 Laura Streit1
Laura Streit1 Emeline Tanguy1
Emeline Tanguy1 Laurent Brunaud2
Laurent Brunaud2 Petra Tóth1
Petra Tóth1 Nicolas Vitale1
Nicolas Vitale1 Stéphane Ory1
Stéphane Ory1 Stéphane Gasman1*
Stéphane Gasman1*- 1Centre National de la Recherche Scientifique, Université de Strasbourg, Institut des Neurosciences Cellulaires et Intégratives, Strasbourg, France
- 2Département de Chirurgie Viscérale, Métabolique et Cancérologique (CVMC), INSERM NGERE-U1256, Université de Lorraine, CHRU Nancy, Hôpital Brabois adultes, Vandoeuvre-lès-Nancy, France
Neuroendocrine tumors (NETs) constitute a heterogeneous group of neoplasms arising from hormone-releasing cells. Secretion of hormones stored in vesicles occurs through calcium-regulated exocytosis, a process that needs to be tightly controlled to avoid unbalanced levels of hormones. A critical feature shared by most of the NETs is a dysfunctional secretory pathway mainly leading to hypersecretion, which often induces clinical complications. In this review, we focus on the cellular process of hormone exocytosis and discuss the potential molecular mechanisms leading to deregulated hormone secretion in various NETs. Particular attention is paid to expression level modifications for genes and proteins involved in the exocytic pathway in NETs.
1 Introduction
Neuroendocrine tumors (NETs) are derived from neuroendocrine cells that control pleiotropic physiological functions by releasing various hormones and neuropeptides. NETs constitute a highly heterogeneous group of neoplasms in terms of function and morphology, mainly because neuroendocrine cells are spread all over the body as dispersed cells (diffuse neuroendocrine system) or concentrated in organs (Figure 1A). However, one of the most common physiological features of these NET family members is the dysfunction of hormone and neuropeptide secretion, leading to hypersecretion and eventually to clinical symptoms. For example, acromegaly often results from excessive secretion of growth hormone by pituitary adenoma (Dineen et al., 2017). Hypersecretion of serotonin by carcinoid tumors from the gastro-intestinal tract can trigger carcinoid syndrome, which is associated with flushing, diarrhea, bronchoconstriction and cardiac valvular disease (Onaitis et al., 2000). Excessive levels of circulating catecholamines in patients with pheochromocytoma, a NET derived from the adrenal medulla chromaffin cells, induce hypertension potentially leading to cardiomyopathies and stroke (Zhang et al., 2017; Y-Hassan and Falhammar, 2020). Moreover, enhanced secretory activity of NET cells may develop over time with negative impact on prognosis. For instance, a silent pituitary adenoma may progress into an actively secreting form, while a non-functional pancreatic tumor can start secreting excessive levels of hormones, thus evolving to a more aggressive tumor phenotype (Brown et al., 2006; Daems et al., 2009). In small cell lung cancer (SCLC), a high-grade malignant cancer, the progressive neuroendocrine nature of cancer cells enhances secretion of a variety of neuropeptides, together with growth factors that significantly accelerate the invasive growth by their autocrine action. Dysfunction of hormone secretion in NETs has been known from the clinical point of view for a long time, but the cellular and molecular mechanisms disturbing secretory pathways in tumor cells are rarely explored and remain poorly understood.
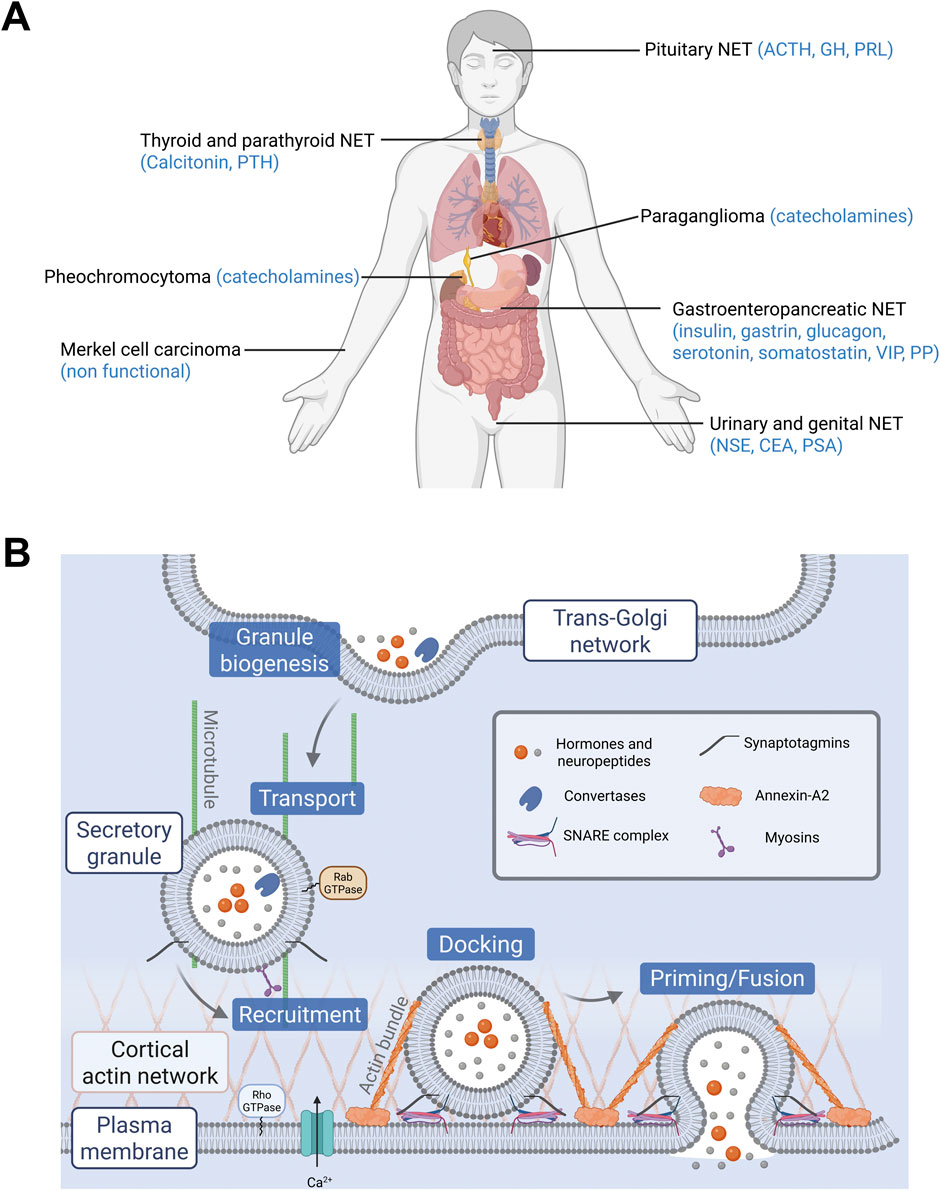
Figure 1. (A) Localization of neuroendocrine tumors (NETs) and their main secreted hormones. Hormones abbreviations: ACTH, adrenocorticotropic hormone; CEA, carcinoembryonic antigen; GH, growth hormone; NSE, neuron specific enolase; PRL, prolactin; PP, pancreatic polypeptide; PSA, prostate specific antigen; PTH, parathyroid hormone; VIP, vasoactive intestinal peptide. (B) Main steps of exocytosis and key proteins involved. Granules containing hormones and neuropeptides are formed at the trans-Golgi network by budding. They are then transported to the cell periphery and recruited to the plasma membrane. Their docking and priming prepare them for the step of fusion. Finally, the granules release their contents following calcium stimulation.
The first idea that usually comes in mind is that the increased level of secreted molecules results from an increased number of hormone-secreting cells within the tumor. However, the simplistic view of “the more secreting cells, the more secreted hormones” fails to consider alternative mechanisms. For example, in human pheochromocytoma, we recently reported using carbon fiber amperometry that individual tumor cells release significantly higher amounts of catecholamines compared to non-tumor chromaffin cells (Houy et al., 2022). The obvious conclusion from these observations is that the process of hormone release itself may be dysregulated in NET cells.
Hormone and neuropeptide secretion by neuroendocrine cells occurs through calcium-regulated exocytosis, a process that has been the subject of extensive studies over the past 3 decades (Anantharam and Kreutzberger, 2019). In these cells, hormones are synthesized and then stored in large dense core vesicles (LDCV) also known as secretory granules. Exocytosis is triggered by extracellular stimuli that increase intracellular calcium level and involves several tightly regulated steps including, i) the recruitment of the secretory granules to the cell periphery, ii) their docking to exocytic sites, which physically attaches them to the plasma membrane, iii) their priming, which prepares the granules for fusion, and finally iv) the fusion of the secretory granule membrane with the plasma membrane, leading to the release of luminal content of secretory granules (Figure 1B). The key questions arising from the observation of hypersecretion at the single cell level are which specific steps of the exocytic pathway are dysregulated in NETs and which proteins might be involved in this amplified secretory activity? Most of the limited data available today comes from “omics” approaches, which reveal changes in gene and protein expression across different NETs. These data are primarily from NETs derived from adrenal medulla, thyroid and pituitary gland. In this review, we focus on studies that investigate gene or protein expression changes specifically in human tumor biopsies. From this literature, we summarize the expression of key proteins involved in distinct steps of the hormone secretion pathway. Table 1 lists these main proteins and genes, highlighting their function in exocytosis and their expression changes in NETs. In the following sections, we examine each step of the hormone secretory pathway, from granule formation and cargo loading to vesicle fusion and membrane remodeling, highlighting the alterations observed in human NET samples.
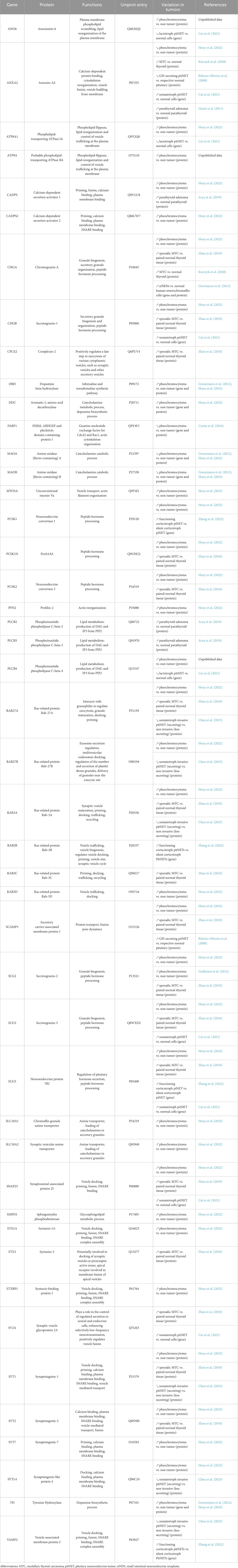
Table 1. List of up- and downregulated proteins or genes in neuroendocrine tumors.
2 Secretory granules biogenesis and maturation
Biogenesis of secretory granules begins at the trans-Golgi network, where prohormones and granin-family proteins (e.g., chromogranins and secretogranins) are selectively sorted and packaged into immature granules (Kim et al., 2006). These granules then undergo a maturation process involving luminal acidification, condensation of their content, proteolytic processing of prohormones, and removal of non-regulated proteins via clathrin-coated vesicles (Tooze et al., 1991; Arvan and Castle, 1998; Kim et al., 2006; Dembla and Becherer, 2021). During this maturation phase, small-molecule hormones such as catecholamines and serotonin are actively transported, from the cytosol into the granules, by vesicular monoamine transporters (VMATs). This uptake is powered by the proton gradients generated by V-ATPases. In parallel, peptide hormones are proteolytically processed within granules by enzymes such as the prohormone convertases PC1/3 and PC2, assisted by the chaperones like secretogranin V (Morvan and Tooze, 2008; Ma et al., 2021). Together, these steps generate mature granules that are competent for stimulus-dependent exocytosis.
The increase in single cell hormone secretion in NETs could arise from the dysregulation of various step of granule biogenesis and maturation. First, it could be the consequence of secretory granules contents overloading. Several studies reported upregulation of mRNAs or proteins involved in hormone synthesis (Table 1). For example, enzymes involved in catecholamines synthesis (Tyrosine Hydroxylase (TH), Dopamine-β-Hydroxylase (DBH), DOPA decarboxylase (DCC)) are overexpressed in pheochromocytoma (Jarrott and Louis, 1977; Isobe et al., 1998; Eisenhofer et al., 2008; Houy et al., 2022). Conversely, reduced expression of enzymes catabolizing catecholamines may further contribute to increase their levels. Accordingly, the monoamine oxydases (MAO-A/B), which deaminate cytosolic catecholamines, are downregulated in pheochromocytoma potentially leading to increased cytoplasmic catecholamine level (Grouzmann et al., 2012; Houy et al., 2022). Finally, expression of transporters such as VMAT1/2 (SLC18A1/A2) responsible for monoamines loading, is also increased (Table 1; Houy et al., 2022). It is, however, currently unknown whether increasing the number of hormone transporter molecules per granule actually leads to a higher hormone content, as there is a physical constraint beyond which additional filling is not possible.
Second, modulating the expression level of prohormone-processing enzymes that convert prohormones into their active forms can also lead to excessive active hormone production in NETs. This is often the case, for example, with neuroendocrine convertases (PCSK-1/2/1N), which are found upregulated in pheochromocytoma (Houy et al., 2022), medullary thyroid carcinoma (Zhan et al., 2018) and pituitary tumor (Cui et al., 2021; Zhang et al., 2022). Interestingly, secretogranin V, a chaperone for prohormone convertase is also overexpressed in various NETs (Table 1).
Third, the overexpression of secretory granule-resident proteins could reflect an increase in the number of secretory granules per cell. Notably, various chromogranin proteins (CHGA/B, SCG2/3; Table 1) or chromogranin-derived peptides, which are key proteins required for granule biogenesis (Kim et al., 2001; Beuret et al., 2004; Elias et al., 2012; Carmon et al., 2020) have also been found to be overexpressed in various NETs (Guillemot et al., 2006; Guerin et al., 2010; Zhan et al., 2018; Kurczyk et al., 2020; Cui et al., 2021; Houy et al., 2022). Whether this reflects an increase of the granin content per granule or an increase of the number of granules per cell or both requires further investigations.
3 Docking, priming and fusion
Our work in pheochromocytoma cells demonstrated that both the number of exocytic events and the kinetic of secretion were enhanced suggesting that late phases of exocytosis (docking, priming and/or fusion) could be modulated (Houy et al., 2022). The core machinery controlling docking, priming and fusion involves SNARE (Soluble N-ethylmaleimide sensitive factor attachment protein receptor) proteins, as well as SNARE-regulating proteins. Fusion between the secretory granule and plasma membrane is energetically unfavorable and requires the assembly of a membrane-bridging complex formed by the SNAREs (Jahn et al., 2024). In most neuroendocrine cells, the SNARE complex mediating the regulated exocytosis of LDCVs typically include SNAP25, Syntaxin1, and VAMP2. The number of SNARE complexes formed at the site of vesicle docking is correlated with the likelihood and speed of vesicle fusion (Mohrmann et al., 2010). Therefore, increasing the number of SNARE proteins can lead to more SNARE complexes, potentially enhancing the exocytosis rate. Accordingly, SNAP25 as well as various Syntaxins and VAMPs are upregulated in several NETs (Table 1) (Lu et al., 2008; Zhan et al., 2018; Cui et al., 2021; Houy et al., 2022; Zhang et al., 2022; Chen et al., 2023).
The assembly of the SNARE complex is not calcium sensitive, even though calcium is the main trigger of exocytosis. Synaptotagmins (Syts), a family of transmembrane vesicular proteins, confer Ca2+ sensitivity to membrane fusion in neuroendocrine cells by sensing changes in intracellular Ca2+ over a wide dynamic range (Pinheiro et al., 2016). Upon Ca2+ binding, synaptotagmins promote SNARE-mediated fusion by lowering the energy barrier primarily through the binding of their C2-domains to anionic phospholipids in the target membrane, which induces local positive curvature and membrane apposition to facilitate membrane merging. Among Syt family members, Syt1 and Syt7 have been implicated in neuroendocrine secretion (Schonn et al., 2008; Mohrmann et al., 2013; Bendahmane et al., 2020; Tawfik et al., 2021). Syt1 is a low-affinity, fast-responding isoform that mediate synchronous exocytosis, whereas Syt7 has higher calcium affinity and contributes to sustained or asynchronous release. Interestingly, Syt 1, 2 or 7 are upregulated in pheochromocytoma, medullary thyroid carcinoma and in somatotroph pituitary tumor (Table 1) (Zhan et al., 2018; Houy et al., 2022; Chen et al., 2023).
Altogether these observations suggest that hormone release might be enhanced either by increasing the number of SNARE complexes, or by modulating calcium sensitivity. Alternatively, the efficiency of exocytosis could also be influenced by proteins that tightly regulate the assembly or disassembly of the SNARE complex such as proteins from Sec1/Munc18-1 family, or even by tethering factors such as Rab GTPases that mediate the initial contact between the secretory granule and the plasma membrane (Baker and Hughson, 2016). For example, syntaxin binding protein 1 (STXBP1, commonly named Munc18-1) is overexpressed in pheochromocytoma (Houy et al., 2022), whereas expression level of the GTPases Rab3a-c or Rab27a-b and granuphilin (sytl4) is modified in pheochromocytoma, medullary thyroid carcinoma and in somatotroph or corticotroph pituitary tumor (Table 1), (Zhan et al., 2018; Houy et al., 2022; Zhang et al., 2022; Chen et al., 2023).
4 Actin cytoskeleton remodelling
In addition to the essential and minimal core machinery mediating secretory granule recruitment, docking, priming and fusion, an important extra-layer of regulation finely tunes calcium-regulated exocytosis, and consequently, the amount of hormone released. Notably, the functional importance of the actin cytoskeleton in regulated exocytosis has been recognized for several decades. Early studies revealed a dense cortical actin layer beneath the plasma membrane in secretory cells such as pancreatic β-cells and adrenal chromaffin cells (Orci et al., 1972; Trifaro et al., 1985; Aunis and Bader, 1988). These observations initially led to the view that cortical actin served primarily as a physical barrier restricting granule access to fusion sites. Accordingly, numerous studies demonstrated that remodeling of this cortical actin barrier facilitates secretory granules transport and fusion during calcium-regulated exocytosis in a variety of secretory cell models (Koffer et al., 1990; Vitale et al., 1991; Vitale et al., 1995; Gasman et al., 1997; Lang et al., 2000; Giner et al., 2007; Kalwat et al., 2013; Uenishi et al., 2013). Subsequent investigations, also demonstrated that actin filaments not only exert inhibitory control but can also play facilitatory roles, depending on the specific step of secretory granule exocytic process. This evolving concept has been discussed in several comprehensive review articles (Trifaro et al., 2008; Papadopulos, 2017; Li et al., 2018; Wu and Chan, 2022). Therefore, actin reorganization is now recognized as a dynamic and tightly regulated process that both constrains and facilitates exocytosis, depending on context and timing.
Members of the Rho GTPases family regulate exocytosis through actin reorganization. Seminal work from our team has demonstrated that the GTPases RhoA and Cdc42 play key regulatory roles on catecholamine release from adrenal chromaffin cells, by differentially influencing actin organization (Gasman et al., 1999; Malacombe et al., 2006a; Momboisse et al., 2011). RhoA has been proposed to actively control the organization of the cortical actin network, which regulates granule positioning and their access to the plasma membrane. In contrast Cdc42 promotes de novo actin nucleation and polymerization at the secretory granule fusion sites, likely controlling the final step of exocytosis such as membrane fusion (Malacombe et al., 2006b; Bretou et al., 2014). Interestingly, we previously showed that Cdc42 activity is inhibited in human pheochromocytoma, suggesting that actin reorganization at the exocytic sites may be affected (Croise et al., 2016; Croise et al., 2017). Although Cdc42 expression remains unchanged compared to non-tumor tissue, we have found that its activity is directly correlated to reduced expression of FARP1, a guanine nucleotide-exchange factor (GEF), that activates Cdc42 (Croise et al., 2016). Moreover, the expression of several GEFs for Rho-GTPases members is modulated in various NETs (for details, see Table 2 in (Streit et al., 2020)). However, whether these changes in Rho-GTPases activity are directly linked to altered hormone release in NETs has not been thoroughly explored.
Annexin-A2 is another important protein that regulates hormone release, and organizes the actin cytoskeleton (Figure 1B). Its expression is altered in various cancers including NETs (Christensen et al., 2018). In adrenal chromaffin cells, annexin-A2 promotes the formation of actin bundles required for efficient secretory granule docking and fusion (Gabel et al., 2015). Changes in annexin-A2 expression have been documented in several NETs, though with some differences. For instance, expression of annexin-A2 is decreased in pheochromocytoma (Houy et al., 2022) and pituitary adenoma (Ribeiro-Oliveira et al., 2008; Cui et al., 2021), but increased in medullary thyroid carcinoma (Kurczyk et al., 2020) and in parathyroid adenoma (Giusti et al., 2011). It would be valuable to study the relationship between the bundling activity of annexin-A2, its expression level and the secretory activity of NET cells. How these actin bundles participate to the exocytic process requires further investigations.
Actin cables may serve to tether, displace secretory granules at the correct location or provide forces to deform membranes. To do so, actin filaments need molecular motors such as myosin proteins (Figure 1B). Myosin II, V and VI constitute the main myosins involved in hormone release by neuroendocrine cells (Papadopulos et al., 2013; Gutierrez and Villanueva, 2018). We found that expression of unconventional myosin-Va is increased in pheochromocytoma (Houy et al., 2022). Interestingly cancer-associated mutations in the MYO5B gene have been described in pheochromocytoma and paraganglioma (Wilzen et al., 2016). Moreover, metastatic pheochromocytoma displays significantly higher expression of Myo5B compared to non-metastatic tumor (Tomic et al., 2020). To our knowledge, no data regarding the expression of Myosins II and VI in human NETs samples has been documented so far.
5 Lipid modifying enzymes
Lipids, with their vast diversity encompassing tens of thousands of species, are essential to cellular function, serving structural, metabolic, and signaling roles (van Meer et al., 2008). Early studies demonstrated that the synthesis, metabolism or transport of various lipids species play a fundamental role in hormone secretion by altering membrane biophysical properties and modulating the activity proteins that governs each step of vesicle transport and fusion; for review see (Tanguy et al., 2016; Gasman and Vitale, 2017). Among these lipids, phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P2) (Martin, 2012), certain fatty acids (Darios et al., 2007), anionic lipid such as phosphatidic acid (PA) (Tanguy et al., 2020), or cholesterol (Lang, 2007) regulate the activity or recruitment at exocytic sites of more than twenty proteins involved in regulated exocytosis (Koch and Holt, 2012). These targets include synaptotagmin (Schiavo et al., 1996), syntaxin (Lam et al., 2008), Ca2+-dependent activator protein for secretion (CAPS) (Loyet et al., 1998), and actin-binding proteins such as scinderin and gelsolin (Trifaro et al., 2008).
Although comprehensive lipidomics for NETs are still lacking, transcriptomic and proteomic studies revealed dysregulation of multiple lipid-metabolic enzymes and lipid transporters, whose activity can influence hormone secretion. For instance, phospholipase C enzymes, which hydrolyze (PtdIns(4,5)P2) to generate diacylglycerol (DAG) and inositol 3-phosphate, are upregulated in NET subtypes. PLC-β2 and -β3 isoforms are overexpressed in parathyroid adenomas, whereas PLC-β4 expression is elevated in pheochromocytoma and in pituitary NETs, suggesting an imbalance in these lipids in tumor cells (Bauer et al., 2007; Arya et al., 2019; Cui et al., 2021). In pheochromocytoma the sphingomyelin phosphodiesterase, which catalyzes the conversion of sphingomyelin into ceramide and phosphatidylcholine is overexpressed, increasing the pool of phosphatidylcholine potentially available to phospholipase-D1 and thereby potentially augmenting PA-driven exocytosis (Vitale et al., 2001; Zeniou-Meyer et al., 2007; Tanguy et al., 2020).
Hormone secretion is further modulated by the asymmetric distribution of phospholipids across the plasma membrane bilayer. Notably, we have shown that calcium-regulated exocytosis requires a transient disruption of the plasma membrane asymmetry near vesicle fusion sites in adrenal chromaffin cells (Ory et al., 2013). This process is regulated by scramblases, which catalyze the bidirectional transport of phospholipids between plasma membrane leaflets. Interestingly, several transbilayer lipid transporters are overexpressed in pituitary NETs and pheochromocytoma, including the scramblase ANO6 and the P4-type ATPases ATP8A1 (Cui et al., 2021; Houy et al., 2022) and ATP9A (unpublished).
Overall, lipid metabolism and transport appear to be altered in several NETs, which could directly affect hormone secretion. However, further investigations are needed to understand how changes in plasma membrane lipid composition and asymmetry precisely affect exocytosis efficiency.
6 Conclusion
Neuroendocrine tumors (NETs) represent a unique class of neoplasms characterized by their aberrant and unregulated hormone secretion, leading to significant clinical consequences. While our understanding of the molecular and cellular mechanisms underlying this hypersecretion is still evolving, recent advances, particularly through “omics” technologies, have begun to shed light on key alterations within the secretory pathway. Current data suggest that disruptions in hormone metabolism and secretory granule trafficking, as well as changes in expression of the regulatory proteins involved in this process, might contribute to the dysfunctional hormone release observed in NETs. Despite this progress, significant gaps in knowledge remain and the next challenge will be to better understand the precise mechanisms by which NET cells acquire such hypersecretory phenotypes. Over the past decade, “omics” technologies have advanced tremendously, enabling extremely fine levels of analysis.
These data, along with the future “omics” research, will be crucial in paving the way for novel therapeutic strategies aimed at targeting the secretory machinery to alleviate the clinical burden of NETs. From a clinical perspective, hormone hypersecretion in NETs is managed through various strategies designed to reduce hormone levels, control tumor growth, and relieve symptoms. Core treatments include somatostatin analogs (SSA), which inhibit hormone release by targeting somatostatin receptors, and peptide receptor radionuclide therapy (PRRT), which delivers targeted radiotherapy to receptor-expressing tumor cells. Surgical resection remains the treatment of choice when feasible, while chemotherapy and targeted agents such as everolimus or sunitinib are used in more advanced stages. Numerous in-depth reviews have covered these therapeutic options extensively (Stueven et al., 2019; Das et al., 2021; Zappi et al., 2023; Faggiano, 2024), including specifically in the NET types discussed in this review, which show significant changes in the expression of proteins controlling exocytosis, such as pheochromocytoma, medullary thyroid carcinoma, and pituitary adenomas (Varlamov et al., 2019; Kim and Kim, 2021; Bihain et al., 2022; Petersenn et al., 2023; Sharma and Fishbein, 2023; Casey et al., 2024). However, these treatments rarely target the secretory pathway itself, and their efficacy can diminish over time. By focusing on the molecular mechanisms of hormone secretion, this review complements the clinical literature and highlights the potential of directly targeting secretory machinery to reduce hormone burden in patients with persistent or refractory symptoms. Finally, despite the availability of several biochemical and imaging biomarkers such as Chromogranin A (CgA), neuron-specific enolase (NSE), and fluorodeoxyglucose positron emission tomography (FDG-PET), the diagnosis of NETs remains challenging. For instance, CgA is the most widely used serum marker, but its limited specificity and frequent false negatives reduce its diagnostic value (Nobels et al., 1997; Nehar et al., 2004). Identifying changes in the expression of secretory granule cargo proteins will certainly help the discovery of new accurate biomarkers.
Author contributions
LS: Conceptualization, Formal Analysis, Investigation, Writing – review and editing. ET: Writing – review and editing. LB: Resources, Writing – review and editing. PT: Writing – review and editing. NV: Writing – review and editing. SO: Formal Analysis, Investigation, Methodology, Supervision, Validation, Writing – review and editing. SG: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Part of the author’s work performed in pheochromocytoma and discussed here has been supported by grants from ITMO Cancer AVIESAN (Alliance Nationale pour les Sciences de la Vie et de la Santé, National Alliance for Life Sciences & Health) within the framework of the Cancer Plan to SG and LB (Single Cell 2018 N◦ 19CS004-00); by the University of Strasbourg Institute for Advanced Study (USIAS) for a Fellowship, within the French national programme “Investment for the future” (IdEx-Unistra) and by Ligue Contre le Cancer (CCIR-GE) to SG.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anantharam, A., and Kreutzberger, A. J. B. (2019). Unraveling the mechanisms of calcium-dependent secretion. J. Gen. Physiol. 151, 417–434. doi:10.1085/jgp.201812298
Arvan, P., and Castle, D. (1998). Sorting and storage during secretory granule biogenesis: looking backward and looking forward. Biochem. J. 332 (Pt 3), 593–610. doi:10.1042/bj3320593
Arya, A. K., Bhadada, S. K., Singh, P., Dahiya, D., Kaur, G., Sharma, S., et al. (2019). Quantitative proteomics analysis of sporadic parathyroid adenoma tissue samples. J. Endocrinol. Invest 42, 577–590. doi:10.1007/s40618-018-0958-1
Aunis, D., and Bader, M. F. (1988). The cytoskeleton as a barrier to exocytosis in secretory cells. J. Exp. Biol. 139, 253–266. doi:10.1242/jeb.139.1.253
Baker, R. W., and Hughson, F. M. (2016). Chaperoning SNARE assembly and disassembly. Nat. Rev. Mol. Cell Biol. 17, 465–479. doi:10.1038/nrm.2016.65
Bauer, C. S., Woolley, R. J., Teschemacher, A. G., and Seward, E. P. (2007). Potentiation of exocytosis by phospholipase C-coupled G-protein-coupled receptors requires the priming protein Munc13-1. J. Neurosci. 27, 212–219. doi:10.1523/JNEUROSCI.4201-06.2007
Bendahmane, M., Morales, A., Kreutzberger, A. J. B., Schenk, N. A., Mohan, R., Bakshi, S., et al. (2020). Synaptotagmin-7 enhances calcium-sensing of chromaffin cell granules and slows discharge of granule cargos. J. Neurochem. 154, 598–617. doi:10.1111/jnc.14986
Beuret, N., Stettler, H., Renold, A., Rutishauser, J., and Spiess, M. (2004). Expression of regulated secretory proteins is sufficient to generate granule-like structures in constitutively secreting cells. J. Biol. Chem. 279, 20242–20249. doi:10.1074/jbc.M310613200
Bihain, F., Nomine-Criqui, C., Guerci, P., Gasman, S., Klein, M., and Brunaud, L. (2022). Management of patients with treatment of pheochromocytoma: a critical appraisal. Cancers (Basel) 14, 3845. doi:10.3390/cancers14163845
Bretou, M., Jouannot, O., Fanget, I., Pierobon, P., Larochette, N., Gestraud, P., et al. (2014). Cdc42 controls the dilation of the exocytotic fusion pore by regulating membrane tension. Mol. Biol. Cell 25, 3195–3209. doi:10.1091/mbc.E14-07-1229
Brown, R. L., Muzzafar, T., Wollman, R., and Weiss, R. E. (2006). A pituitary carcinoma secreting TSH and prolactin: a non-secreting adenoma gone awry. Eur. J. Endocrinol. 154, 639–643. doi:10.1530/eje.1.02141
Carmon, O., Laguerre, F., Riachy, L., Delestre-Delacour, C., Wang, Q., Tanguy, E., et al. (2020). Chromogranin A preferential interaction with Golgi phosphatidic acid induces membrane deformation and contributes to secretory granule biogenesis. FASEB J. 34, 6769–6790. doi:10.1096/fj.202000074R
Casey, R. T., Hendriks, E., Deal, C., Waguespack, S. G., Wiegering, V., Redlich, A., et al. (2024). International consensus statement on the diagnosis and management of phaeochromocytoma and paraganglioma in children and adolescents. Nat. Rev. Endocrinol. 20, 729–748. doi:10.1038/s41574-024-01024-5
Chen, M., Duan, L., Sun, W., Guo, Z., Miao, H., Yu, N., et al. (2023). Clinical and proteomic-based molecular characterizations of invasive and noninvasive somatotroph PitNETs. Neuroendocrinology 113, 971–986. doi:10.1159/000531200
Christensen, M. V., Hogdall, C. K., Jochumsen, K. M., and Hogdall, E. V. S. (2018). Annexin A2 and cancer: a systematic review. Int. J. Oncol. 52, 5–18. doi:10.3892/ijo.2017.4197
Croise, P., Brunaud, L., Toth, P., Gasman, S., and Ory, S. (2017). Inhibition of Cdc42 and Rac1 activities in pheochromocytoma, the adrenal medulla tumor. Small GTPases 8, 122–127. doi:10.1080/21541248.2016.1202634
Croise, P., Houy, S., Gand, M., Lanoix, J., Calco, V., Toth, P., et al. (2016). Cdc42 and Rac1 activity is reduced in human pheochromocytoma and correlates with FARP1 and ARHGEF1 expression. Endocr. Relat. Cancer 23, 281–293. doi:10.1530/ERC-15-0502
Cui, Y., Li, C., Jiang, Z., Zhang, S., Li, Q., Liu, X., et al. (2021). Single-cell transcriptome and genome analyses of pituitary neuroendocrine tumors. Neuro Oncol. 23, 1859–1871. doi:10.1093/neuonc/noab102
Daems, T., Verhelst, J., Michotte, A., Abrams, P., De Ridder, D., and Abs, R. (2009). Modification of hormonal secretion in clinically silent pituitary adenomas. Pituitary 12, 80–86. doi:10.1007/s11102-008-0085-7
Darios, F., Connell, E., and Davletov, B. (2007). Phospholipases and fatty acid signalling in exocytosis. J. Physiol. 585, 699–704. doi:10.1113/jphysiol.2007.136812
Das, S., Al-Toubah, T., and Strosberg, J. (2021). Chemotherapy in neuroendocrine tumors. Cancers (Basel) 13, 4872. doi:10.3390/cancers13194872
Dembla, E., and Becherer, U. (2021). Biogenesis of large dense core vesicles in mouse chromaffin cells. Traffic 22, 78–93. doi:10.1111/tra.12783
Dineen, R., Stewart, P. M., and Sherlock, M. (2017). Acromegaly. QJM 110, 411–420. doi:10.1093/qjmed/hcw004
Eisenhofer, G., Huynh, T. T., Elkahloun, A., Morris, J. C., Bratslavsky, G., Linehan, W. M., et al. (2008). Differential expression of the regulated catecholamine secretory pathway in different hereditary forms of pheochromocytoma. Am. J. Physiol. Endocrinol. Metab. 295, E1223–E1233. doi:10.1152/ajpendo.90591.2008
Elias, S., Delestre, C., Ory, S., Marais, S., Courel, M., Vazquez-Martinez, R., et al. (2012). Chromogranin A induces the biogenesis of granules with calcium- and actin-dependent dynamics and exocytosis in constitutively secreting cells. Endocrinology 153, 4444–4456. doi:10.1210/en.2012-1436
Faggiano, A. (2024). Long-acting somatostatin analogs and well differentiated neuroendocrine tumors: a 20-year-old story. J. Endocrinol. Invest 47, 35–46. doi:10.1007/s40618-023-02170-9
Gabel, M., Delavoie, F., Demais, V., Royer, C., Bailly, Y., Vitale, N., et al. (2015). Annexin A2-dependent actin bundling promotes secretory granule docking to the plasma membrane and exocytosis. J. Cell Biol. 210, 785–800. doi:10.1083/jcb.201412030
Gasman, S., Chasserot-Golaz, S., Popoff, M. R., Aunis, D., and Bader, M. F. (1997). Trimeric G proteins control exocytosis in chromaffin cells. Go regulates the peripheral actin network and catecholamine secretion by a mechanism involving the small GTP-binding protein Rho. J. Biol. Chem. 272, 20564–20571. doi:10.1074/jbc.272.33.20564
Gasman, S., Chasserot-Golaz, S., Popoff, M. R., Aunis, D., and Bader, M. F. (1999). Involvement of Rho GTPases in calcium-regulated exocytosis from adrenal chromaffin cells. J. Cell Sci. 112 (Pt 24), 4763–4771. doi:10.1242/jcs.112.24.4763
Gasman, S., and Vitale, N. (2017). Lipid remodelling in neuroendocrine secretion. Biol. Cell 109, 381–390. doi:10.1111/boc.201700030
Giner, D., Lopez, I., Villanueva, J., Torres, V., Viniegra, S., and Gutierrez, L. M. (2007). Vesicle movements are governed by the size and dynamics of F-actin cytoskeletal structures in bovine chromaffin cells. Neuroscience 146, 659–669. doi:10.1016/j.neuroscience.2007.02.039
Giovinazzo, F., Schimmack, S., Svejda, B., Alaimo, D., Pfragner, R., Modlin, I., et al. (2013). Chromogranin A and its fragments as regulators of small intestinal neuroendocrine neoplasm proliferation. PLoS One 8, e81111. doi:10.1371/journal.pone.0081111
Giusti, L., Cetani, F., Ciregia, F., Da Valle, Y., Donadio, E., Giannaccini, G., et al. (2011). A proteomic approach to study parathyroid glands. Mol. Biosyst. 7, 687–699. doi:10.1039/c0mb00191k
Grouzmann, E., Matter, M., Bilz, S., Herren, A., Triponez, F., Henzen, C., et al. (2012). Monoamine oxidase A down-regulation contributes to high metanephrine concentration in pheochromocytoma. J. Clin. Endocrinol. Metab. 97, 2773–2781. doi:10.1210/jc.2012-1557
Guerin, M., Guillemot, J., Thouennon, E., Pierre, A., El-Yamani, F. Z., Montero-Hadjadje, M., et al. (2010). Granins and their derived peptides in normal and tumoral chromaffin tissue: implications for the diagnosis and prognosis of pheochromocytoma. Regul. Pept. 165, 21–29. doi:10.1016/j.regpep.2010.06.003
Guillemot, J., Anouar, Y., Montero-Hadjadje, M., Grouzmann, E., Grumolato, L., Roshmaninho-Salgado, J., et al. (2006). Circulating EM66 is a highly sensitive marker for the diagnosis and follow-up of pheochromocytoma. Int. J. Cancer 118, 2003–2012. doi:10.1002/ijc.21571
Guillemot, J., Thouennon, E., Guerin, M., Vallet-Erdtmann, V., Ravni, A., Montero-Hadjadje, M., et al. (2012). Differential expression and processing of secretogranin II in relation to the status of pheochromocytoma: implications for the production of the tumoral marker EM66. J. Mol. Endocrinol. 48, 115–127. doi:10.1530/JME-11-0077
Gutierrez, L. M., and Villanueva, J. (2018). The role of F-actin in the transport and secretion of chromaffin granules: an historic perspective. Pflugers Arch. 470, 181–186. doi:10.1007/s00424-017-2040-9
Houy, S., Streit, L., Drissa, I., Rame, M., Decraene, C., Moog, S., et al. (2022). Dysfunction of calcium-regulated exocytosis at a single-cell level causes catecholamine hypersecretion in patients with pheochromocytoma. Cancer Lett. 543, 215765 doi:10.1016/j.canlet.2022.215765
Isobe, K., Nakai, T., Yukimasa, N., Nanmoku, T., Takekoshi, K., and Nomura, F. (1998). Expression of mRNA coding for four catecholamine-synthesizing enzymes in human adrenal pheochromocytomas. Eur. J. Endocrinol. 138, 383–387. doi:10.1530/eje.0.1380383
Jahn, R., Cafiso, D. C., and Tamm, L. K. (2024). Mechanisms of SNARE proteins in membrane fusion. Nat. Rev. Mol. Cell Biol. 25, 101–118. doi:10.1038/s41580-023-00668-x
Jarrott, B., and Louis, W. J. (1977). Abnormalities in enzymes involved in catecholamine synthesis and catabolism in phaeochromocytoma. Clin. Sci. Mol. Med. 53, 529–535. doi:10.1042/cs0530529
Kalwat, M. A., Yoder, S. M., Wang, Z., and Thurmond, D. C. (2013). A p21-activated kinase (PAK1) signaling cascade coordinately regulates F-actin remodeling and insulin granule exocytosis in pancreatic beta cells. Biochem. Pharmacol. 85, 808–816. doi:10.1016/j.bcp.2012.12.003
Kim, M., and Kim, B. H. (2021). Current guidelines for management of medullary thyroid carcinoma. Endocrinol. Metab. Seoul. 36, 514–524. doi:10.3803/EnM.2021.1082
Kim, T., Gondre-Lewis, M. C., Arnaoutova, I., and Loh, Y. P. (2006). Dense-core secretory granule biogenesis. Physiol. (Bethesda) 21, 124–133. doi:10.1152/physiol.00043.2005
Kim, T., Tao-Cheng, J. H., Eiden, L. E., and Loh, Y. P. (2001). Chromogranin A, an “on/off” switch controlling dense-core secretory granule biogenesis. Cell 106, 499–509. doi:10.1016/s0092-8674(01)00459-7
Koch, M., and Holt, M. (2012). Coupling exo- and endocytosis: an essential role for PIP₂ at the synapse. Biochim. Biophys. Acta 1821, 1114–1132. doi:10.1016/j.bbalip.2012.02.008
Koffer, A., Tatham, P. E., and Gomperts, B. D. (1990). Changes in the state of actin during the exocytotic reaction of permeabilized rat mast cells. J. Cell Biol. 111, 919–927. doi:10.1083/jcb.111.3.919
Kurczyk, A., Gawin, M., Chekan, M., Wilk, A., Lakomiec, K., Mrukwa, G., et al. (2020). Classification of thyroid tumors based on mass spectrometry imaging of tissue microarrays; a single-pixel approach. Int. J. Mol. Sci. 21, 6289. doi:10.3390/ijms21176289
Lam, A. D., Tryoen-Toth, P., Tsai, B., Vitale, N., and Stuenkel, E. L. (2008). SNARE-catalyzed fusion events are regulated by Syntaxin1A-lipid interactions. Mol. Biol. Cell 19, 485–497. doi:10.1091/mbc.e07-02-0148
Lang, T. (2007). SNARE proteins and 'membrane rafts. J. Physiol. 585, 693–698. doi:10.1113/jphysiol.2007.134346
Lang, T., Wacker, I., Wunderlich, I., Rohrbach, A., Giese, G., Soldati, T., et al. (2000). Role of actin cortex in the subplasmalemmal transport of secretory granules in PC-12 cells. Biophys. J. 78, 2863–2877. doi:10.1016/S0006-3495(00)76828-7
Li, P., Bademosi, A. T., Luo, J., and Meunier, F. A. (2018). Actin remodeling in regulated exocytosis: toward a mesoscopic view. Trends Cell Biol. 28, 685–697. doi:10.1016/j.tcb.2018.04.004
Loyet, K. M., Kowalchyk, J. A., Chaudhary, A., Chen, J., Prestwich, G. D., and Martin, T. F. (1998). Specific binding of phosphatidylinositol 4,5-bisphosphate to calcium-dependent activator protein for secretion (CAPS), a potential phosphoinositide effector protein for regulated exocytosis. J. Biol. Chem. 273, 8337–8343. doi:10.1074/jbc.273.14.8337
Lu, M., Forsberg, L., Hoog, A., Juhlin, C. C., Vukojevic, V., Larsson, C., et al. (2008). Heterogeneous expression of SNARE proteins SNAP-23, SNAP-25, Syntaxin1 and VAMP in human parathyroid tissue. Mol. Cell Endocrinol. 287, 72–80. doi:10.1016/j.mce.2008.01.028
Ma, C. J., Burgess, J., and Brill, J. A. (2021). Maturing secretory granules: where secretory and endocytic pathways converge. Adv. Biol. Regul. 80, 100807. doi:10.1016/j.jbior.2021.100807
Malacombe, M., Bader, M. F., and Gasman, S. (2006a). Exocytosis in neuroendocrine cells: new tasks for actin. Biochim. Biophys. Acta 1763, 1175–1183. doi:10.1016/j.bbamcr.2006.09.004
Malacombe, M., Ceridono, M., Calco, V., Chasserot-Golaz, S., Mcpherson, P. S., Bader, M. F., et al. (2006b). Intersectin-1L nucleotide exchange factor regulates secretory granule exocytosis by activating Cdc42. Embo J. 25, 3494–3503. doi:10.1038/sj.emboj.7601247
Martin, T. F. (2012). Role of PI(4,5)P(2) in vesicle exocytosis and membrane fusion. Subcell. Biochem. 59, 111–130. doi:10.1007/978-94-007-3015-1_4
Mohrmann, R., De Wit, H., Connell, E., Pinheiro, P. S., Leese, C., Bruns, D., et al. (2013). Synaptotagmin interaction with SNAP-25 governs vesicle docking, priming, and fusion triggering. J. Neurosci. 33, 14417–14430. doi:10.1523/JNEUROSCI.1236-13.2013
Mohrmann, R., De Wit, H., Verhage, M., Neher, E., and Sorensen, J. B. (2010). Fast vesicle fusion in living cells requires at least three SNARE complexes. Science 330, 502–505. doi:10.1126/science.1193134
Momboisse, F., Houy, S., Ory, S., Calco, V., Bader, M. F., and Gasman, S. (2011). How important are Rho GTPases in neurosecretion? J. Neurochem. 117, 623–631. doi:10.1111/j.1471-4159.2011.07241.x
Morvan, J., and Tooze, S. A. (2008). Discovery and progress in our understanding of the regulated secretory pathway in neuroendocrine cells. Histochem Cell Biol. 129, 243–252. doi:10.1007/s00418-008-0377-z
Nehar, D., Lombard-Bohas, C., Olivieri, S., Claustrat, B., Chayvialle, J. A., Penes, M. C., et al. (2004). Interest of Chromogranin A for diagnosis and follow-up of endocrine tumours. Clin. Endocrinol. (Oxf) 60, 644–652. doi:10.1111/j.1365-2265.2004.02030.x
Nobels, F. R., Kwekkeboom, D. J., Coopmans, W., Schoenmakers, C. H., Lindemans, J., De Herder, W. W., et al. (1997). Chromogranin A as serum marker for neuroendocrine neoplasia: comparison with neuron-specific enolase and the alpha-subunit of glycoprotein hormones. J. Clin. Endocrinol. Metab. 82, 2622–2628. doi:10.1210/jcem.82.8.4145
Onaitis, M. W., Kirshbom, P. M., Hayward, T. Z., Quayle, F. J., Feldman, J. M., Seigler, H. F., et al. (2000). Gastrointestinal carcinoids: characterization by site of origin and hormone production. Ann. Surg. 232, 549–556. doi:10.1097/00000658-200010000-00010
Orci, L., Gabbay, K. H., and Malaisse, W. J. (1972). Pancreatic beta-cell web: its possible role in insulin secretion. Science 175, 1128–1130. doi:10.1126/science.175.4026.1128
Ory, S., Ceridono, M., Momboisse, F., Houy, S., Chasserot-Golaz, S., Heintz, D., et al. (2013). Phospholipid scramblase-1-induced lipid reorganization regulates compensatory endocytosis in neuroendocrine cells. J. Neurosci. 33, 3545–3556. doi:10.1523/JNEUROSCI.3654-12.2013
Papadopulos, A. (2017). Membrane shaping by actin and myosin during regulated exocytosis. Mol. Cell Neurosci. 84, 93–99. doi:10.1016/j.mcn.2017.05.006
Papadopulos, A., Tomatis, V. M., Kasula, R., and Meunier, F. A. (2013). The cortical acto-myosin network: from diffusion barrier to functional gateway in the transport of neurosecretory vesicles to the plasma membrane. Front. Endocrinol. (Lausanne) 4, 153. doi:10.3389/fendo.2013.00153
Petersenn, S., Fleseriu, M., Casanueva, F. F., Giustina, A., Biermasz, N., Biller, B. M. K., et al. (2023). Diagnosis and management of prolactin-secreting pituitary adenomas: a Pituitary Society international Consensus Statement. Nat. Rev. Endocrinol. 19, 722–740. doi:10.1038/s41574-023-00886-5
Pinheiro, P. S., Houy, S., and Sorensen, J. B. (2016). C2-domain containing calcium sensors in neuroendocrine secretion. J. Neurochem. 139, 943–958. doi:10.1111/jnc.13865
Ribeiro-Oliveira, A., Franchi, G., Kola, B., Dalino, P., Pinheiro, S. V., Salahuddin, N., et al. (2008). Protein western array analysis in human pituitary tumours: insights and limitations. Endocr. Relat. Cancer 15, 1099–1114. doi:10.1677/ERC-08-0003
Schiavo, G., Gu, Q. M., Prestwich, G. D., Sollner, T. H., and Rothman, J. E. (1996). Calcium-dependent switching of the specificity of phosphoinositide binding to synaptotagmin. Proc. Natl. Acad. Sci. U. S. A. 93, 13327–13332. doi:10.1073/pnas.93.23.13327
Schonn, J. S., Maximov, A., Lao, Y., Sudhof, T. C., and Sorensen, J. B. (2008). Synaptotagmin-1 and -7 are functionally overlapping Ca2+ sensors for exocytosis in adrenal chromaffin cells. Proc. Natl. Acad. Sci. U. S. A. 105, 3998–4003. doi:10.1073/pnas.0712373105
Sharma, S., and Fishbein, L. (2023). Diagnosis and management of pheochromocytomas and paragangliomas: a guide for the clinician. Endocr. Pract. 29, 999–1006. doi:10.1016/j.eprac.2023.07.027
Streit, L., Brunaud, L., Vitale, N., Ory, S., and Gasman, S. (2020). Hormones secretion and Rho GTPases in neuroendocrine tumors. Cancers (Basel) 12, 1859. doi:10.3390/cancers12071859
Stueven, A. K., Kayser, A., Wetz, C., Amthauer, H., Wree, A., Tacke, F., et al. (2019). Somatostatin analogues in the treatment of neuroendocrine tumors: past, present and future. Int. J. Mol. Sci. 20, 3049. doi:10.3390/ijms20123049
Tanguy, E., Carmon, O., Wang, Q., Jeandel, L., Chasserot-Golaz, S., Montero-Hadjadje, M., et al. (2016). Lipids implicated in the journey of a secretory granule: from biogenesis to fusion. J. Neurochem. 137, 904–912. doi:10.1111/jnc.13577
Tanguy, E., Coste De Bagneaux, P., Kassas, N., Ammar, M. R., Wang, Q., Haeberle, A. M., et al. (2020). Mono- and poly-unsaturated phosphatidic acid regulate distinct steps of regulated exocytosis in neuroendocrine cells. Cell Rep. 32, 108026. doi:10.1016/j.celrep.2020.108026
Tawfik, B., Martins, J. S., Houy, S., Imig, C., Pinheiro, P. S., Wojcik, S. M., et al. (2021). Synaptotagmin-7 places dense-core vesicles at the cell membrane to promote Munc13-2- and Ca(2+)-dependent priming. Elife 10, e64527. doi:10.7554/eLife.64527
Tomic, T. T., Olausson, J., Rehammar, A., Deland, L., Muth, A., Ejeskar, K., et al. (2020). MYO5B mutations in pheochromocytoma/paraganglioma promote cancer progression. PLoS Genet. 16, e1008803. doi:10.1371/journal.pgen.1008803
Tooze, S. A., Flatmark, T., Tooze, J., and Huttner, W. B. (1991). Characterization of the immature secretory granule, an intermediate in granule biogenesis. J. Cell Biol. 115, 1491–1503. doi:10.1083/jcb.115.6.1491
Trifaro, J. M., Bader, M. F., and Doucet, J. P. (1985). Chromaffin cell cytoskeleton: its possible role in secretion. Can. J. Biochem. Cell Biol. 63, 661–679. doi:10.1139/o85-084
Trifaro, J. M., Gasman, S., and Gutierrez, L. M. (2008). Cytoskeletal control of vesicle transport and exocytosis in chromaffin cells. Acta Physiol. (Oxf) 192, 165–172. doi:10.1111/j.1748-1716.2007.01808.x
Uenishi, E., Shibasaki, T., Takahashi, H., Seki, C., Hamaguchi, H., Yasuda, T., et al. (2013). Actin dynamics regulated by the balance of neuronal Wiskott-Aldrich syndrome protein (N-WASP) and cofilin activities determines the biphasic response of glucose-induced insulin secretion. J. Biol. Chem. 288, 25851–25864. doi:10.1074/jbc.M113.464420
Van Meer, G., Voelker, D. R., and Feigenson, G. W. (2008). Membrane lipids: where they are and how they behave. Nat. Rev. Mol. Cell Biol. 9, 112–124. doi:10.1038/nrm2330
Varlamov, E. V., Mccartney, S., and Fleseriu, M. (2019). Functioning pituitary adenomas - current treatment options and emerging medical therapies. Eur. Endocrinol. 15, 30–40. doi:10.17925/EE.2019.15.1.30
Vitale, M. L., Rodriguez Del Castillo, A., Tchakarov, L., and Trifaro, J. M. (1991). Cortical filamentous actin disassembly and scinderin redistribution during chromaffin cell stimulation precede exocytosis, a phenomenon not exhibited by gelsolin. J. Cell Biol. 113, 1057–1067. doi:10.1083/jcb.113.5.1057
Vitale, M. L., Seward, E. P., and Trifaro, J. M. (1995). Chromaffin cell cortical actin network dynamics control the size of the release-ready vesicle pool and the initial rate of exocytosis. Neuron 14, 353–363. doi:10.1016/0896-6273(95)90291-0
Vitale, N., Caumont, A. S., Chasserot-Golaz, S., Du, G., Wu, S., Sciorra, V. A., et al. (2001). Phospholipase D1: a key factor for the exocytotic machinery in neuroendocrine cells. Embo J. 20, 2424–2434. doi:10.1093/emboj/20.10.2424
Wilzen, A., Rehammar, A., Muth, A., Nilsson, O., Tesan Tomic, T., Wangberg, B., et al. (2016). Malignant pheochromocytomas/paragangliomas harbor mutations in transport and cell adhesion genes. Int. J. Cancer 138, 2201–2211. doi:10.1002/ijc.29957
Wu, L. G., and Chan, C. Y. (2022). Multiple roles of actin in exo- and endocytosis. Front. Synaptic Neurosci. 14, 841704. doi:10.3389/fnsyn.2022.841704
Y-Hassan, S., and Falhammar, H. (2020). Cardiovascular manifestations and complications of pheochromocytomas and paragangliomas. J. Clin. Med. 9, 2435. doi:10.3390/jcm9082435
Zappi, A., Persano, I., Galvani, L., Parlagreco, E., Andrini, E., Campana, D., et al. (2023). Chemotherapy in well differentiated neuroendocrine tumors (NET) G1, G2, and G3: a narrative review. J. Clin. Med. 12, 717. doi:10.3390/jcm12020717
Zeniou-Meyer, M., Zabari, N., Ashery, U., Chasserot-Golaz, S., Haeberle, A. M., Demais, V., et al. (2007). Phospholipase D1 production of phosphatidic acid at the plasma membrane promotes exocytosis of large dense-core granules at a late stage. J. Biol. Chem. 282, 21746–21757. doi:10.1074/jbc.M702968200
Zhan, S., Li, J., Wang, T., and Ge, W. (2018). Quantitative proteomics analysis of sporadic medullary thyroid cancer reveals FN1 as a potential novel candidate prognostic biomarker. Oncologist 23, 1415–1425. doi:10.1634/theoncologist.2017-0399
Zhang, D., Hugo, W., Bergsneider, M., Wang, M. B., Kim, W., Vinters, H. V., et al. (2022). Single-cell RNA sequencing in silent corticotroph tumors confirms impaired POMC processing and provides new insights into their invasive behavior. Eur. J. Endocrinol. 187, 49–64. doi:10.1530/EJE-21-1183
Keywords: neuroendocrine tumors, hormone, secretion, vesicular trafficking, exocytosis, dysregulated genes and proteins
Citation: Streit L, Tanguy E, Brunaud L, Tóth P, Vitale N, Ory S and Gasman S (2025) Cellular mechanisms of hormone secretion in neuroendocrine tumors: what goes wrong?. Front. Cell Dev. Biol. 13:1527083. doi: 10.3389/fcell.2025.1527083
Received: 12 November 2024; Accepted: 05 June 2025;
Published: 01 July 2025.
Edited by:
Georg Haase, Aix Marseille Université, FranceReviewed by:
Cedric Asensio, University of Denver, United StatesCopyright © 2025 Streit, Tanguy, Brunaud, Tóth, Vitale, Ory and Gasman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphane Gasman, Z2FzbWFuQHVuaXN0cmEuZnI=