M., Fiordaliso, F., Bisighini, C., Pasetto, L., et al. (2013). Mutant copper-zinc superoxide dismutase (SOD1) induces protein secretion pathway alterations and exosome release in astrocytes: implications for disease spreading and motor neuron pathology in amyotrophic lateral sclerosis. J. Biol. Chem. 288, 15699–15711. doi: 10.1074/jbc.M112.425066
Batagov, A. O., Kuznetsov, V. A., and Kurochkin, I. V. (2011). Identification of nucleotide patterns enriched in secreted RNAs as putative cis-acting elements targeting them to exosome nano-vesicles. BMC Genom. 12, (Suppl. 3):S18. doi: 10.1186/1471-2164-12-S3-S18
Bátiz, L. F., Castro, M. A., Burgos, P. V., Velásquez, Z. D., Muñoz, R. I., Lafourcade, C. A., et al. (2015). Exosomes as novel regulators of adult neurogenic niches. Front. Cell Neurosci. 9:501. doi: 10.3389/fncel.2015.00501
Becker, A., Thakur, B. K., Weiss, J. M., Kim, H. S., Peinado, H., and Lyden, D. (2016). Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell 30, 836–848. doi: 10.1016/j.ccell.2016.10.009
Beretta, C., Nikitidou, E., Streubel-Gallasch, L., Ingelsson, M., Sehlin, D., and Erlandsson, A. (2020). Extracellular vesicles from amyloid-β exposed cell cultures induce severe dysfunction in cortical neurons. Sci. Rep. 10:19656. doi: 10.1038/s41598-020-72355-72352
Bolukbasi, M. F., Mizrak, A., Ozdener, G. B., Madlener, S., Strobel, T., Erkan, E. P., et al. (2012). miR-1289 and “Zipcode”-like sequence enrich mRNAs in microvesicles. Mol. Ther. Nucleic Acids 1:e10. doi: 10.1038/mtna.2011.2
Boura, E., Ivanov, V., Carlson, L. A., Mizuuchi, K., and Hurley, J. H. (2012). Endosomal sorting complex required for transport (ESCRT) complexes induce phase-separated microdomains in supported lipid bilayers. J. Biol. Chem. 287, 28144–28151. doi: 10.1074/jbc.M112.378646
Burke, M., Choksawangkarn, W., Edwards, N., Ostrand-Rosenberg, S., and Fenselau, C. (2014). Exosomes from myeloid-derived suppressor cells carry biologically active proteins. J. Proteome Res. 13, 836–843. doi: 10.1021/pr400879c
Buschow, S. I., Liefhebber, J. M., Wubbolts, R., and Stoorvogel, W. (2005). Exosomes contain ubiquitinated proteins. Blood Cells Mol. Dis. 35, 398–403. doi: 10.1016/j.bcmd.2005.08.005
Cao, X.-Y., Lu, J.-M., Zhao, Z.-Q., Li, M.-C., Lu, T., An, X.-S., et al. (2017). MicroRNA biomarkers of Parkinson’s disease in serum exosome-like microvesicles. Neurosci. Lett. 644, 94–99. doi: 10.1016/j.neulet.2017.02.045
Cha, D. J., Franklin, J. L., Dou, Y., Liu, Q., Higginbotham, J. N., Demory Beckler, M., et al. (2015). KRAS-dependent sorting of miRNA to exosomes. eLife 4:e07197. doi: 10.7554/eLife.07197
Chang, C., Lang, H., Geng, N., Wang, J., Li, N., and Wang, X. (2013a). Exosomes of BV-2 cells induced by alpha-synuclein: important mediator of neurodegeneration in PD. Neurosci. Lett. 548, 190–195. doi: 10.1016/j.neulet.2013.06.009
Chang, L. W., Viader, A., Varghese, N., Payton, J. E., Milbrandt, J., and Nagarajan, R. (2013b). An integrated approach to characterize transcription factor and microRNA regulatory networks involved in Schwann cell response to peripheral nerve injury. BMC Genom. 14:84. doi: 10.1186/1471-2164-14-84
Cheng, L., Vella, L. J., Barnham, K. J., McLean, C., Masters, C. L., and Hill, A. F. (2020). Small RNA fingerprinting of Alzheimer’s disease frontal cortex extracellular vesicles and their comparison with peripheral extracellular vesicles. J. Extracell Vesicles 9:1766822. doi: 10.1080/20013078.2020.1766822
Ching, R. C., Wiberg, M., and Kingham, P. J. (2018). Schwann cell-like differentiated adipose stem cells promote neurite outgrowth via secreted exosomes and RNA transfer. Stem Cell Res. Ther. 9:266. doi: 10.1186/s13287-018-1017-1018
Chivet, M., Javalet, C., Laulagnier, K., Blot, B., Hemming, F. J., and Sadoul, R. (2014). Exosomes secreted by cortical neurons upon glutamatergic synapse activation specifically interact with neurons. J. Extracell Vesicles 3:24722. doi: 10.3402/jev.v3.24722
Choi, D. S., Kim, D. K., Kim, Y. K., and Gho, Y. S. (2013). Proteomics, transcriptomics and lipidomics of exosomes and ectosomes. Proteomics 13, 1554–1571. doi: 10.1002/pmic.201200329
Cicchetti, F., Lacroix, S., Cisbani, G., Vallières, N., Saint-Pierre, M., St-Amour, I., et al. (2014). Mutant huntingtin is present in neuronal grafts in Huntington disease patients. Ann. Neurol. 76, 31–42. doi: 10.1002/ana.24174
Colombo, M., Raposo, G., and Thery, C. (2014). Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 30, 255–289. doi: 10.1146/annurev-cellbio-101512-122326
Crescitelli, R., Lässer, C., Szabó, T. G., Kittel, A., Eldh, M., Dianzani, I., et al. (2013). Distinct RNA profiles in subpopulations of extracellular vesicles: apoptotic bodies, microvesicles and exosomes. J. Extracell Vesicles 2, doi: 10.3402/jev.v2i0.20677
Crotti, A., Sait, H. R., McAvoy, K. M., Estrada, K., Ergun, A., Szak, S., et al. (2019). BIN1 favors the spreading of Tau via extracellular vesicles. Sci. Rep. 9:9477. doi: 10.1038/s41598-019-45676-45670
Dalvi, P., Sun, B., Tang, N., and Pulliam, L. (2017). Immune activated monocyte exosomes alter microRNAs in brain endothelial cells and initiate an inflammatory response through the TLR4/MyD88 pathway. Sci. Rep. 7:9954. doi: 10.1038/s41598-017-10449-10440
Danzer, K. M., Kranich, L. R., Ruf, W. P., Cagsal-Getkin, O., Winslow, A. R., Zhu, L., et al. (2012). Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol. Neurodegenerat. 7:42. doi: 10.1186/1750-1326-7-42
de Gassart, A., Geminard, C., Fevrier, B., Raposo, G., and Vidal, M. (2003). Lipid raft-associated protein sorting in exosomes. Blood 102, 4336–4344. doi: 10.1182/blood-2003-03-0871
Delenclos, M., Trendafilova, T., Mahesh, D., Baine, A. M., Moussaud, S., Yan, I. K., et al. (2017). Investigation of endocytic pathways for the internalization of exosome-associated oligomeric alpha-synuclein. Front. Neurosci. 11:172. doi: 10.3389/fnins.2017.00172
Denis, H. L., Lamontagne-Proulx, J., St-Amour, I., Mason, S. L., Weiss, A., Chouinard, S., et al. (2018). Platelet-derived extracellular vesicles in Huntington’s disease. J. Neurol. 265, 2704–2712. doi: 10.1007/s00415-018-9022-9025
Diaz-Hidalgo, L., Altuntas, S., Rossin, F., D’Eletto, M., Marsella, C., Farrace, M. G., et al. (2016). Transglutaminase type 2-dependent selective recruitment of proteins into exosomes under stressful cellular conditions. Biochim. et Biophys. Acta (BBA)-Molecular Cell Res. 1863, 2084–2092. doi: 10.1016/j.bbamcr.2016.05.005
Ding, X., Ma, M., Teng, J., Teng, R. K. F., Zhou, S., Yin, J., et al. (2015). Exposure to ALS-FTD-CSF generates TDP-43 aggregates in glioblastoma cells through exosomes and TNTs-like structure. Oncotarget 6:24178. doi: 10.18632/oncotarget.4680
Dinkins, M. B., Dasgupta, S., Wang, G., Zhu, G., and Bieberich, E. (2014). Exosome reduction in vivo is associated with lower amyloid plaque load in the 5XFAD mouse model of Alzheimer’s disease. Neurobiol. Aging 35, 1792–1800. doi: 10.1016/j.neurobiolaging.2014.02.012
Eitan, E., Hutchison, E. R., Marosi, K., Comotto, J., Mustapic, M., Nigam, S. M., et al. (2016). Extracellular vesicle-associated Aβ mediates trans-neuronal bioenergetic and Ca 2+-handling deficits in Alzheimer’s disease models. NPJ Aging Mechan. Dis. 2, 1–11. doi: 10.1038/npjamd.2016.19
Emmanouilidou, E., Melachroinou, K., Roumeliotis, T., Garbis, S. D., Ntzouni, M., Margaritis, L. H., et al. (2010). Cell-produced α-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J. Neurosci. 30, 6838–6851. doi: 10.1523/JNEUROSCI.5699-09.2010
Escola, J. M., Kleijmeer, M. J., Stoorvogel, W., Griffith, J. M., Yoshie, O., and Geuze, H. J. (1998). Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human B-lymphocytes. J. Biol. Chem. 273, 20121–20127. doi: 10.1074/jbc.273.32.20121
Escrevente, C., Keller, S., Altevogt, P., and Costa, J. (2011). Interaction and uptake of exosomes by ovarian cancer cells. BMC Cancer 11:108. doi: 10.1186/1471-2407-11-108
Escudero, C. A., Lazo, O. M., Galleguillos, C., Parraguez, J. I., Lopez-Verrilli, M. A., Cabeza, C., et al. (2014). The p75 neurotrophin receptor evades the endolysosomal route in neuronal cells, favouring multivesicular bodies specialised for exosomal release. J. Cell Sci. 127(Pt 9), 1966–1979. doi: 10.1242/jcs.141754
Falker, C., Hartmann, A., Guett, I., Dohler, F., Altmeppen, H., Betzel, C., et al. (2016). Exosomal cellular prion protein drives fibrillization of amyloid beta and counteracts amyloid beta-mediated neurotoxicity. J. Neurochem. 137, 88–100. doi: 10.1111/jnc.13514
Faure, J., Lachenal, G., Court, M., Hirrlinger, J., Chatellard-Causse, C., Blot, B., et al. (2006). Exosomes are released by cultured cortical neurones. Mol. Cell Neurosci. 31, 642–648. doi: 10.1016/j.mcn.2005.12.003
Fiandaca, M. S., Kapogiannis, D., Mapstone, M., Boxer, A., Eitan, E., Schwartz, J. B., et al. (2015). Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally derived blood exosomes: a case-control study. Alzheimers Dement 11, 600–607e601. doi: 10.1016/j.jalz.2014.06.008
Fitzner, D., Schnaars, M., van Rossum, D., Krishnamoorthy, G., Dibaj, P., Bakhti, M., et al. (2011). Selective transfer of exosomes from oligodendrocytes to microglia by macropinocytosis. J. Cell Sci. 124(Pt 3), 447–458. doi: 10.1242/jcs.074088
Frohlich, D., Kuo, W. P., Fruhbeis, C., Sun, J. J., Zehendner, C. M., Luhmann, H. J., et al. (2014). Multifaceted effects of oligodendroglial exosomes on neurons: impact on neuronal firing rate, signal transduction and gene regulation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369:20130510. doi: 10.1098/rstb.2013.0510
Fruhbeis, C., Frohlich, D., and Kramer-Albers, E. M. (2012). Emerging roles of exosomes in neuron-glia communication. Front. Physiol. 3:119. doi: 10.3389/fphys.2012.00119
Fruhbeis, C., Frohlich, D., Kuo, W. P., Amphornrat, J., Thilemann, S., Saab, A. S., et al. (2013). Neurotransmitter-triggered transfer of exosomes mediates oligodendrocyte-neuron communication. PLoS Biol. 11:e1001604. doi: 10.1371/journal.pbio.1001604
Fu, Y., Jiang, C., Tofaris, G. K., and Davis, J. J. (2020). A facile impedimetric analysis of neuronal exosome markers in Parkinson’s disease diagnostics. Anal. Chem. 92, 13647–13651. doi: 10.1021/acs.analchem.0c03092
Gibbings, D. J., Ciaudo, C., Erhardt, M., and Voinnet, O. (2009). Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity. Nat. Cell Biol. 11, 1143–1149. doi: 10.1038/ncb1929
Glebov, K., Löchner, M., Jabs, R., Lau, T., Merkel, O., Schloss, P., et al. (2015). Serotonin stimulates secretion of exosomes from microglia cells. Glia 63, 626–634. doi: 10.1002/glia.22772
Goetzl, E. J., Abner, E. L., Jicha, G. A., Kapogiannis, D., and Schwartz, J. B. (2018). Declining levels of functionally specialized synaptic proteins in plasma neuronal exosomes with progression of Alzheimer’s disease. FASEB J. 32, 888–893. doi: 10.1096/fj.201700731R
Goetzl, E. J., Boxer, A., Schwartz, J. B., Abner, E. L., Petersen, R. C., Miller, B. L., et al. (2015). Low neural exosomal levels of cellular survival factors in Alzheimer’s disease. Ann. Clin. Trans. Neurol. 2, 769–773. doi: 10.1002/acn3.211
Goetzl, E. J., Kapogiannis, D., Schwartz, J. B., Lobach, I. V., Goetzl, L., Abner, E. L., et al. (2016). Decreased synaptic proteins in neuronal exosomes of frontotemporal dementia and Alzheimer’s disease. FASEB J. 30, 4141–4148. doi: 10.1096/fj.201600816R
Goldie, B. J., Dun, M. D., Lin, M., Smith, N. D., Verrills, N. M., Dayas, C. V., et al. (2014). Activity-associated miRNA are packaged in Map1b-enriched exosomes released from depolarized neurons. Nucleic Acids Res. 42, 9195–9208. doi: 10.1093/nar/gku594
Gomes, C., Keller, S., Altevogt, P., and Costa, J. (2007). Evidence for secretion of Cu, Zn superoxide dismutase via exosomes from a cell model of amyotrophic lateral sclerosis. Neurosci. Lett. 428, 43–46. doi: 10.1016/j.neulet.2007.09.024
Grad, L. I., Yerbury, J. J., Turner, B. J., Guest, W. C., Pokrishevsky, E., O’Neill, M. A., et al. (2014). Intercellular propagated misfolding of wild-type Cu/Zn superoxide dismutase occurs via exosome-dependent and-independent mechanisms. Proc. Natl. Acad. Sci. U S A. 111, 3620–3625. doi: 10.1073/pnas.1312245111
Granseth, B., Odermatt, B., Royle, S. J., and Lagnado, L. (2006). Clathrin-mediated endocytosis is the dominant mechanism of vesicle retrieval at hippocampal synapses. Neuron 51, 773–786. doi: 10.1016/j.neuron.2006.08.029
Grey, M., Dunning, C. J., Gaspar, R., Grey, C., Brundin, P., Sparr, E., et al. (2015). Acceleration of α-synuclein aggregation by exosomes. J. Biol. Chem. 290, 2969–2982. doi: 10.1074/jbc.M114.585703
Guduric-Fuchs, J., O’Connor, A., Camp, B., O’Neill, C. L., Medina, R. J., and Simpson, D. A. (2012). Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genom. 13:357. doi: 10.1186/1471-2164-13-357
Guescini, M., Genedani, S., Stocchi, V., and Agnati, L. F. (2010). Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J. Neural Transm. (Vienna) 117, 1–4. doi: 10.1007/s00702-009-0288-288
Guitart, K., Loers, G., Buck, F., Bork, U., Schachner, M., and Kleene, R. (2016). Improvement of neuronal cell survival by astrocyte-derived exosomes under hypoxic and ischemic conditions depends on prion protein. Glia 64, 896–910. doi: 10.1002/glia.22963
Guix, F. X., Corbett, G. T., Cha, D. J., Mustapic, M., Liu, W., Mengel, D., et al. (2018). Detection of aggregation-competent tau in neuron-derived extracellular vesicles. Int. J. Mol. Sci. 19:663. doi: 10.3390/ijms19030663
Guix, F. X., Sannerud, R., Berditchevski, F., Arranz, A. M., Horré, K., Snellinx, A., et al. (2017). Tetraspanin 6: a pivotal protein of the multiple vesicular body determining exosome release and lysosomal degradation of amyloid precursor protein fragments. Mol. Neurodegeneration 12:25. doi: 10.1186/s13024-017-0165-160
Han, C., Xiong, N., Guo, X., Huang, J., Ma, K., Liu, L., et al. (2019). Exosomes from patients with Parkinson’s disease are pathological in mice. J. Mol. Med. 97, 1329–1344. doi: 10.1007/s00109-019-01810-z
Hao, Z. C., Lu, J., Wang, S. Z., Wu, H., Zhang, Y. T., and Xu, S. G. (2017). Stem cell-derived exosomes: a promising strategy for fracture healing. Cell Prolif. 50:e12359. doi: 10.1111/cpr.12359
Henne, W. M., Buchkovich, N. J., and Emr, S. D. (2011). The ESCRT pathway. Dev. Cell 21, 77–91. doi: 10.1016/j.devcel.2011.05.015
Ho, D. H., Yi, S., Seo, H., Son, I., and Seol, W. (2014). Increased DJ-1 in urine exosome of Korean males with Parkinson’s disease. BioMed. Res. Int. 2014:704678. doi: 10.1155/2014/704678
Hong, Y., Zhao, T., Li, X.-J., and Li, S. (2017). Mutant huntingtin inhibits αB-crystallin expression and impairs exosome secretion from astrocytes. J. Neurosci. 37, 9550–9563. doi: 10.1523/JNEUROSCI.1418-17.2017
Hsu, C., Morohashi, Y., Yoshimura, S., Manrique-Hoyos, N., Jung, S., Lauterbach, M. A., et al. (2010). Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J. Cell Biol. 189, 223–232. doi: 10.1083/jcb.200911018
Huntington’s Disease Collaborative Research Group (1993). A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. the Huntington’s disease collaborative research group. Cell 72, 971–983.
Huotari, J., and Helenius, A. (2011). Endosome maturation. EMBO J. 30, 3481–3500. doi: 10.1038/emboj.2011.286
Ibrahim, A. G., Cheng, K., and Marbán, E. (2014). Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep. 2, 606–619. doi: 10.1016/j.stemcr.2014.04.006
Itakura, E., Kishi-Itakura, C., and Mizushima, N. (2012). The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell 151, 1256–1269. doi: 10.1016/j.cell.2012.11.001
Izquierdo-Useros, N., Naranjo-Gomez, M., Archer, J., Hatch, S. C., Erkizia, I., Blanco, J., et al. (2009). Capture and transfer of HIV-1 particles by mature dendritic cells converges with the exosome-dissemination pathway. Blood 113, 2732–2741. doi: 10.1182/blood-2008-05-158642
Jaiswal, J. K., Andrews, N. W., and Simon, S. M. (2002). Membrane proximal lysosomes are the major vesicles responsible for calcium-dependent exocytosis in nonsecretory cells. J. Cell Biol. 159, 625–635. doi: 10.1083/jcb.200208154
Janas, T., Janas, T., and Yarus, M. (2004). A membrane transporter for tryptophan composed of RNA. RNA 10, 1541–1549. doi: 10.1261/rna.7112704
Jeon, I., Cicchetti, F., Cisbani, G., Lee, S., Li, E., Bae, J., et al. (2016). Human-to-mouse prion-like propagation of mutant huntingtin protein. Acta Neuropathol. 132, 577–592. doi: 10.1007/s00401-016-1582-1589
Jeppesen, D. K., Fenix, A. M., Franklin, J. L., Higginbotham, J. N., Zhang, Q., Zimmerman, L. J., et al. (2019). Reassessment of exosome composition. Cell 177, 428-445.18. doi: 10.1016/j.cell.2019.02.029
Jia, L., Zhu, M., Kong, C., Pang, Y., Zhang, H., Qiu, Q., et al. (2020). Blood neuro-exosomal synaptic proteins predict Alzheimer’s disease at the asymptomatic stage. Alzheimers Dement 17, 49–60. doi: 10.1002/alz.12166
Jiang, C., Hopfner, F., Katsikoudi, A., Hein, R., Catli, C., Evetts, S., et al. (2020). Serum neuronal exosomes predict and differentiate Parkinson’s disease from atypical parkinsonism. J. Neurol. Neurosurg. Psychiatry 91, 720–729. doi: 10.1136/jnnp-2019-322588
Jiang, P., Nishimura, T., Sakamaki, Y., Itakura, E., Hatta, T., Natsume, T., et al. (2014). The HOPS complex mediates autophagosome-lysosome fusion through interaction with syntaxin 17. Mol. Biol. Cell 25, 1327–1337. doi: 10.1091/mbc.E13-08-0447
Kalra, H., Simpson, R. J., Ji, H., Aikawa, E., Altevogt, P., Askenase, P., et al. (2012). Vesiclepedia: a compendium for extracellular vesicles with continuous community annotation. PLoS Biol. 10:e1001450. doi: 10.1371/journal.pbio.1001450
Katsu, M., Hama, Y., Utsumi, J., Takashina, K., Yasumatsu, H., Mori, F., et al. (2019). MicroRNA expression profiles of neuron-derived extracellular vesicles in plasma from patients with amyotrophic lateral sclerosis. Neurosci. Lett. 708:134176. doi: 10.1016/j.neulet.2019.03.048
Kim, D. K., Kang, B., Kim, O. Y., Choi, D. S., Lee, J., Kim, S. R., et al. (2013). EVpedia: an integrated database of high-throughput data for systemic analyses of extracellular vesicles. J. Extracell. Vesicles 2:20384. doi: 10.3402/jev.v2i0.20384
Kong, S. M. Y., Chan, B. K. K., Park, J.-S., Hill, K. J., Aitken, J. B., Cottle, L., et al. (2014). Parkinson’s disease-linked human PARK9/ATP13A2 maintains zinc homeostasis and promotes α-Synuclein externalization via exosomes. Hum. Mol. Genet. 23, 2816–2833. doi: 10.1093/hmg/ddu099
Koppers-Lalic, D., Hackenberg, M., Bijnsdorp, I. V., van Eijndhoven, M. A. J., Sadek, P., Sie, D., et al. (2014). Nontemplated nucleotide additions distinguish the small RNA composition in cells from exosomes. Cell Rep. 8, 1649–1658. doi: 10.1016/j.celrep.2014.08.027
Kordower, J. H., Chu, Y., Hauser, R. A., Freeman, T. B., and Olanow, C. W. (2008). Lewy body–like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat. Med. 14, 504–506. doi: 10.1038/nm1747
Korkut, C., Ataman, B., Ramachandran, P., Ashley, J., Barria, R., Gherbesi, N., et al. (2009). Trans-synaptic transmission of vesicular Wnt signals through Evi/Wntless. Cell 139, 393–404. doi: 10.1016/j.cell.2009.07.051
Korkut, C., Li, Y., Koles, K., Brewer, C., Ashley, J., Yoshihara, M., et al. (2013). Regulation of postsynaptic retrograde signaling by presynaptic exosome release. Neuron 77, 1039–1046. doi: 10.1016/j.neuron.2013.01.013
Kowal, J., Arras, G., Colombo, M., Jouve, M., Morath, J. P., Primdal-Bengtson, B., et al. (2016). Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. U S A. 113, E968–E977. doi: 10.1073/pnas.1521230113
Kowal, J., Tkach, M., and Thery, C. (2014). Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 29, 116–125. doi: 10.1016/j.ceb.2014.05.004
Kramer-Albers, E. M., Bretz, N., Tenzer, S., Winterstein, C., Mobius, W., Berger, H., et al. (2007). Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: trophic support for axons? Proteom. Clin. Appl. 1, 1446–1461. doi: 10.1002/prca.200700522
Kramer-Albers, E. M., and White, R. (2011). From axon-glial signalling to myelination: the integrating role of oligodendroglial Fyn kinase. Cell Mol. Life Sci. 68, 2003–2012. doi: 10.1007/s00018-010-0616-z
Kufareva, I., Lenoir, M., Dancea, F., Sridhar, P., Raush, E., Bissig, C., et al. (2014). Discovery of novel membrane binding structures and functions. Biochem. Cell Biol. 92, 555–563. doi: 10.1139/bcb-2014-2074
Lachenal, G., Pernet-Gallay, K., Chivet, M., Hemming, F. J., Belly, A., Bodon, G., et al. (2011). Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol. Cell Neurosci. 46, 409–418. doi: 10.1016/j.mcn.2010.11.004
Lafourcade, C., Ramirez, J. P., Luarte, A., Fernandez, A., and Wyneken, U. (2016). MiRNAs in astrocyte-derived exosomes as possible mediators of neuronal plasticity. J. Exp. Neurosci. 10, (Suppl. 1), 1–9. doi: 10.4137/JEN.S39916
Laulagnier, K., Javalet, C., Hemming, F. J., Chivet, M., Lachenal, G., Blot, B., et al. (2018). Amyloid precursor protein products concentrate in a subset of exosomes specifically endocytosed by neurons. Cell. Mol. Life Sci. 75, 757–773. doi: 10.1007/s00018-017-2664-2660
Lin, M. C., Chen, S. Y., Tsai, H. M., He, P. L., Lin, Y. C., Herschman, H., et al. (2017). PGE2/EP4 signaling controls the transfer of the mammary stem cell state by lipid rafts in extracellular vesicles. Stem Cells 35, 425–444. doi: 10.1002/stem.2476
Liu, W., Bai, X., Zhang, A., Huang, J., Xu, S., and Zhang, J. (2019). Role of exosomes in central nervous system diseases. Front. Mol. Neurosci. 12:240. doi: 10.3389/fnmol.2019.00240
Llorente, A., Skotland, T., Sylvanne, T., Kauhanen, D., Rog, T., Orlowski, A., et al. (2013). Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim Biophys. Acta 1831, 1302–1309. doi: 10.1016/j.bbalip.2013.04.011
Llorente, A., van Deurs, B., and Sandvig, K. (2007). Cholesterol regulates prostasome release from secretory lysosomes in PC-3 human prostate cancer cells. Eur. J. Cell Biol. 86, 405–415. doi: 10.1016/j.ejcb.2007.05.001
Lopes, C., Tang, Y., Anjo, S. I., Manadas, B., Onofre, I., De Almeida, L. P., et al. (2020). Mitochondrial and redox modifications in huntington disease induced pluripotent stem cells rescued by CRISPR/Cas9 CAGs targeting. Front. Cell Dev. Biol. 8:576592. doi: 10.3389/fcell.2020.576592
Lopez-Verrilli, M. A., Picou, F., and Court, F. A. (2013). Schwann cell-derived exosomes enhance axonal regeneration in the peripheral nervous system. Glia 61, 1795–1806. doi: 10.1002/glia.22558
Lötvall, J., Hill, A. F., Hochberg, F., Buzás, E. I., Di Vizio, D., Gardiner, C., et al. (2014). Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the international society for extracellular vesicles. J. Extracell. Vesicles. 3:26913. doi: 10.3402/jev.v3.26913
Luarte, A., Cisternas, P., Caviedes, A., Batiz, L. F., Lafourcade, C., Wyneken, U., et al. (2017). Astrocytes at the hub of the stress response: potential modulation of neurogenesis by miRNAs in astrocyte-derived exosomes. Stem Cells Int. 2017:1719050. doi: 10.1155/2017/1719050
Ludwig, N., Whiteside, T. L., and Reichert, T. E. (2019). Challenges in exosome isolation and analysis in health and disease. Int. J. Mol. Sci. 20:4684. doi: 10.3390/ijms20194684
Ma, Y., Li, C., Huang, Y., Wang, Y., Xia, X., and Zheng, J. C. (2019). Exosomes released from neural progenitor cells and induced neural progenitor cells regulate neurogenesis through miR-21a. Cell Commun. Signal. 17:96. doi: 10.1186/s12964-019-0418-413
MacDonald, C., Buchkovich, N. J., Stringer, D. K., Emr, S. D., and Piper, R. C. (2012). Cargo ubiquitination is essential for multivesicular body intralumenal vesicle formation. EMBO Rep. 13, 331–338. doi: 10.1038/embor.2012.18
Mathivanan, S., Ji, H., and Simpson, R. J. (2010). Exosomes: extracellular organelles important in intercellular communication. J. Proteomics 73, 1907–1920. doi: 10.1016/j.jprot.2010.06.006
Mathivanan, S., and Simpson, R. J. (2009). ExoCarta: a compendium of exosomal proteins and RNA. Proteomics 9, 4997–5000. doi: 10.1002/pmic.200900351
McKenzie, A. J., Hoshino, D., Hong, N. H., Cha, D. J., Franklin, J. L., Coffey, R. J., et al. (2016). KRAS-MEK signaling controls Ago2 sorting into exosomes. Cell Rep. 15, 978–987. doi: 10.1016/j.celrep.2016.03.085
Mears, R., Craven, R. A., Hanrahan, S., Totty, N., Upton, C., Young, S. L., et al. (2004). Proteomic analysis of melanoma-derived exosomes by two-dimensional polyacrylamide gel electrophoresis and mass spectrometry. Proteomics 4, 4019–4031. doi: 10.1002/pmic.200400876
Mejzini, R., Flynn, L. L., Pitout, I. L., Fletcher, S., Wilton, S. D., and Akkari, P. A. (2019). ALS genetics. mechanisms, and therapeutics: where are we now? Front. Neurosci. 13:1310. doi: 10.3389/fnins.2019.01310
Melo, S. A., Sugimoto, H., O’Connell, J. T., Kato, N., Villanueva, A., Vidal, A., et al. (2014). Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell 26, 707–721. doi: 10.1016/j.ccell.2014.09.005
Men, Y., Yelick, J., Jin, S., Tian, Y., Chiang, M. S. R., Higashimori, H., et al. (2019). Exosome reporter mice reveal the involvement of exosomes in mediating neuron to astroglia communication in the CNS. Nat. Commun. 10:4136. doi: 10.1038/s41467-019-11534-w
Menck, K., Sonmezer, C., Worst, T. S., Schulz, M., Dihazi, G. H., Streit, F., et al. (2017). Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J. Extracell. Vesicles 6:1378056. doi: 10.1080/20013078.2017.1378056
Mendt, M., Rezvani, K., and Shpall, E. (2019). Mesenchymal stem cell-derived exosomes for clinical use. Bone Marrow Transplant 54, (Suppl. 2), 789–792. doi: 10.1038/s41409-019-0616-z
Miranda, A. M., Lasiecka, Z. M., Xu, Y., Neufeld, J., Shahriar, S., Simoes, S., et al. (2018). Neuronal lysosomal dysfunction releases exosomes harboring APP C-terminal fragments and unique lipid signatures. Nat. Commun. 9:291. doi: 10.1038/s41467-017-02533-w
Mittelbrunn, M., Vicente-Manzanares, M., and Sánchez-Madrid, F. (2015). Organizing polarized delivery of exosomes at synapses. Traffic 16, 327–337. doi: 10.1111/tra.12258
Morel, L., Regan, M., Higashimori, H., Ng, S. K., Esau, C., Vidensky, S., et al. (2013). Neuronal exosomal miRNA-dependent translational regulation of astroglial glutamate transporter GLT1. J. Biol. Chem. 288, 7105–7116. doi: 10.1074/jbc.M112.410944
Morelli, A. E., Larregina, A. T., Shufesky, W. J., Sullivan, M. L., Stolz, D. B., Papworth, G. D., et al. (2004). Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood 104, 3257–3266. doi: 10.1182/blood-2004-03-0824
Morton, M. C., Neckles, V. N., Seluzicki, C. M., Holmberg, J. C., and Feliciano, D. M. (2018). Neonatal subventricular zone neural stem cells release extracellular vesicles that act as a microglial morphogen. Cell Rep. 23, 78–89. doi: 10.1016/j.celrep.2018.03.037
Mulcahy, L. A., Pink, R. C., and Carter, D. R. (2014). Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 3. doi: 10.3402/jev.v3.24641
Munich, S., Sobo-Vujanovic, A., Buchser, W. J., Beer-Stolz, D., and Vujanovic, N. L. (2012). Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. Oncoimmunology 1, 1074–1083. doi: 10.4161/onci.20897
Nanbo, A., Kawanishi, E., Yoshida, R., and Yoshiyama, H. (2013). Exosomes derived from Epstein-Barr virus-infected cells are internalized via caveola-dependent endocytosis and promote phenotypic modulation in target cells. J. Virol. 87, 10334–10347. doi: 10.1128/JVI.01310-1313
Niu, M., Li, Y., Li, G., Zhou, L., Luo, N., Yao, M., et al. (2020). A longitudinal study on α-synuclein in plasma neuronal exosomes as a biomarker for Parkinson’s disease development and progression. Eur. J. Neurol. 27, 967–974. doi: 10.1111/ene.14208
Oh, M., Lee, J., Kim, Y. J., Rhee, W. J., and Park, J. H. (2018). Exosomes derived from human induced pluripotent stem cells ameliorate the aging of skin fibroblasts. Int. J. Mol. Sci. 19:1715. doi: 10.3390/ijms19061715
Ohmichi, T., Mitsuhashi, M., Tatebe, H., Kasai, T., El-Agnaf, O. M. A., and Tokuda, T. (2019). Quantification of brain-derived extracellular vesicles in plasma as a biomarker to diagnose Parkinson’s and related diseases. Parkinson. Related Dis. 61, 82–87. doi: 10.1016/j.parkreldis.2018.11.021
Ostrowski, M., Carmo, N. B., Krumeich, S., Fanget, I., Raposo, G., Savina, A., et al. (2010). Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 12, su11–su13. doi: 10.1038/ncb2000
Palmqvist, S., Schöll, M., Strandberg, O., Mattsson, N., Stomrud, E., Zetterberg, H., et al. (2017). Earliest accumulation of β-amyloid occurs within the default-mode network and concurrently affects brain connectivity. Nat. Commun. 8:1214. doi: 10.1038/s41467-017-01150-x
Papadopoulos, V. E., Nikolopoulou, G., Antoniadou, I., Karachaliou, A., Arianoglou, G., Emmanouilidou, E., et al. (2018). Modulation of β-glucocerebrosidase increases α-synuclein secretion and exosome release in mouse models of Parkinson’s disease. Hum. Mol. Genet. 27, 1696–1710. doi: 10.1093/hmg/ddy075
Parolini, I., Federici, C., Raggi, C., Lugini, L., Palleschi, S., De Milito, A., et al. (2009). Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 284, 34211–34222. doi: 10.1074/jbc.M109.041152
Perez-Gonzalez, R., Gauthier, S. A., Kumar, A., and Levy, E. (2012). The exosome secretory pathway transports amyloid precursor protein carboxyl-terminal fragments from the cell into the brain extracellular space. J. Biol. Chem. 287, 43108–43115. doi: 10.1074/jbc.M112.404467
Peruzzotti-Jametti, L., Bernstock, J. D., Manferrari, G., Rogall, R., Fernandez-Vizarra, E., Williamson, J. C., et al. (2020). Neural stem cells traffic functional mitochondria via extracellular vesicles to correct mitochondrial dysfunction in target cells. bioRxiv [preprint] doi: 10.1101/2020.01.29.923441
Pinto, S., Cunha, C., Barbosa, M., Vaz, A. R., and Brites, D. (2017). Exosomes from NSC-34 cells transfected with hSOD1-G93A are enriched in miR-124 and drive alterations in microglia phenotype. Front. Neurosci. 11:273. doi: 10.3389/fnins.2017.00273
Polanco, J. C., Li, C., Durisic, N., Sullivan, R., and Gotz, J. (2018). Exosomes taken up by neurons hijack the endosomal pathway to spread to interconnected neurons. Acta Neuropathol. Commun. 6:10. doi: 10.1186/s40478-018-0514-514
Pols, M. S., and Klumperman, J. (2009). Trafficking and function of the tetraspanin CD63. Exp Cell Res. 315, 1584–1592. doi: 10.1016/j.yexcr.2008.09.020
Potolicchio, I., Carven, G. J., Xu, X., Stipp, C., Riese, R. J., Stern, L. J., et al. (2005). Proteomic analysis of microglia-derived exosomes: metabolic role of the aminopeptidase CD13 in neuropeptide catabolism. J. Immunol. 175, 2237–2243. doi: 10.4049/jimmunol.175.4.2237
Putz, U., Howitt, J., Lackovic, J., Foot, N., Kumar, S., Silke, J., et al. (2008). Nedd4 family-interacting protein 1 (Ndfip1) is required for the exosomal secretion of Nedd4 family proteins. J. Biol. Chem. 283, 32621–32627. doi: 10.1074/jbc.M804120200
Rajendran, L., Honsho, M., Zahn, T. R., Keller, P., Geiger, K. D., Verkade, P., et al. (2006). Alzheimer’s disease β-amyloid peptides are released in association with exosomes. Proc. Natl. Acad. Sci. U S A. 103, 11172–11177. doi: 10.1073/pnas.0603838103
Raposo, G., and Stoorvogel, W. (2013). Extracellular vesicles: exosomes, microvesicles, and friends. J. Cell Biol. 200, 373–383. doi: 10.1083/jcb.201211138
Record, M., Carayon, K., Poirot, M., and Silvente-Poirot, S. (2014). Exosomes as new vesicular lipid transporters involved in cell-cell communication and various pathophysiologies. Biochim Biophys. Acta 1841, 108–120. doi: 10.1016/j.bbalip.2013.10.004
Ridder, K., Keller, S., Dams, M., Rupp, A. K., Schlaudraff, J., Del Turco, D., et al. (2014). Extracellular vesicle-mediated transfer of genetic information between the hematopoietic system and the brain in response to inflammation. PLoS Biol. 12:e1001874. doi: 10.1371/journal.pbio.1001874
Rong, Y., Liu, W., Wang, J., Fan, J., Luo, Y., Li, L., et al. (2019). Neural stem cell-derived small extracellular vesicles attenuate apoptosis and neuroinflammation after traumatic spinal cord injury by activating autophagy. Cell Death Dis. 10:340. doi: 10.1038/s41419-019-1571-1578
Roos, R. A. (2010). Huntington’s disease: a clinical review. Orphanet J. Rare Dis. 5:40. doi: 10.1186/1750-1172-5-40
Ruan, Z., Pathak, D., Venkatesan Kalavai, S., Yoshii-Kitahara, A., Muraoka, S., Bhatt, N., et al. (2021). Alzheimer’s disease brain-derived extracellular vesicles spread tau pathology in interneurons. Brain 144, 288–309. doi: 10.1093/brain/awaa376
Russo, F., Di Bella, S., Vannini, F., Berti, G., Scoyni, F., Cook, H. V., et al. (2018). miRandola 2017: a curated knowledge base of non-invasive biomarkers. Nucleic Acids Res. 46, D354–D359. doi: 10.1093/nar/gkx854
Saito, N., Okada, Y., Noda, Y., Kinoshita, Y., Kondo, S., and Hirokawa, N. (1997). KIFC2 is a novel neuron-specific C-terminal type kinesin superfamily motor for dendritic transport of multivesicular body-like organelles. Neuron 18, 425–438. doi: 10.1016/s0896-6273(00)81243-x
Saman, S., Kim, W., Raya, M., Visnick, Y., Miro, S., Saman, S., et al. (2012). Exosome-associated tau is secreted in tauopathy models and is selectively phosphorylated in cerebrospinal fluid in early Alzheimer disease. J. Biol. Chem. 287, 3842–3849. doi: 10.1074/jbc.M111.277061
Sansone, P., Savini, C., Kurelac, I., Chang, Q., Amato, L. B., Strillacci, A., et al. (2017). Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy-resistant breast cancer. Proc. Natl. Acad. Sci. U S A. 114, E9066–E9075. doi: 10.1073/pnas.1704862114
Sharma, P., Mesci, P., Carromeu, C., McClatchy, D. R., Schiapparelli, L., Yates, J. R., et al. (2019). Exosomes regulate neurogenesis and circuit assembly. Proc. Natl. Acad. Sci. U S A. 116, 16086–16094. doi: 10.1073/pnas.1902513116
Sharples, R. A., Vella, L. J., Nisbet, R. M., Naylor, R., Perez, K., Barnham, K. J., et al. (2008). Inhibition of gamma-secretase causes increased secretion of amyloid precursor protein C-terminal fragments in association with exosomes. FASEB J. 22, 1469–1478. doi: 10.1096/fj.07-9357com
Shi, M., Liu, C., Cook, T. J., Bullock, K. M., Zhao, Y., Ginghina, C., et al. (2014). Plasma exosomal α-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol. 128, 639–650. doi: 10.1007/s00401-014-1314-y
Silverman, J. M., Christy, D., Shyu, C. C., Moon, K.-M., Fernando, S., Gidden, Z., et al. (2019). CNS-derived extracellular vesicles from superoxide dismutase 1 (SOD1) G93A ALS mice originate from astrocytes and neurons and carry misfolded SOD1. J. Biol. Chem. 294, 3744–3759. doi: 10.1074/jbc.RA118.004825
Simpson, R. J., Jensen, S. S., and Lim, J. W. (2008). Proteomic profiling of exosomes: current perspectives. Proteomics 8, 4083–4099. doi: 10.1002/pmic.200800109
Sinha, M. S., Ansell-Schultz, A., Civitelli, L., Hildesjö, C., Larsson, M., Lannfelt, L., et al. (2018). Alzheimer’s disease pathology propagation by exosomes containing toxic amyloid-beta oligomers. Acta Neuropathol. 136, 41–56. doi: 10.1007/s00401-018-1868-1861
Skotland, T., Sandvig, K., and Llorente, A. (2017). Lipids in exosomes: current knowledge and the way forward. Prog. Lipid Res. 66, 30–41. doi: 10.1016/j.plipres.2017.03.001
Soubannier, V., McLelland, G. L., Zunino, R., Braschi, E., Rippstein, P., Fon, E. A., et al. (2012). A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol. 22, 135–141. doi: 10.1016/j.cub.2011.11.057
Sperling, R., Mormino, E., and Johnson, K. (2014). The evolution of preclinical Alzheimer’s disease: implications for prevention trials. Neuron 84, 608–622. doi: 10.1016/j.neuron.2014.10.038
Stevanato, L., Thanabalasundaram, L., Vysokov, N., and Sinden, J. D. (2016). Investigation of content, stoichiometry and transfer of miRNA from human neural stem cell line derived exosomes. PLoS One 11:e0146353. doi: 10.1371/journal.pone.0146353
Strauss, K., Goebel, C., Runz, H., Mobius, W., Weiss, S., Feussner, I., et al. (2010). Exosome secretion ameliorates lysosomal storage of cholesterol in Niemann-Pick type C disease. J. Biol. Chem. 285, 26279–26288. doi: 10.1074/jbc.M110.134775
Stuendl, A., Kunadt, M., Kruse, N., Bartels, C., Moebius, W., Danzer, K. M., et al. (2016). Induction of α-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain 139, 481–494. doi: 10.1093/brain/awv346
Sun, T., Ding, Z.-X., Luo, X., Liu, Q.-S., and Cheng, Y. (2020). Blood exosomes have neuroprotective effects in a mouse model of Parkinson’s disease. Oxid. Med. Cell. Longevity 2020:3807476. doi: 10.1155/2020/3807476
Svensson, K. J., Christianson, H. C., Wittrup, A., Bourseau-Guilmain, E., Lindqvist, E., Svensson, L. M., et al. (2013). Exosome uptake depends on ERK1/2-heat shock protein 27 signaling and lipid Raft-mediated endocytosis negatively regulated by caveolin-1. J. Biol. Chem. 288, 17713–17724. doi: 10.1074/jbc.M112.445403
Takahashi, A., Okada, R., Nagao, K., Kawamata, Y., Hanyu, A., Yoshimoto, S., et al. (2017). Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun. 8:15287. doi: 10.1038/ncomms15287
Takahashi, R. H., Milner, T. A., Li, F., Nam, E. E., Edgar, M. A., Yamaguchi, H., et al. (2002). Intraneuronal Alzheimer Aβ42 accumulates in multivesicular bodies and is associated with synaptic pathology. Am. J. Pathol. 161, 1869–1879. doi: 10.1016/s0002-9440(10)64463-x
Takahashi, Y., Nishikawa, M., Shinotsuka, H., Matsui, Y., Ohara, S., Imai, T., et al. (2013). Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J. Biotechnol. 165, 77–84. doi: 10.1016/j.jbiotec.2013.03.013
Tamboli, I. Y., Barth, E., Christian, L., Siepmann, M., Kumar, S., Singh, S., et al. (2010). Statins promote the degradation of extracellular amyloid β-peptide by microglia via stimulation of exosome-associated insulin-degrading enzyme (IDE) secretion. J. Biol. Chem. 285, 37405–37414. doi: 10.1074/jbc.M110.149468
Taylor, A. R., Robinson, M. B., Gifondorwa, D. J., Tytell, M., and Milligan, C. E. (2007). Regulation of heat shock protein 70 release in astrocytes: role of signaling kinases. Dev. Neurobiol. 67, 1815–1829. doi: 10.1002/dneu.20559
Thakur, B. K., Zhang, H., Becker, A., Matei, I., Huang, Y., Costa-Silva, B., et al. (2014). Double-stranded DNA in exosomes: a novel biomarker in cancer detection. Cell Res. 24, 766–769. doi: 10.1038/cr.2014.44
Théry, C., Witwer, K. W., Aikawa, E., Alcaraz, M. J., Anderson, J. D., Andriantsitohaina, R., et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 7:1535750. doi: 10.1080/20013078.2018.1535750
Tian, T., Zhu, Y. L., Hu, F. H., Wang, Y. Y., Huang, N. P., and Xiao, Z. D. (2013). Dynamics of exosome internalization and trafficking. J. Cell Physiol. 228, 1487–1495. doi: 10.1002/jcp.24304
Tomlinson, P. R., Zheng, Y., Fischer, R., Heidasch, R., Gardiner, C., Evetts, S., et al. (2015). Identification of distinct circulating exosomes in Parkinson’s disease. Ann. Clin. Trans. Neurol. 2, 353–361. doi: 10.1002/acn3.175
Trajkovic, K., Hsu, C., Chiantia, S., Rajendran, L., Wenzel, D., Wieland, F., et al. (2008). Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 319, 1244–1247. doi: 10.1126/science.1153124
Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J. J., and Lotvall, J. O. (2007). Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654–659. doi: 10.1038/ncb1596
Van Niel, G., Charrin, S., Simoes, S., Romao, M., Rochin, L., Saftig, P., et al. (2011). The tetraspanin CD63 regulates ESCRT-independent and-dependent endosomal sorting during melanogenesis. Dev. Cell 21, 708–721. doi: 10.1016/j.devcel.2011.08.019
Varcianna, A., Myszczynska, M. A., Castelli, L. M., O’Neill, B., Kim, Y., Talbot, J., et al. (2019). Micro-RNAs secreted through astrocyte-derived extracellular vesicles cause neuronal network degeneration in C9orf72 ALS. EBioMedicine 40, 626–635. doi: 10.1016/j.ebiom.2018.11.067
Venturini, A., Passalacqua, M., Pelassa, S., Pastorino, F., Tedesco, M., Cortese, K., et al. (2019). Exosomes from astrocyte processes: signaling to neurons. Front. Pharmacol. 10:1452. doi: 10.3389/fphar.2019.01452
Villarroya-Beltri, C., Baixauli, F., Gutierrez-Vazquez, C., Sanchez-Madrid, F., and Mittelbrunn, M. (2014). Sorting it out: regulation of exosome loading. Semin. Cancer Biol. 28, 3–13. doi: 10.1016/j.semcancer.2014.04.009
Villarroya-Beltri, C., Gutierrez-Vazquez, C., Sanchez-Cabo, F., Perez-Hernandez, D., Vazquez, J., Martin-Cofreces, N., et al. (2013). Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 4:2980. doi: 10.1038/ncomms3980
Vingtdeux, V., Hamdane, M., Loyens, A., Gele, P., Drobeck, H., Begard, S., et al. (2007). Alkalizing drugs induce accumulation of amyloid precursor protein by-products in luminal vesicles of multivesicular bodies. J. Biol. Chem. 282, 18197–18205. doi: 10.1074/jbc.M609475200
Waldenstrom, A., Genneback, N., Hellman, U., and Ronquist, G. (2012). Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS One 7:e34653. doi: 10.1371/journal.pone.0034653
Walsh, D. M., Cha, D. J., Mengel, D., Mustapic, M., Liu, W., Selkoe, D. J., et al. (2019). miR-212 and miR-132 are down-regulated in neurally-derived plasma exosomes of Alzheimer’s patients. Front. Neurosci. 13:1208. doi: 10.3389/fnins.2019.01208
Wang, G., Dinkins, M., He, Q., Zhu, G., Poirier, C., Campbell, A., et al. (2012). Astrocytes secrete exosomes enriched with proapoptotic ceramide and prostate apoptosis response 4 (PAR-4) potential mechanism of apoptosis induction in Alzheimer disease (AD). J. Biol. Chem. 287, 21384–21395. doi: 10.1074/jbc.M112.340513
Wang, S., Cesca, F., Loers, G., Schweizer, M., Buck, F., Benfenati, F., et al. (2011). Synapsin I is an oligomannose-carrying glycoprotein, acts as an oligomannose-binding lectin, and promotes neurite outgrowth and neuronal survival when released via glia-derived exosomes. J. Neurosci. 31, 7275–7290. doi: 10.1523/JNEUROSCI.6476-10.2011
Wang, X., Weidling, I., Koppel, S., Menta, B., Perez Ortiz, J., Kalani, A., et al. (2020). Detection of mitochondria-pertinent components in exosomes. Mitochondrion 55, 100–110. doi: 10.1016/j.mito.2020.09.006
Wang, Y., Balaji, V., Kaniyappan, S., Krüger, L., Irsen, S., Tepper, K., et al. (2017). The release and trans-synaptic transmission of Tau via exosomes. Mol. Neurodegeneration 12, 1–25. doi: 10.1186/s13024-016-0143-y
Wang, Y., Zhang, L., Li, Y., Chen, L., Wang, X., Guo, W., et al. (2015). Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. Int. J. Cardiol. 192, 61–69. doi: 10.1016/j.ijcard.2015.05.020
Webb, R. L., Kaiser, E. E., Jurgielewicz, B. J., Spellicy, S., Scoville, S. L., Thompson, T. A., et al. (2018a). Human neural stem cell extracellular vesicles improve recovery in a porcine model of ischemic stroke. Stroke 49, 1248–1256. doi: 10.1161/STROKEAHA.117.020353
Webb, R. L., Kaiser, E. E., Scoville, S. L., Thompson, T. A., Fatima, S., Pandya, C., et al. (2018b). Human neural stem cell extracellular vesicles improve tissue and functional recovery in the murine thromboembolic stroke model. Transl. Stroke Res. 9, 530–539. doi: 10.1007/s12975-017-0599-592
Witwer, K. W., and Thery, C. (2019). Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell. Vesicles 8:1648167. doi: 10.1080/20013078.2019.1648167
Wyman, S. K., Knouf, E. C., Parkin, R. K., Fritz, B. R., Lin, D. W., Dennis, L. M., et al. (2011). Post-transcriptional generation of miRNA variants by multiple nucleotidyl transferases contributes to miRNA transcriptome complexity. Genome Res. 21, 1450–1461. doi: 10.1101/gr.118059.110
Xia, Y., Zhang, G., Han, C., Ma, K., Guo, X., Wan, F., et al. (2019). Microglia as modulators of exosomal alpha-synuclein transmission. Cell Death Dis. 10:174. doi: 10.1038/s41419-019-1404-1409
Xie, J.-C., Ma, X.-Y., Liu, X.-H., Yu, J., Zhao, Y.-C., Tan, Y., et al. (2018). Hypoxia increases amyloid-β level in exosomes by enhancing the interaction between CD147 and Hook1. Am. J. Transl. Res. 10:150.
Xin, H., Li, Y., Buller, B., Katakowski, M., Zhang, Y., Wang, X., et al. (2012). Exosome-mediated transfer of miR-133b from multipotent mesenchymal stromal cells to neural cells contributes to neurite outgrowth. Stem Cells 30, 1556–1564. doi: 10.1002/stem.1129
Yanez-Mo, M., Siljander, P. R., Andreu, Z., Zavec, A. B., Borras, F. E., Buzas, E. I., et al. (2015). Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 4:27066. doi: 10.3402/jev.v4.27066
Yang, S., Che, S. P., Kurywchak, P., Tavormina, J. L., Gansmo, L. B., and Correa, et al. (2017). Detection of mutant KRAS and TP53 DNA in circulating exosomes from healthy individuals and patients with pancreatic cancer. Cancer Biol. Ther. 18, 158–165. doi: 10.1080/15384047.2017.1281499
Yelick, J., Men, Y., Jin, S., Seo, S., Espejo-Porras, F., and Yang, Y. (2020). Elevated exosomal secretion of miR-124-3p from spinal neurons positively associates with disease severity in ALS. Exp. Neurol. 333:113414. doi: 10.1016/j.expneurol.2020.113414
Yuyama, K., Sun, H., Mitsutake, S., and Igarashi, Y. (2012). Sphingolipid-modulated exosome secretion promotes clearance of amyloid-beta by microglia. J. Biol. Chem. 287, 10977–10989. doi: 10.1074/jbc.M111.324616
Yuyama, K., Sun, H., Sakai, S., Mitsutake, S., Okada, M., Tahara, H., et al. (2014). Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J. Biol. Chem. 289, 24488–24498. doi: 10.1074/jbc.M114.577213
Yuyama, K., Yamamoto, N., and Yanagisawa, K. (2008). Accelerated release of exosome-associated GM1 ganglioside (GM1) by endocytic pathway abnormality: another putative pathway for GM1-induced amyloid fibril formation. J. Neurochem. 105, 217–224. doi: 10.1111/j.1471-4159.2007.05128.x
Zhang, N., Gu, D., Meng, M., and Gordon, M. L. (2020). TDP-43 is elevated in plasma neuronal-derived exosomes of patients with Alzheimer’s disease. Front. Aging Neurosci. 12:166. doi: 10.3389/fnagi.2020.00166
Zhang, X., Abels, E. R., Redzic, J. S., Margulis, J., Finkbeiner, S., and Breakefield, X. O. (2016). Potential transfer of polyglutamine and CAG-repeat RNA in extracellular vesicles in Huntington’s disease: background and evaluation in cell culture. Cell. Mol. Neurobiol. 36, 459–470. doi: 10.1007/s10571-016-0350-357
Zhang, Y., Chopp, M., Liu, X. S., Katakowski, M., Wang, X., Tian, X., et al. (2017a). Exosomes derived from mesenchymal stromal cells promote axonal growth of cortical neurons. Mol. Neurobiol. 54, 2659–2673. doi: 10.1007/s12035-016-9851-9850
Zhang, Y., Kim, M. S., Jia, B., Yan, J., Zuniga-Hertz, J. P., Han, C., et al. (2017b). Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature 548, 52–57. doi: 10.1038/nature23282
Zhao, A., Li, Y., Yan, Y., Qiu, Y., Li, B., Xu, W., et al. (2020). Increased prediction value of biomarker combinations for the conversion of mild cognitive impairment to Alzheimer’s dementia. Transl. Neurodegener. 9:30. doi: 10.1186/s40035-020-00210-215
Keywords: exosomes, central nervous system, neurodegenerative diseases, biomarkers, neural-derived exosomes
Citation: Beatriz M, Vilaça R and Lopes C (2021) Exosomes: Innocent Bystanders or Critical Culprits in Neurodegenerative Diseases. Front. Cell Dev. Biol. 9:635104. doi: 10.3389/fcell.2021.635104
Received: 29 November 2020; Accepted: 06 April 2021;
Published: 13 May 2021.
Edited by:
Oleg Shupliakov, Karolinska Institutet (KI), SwedenReviewed by:
Graca Raposo, Institut Curie, FranceJafar Rezaie, Urmia University of Medical Sciences, Iran
Claudia Verderio, National Research Council, Consiglio Nazionale delle Ricerche (CNR), Italy
Copyright © 2021 Beatriz, Vilaça and Lopes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carla Lopes, Y2FybGFsb3BlczA5QGdtYWlsLmNvbQ==; Y2FybGEubG9wZXNAY25jLnVjLnB0
†These authors have contributed equally to this work