Abstract
Post-translational modification (PTM) has a significant impact on cellular signaling and function regulation. In pancreatic β cells, PTMs are involved in insulin secretion, cell development, and viability. The dysregulation of PTM in β cells is clinically associated with the development of diabetes mellitus. Here, we summarized current findings on major PTMs occurring in β cells and their roles in insulin secretion. Our work provides comprehensive insight into understanding the mechanisms of insulin secretion and potential therapeutic targets for diabetes from the perspective of protein PTMs.
1 Introduction
Insulin is an anabolic hormone released from pancreatic islet β cells with the distinct capacity to maintain blood glucose homeostasis (Cabrera et al., 2006; Da Silva Xavier, 2018). The secretion of insulin in response to glucose and other nutrients (such as amino acids and free fatty acids) is a complex process involving the coordination of multiple signaling pathways (Nolan et al., 2006; Henquin, 2011). Impairment of this process is directly associated with the development of diabetes mellitus (Schwartz et al., 2013). Thus, the insulin secretion process is considered as a promising target for the treatment of diabetes mellitus (DeFronzo et al., 2014). But the detail of insulin secretion regulation is still an unanswered question.
Post-translational modification (PTM) is the covalent modification with addition or removal of chemical groups on proteins (Walsh et al., 2005). It is closely associated with almost all physiological and pathological processes by regulating protein localization, degradation, and functions (Walsh and Jefferis, 2006; Khan et al., 2016; Morales-Tarre et al., 2021; Zhu and Hart, 2021). Accumulating evidence suggests that PTMs are extensively involved in the insulin secretion process, Currently, at least eight types of PTMs are known associated with insulin secretion. For example, phosphorylation is required for signaling cascades mediating insulin secretion (Campbell and Newgard, 2021). SUMOylation and palmitoylation have been reported to regulate insulin secretion at multiple stages (Davey et al., 2019; Chamberlain et al., 2021). Acetylation, ubiquitination and O-GlcNAcylation are involved in insulin gene transcription (Mounier and Posner, 2006; Ozcan et al., 2010; Sampley and Ozcan, 2012). Even some understudied PTMs such as citrullination and deamidation are recently reported to be linked with insulin secretion. These indicate that protein PTM plays a critical role in the regulation of insulin secretion in β cells. However, a systematic review of current findings of PTM in insulin secretion is still missing. Here, we review the current understanding of the functional roles of these PTMs in the insulin secretion process.
2 Insulin secretion
The main physiological stimulus for insulin secretion is blood glucose. Activation of insulin secretion by elevated glucose concentration is called glucose-stimulated insulin secretion (GSIS) (Figure 1). It includes two tandem pathways, the triggering pathway and the amplifying pathway. In the triggering pathway, glucose is transported into islet β cells via glucose transporters (GLUTs) and converted to glucose-6-phosphate (G-6-P) by glucokinase (GK). G-6-P further enters the tricarboxylic acid (TCA) cycle through glycolysis, leading to the production of adenosine triphosphate (ATP). Increased ATP/adenosine diphosphate (ADP) ratio in the cytoplasm causes the closure of the KATP channel (composed of SUR1 subunits and Kir6.2 subunits) in the cell membrane (Seino, 1999). This leads to the generation of electro-voltage between the inside and outside of the cell membrane, opening voltage-dependent Ca2+ channels (VDCCs) and inducing calcium influx (Lim et al., 2009). Subsequently, calcium influx is sensed by multiple calcium-binding proteins and triggers insulin granules exocytosis. The amplifying pathway increases the sensitivity of insulin secretion to the induced calcium influx, which is independent of the KATP channel (Kalwat and Cobb, 2017). Therefore, the amplifying pathway is also referred to the KATP-independent pathway. However, the exact mechanisms of action of the amplifying pathway are not fully clear (Kalwat and Cobb, 2017). Store-operated calcium channels (SOCs) are believed as a critical compartment of the amplifying pathway. SOCs can be activated by Ca2+ depletion from the endoplasmic reticulum (ER) (Prakriya and Lewis, 2015; Lopez et al., 2020). Upon the depletion of ER Ca2+ store, stromal interaction molecule 1 (STIM1) aggregates and translocates to the plasma membrane, where it interacts directly with Calcium Release-Activated Calcium Modulator 1 (Orai1) and transient receptor potential canonical channel-1 (TRPC1). This interaction opens SOCs and initiates Ca2+ influx, which induces insulin secretion (Tian et al., 2012b). Another critical compartment in the amplifying pathway is the second messenger cyclic adenosine monophosphate (cAMP) (Tian et al., 2015). The cAMP signaling mediates insulin secretion induced by glucose, free fatty acid (FFA) and Glucagon and glucagon-like peptide-1 (GLP-1) (Dyachok et al., 2008; Tian et al., 2011; Tian et al., 2012a).
FIGURE 1
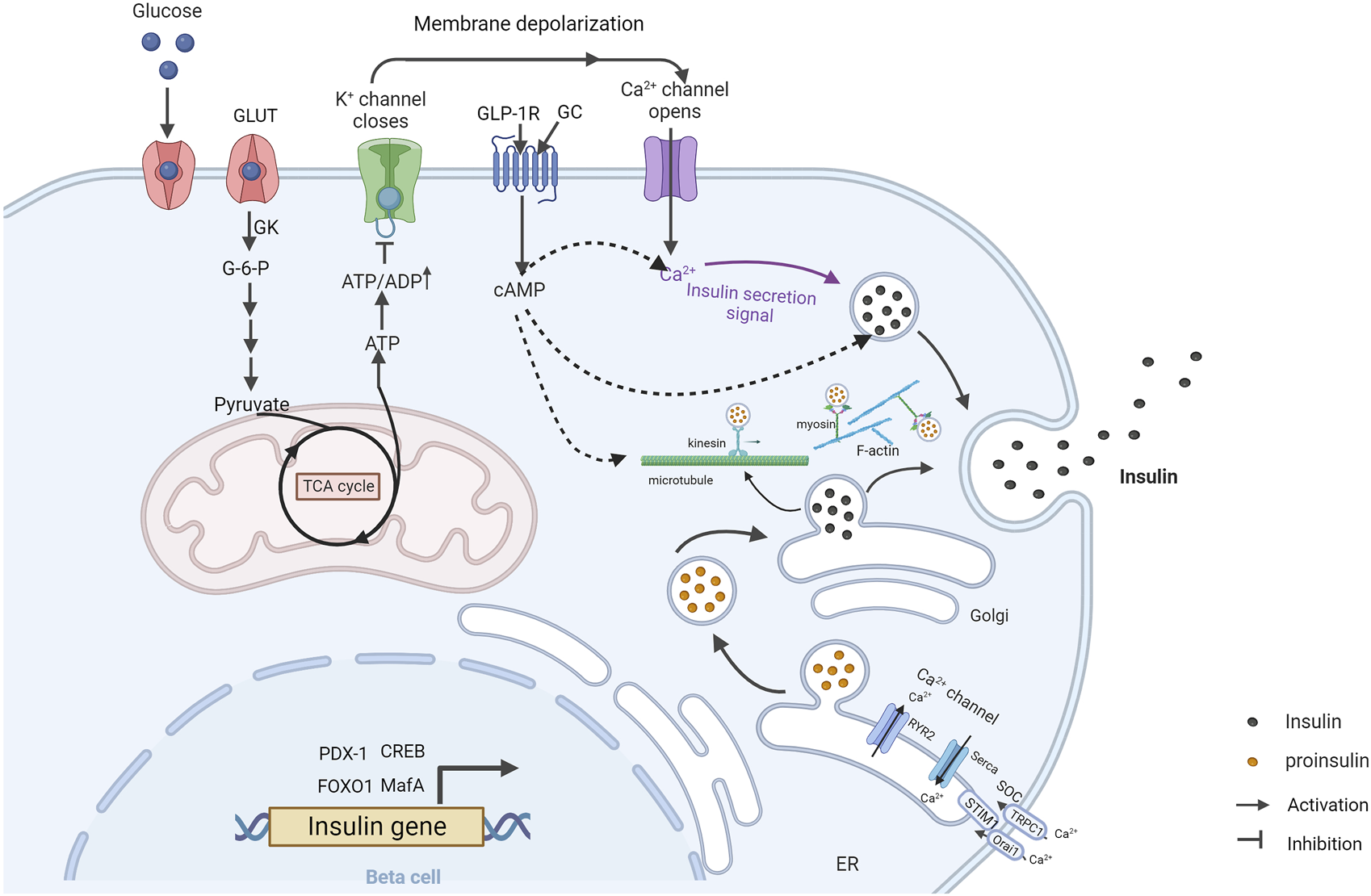
Overview of glucose-stimulated insulin secretion in β cells (Created with BioRender.com). GSIS is mediated by the triggering pathway (solid arrows) and amplification pathways (dashed arrows). At high blood glucose levels, glucose is taken up and converted to glucose-6-phosphate through GK. Glucose-6-phosphate is metabolized by glycolysis and subsequently enters the tricarboxylic acid cycle, triggering an increase in intracellular ATP. Increasing ATP/ADP ratio leads to the closure of the KATP channel and the opening of Ca2+ channels, allowing calcium influx and triggering insulin granules exocytosis. Calcium depletion from ER causes STIM1 to accumulate at ER-plasma membrane junctions where it gates Orai1 and TRPC1, leading to calcium influx and subsequent insulin granules exocytosis. GLP-1 stimulates insulin secretion through cAMP signal pathway. Insulin-vesicles are transported by motor proteins along cytoskeletal biopolymers. Insulin gene transcription is regulated by multiple transcription factors. GC, glucagon; GK, glucokinase; ER, endoplasmic reticulum.
Insulin secretion in response to glucose stimulation occurs in a biphasic manner composed of a transient first phase and followed a prolonged second phase. Released insulin granules in the first-phase are mainly from a “readily releasable pool” (RRP) in the vicinity of the plasma membrane. Released insulin granules in the second phase are mainly recruited from intracellular storage pools. Actin filaments and microtubules are two main transport routes for insulin granules in β cells (Thurmond et al., 2003; Varadi et al., 2003; Varadi et al., 2005; Cui et al., 2011). Actin filaments mainly contribute to the short-range movement near the plasma membrane, whereas microtubules contribute to the long-range movements (Omar-Hmeadi and Idevall-Hagren, 2021). The remodeling of F-actin cytoskeleton and microtubule network is critical for GSIS in β cells (Thurmond et al., 2003; Bracey et al., 2020; Ho et al., 2020; Wang et al., 2020). Moreover, the formation of SNARE complex which is composed of t-SNARE proteins (SNAP-25 and syntaxin) in the plasma membrane and v-SNARE protein (VAMP) on the insulin granules membrane, is also important for insulin exocytosis.
3 Regulatory effect of PTMs on insulin secretion
Various PTMs related to insulin secretion have been reported. In the following sections, we will outline currently known PTMs and related substrates in β cells (Table 1), and discuss the roles of individual PTMs in the regulation of insulin secretion (Figure 2).
TABLE 1
| Protein name | Protein description | Modification type | Modification function | |
|---|---|---|---|---|
| Regulatory direction on insulin secretion | Details | |||
| CRTC1 | Transcription factor; Regulate insulin gene transcription | Phosphorylation | negative | CRTC is phosphorylated and resides in the cytoplasm |
| FOXO1 | Acetylation | negative | FOXO1 is acetylated and resides in the cytoplasm | |
| CREB | Ubiquitination | negative | Ubiquitination of CREB induces its degradation | |
| PDX-1 | Ubiquitination | negative | Ubiquitination of PDX-1 induces its degradation | |
| SUMOylation | positive | SUMOylation protects PDX-1 from proteasomal degradation | ||
| O-GlcNAcylation | positive | O-GlcNAcylation increase DNA binding activity of PDX-1 | ||
| MafA | SUMOylation | negative | SUMOylation of MafA results in reduced transcriptional activity | |
| Ubiquitination | negative | Ubiquitination of MafA induces its degradation | ||
| NeuroD1 | O-GlcNAcylation | positive | NeuroD1 is O-GlcNAcylated and transfers into the nucleus | |
| histone H3 | Core component of nucleosome; Regulate insulin gene transcription | Acetylation | positive | Acetylation increase histone H3 transcriptional activation |
| O-GlcNAcylation | positive | O-GlcNAcylation elevates histone H3 transcriptional activation | ||
| GK | Glucokinase; Convert glucose to G-6-P, promote TCA cycle and ATP production | Ubiquitination | negative | Ubiquitination of GK induces its degradation |
| SUMOylation | positive | SUMOylation protects GK from ubiquitination and degradation | ||
| Citrullination | negative | Citrullination reduces substrate binding affinity of GK | ||
| Kir6.2 | Component of KATP channel; Close KATP channel and induce electro-voltage generation | Phosphorylation | positive | Phosphorylation of Kir6.2 induces inhibition of the KATP channel conductance |
| Palmitoylation | negative | Palmitoylated Kir6.2 increases the open state of KATP | ||
| β2a | An important auxiliary subunit of VDCCs; Open VDCCs and induce calcium influx | Palmitoylation | positive | Palmitoylation increases the plasma membrane trafficking and location of β2a |
| CDK5R1 (p35) | A cell cycle-dependent protein kinase; Phosphorylates α1C subunit of L-VDCC and inhibits L-VDCC activity | Phosphorylation | positive | Phosphorylation of CDK5R1 triggers its ubiquitination and degradation |
| Syt7 | Cytoplasmic Ca2+ sensor; Trigger Ca2+-dependent insulin secretion | Phosphorylation | positive | Phosphorylation of Syt7 enhances GLP-1–dependent insulin secretion |
| RyR2 | Component of ER intracellular calcium channels; Mediates the release of Ca2+ from ER into the cytoplasm | Phosphorylates | positive | Phosphorylation of RyR2 promotes Ca2+-dependent insulin secretion |
| TRPC1 | Component of SOCs; Cooperate with Orai1 and STIM1, induce SOCE | Phosphorylation | positive | Phosphorylation of TRPC1 enhances insulin secretion |
| tau | Microtubule-associated protein; Mediates insulin vesicles movement | Phosphorylation | positive | Tau is phosphorylated and dissociates from microtubules, which promotes microtubules turning over |
| Myosin | Actin-based molecular motor; Regulate insulin granules trafficking | Phosphorylation | positive | Phosphorylation of MHC and MLC increases insulin secretory granules translocation |
| β-catenin | A cell-cell adhesion protein; Regulate insulin vesicles trafficking | Phosphorylation | positive | Phosphorylation of β-catenin promotes the rearrangement of the actin cytoskeleton |
| syntaxin 1A | Part of the SNARE; Participate in the insulin exocytosis | SUMOylation | negative | SUMOylation enhances the binding between syntaxin 1A and tomosyn-1A |
| tomosyn-1A | Syntaxin 1A binding proteins; Bind with syntaxin 1A to inhibit SNARE complex formation | SUMOylation | negative | |
| tomosyn-2 | Phosphorylation/Ubiquitination | positive | Phosphorylation of tomosyn-2 leads to its ubiquitination and degradation | |
| SNAP-25 | A component of SNARE complex; Participate in insulin exocytosis | Palmitoylation | positive | Palmitoylation of SNAP-25 increase its membrane localization |
| Scamp1 | A recycling carrier to the cell surface in post-Golgi recycling pathways; Involved in insulin secretion | Palmitoylation | positive | Palmitoylation of Scamp1 prolonged vesicular kiss-and-run or cavicapture events |
| PAK1 | A protein kinase; Regulate insulin exocytosis as an effector of Rho GTPases | Phosphorylation | positive | Phosphorylation of PAK1 stimulates cytoskeletal remodeling required for insulin exocytosis |
| PTB1 | A mRNA binding protein; Regulate secretory granule proteins level | Phosphorylation | positive | Phosphorylation enhances the binding activity of PTB1 to the 3′UTR of mRNAs encoding secretory granule proteins |
| GLP-1R | GLP-1 reporter; Regulating insulin secretion in response to GLP-1 | SUMOylation | negative | SUMOylation attenuates cell surface trafficking of GLP-1R |
| Palmitoylation | positive | Palmitoylation induces GLP-1R clustering, nanodomain signaling, and internalization | ||
| GDIα | Locks Rho GTPases in an inactive GDP-bound form and inhibits GSIS | Phosphorylation | positive | GDIα is phosphorylated and dissociates from Rho GTPases, leading to the onset of GSIS |
| PLD2 | A phospholipase; Hydrolyzes phosphatidyl choline to generate phosphatidic acid, a mediator of insulin exocytosis | Phosphorylation | positive | Phosphorylation of PLD2 activates EGF-dependent insulin secretion |
| FADD | An apoptotic adaptor molecule | Phosphorylation | negative | Phosphorylation of FADD induces impaired GSIS |
| IAPP | Islet amyloid polypeptide; Co-expresses and secretes with insulin; Induces toxicity toward β cells and inhibits insulin secretion | Deamidation | negative | Deamidation modulates IAPP amyloid formation and fibril morphology, inducing its cytotoxicity |
Summary of PTM regulation in insulin secretion.
FIGURE 2
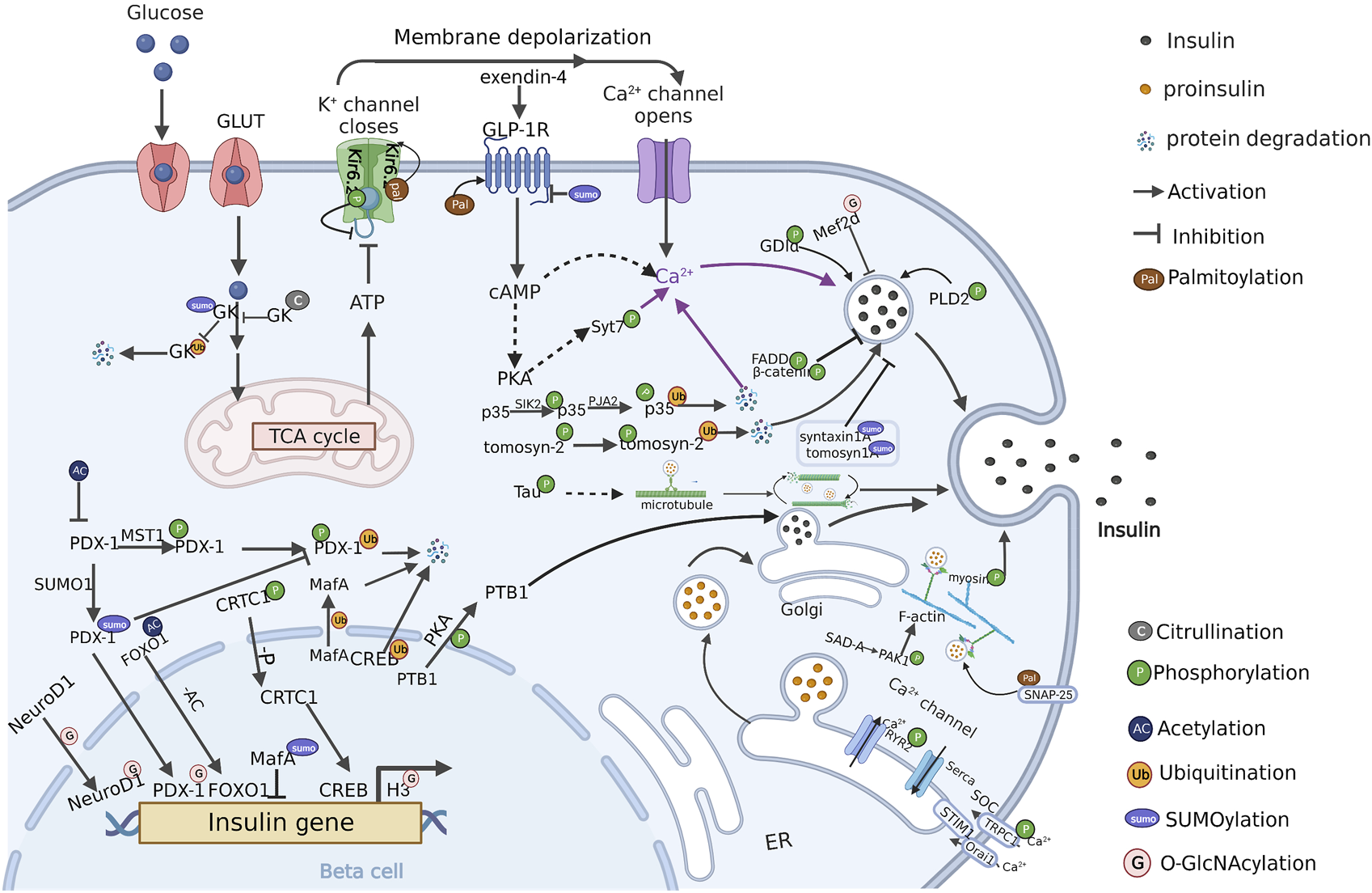
PTM regulation of insulin secretion in β cells (Created with BioRender.com). PTMs are ubiquitously engaged in the various stages of insulin secretion, including signal regulation, insulin gene transcription, insulin-vesicles transportation. Multiple PTMs synergistically regulate insulin secretion in pancreatic β cells. The crosstalk between these PTMs and the modified protein substrates currently known is present in the figure. P: Phosphorylation, AC: Acetylation, Ub: Ubiquitination, SUMO: SUMOylation, G: O-GlcNAcylation, Pal: Palmitoylation, C: Citrullination.
3.1 Phosphorylation
Phosphorylation is a ubiquitous PTM regulated by protein kinases and phosphatases that transfer and remove phosphate groups from ATP or GTP to amino acid residues (Ser, Tyr, Thr) of proteins, respectively. It is widely involved in signal transduction pathways and associated with various physiological and pathological processes by regulating cell proliferation, development, differentiation, apoptosis, and other cell processes (Humphrey et al., 2015). Phosphorylation mediated signaling cascades play an important role in maintaining glucose homeostasis (Roder et al., 2016). The insulin/insulin receptor and their downstream proteins, such as PI3K/AKT and ERK, regulate various metabolic pathways including improving glucose tolerance and protecting against insulin resistance (Dall'Agnese et al., 2022). In the insulin secretion process, phosphorylation is wildly existing in the insulin biosynthesis and signaling cascade of insulin exocytosis (Sacco et al., 2016).
Transcription factors are required in insulin gene transcription, and regulated by phosphorylation (Knoch et al., 2006; Ch’ng et al., 2012; Malm et al., 2016). cAMP-Regulated Transcriptional Co-activator-1 (CRTC1) is a cAMP-responsive element binding protein (CREB)-mediated transcription regulator. Under basal conditions, CRTC is phosphorylated by Salt Inducible Kinase (SIK) and binds to 14-3-3 proteins, sequestering in the cytoplasm (Ch’ng et al., 2012; Malm et al., 2016). Dephosphorylation of CRTC by activating CRTC phosphatase or inhibiting CRTC kinases triggers its dissociation from 14-3-3 and translocation into the nucleus, where it activates CREB-mediated transcription of Ins1 gene (Oetjen et al., 1994; Altarejos et al., 2008; Altarejos and Montminy, 2011). It has been reported that this process is involved in GLP-1-stimulated insulin secretion (Shin et al., 2014). Cytosolic polypyrimidine tract-binding protein 1 (PTB1) binds and stabilizes mRNA encoding secretory granules (SGs) (Knoch et al., 2004). PKA-dependent PTB1 phosphorylation is promoted by elevated cAMP and in turn promotes SG expression and insulin secretion (Knoch et al., 2006).
In β cells, Ca2+ is considered as triggering signal of insulin exocytosis. Phosphorylation is involved in the depolarization of membrane potential and calcium influx. Kir6.2, the KATP channel component, can be phosphorylated on Ser385 by AMPK. Phosphorylated Kir6.2 restrains KATP channel activity and leads to depolarization of membrane potential. This depolarization results in the opening of VDCCs, leading to elevated intracellular calcium levels and ultimately induced insulin secretion (Chang et al., 2009). Synaptotagmin-7 (Syt7) is a major cytoplasmic Ca2+ sensor for exocytosis by triggering secretory granule fusion and insulin secretion (Gao et al., 2000; Gauthier et al., 2008). It is reported that GLP-1 stimulates PKA-dependent phosphorylation of Syt7 at Ser103. Such modification enhances Ca2+-triggered exocytosis, whereas dephosphorylation of Syt7 disrupts GLP-1 potentiation of insulin secretion (Wu et al., 2015). Besides cytoplasmic Ca2+ balance, calcium homeostasis in the ER also has a pivotal role in insulin secretion (Sabourin et al., 2015; Li et al., 2021). Ryanodine receptors (RyRs) are vital components of ER intracellular calcium channels which mediate Ca2+ release from ER into the cytoplasm (Doser et al., 2020). RyR2 is one of the RyRs isoforms that is expressed in β cells (Johnson et al., 2004; Dror et al., 2008; Takasawa et al., 2010). It has been shown that sufficient phosphorylation of RyR2 and subsequent Ca2+ release are essential steps in GSIS (Dixit et al., 2013; Llanos et al., 2015). However, hyperphosphorylation of RyR2 induced by CaMKII leads to glucose intolerance, impaired GSIS and lowered [(Ca2+)]cyt transients. This is due to the increased basal RyR2-mediated Ca2+ leak and basal hyperinsulinemia (Dixit et al., 2013). Moreover, components of SOCs can be phosphorylated. TRPC1, as a component of SOCs, can be phosphorylated by protein kinase C (PKC). Inhibition of PKC activity reduces TRPC1 phosphorylation and decreases insulin secretion rapidly, which could be restored by the TRPC1 activator (Xu et al., 2019).
Insulin granules transport and release also can be regulated by protein phosphorylation (Ho et al., 2020). Myosin is a motor protein that is responsible for insulin granules transportation dependent on F-actin (Varadi et al., 2005; Sweeney and Holzbaur, 2018; Omar-Hmeadi and Idevall-Hagren, 2021). It is composed of heavy chains (MHC) and two types of light chains (MLC) (Penn et al., 1982; Niki et al., 1993). Both MHC and MLC can be phosphorylated by multiple protein kinase including Myosin light-chain kinase (MLCK), and promote insulin secretion (Tan et al., 1992; Iida et al., 1997; Wilson et al., 1999). Tau is a microtubule-associated protein (MAP) involved with insulin secretion and glycemic control (Maj et al., 2016; Wijesekara et al., 2018). High glucose induces hyper-phosphorylation of tau by multiple kinases including GSK3, PKA, PKC, and CDK5, which enhances microtubule turnover to acutely induces GSIS (Ho et al., 2020). β-catenin, as a cell-cell adhesion protein, increases GSIS through promoting the rearrangement of actin cytoskeleton (Sorrenson et al., 2016). It can be phosphorylated at Ser552 by p21-activated protein kinase-1 (PAK1) in response to glucose and GLP-1 stimulation. A mutation of Ser552 to Ala on β-catenin attenuates GSIS, suggesting a critical role for β-catenin Ser552 phosphorylation in insulin secretion (Sorrenson et al., 2021). Tomosyn is a negative regulator of cellular exocytosis and can hinder insulin secretion (Zhang et al., 2006; Bhatnagar et al., 2011). In response to high glucose, enhanced tomosyn-2 phosphorylation targets tomosyn-2 for Hrd-1-mediated ubiquitination and degradation, causing increased insulin secretion (Bhatnagar et al., 2014). Activated Phospholipase D2 (PLD2) can stimulate insulin secretion (Jones et al., 1999). PLD2 phosphorylation at Ser134 via cell cycle protein-dependent kinase 5 (CDK5) plays an influential role in EGF-dependent insulin secretion (Lee et al., 2008). Brain-selective kinase 2 (BRSK2), also known as SAD-A, is a member of the AMPK-related kinase family. It is abundantly expressed in pancreatic islets β cells and acts a key function in insulin secretion (Lizcano et al., 2004). Overexpression of SAD-A significantly enhances GSIS and further potentiates GLP-1’s effect on GSIS in isolated mouse islets (Nie et al., 2013). Phosphorylation of PAK1 at Thr423 via SAD-A triggers the onset of GSIS in islet β cells (Nie et al., 2012). Phosphorylation of GDP-dissociation inhibitors (GDIα) at Ser174 by SAD-A leads to the dissociation of Rho GTPases from GDIα complexes, culminating in insulin exocytosis (Nie et al., 2018).
Phosphorylation also presents an inhibitory role in insulin secretion. Fas-associated death domain protein (FADD) is a classical adaptor in Fas-FasL signaling, which can regulate islet mass and insulin secretion. In the mouse model of FADD-D (S191D), which mimics the constitutive expression of phosphorylated FADD in mice, the area of pancreatic islets is shrunken, and GSIS is impaired. This suggests that FADD phosphorylation negatively regulates islet development and insulin secretion (Yao et al., 2015).
3.2 Acetylation
Acetylation is a chemical reaction in which an acetyl group is added to a compound in place of a hydrogen atom, which can be regulated by histone acetyltransferases (HATs) and histone deacetylases (HDACs) (Gao et al., 2017). Acetylation not only regulates histones, transcription factors, and epigenetic regulators, but also regulates many enzymes in metabolic pathways such as glycolysis, gluconeogenesis, tricarboxylic acid cycle, and fatty acid oxidation (Guan and Xiong, 2011).
Protein acetylation is tightly linked to insulin secretion and functional regulation of pancreatic β cells (Zhang et al., 2019). As a class of highly conserved deacetylases, Sirtuins (SIRTs) significantly contribute to insulin secretion. Among the SIRT family, SIRT1 is located in the nucleus, and ameliorates hyperglycemia by promoting insulin secretion and β cell expansion (Luu et al., 2013). SIRT2 is the only predominantly cytoplasmic isoform but is also found in the nucleus and mitochondria (Vaquero et al., 2006; Liu et al., 2017). SIRT2 knockout rats exhibit impaired glucose tolerance and decreased GSIS (Moynihan et al., 2005). SIRT3 and SIRT4 are mainly localized in the mitochondria. Overexpression of SIRT3 inhibits acetylation and degradation of trifunctional enzyme subunit alpha (ECHA) that participated in fatty acid β-oxidation, resulting in increased β-oxidation of fatty acid and reduced oxidation of glucose in β cells. SIRT3 knockout mice show increased insulin secretion upon glucose stimulation (Zhang et al., 2019). SIRT4 regulates amino acid catabolism and insulin secretion, maintaining glucose homeostasis during aging (Haigis et al., 2006; Anderson et al., 2017). SIRT6 is mainly localized in the nucleus. Pancreatic β cell-specific SIRT6 knockout mice show significantly reduced GSIS (Xiong et al., 2016).
Most known acetylated proteins contributing to insulin secretion are transcription factors. Pancreatic and duodenal homeobox-1 (PDX-1) and FOXO1 are two transcription factors associated with insulin gene transcription (Sussel et al., 1998; Berneman-Zeitouni et al., 2014). The acetylated form of FOXO1 resides in the cytoplasm while the deacetylated form is mainly in the nucleus. Deacetylation of FOXO1 enhances its transcriptional activity and plays an essential role in insulin signaling (Accili and Arden, 2004). Moreover, insulin secretion is enhanced in deacetylated FOXO1 (6 KR) knock-in mice (Kim-Muller et al., 2016). lncRNA MALAT1 impairs insulin secretion by reducing histone H3 acetylation at the Pdx-1 promoter and subsequently inhibiting Pdx-1 expression (Ding et al., 2020).
3.3 Ubiquitination
Ubiquitination is regulated by a three-step cascade: ubiquitin activation by E1 enzymes, conjugation by E2 enzymes, and ligation by E3 ligases (Komander and Rape, 2012). The Ubiquitin-proteasome system (UPS) is the main pathway for intracellular protein degradation. It plays a crucial role in the regulation of glucose homeostasis, insulin secretion, and the pathogenesis of diabetes mellitus (Price et al., 1996; Kawaguchi et al., 2006; Song et al., 2013; Uruno et al., 2013; Al-Khalili et al., 2014; Yang et al., 2016; Balaji et al., 2018; Yamada et al., 2018).
A group of critical proteins involved in insulin secretion is regulated by ubiquitination-mediated degradation. GK degradation by ubiquitination inhibits insulin secretion by decreasing G-6-P production (Cho et al., 2020). The proteasome inhibitor lactacystin has been shown to enhance GSIS in a 2-h short-term treatment (Lopez-Avalos et al., 2006). The v-maf musculoaponeurotic fibrosarcoma oncogene homolog A (MafA) is a key transcription factor required for β cells formation and function (Zhang et al., 2005; Andrali et al., 2008; Kaneto and Matsuoka, 2015). MafA deletion in mice causes glucose intolerance and induces diabetes. The E3 ubiquitin ligase Hrd-1 targets MafA for ubiquitination and degradation in β cells, which results in cytoplasmic accumulation of MafA. Such accumulation impairs its function in the nucleus, causing reduced insulin secretion (Wu et al., 2020). CREB is a transcription factor that is essential for glucose homeostasis and β cell survival (Jambal et al., 2003; Jhala et al., 2003; Sarkar et al., 2007). Chronic hyperglycemia increases CREB ubiquitination and decreases protein expression which ultimately inhibits insulin secretion (Costes et al., 2009). Somatostatin receptor subtype 5 (SSTR5) inhibits PDX-1 expression by downregulating Pdx-1 transcription and enhancing PDX-1 ubiquitination at post-translational level, thus reducing insulin secretion (Zhou et al., 2012). Phosphorylation of PDX-1 at Thr11 by Macrophage Stimulating 1 (MST1) in β cells leads to PDX-1 ubiquitination and degradation, resulting in impaired insulin secretion (Ardestani et al., 2014). A previous report showed that silencing CDK5 activator CDK5R1 (also known as p35) enhances insulin secretion in MIN6 cells in high glucose culture (Wei et al., 2005). Subsequent research showed that p35 phosphorylation via Salt-inducible kinase 2 (SIK2) induces p35 ubiquitylation. The modification is mediated by the E3 ubiquitin ligase PJA2, which leads to the activation of calcium entry and insulin secretion (Sakamaki et al., 2014).
3.4 SUMOylation
SUMOylation is a post-translational modification of small SUMO proteins that is catalyzed by activating enzyme (E1), conjugating enzyme (E2), and ligase (E3), and reversed by specific proteases such as Sentrin-specific SUMO proteases (SENPs) (Li et al., 2005; Flotho and Melchior, 2013). It plays an important role in modulating protein activity, protein-protein interactions, and subcellular localization (Shao and Cobb, 2009; Gareau and Lima, 2010). It has been reported that the homeostasis of SUMOylation plays an essential role in maintaining β cell function (Dai et al., 2011; Rajan et al., 2012; Vergari et al., 2012; Davey et al., 2019).
E2 SUMO-conjugating enzyme (UBC9) is the only conjugating enzyme essential for the SUMO system. Mice lacking UBC9 in β cells exhibit decreased insulin content and loss of β cell mass. In contrast, overexpression of UBC9 in β cells leads to an increased antioxidant ability but impaired insulin secretion (He et al., 2018). Protein deSUMOylation by SENPs regulates the conjugation/deconjugation balance of target proteins. Studies on islet-specific SENP1 deletion in mice further demonstrated that the knockdown of SENP1 reduces Ca2+-triggered β cell exocytosis. Conversely, overexpression of SENP1 augments β cell exocytosis (Vergari et al., 2012). It suggests a key role for SUMOylation/deSUMOylation balance in GSIS.
SUMOylation regulates insulin secretion at multiple stages. In the insulin gene transcription process, SUMOylation reduced transcriptional activity of MafA toward the insulin gene promoter in low glucose (2 mm) or exposure to hydrogen peroxide (Shao and Cobb, 2009). However, SUMOylation protects PDX-1 from proteasomal degradation and promotes its entry into the nucleus, where it activates insulin gene transcription (Kishi et al., 2003). SUMOylation also participates in triggering insulin secretion pathway. Overexpression of SUMO-1 in β cells increases the stability and activity of GK to induce the closure of the KATP channel (Aukrust et al., 2013), meanwhile exert a strong inhibitory action on the Kv2.1 voltage-dependent K+ channel (MacDonald et al., 2001; MacDonald et al., 2002; Dai et al., 2009). The depolarization of cell membrane promotes the activation of calcium channels and subsequent insulin secretion. GLP-1 activates the GLP-1 signaling by the interaction with GLP1 receptor (GLP-1R) in β cells, resulting in a rapid increase in intracellular cAMP that promotes insulin secretion. Overexpression of SUMO-1 attenuates GLP-1R function by preventing GLP-1R oligomerization which is essential for forward trafficking, leading to a significant reduction in insulin secretion (Rajan et al., 2012). SUMOylation has been proposed to act as a ‘brake’ on insulin exocytosis. Some proteins associated with insulin exocytosis have been reported to be SUMOylated such as tomosyn1A and syntaxin 1A. SUMOylation increases the interaction between tomosyn1A and syntaxin 1A, which sequesters syntaxin 1A to inhibits the formation of SNARE complex and subsequent insulin exocytosis (Ferdaoussi et al., 2017; Davey et al., 2019). Moreover, SUMOylation suppresses Syt7-mediated insulin secretion, which is transiently lost upon glucose stimulation and returns within 30–60 min (Dai et al., 2011).
3.5 O-GlcNAcylation
O-GlcNAcylation is a unique protein glycosylation that is controlled bidirectionally by two enzymes: the writer O-GlcNAc transferase (OGT) and the eraser O-GlcNAcase (OGA). As a nutrition sensor, O-GlcNAcylation is closely associated with type 2 diabetes and associated complications (Dias and Hart, 2007).
O-GlcNAc homeostasis in β cells plays a notable role in insulin secretion (Sireesh et al., 2014; Humphrey et al., 2015; Wende, 2015; Yang and Qian, 2017; Yoshida et al., 2022). OGT is expressed virtually in all cell types but is particularly high in pancreatic β cells (Hanover et al., 1999; Hart and Akimoto, 2009). β cell-specific OGT deletion leads to reduced insulin secretion, and this effect is more pronounced in high fat diet-fed mice (Lockridge et al., 2020). OGA acts in opposition to OGT to regulate protein O-GlcNAcylation. Research has shown that overexpression of OGA in β cells decreases insulin secretion and impairs glucose tolerance in mice (Soesanto et al., 2011). Paradoxically, OGA deletion in pancreatic β cells also impairs insulin secretion in vivo and in vitro (Yoshida et al., 2022).
Currently, most identified substrates of O-GlcNAcylation are related to insulin gene transcription. Increased nuclear O-GlcNAcylation increases intracellular insulin levels and reserves GSIS in part by boosting histone H3 transcriptional activation to promote Ins1 and Ins2 gene transcription in Min6 cells (Durning et al., 2016). Glucose induced PDX-1 O-GlcNAcylation leads to increased PDX-1 DNA binding activity and insulin secretion in Min6 cells (Gao et al., 2003). Further studies demonstrated that OGT interacts with phosphatidylinositol 3,4,5-trisphosphate (PIP3), which enables OGT to catalyze O-GlcNAcylation of nuclear proteins, including PDX-1 (Yang et al., 2008; Kebede et al., 2012). The localization of NeuroD1, a transcription factor of the insulin gene, is regulated by O-GlcNAcylation (Peterson and Hart, 2016). Under low glucose conditions, NeuroD1 is mainly in the cytosol. However, OGA inhibitor treatment induces NeuroD1 translocation into the nucleus, leading to enhanced insulin expression (Andrali et al., 2007). Another transcription factor, myocyte enhancer factor 2D (Mef2d), has been reported to negatively regulate insulin secretion through O-GlcNAcylation (Yoshida et al., 2022).
3.6 Palmitoylation
Palmitoylation, the attachment of fatty acyl chains to cysteine residues, is a reversible process mediated by the opposing activities of acyltransferases and thioesterases (Chamberlain and Shipston, 2015). Palmitoylation is a recently discovered PTM that plays a significant role in the regulation of cellular functions (Blaskovic et al., 2014; Chamberlain and Shipston, 2015). Recent studies have revealed the emerging importance of palmitoylation in insulin secretion and insulin response pathways (Chamberlain et al., 2021; Dong et al., 2023). Acyl-protein thioesterase 1 (APT1) is a depalmitoylation enzyme. The function of APT1 was blocked in chronic hyperglycemia, leading to defective insulin secretion (Chamberlain et al., 2021). APT1 knockdown in islets caused insulin hypersecretion (Dong et al., 2023).
In β-cells, palmitoylation are widely present in ion channels and insulin exocytosis. Kir6.2 is palmitoylated at cysteine-166, which increase the open state of the KATP channel and lead to inhibitory insulin secretion (Chamberlain et al., 2021). β2a, an important auxiliary subunit of VDCCs, is palmitoylated at cysteine-3/4, thus increasing plasma membrane trafficking of β2a and Cav subunits of VDCCs. However, excessive number of palmitoylated CaVβ2a leads to Ca2+ overload and β cell death (Kazim et al., 2017). Palmitoylation of SNAP-25 in the central region increase its membrane localization, which may help insulin exocytosis (Gonelle-Gispert et al., 2000). Scamp1, which is localized in insulin secretory granules, can be palmitoylated at Cys132. Palmitoylation-defective Scamp1 mutant C132S rescues insulin hypersecretion and nutrient-induced apoptosis in APT1-deficient cells (Dong et al., 2023). GLP-1R is palmitoylated mainly at Cys438 in response to agonists, inducing GLP-1R clustering, nanodomain signaling, and internalization. These result in enhanced GLP-1R signaling and insulin secretion (Buenaventura et al., 2019).
3.7 Other PTMs
Besides the PTM types described above, a group of less-studied PTM types is also related to insulin secretion. Citrullination is mediated by calcium-dependent peptidyl arginine deiminases (PADs), which catalyze deimination, the conversion of arginine into non-classical amino acid citrulline (Kunieda et al., 2018). A major role for PADs and citrullinated proteins has emerged in type 1 diabetes (Yang et al., 2021). Citrullination alters the enzyme kinetics of GK and suppresses GSIS (Yang et al., 2022).
Deamidation is the conversion of glutamine to glutamic acid by transglutaminase (TGM) enzymes (Callebaut et al., 2022). Deamidated peptides have been reported as autoantigens in type 1 diabetes (Callebaut et al., 2022). Islet amyloid polypeptide (IAPP) accelerates the pathogenesis of type 2 diabetes by exacerbating β cell degeneration and ultimately compromising insulin secretion. Studies have revealed that deamidation can modulate IAPP amyloid formation and fibril morphology, which induces its cytotoxicity (Nguyen et al., 2017).
4 Conclusion and perspectives
In this review, we summarized the current progress of PTM regulation in insulin secretion. Eight PTMs and at least twenty-nine modified protein substrates have been reported to be associated with insulin secretion, suggesting that insulin secretion is extensively regulated by PTMs. However, there remains numerous unanswered questions about the role of PTMs in insulin secretion. For example, currently more than 600 PTMs have been identified in eukaryotes (Bradley, 2022). Whether there are more PTMs and more modified substrates present in β cells? What’s the function? Furthermore, it is noticed that some key proteins are regulated by multiple PTMs with consistent or contrary effect. What’s the crosstalk between the different PTMs? How these PTMs synergistically regulate the function of protein? At last, defective or excessive PTMs may induce insulin deficiency, or insulin hypersecretion which may lead to β cells failure, resulting impaired insulin secretion. How the PTMs are dynamically regulated in the physiologic or pathological insulin secretion process? Moreover, several drug candidates for T2DM have been proved to regulate protein PTMs. For example, Glucokinase activators (GKAs), including dorzagliatin, MK-0941 and AZD1656, also promotes SUMOylation of pancreatic glucokinase, (Wu et al., 2023), implying PTM can be the potential drug targets for T2DM. Thus, further in-depth studies of PTMs in insulin secretion are in need.
Statements
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Accili D. Arden K. C. (2004). FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell117 (4), 421–426. 10.1016/s0092-8674(04)00452-0
2
Al-Khalili L. de Castro Barbosa T. Ostling J. Massart J. Cuesta P. G. Osler M. E. et al (2014). Proteasome inhibition in skeletal muscle cells unmasks metabolic derangements in type 2 diabetes. Am. J. Physiol. Cell Physiol.307 (9), C774–C787. 10.1152/ajpcell.00110.2014
3
Altarejos J. Y. Goebel N. Conkright M. D. Inoue H. Xie J. Arias C. M. et al (2008). The Creb1 coactivator Crtc1 is required for energy balance and fertility. Nat. Med.14 (10), 1112–1117. 10.1038/nm.1866
4
Altarejos J. Y. Montminy M. (2011). CREB and the CRTC co-activators: Sensors for hormonal and metabolic signals. Nat. Rev. Mol. Cell Biol.12 (3), 141–151. 10.1038/nrm3072
5
Anderson K. A. Huynh F. K. Fisher-Wellman K. Stuart J. D. Peterson B. S. Douros J. D. et al (2017). SIRT4 is a lysine deacylase that controls leucine metabolism and insulin secretion. Cell Metab.25 (4), 838–855.e15. 10.1016/j.cmet.2017.03.003
6
Andrali S. S. Qian Q. Ozcan S. (2007). Glucose mediates the translocation of NeuroD1 by O-linked glycosylation. J. Biol. Chem.282 (21), 15589–15596. 10.1074/jbc.M701762200
7
Andrali S. S. Sampley M. L. Vanderford N. L. Ozcan S. (2008). Glucose regulation of insulin gene expression in pancreatic beta-cells. Biochem. J.415 (1), 1–10. 10.1042/BJ20081029
8
Ardestani A. Paroni F. Azizi Z. Kaur S. Khobragade V. Yuan T. et al (2014). MST1 is a key regulator of beta cell apoptosis and dysfunction in diabetes. Nat. Med.20 (4), 385–397. 10.1038/nm.3482
9
Aukrust I. Bjorkhaug L. Negahdar M. Molnes J. Johansson B. B. Muller Y. et al (2013). SUMOylation of pancreatic glucokinase regulates its cellular stability and activity. J. Biol. Chem.288 (8), 5951–5962. 10.1074/jbc.M112.393769
10
Balaji V. Pokrzywa W. Hoppe T. (2018). Ubiquitylation pathways in insulin signaling and organismal homeostasis. Bioessays40 (5), e1700223. 10.1002/bies.201700223
11
Berneman-Zeitouni D. Molakandov K. Elgart M. Mor E. Fornoni A. Dominguez M. R. et al (2014). The temporal and hierarchical control of transcription factors-induced liver to pancreas transdifferentiation. PLoS One9 (2), e87812. 10.1371/journal.pone.0087812
12
Bhatnagar S. Oler A. T. Rabaglia M. E. Stapleton D. S. Schueler K. L. Truchan N. A. et al (2011). Positional cloning of a type 2 diabetes quantitative trait locus; tomosyn-2, a negative regulator of insulin secretion. PLoS Genet.7 (10), e1002323. 10.1371/journal.pgen.1002323
13
Bhatnagar S. Soni M. S. Wrighton L. S. Hebert A. S. Zhou A. S. Paul P. K. et al (2014). Phosphorylation and degradation of tomosyn-2 de-represses insulin secretion. J. Biol. Chem.289 (36), 25276–25286. 10.1074/jbc.M114.575985
14
Blaskovic S. Adibekian A. Blanc M. van der Goot G. F. (2014). Mechanistic effects of protein palmitoylation and the cellular consequences thereof. Chem. Phys. Lipids180, 44–52. 10.1016/j.chemphyslip.2014.02.001
15
Bracey K. M. Ho K. H. Yampolsky D. Gu G. Kaverina I. Holmes W. R. (2020). Microtubules regulate localization and availability of insulin granules in pancreatic beta cells. Biophys. J.118 (1), 193–206. 10.1016/j.bpj.2019.10.031
16
Bradley D. (2022). The evolution of post-translational modifications. Curr. Opin. Genet. Dev.76, 101956. 10.1016/j.gde.2022.101956
17
Buenaventura T. Bitsi S. Laughlin W. E. Burgoyne T. Lyu Z. Oqua A. I. et al (2019). Agonist-induced membrane nanodomain clustering drives GLP-1 receptor responses in pancreatic beta cells. PLoS Biol.17 (8), e3000097. 10.1371/journal.pbio.3000097
18
Cabrera O. Berman D. M. Kenyon N. S. Ricordi C. Berggren P. O. Caicedo A. (2006). The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. U. S. A.103 (7), 2334–2339. 10.1073/pnas.0510790103
19
Callebaut A. Bruggeman Y. Zamit C. Sodre F. M. C. Irla M. Mathieu C. et al (2022). Aberrant expression of transglutaminase 2 in pancreas and thymus of NOD mice underscores the importance of deamidation in neoantigen generation. Front. Endocrinol. (Lausanne)13, 908248. 10.3389/fendo.2022.908248
20
Campbell J. E. Newgard C. B. (2021). Mechanisms controlling pancreatic islet cell function in insulin secretion. Nat. Rev. Mol. Cell Biol.22 (2), 142–158. 10.1038/s41580-020-00317-7
21
Ch'ng T. H. Uzgil B. Lin P. Avliyakulov N. K. O'Dell T. J. Martin K. C. (2012). Activity-dependent transport of the transcriptional coactivator CRTC1 from synapse to nucleus. Cell150 (1), 207–221. 10.1016/j.cell.2012.05.027
22
Chamberlain L. H. Shipston M. J. Gould G. W. (2021). Regulatory effects of protein S-acylation on insulin secretion and insulin action. Open Biol.11 (3), 210017. 10.1098/rsob.210017
23
Chamberlain L. H. Shipston M. J. (2015). The physiology of protein S-acylation. Physiol. Rev.95 (2), 341–376. 10.1152/physrev.00032.2014
24
Chang T. J. Chen W. P. Yang C. Lu P. H. Liang Y. C. Su M. J. et al (2009). Serine-385 phosphorylation of inwardly rectifying K+ channel subunit (Kir6.2) by AMP-dependent protein kinase plays a key role in rosiglitazone-induced closure of the K(ATP) channel and insulin secretion in rats. Diabetologia52 (6), 1112–1121. 10.1007/s00125-009-1337-4
25
Cho J. Horikawa Y. Enya M. Takeda J. Imai Y. Imai Y. et al (2020). L-Arginine prevents cereblon-mediated ubiquitination of glucokinase and stimulates glucose-6-phosphate production in pancreatic beta-cells. Commun. Biol.3 (1), 497. 10.1038/s42003-020-01226-3
26
Costes S. Vandewalle B. Tourrel-Cuzin C. Broca C. Linck N. Bertrand G. et al (2009). Degradation of cAMP-responsive element-binding protein by the ubiquitin-proteasome pathway contributes to glucotoxicity in beta-cells and human pancreatic islets. Diabetes58 (5), 1105–1115. 10.2337/db08-0926
27
Cui J. Wang Z. Cheng Q. Lin R. Zhang X. M. Leung P. S. et al (2011). Targeted inactivation of kinesin-1 in pancreatic beta-cells in vivo leads to insulin secretory deficiency. Diabetes60 (1), 320–330. 10.2337/db09-1078
28
Da Silva Xavier G. (2018). The cells of the islets of langerhans. J. Clin. Med.7 (3), 54. 10.3390/jcm7030054
29
Dai X. Q. Kolic J. Marchi P. Sipione S. Macdonald P. E. (2009). SUMOylation regulates Kv2.1 and modulates pancreatic beta-cell excitability. J. Cell Sci.122 (Pt 6), 775–779. 10.1242/jcs.036632
30
Dai X. Q. Plummer G. Casimir M. Kang Y. Hajmrle C. Gaisano H. Y. et al (2011). SUMOylation regulates insulin exocytosis downstream of secretory granule docking in rodents and humans. Diabetes60 (3), 838–847. 10.2337/db10-0440
31
Dall'Agnese A. Platt J. M. Zheng M. M. Friesen M. Dall'Agnese G. Blaise A. M. et al (2022). The dynamic clustering of insulin receptor underlies its signaling and is disrupted in insulin resistance. Nat. Commun.13 (1), 7522. 10.1038/s41467-022-35176-7
32
Davey J. S. Carmichael R. E. Craig T. J. (2019). Protein SUMOylation regulates insulin secretion at multiple stages. Sci. Rep.9 (1), 2895. 10.1038/s41598-019-39681-6
33
DeFronzo R. A. Triplitt C. L. Abdul-Ghani M. Cersosimo E. (2014). Novel agents for the treatment of type 2 diabetes. Diabetes Spectr.27 (2), 100–112. 10.2337/diaspect.27.2.100
34
Dias W. B. Hart G. W. (2007). O-GlcNAc modification in diabetes and Alzheimer's disease. Mol. Biosyst.3 (11), 766–772. 10.1039/b704905f
35
Ding H. Wang F. Shi X. Ma H. Du Y. Hou L. et al (2020). LncRNA MALAT1 induces the dysfunction of beta cells via reducing the histone acetylation of the PDX-1 promoter in type 1 diabetes. Exp. Mol. Pathol.114, 104432. 10.1016/j.yexmp.2020.104432
36
Dixit S. S. Wang T. Manzano E. J. Yoo S. Lee J. Chiang D. Y. et al (2013). Effects of CaMKII-mediated phosphorylation of ryanodine receptor type 2 on islet calcium handling, insulin secretion, and glucose tolerance. PLoS One8 (3), e58655. 10.1371/journal.pone.0058655
37
Dong G. Adak S. Spyropoulos G. Zhang Q. Feng C. Yin L. et al (2023). Palmitoylation couples insulin hypersecretion with beta cell failure in diabetes. Cell Metab.35 (2), 332–344.e7. 10.1016/j.cmet.2022.12.012
38
Doser R. L. Amberg G. C. Hoerndli F. J. (2020). Reactive oxygen species modulate activity-dependent AMPA receptor transport in C. elegans. J. Neurosci.40 (39), 7405–7420. 10.1523/JNEUROSCI.0902-20.2020
39
Dror V. Kalynyak T. B. Bychkivska Y. Frey M. H. Tee M. Jeffrey K. D. et al (2008). Glucose and endoplasmic reticulum calcium channels regulate HIF-1beta via presenilin in pancreatic beta-cells. J. Biol. Chem.283 (15), 9909–9916. 10.1074/jbc.M710601200
40
Durning S. P. Flanagan-Steet H. Prasad N. Wells L. (2016). O-linked beta-N-acetylglucosamine (O-GlcNAc) acts as a glucose sensor to epigenetically regulate the insulin gene in pancreatic beta cells. J. Biol. Chem.291 (5), 2107–2118. 10.1074/jbc.M115.693580
41
Dyachok O. Idevall-Hagren O. Sagetorp J. Tian G. Wuttke A. Arrieumerlou C. et al (2008). Glucose-induced cyclic AMP oscillations regulate pulsatile insulin secretion. Cell Metab.8 (1), 26–37. 10.1016/j.cmet.2008.06.003
42
Ferdaoussi M. Fu J. Dai X. Manning Fox J. E. Suzuki K. Smith N. et al (2017). SUMOylation and calcium control syntaxin-1A and secretagogin sequestration by tomosyn to regulate insulin exocytosis in human ß cells. Sci. Rep.7 (1), 248. 10.1038/s41598-017-00344-z
43
Flotho A. Melchior F. (2013). Sumoylation: A regulatory protein modification in health and disease. Annu. Rev. Biochem.82, 357–385. 10.1146/annurev-biochem-061909-093311
44
Gao B. Kong Q. Zhang Y. Yun C. Dent S. Y. R. Song J. et al (2017). The histone acetyltransferase Gcn5 positively regulates T cell activation. J. Immunol.198 (10), 3927–3938. 10.4049/jimmunol.1600312
45
Gao Y. Miyazaki J. Hart G. W. (2003). The transcription factor PDX-1 is post-translationally modified by O-linked N-acetylglucosamine and this modification is correlated with its DNA binding activity and insulin secretion in min6 beta-cells. Arch. Biochem. Biophys.415 (2), 155–163. 10.1016/s0003-9861(03)00234-0
46
Gao Z. Reavey-Cantwell J. Young R. A. Jegier P. Wolf B. A. (2000). Synaptotagmin III/VII isoforms mediate Ca2+-induced insulin secretion in pancreatic islet beta -cells. J. Biol. Chem.275 (46), 36079–36085. 10.1074/jbc.M004284200
47
Gareau J. R. Lima C. D. (2010). The SUMO pathway: Emerging mechanisms that shape specificity, conjugation and recognition. Nat. Rev. Mol. Cell Biol.11 (12), 861–871. 10.1038/nrm3011
48
Gauthier B. R. Duhamel D. L. Iezzi M. Theander S. Saltel F. Fukuda M. et al (2008). Synaptotagmin VII splice variants alpha, beta, and delta are expressed in pancreatic beta-cells and regulate insulin exocytosis. FASEB J.22 (1), 194–206. 10.1096/fj.07-8333com
49
Gonelle-Gispert C. Molinete M. Halban P. A. Sadoul K. (2000). Membrane localization and biological activity of SNAP-25 cysteine mutants in insulin-secreting cells. J. Cell Sci.113 (Pt 18), 3197–3205. 10.1242/jcs.113.18.3197
50
Guan K. L. Xiong Y. (2011). Regulation of intermediary metabolism by protein acetylation. Trends Biochem. Sci.36 (2), 108–116. 10.1016/j.tibs.2010.09.003
51
Haigis M. C. Mostoslavsky R. Haigis K. M. Fahie K. Christodoulou D. C. Murphy A. J. et al (2006). SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell126 (5), 941–954. 10.1016/j.cell.2006.06.057
52
Hanover J. A. Lai Z. Lee G. Lubas W. A. Sato S. M. (1999). Elevated O-linked N-acetylglucosamine metabolism in pancreatic beta-cells. Arch. Biochem. Biophys.362 (1), 38–45. 10.1006/abbi.1998.1016
53
Hart G. W. Akimoto Y. (2009). “The O-GlcNAc modification,” in Essentials of glycobiology. Editors VarkiA.CummingsR. D.EskoJ. D.FreezeH. H.StanleyP.BertozziC. R.et al2nd ed (New York, NY: Cold Spring Harbor). (NY)).
54
He X. Lai Q. Chen C. Li N. Sun F. Huang W. et al (2018). Both conditional ablation and overexpression of E2 SUMO-conjugating enzyme (UBC9) in mouse pancreatic beta cells result in impaired beta cell function. Diabetologia61 (4), 881–895. 10.1007/s00125-017-4523-9
55
Henquin J. C. (2011). The dual control of insulin secretion by glucose involves triggering and amplifying pathways in beta-cells. Diabetes Res. Clin. Pract.93 (Suppl. 1), S27–S31. 10.1016/S0168-8227(11)70010-9
56
Ho K. H. Yang X. Osipovich A. B. Cabrera O. Hayashi M. L. Magnuson M. A. et al (2020). Glucose regulates microtubule disassembly and the dose of insulin secretion via tau phosphorylation. Diabetes69 (9), 1936–1947. 10.2337/db19-1186
57
Humphrey S. J. James D. E. Mann M. (2015). Protein phosphorylation: A major switch mechanism for metabolic regulation. Trends Endocrinol. Metab.26 (12), 676–687. 10.1016/j.tem.2015.09.013
58
Iida Y. Senda T. Matsukawa Y. Onoda K. Miyazaki J. I. Sakaguchi H. et al (1997). Myosin light-chain phosphorylation controls insulin secretion at a proximal step in the secretory cascade. Am. J. Physiol.273 (4), E782–E789. 10.1152/ajpendo.1997.273.4.E782
59
Jambal P. Masterson S. Nesterova A. Bouchard R. Bergman B. Hutton J. C. et al (2003). Cytokine-mediated down-regulation of the transcription factor cAMP-response element-binding protein in pancreatic beta-cells. J. Biol. Chem.278 (25), 23055–23065. 10.1074/jbc.M212450200
60
Jhala U. S. Canettieri G. Screaton R. A. Kulkarni R. N. Krajewski S. Reed J. et al (2003). cAMP promotes pancreatic beta-cell survival via CREB-mediated induction of IRS2. Genes Dev.17 (13), 1575–1580. 10.1101/gad.1097103
61
Johnson J. D. Kuang S. Misler S. Polonsky K. S. (2004). Ryanodine receptors in human pancreatic beta cells: Localization and effects on insulin secretion. FASEB J.18 (7), 878–880. 10.1096/fj.03-1280fje
62
Jones D. Morgan C. Cockcroft S. (1999). Phospholipase D and membrane traffic. Potential roles in regulated exocytosis, membrane delivery and vesicle budding. Biochim. Biophys. Acta1439 (2), 229–244. 10.1016/s1388-1981(99)00097-9
63
Kalwat M. A. Cobb M. H. (2017). Mechanisms of the amplifying pathway of insulin secretion in the beta cell. Pharmacol. Ther.179, 17–30. 10.1016/j.pharmthera.2017.05.003
64
Kaneto H. Matsuoka T. A. (2015). Role of pancreatic transcription factors in maintenance of mature beta-cell function. Int. J. Mol. Sci.16 (3), 6281–6297. 10.3390/ijms16036281
65
Kawaguchi M. Minami K. Nagashima K. Seino S. (2006). Essential role of ubiquitin-proteasome system in normal regulation of insulin secretion. J. Biol. Chem.281 (19), 13015–13020. 10.1074/jbc.M601228200
66
Kazim A. S. Storm P. Zhang E. Renstrom E. (2017). Palmitoylation of Ca2+ channel subunit CaVβ2a induces pancreatic beta-cell toxicity via Ca2+ overload. Biochem. Biophys. Res. Commun.491 (3), 740–746. 10.1016/j.bbrc.2017.07.117
67
Kebede M. Ferdaoussi M. Mancini A. Alquier T. Kulkarni R. N. Walker M. D. et al (2012). Glucose activates free fatty acid receptor 1 gene transcription via phosphatidylinositol-3-kinase-dependent O-GlcNAcylation of pancreas-duodenum homeobox-1. Proc. Natl. Acad. Sci. U. S. A.109 (7), 2376–2381. 10.1073/pnas.1114350109
68
Khan F. A. Pandupuspitasari N. S. Huang C. J. Hao X. Zhang S. (2016). SUMOylation: A link to future therapeutics. Curr. Issues Mol. Biol.18, 49–56. 10.21775/cimb.018.049
69
Kim-Muller J. Y. Kim Y. J. Fan J. Zhao S. Banks A. S. Prentki M. et al (2016). FoxO1 deacetylation decreases fatty acid oxidation in beta-cells and sustains insulin secretion in diabetes. J. Biol. Chem.291 (19), 10162–10172. 10.1074/jbc.M115.705608
70
Kishi A. Nakamura T. Nishio Y. Maegawa H. Kashiwagi A. (2003). Sumoylation of Pdx1 is associated with its nuclear localization and insulin gene activation. Am. J. Physiol. Endocrinol. Metab.284 (4), E830–E840. 10.1152/ajpendo.00390.2002
71
Knoch K. P. Bergert H. Borgonovo B. Saeger H. D. Altkruger A. Verkade P. et al (2004). Polypyrimidine tract-binding protein promotes insulin secretory granule biogenesis. Nat. Cell Biol.6 (3), 207–214. 10.1038/ncb1099
72
Knoch K. P. Meisterfeld R. Kersting S. Bergert H. Altkruger A. Wegbrod C. et al (2006). cAMP-dependent phosphorylation of PTB1 promotes the expression of insulin secretory granule proteins in beta cells. Cell Metab.3 (2), 123–134. 10.1016/j.cmet.2005.12.008
73
Komander D. Rape M. (2012). The ubiquitin code. Annu. Rev. Biochem.81, 203–229. 10.1146/annurev-biochem-060310-170328
74
Kunieda K. Yamauchi H. Kawaguchi M. Ieda N. Nakagawa H. (2018). Development of a fluorescent probe for detection of citrulline based on photo-induced electron transfer. Bioorg Med. Chem. Lett.28 (5), 969–973. 10.1016/j.bmcl.2018.01.026
75
Lee H. Y. Jung H. Jang I. H. Suh P. G. Ryu S. H. (2008). Cdk5 phosphorylates PLD2 to mediate EGF-dependent insulin secretion. Cell Signal20 (10), 1787–1794. 10.1016/j.cellsig.2008.06.009
76
Li M. Guo D. Isales C. M. Eizirik D. L. Atkinson M. She J. X. et al (2005). SUMO wrestling with type 1 diabetes. J. Mol. Med. Berl.83 (7), 504–513. 10.1007/s00109-005-0645-5
77
Li M. Shao F. Qian Q. Yu W. Zhang Z. Chen B. et al (2021). A putative long noncoding RNA-encoded micropeptide maintains cellular homeostasis in pancreatic beta cells. Mol. Ther. Nucleic Acids26, 307–320. 10.1016/j.omtn.2021.06.027
78
Lim A. Park S. H. Sohn J. W. Jeon J. H. Park J. H. Song D. K. et al (2009). Glucose deprivation regulates KATP channel trafficking via AMP-activated protein kinase in pancreatic beta-cells. Diabetes58 (12), 2813–2819. 10.2337/db09-0600
79
Liu G. Park S. H. Imbesi M. Nathan W. J. Zou X. Zhu Y. et al (2017). Loss of NAD-dependent protein deacetylase sirtuin-2 alters mitochondrial protein acetylation and dysregulates mitophagy. Antioxid. Redox Signal26 (15), 849–863. 10.1089/ars.2016.6662
80
Lizcano J. M. Goransson O. Toth R. Deak M. Morrice N. A. Boudeau J. et al (2004). LKB1 is a master kinase that activates 13 kinases of the AMPK subfamily, including MARK/PAR-1. EMBO J.23 (4), 833–843. 10.1038/sj.emboj.7600110
81
Llanos P. Contreras-Ferrat A. Barrientos G. Valencia M. Mears D. Hidalgo C. (2015). Glucose-dependent insulin secretion in pancreatic beta-cell islets from male rats requires Ca2+ release via ROS-stimulated ryanodine receptors. PLoS One10 (6), e0129238. 10.1371/journal.pone.0129238
82
Lockridge A. Jo S. Gustafson E. Damberg N. Mohan R. Olson M. et al (2020). Islet O-GlcNAcylation is required for lipid potentiation of insulin secretion through SERCA2. Cell Rep.31 (5), 107609. 10.1016/j.celrep.2020.107609
83
Lopez J. J. Jardin I. Sanchez-Collado J. Salido G. M. Smani T. Rosado J. A. (2020). TRPC channels in the SOCE scenario. Cells9 (1), 126. 10.3390/cells9010126
84
Lopez-Avalos M. D. Duvivier-Kali V. F. Xu G. Bonner-Weir S. Sharma A. Weir G. C. (2006). Evidence for a role of the ubiquitin-proteasome pathway in pancreatic islets. Diabetes55 (5), 1223–1231. 10.2337/db05-0450
85
Luu L. Dai F. F. Prentice K. J. Huang X. Hardy A. B. Hansen J. B. et al (2013). The loss of Sirt1 in mouse pancreatic beta cells impairs insulin secretion by disrupting glucose sensing. Diabetologia56 (9), 2010–2020. 10.1007/s00125-013-2946-5
86
MacDonald P. E. Ha X. F. Wang J. Smukler S. R. Sun A. M. Gaisano H. Y. et al (2001). Members of the Kv1 and Kv2 voltage-dependent K(+) channel families regulate insulin secretion. Mol. Endocrinol.15 (8), 1423–1435. 10.1210/mend.15.8.0685
87
MacDonald P. E. Sewing S. Wang J. Joseph J. W. Smukler S. R. Sakellaropoulos G. et al (2002). Inhibition of Kv2.1 voltage-dependent K+ channels in pancreatic beta-cells enhances glucose-dependent insulin secretion. J. Biol. Chem.277 (47), 44938–44945. 10.1074/jbc.M205532200
88
Maj M. Hoermann G. Rasul S. Base W. Wagner L. Attems J. (2016). The microtubule-associated protein tau and its relevance for pancreatic beta cells. J. Diabetes Res.2016, 1964634. 10.1155/2016/1964634
89
Malm H. A. Mollet I. G. Berggreen C. Orho-Melander M. Esguerra J. L. Goransson O. et al (2016). Transcriptional regulation of the miR-212/miR-132 cluster in insulin-secreting beta-cells by cAMP-regulated transcriptional co-activator 1 and salt-inducible kinases. Mol. Cell Endocrinol.424, 23–33. 10.1016/j.mce.2016.01.010
90
Morales-Tarre O. Alonso-Bastida R. Arcos-Encarnacion B. Perez-Martinez L. Encarnacion-Guevara S. (2021). Protein lysine acetylation and its role in different human pathologies: A proteomic approach. Expert Rev. Proteomics18 (11), 949–975. 10.1080/14789450.2021.2007766
91
Mounier C. Posner B. I. (2006). Transcriptional regulation by insulin: From the receptor to the gene. Can. J. Physiol. Pharmacol.84 (7), 713–724. 10.1139/y05-152
92
Moynihan K. A. Grimm A. A. Plueger M. M. Bernal-Mizrachi E. Ford E. Cras-Meneur C. et al (2005). Increased dosage of mammalian Sir2 in pancreatic beta cells enhances glucose-stimulated insulin secretion in mice. Cell Metab.2 (2), 105–117. 10.1016/j.cmet.2005.07.001
93
Nguyen P. T. Zottig X. Sebastiao M. Bourgault S. (2017). Role of site-specific asparagine deamidation in islet amyloid polypeptide amyloidogenesis: Key contributions of residues 14 and 21. Biochemistry56 (29), 3808–3817. 10.1021/acs.biochem.7b00209
94
Nie J. Lilley B. N. Pan Y. A. Faruque O. Liu X. Zhang W. et al (2013). SAD-A potentiates glucose-stimulated insulin secretion as a mediator of glucagon-like peptide 1 response in pancreatic beta cells. Mol. Cell Biol.33 (13), 2527–2534. 10.1128/MCB.00285-13
95
Nie J. Sun C. Chang Z. J. Musi N. Shi Y. G. (2018). SAD-A promotes glucose-stimulated insulin secretion through phosphorylation and inhibition of GDIα in male islet β cells. Endocrinology159 (8), 3036–3047. 10.1210/en.2017-03243
96
Nie J. Sun C. Faruque O. Ye G. Li J. Liang Q. et al (2012). Synapses of amphids defective (SAD-A) kinase promotes glucose-stimulated insulin secretion through activation of p21-activated kinase (PAK1) in pancreatic beta-Cells. J. Biol. Chem.287 (31), 26435–26444. 10.1074/jbc.M112.378372
97
Niki I. Okazaki K. Saitoh M. Niki A. Niki H. Tamagawa T. et al (1993). Presence and possible involvement of Ca/calmodulin-dependent protein kinases in insulin release from the rat pancreatic beta cell. Biochem. Biophys. Res. Commun.191 (1), 255–261. 10.1006/bbrc.1993.1210
98
Nolan C. J. Madiraju M. S. Delghingaro-Augusto V. Peyot M. L. Prentki M. (2006). Fatty acid signaling in the beta-cell and insulin secretion. Diabetes55 (Suppl. 2), S16–S23. 10.2337/db06-s003
99
Oetjen E. Diedrich T. Eggers A. Eckert B. Knepel W. (1994). Distinct properties of the cAMP-responsive element of the rat insulin I gene. J. Biol. Chem.269 (43), 27036–27044. 10.1016/s0021-9258(18)47122-0
100
Omar-Hmeadi M. Idevall-Hagren O. (2021). Insulin granule biogenesis and exocytosis. Cell Mol. Life Sci.78 (5), 1957–1970. 10.1007/s00018-020-03688-4
101
Ozcan S. Andrali S. S. Cantrell J. E. (2010). Modulation of transcription factor function by O-GlcNAc modification. Biochim. Biophys. Acta1799 (5-6), 353–364. 10.1016/j.bbagrm.2010.02.005
102
Penn E. J. Brocklehurst K. W. Sopwith A. M. Hales C. N. Hutton J. C. (1982). Ca2+--Calmodulin dependent myosin light-chain phosphorylating activity in insulin-secreting tissues. FEBS Lett.139 (1), 4–8. 10.1016/0014-5793(82)80474-2
103
Peterson S. B. Hart G. W. (2016). New insights: A role for O-GlcNAcylation in diabetic complications. Crit. Rev. Biochem. Mol. Biol.51 (3), 150–161. 10.3109/10409238.2015.1135102
104
Prakriya M. Lewis R. S. (2015). Store-operated calcium channels. Physiol. Rev.95 (4), 1383–1436. 10.1152/physrev.00020.2014
105
Price S. R. Bailey J. L. Wang X. Jurkovitz C. England B. K. Ding X. et al (1996). Muscle wasting in insulinopenic rats results from activation of the ATP-dependent, ubiquitin-proteasome proteolytic pathway by a mechanism including gene transcription. J. Clin. Invest.98 (8), 1703–1708. 10.1172/JCI118968
106
Rajan S. Torres J. Thompson M. S. Philipson L. H. (2012). SUMO downregulates GLP-1-stimulated cAMP generation and insulin secretion. Am. J. Physiol. Endocrinol. Metab.302 (6), E714–E723. 10.1152/ajpendo.00486.2011
107
Roder P. V. Wu B. Liu Y. Han W. (2016). Pancreatic regulation of glucose homeostasis. Exp. Mol. Med.48 (3), e219. 10.1038/emm.2016.6
108
Sabourin J. Le Gal L. Saurwein L. Haefliger J. A. Raddatz E. Allagnat F. (2015). Store-operated Ca2+ entry mediated by Orai1 and TRPC1 participates to insulin secretion in rat beta-cells. J. Biol. Chem.290 (51), 30530–30539. 10.1074/jbc.M115.682583
109
Sacco F. Humphrey S. J. Cox J. Mischnik M. Schulte A. Klabunde T. et al (2016). Glucose-regulated and drug-perturbed phosphoproteome reveals molecular mechanisms controlling insulin secretion. Nat. Commun.7, 13250. 10.1038/ncomms13250
110
Sakamaki J. Fu A. Reeks C. Baird S. Depatie C. Al Azzabi M. et al (2014). Role of the SIK2-p35-PJA2 complex in pancreatic beta-cell functional compensation. Nat. Cell Biol.16 (3), 234–244. 10.1038/ncb2919
111
Sampley M. L. Ozcan S. (2012). Regulation of insulin gene transcription by multiple histone acetyltransferases. DNA Cell Biol.31 (1), 8–14. 10.1089/dna.2011.1336
112
Sarkar S. A. Gunter J. Bouchard R. Reusch J. E. Wiseman A. Gill R. G. et al (2007). Dominant negative mutant forms of the cAMP response element binding protein induce apoptosis and decrease the anti-apoptotic action of growth factors in human islets. Diabetologia50 (8), 1649–1659. 10.1007/s00125-007-0707-z
113
Schwartz M. W. Seeley R. J. Tschop M. H. Woods S. C. Morton G. J. Myers M. G. et al (2013). Cooperation between brain and islet in glucose homeostasis and diabetes. Nature503 (7474), 59–66. 10.1038/nature12709
114
Seino S. (1999). ATP-Sensitive potassium channels: A model of heteromultimeric potassium channel/receptor assemblies. Annu. Rev. Physiol.61, 337–362. 10.1146/annurev.physiol.61.1.337
115
Shao C. Cobb M. H. (2009). Sumoylation regulates the transcriptional activity of MafA in pancreatic beta cells. J. Biol. Chem.284 (5), 3117–3124. 10.1074/jbc.M806286200
116
Shin S. Le Lay J. Everett L. J. Gupta R. Rafiq K. Kaestner K. H. (2014). CREB mediates the insulinotropic and anti-apoptotic effects of GLP-1 signaling in adult mouse beta-cells. Mol. Metab.3 (8), 803–812. 10.1016/j.molmet.2014.08.001
117
Sireesh D. Bhakkiyalakshmi E. Ramkumar K. M. Rathinakumar S. Jennifer P. S. Rajaguru P. et al (2014). Targeting SUMOylation cascade for diabetes management. Curr. Drug Targets15 (12), 1094–1106. 10.2174/1389450115666140915124747
118
Soesanto Y. Luo B. Parker G. Jones D. Cooksey R. C. McClain D. A. (2011). Pleiotropic and age-dependent effects of decreased protein modification by O-linked N-acetylglucosamine on pancreatic beta-cell function and vascularization. J. Biol. Chem.286 (29), 26118–26126. 10.1074/jbc.M111.249508
119
Song R. Peng W. Zhang Y. Lv F. Wu H. K. Guo J. et al (2013). Central role of E3 ubiquitin ligase MG53 in insulin resistance and metabolic disorders. Nature494 (7437), 375–379. 10.1038/nature11834
120
Sorrenson B. Cognard E. Lee K. L. Dissanayake W. C. Fu Y. Han W. et al (2016). A critical role for beta-catenin in modulating levels of insulin secretion from beta-cells by regulating actin cytoskeleton and insulin vesicle localization. J. Biol. Chem.291 (50), 25888–25900. 10.1074/jbc.M116.758516
121
Sorrenson B. Dissanayake W. C. Hu F. Lee K. L. Shepherd P. R. (2021). A role for PAK1 mediated phosphorylation of beta-catenin Ser552 in the regulation of insulin secretion. Biochem. J.478 (8), 1605–1615. 10.1042/BCJ20200862
122
Sussel L. Kalamaras J. Hartigan-O'Connor D. J. Meneses J. J. Pedersen R. A. Rubenstein J. L. et al (1998). Mice lacking the homeodomain transcription factor Nkx2.2 have diabetes due to arrested differentiation of pancreatic beta cells. Development125 (12), 2213–2221. 10.1242/dev.125.12.2213
123
Sweeney H. L. Holzbaur E. L. F. (2018). Motor proteins. Cold Spring Harb. Perspect. Biol.10 (5), a021931. 10.1101/cshperspect.a021931
124
Takasawa S. Kuroki M. Nata K. Noguchi N. Ikeda T. Yamauchi A. et al (2010). A novel ryanodine receptor expressed in pancreatic islets by alternative splicing from type 2 ryanodine receptor gene. Biochem. Biophys. Res. Commun.397 (2), 140–145. 10.1016/j.bbrc.2010.05.051
125
Tan J. L. Ravid S. Spudich J. A. (1992). Control of nonmuscle myosins by phosphorylation. Annu. Rev. Biochem.61, 721–759. 10.1146/annurev.bi.61.070192.003445
126
Thurmond D. C. Gonelle-Gispert C. Furukawa M. Halban P. A. Pessin J. E. (2003). Glucose-stimulated insulin secretion is coupled to the interaction of actin with the t-SNARE (target membrane soluble N-ethylmaleimide-sensitive factor attachment protein receptor protein) complex. Mol. Endocrinol.17 (4), 732–742. 10.1210/me.2002-0333
127
Tian G. Sagetorp J. Xu Y. Shuai H. Degerman E. Tengholm A. (2012a). Role of phosphodiesterases in the shaping of sub-plasma-membrane cAMP oscillations and pulsatile insulin secretion. J. Cell Sci.125 (Pt 21), 5084–5095. 10.1242/jcs.107201
128
Tian G. Sandler S. Gylfe E. Tengholm A. (2011). Glucose- and hormone-induced cAMP oscillations in α- and β-cells within intact pancreatic islets. Diabetes60 (5), 1535–1543. 10.2337/db10-1087
129
Tian G. Sol E. R. Xu Y. Shuai H. Tengholm A. (2015). Impaired cAMP generation contributes to defective glucose-stimulated insulin secretion after long-term exposure to palmitate. Diabetes64 (3), 904–915. 10.2337/db14-1036
130
Tian G. Tepikin A. V. Tengholm A. Gylfe E. (2012b). cAMP induces stromal interaction molecule 1 (STIM1) puncta but neither Orai1 protein clustering nor store-operated Ca2+ entry (SOCE) in islet cells. J. Biol. Chem.287 (13), 9862–9872. 10.1074/jbc.M111.292854
131
Uruno A. Furusawa Y. Yagishita Y. Fukutomi T. Muramatsu H. Negishi T. et al (2013). The Keap1-Nrf2 system prevents onset of diabetes mellitus. Mol. Cell Biol.33 (15), 2996–3010. 10.1128/MCB.00225-13
132
Vaquero A. Scher M. B. Lee D. H. Sutton A. Cheng H. L. Alt F. W. et al (2006). SirT2 is a histone deacetylase with preference for histone H4 Lys 16 during mitosis. Genes Dev.20 (10), 1256–1261. 10.1101/gad.1412706
133
Varadi A. Tsuboi T. Johnson-Cadwell L. I. Allan V. J. Rutter G. A. (2003). Kinesin I and cytoplasmic dynein orchestrate glucose-stimulated insulin-containing vesicle movements in clonal MIN6 beta-cells. Biochem. Biophys. Res. Commun.311 (2), 272–282. 10.1016/j.bbrc.2003.09.208
134
Varadi A. Tsuboi T. Rutter G. A. (2005). Myosin Va transports dense core secretory vesicles in pancreatic MIN6 beta-cells. Mol. Biol. Cell16 (6), 2670–2680. 10.1091/mbc.e04-11-1001
135
Vergari E. Plummer G. Dai X. MacDonald P. E. (2012). DeSUMOylation controls insulin exocytosis in response to metabolic signals. Biomolecules2 (2), 269–281. 10.3390/biom2020269
136
Walsh C. T. Garneau-Tsodikova S. Gatto G. J. Jr. (2005). Protein posttranslational modifications: The chemistry of proteome diversifications. Angew. Chem. Int. Ed. Engl.44 (45), 7342–7372. 10.1002/anie.200501023
137
Walsh G. Jefferis R. (2006). Post-translational modifications in the context of therapeutic proteins. Nat. Biotechnol.24 (10), 1241–1252. 10.1038/nbt1252
138
Wang B. Lin H. Li X. Lu W. Kim J. B. Xu A. et al (2020). The adaptor protein APPL2 controls glucose-stimulated insulin secretion via F-actin remodeling in pancreatic beta-cells. Proc. Natl. Acad. Sci. U. S. A.117 (45), 28307–28315. 10.1073/pnas.2016997117
139
Wei F. Y. Nagashima K. Ohshima T. Saheki Y. Lu Y. F. Matsushita M. et al (2005). Cdk5-dependent regulation of glucose-stimulated insulin secretion. Nat. Med.11 (10), 1104–1108. 10.1038/nm1299
140
Wende A. R. (2015). Unsticking the broken diabetic heart: O-GlcNAcylation and calcium sensitivity. Diabetes64 (10), 3339–3341. 10.2337/dbi15-0001
141
Wijesekara N. Goncalves R. A. Ahrens R. De Felice F. G. Fraser P. E. (2018). Tau ablation in mice leads to pancreatic beta cell dysfunction and glucose intolerance. FASEB J.32 (6), 3166–3173. 10.1096/fj.201701352
142
Wilson J. R. Biden T. J. Ludowyke R. I. (1999). Increases in phosphorylation of the myosin II heavy chain, but not regulatory light chains, correlate with insulin secretion in rat pancreatic islets and RINm5F cells. Diabetes48 (12), 2383–2389. 10.2337/diabetes.48.12.2383
143
Wu B. Wei S. Petersen N. Ali Y. Wang X. Bacaj T. et al (2015). Synaptotagmin-7 phosphorylation mediates GLP-1-dependent potentiation of insulin secretion from beta-cells. Proc. Natl. Acad. Sci. U. S. A.112 (32), 9996–10001. 10.1073/pnas.1513004112
144
Wu T. Zhang S. Xu J. Zhang Y. Sun T. Shao Y. et al (2020). HRD1, an important player in pancreatic beta-cell failure and therapeutic target for type 2 diabetic mice. Diabetes69 (5), 940–953. 10.2337/db19-1060
145
Wu X. Xu M. Geng M. Chen S. Little P. J. Xu S. et al (2023). Targeting protein modifications in metabolic diseases: Molecular mechanisms and targeted therapies. Signal Transduct. Target Ther.8 (1), 220. 10.1038/s41392-023-01439-y
146
Xiong X. Wang G. Tao R. Wu P. Kono T. Li K. et al (2016). Sirtuin 6 regulates glucose-stimulated insulin secretion in mouse pancreatic beta cells. Diabetologia59 (1), 151–160. 10.1007/s00125-015-3778-2
147
Xu J. Zhang W. Cui W. Shi B. Wang H. (2019). PKCα promotes insulin secretion via TRPC1 phosphorylation in INS-1E cells. Biosci. Biotechnol. Biochem.83 (9), 1676–1682. 10.1080/09168451.2019.1617106
148
Yamada T. Murata D. Adachi Y. Itoh K. Kameoka S. Igarashi A. et al (2018). Mitochondrial stasis reveals p62-mediated ubiquitination in parkin-independent mitophagy and mitigates nonalcoholic fatty liver disease. Cell Metab.28 (4), 588–604.e5. 10.1016/j.cmet.2018.06.014
149
Yang M. L. Horstman S. Gee R. Guyer P. Lam T. T. Kanyo J. et al (2022). Citrullination of glucokinase is linked to autoimmune diabetes. Nat. Commun.13 (1), 1870. 10.1038/s41467-022-29512-0
150
Yang M. L. Sodre F. M. C. Mamula M. J. Overbergh L. (2021). Citrullination and PAD enzyme Biology in type 1 diabetes - regulators of inflammation, autoimmunity, and pathology. Front. Immunol.12, 678953. 10.3389/fimmu.2021.678953
151
Yang X. D. Xiang D. X. Yang Y. Y. (2016). Role of E3 ubiquitin ligases in insulin resistance. Diabetes Obes. Metab.18 (8), 747–754. 10.1111/dom.12677
152
Yang X. Ongusaha P. P. Miles P. D. Havstad J. C. Zhang F. So W. V. et al (2008). Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature451 (7181), 964–969. 10.1038/nature06668
153
Yang X. Qian K. (2017). Protein O-GlcNAcylation: Emerging mechanisms and functions. Nat. Rev. Mol. Cell Biol.18 (7), 452–465. 10.1038/nrm.2017.22
154
Yao C. Zhuang H. Cheng W. Lin Y. Du P. Yang B. et al (2015). FADD phosphorylation impaired islet morphology and function. J. Cell Physiol.230 (7), 1448–1456. 10.1002/jcp.24885
155
Yoshida M. Yokoi N. Takahashi H. Hatano N. Hayami T. Ogawa W. et al (2022). O-GlcNAcylation of myocyte-specific enhancer factor 2D negatively regulates insulin secretion from pancreatic beta-cells. Biochem. Biophys. Res. Commun.605, 90–96. 10.1016/j.bbrc.2022.03.036
156
Zhang C. Moriguchi T. Kajihara M. Esaki R. Harada A. Shimohata H. et al (2005). MafA is a key regulator of glucose-stimulated insulin secretion. Mol. Cell Biol.25 (12), 4969–4976. 10.1128/MCB.25.12.4969-4976.2005
157
Zhang W. Lilja L. Mandic S. A. Gromada J. Smidt K. Janson J. et al (2006). Tomosyn is expressed in beta-cells and negatively regulates insulin exocytosis. Diabetes55 (3), 574–581. 10.2337/diabetes.55.03.06.db05-0015
158
Zhang Y. Zhou F. Bai M. Liu Y. Zhang L. Zhu Q. et al (2019). The pivotal role of protein acetylation in linking glucose and fatty acid metabolism to beta-cell function. Cell Death Dis.10 (2), 66. 10.1038/s41419-019-1349-z
159
Zhou G. Liu S. H. Shahi K. M. Wang H. Duan X. Lin X. et al (2012). Negative regulation of pancreatic and duodenal homeobox-1 by somatostatin receptor subtype 5. Mol. Endocrinol.26 (7), 1225–1234. 10.1210/me.2012-1095
160
Zhu Y. Hart G. W. (2021). Targeting O-GlcNAcylation to develop novel therapeutics. Mol. Asp. Med.79, 100885. 10.1016/j.mam.2020.100885
Summary
Keywords
PTMs, phosphorylation, acetylation, ubiquitination, sumoylation, O-GlcNAcylation, palmitoylation, insulin secretion
Citation
Yang C, Wei M, Zhao Y, Yang Z, Song M, Mi J, Yang X and Tian G (2023) Regulation of insulin secretion by the post-translational modifications. Front. Cell Dev. Biol. 11:1217189. doi: 10.3389/fcell.2023.1217189
Received
05 May 2023
Accepted
24 July 2023
Published
04 August 2023
Volume
11 - 2023
Edited by
Venkateswarlu Kanamarlapudi, Swansea University Medical School, United Kingdom
Reviewed by
Hui-Ying Lim, University of Oklahoma Health Sciences Center, United States
Rohit Mahar, Hemwati Nandan Bahuguna Garhwal University, India
Suresh Kumar Mohankumar, Swansea University Medical School, United Kingdom
Updates
Copyright
© 2023 Yang, Wei, Zhao, Yang, Song, Mi, Yang and Tian.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Geng Tian, tiangeng@bzmc.edu.cn; Xiaoyong Yang, xiaoyong.yang@yale.edu
†These authors have contributed equally to this work and share first authorship
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.