 Jeferson Jantsch1†
Jeferson Jantsch1† Isadora D’Ávila Tassinari2†
Isadora D’Ávila Tassinari2† Márcia Giovenardi1,3
Márcia Giovenardi1,3 Victorio Bambini-Junior4,5
Victorio Bambini-Junior4,5 Renata Padilha Guedes1,3
Renata Padilha Guedes1,3 Luciano Stürmer de Fraga2*
Luciano Stürmer de Fraga2*- 1Programa de Pós-Graduação em Biociências, Universidade Federal de Ciências da Saúde de Porto Alegre (UFCSPA), Porto Alegre, Brazil
- 2Programa de Pós-Graduação em Ciências Biológicas: Fisiologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, Brazil
- 3Programa de Pós-Graduação em Ciências da Saúde, Universidade Federal de Ciências da Saúde de Porto Alegre (UFCSPA), Porto Alegre, Brazil
- 4School of Pharmacy and Biomedical Sciences, University of Central Lancashire (UCLan), Preston, United Kingdom
- 5Division of Biomedical and Life Sciences, Faculty of Health and Medicine, Lancaster University, Lancaster, United Kingdom
Since the first evidence suggesting that maternal nutrition can impact the development of diseases in the offspring, much has been elucidated about its effects on the offspring’s nervous system. Animal studies demonstrated that maternal obesity can predispose the offspring to greater chances of metabolic and neurodevelopmental diseases. However, the mechanisms underlying these responses are not well established. In recent years, the role of the gut-brain axis in the development of anxiety and depression in people with obesity has emerged. Studies investigating changes in the maternal microbiota during pregnancy and also in the offspring demonstrate that conditions such as maternal obesity can modulate the microbiota, leading to long-term outcomes in the offspring. Considering that maternal obesity has also been linked to the development of psychiatric conditions (anxiety and depression), the gut-brain axis is a promising target to be further explored in these neuropsychiatric contexts. In the present study, we review the relationship between maternal obesity and anxious and depressive features, exploring the gut-brain axis as a potential mechanism underlying this relationship.
Introduction
The Developmental Origins of Health and Disease (DOHaD) theory brought by Barker and Osmont (1986) states that prenatal environmental factors influence the offspring’s health in adulthood (Barker and Osmond, 1986). Both malnutrition and overnutrition are determinants of metabolic and neurological dysfunctions in the offspring, such as impairments in the brain reward circuitry, neuroinflammatory states, and behavioural disorders (Smith and Reyes, 2017).
Neurodevelopmental disorders are well documented in the offspring of obese mothers in humans and animal models (Edlow, 2017; Tong and Kalish, 2021). However, observational epidemiological studies have struggled to establish a direct cause-effect relationship between gestational obesity and neurological diseases in the progeny. Nevertheless, in preclinical studies, there is a substantial body of evidence showing a mechanistic link between maternal obesity and neurological dysfunctions in the offspring (Edlow, 2017). For example, long-term hippocampal insulin resistance in obese mice dams was followed by a decrease in the expression of neurogenesis markers and synaptic plasticity in the offspring (Schmitz et al., 2018). Moreover, the impact of maternal obesity is multigenerational, impairing synaptic function in the hippocampus until the third generation (Fusco et al., 2019).
Considering that obesity during pregnancy is increasing globally (Chen et al., 2018), as well as the prevalence of mood disorders such as depression and anxiety (WHO, 2017), it is important to unravel the mechanisms linking these conditions. Maternal obesity alters intestinal microbiota (Chu et al., 2016), which has emerged as a key regulator of metabolism and behaviour. Thus, understanding how the maternal intestinal microbiota influences fetal neurodevelopment is fundamental to comprehend the repercussions of maternal obesity on the mental health of the offspring (Kumpulainen et al., 2018).
This review will discuss the influence of maternal overnutrition on the development of depression and anxiety in the offspring. We will emphasize the role of the gut-brain axis in the mechanisms linking maternal obesity to the harmful neurodevelopment of the progeny.
Depression
Intrauterine overnutrition could affect the fetal programming, increasing the risk of the newborn developing neuropsychiatric conditions such as depression (McDonald et al., 2015; Cárdenas-Tueme et al., 2020; Cattane et al., 2021).
Offspring of rodent mothers fed with a high-fat diet (HFD) during pregnancy and lactation shows increased depressive-like behaviour in adolescence and adulthood (Gawlinska et al., 2019), even if the offspring was fed a standard diet. Although there is some controversy in the literature [for review see (Ortiz-Valladares et al., 2021)], it reinforces that prenatal exposure to maternal HFD is a decisive factor that can lead to depressive symptoms later in life (Gawlinska et al., 2019). Transcriptional changes in the prefrontal cortex (Gawlińska et al., 2021), reduction in the number and morphology of dendrites and spines in the hippocampus, amygdala, and somatosensory cortex (Hatanaka et al., 2017), lower volume of brain areas such as the thalamus, hippocampus, nucleus accumbens, and hypothalamus, in addition to changes in synaptophysin and GFAP (glial fibrillary acidic protein) expression (Trujillo-Villarreal et al., 2021) may be part of the mechanisms underlying the behavioural changes seen in the adult offspring.
Maternal exposure to HFD, or a carbohydrate-rich diet, also affects the dopaminergic (DA) system by changing the amount of dopamine and its metabolite DOPAC (3,4-dihydroxyphenylacetic acid) in the nucleus accumbens and the expression of dopamine receptors D1/D2 in the ventral tegmental area (Naef et al., 2008; Naef et al., 2011; Sullivan et al., 2015; Paradis et al., 2017). Dysregulation of these circuits increases the risk of mood disorders and altered function of the mesolimbic DA system, as well as the dopamine receptor in limbic structures, has already been observed in different models of depression (Winter et al., 2007; Tye et al., 2013; Belujon and Grace, 2017).
Clinical studies have reported that elevated levels of proinflammatory cytokines in obese pregnant women can lead to placental dysfunction and are related to neurodevelopmental and neuropsychiatric disorders in the offspring [for review see (Howell and Powell, 2017)]. Evidence from humans and animals also links depressive-like behaviour with immune system disturbances or neuroinflammatory responses. Proinflammatory mediators such as C-reactive protein, interleukin (IL)-6, IL-1β, and tumor necrosis factor (TNF)-α affect the development of neural circuits that are critical for regulation of behaviour, providing evidence that inflammation may also be a potential factor leading to increased risk of mood disorders (Rivera et al., 2015; Rasmussen et al., 2019).
Furthermore, emotional changes in the progeny were associated with maternal nutritional status (Ortiz-Valladares et al., 2021) and high maternal body mass index with a decline in psychological development in the early stages of childhood (Bergmann et al., 2016; Aubuchon-Endsley et al., 2017). In summary, poor maternal nutrition during pregnancy negatively impacts neurodevelopmental outcomes in the first years of life and plays an important role in inducing mental disorders in the offspring.
Anxiety
Maternal obesity has been associated with offspring’s anxiety independently of sex, species, and age (Menting et al., 2019). However, the mechanisms linking maternal obesity to the development of anxiety in the offspring remain a gap in the field. The available evidence will be briefly scrutinized here.
During adolescence, there is an increase in anxiety in the offspring of HFD-fed dams, shown in the light-dark transition task, but a decrease in anxiety in the open field and elevated plus-maze, which could be interpreted as an increased risky behaviour rather than decreased anxiety. Importantly, an increase in the expression of nuclear factor kappa B (NF-kB) and IL-6 genes, responsible for inflammatory responses, was found in the hippocampus of adolescent offspring of dams fed with a hypercaloric diet. But an increase in the expression of anti-inflammatory genes such as IkBα and mitogen-activated protein kinase MKP-1 was also seen, suggesting a disturbance in the expression of inflammatory mediators in the brain of these animals (Sasaki et al., 2014). On the other hand, when analyzing the offspring of HFD-fed dams during adult life, an increase in anxiety-like behaviours was observed in the open field, elevated plus-maze, and light-dark transition tasks. The maternal diet also changed the concentration of plasma corticosteroids after stress challenge and the expression of pro-inflammatory genes in the amygdala, such as NF-kB or IL-6, with a notable sex-specific influence (Sasaki et al., 2013). Although the effect of maternal diet on anxiety did not appear to be influenced by sex or age in the systematic review by Menting et al. (2019), it is clear that this claim is controversial and needs further investigation.
Most studies involving maternal consumption of obesogenic diets use the HFD model. However, the cafeteria diet (CAF) model provides animals with ultra-processed human food consumption and has translational potential (Matuszewska et al., 2021). Wright et al. showed that maternal consumption of a CAF impacts offspring behaviour, regardless of whether consumption occurs in the preconception period, during pregnancy, lactation, or all these periods (Wright et al., 2011). Interestingly, CAF during lactation caused a consistent anxiolytic effect in males as observed by the increase of entries into and more time spent on open arms in the elevated plus-maze and the reduced latency to enter the center in the open field test. It reinforces that the sex of the offspring is an important factor and should be taken into account when studying the effects of maternal obesity (Wright et al., 2011).
Otherwise, consuming CAF exclusively during pregnancy increased maternal behaviours such as licking pups, arched nursing, and nest building after 2- and 8-days following birth. Interestingly, even with increased maternal behaviour, the offspring demonstrated behavioural disturbances in adolescence such as increased anxiety-like manifestations during light-dark tests (Ribeiro et al., 2018).
Table 1 summarizes preclinical studies demonstrating maternal diet’s impact on depressive and anxiety-like behaviours in the progeny.
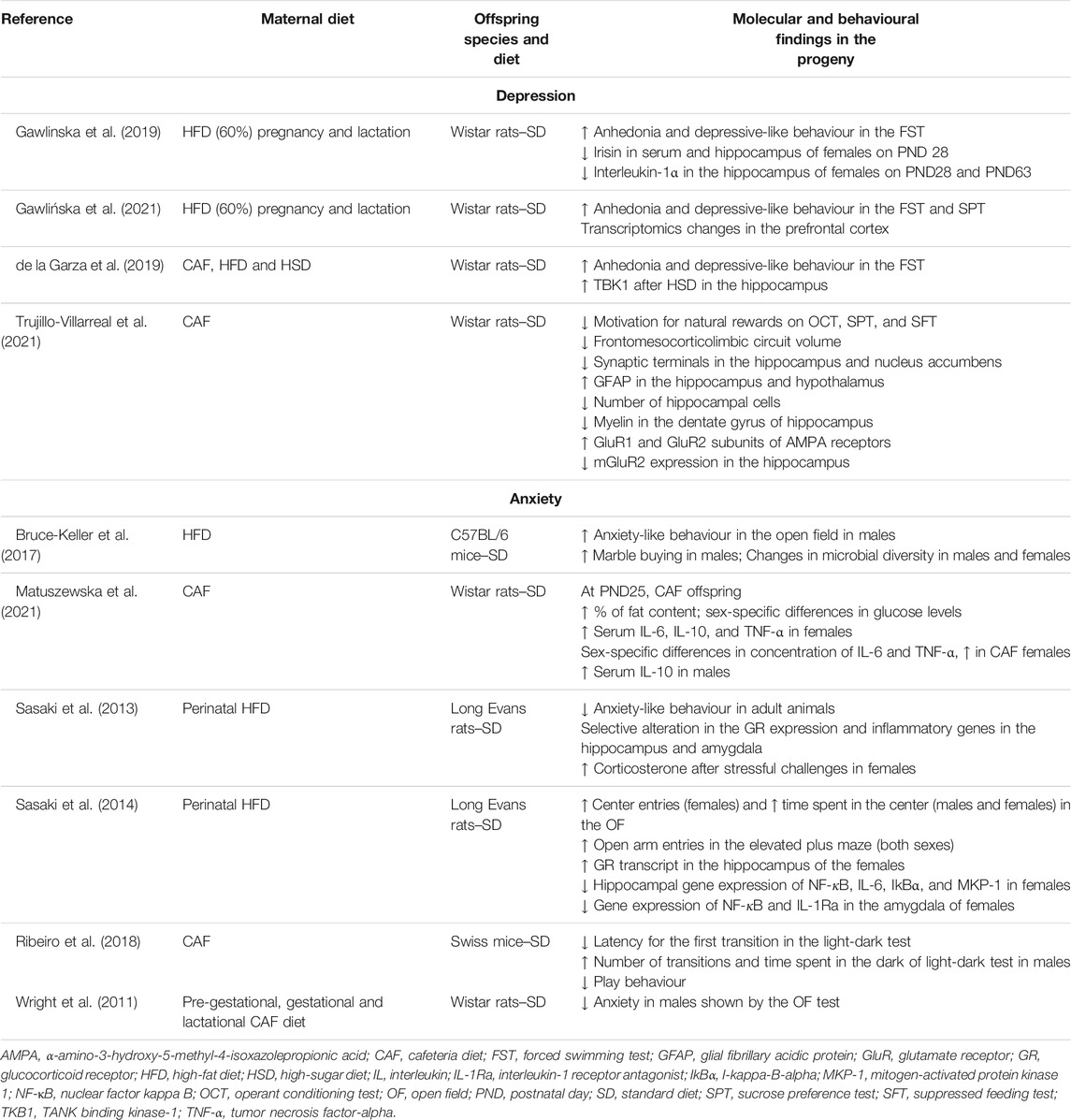
TABLE 1. Effects of maternal nutrition on depressive and anxious-like behaviour and reported mechanisms in preclinical studies.
The Gut-Brain Axis During Fetal Development
Data presented so far reinforce a causative relationship between maternal diet and neurologic programming in offspring. However, the underlying mechanism involved in this phenomenon remains elusive. Despite that, the influence of maternal microbiota on the development of anxious behaviours in the offspring has emerged as a mechanism that is sufficient to disrupt behavioural function in murine offspring in a sex-specific manner (Bruce-Keller et al., 2017). It suggests that intestinal dysbiosis could link the effects of unhealthy modern diets to the increased prevalence of neurodevelopmental and childhood disorders.
In addition to fermenting non-digestible fibers from the diet, which increases energy extraction from food, the influence of the gut microbiota on the host’s physiology has been gaining notoriety from studies that demonstrate a bi-directional relationship with the brain, a pathway coined as the gut-brain axis (Martin et al., 2018). Although this communication is not completely clear, three main pathways are proposed: neural signals carried out by the vagus nerve; chemical signals synthesized by the microbiota, as short-chain fatty acids (SCFA) or neurotransmitters; and immunomodulation exerted by microbiota and its products (Morais et al., 2021).
The main phyla of bacteria that colonize the human gut are Firmicutes and Bacteroidetes (Eckburg et al., 2005), but microbiota composition is extremely dynamic and altered even in physiological contexts such as pregnancy. For example, in the first trimester, the maternal gut microbiota is more similar to that of men and non-pregnant women, while Proteobacteria and Actinobacteria are expanded during the third trimester (Koren et al., 2012). Conversely, analysis from 1479 pregnant women found no differences compared to age-matched non-pregnant women, suggesting individual heterogeneity as the most relevant element for changes in gut microbiota during pregnancy (Yang et al., 2020). Even though, in animal studies, mice that received faecal transplantation from women in the third trimester, but not in the first, develop weight gain and insulin resistance (Koren et al., 2012). Also, an increase in Akkermansia, Clostridium, Bacteroides, and Bifidobacterium genera was found in C57BL/6 pregnant mice, supporting the idea of gut microbiome changes during pregnancy (Gohir et al., 2015). These changes during pregnancy do not influence only the mother, but also impact the offspring’s neurodevelopment (Al Rubaye et al., 2021), e.g., altering the offspring’s microglial gene signature: male offspring of germ-free dams showed changes in the expression of 1216 microglial genes near birth. In contrast, the major impact on female offspring was observed in adulthood with alterations in 433 genes expressed by microglia compared to the offspring of dams with intact gut microbiota (Thion et al., 2018). Therefore, the maternal gut microbiota can also influence the offspring’s brain.
Changes in the microbiota are not limited to physiological situations such as pregnancy. In metabolic diseases such as obesity, there is evidence demonstrating changes in the bacterial profile, such as an increase in Firmicutes and a decrease in Bacteroidetes compared to their lean counterparts both in obese humans (Crovesy et al., 2020) and in HFD-fed animals (Jo et al., 2021). Indeed, the interaction between obesity and pregnancy causes specific changes that impact both mother and offspring’s microbiota. Mice exposed to HFD before mating and during gestation had increased Akkermansia (phylum Firmicutes) and Bifidobacterium compared to lean pregnant animals (Gohir et al., 2015). Previous studies show that the type of birth delivery can profoundly impact the bacterial profile in children (Ferretti et al., 2018; García-Mantrana et al., 2020), even though bacterial colonization does not occur in the uterus (Perez-Muñoz et al., 2017). The cohort study carried out by Chu et al. (2016) showed differences in the intestinal microbiota of children of mothers who consumed HFD compared to balanced diets, which demonstrated that the maternal diet influences both the mother and the infant’s bacterial profile.
Advances in the field have been showing how gut microbiota could influence the development of mood disorders such as anxiety and depression (Peirce and Alviña, 2019; Huang and Wu, 2021). In a recent study, pregnant women had their microbiota sequenced through stool samples during the third trimester of pregnancy, and children’s behaviour was analyzed at 2 years of age. Interestingly, increased alpha diversity, a measure of microbiota diversity, was related to less internalizing behaviour at 2 years of age, which can be associated with decreased anxiety behaviour. Furthermore, the butyrate-producing families Lachnospiraceae and Ruminococcaceae were found in mothers from children with normal behaviour. Prenatal exposure to healthy diets was also linked to the development of healthy behaviour in children (Dawson et al., 2021). However, few studies explore this mechanism in the context of maternal overnutrition and offspring’s behaviour.
Animal models support that maternal consumption of unhealthy diets during pregnancy, such as HFD, is related to gut microbiota changes that can predispose behaviours related to anxiety and depression in the offspring (Buffington et al., 2016; Bruce-Keller et al., 2017). To isolate the effects of obesity-modified microbiota on offspring behaviour, gut microbiota was transplanted from HFD-fed animals to female mice with depleted microbiota before copulation. It resulted in altered offspring’s microbiota and primed anxiety-like behaviour in males, but not in females. Male offspring from HFD transplanted dams also showed a pattern of depressive-like behaviour. Therefore, maternal microbiota influenced by the diet can lead to depressive and anxiety-like behaviours in the offspring in a sex-dependent manner (Bruce-Keller et al., 2017). In another preclinical study, co-housing the offspring of HFD dams with offspring of a control dam reversed changes in the offspring’s microbiota caused by maternal obesogenic diet and alleviated the behavioural abnormalities. Moreover, faecal transplantation from offspring of HFD dams to germ-free mice led to behavioural effects similar to those observed in the offspring of obese dams (Buffington et al., 2016).
Among the proposed mechanisms, a decrease in fermenting bacteria responsible for SCFA production such as butyrate, propionate, and acetate can lead to a leaky gut, impairing the intestinal barrier and allowing immunogenic molecules from the gut, e.g., lipopolysaccharide (LPS) reach the circulation, where they activate inflammatory responses that can become chronic and lead to psychiatric manifestations (Dash et al., 2015; Winter et al., 2018). Yucatan pigs dams consuming CAF during pregnancy and lactation showed a decrease in SCFA in both dams’ and offspring’s faeces 100 days after birth. A decrease in neurogenesis markers in the offspring’s hippocampus was also found, evidencing the involvement of the gut-brain axis in this context (Val-Laillet et al., 2017). Also, altered microbiota during gestation can trigger maternal immune activation, which leads to conditions in the offspring that also have anxiety as a feature, e.g., autism spectrum disorder (Careaga et al., 2017; Gottfried and Bambini-Junior, 2018). Important to mention, gut microbiota changes impact the synthesis of neurotransmitters such as serotonin, dopamine, and norepinephrine [for review see (Huang and Wu, 2021)], which also might influence the development of mood disorders. Moreover, diet-induced maternal dysbiosis has also been proposed to increase hypothalamus-pituitary-adrenal (HPA) axis activation (Farzi et al., 2018; Di Gesù et al., 2021), whilst the activation of the HPA axis has an established relationship with psychiatric disorders such as anxiety (Frankiensztajn et al., 2020) (Figure 1).
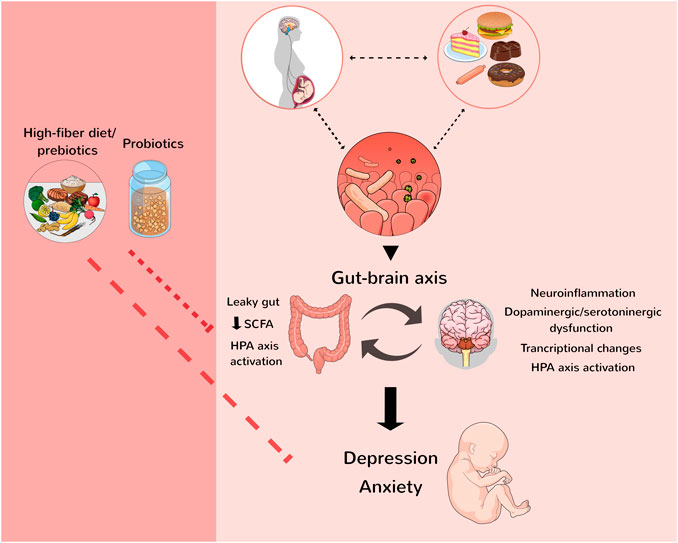
FIGURE 1. Maternal diet can affect the offspring, increasing the chances of metabolic and neurodevelopmental diseases. The gut-brain axis modulates anxiety and depression development since maternal microbiota influence the offspring’s behaviour. Some mechanisms showed in the figure are proposed to influence the development of neurological disorders such as psychiatric manifestations. The consumption of a high-fiber diet, prebiotics, and probiotics is helpful to modulate the offspring’s microbiome and, thus, can diminish the risk of mental illnesses. HPA, hypothalamus-pituitary-adrenal; SCFA, short-chain fatty acids. Figure created by the authors with the assistance from Mind the Graph (mindthegraph.com).
Therefore, microbiota and its metabolites may provide an interesting mechanism of how the gut-brain axis is involved in the effects of obesity on the offspring’s brain and also provide a potential therapeutic target. The use of prebiotics and probiotics has provoked interest in the scientific community in recent years. While probiotics can modulate the composition of intestinal bacteria, prebiotics are the substrate used for fermentation and production of metabolites with an interest to improve host health. Recent research data indicate positive impacts of these strategies regarding metabolic diseases and also in the CNS, including improvement of cognitive impairment, anxiety, and depression [for a review see (Liu et al., 2019; Paiva et al., 2020)]. The gestation period could be a suitable and opportune moment to introduce, or even prioritize the probiotics therapy to benefit the offspring. In an animal model, consumption of probiotics concomitant with an HFD started before pregnancy and continued until lactation was able to protect mice offspring against metabolic disturbances compared to offspring of dams on HFD alone (Guo et al., 2018). On the other hand, its effect in pregnant women is controversial since promising literature (Desai et al., 2021) contrasts with evidence such as a randomized clinical trial that found no benefits with Lactobacillus rhamnosus and Bifidobacterium lactis supplementation in obese pregnant women regarding depression and well-being (Dawe et al., 2020). Therefore, future studies are needed to robustly assert whether probiotic supplementation during pregnancy would be useful to prevent the development of mood disorders such as depression and anxiety in the progeny.
Current Research Gaps and Conclusion
As already mentioned, dysbiosis may be related to the genesis of chronic inflammation in obesity and, through the gut-brain axis, trigger neuroinflammation and the development of psychiatric manifestations (Peirce and Alviña, 2019; Milano et al., 2020; Shi et al., 2020). Maternal diet influences offspring’s brain development, e.g., affecting microglial maturation and gene expression (Thion et al., 2018) and also gut microbiota composition (Buffington et al., 2016). Moreover, brain cells such as microglia can be influenced by the gut-brain axis acquiring a more pro-inflammatory profile and triggering behavioural disturbances (Duan et al., 2021).
In this review, we highlighted the influence of maternal overnutrition and obesity on offspring’s CNS manifestations, mainly on depression and anxiety, psychiatric disorders increasingly prevalent in the world. We also show how the gut-brain axis is altered during pregnancy and obesity. Accordingly, healthier food consumption during pregnancy must be encouraged to prevent this global health issue (Hanson et al., 2017; Bhagavata Srinivasan et al., 2018).
Depression and anxiety may manifest at different stages of life, either in childhood, adulthood, or aging. Whenever it happens, the causes regarding maternal exposure cannot be excluded. Thus, a particularly productive focus for research would be to better understand the influence of the microbiota of obese mothers on the offspring’s brain, allowing the establishment of the importance of therapeutic interventions in this context. Modulating the microbiota through a healthier diet can be beneficial to the offspring, as maternal and offspring dietary fiber consumption was demonstrated to diminish cognitive and social disabilities while improving synaptic function and microglia maturation in the offspring (Liu et al., 2021). The use of prebiotics and probiotics has shown promising results to modulate the maternal and offspring microbiota, and it could be an interesting strategy to prevent psychiatric conditions, such as anxiety and depression (Sanders et al., 2019; Lee, 2021). However, conflicting results and the lack of knowledge about the mechanisms of action of these interventions should encourage future research focused on this subject.
Author Contributions
Conceptualization, MG, VBJ, RPG, and LSF; writing and literature review, all the authors; critical revision of the manuscript, VB-J, RPG, and LSF; figure and table preparation, JJ and IDT. All authors have read, edited and approved the final version of the article.
Funding
Our research groups received financial support, over the years, from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS), Fundo de Incentivo à Pesquisa e Eventos do Hospital de Clínicas de Porto Alegre (FIPE/HCPA), Universidade Federal de Ciências da Saúde de Porto Alegre (UFCSPA), National Institute of Science and Technology on Neuroimmunomodulation (INCT-NIM) and Erasmus+ Funding Programme.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Al Rubaye, H., Adamson, C. C., and Jadavji, N. M. (2021). The Role of Maternal Diet on Offspring Gut Microbiota Development: A Review. J. Neurosci. Res. 99, 284–293. doi:10.1002/jnr.24605
Aubuchon-Endsley, N., Morales, M., Giudice, C., Bublitz, M. H., Lester, B. M., Salisbury, A. L., et al. (2017). Maternal Pre-pregnancy Obesity and Gestational Weight Gain Influence Neonatal Neurobehaviour. Matern. Child Nutr. 13, e12317. doi:10.1111/mcn.12317
Barker, D., and Osmond, C. (1986). Infant Mortality, Childhood Nutrition, and Ischaemic Heart Disease in England and Wales. The Lancet 327, 1077–1081. doi:10.1016/s0140-6736(86)91340-1
Belujon, P., and Grace, A. A. (2017). Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 20, 1036–1046. doi:10.1093/ijnp/pyx056
Bergmann, S., Schlesier-Michel, A., Wendt, V., Grube, M., Keitel-Korndörfer, A., Gausche, R., et al. (2016). Maternal Weight Predicts Children's Psychosocial Development via Parenting Stress and Emotional Availability. Front. Psychol. 7, 1156. doi:10.3389/fpsyg.2016.01156
Bhagavata Srinivasan, S. P., Raipuria, M., Bahari, H., Kaakoush, N. O., and Morris, M. J. (2018). Impacts of Diet and Exercise on Maternal Gut Microbiota Are Transferred to Offspring. Front. Endocrinol. 9, 716. doi:10.3389/fendo.2018.00716
Bruce-Keller, A. J., Fernandez-Kim, S.-O., Townsend, R. L., Kruger, C., Carmouche, R., Newman, S., et al. (2017). Maternal Obese-type Gut Microbiota Differentially Impact Cognition, Anxiety and Compulsive Behavior in Male and Female Offspring in Mice. PLoS One 12, e0175577. doi:10.1371/journal.pone.0175577
Buffington, S. A., Di Prisco, G. V., Auchtung, T. A., Ajami, N. J., Petrosino, J. F., and Costa-Mattioli, M. (2016). Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 165, 1762–1775. doi:10.1016/j.cell.2016.06.001
Cárdenas-Tueme, M., Montalvo-Martínez, L., Maldonado-Ruiz, R., Camacho-Morales, A., and Reséndez-Pérez, D. (2020). Neurodegenerative Susceptibility during Maternal Nutritional Programing: Are Central and Peripheral Innate Immune Training Relevant. Front. Neurosci. 14, 13. doi:10.3389/fnins.2020.00013
Careaga, M., Murai, T., and Bauman, M. D. (2017). Maternal Immune Activation and Autism Spectrum Disorder: From Rodents to Nonhuman and Human Primates. Biol. Psychiatry 81, 391–401. doi:10.1016/j.biopsych.2016.10.020
Cattane, N., Räikkönen, K., Anniverno, R., Mencacci, C., Riva, M. A., Pariante, C. M., et al. (2021). Depression, Obesity and Their Comorbidity during Pregnancy: Effects on the Offspring's Mental and Physical Health. Mol. Psychiatry 26, 462–481. doi:10.1038/s41380-020-0813-6
Chen, C., Xu, X., and Yan, Y. (2018). Estimated Global Overweight and Obesity burden in Pregnant Women Based on Panel Data Model. PLoS One 13, e0202183. doi:10.1371/journal.pone.0202183
Chu, D. M., Antony, K. M., Ma, J., Prince, A. L., Showalter, L., Moller, M., et al. (2016). The Early Infant Gut Microbiome Varies in Association with a Maternal High-Fat Diet. Genome Med. 8, 77. doi:10.1186/s13073-016-0330-z
Crovesy, L., Masterson, D., and Rosado, E. L. (2020). Profile of the Gut Microbiota of Adults with Obesity: a Systematic Review. Eur. J. Clin. Nutr. 74, 1251–1262. doi:10.1038/s41430-020-0607-6
Dash, S., Clarke, G., Berk, M., and Jacka, F. N. (2015). The Gut Microbiome and Diet in Psychiatry. Curr. Opin. Psychiatry 28, 1–6. doi:10.1097/YCO.0000000000000117
Dawe, J. P., McCowan, L. M. E., Wilson, J., Okesene-Gafa, K. A. M., and Serlachius, A. S. (2020). Probiotics and Maternal Mental Health: A Randomised Controlled Trial Among Pregnant Women with Obesity. Sci. Rep. 10, 1291. doi:10.1038/s41598-020-58129-w
Dawson, S. L., O'Hely, M., Jacka, F. N., Ponsonby, A.-L., Symeonides, C., Loughman, A., et al. (2021). Maternal Prenatal Gut Microbiota Composition Predicts Child Behaviour. EBioMedicine 68, 103400. doi:10.1016/j.ebiom.2021.103400
de la Garza, A., Garza-Cuellar, M., Silva-Hernandez, I., Cardenas-Perez, R., Reyes-Castro, L., Zambrano, E., et al. (2019). Maternal Flavonoids Intake Reverts Depression-like Behaviour in Rat Female Offspring. Nutrients 11, 572. doi:10.3390/nu11030572
Desai, V., Kozyrskyj, A. L., Lau, S., Sanni, O., Dennett, L., Walter, J., et al. (2021). Effectiveness of Probiotic, Prebiotic, and Synbiotic Supplementation to Improve Perinatal Mental Health in Mothers: A Systematic Review and Meta-Analysis. Front. Psychiatry 12, 622181. doi:10.3389/fpsyt.2021.622181
Di Gesù, C. M., Matz, L. M., and Buffington, S. A. (2021). Diet-induced Dysbiosis of the Maternal Gut Microbiome in Early Life Programming of Neurodevelopmental Disorders. Neurosci. Res. 168, 3–19. doi:10.1016/j.neures.2021.05.003
Duan, C., Huang, L., Zhang, C., Zhang, L., Xia, X., Zhong, Z., et al. (2021). Gut Commensal-Derived Butyrate Reverses Obesity-Induced Social Deficits and Anxiety-like Behaviors via Regulation of Microglial Homeostasis. Eur. J. Pharmacol. 908, 174338. doi:10.1016/j.ejphar.2021.174338
Eckburg, P. B., Bik, E. M., Bernstein, C. N., Purdom, E., Dethlefsen, L., Sargent, M., et al. (2005). Diversity of the Human Intestinal Microbial flora. Science 308, 1635–1638. doi:10.1126/science.1110591
Edlow, A. G. (2017). Maternal Obesity and Neurodevelopmental and Psychiatric Disorders in Offspring. Prenat. Diagn. 37, 95–110. doi:10.1002/pd.4932
Farzi, A., Fröhlich, E. E., and Holzer, P. (2018). Gut Microbiota and the Neuroendocrine System. Neurotherapeutics 15, 5–22. doi:10.1007/s13311-017-0600-5
Ferretti, P., Pasolli, E., Tett, A., Asnicar, F., Gorfer, V., Fedi, S., et al. (2018). Mother-to-Infant Microbial Transmission from Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host & Microbe 24, 133–145. e5. doi:10.1016/j.chom.2018.06.005
Frankiensztajn, L. M., Elliott, E., and Koren, O. (2020). The Microbiota and the Hypothalamus-Pituitary-Adrenocortical (HPA) axis, Implications for Anxiety and Stress Disorders. Curr. Opin. Neurobiol. 62, 76–82. doi:10.1016/j.conb.2019.12.003
Fusco, S., Spinelli, M., Cocco, S., Ripoli, C., Mastrodonato, A., Natale, F., et al. (2019). Maternal Insulin Resistance Multigenerationally Impairs Synaptic Plasticity and Memory via Gametic Mechanisms. Nat. Commun. 10, 4799. doi:10.1038/s41467-019-12793-3
García-Mantrana, I., Selma-Royo, M., González, S., Parra-Llorca, A., Martínez-Costa, C., and Collado, M. C. (2020). Distinct Maternal Microbiota Clusters Are Associated with Diet during Pregnancy: Impact on Neonatal Microbiota and Infant Growth during the First 18 Months of Life. Gut Microbes 11, 962–978. doi:10.1080/19490976.2020.1730294
Gawlinska, K., Gawlinski, D., Przegalinski, E., and Filip, M. (2019). Maternal High-Fat Diet during Pregnancy and Lactation Provokes Depressive-like Behavior and Influences the Irisin/brain-Derived Neurotrophic Factor axis and Inflammatory Factors in Male and Female Offspring in Rats. J. Physiol. Pharmacol. 70 (3), 407–417. doi:10.26402/jpp.2019.3.07
Gawlińska, K., Gawliński, D., Korostyński, M., Borczyk, M., Frankowska, M., Piechota, M., et al. (2021). Maternal Dietary Patterns Are Associated with Susceptibility to a Depressive-like Phenotype in Rat Offspring. Develop. Cogn. Neurosci. 47, 100879. doi:10.1016/j.dcn.2020.100879
Gohir, W., Whelan, F. J., Surette, M. G., Moore, C., Schertzer, J. D., and Sloboda, D. M. (2015). Pregnancy-related Changes in the Maternal Gut Microbiota Are Dependent upon the Mother's Periconceptional Diet. Gut Microbes 6, 310–320. doi:10.1080/19490976.2015.1086056
Gottfried, C., and Bambini-Junior, V. (2018). Insights into the Relationship of the Immune System with Neurodevelopmental and Psychiatric Disorders. Neuroimmunomodulation 25, 243–245. doi:10.1159/000496180
Guo, Y., Wang, Z., Chen, L., Tang, L., Wen, S., Liu, Y., et al. (2018). Diet Induced Maternal Obesity Affects Offspring Gut Microbiota and Persists into Young Adulthood. Food Funct. 9, 4317–4327. doi:10.1039/c8fo00444g
Hanson, M., Barker, M., Dodd, J. M., Kumanyika, S., Norris, S., Steegers, E., et al. (2017). Interventions to Prevent Maternal Obesity before conception, during Pregnancy, and post Partum. Lancet Diabetes Endocrinol. 5, 65–76. doi:10.1016/S2213-8587(16)30108-5
Hatanaka, Y., Kabuta, T., and Wada, K. (2017). Disturbance in Maternal Environment Leads to Abnormal Synaptic Instability during Neuronal Circuitry Development. Front. Neurosci. 11, 35. doi:10.3389/fnins.2017.00035
Howell, K. R., and Powell, T. L. (2017). Effects of Maternal Obesity on Placental Function and Fetal Development. Reproduction 153, R97–R108. doi:10.1530/REP-16-0495
Huang, F., and Wu, X. (2021). Brain Neurotransmitter Modulation by Gut Microbiota in Anxiety and Depression. Front. Cel Dev. Biol. 9, 649103. doi:10.3389/fcell.2021.649103
Jo, J.-K., Seo, S.-H., Park, S.-E., Kim, H.-W., Kim, E.-J., Kim, J.-S., et al. (2021). Gut Microbiome and Metabolome Profiles Associated with High-Fat Diet in Mice. Metabolites 11, 482. doi:10.3390/metabo11080482
Koren, O., Goodrich, J. K., Cullender, T. C., Spor, A., Laitinen, K., Kling Bäckhed, H., et al. (2012). Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 150, 470–480. doi:10.1016/j.cell.2012.07.008
Kumpulainen, S. M., Girchenko, P., Lahti-Pulkkinen, M., Reynolds, R. M., Tuovinen, S., Pesonen, A.-K., et al. (2018). Maternal Early Pregnancy Obesity and Depressive Symptoms during and after Pregnancy. Psychol. Med. 48, 2353–2363. doi:10.1017/S0033291717003889
Lee, S.-K. (2021). Don't Worry, Heavy Moms; Just Eat Your Broccoli(or Kimchi)!. Mol. Cell 44, 422–424. doi:10.14348/molcells.2021.0118
Liu, R. T., Walsh, R. F. L., and Sheehan, A. E. (2019). Prebiotics and Probiotics for Depression and Anxiety: A Systematic Review and Meta-Analysis of Controlled Clinical Trials. Neurosci. Biobehavioral Rev. 102, 13–23. doi:10.1016/j.neubiorev.2019.03.023
Liu, X., Li, X., Xia, B., Jin, X., Zou, Q., Zeng, Z., et al. (2021). High-fiber Diet Mitigates Maternal Obesity-Induced Cognitive and Social Dysfunction in the Offspring via Gut-Brain axis. Cel Metab. 33, 923–938. e6. doi:10.1016/j.cmet.2021.02.002
Martin, C. R., Osadchiy, V., Kalani, A., and Mayer, E. A. (2018). The Brain-Gut-Microbiome Axis. Cell Mol. Gastroenterol. Hepatol. 6, 133–148. doi:10.1016/j.jcmgh.2018.04.003
Matuszewska, J., Zalewski, T., Klimaszyk, A., Ziarniak, K., Jurga, S., Chmurzynska, A., et al. (2021). Mothers' Cafeteria Diet Induced Sex-specific Changes in Fat Content, Metabolic Profiles, and Inflammation Outcomes in Rat Offspring. Sci. Rep. 11, 18573. doi:10.1038/s41598-021-97487-x
McDonald, S. D., McKinney, B., Foster, G., Taylor, V., Lutsiv, O., and Pullenayegum, E. (2015). The Combined Effects of Maternal Depression and Excess Weight on Neonatal Outcomes. Int. J. Obes. 39, 1033–1040. doi:10.1038/ijo.2015.44
Menting, M. D., van de Beek, C., Mintjens, S., Wever, K. E., Korosi, A., Ozanne, S. E., et al. (2019). The Link between Maternal Obesity and Offspring Neurobehavior: A Systematic Review of Animal Experiments. Neurosci. Biobehavioral Rev. 98, 107–121. doi:10.1016/j.neubiorev.2018.12.023
Milano, W., Ambrosio, P., Carizzone, F., De Biasio, V., Di Munzio, W., Foia, M. G., et al. (2020). Depression and Obesity: Analysis of Common Biomarkers. Diseases 8, 23. doi:10.3390/diseases8020023
Morais, L. H., Schreiber, H. L., Mazmanian, S. K., and Mazmanian, S. K. (2021). The Gut Microbiota-Brain axis in Behaviour and Brain Disorders. Nat. Rev. Microbiol. 19, 241–255. doi:10.1038/s41579-020-00460-0
Naef, L., Moquin, L., Dal Bo, G., Giros, B., Gratton, A., and Walker, C.-D. (2011). Maternal High-Fat Intake Alters Presynaptic Regulation of Dopamine in the Nucleus Accumbens and Increases Motivation for Fat Rewards in the Offspring. Neuroscience 176, 225–236. doi:10.1016/j.neuroscience.2010.12.037
Naef, L., Srivastava, L., Gratton, A., Hendrickson, H., Owens, S. M., and Walker, C.-D. (2008). Maternal High Fat Diet during the Perinatal Period Alters Mesocorticolimbic Dopamine in the Adult Rat Offspring: Reduction in the Behavioral Responses to Repeated Amphetamine Administration. Psychopharmacology 197, 83–94. doi:10.1007/s00213-007-1008-4
Ortiz-Valladares, M., Pedraza-Medina, R., Pinto-González, M. F., Muñiz, J. G., Gonzalez-Perez, O., and Moy-López, N. A. (2021). Neurobiological Approaches of High-Fat Diet Intake in Early Development and Their Impact on Mood Disorders in Adulthood: A Systematic Review. Neurosci. Biobehavioral Rev. 129, 218–230. doi:10.1016/j.neubiorev.2021.07.028
Paiva, I. H. R., Duarte-Silva, E., and Peixoto, C. A. (2020). The Role of Prebiotics in Cognition, Anxiety, and Depression. Eur. Neuropsychopharmacol. 34, 1–18. doi:10.1016/j.euroneuro.2020.03.006
Paradis, J., Boureau, P., Moyon, T., Nicklaus, S., Parnet, P., and Paillé, V. (2017). Perinatal Western Diet Consumption Leads to Profound Plasticity and GABAergic Phenotype Changes within Hypothalamus and Reward Pathway from Birth to Sexual Maturity in Rat. Front. Endocrinol. 8, 216. doi:10.3389/fendo.2017.00216
Peirce, J. M., and Alviña, K. (2019). The Role of Inflammation and the Gut Microbiome in Depression and Anxiety. J. Neurosci. Res. 97, 1223–1241. doi:10.1002/jnr.24476
Perez-Muñoz, M. E., Arrieta, M.-C., Ramer-Tait, A. E., and Walter, J. (2017). A Critical Assessment of the “Sterile Womb” and “In Utero Colonization” Hypotheses: Implications for Research on the pioneer Infant Microbiome. Microbiome 5, 48. doi:10.1186/s40168-017-0268-4
Rasmussen, J. M., Graham, A. M., Entringer, S., Gilmore, J. H., Styner, M., Fair, D. A., et al. (2019). Maternal Interleukin-6 Concentration during Pregnancy Is Associated with Variation in Frontolimbic white Matter and Cognitive Development in Early Life. Neuroimage 185, 825–835. doi:10.1016/j.neuroimage.2018.04.020
Ribeiro, A. C. A. F., Batista, T. H., Veronesi, V. B., Giusti-Paiva, A., and Vilela, F. C. (2018). Cafeteria Diet during the Gestation Period Programs Developmental and Behavioral Courses in the Offspring. Int. J. Dev. Neurosci. 68, 45–52. doi:10.1016/j.ijdevneu.2018.05.001
Rivera, H. M., Christiansen, K. J., and Sullivan, E. L. (2015). The Role of Maternal Obesity in the Risk of Neuropsychiatric Disorders. Front. Neurosci. 9, 194. doi:10.3389/fnins.2015.00194
Sanders, A., Rackers, H., and Kimmel, M. (2019). A Role for the Microbiome in Mother-Infant Interaction and Perinatal Depression. Int. Rev. Psychiatry 31, 280–294. doi:10.1080/09540261.2018.1548431
Sasaki, A., de Vega, W. C., St-Cyr, S., Pan, P., and McGowan, P. O. (2013). Perinatal High Fat Diet Alters Glucocorticoid Signaling and Anxiety Behavior in Adulthood. Neuroscience 240, 1–12. doi:10.1016/j.neuroscience.2013.02.044
Sasaki, A., de Vega, W., Sivanathan, S., St-Cyr, S., and McGowan, P. O. (2014). Maternal High-Fat Diet Alters Anxiety Behavior and Glucocorticoid Signaling in Adolescent Offspring. Neuroscience 272, 92–101. doi:10.1016/j.neuroscience.2014.04.012
Schmitz, L., Kuglin, R., Bae-Gartz, I., Janoschek, R., Appel, S., Mesaros, A., et al. (2018). Hippocampal Insulin Resistance Links Maternal Obesity with Impaired Neuronal Plasticity in Adult Offspring. Psychoneuroendocrinology 89, 46–52. doi:10.1016/j.psyneuen.2017.12.023
Shi, H., Wang, Q., Zheng, M., Hao, S., Lum, J. S., Chen, X., et al. (2020). Supplement of Microbiota-Accessible Carbohydrates Prevents Neuroinflammation and Cognitive Decline by Improving the Gut Microbiota-Brain axis in Diet-Induced Obese Mice. J. Neuroinflammation 17, 77. doi:10.1186/s12974-020-01760-1
Smith, B. L., and Reyes, T. M. (2017). Offspring Neuroimmune Consequences of Maternal Malnutrition: Potential Mechanism for Behavioral Impairments that Underlie Metabolic and Neurodevelopmental Disorders. Front. Neuroendocrinology 47, 109–122. doi:10.1016/j.yfrne.2017.07.007
Sullivan, E. L., Riper, K. M., Lockard, R., and Valleau, J. C. (2015). Maternal High-Fat Diet Programming of the Neuroendocrine System and Behavior. Horm. Behav. 76, 153–161. doi:10.1016/j.yhbeh.2015.04.008
Thion, M. S., Low, D., Silvin, A., Chen, J., Grisel, P., Schulte-Schrepping, J., et al. (2018). Microbiome Influences Prenatal and Adult Microglia in a Sex-specific Manner. Cell 172, 500–516. e16. doi:10.1016/j.cell.2017.11.042
Tong, L., and Kalish, B. T. (2021). The Impact of Maternal Obesity on Childhood Neurodevelopment. J. Perinatol. 41, 928–939. doi:10.1038/s41372-020-00871-0
Trujillo-Villarreal, L. A., Romero-Díaz, V. J., Marino-Martínez, I. A., Fuentes-Mera, L., Ponce-Camacho, M. A., Devenyi, G. A., et al. (2021). Maternal Cafeteria Diet Exposure Primes Depression-like Behavior in the Offspring Evoking Lower Brain Volume Related to Changes in Synaptic Terminals and Gliosis. Transl. Psychiatry 11, 53. doi:10.1038/s41398-020-01157-x
Tye, K. M., Mirzabekov, J. J., Warden, M. R., Ferenczi, E. A., Tsai, H.-C., Finkelstein, J., et al. (2013). Dopamine Neurons Modulate Neural Encoding and Expression of Depression-Related Behaviour. Nature 493, 537–541. doi:10.1038/nature11740
Val-Laillet, D., Besson, M., Guérin, S., Coquery, N., Randuineau, G., Kanzari, A., et al. (2017). A Maternal Western Diet during Gestation and Lactation Modifies Offspring's Microbiota Activity, Blood Lipid Levels, Cognitive Responses, and Hippocampal Neurogenesis in Yucatan Pigs. FASEB j. 31, 2037–2049. doi:10.1096/fj.201601015R
WHO (2017). Depression and Other Common Mental Disorders: Global Health Estimates. World Health Organization. Available at: https://apps.who.int/iris/bitstream/handle/10665/254610/W?sequence=1 (Accessed October 7, 2021).
Winter, C., von Rumohr, A., Mundt, A., Petrus, D., Klein, J., Lee, T., et al. (2007). Lesions of Dopaminergic Neurons in the Substantia Nigra Pars Compacta and in the Ventral Tegmental Area Enhance Depressive-like Behavior in Rats. Behav. Brain Res. 184, 133–141. doi:10.1016/j.bbr.2007.07.002
Winter, G., Hart, R. A., Charlesworth, R. P. G., and Sharpley, C. F. (2018). Gut Microbiome and Depression: what We Know and what We Need to Know. Rev. Neurosci. 29, 629–643. doi:10.1515/revneuro-2017-0072
Wright, T., Langley-Evans, S. C., and Voigt, J.-P. (2011). The Impact of Maternal Cafeteria Diet on Anxiety-Related Behaviour and Exploration in the Offspring. Physiol. Behav. 103, 164–172. doi:10.1016/j.physbeh.2011.01.008
Keywords: maternal obesity, gut-brain axis, anxiety, depression, microbiota
Citation: Jantsch J, Tassinari ID, Giovenardi M, Bambini-Junior V, Guedes RP and de Fraga LS (2022) Mood Disorders Induced by Maternal Overnutrition: The Role of the Gut-Brain Axis on the Development of Depression and Anxiety. Front. Cell Dev. Biol. 10:795384. doi: 10.3389/fcell.2022.795384
Received: 15 October 2021; Accepted: 10 January 2022;
Published: 26 January 2022.
Edited by:
Patricia Ybot-Gonzalez, Institute of Biomedicine of Seville (CSIC), SpainReviewed by:
Evelyn Nunes Goulart da Silva Pereira, Oswaldo Cruz Foundation (Fiocruz), BrazilCopyright © 2022 Jantsch, Tassinari, Giovenardi, Bambini-Junior, Guedes and de Fraga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luciano Stürmer de Fraga, bHVjaWFub2ZAdWZyZ3MuYnI=
†These authors have contributed equally to this work and share first authorship