 Ping Li1
Ping Li1 Hong Wei
Hong Wei Tingtao Chen
Tingtao Chen- 1School of Life Sciences, Institute of Translational Medicine, Nanchang University, Nanchang, China
- 2Department of Obstetrics and Gynecology, The Ninth Hospital of Nanchang, Nanchang, China
- 3Department of Obstetrics and Gynecology, The Second Affiliated Hospital of Nanchang University, Nanchang, China
- 4Institute of Precision Medicine, The First Affiliated Hospital, Sun Yat-sen University, Guangzhou, China
Background: The vaginal microbiota is associated with the health of the female reproductive system and the offspring. Lactobacillus crispatus belongs to one of the most important vaginal probiotics, while its role in the agglutination and immobilization of human sperm, fertility, and offspring health is unclear.
Methods: Adherence assays, sperm motility assays, and Ca2+-detecting assays were used to analyze the adherence properties and sperm motility of L. crispatus Lcr-MH175, attenuated Salmonella typhimurium VNP20009, engineered S. typhimurium VNP20009 DNase I, and Escherichia coli O157:H7 in vitro. The rat reproductive model was further developed to study the role of L. crispatus on reproduction and offspring health, using high-throughput sequencing, real-time PCR, and molecular biology techniques.
Results: Our results indicated that L. crispatus, VNP20009, VNP20009 DNase I, and E. coli O157:H7 significantly inhibited the sperm motility in vitro via adversely affecting the sperm intracellular Ca2+ concentration and showed a high adhesion to sperms. The in vivo results indicated that L. crispatus and other tested bacteria greatly reduced the pregnancy rates, but L. crispatus had a positive effect on maternal health and offspring development. Moreover, the transplantation of L. crispatus could sustain a normal bacterial composition of the vaginal microbiota in healthy rats and markedly reduced the expression of uterine inflammatory factors (toll-like receptor-4/nuclear factor kappa-B, tumor necrosis factor-α, production of interleukin-1β, etc.) and apoptosis factors (Fas Ligand, Bcl-2-associated X protein/B cell lymphoma-2, etc.) compared with the other tested strains.
Conclusion: Our study demonstrated that the vaginal probiotic L. crispatus greatly affected the sperm activity and could also reduce pregnancies through its adhesion property, which might account for some unexplained infertility. Therefore, more caution should be paid when using L. crispatus as a vaginal viable preparation in women of child-bearing age, especially for women whose partners have abnormal sperms.
Introduction
Multicellular organisms co-evolved with a complex coalition of acellular viruses, bacteria, fungi, and protist, collectively known as the microbiota (Belkaid and Hand, 2014). In mammals, the microbiota that reside in the vagina may interact with the host in a complex series of physiologic processes (Chen et al., 2017), including development, metabolism, and immune function (Cohen, 2016; Halmos and Suba, 2016). In recent years, there is an increasing attention on the vaginal microbiota specific to female reproductive and offspring health (Chen et al., 2017), and published studies have indicated that the vaginal microbiota plays a crucial role not only in health and disease but also in fertilization and pregnancy (Younes et al., 2018).
The human vagina is an important channel for women to connect the internal and the external genitalia, for sexual intercourse, for menstrual blood to flow out, and for fetal delivery (Pellati et al., 2008). The vaginal discharge provides nutrients for the microbiota, among which the healthy vaginal microbiota is mainly dominated by Lactobacillus spp., such as Lactobacillus iners, Lactobacillus crispatus, Lactobacillus jensenii, and Lactobacillus gasseri (Ling et al., 2010). They exert important promoting effects in maintaining the health condition of the female genital system (Petrova et al., 2015). L. crispatus appears to be substantially superior to the other Lactobacillus species (Shipitsyna et al., 2013) for its benefits of promoting the stability of normal vaginal microbiota (Verstraelen et al., 2009). Apart from that, it also has some other probiotic properties, including inhibiting various genital tract pathogenic bacteria (e.g., Gardnerella vaginalis) and fungi (e.g., Candida albicans) (Reid, 2017), increasing the antibiotic sensitivity, and reducing the risk of sexually transmitted diseases such as HIV (Tuddenham and Ghanem, 2017). These properties of L. crispatus can be attributed to the production of bactericidal substances (e.g., lactic acid and hydrogen peroxide) (Gong et al., 2014), ecological niche occupation (Srinivasan et al., 2009; Sgibnev and Kremleva, 2017), and immunomodulatory effects (Reid, 2017). Additionally, L. crispatus can prevent the colonization of pathogens via adhering to host cells (Younes et al., 2018), and the strong adherent effect of L. crispatus is considered to be more beneficial to the human body (Rizzo et al., 2015), but few studies focus on the possible negative effects caused by this characteristic.
As we have known, infertility is the condition of being unable to produce offspring and is defined by the World Health Organization as “a disease of the reproductive system defined by the failure to achieve a clinical pregnancy after 12 months or more of regular unprotected sexual intercourse” (Zegers-Hochschild et al., 2017). The common causes of infertility include genetic, immunological, and anatomical reasons (Mandar et al., 2017). Notably, the microbiological factors account for about 15% of overall couple infertility, and Escherichia coli has been implicated as one of the most common ones (Monteiro et al., 2018; Dutta et al., 2020). As early as in 1931, Rosenthal L demonstrated the agglutination effect of E. coli on sperm; subsequent studies also proved its immobilization effect. Both of the effects can remarkably reduce the vitality of sperm and cause infertility (Answal and Prabha, 2018). As one of the dominant species of the vaginal microbiota, the influence and the mechanism of L. crispatus on fertility are still unclear.
Our previous work indicated that some of the Lactobacillus spp. studied (L. crispatus, Lactobacillus acidophilus, Lactobacillus salivarius, Lactobacillus helveticus, and L. gasseri) could effectively adhere to sperm and greatly reduce the sperm motility. Of these, L. crispatus could greatly reduce the ability of sperm to penetrate the viscous medium in vitro, which suggest that this resident may influence the reproductive outcomes via some ways (Wang et al., 2020). Hence, the effect of L. crispatus as the vaginal probiotic product should be reconsidered, especially for some couples whose men are in a sub-fertile state.
In the present study, we verified whether L. crispatus had the potential to inhibit sperm motility in vitro, and we also evaluated the ability of L. crispatus in regulating the vaginal microbiota and affecting the reproductive results as a new vaginal agent both in maternal and offspring rat compared with other pathogenic bacteria. This study serves as a helpful warning and a theoretical basis for women preparing for a pregnancy which requires L. crispatus as a vaginal probiotic agent. Simultaneously, it is significant to further study and evaluate the probiotic activity of L. crispatus while avoiding the reduction of sperm motility.
Materials and Methods
Semen Sample Collection and Treatment
Semen samples of male subjects were collected from March 2019 to June 2019 after signing the informed consent (age = 22–38 years, without leukocytospermia), which was approved by the Institutional Ethics Committee on human subjects of The Second Affiliated Hospital of Nanchang University (ChiCTR2000036420). Donors who had been proven to be fertile and had normal sperm parameters according to the WHO criteria (Cao et al., 2011) were recruited. The subjects with abnormal karyotype and genital injuries were excluded. The subjects masturbated and ejaculated the semen into a 15-ml sterile collection tube. The semen samples were examined under a microscope and incubated in 37°C water bath for 25–45 min until they were liquefied, and then the purified supernatant sperms were harvested by direct swim-up in human tubal fluid (HTF) medium (Millipore) for subsequent in vitro experiments (Alasmari et al., 2013). The semen samples obtained on the same day were used immediately, and the sampling process continued until the end of the in vitro experiment.
Bacterial Strain Treatment
VNP20009 (ATCC 202165, an attenuated Salmonella typhimurium VNP20009), VNP20009 DNase I (an engineering bacteria, which was constructed by electrically transforming plasmid pLIVE-DNase I into attenuated S. typhimurium VNP20009, DNase I sequence number NM_005223.3, which was generated at Kingsy Biotechnology Co., Chen et al., 2019), E. coli O157:H7 (ATCC 12806), and L. crispatus (L. crispatus Lcr-MH175, number CGMCC 15938, identified by Harbin Meihua Biotechnology Co., Ltd., Harbin, Heilongjiang, China) were stored at −80°C. These strains were cultivated in a proper medium [L. crispatus was cultivated in De Man, Rogosa, Sharpe Broth (MRS, Hopebio, HB0384-1, Qingdao); VNP20009 and E. coli O157:H7 were cultivated in Lysogeny Broth (LB, Hopebio, HB0384-1, Qingdao), and VNP20009 DNase I was cultivated in LB medium containing 50 μg/ml ampicillin] for 24 h in an incubator at 37°C and activated twice; the bacterial growth density of each strain was approximately 109 CFU/ml. Taking 50 μl, 108 sperms/ml purified sperm samples was mixed with HTF medium containing different bacterial strains in equal volumes, and the number ratio of sperm to bacteria was 1:10. Subsequently, the mixed samples were incubated in an incubator at 37°C and 5% CO2 for different times. The experiment was carried out according to the following procedures.
Adhesion Assays
After incubation for 2, 4, and 6 h, the mixture of the sperm samples and bacterial strains was washed with sterile HTF solution four times. Briefly, the mixed samples were centrifuged at 400 × g for 5 min; the supernatant was removed and resuspended with 100 μl sterile HTF solution. A mixture of 10 μl was placed on a slide to dry, then fixed with methanol for 30 min, stained using the Gram staining procedure, and examined under a ×40 objective on an optical microscope (Olympus BX63). The number of adhesion bacteria in 100 sperms was measured from 25 microscopic fields which were randomly selected. Each adhesion test was repeated three times.
Assessment of Sperm Motility
A computer-assisted sperm analysis system (WLJY-9000, WeiLi Co., Ltd., China) was used to detect and analyze sperm motility after incubation with the tested bacteria for 2, 4, and 6 h. The sperm parameters related to total motility, progressive motility, linear velocity, and immotile sperm were measured. At least 200 sperms were examined in each trial. Each sperm motility assay was performed three times.
Sperm Penetration Assay
According to the published experimental method of Ivic et al. (2002), a 1% (w/v) methylcellulose solution was used to simulate the viscous environment of the female reproductive tract. Methylcellulose was prepared using HTF medium and inhaled into a flat capillary tube with a length of 7.5 cm and an inner diameter of 1.0 mm, one end of which was sealed with plasticine. Human sperms and tested bacteria were incubated in an incubator at 37°C, 5% CO2, for 2 h, and the open end of the capillary tube was inserted into the incubation mixture. After 1 h, the tubes were removed, and the penetration of methylcellulose was recorded using a Leica DM2500 upright microscope. The average number of sperms in the three sperm fields (10×) was counted at 1 and 2 cm from the base of the tube, respectively.
Measurement of Intracellular Ca2+ Concentrations in Sperm Samples
The Ca2+ signaling in human sperm was examined by sperm [Ca2+]i imaging (Nash et al., 2010). After the human sperms were loaded with 5 μM Fluo-4 AM (Molecular Probes, United States) and 0.05% pluronic F-127 (Molecular Probes, United States) at room temperature in the dark for 30 min, these were then washed in HTF medium. The washed sperms were loaded on a 96-well cell culture dish (Nest Biotechnology Co., Ltd.) and quickly added with the tested bacteria, respectively. A Polychrome V chromator (TILL Photonics GmbH, Germany) was used to image the sperm [Ca2+]i signaling. The generated 488-nm excitation light and the emissions (515–565 nm) were bandpass-filtered (HQ540/50, Chroma), which were collected with a cooled CCD camera (CoolSNAP HQ, Roper Scientific). The mixture was recorded for 200 s (50 ms of exposure time and 2 s of time interval).
Animal Set
Sexually mature and healthy 10- to 11-week-old male (n = 18, 313.3 ± 21.3 g) and 9- to 10-week-old female Sprague–Dawley (SD) rats (n = 72, 213.3 ± 21.3 g) were purchased from Nanchang Royo Biotech Co., Ltd. The animals were habituated to the animal facility for 2 weeks before the experiments under a 12-h light/dark cycle, with temperature of 21 ± 1°C and humidity of 55 ± 10%. Food and water were given ad libitum.
Reproductive Studies
In the first experimental set, the animals were randomly divided into five groups, i.e., control group (C group, n = 8), VNP20009 group (V group, n = 8), VNP20009 DNase I group (VD group, n = 8), E. coli O157:H7 group (E group, n = 8), and L. crispatus group (L group, n = 8). For the C group, the rats were held in a supine position during the application, and then the medical absorbable gelatine sponges (0.5 cm × 0.5 cm) which were soaked in 50 μl of phosphate-buffered saline (PBS) were delivered into the vaginal cavities of the rats once a day; the entire administration lasted 25 days. Similar to the C group, the V group, VD group, E group, and L group were treated with the bacterial strains of VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus, respectively (the concentration of each bacterial strain was about 5 × 104 CFU/ml). All females were immediately cohabited with males in a ratio of 4:1 after the sixth administration. When mating was confirmed by the presence of a vaginal plug or a pregnancy outcome, the females were separated from the males. On the 1st day, the administration of bacteria was stopped for 1 day, and the vaginal secretions in all groups were collected from donor dams with 200 μl sterile saline, and the entire contents of the pipette were expelled into the tube, which was conserved at −80°C for further use. On the 21st day, all the females were separated from the males and killed; their vaginas were collected for subsequent experiments.
Offspring Health Studies
In this experiment, the animals were randomly divided into three groups, i.e., control group (C group, n = 8), wide-spectrum antibiotic group (ABX group, n = 12), and L. crispatus group (L group, n = 12). For the C group, medical absorbable gelatine sponges (0.5 cm × 0.5 cm) were soaked in 50 μl PBS, and then delivered into the vaginal cavities each day as previously described. Similar to the C group, the L group was treated with the bacterial strain of L. crispatus whose concentration was about 5 × 104 CFU/ml, and the ABX group was treated with a wide-spectrum antibiotic [50-μl mixture of ampicillin (100 mg/ml), metronidazole (100 mg/ml), neomycin (100 mg/ml), and vancomycin (50 mg/ml)]. All females were immediately cohabited with males in the ratio of 4:1 after the sixth administration. On the 21st day, all the females were separated from the males, and they waited for parturition. When the pups were all delivered, six pups at postnatal day (P) 7 were randomly selected from each group for their colon and vaginal samples to be collected. The body weight of the remaining pups was measured from P 7 to P 42. After 2 weeks, the pups underwent the open-field test and the pole test. Finally, all the pups were sacrificed after 6 weeks; their tissues were collected and stored at −80°C for further use.
Fetal Health Assessment
The determination of live, dead, absorbed, and malformed fetus observed the following criteria: a live fetus showed a fleshy red color, fully formed, with natural movement, with motor response to mechanical stimulation, with a red placenta, large, and with its heart beating. A dead fetus demonstrated a grayish-red or dark purple color, fully formed, with recognizable toes of the limbs, with a body weight of ≤0.8 g, with no natural movement or response to mechanical stimulation, with the placenta grayish-red in color, and slightly smaller. An absorbed fetus revealed a dark purple or pale white color, not fully formed, with visible limb buds but no toes, with autolysis or softening, with no natural movement, with the placenta dark purplish red in color, and small. A malformed fetus evidenced the toes of the limbs being well formed, but the head, limbs, or tail was missing.
Behavioral Assessment
The pole test and the open-field test were performed to evaluate the development of the offspring. The pole test was used to determine the ability of movement balance and body coordination (Meligy et al., 2019), and the open-field test was used to evaluate the capacity for exploratory activities (Vakhnin and Briukhin, 2014). In the pole test, the rats were placed facing upward at the top of a rough wooden pole. Each rat got training (3–5 min/day) in the pole-descending behavior for 3 days, and only those rats revealing a time <18 s in the pre-test trial were used to participate in the formal trials. In the open-field test, each rat was individually placed in a chamber of size 100 cm × 100 cm (16 peripheral and nine internal). Each rat was placed in the center of the area to acclimate for 10 min, and then its behavioral parameters were monitored for 10 min with the ethovision video tracking software (Noldus, Wageningen, Netherlands). At the end of each trial, the equipment was cleaned with 75% ethanol to minimize odor interference to the next rat.
High-Throughput Sequencing Analysis
The genomic DNA of vaginal secretions was extracted using a DNA extraction kit (Tiangen Biotech Co., Ltd., Beijing, Cat#DP302-02), and the V4 region of the 16S rRNA genes was amplified using the primer 515F/806R (Walters et al., 2016). An Illumina HiSeq 2000 platform was applied to sequence these PCR products (GenBank accession number: PRJNA637679) (Edgar, 2013). Sequence analysis was subsequently processed using the UPARSE software package (version 7.0.100). Then, the QIIME software (version 1.9.1) was used to analyze the α diversity [indexes of observed operational taxonomic units (OTUs), Chao1, Shannon, Simpson, ACE, and goods coverage], β diversity [principal component analysis, principal coordinate analysis, and non-metric multidimensional scaling (NMDS)], differently abundant taxa identifications (Metastats and LEfSe analysis), and cluster analysis, and partial least squares discriminant analysis was performed using SIMCA-P software (version 11.5).
Histopathological Analysis
After dissecting the maternal rats, the fresh uterine tissue was put into the prepared 4% paraformaldehyde for fixation. Dehydration and transparent treatment were carried out, paraffin embedding was carried out, and the tissue was sliced into 5-μm segments. Before staining, these were rehydrated by xylene and ethanol for 5–6 min and washed three times for final hematoxylin and eosin (H&E) staining. Rabbit anti-Fas Ligand (FasL) antibody [1:1,000, Cell Signaling Technology (CST), Cat#ab15285] was used for immunohistochemical staining. Then, an antigen repair was performed, and the subsequent antibody binding and color rendering were completed according to the antibody instructions.
QRT-PCR and ELISA
In order to evaluate the mRNA expression level of cytokines associated with inflammation in female rat uterine tissues, the total RNA extracted from uterine tissues was reverse-transcribed into cDNA using a commercial reverse transcription kit (Takara, RR047A) (Sawant et al., 2021). Then, qRT-PCR was performed using Takara TB Green® Premix Ex TaqTM (Tli RNaseH Plus) Kit (Takara, RR420A) to detect the expressions of tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), interleukin 1β (IL-1β), and the housekeeping gene glyceraldehyde 3 phosphate dehydrogenase (GAPDH). After normalization against GAPDH, the relative amount of target gene transcripts in each cDNA sample was measured (Wang and Liu, 2020). The relative expression levels of these cytokines were analyzed by the 2−ΔΔCt method. The primer sequences were as follows:
TNF-α: forward primer – 5′-GTGGAACTGGCAGAAGA GGCA-3′, reverse primer – 5′-AGAGGGAGGCCATTTGG GAAC-3′;
IL-6: forward primer – 5′-GAAATCGTGGAAATGAGA- 3′, reverse primer – 5′-GCTTAGGCATAACGCACT-3′;
IL-1β: forward primer – 5′-GTGTCTTTCCCGTGGAC CTTC-3′, reverse primer – 5′-TCATCTCGGAGCCTGTA GTGC-3′;
GAPDH: forward primer – 5′-CTCGTGGAGTCTACTGG TGT-3′, reverse primer 5′-GTCATCATACTTGGCAG GTT-3′.
The collection of serum samples of female rats was via retro-orbital hemorrhage. For the determination of inflammatory cytokine levels in serum samples, ELISA kits for TNF-α (Cloud Clone Crop, United States, Cat #SEA133Mu), IL-6 (Cloud Clone Crop, United States, Cat #SEA079Mu), and IL-1β (Cloud Clone Crop, United States, Cat #SEA563Mu) were used. The inflammatory factors in serum bound to the biotinylated anti-rat monoclonal antibody and the immune complex could be formed after adding the secondary antibody, which combined to streptavidin labeled with horseradish peroxides. The addition of tetramethylbenzidine can make it colored, and the expression of the concentration of inflammatory factors in serum could be calculated according to its OD value.
Western Blotting Analysis
Total proteins were extracted from lysed tissues using a cell lysis buffer (Solarbio, R0010) containing protease inhibitors (MedChemExpress, HY-K0010). After the protein samples were denatured in boiling water, sodium dodecyl-sulfate polyacrylamide gel electrophoresis was completed using the electrophoresis apparatus. At this time, the total proteins in the samples were separated and arranged in the gel, and then the separated proteins were transferred to the polyvinylidene fluoride membrane by wet transfer. Subsequently, the membranes were sealed in 5% milk to block the non-specific site, and this was incubated overnight with the corresponding primary antibody at 4°C and then incubated with the secondary antibody conjugated with horseradish peroxidase (Chen et al., 2019). Finally, the membranes were observed and photographed using a chemiluminescence detection system (Tanon-5200, 20T12NPFLI6-10111). The primary antibodies including anti-β-actin (1:1,000, rabbit, CST, Cat#4970S), anti-transforming growth factor β1 (TGF-β1) (1:1,000, rabbit, CST, Cat#3711), anti-Bcl-2-associated X protein (Bax) (1:1,000, rabbit, CST, Cat#5023), anti-B cell lymphoma-2 (Bcl-2; 1:1,000, rabbit, CST, Cat#3498), anti-toll-like receptor 4 (TLR-4; 1:1,000, mouse, Santa Cruz, Cat#sc-293072), anti-phosphorylated nuclear factor kappa-B p65 (p-p65) (1:1,000, rabbit, Abcam, Cat#ab86299), anti-p65 (1:1,000, rabbit, CST, Cat#8242S), anti-zonula occludens-1 (ZO-1; 1:1,000, rabbit, CST, Cat#5406S), and anti-Occludin (1:1,000, rabbit, CST, Cat#91131).
Statistical Analysis
All values were expressed as mean ± standard deviation (SD). The differences of the experimental data between the control and samples were assessed with one- or two-way analysis of variance (ANOVA). Data handling, analyses, and graphical representations were performed using the GraphPad Prism software, version 7.0 (San Diego, CA, United States; RRID: SCR_004812). A P-value < 0.05 was regarded as statistically significant.
Results
Effect of L. crispatus on Human Sperm Motility Compared With Other Strains
To evaluate whether the probiotic L. crispatus had a better health effect compared to potential pathogens, we used E. coli O157:H7, a pathogenic bacterium capable of impairing sperm viability, as a positive control. In addition, we engineered the S. typhimurium VNP20009 DNase I strain, which has plasmid-containing DNase I to express DNase I toxin to harm the sperm activity, and S. typhimurium VNP20009 was used to serve as a control for S. typhimurium VNP20009 DNase I strain. Therefore, the three types of pathogenic bacteria, VNP20009, VNP20009 DNase I, and E. coli O157:H7, were compared with the probiotic L. crispatus to investigate their effects on sperm activity.
After conducting adherence assays, we found that all tested bacteria could adhere to sperm, with adhesive indices of approximately 65, 83, 206, and 521 per 100 sperms for VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus, respectively (Figure 1A). When analyzing the sperm motility, our results indicated that the adhered bacteria significantly decreased all parameters, reducing the total sperm motility (<40%), progressive motility (<30%), and linear velocity (<40 μm/s). In fact, an increase in the immotile sperm was also observed (>60%) when compared with the control at 4 and 6 h (Figures 1B–E). It is important to mention that L. crispatus could efficiently adhere to sperm and reduce the sperm motility in vitro.
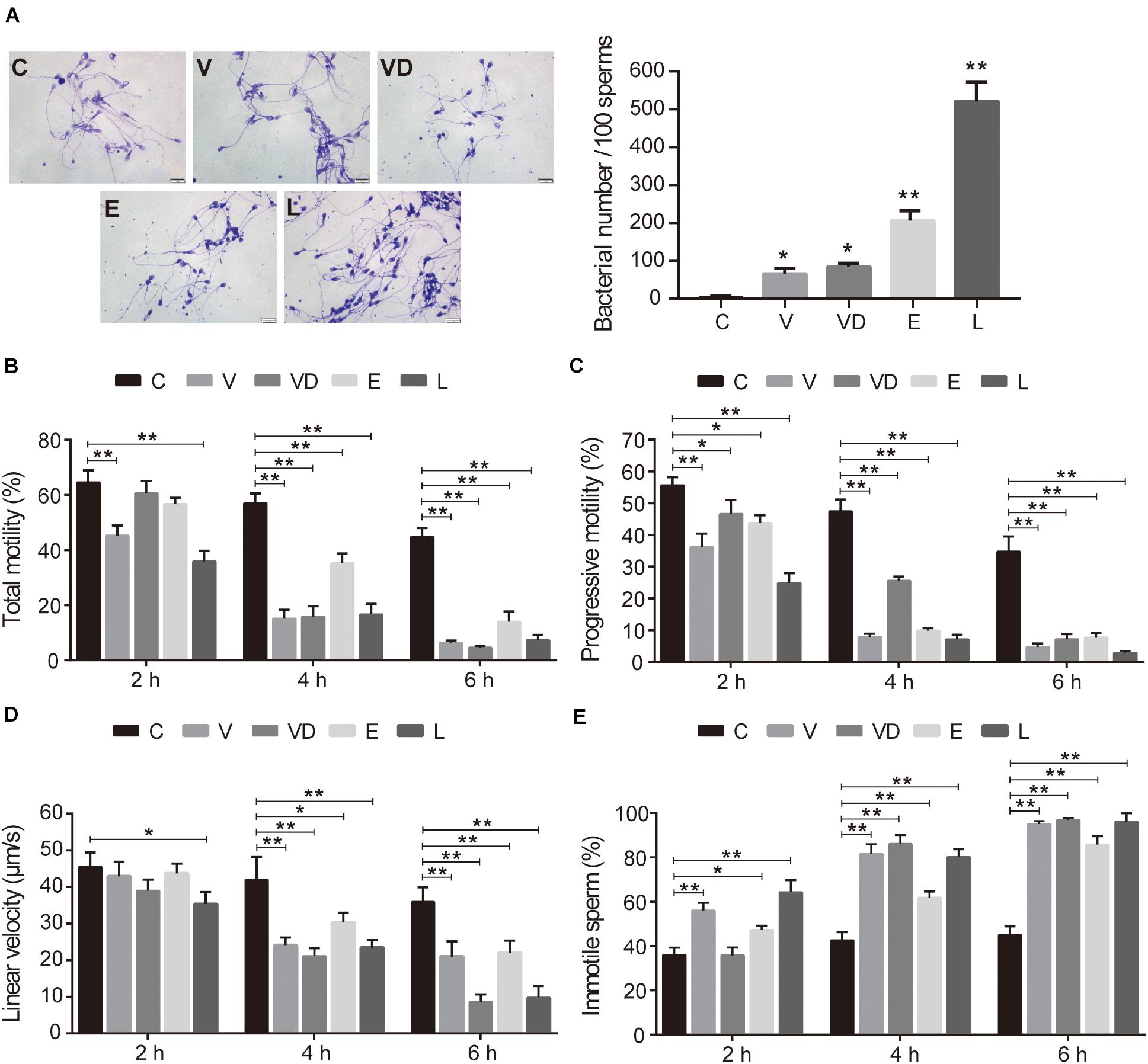
Figure 1. The effect of tested bacteria on sperm motility and the adhesion to sperm in vitro. (A) The gram stain results (magnification, ×100) and adhesion numbers of PBS, VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus. The initial sperm/bacteria ratio was 1:10, and the bacteria were available in-house. Human ejaculated sperm were incubated with a single species of bacteria in HTF medium in an incubator at 37°C and 5% CO2 for 2, 4, and 6 h, respectively. The motion parameters including (B) total motility, (C) progressive motility, (D) linear velocity, and (E) immotile sperm were analyzed by computer-assisted sperm analysis. For exactly assessing the sperm viability, a minimum of 200 sperms were counted for each assay. The data represent means ± SD of three independent experiments. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; V, VNP20009; VD, VNP20009 DNase I; E, E. coli O157:H7; L, L. crispatus.
Reduction of Intracellular Ca2+ Concentration in vitro
As previously reported, [Ca2+]i can regulate the chemotactic behavior of sperm and is pivotal to its function (Publicover et al., 2008). Thus, we further studied the effect of adhered bacteria on [Ca2+]i. Our results indicated that the adhered bacteria caused a reduction of human sperm [Ca2+]i within 1 min, and L group had the lowest sperm [Ca2+]i compared with the other groups at 2 min (Figures 2A,B). As shown in Figures 2C,D, the mean sperm numbers of penetrating 1% methylcellulose medium at 1 and 2 cm away from base of the tube for the controls were 489 and 166, respectively. VNP20009 DNase I and E. coli O157:H7 reduced 50 and 80% of the sperm numbers compared with the control group at 2 cm, while L. crispatus reduced the sperm numbers by approximately 90%.
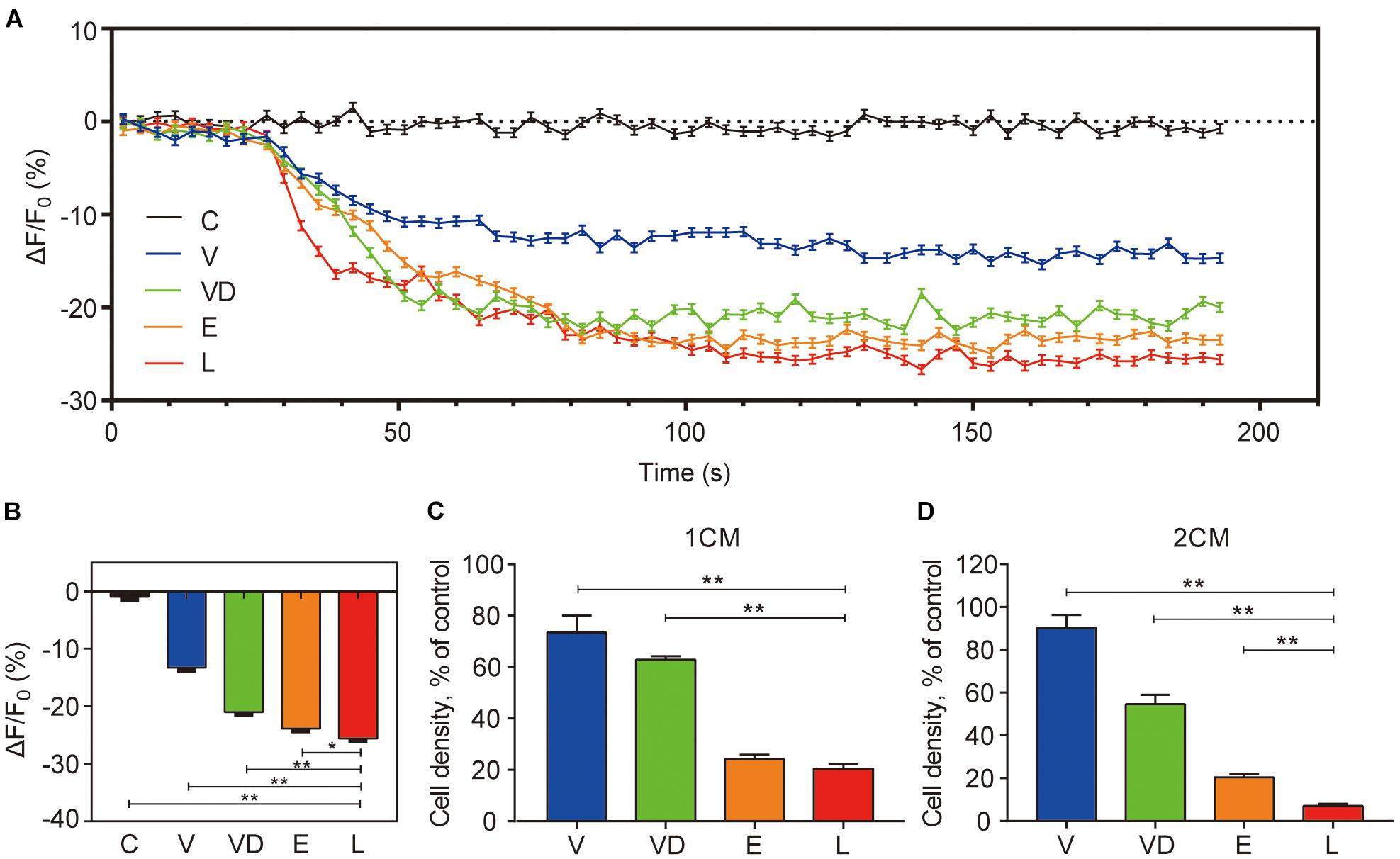
Figure 2. The effect of tested bacteria on the sperm of the intracellular Ca2+ concentration ([Ca2+]i) and the ability of penetration into viscous medium. (A) Sperm [Ca2+]i was monitored after loading sperms with 5 μM Fluo-4 AM, and the fluorescence intensity of the sperm head was visualized and detected before and after adding the different species of bacteria. (B) The statistical analysis of the effects of different species of bacteria on sperm [Ca2+]i at the time frame of 120 s as indicated in (A). Human sperms were treated with phosphate-buffered saline as control and were exposed to different species of bacteria to assess the penetrating ability of sperms into a viscous medium. Cell density (percentage of control) of (C) 1 cm and (D) 2 cm into the methylcellulose are shown. The initial sperm/bacteria ratio was 1:10. The data represent means ± SD of three independent experiments. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; V, VNP20009; VD, VNP20009 DNase I; E, E. coli O157:H7; L, L. crispatus.
Effect of L. crispatus on Reproduction by Inhibiting Sperm Motility
A rat reproductive model was developed to study the potential adverse effect of tested bacteria on the conception of females. PBS, VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus were transplanted into rat vaginal tracts, respectively (Figure 3A). After 25 days of continuous administration, our results showed that all the adhered bacteria resulted in a decrease of conception, and the pregnancy rates of the V, VD, E, and L group were 62.5, 50, 75, and 62.5%, respectively (Figure 3B). Comparing with the pregnant rats of the C group, the number of embryos in the V, VD, and E groups had a significant reduction, and there were also those that caused dead, absorbed, and malformed fetus, but the L group had no effect on the embryo health (Figure 3C and Supplementary Figure 1). The uterine morphology of unpregnant rats shown in the V, VD, and E groups were markedly swollen with a serious hyperemia, a bulbous expansion at the end of the uterine cavity, a thinner uterine wall containing a clear liquid in the lumen, and an increased uterine organ coefficient, but the morphology and the weight of the L group uterine were normal (Figures 3D,E), showing that VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus exerted the inhibition of conception, whereas L. crispatus had no toxicity to the maternal uterine and offspring.
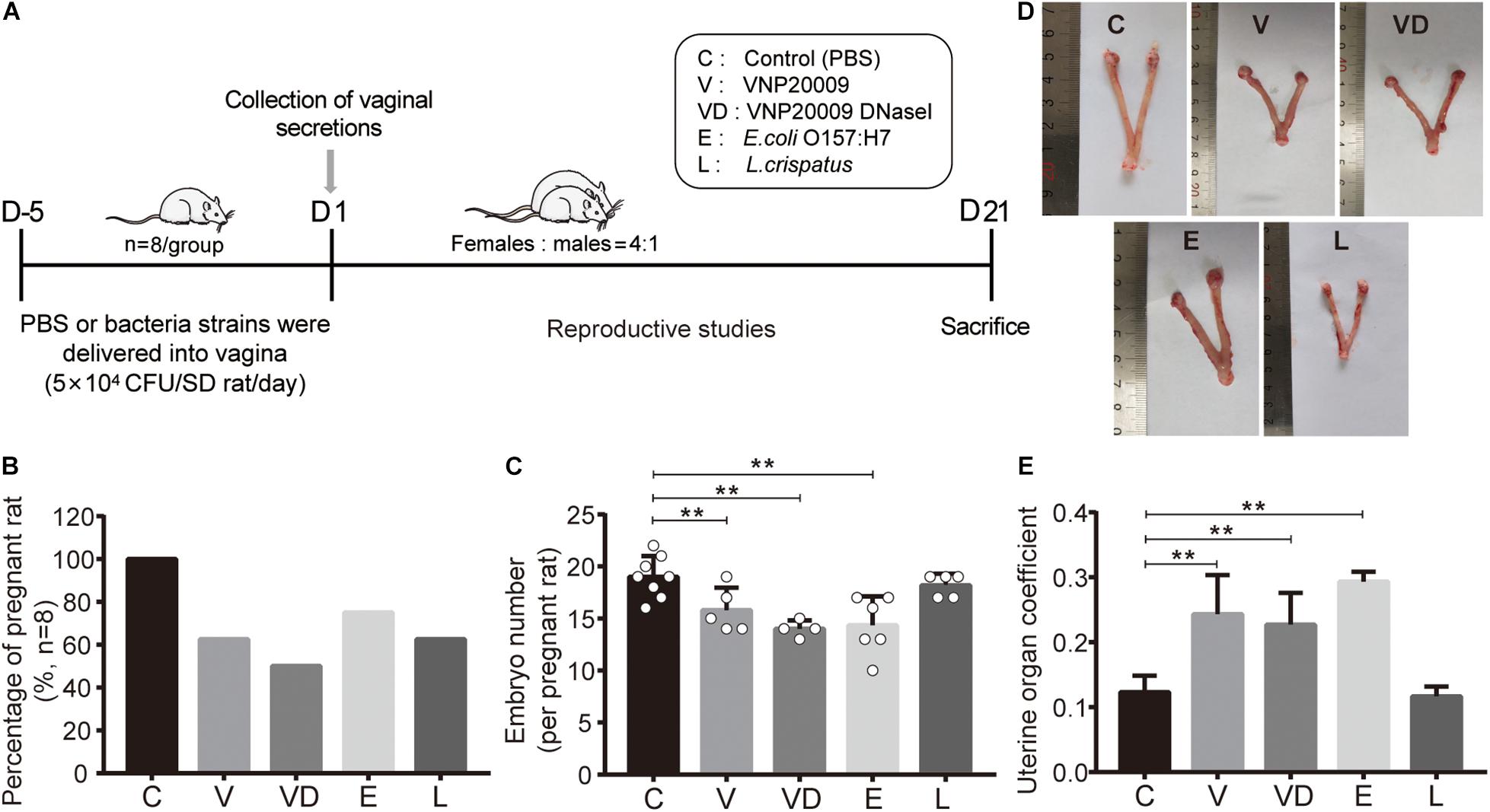
Figure 3. The inhibitory effect of L. crispatus on sperm reduced the pregnancy rates and had no reproductive impairment. (A) Scheme of experimental timeline and procedures. (B) Fertility outcome about pregnancy rates and (C) embryo number about the female rats of the indicated treatment groups. (D) Representative gross images and (E) organ coefficients of the uterus are shown in different treatment groups. The data represent means ± SD of eight to 10 animals. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; V, VNP20009; VD, VNP20009 DNase I; E, E. coli O157:H7; L, L. crispatus.
Effect of Tested Bacteria on Uterine Tissue Proliferation and Inflammation
To further explore the mechanisms of adhered bacteria on conception, H&E staining was used, and it was observed that the uterine wall showed a clear demarcation of the different layers. Intact endometrial epithelial cells were observed in the C group and the L group without hyperemia and inflammatory cell infiltration, while a damaged uterine wall and inflammatory infiltration were observed in the V, VD, and E groups (Figure 4A). Furthermore, the immunohistochemistry results showed that the V, VD, and E groups clearly induced the production of Fas Ligand (FasL) (Figure 4B). Then, western blotting analysis was used to detect key proteins associated with uterine proliferation and inflammation, and significant increases of transforming growth factor β1 (TGF-β1), Bax/Bcl-2, toll-like receptor-4 (TLR-4), and p-p65/p65 ratio in the uterus were observed in the V, VD, and E groups (Figures 4C–E). Moreover, the mRNA expressions of tumor necrosis factor-α (TNF-α) and cytokines interleukin-6 (IL-6) and cytokines interleukin-1β (IL-1β) in the uterus were greatly downregulated compared with the C group, respectively (Figures 4F,G). For L. crispatus, it sustained the normal expressions of cytokines related to uterine tissue proliferation and inflammation compared with other pathogens.
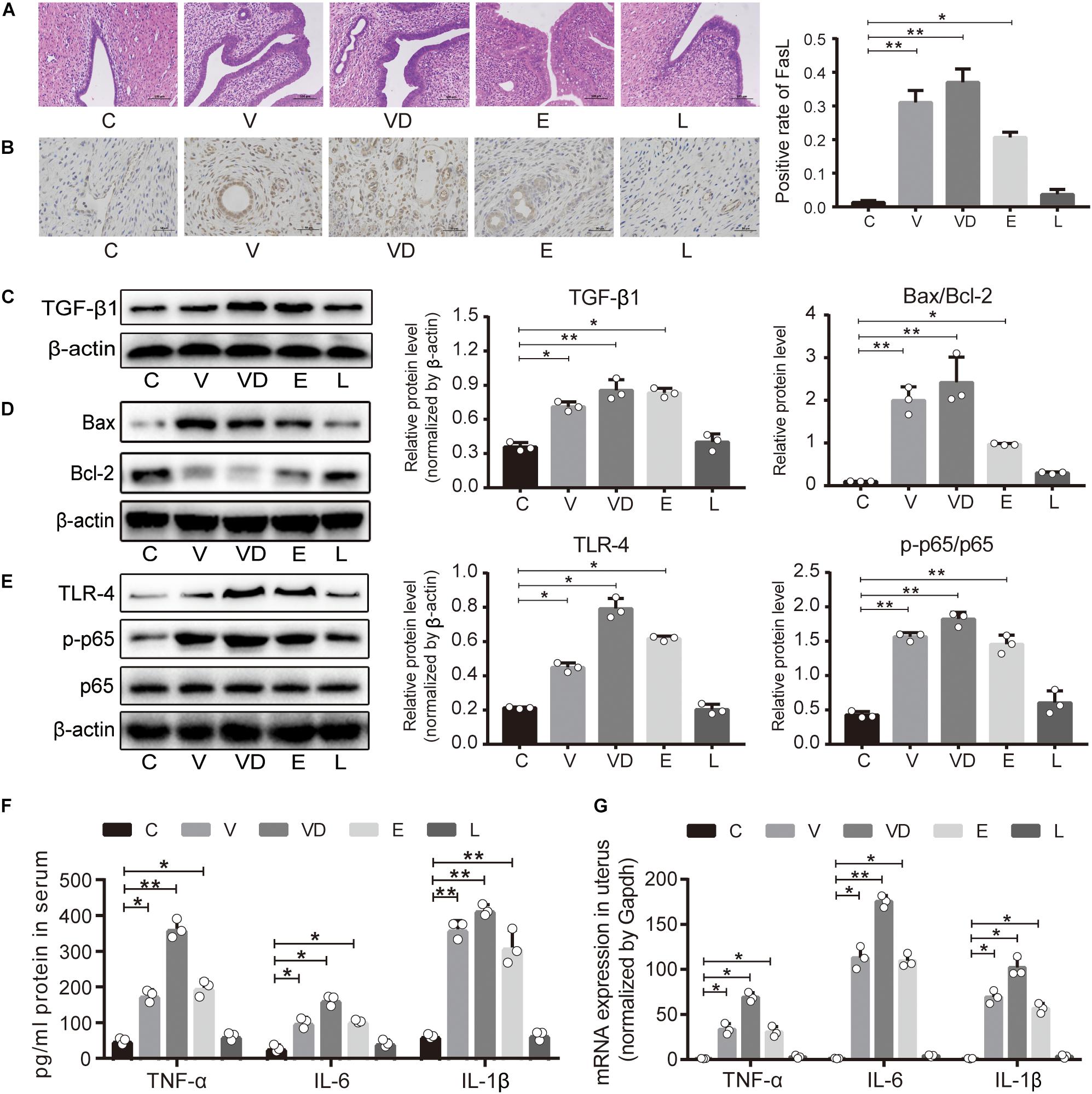
Figure 4. Treated vagina with L. crispatus had no influence on uterine impairment and vaginal inflammation. (A) Uterine histopathological examination was performed by H&E staining with different treatments; the bar represents 100 μm. (B) The immunohistochemistry images (left) of FasL expression were determined in the uterus from the indicated treatment groups; the graph on the right shows a positive rate counted in at least five different areas in the images; the bar represents 50 μm. The relative expression of (C) TGF-β1, (D) Bax/Bcl-2, and (E) TLR-4/NFκB was analyzed by western blotting in the vagina. (F) ELISA-based quantification of TNF-α, IL-6, and IL-1β levels in serum from the indicated treatment groups. (G) The mRNA expressions of TNF-α, IL-6, and IL-1β were determined by qPCR in the uterus. The data represent means ± SD of three independent experiments. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; V, VNP20009; VD, VNP20009 DNase I; E, E. coli O157:H7; L, L. crispatus.
Effect of Tested Bacteria on the Composition of the Vaginal Microbiota
To determine the effect of bacterial transplantation on vaginal microbial composition, we performed 16S rRNA gene sequencing of DNA isolated from vaginal secretion samples from the C, V, VD, E, and L groups. As shown in Figures 5A,B, the alpha-diversity of microorganism (Shannon and Simpson) was higher in the C and L groups compared with the V, VD, and E groups. The NMDS analysis (beta diversity) indicated that the samples in the C and L groups were clustered together, while the samples in the V, VD, and E groups were diverged from the C group (Figure 5C). Besides this, the Venn diagram showed that 761 common OTUs were identified in all groups, among which 48 OTUs were identified in the C and L groups, 17 OTUs were identified in the C and V groups, 28 OTUs were identified in the C and VD groups, and 13 OTUs were identified in the C and E groups (Figure 5D). At different taxonomic levels, we found that the number of microbial groups contained in the V, VD, and E groups were reduced compared with the C and L groups (Supplementary Figure 2).
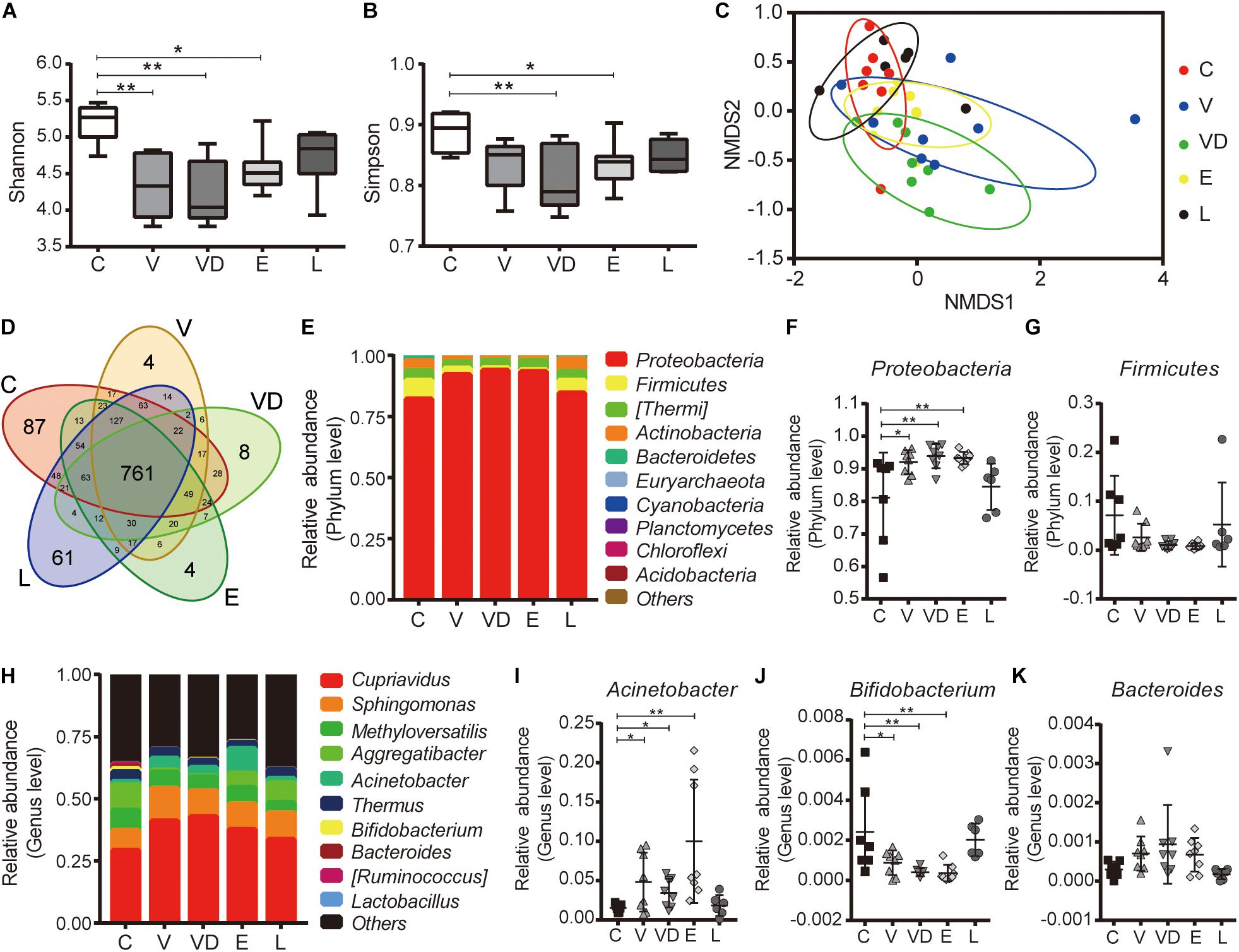
Figure 5. The effect of the tested bacteria on the vaginal microbiota diversity of rats. (A) Chao1 index and (B) Shannon index among the indicated treatment groups. (C) NMDS analysis of the indicated treatment groups. (D) Venn diagram of the vaginal microbiota among the indicated treatment groups. (E) Microbial composition at the phylum level. Relative abundance of (F) Proteobacteria and (G) Firmicutes of vaginal bacterial communities in phylum level among the indicated treatment groups. (H) Microbial composition at the genus level. Relative abundance of (I) Acinetobacter, (J) Bifidobacterium, and (K) Bacteroides of vaginal bacterial communities at the genus level among the indicated treatment groups. The data represent means ± SD of six to eight animals. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; V, VNP20009; VD, VNP20009 DNase I; E, E. coli O157:H7; L, L. crispatus; NMDS, non-metric multidimensional scaling.
Furthermore, the microbial community structure was also changed at the phylum and genus levels (Figures 5E,H). At the phylum level, a higher abundance of Proteobacteria was observed in the V, VD, and E groups (C vs. V, P = 0.019; C vs. VD, P = 0.0055; C vs. E, P = 0.0081). The abundance of Firmicutes did not show significant differences (C vs. V, P = 0.3297; C vs. VD, P = 0.1024; C vs. E, P = 0.0867), but we could also find a decrease in the V, VD, and E groups (Figures 5E–G). We next examined microbiota at the genus level. The abundance of Bifidobacterium was reduced in the V, VD, and E groups (C vs. V, P = 0.0275; C vs. VD, P = 0.0029; C vs. E, P = 0.0024), corresponding to the change of Firmicutes at the phylum level, and a higher abundance of Acinetobacter (C vs. V, P = 0.0349; C vs. VD, P = 0.0278; C vs. E, P = 0.0018) and Bacteroides (C vs. V, P = 0.5224 C vs. VD, P = 0.1303; C vs. E, P = 0.6012) was observed in the V, VD, and E groups, also corresponding to the change of Proteobacteria at the phylum level, although without statistical significance in Bacteroides genus level (P > 0.05) (Figures 5I–K). In addition, the LEfSe and Metastats analysis indicated that some taxa abundances were also significantly changed in the V, VD, and E groups compared with the C and L groups, for example, the abundance of Lactobacillaceae was significantly increased in the L group (Supplementary Figures 3A,B).
Effect of Vaginal Transplantation of L. crispatus on Offspring Health
To further assess whether L. crispatus mediated the spermicidal effect of bacteria on the offspring, we compared the offspring whose maternal vaginal tracts were transplanted with PBS, broad-spectrum antibiotics (to exclude the influence of vaginal microbiota on sperm), and L. crispatus. As presented in Supplementary Figure 4, ABX greatly reduced the overall microbial abundance including the observed species (C vs. ABX, P = 0.0010; ABX vs. L, P = 0.0340), Shannon (C vs. ABX, P = 0.0001; ABX vs. L, P = 0.0436; C vs. L, P = 0.0082), and Simpson (C vs. ABX, P = 0.0024) indices, increased the relative abundance of Firmicutes and Actinobacteria at the phylum level, increased the relative abundance of Mannheimia at the genus level, and made a significant division of the ABX group from the C group.
Then, we compared the fertility outcomes in the C, ABX, and L groups. As shown in Figures 6A–C, the use of broad-spectrum antibiotics and L. crispatus both reduced the conception rates of rats (ABX vs. L, 7 vs. 3), and L. crispatus had more pup number after conception compared with the ABX group (ABX vs. L, 10 vs. 17, P = 0.0016), and no obvious difference was observed between C and L groups.
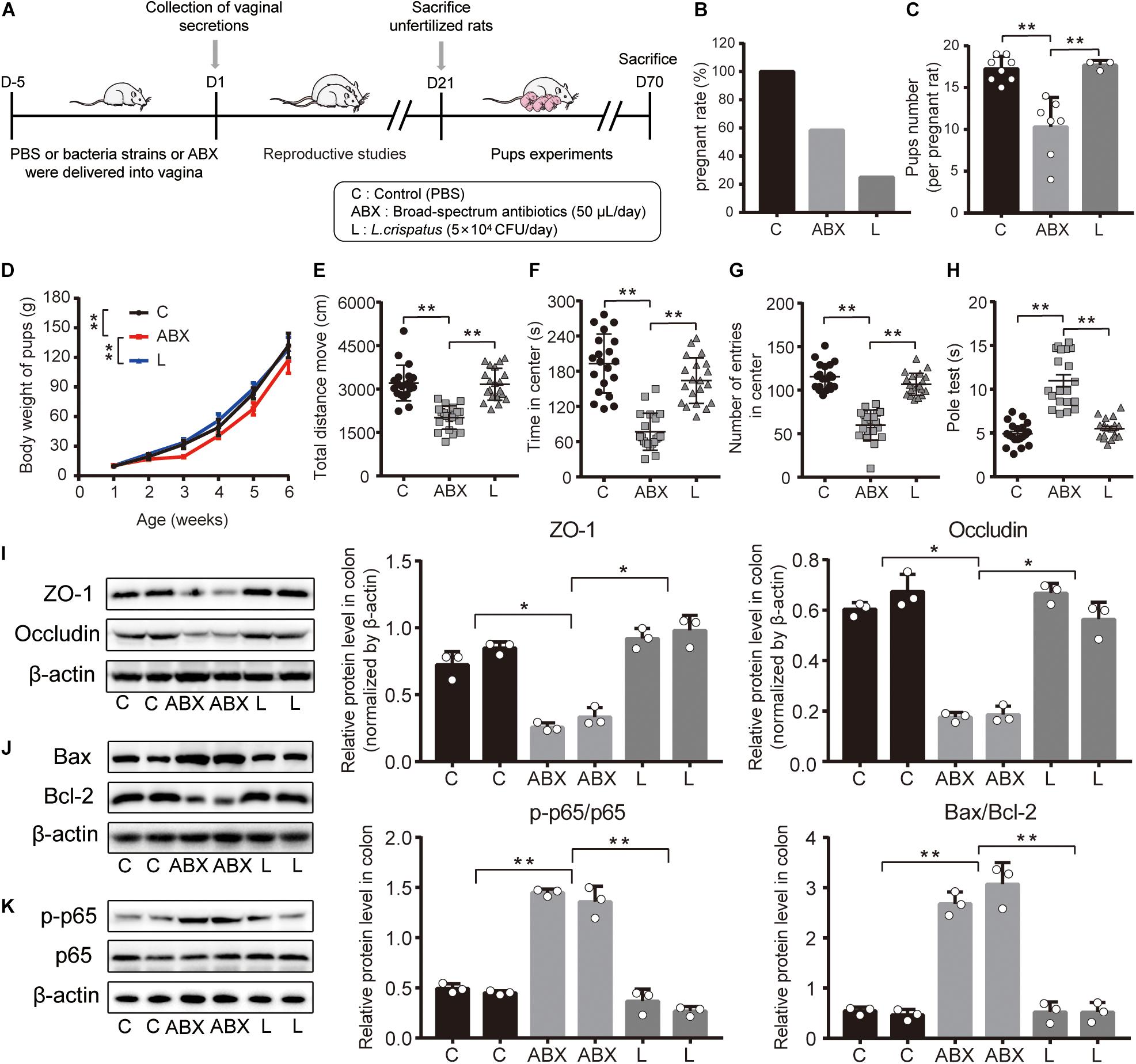
Figure 6. Treated vagina with L. crispatus had no reproductive and developmental toxicity on maternal and offspring rats. (A) Scheme of experimental timeline and procedures. (B) Fertility outcomes about pregnancy rates and (C) number of pups from the female rats of the indicated treatment groups. (D) Comparison of the body weight of pups among C, ABX, and L groups from P 7 to P 42. (E) Total moving distance in the open-field test. (F) The time of pups in center total moving distance in the open-field test. (G) The number of entries in the center in the open-field test. (H) The time of pups climbing pole in the pole test. (I) Expression level of intestinal tight junction proteins, ZO-1, and Occludin. (J) Protein expression level of apoptosis-associated factors, Bcl-2 and Bax. (K) Protein expression level of p-p65 and p65. The data represent means ± SD of eight to 20 animals or three independent experiments. *P < 0.05, **P < 0.01. SD, standard deviation; C, control; ABX, wide-spectrum antibiotic; L, L. crispatus.
Then, we studied the effects of ABX and L. crispatus on offspring health and development, and our results indicated that the use of ABX greatly reduced the body weight growth from postnatal day (P) 7 to P 42 (P 42, ABX vs. L, 117.5 vs. 128.59 g) (Figure 6D). Additionally, offspring anxiety-like behavior, motor coordination, and cognitive deficits were also evaluated by open-field test and polo test, and the results showed that the pups in groups C and L moved the longer mean moving distance (C vs. ABX vs. L, 3,206.699 cm vs. 2,017.157 cm vs. 3,167.3345 cm), stayed a longer time in the center (C vs. ABX vs. L, 193.2325 s vs. 76.905 s vs. 164.08 s), significantly increased the number of entering to the center (C vs. ABX vs. L, 116 vs. 60 vs. 107), and had the lowest latency time (C vs. ABX vs. L, 4.9175 s vs. 10.9955 s vs. 5.5145 s) compared with the ABX group, respectively (Figures 6E–H). We subsequently examined the tight junction proteins, apoptosis factors, and inflammatory-related factors in the colon of pups, and we found that the ABX group exhibited lower expression levels of ZO-1 and Occludin (Figure 6I) and higher expression levels of Bax/Bcl-2 and p-p65/p65 (Figures 6J,K) compared with the C group. For the L group, it seemed that the use of L. crispatus would not affect offspring development, while excluded vaginal microbiota using broad-spectrum antibiotics poses a negative effect on offspring development.
Discussion
A healthy vaginal microbiome is dominated by species of Lactobacillus that have a protective effect and may have therapeutic potential (Codagnone et al., 2019). As with the gut microbiome, disruption of the vaginal microbiome may affect the immunity of the body or lead to an increase in pathogens, potentially leading to the occurrence of diseases (Brown et al., 2019). Although some studies have demonstrated the relationship between bacteria and fertilization (Fredricks et al., 2005; Srinivasan et al., 2012), the underlying mechanisms involved in it is unclear. Our work indicated that L. crispatus not only has the sperm impairment effect in vitro but also may influence the maternal reproductive ability in vivo.
Motility is the key factor of sperm function which can be used to predict the fertilization potential of sperm in vitro, while the maintenance of sperm function is mainly dependent on [Ca2+]i (Agnihotri et al., 2016). Our study found that L. crispatus greatly reduced the total sperm motility, progressive motility, linear velocity, immotile sperm, [Ca2+]i, and the ability of sperm to penetrate the viscous medium, which give us an insight that L. crispatus has the potential to affect fertilization (Figures 1, 2). It was also implying that the inhibitory effect of adhered bacteria on sperm motility and their ability to penetrate the methylcellulose medium may result from the reduction of sperm [Ca2+]i in vitro. Nevertheless, whether L. crispatus has the same action in vivo is not clear. Therefore, the in vivo sperm inhibitory efficacy of bacteria was assessed using the rat model.
Fertility outcome experiments showed that female rats treated with VNP20009, VNP20009 DNase I, E. coli O157:H7, and L. crispatus had a reduction of conception rates, respectively. However, L. crispatus did not affect fetal development and survival. Other bacteria reduced the fetal survival rate after pregnancy and led to a dead, malformed, or absorbed fetus (Supplementary Figure 1). The V, VD, and E groups also showed a serious inflammation reaction and tissue damage. Moreover, the production of apoptosis factors (FasL) (Krzyzowska et al., 2011) was obviously detected in uterus slices, which indicated that these bacteria had reproductive toxicity, but L. crispatus protected the uterine health of rats (Figure 3).
Recently, apoptosis has been implicated in regulating various reproductive tissues, including the uterus, ovary, placenta, and fetal membrane (Mu et al., 2002; Mitchell et al., 2015). In addition to FasL, we found that the expressions of Bax/Bcl-2 in the L group were also significantly ameliorated compared with the other experimental groups and mainly occurred at the protein level. When we explored the body immune status, we found that the balance is disturbed, such that the TNF-α, IL-6, and IL-1β cytokines in the serum and the uterine tissues were higher in the V, VD, and E groups, which were coincident with the levels of inflammation found in uterus slices. As TNF-α, IL-6, and IL-1β are the three prominent pro-inflammatory cytokines that can activate NFκB transcription, the TLR/NFκB pathway was further studied (Joo et al., 2012). As expected, the expression of key proteins in the TLR/NFκB pathway including TLR-4 and p-p65/p65 was also significantly increased in the pathogen group, while no obvious changes were observed between the L group and the C group. Then, we also measured the expression of TGF-β1, a protein whose disordered expression in the maternal uterus might be associated with embryo implantation failure (Rajaei et al., 2011; Guo et al., 2018). We found that the uterus in the L group had normal TGF-β1 level that was consistent with the C group but which in the V, VD, and E groups were aberrant (Figure 4). The abnormal expression of TGF-β1 leads to an imbalance between apoptosis and proliferation of uterine tissues, resulting in the occurrence of abortion and other adverse pregnancy outcomes (Latifi et al., 2019). These data together suggested that the interventions of L. crispatus kept the uterine healthy and avoided the apoptosis and inflammation of the uterus compared with other pathogens.
Communities of microbiota have been proved to be associated with numerous health outcomes (Younes et al., 2018). We integrated vaginal microbiome datasets using the 16S rRNA gene high-throughput sequencing to reveal the functional impact of microbiome on genital health with the rat model. Regarding the vaginal microbiota at the phylum level, we observed a lower relative abundance of Proteobacteria and a higher relative abundance of Firmicutes in the C and L groups. Moreover, our results showed that VNP20009, VNP20009 DNase I, and E. coli O157:H7 could cause vaginal dysbiosis by downregulating Bifidobacterium and upregulating Acinetobacter (opportunistic pathogen) and Bacteroides at the genus level. These specific changes may contribute to the development of inflammatory-related diseases and are associated with the pathogenesis of bacterial vaginosis (Anstey Watkins et al., 2019; Figure 5).
In this study, we found that the vaginal microbiota, reproductive health, and inflammation were closely correlated. The abundance of beneficial bacteria (Firmicutes and Bifidobacterium) was negatively associated with pro-inflammatory indicators to some extent. Reversely, Proteobacteria, Acinetobacter, and Bacteroides were positively associated with pro-inflammatory indicators. VNP20009, VNP20009 DNase I, and E. coli O157:H7 did not colonize the rat vagina; they all showed a serious destruction to the maternal vaginal microbiota, leading to an imbalance in the immune environment. Although L. crispatus showed a potential to inhibit sperm motility, it maintained the normal vaginal microbiota homeostasis and did not adversely affect the reproductive health of rats.
Recent reports have demonstrated that the pioneer community of maternal vaginal microbiota that colonizes the offspring gastrointestinal tract at birth has a lasting impact on the gut and brain (Peterson and Artis, 2014; Sakwinska et al., 2017; Jasarevic et al., 2018). Our results demonstrated that broad-spectrum antibiotics greatly reduced the maternal vaginal microbiota diversity and overall abundance (Supplementary Figure 4). In addition, the rats transplanted with broad-spectrum antibiotics exhibited an intermediate inhibitory effect of conception and pup number, and their offspring also had a lower body weight curve, anxiety-like behavior, uncoordinated movement, and cognitive impairment, indicating that the use of antibiotics not only greatly reduced the vaginal microbiota but also posed a serious negative effect on offspring health. The experiments on the intestinal health of the offspring have also shown that the colonic tight junction (TJ) proteins (Feng et al., 2019) – ZO-1 and Occludin – were reduced in the ABX group, and the expression of inflammation factors (p-p65/p65) and apoptosis factors (Bax/Bcl-2) in the colon tissue was also increased in the ABX group, which suggested that the intestinal health of offspring from the ABX group was at risk (Figure 6). In summary, L. crispatus could sustain the maternal vaginal composition and had no effect on the healthy development during the early life of the offspring as the normal female rats.
In obstetrics and gynecology, Lactobacillus-based probiotics have been used to restore the physiologic vaginal microbiota to alter the maternal and neonatal microbiome (Czaja et al., 2007; Srikrishna and Cardozo, 2013), thus improving pregnancy and neonatal outcomes (Stone, 2018). Our study found that L. crispatus protected vaginal health and offspring development, which provided the foundation for vaginal microbiome transplantation, and also gave a theoretical basis for the development and application of L. crispatus as a vaginal probiotic product in the future. However, the limitations of L. crispatus in inhibiting sperm motility also provide challenges, including achieving a stable colonization of the desired probiotic species, avoiding over-transplanting of probiotic species in the face of intrinsic and extrinsic vaginal disturbances, as well as determining the ideal species or strains for different women individuals.
Conclusion
In the present study, we evaluated the adhesion ability of L. crispatus Lcr-MH175 to sperm. All bacteria could adhere to sperms effectively and reduce their motility, which were particularly obvious for the probiotic L. crispatus. This property of L. crispatus reduced the pregnancy rate in a rat reproduction model but did not affect maternal health or offspring development. We postulate that the reduction in motility caused by the adhesion of L. crispatus may be beneficial for healthy couples and offspring health as it could reduce the chance of a poor-quality sperm combining with the egg and provide a unique sperm selection mechanism. Additionally, the strong adherent effect of L. crispatus may be detrimental for men with severe asthenospermia, oligospermia, or aspermia as L. crispatus could lower the potential of the sperm combining with the egg. Therefore, although L. crispatus is a probiotic species, more caution should be paid when using it as a vaginal viable preparation for women of child-bearing age, such as the dosage. Taken together, this study focuses exclusively on the role of L. crispatus on sperm activity and the development of offspring, so more work is needed to explore the mechanistic insights of L. crispatus and other Lactobacillus spp. on sperm functions.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics Statement
The animal study was reviewed and approved by the Laboratory Animal Ethics Committee of Nanchang Royo Biotech Co., Ltd. (RYE2019040801).
Author Contributions
PL contributed to the methodology, investigation, formal analysis, visualization, and writing – original draft. KW contributed to the methodology, investigation, and visualization. XH contributed to the methodology. LZ and JW contributed to the formal analysis. ZL and XC contributed to the investigation. HW contributed to the resources and conceptualization. TC contributed to the conceptualization, funding acquisition, supervision, and writing – review and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the National Natural Science Foundation of China (Grant No. 82060638), Academic and Technical Leaders of Major Disciplines in Jiangxi Province (Grant No. 20194BCJ22032), and Double Thousand Plan of Jiangxi Province (High-End Talents Project of Scientific and Technological Innovation).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2021.705690/full#supplementary-material
References
Agnihotri, S. K., Agrawal, A. K., Hakim, B. A., Vishwakarma, A. L., Narender, T., Sachan, R., et al. (2016). Mitochondrial membrane potential (MMP) regulates sperm motility. Vitro Cell Dev. Biol. Anim. 52, 953–960. doi: 10.1007/s11626-016-0061-x
Alasmari, W., Costello, S., Correia, J., Oxenham, S. K., Morris, J., Fernandes, L., et al. (2013). Ca2+ signals generated by CatSper and Ca2+ stores regulate different behaviors in human sperm. J. Biol. Chem. 288, 6248–6258. doi: 10.1074/jbc.M112.439356
Anstey Watkins, J., Ross, J. D. C., Thandi, S., Brittain, C., Kai, J., and Griffiths, F. (2019). Acceptability of and treatment preferences for recurrent bacterial vaginosis-topical lactic acid gel or oral metronidazole antibiotic: qualitative findings from the VITA trial. PLoS One 14:e0224964. doi: 10.1371/journal.pone.0224964
Answal, M., and Prabha, V. (2018). Escherichia coli recombinant sperm immobilizing factor RecX as a potential vaginal contraceptive. Reprod. Biol. Endocrinol. 16:88. doi: 10.1186/s12958-018-0407-1
Belkaid, Y., and Hand, T. W. (2014). Role of the microbiota in immunity and inflammation. Cell 157, 121–141. doi: 10.1016/j.cell.2014.03.011
Brown, R. G., Al-Memar, M., Marchesi, J. R., Lee, Y. S., Smith, A., Chan, D., et al. (2019). Establishment of vaginal microbiota composition in early pregnancy and its association with subsequent preterm prelabor rupture of the fetal membranes. Transl. Res. 207, 30–43. doi: 10.1016/j.trsl.2018.12.005
Cao, X. W., Lin, K., Li, C. Y., and Yuan, C. W. (2011). [A review of WHO laboratory manual for the examination and processing of human semen (5th edition)]. Zhonghua Nan Ke Xue 17, 1059–1063.
Chen, C., Song, X., Wei, W., Zhong, H., Dai, J., Lan, Z., et al. (2017). The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 8:875. doi: 10.1038/s41467-017-00901-0
Chen, T., Zhao, X., Ren, Y., Wang, Y., Tang, X., Tian, P., et al. (2019). Triptolide modulates tumour-colonisation and anti-tumour effect of attenuated Salmonella encoding DNase I. Appl. Microbiol. Biotechnol. 103, 929–939. doi: 10.1007/s00253-018-9481-8
Codagnone, M. G., Spichak, S., O’Mahony, S. M., O’Leary, O. F., Clarke, G., Stanton, C., et al. (2019). Programming bugs: microbiota and the developmental origins of brain health and disease. Biol. Psychiatry 85, 150–163. doi: 10.1016/j.biopsych.2018.06.014
Cohen, J. (2016). INFECTIOUS DISEASE. Vaginal microbiome affects HIV risk. Science 353:331. doi: 10.1126/science.353.6297.331
Czaja, C. A., Stapleton, A. E., Yarova-Yarovaya, Y., and Stamm, W. E. (2007). Phase I trial of a Lactobacillus crispatus vaginal suppository for prevention of recurrent urinary tract infection in women. Infect. Dis. Obstet. Gynecol. 2007:35387. doi: 10.1155/2007/35387
Dutta, S., Sengupta, P., Izuka, E., Menuba, I., Jegasothy, R., and Nwagha, U. (2020). Staphylococcal infections and infertility: mechanisms and management. Mol. Cell Biochem. 474, 57–72. doi: 10.1007/s11010-020-03833-4
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Feng, Y., Huang, Y., Wang, Y., Wang, P., Song, H., and Wang, F. (2019). Antibiotics induced intestinal tight junction barrier dysfunction is associated with microbiota dysbiosis, activated NLRP3 inflammasome and autophagy. PLoS One 14:e0218384. doi: 10.1371/journal.pone.0218384
Fredricks, D. N., Fiedler, T. L., and Marrazzo, J. M. (2005). Molecular identification of bacteria associated with bacterial vaginosis. N. Engl. J. Med. 353, 1899–1911. doi: 10.1056/NEJMoa043802
Gong, Z., Luna, Y., Yu, P., and Fan, H. (2014). Lactobacilli inactivate Chlamydia trachomatis through lactic acid but not H2O2. PLoS One 9:e107758. doi: 10.1371/journal.pone.0107758
Guo, F., Si, C., Zhou, M., Wang, J., Zhang, D., Leung, P. C. K., et al. (2018). Decreased PECAM1-mediated TGF-beta1 expression in the mid-secretory endometrium in women with recurrent implantation failure. Hum. Reprod. 33, 832–843. doi: 10.1093/humrep/dey022
Halmos, T., and Suba, I. (2016). [Physiological patterns of intestinal microbiota. The role of dysbacteriosis in obesity, insulin resistance, diabetes and metabolic syndrome]. Orv. Hetil. 157, 13–22. doi: 10.1556/650.2015.30296
Ivic, A., Onyeaka, H., Girling, A., Brewis, I. A., Ola, B., Hammadieh, N., et al. (2002). Critical evaluation of methylcellulose as an alternative medium in sperm migration tests. Hum. Reprod. 17, 143–149. doi: 10.1093/humrep/17.1.143
Jasarevic, E., Howard, C. D., Morrison, K., Misic, A., Weinkopff, T., Scott, P., et al. (2018). The maternal vaginal microbiome partially mediates the effects of prenatal stress on offspring gut and hypothalamus. Nat. Neurosci. 21, 1061–1071. doi: 10.1038/s41593-018-0182-5
Joo, H. M., Kim, K. A., Myoung, K. S., Ahn, Y. T., Lee, J. H., Huh, C. S., et al. (2012). Lactobacillus helveticus HY7801 ameliorates vulvovaginal candidiasis in mice by inhibiting fungal growth and NF-kappaB activation. Int. Immunopharmacol. 14, 39–46. doi: 10.1016/j.intimp.2012.05.023
Krzyzowska, M., Shestakov, A., Eriksson, K., and Chiodi, F. (2011). Role of Fas/FasL in regulation of inflammation in vaginal tissue during HSV-2 infection. Cell Death Dis. 2:e132. doi: 10.1038/cddis.2011.14
Latifi, Z., Nejabati, H. R., Abroon, S., Mihanfar, A., Farzadi, L., Hakimi, P., et al. (2019). Dual role of TGF-beta in early pregnancy: clues from tumor progression. Biol. Reprod. 100, 1417–1430. doi: 10.1093/biolre/ioz024
Ling, Z., Kong, J., Liu, F., Zhu, H., Chen, X., Wang, Y., et al. (2010). Molecular analysis of the diversity of vaginal microbiota associated with bacterial vaginosis. BMC Genomics 11:488. doi: 10.1186/1471-2164-11-488
Mandar, R., Punab, M., Korrovits, P., Turk, S., Ausmees, K., Lapp, E., et al. (2017). Seminal microbiome in men with and without prostatitis. Int. J. Urol. 24, 211–216. doi: 10.1111/iju.13286
Meligy, F. Y., Elgamal, D. A., Abd Allah, E. S. H., Idriss, N. K., Ghandour, N. M., Bayoumy, E. M. R., et al. (2019). Testing alternatives: the use of adipose-derived mesenchymal stem cells to slow neurodegeneration in a rat model of Parkinson’s disease. Mol. Biol. Rep. 46, 5841–5858. doi: 10.1007/s11033-019-05018-9
Mitchell, M. D., Peiris, H. N., Kobayashi, M., Koh, Y. Q., Duncombe, G., Illanes, S. E., et al. (2015). Placental exosomes in normal and complicated pregnancy. Am. J. Obstet. Gynecol. 213, S173–S181. doi: 10.1016/j.ajog.2015.07.001
Monteiro, C., Marques, P. I., Cavadas, B., Damiao, I., Almeida, V., Barros, N., et al. (2018). Characterization of microbiota in male infertility cases uncovers differences in seminal hyperviscosity and oligoasthenoteratozoospermia possibly correlated with increased prevalence of infectious bacteria. Am. J. Reprod. Immunol. 79:e12838. doi: 10.1111/aji.12838
Mu, J., Kanzaki, T., Si, X., Tomimatsu, T., Fukuda, H., Fujii, E., et al. (2002). Apoptosis and related proteins during parturition in prostaglandin F receptor-deficient mice. Biochem. Biophys. Res. Commun. 292, 675–681. doi: 10.1006/bbrc.2002.6698
Nash, K., Lefievre, L., Peralta-Arias, R., Morris, J., Morales-Garcia, A., Connolly, T., et al. (2010). Techniques for imaging Ca2+ signaling in human sperm. J Vis Exp., 40:e1996. doi: 10.3791/1996
Pellati, D., Mylonakis, I., Bertoloni, G., Fiore, C., Andrisani, A., Ambrosini, G., et al. (2008). Genital tract infections and infertility. Eur. J. Obstet. Gynecol. Reprod. Biol. 140, 3–11. doi: 10.1016/j.ejogrb.2008.03.009
Peterson, L. W., and Artis, D. (2014). Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 14, 141–153. doi: 10.1038/nri3608
Petrova, M. I., Lievens, E., Malik, S., Imholz, N., and Lebeer, S. (2015). Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 6:81. doi: 10.3389/fphys.2015.00081
Publicover, S. J., Giojalas, L. C., Teves, M. E., de Oliveira, G. S., Garcia, A. A., Barratt, C. L., et al. (2008). Ca2+ signalling in the control of motility and guidance in mammalian sperm. Front. Biosci. 13:5623–5637. doi: 10.2741/3105
Rajaei, S., Zarnani, A. H., Jeddi-Tehrani, M., Tavakoli, M., Mohammadzadeh, A., Dabbagh, A., et al. (2011). Cytokine profile in the endometrium of normal fertile and women with repeated implantation failure. Iran. J. Immunol. 8, 201–208.
Reid, G. (2017). The development of probiotics for women’s health. Can. J. Microbiol. 63, 269–277. doi: 10.1139/cjm-2016-0733
Rizzo, A., Fiorentino, M., Buommino, E., Donnarumma, G., Losacco, A., and Bevilacqua, N. (2015). Lactobacillus crispatus mediates anti-inflammatory cytokine interleukin-10 induction in response to Chlamydia trachomatis infection in vitro. Int. J. Med. Microbiol. 305, 815–827. doi: 10.1016/j.ijmm.2015.07.005
Sakwinska, O., Foata, F., Berger, B., Brussow, H., Combremont, S., Mercenier, A., et al. (2017). Does the maternal vaginal microbiota play a role in seeding the microbiota of neonatal gut and nose? Benef. Microbes 8, 763–778. doi: 10.3920/BM2017.0064
Sawant, S., Dugad, J., Parikh, D., and Singh, H. (2021). Absolute quantitation of oral bacteria involved in oral cancer by real-time PCR. Med. Microecol. 7:100034. doi: 10.1016/j.medmic.2021.100034
Sgibnev, A., and Kremleva, E. (2017). Influence of hydrogen peroxide, lactic acid, and surfactants from vaginal lactobacilli on the antibiotic sensitivity of opportunistic bacteria. Probiotics Antimicrob. Proteins 9, 131–141. doi: 10.1007/s12602-016-9238-6
Shipitsyna, E., Roos, A., Datcu, R., Hallen, A., Fredlund, H., Jensen, J. S., et al. (2013). Composition of the vaginal microbiota in women of reproductive age–sensitive and specific molecular diagnosis of bacterial vaginosis is possible? PLoS One 8:e60670. doi: 10.1371/journal.pone.0060670
Srikrishna, S., and Cardozo, L. (2013). The vagina as a route for drug delivery: a review. Int. Urogynecol. J. 24, 537–543. doi: 10.1007/s00192-012-2009-3
Srinivasan, S., Hoffman, N. G., Morgan, M. T., Matsen, F. A., Fiedler, T. L., Hall, R. W., et al. (2012). Bacterial communities in women with bacterial vaginosis: high resolution phylogenetic analyses reveal relationships of microbiota to clinical criteria. PLoS One 7:e37818. doi: 10.1371/journal.pone.0037818
Srinivasan, U., Misra, D., Marazita, M. L., and Foxman, B. (2009). Vaginal and oral microbes, host genotype and preterm birth. Med. Hypotheses 73, 963–975. doi: 10.1016/j.mehy.2009.06.017
Stone, L. (2018). Infection: vaginal microbiota and infectious infertility. Nat. Rev. Urol. 15:136. doi: 10.1038/nrurol.2018.11
Tuddenham, S., and Ghanem, K. G. (2017). A microbiome variable in the HIV-prevention equation. Science 356, 907–908. doi: 10.1126/science.aan6103
Vakhnin, V. A., and Briukhin, G. V. (2014). [Influence of environmental enrichment on parameters of behavior in open field test in the rats born from females with chronic alcoholization]. Ross. Fiziol. Zh. Im. I M Sechenova 100, 406–417.
Verstraelen, H., Verhelst, R., Claeys, G., De Backer, E., Temmerman, M., and Vaneechoutte, M. (2009). Longitudinal analysis of the vaginal microflora in pregnancy suggests that L. crispatus promotes the stability of the normal vaginal microflora and that L. gasseri and/or L. iners are more conducive to the occurrence of abnormal vaginal microflora. BMC Microbiol. 9:116. doi: 10.1186/1471-2180-9-116
Walters, W., Hyde, E. R., Berg-Lyons, D., Ackermann, G., Humphrey, G., Parada, A., et al. (2016). Improved bacterial 16S rRNA Gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 1:e00009-15. doi: 10.1128/mSystems.00009-15
Wang, H., Chen, T., Chen, Y., Luo, T., Tan, B., Chen, H., et al. (2020). Evaluation of the inhibitory effects of vaginal microorganisms on sperm motility in vitro. Exp. Ther. Med. 19, 535–544. doi: 10.3892/etm.2019.8237
Wang, X.-W., and Liu, Y.-Y. (2020). Comparative study of classifiers for human microbiome data. Med. Microecol. 4:100013. doi: 10.1016/j.medmic.2020.100013
Younes, J. A., Lievens, E., Hummelen, R., van der Westen, R., Reid, G., and Petrova, M. I. (2018). Women and their microbes: the unexpected friendship. Trends Microbiol. 26, 16–32. doi: 10.1016/j.tim.2017.07.008
Keywords: Lactobacillus crispatus, vaginal microbiome, sperm, animal model, reproductive health, offspring
Citation: Li P, Wei K, He X, Zhang L, Liu Z, Wei J, Chen X, Wei H and Chen T (2021) Vaginal Probiotic Lactobacillus crispatus Seems to Inhibit Sperm Activity and Subsequently Reduces Pregnancies in Rat. Front. Cell Dev. Biol. 9:705690. doi: 10.3389/fcell.2021.705690
Received: 06 May 2021; Accepted: 13 July 2021;
Published: 13 August 2021.
Edited by:
Bechan Sharma, University of Allahabad, IndiaReviewed by:
António Machado, Universidad San Francisco de Quito, EcuadorMargarita Aguilera, University of Granada, Spain
Copyright © 2021 Li, Wei, He, Zhang, Liu, Wei, Chen, Wei and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tingtao Chen, Y2hlbnRpbmd0YW8xOTg0QDE2My5jb20=; Hong Wei, d2VpaG9uZzYzNTI4QDE2My5jb20=; d2VpaDI2QG1haWwuc3lzdS5lZHUuY24=