Abstract
Mast cells (MCs) are sentinel cells of the immune system that play important protective roles in innate host defenses but are also key effectors of allergic responses and chronic inflammatory diseases. Both physiological and pathophysiological responses of MCs are mediated by the release of inflammatory mediators, many of which are stored, preformed, in secretory granules (SGs), and released by regulated exocytosis in response to multiple stimuli. MC SGs belong to the family of lysosome related organelles (LROs), as indicated by their content of lysosomal hydrolases, lysosomal membrane proteins and acidic pH. The SGs derive from the Golgi and increase in size in a quantal manner by their fusion with additional SGs. They have access to external cargo, which they acquire by fusion with endosomes and contain LC3, which they acquire by fusion with amphisomes. This review discusses the underlying mechanisms of MC SG biogenesis and remodeling.
1 Introduction
Mast cells (MCs) are key regulatory cells of the immune system (Dahlin et al., 2022). Though best known for their critical role in allergy and anaphylaxis (Vitte et al., 2022), MCs also contribute to innate defense against infections and play significant roles in inflammatory conditions associated with autoimmunity, cancer, and neurodegenerative diseases (Theoharides et al., 2015; Segura-Villalobos et al., 2022; Hendriksen et al., 2017; Lin et al., 2023; Jiménez et al., 2021; St John and Abraham, 2013). MCs are particularly abundant at the interfaces between the external environment and the internal milieu, such as the skin, mucosa of the lungs, digestive tract, mouth, conjunctiva, and nose (Prussin and Metcalfe, 2006). Their presence in the brain has also been documented (Theoharides et al., 2024). In these locations, MCs are found in close proximity to blood vessels and sensory neurons. MCs originate from CD34+/CD117+ pluripotent progenitor cells in the bone marrow (Kirshenbaum et al., 1999). These progenitors migrate into peripheral tissues, where they mature and undergo terminal differentiation under the influence of local cytokines (Metcalfe et al., 1997; Prussin and Metcalfe, 2006). In connective tissues such as the skin, bone marrow–derived MCs progressively replace MCs that originated from extra-embryonic yolk sac and fetal liver (Chia et al., 2023).
MCs have been categorized into subsets based on their localization and protease expression profiles. In rodents, they are classified as mucosal MCs (MMCs) or connective tissue type MCs (CTMCs), whereas in humans, they are distinguished by their protease content: MCTC, which co-express tryptase and chymase among other proteases, and MCT, which express only tryptase (Reber et al., 2015). For both MC types, the transition from progenitor to mature MCs depends on activation of the c-KIT receptor upon binding of its ligand, stem cell factor (SCF) (Mekori et al., 1995). Additionally, both subsets express FcεRI, the high-affinity receptor for immunoglobulin E (IgE), which triggers MC activation upon allergen-induced crosslinking of cell-bound IgE (Dema et al., 2014; Nagata and Suzuki, 2022; Blank et al., 2021).
MC subsets differ not only in their protease expression profiles but also in their expression of Mrgprs, a family of G protein-coupled receptors selectively expressed in CTMCs or MCTC (West and Bulfone-Paus, 2022; Ali, 2021; McNeil et al., 2015). These receptors enable IgE-independent activation in response to various ligands, previously termed MC basic secretagogues due to their positive charge (West and Bulfone-Paus, 2022; Ali, 2021; McNeil et al., 2015). These ligands include exogenous molecules such as toxins (e.g., the wasp venom peptide mastoparan), a wide range of FDA-approved drugs, such as vancomycin, and endogenous ligands such as neuropeptides (e.g., substance P) and antimicrobial peptides. Their activation of MCs can trigger pseudo-allergic reactions but also rapid innate immune and neorogenic responses (Subramanian et al., 2016). Notably, recent transcriptomic analyses of MCs from different tissues indicate that MC heterogeneity extends beyond their classification as MMCs or CTMCs, emphasizing the critical role of their microenvironment (Tauber et al., 2023; Akula et al., 2020). This aligns with evidence that MC activation is influenced by crosstalk with neighboring cells, including stromal cells, immune cells and neurons (Wang et al., 2024; Gri et al., 2012; Toyoshima and Okayama, 2022; Bao and Abraham, 2024). Interestingly, MCs are also present in the brain, where they interact with microglia, further highlighting their role in neuroimmune regulation (Huang et al., 2024; Hendriksen et al., 2017; Sandhu and Kulka, 2021).
Both the physiological immune responses of the MCs and their pathological functions in allergy and inflammation are primarily mediated by the release of inflammatory mediators, part of which, such as histamine, proteoglycans and proteases, are preformed and stored in secretory granules (SGs), that release their content immediately after activation by exocytosis. Others, such as prostaglandins, leukotrienes, cytokines and chemokines are synthesized de novo and released thereafter (Moon et al., 2014; Wernersson and Pejler, 2014; Blank et al., 2014; Gordon and Galli, 1990). Collectively, these mediators initiate early and late inflammatory responses.
2 The MC SGs
MC SGs belong to the family of lysosome related organelles (LROs), a specialized subset of SGs that exhibit lysosomal characteristics (Delevoye et al., 2019; Marks et al., 2013; Dell’Angelica et al., 2000; Luzio et al., 2014). The latter include secretory organelles of other immune cells, such as neutrophils, natural killer cells, and cytotoxic T lymphocytes, as well as melanocytes and osteoclasts, in which LROs play a role in pathogen killing, pigmentation and bone absorption (Delevoye et al., 2019; Marks et al., 2013; Dell'Angelica et al., 2000; Luzio et al., 2014). In neurons LROs are precursors of synaptic vesicles (Vukoja et al., 2018). Indeed, the MC SGs contain in addition to their inflammatory mediators, lysosomal enzymes (Schwartz and Austen, 1980) and lysosomal membrane proteins (LIMPs) (Suárez-Quian, 1987) and also contain an acidic luminal pH (Johnson et al., 1980) (Figure 1). The SGs also receive and exocytose in a regulated fashion endocytic cargo (Xu et al., 1998), recycle SG proteins (Bonifacino et al., 1989), and are regulated by endocytic recycling controlling synaptotagmins (Grimberg et al., 2003; Haberman et al., 2007). Based on electron microscopy (EM) analyses (Raposo et al., 1997), differential regulation by Soluble N-ethylmaleimide-sensitive-factor Attachment protein REceptors (SNAREs) (Adhikari et al., 2023; Xu et al., 2018; Lorentz et al., 2012), and fractionation data (Baram et al., 1999; Grimberg et al., 2003), MCs contain discrete types of SGs, which differ in their morphology (Raposo et al., 1997) and content composition (Puri and Roche, 2008). Specifically, based on their transmission EM features, the SGs were classified into three types: Type I granules, which contain intraluminal vesicles (ILVs), reminiscent of multivesicular bodies (MVBs), and become accessible to external cargo after a 20-min lag, Type II granules which display a serotonin-rich electron-dense core surrounded by ILVs and become accessible to endocytic cargo at a later stage, and Type III granules, which are electron-dense and lack ILVs (Raposo et al., 1997). Both Type I and Type II granules also contain MHC class II molecules, mannose-6-phosphate receptors, and lysosomal membrane proteins (Lamp1 and Lamp2), which localize to the small intraluminal vesicles (Raposo et al., 1997).
FIGURE 1

MC lysosome related SGs. MCs contain SGs which store preformed inflammatory mediators, including biogenic amines, such as histamine, serotonin and polyamines, proteoglycans, such as heparin and chondroitin sulfates, proteases such as tryptase, chymase and carboxypeptidase A3 and some cytokines, such as TNF-α. MC SGs also contain lysosomal enzymes, such as β-hexosaminidase and β-glucuronidase and lysosomal membrane proteins, such as CD63. In response to cell activation, for example, via the FcεRI, the high affinity receptor of IgE, via the binding of an allergen to cell bound IgE, or by ligands that bind to Mrgprs, G protein coupled receptors expressed in a subset of MCs, the SGs’ contents are released by exocytosis, a process referred to as degranulation. The inset is the enlargement of the boxed area. * “Created with BioRender.com”.
3 The relationship between lysosomes and the lysosome related SGs in MCs
Previous studies have demonstrated the presence of acid phosphatase in two populations of granules, one which comprised most granules, also exocytosed, while the other smaller one, was retained in triggered cells (Jamur and Vugman, 1990). Similarly, the enzyme Dipeptidyl aminopeptidase II (DAP II) was found to reside in few granules that reside near the nucleus and are retained in MCs that are triggered to degranulate (Sannes and Spicer, 1979). By cell fractionation, we have demonstrated the existence of two types of β-hexosaminidase containing fractions, one that also contains histamine, while the other is histamine-free (Baram et al., 1999). However, while these studies may imply the existence of lysosomes that are distinct from the SGs, fragments of IgE, that was bound to the IgE receptor, were shown to be released during exocytosis (Xu et al., 1998), suggesting that endocytosed IgE is degraded and delivered to the SGs or degraded in the SGs. Therefore, the precise relationship between degradative endolysosomes and the lysosomal related SGs is still poorly resolved (Figure 2).
FIGURE 2
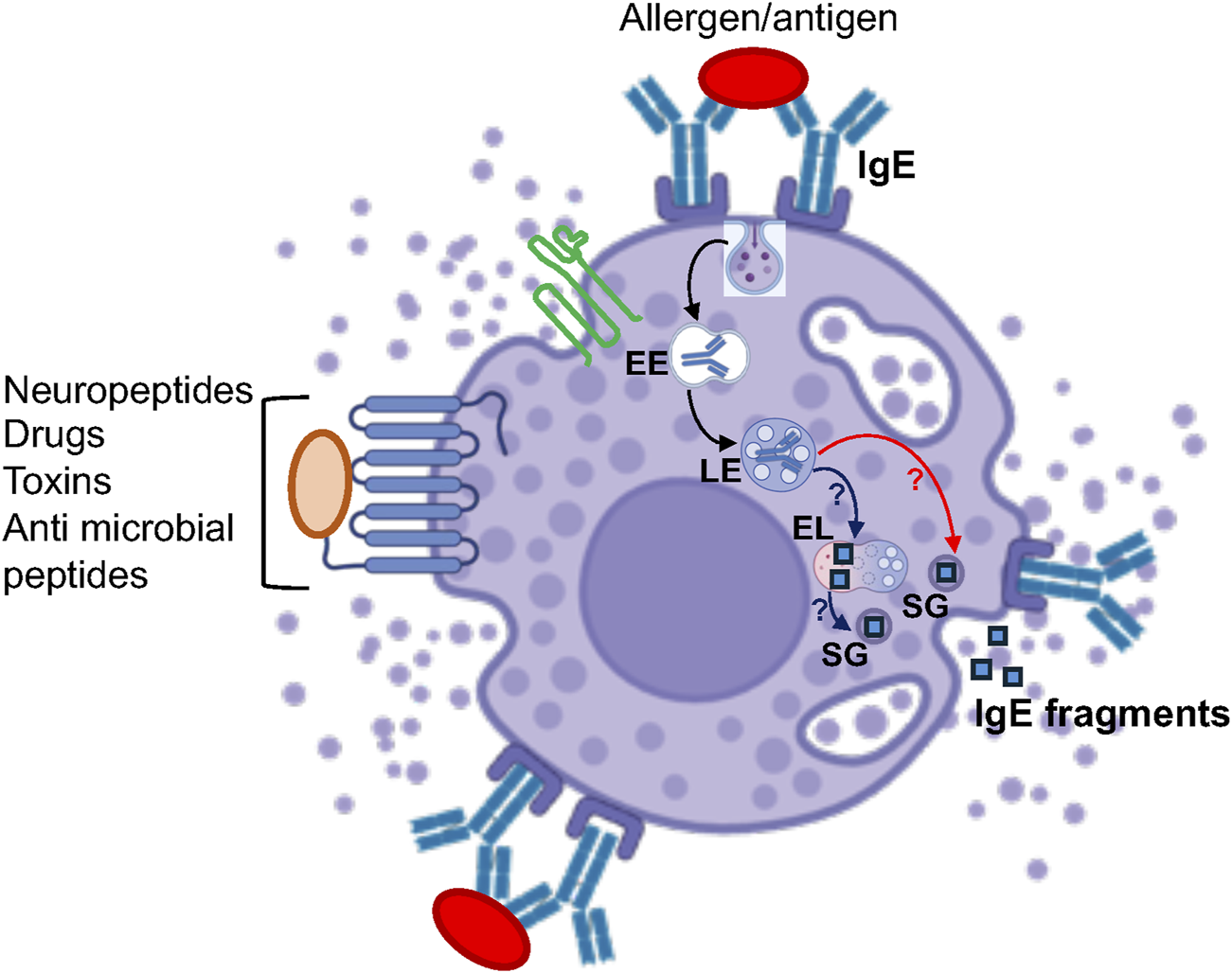
Model of the potential crosstalk between MC SGs and the endolysosomal system. According to this model, after internalization, external cargo such as IgE travels from early endosomes (EE) to late endosomes (LE) and is degraded in endolysosomes (EL). This degraded cargo may then be delivered to the SGs, possibly via SG-EL fusion (blue arrows), followed by its release during exocytosis. Alternatively, internalized cargo might be directly delivered from late endosomes to the SGs, which are proteolytically active, where it is degraded before being released. * “Created with BioRender.com”.
4 The crosstalk between MC SGs and the autophagic system
Further complexity in the mechanisms underlying MC SG biogenesis is highlighted by the presence of LC3 within SGs, where it colocalizes with the tetraspanin protein CD63 and is released during exocytosis (Nakano and Ushio, 2011). This observation indicates the existence of crosstalk between the autophagic system, which is constitutively active in MCs (Nakano and Ushio, 2011), and the SGs. Indeed, we have recently demonstrated dynamic interactions between SGs and the autophagic system, as evidenced by the targeting of MRGPRX2, the human member of the Mrgpr receptor family, to LC3-positive SGs following its substance P-induced internalization. Moreover, the number of LC3-positive SGs increases in response to receptor internalization (Lazki-Hagenbach et al., 2022). While the precise nature of these interactions remains to be fully elucidated, at least one mechanism involves SG fusion with amphisomes, as discussed below.
5 The biogenesis of MC SGs
5.1 MC SGs undergo dynamic remodelling
Early EM analyses of MCs (Combs, 1971), further supported by our analyses of the route of trafficking of Neuropeptide Y (NPY)-mRFP to the SGs (Azouz et al., 2012), demonstrate that the SGs derive from the Golgi, implying dynamic interactions between Golgi derived granules and the endocytic and autophagic systems. Subsequent analyses of EM images implicated fusion between SGs as a key post-Golgi mechanism for generating mature SGs (Hammel et al., 2010). In agreement with this model, we found that expression of a constitutively active mutant of the small GTPase Rab5 leads to SG enlargement that is linked with a reduction in SG number (Azouz et al., 2014). Conversely, Rab5 knockdown reduces the SG size while increasing the SG number (Azouz et al., 2014). These findings implicate Rab5 in the regulation of SG fusion. Furthermore, we have demonstrated that Rab5 also facilitates SG fusion with endosomes, allowing the incorporation of CD63 into the SGs (Azouz et al., 2014). This reinforces the idea that fusion events between Golgi-derived SGs contribute to their enlargement and the incorporation of both external and endogenous endocytic cargo. Further analysis of Rab5-mediated SG fusion has unveiled key steps in this process, clarifying several previously enigmatic observations (Omari et al., 2024). Specifically, we found that homotypic SG fusion is a multistep process that requires CD63 and depends on Rab5-regulated CD63 internalization. Additionally, SG enlargement necessitates fusion with amphisomes, hybrid organelles formed by the fusion of late endosomes with autophagosomes, highlighting further the close interplay between the SGs, late endosomes and autophagosomes (Figure 3). This mechanism explains the colocalization of LC3 with CD63 at SGs, as well as prior observations, which documented the release of mitochondrial fragments during MC exocytosis (Zhang et al., 2012). In addition to its dependence on CD63, SG fusion with amphisomes is regulated by phosphatidylinositol (PI) 3- and 4-kinases, as well as the protein tyrosine phosphatase PTPN9 (PTP-MEG2), which has previously been implicated in granule fusion (Omari et al., 2024). SG fusion is also associated with an enrichment of PI(3)P, PI(4)P, and PI(3,4,5)P3 in the SG membrane, of which the latter may be required for the activation of PTPN9, as its interaction with phosphoinositides is essential for relieving its autoinhibition (Kruger et al., 2002). Strikingly, we found that fusion with amphisomes not only enlarges SGs, allowing them to store greater amounts of secretory cargo that is “ready to go” during degranulation, but also endows SGs with the ability to release exosomes (Omari et al., 2024) (Figure 3).
FIGURE 3

Model of the biogenesis of MC SGs. According to this model, Golgi derived SGs [SG(I)] incorporate endocytic cargo, including CD63, which internalized from the plasma membrane by a Rab5-regulated mechanism, by fusion with early endosomes (EE). The fused SGs [SG (II)] can further fuse by a mechanism dependent on the protein tyrosine phosphatase PTPN9, CD63 and phosphatidylinositol-4-kinase (PI4K), with amphisomes (AMFs), which form by the fusion of late endosomes (LE) with autophagosomes (AFs), forming large and LC3-positive SGs [SGIII)] that also contain intraluminal vesicles. Both SG (II) and SG (III) are exocytosis competent. However, in response to an external trigger, SG (II) release prestored inflammatory mediators, while SG (III) additionally release CD63-positive exosomes. SG (III) can revert to SG (II) by undergoing dynamin-mediated fission. Dynamic cycling of the SGs between fusion and fission events is regulated by phosphoinositides interconversion between PIP3 and PIP2. * “Created with BioRender.com”.
Unexpectedly, we discovered that SGs also undergo fission through a dynamin-mediated mechanism, which is triggered by a switch from SG PI(3,4,5)P3 to PI(4,5)P2 (Omari et al., 2024) (Figure 3). Taken together, these findings suggest that MCs may harbor SGs of varying sizes and contents, depending on their fusion and fission events. Some smaller SGs may only release soluble mediators, potentially through kiss-and-run exocytosis, while larger SGs may release both soluble mediators and exosomes, likely via compound exocytosis, which requires a more stable fusion pore opening (Flašker et al., 2013). The relative abundance of distinct SG subtypes is likely regulated by factors such as the activation state of relevant lipid kinases and phosphatases, which determine the SG phosphoinositide composition, as well as the extent of internalization of CD63 and the cellular levels of amphisomes (Figure 3).
5.2 The role of protein recycling
5.2.1 The role of LYST
Chediak-Higashi syndrome (CHS) is an autosomal recessive disorder caused by mutations in the Lysosomal Trafficking Regulator (LYST) gene, which lead to a broad range of clinical manifestations associated with the enlargement of lysosomes and LROs, including MC SGs (Turner et al., 2024; Shiflett et al., 2002; Kiyoi et al., 2019). Mechanistically, LYST has been implicated in regulating lysosome/LRO size by promoting fusion or inhibiting fission. It has also been linked to controlling the movement of lysosomes and LROs (Turner et al., 2024; Serra-Vinardell et al., 2023). However, the precise mechanism underlying LYST’s function remains unresolved. Interestingly, unlike the functional impact of LYST mutations on the lytic granules of cytotoxic T lymphocytes or NK cells, which results in reduced cytotoxicity (Turner et al., 2024), analyses of skin and peritoneal MCs (i.e., CTMCs) and bone marrow-derived MCs (i.e., BMMCs) from homozygous Beige mice carrying a mutation in LYST revealed enlarged SGs, which despite this enlargement, preserved their exocytosis competence (Kiyoi et al., 2019). This observation is consistent with our findings showing that the SG size has no impact on their exocytosis competence (Omari et al., 2024).
5.2.2 Regulation by synaptotagmins
A role for endocytic recycling in the biogenesis of MC SGs is suggested by the influence of certain members of the synaptotagmin (Syt) family of proteins on their biogenesis. Seventeen members of this family have been identified based on their common structural features, which include a short lumenal/extracellular domain, a transmembrane or membrane association domain (for Syt16 and Syt17), and two cytoplasmic C2 calcium-binding domains (Wolfes and Dean, 2020). Syts have been implicated in regulating protein trafficking along both exocytic and endocytic routes (Wolfes and Dean, 2020). In MCs, knockdown of Syt III, which interfered with the transport of internalized transferrin to the endocytic recycling compartment (ERC), induced a Chediak-Higashi-like phenotype, characterized by a significant increase in the number of giant SGs (Grimberg et al., 2003). Knockdown of Syt IX, which disrupted the recycling of transferrin (Tfn) from the ERC to the plasma membrane (Haberman et al., 2003), led to the mistargeting of TGN38 to the SGs (Haberman et al., 2007). These findings support a model in which endocytic recycling plays a role in segregating endosomal cargo, preventing its accumulation in SGs. Spillover of cargo from the ERC to late endosomes may result in mistargeting of TGN cargo to the SGs, possibly via the formation of amphisomes.
5.3 The role of Hermansky-Pudlak Syndrome genes
Hermansky-Pudlak Syndrome is a group of autosomal recessive disorders characterized by oculocutaneous albinism, bleeding disorders, innate immune deficiency and pulmonary fibrosis, all of which are associated with abnormalities in LRO biogenesis in melanocytes, platelets, neutrophils, natural killer cells and cytotoxic T lymphocytes (Bowman et al., 2019; Wei, 2006; Banushi and Simpson, 2022; Starcevic et al., 2002). The disease is caused by genetic defects in 11 different genes that encode subunits of protein complexes involved in the biogenesis of LROs (Banushi and Simpson, 2022). These include subunits of the Biogenesis of Lysosome-related Organelles Complexes (BLOC)-1, -2, and -3, as well as the β subunit of the adaptor complex AP-3 (Dell’Angelica, 2009; Banushi and Simpson, 2022). Analysis of dermal MCs and an HPS-1-derived MC culture revealed abnormalities in SG morphology and an increase in activation markers (Kirshenbaum et al., 2016). These findings suggest a role for HPS-1, a subunit of BLOC-3, in the biogenesis of MC SGs, although the precise mechanism remains to be further investigated. In a separate study, the role of the AP-3 complex was examined. This complex is part of the AP-1 to AP-5 family of adaptor protein complexes, which mediate the transport of distinct types of vesicles (Dacks and Robinson, 2017; Begley et al., 2024). The complexes are structurally related consisting of α/γ/δ/ε/ζ, β1-5, μ1-5, and σ1-5 subunits (Begley et al., 2024). Among these adaptor complexes, the AP-3 complex has been implicated in regulating transport from endosomes to lysosomes and LROs (Begley et al., 2024). shRNA-mediated knockdown of the δ subunit of AP-3 in RBL-2H3, a mast cell line widely used as a model for MC exocytosis (Falcone et al., 2018), destabilized the complex, leading to its depletion (da Silva et al., 2017). Morphometric evaluation of the SGs by EM revealed an increase in SG size (da Silva et al., 2017), like the phenotype observed with Syt III depletion (Grimberg et al., 2003). AP-3 may thus play a role in the cellular transport of Syt III and potentially other membrane proteins to the SGs.
Two other proteins linked to HPS are the Rab GTPases Rab32 and Rab38, for which the BLOC-3 complex displays GEF activity. Indeed, both Rabs have been implicated in the biogenesis of LROs in several cell types (Bultema and Di Pietro, 2013). In MCs, expression of a constitutively active mutant of Rab38, but not Rab32, selectively inhibited IgE-mediated degranulation, while it had no effect on MRGPRX2-or calcium ionophore and phorbol ester-induced release (Azouz et al., 2012). However, expression of neither Rab32 nor Rab38 affected the SG size (Azouz et al., 2012; Lazki-Hagenbach et al., 2024).
6 The regulation of MC SG transport
Similar to lysosomes and LROs in other cell types, the SGs in MCs move bidirectionally (Smith et al., 2003). Additionally, similar to other LROs, the anterograde transport of MC SGs is regulated by Rab27. However, in cytotoxic T lymphocytes, the anterograde movement of lytic granules to the immune synapse is mediated by Rab27a, which recruits kinesin-1 through its effector synaptotagmin-like protein 3 (Slp3) (Kurowska et al., 2012). In contrast, in MCs, kinesin-1 is recruited to SGs via the Rab27b–Slp3 complex (Munoz et al., 2016). Furthermore, in MCs this recruitment is dependent on PI3K activity and accordingly occurs only in activated cells (Munoz et al., 2016). Another regulator of kinesin-1-mediated translocation of the SGs to the plasma membrane in activated cells is the large GTPase Rab44 (Longé et al., 2022). The precise relationship between these two mechanisms of SG transport remains unknown. Finally, and most intriguingly, SG trafficking to the plasma membrane in activated cells was shown to require the association of inflammasome components with SGs (Mencarelli et al., 2024). Moreover, this mechanism also involves the motor protein dynein (Mencarelli et al., 2024), which has previously been implicated in the retrograde transport of MC SGs (Efergan et al., 2016). Notably, MC SGs differ from other LROs in their mechanism of retrograde transport. While Rab7 and Rab36 mediate the recruitment of the RILP–dynein complex to other LROs (Daniele et al., 2011; Matsui et al., 2012), in MCs this role is fulfilled by Rab12 (Efergan et al., 2016). Intriguingly, Rab12 also stimulates SG translocation to cell tips in activated cells (Efergan et al., 2016). How Rab12 and dynein can simultaneously promote perinuclear accumulation of a subset of SGs while driving translocation of another subset to the cell surface remains unknown. It is noteworthy that this type of dual regulation is not without precedent: Rab7 has been shown to drive lysosome movement in either direction by binding different effectors, depending on the cellular concentration of cholesterol (Rocha et al., 2009). Whether Rab12 controls the anterograde transport of SGs through effectors other than RILP, and how Rab12, Rab27b, Rab44 and inflammasome-regulated transport are functionally related, remain open questions. It is also worth noting that Rab12 is one of the physiological substrates of the leucine-rich repeat kinase 2 (LRRK2), a kinase whose hyperactivation is linked to Parkinson’s and Crohn’s diseases (Hui et al., 2018). However, whether phosphorylation of Rab12 plays a role in regulating MC functions is currently unknown.
7 SG homeostasis
7.1 The role of serglycin
Proteoglycans containing glycosaminoglycan (GAG) side chains of either heparin or chondroitin sulfate are central components of MC SGs, with serglycin serving as the core protein. Serglycin features an extended Ser-Gly repeat region, in which each Ser-Gly unit provides a potential GAG attachment site (Rönnberg et al., 2012). Strikingly, knockout of serglycin impairs the storage of proteases and histamine leading to disorganized SGs (Abrink et al., 2004), highlighting serglycin’s key role in the retention of proteases and histamine within SGs. Interactions between serglycin and SG proteins prevent premature degradation and regulate the kinetics of their diffusion into the extracellular milieu following triggered exocytosis. Cargo with a high affinity for serglycin is retained near the SG, while low-affinity cargo, such as β-hexosaminidase, diffuses away into the circulation. Based on our findings on SG fission, we hypothesize that serglycin may also contribute to the well-documented heterogeneity of SGs. This could occur through the unequal distribution of granule contents between budding SGs, driven by differential binding affinities of cargo molecules to serglycin (Rönnberg et al., 2012). Interestingly, deficiency of serglycin-dependent proteases reduces the amount of heparin, replicating the phenotype of serglycin deficiency by causing a major distortion in SG integrity, presumably due to a disruption in the SG’s electric charge balance (Grujic et al., 2013).
7.2 The role of acidic pH
Significant morphological changes were also observed in MCs treated with bafilomycin A1, an inhibitor of the vacuolar-type ATPase proton pump (Pejler et al., 2017). The granules became swollen and acquired a vacuole-like morphology (Pejler et al., 2017). Bafilomycin A1 also had selective effects on SG cargo, altering the processing of pro-carboxypeptidase A3, reducing the level of SG-stored histamine, and enhancing the autoproteolysis of tryptase (Pejler et al., 2017). In contrast, the storage of β-hexosaminidase was unaffected. Therefore, a low SG pH is essential for maintaining the homeostasis of MC SGs (Pejler et al., 2017).
8 The impact of aging
MC numbers increase in aged tissue and changes have been recorded in their responsiveness and ability to degranulate (Chatterjee and Gashev, 2012; Pilkington et al., 2019). To gain insights into the autonomous changes that may occur in MCs during aging, we recently established a novel model of inducible senescence in MCs as a paradigm of aging (Kleeblatt et al., 2024). This model is based on the inducible upregulation of the cell cycle inhibitor p16INK4A, which we have also shown to be upregulated in human skin derived from elderly donors and in peritoneal MCs derived from old mice (Kleeblatt et al., 2024). Analyses of in vitro-differentiated MCs derived from the bone marrow of these transgenic mice revealed significant morphological and functional differences in the SGs of senescent MCs. These differences were reflected in a significant increase in large SGs containing intraluminal vesicles (ILVs), which was associated with a shift towards the regulated release of smaller, CD63-enriched extracellular vesicles (EVs), reminiscent of the functional changes observed following SG fusion with amphisomes (Omari et al., 2024). Interestingly, this increase in the release of small CD63-positive EVs was also associated with an increase in proteoglycan exteriorisation, while the ability to release β-hexosaminidase decreased during prolonged senescence (Kleeblatt et al., 2024).
9 Conclusion and perspectives
LROs were traditionally defined as a subtype of SGs that exhibit lysosomal features. However, this group of organelles encompasses a variety of structures that appear to differ in their mechanisms of biogenesis. Furthermore, the boundaries between conventional SGs, LROs and lysosomes have become more fluid, as accumulating data demonstrate the involvement of both the endocytic and autophagic systems in the biogenesis of endocrine and exocrine SGs (Patel et al., 2013; Bel et al., 2017; Morishita et al., 2020; Li et al., 2022). Furthermore, classical lysosomes can also undergo exocytosis (Trojani et al., 2024). Therefore, while LROs may not necessarily share common mechanisms for their biogenesis, some of these mechanisms might be shared with conventional SGs, classical lysosomes, or autolysosomes. Deciphering the molecular details of the interactions between MC SGs and the endolysosomal and autophagic systems could contribute to our understanding of these processes in other cells. In this review, we primarily focused on factors shown to play a role in the biogenesis of MC SGs. However, many questions remain unanswered. For example, how are proteins targeted to the SGs? Do specific sorting signals direct SG cargo to SGs rather than lysosomes, or do lipid phase separations play a role in cargo targeting? What is the precise role of CD63, PI4K, and PTPN9 in mediating SG fusion? What is the exact role of Lyst or synaptotagmins? What is the precise mechanism of SG fission, and which lipid kinases and phosphatases control the switch between fusion and fission? What is the relationship between SGs and degradative endolysosomes or autolysosomes? How do Rab GTPases that affect SG size or exocytosis execute their regulatory functions? What is the precise mechanism of SG recapture and regranulation? Finally, what is the precise mechanism of SG exocytosis? a process that remains incompletely understood. Future studies, leveraging novel technologies and tools, will need to address these fundamental questions to better understand the functions of MCs in health and disease.
Statements
Author contributions
RS-E: Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by grant 1600/19 from the Israel Science Foundation, founded by the Israel Academy for Sciences (to R. Sagi-Eisenberg), and grants 2017182 (to R. Sagi-Eisenberg and S. J. Galli) and 2021172 (to R. Sagi-Eisenberg and H. Ali) by the United States–Israel Binational Science Foundation.
Acknowledgments
I am grateful to my lab members (Yaara Gorzalczany, Sewar Omari, Priyangana Deb, Daniele Ventriglia, Shahar Doron and Ran Perlman) for proof-reading the article.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Abrink M. Grujic M. Pejler G. (2004). Serglycin is essential for maturation of mast cell secretory granule. J. Biol. Chem.279, 40897–40905. 10.1074/jbc.M405856200
2
Adhikari P. Ayo T. E. Vines J. C. Sugita S. Xu H. (2023). Exocytic machineries differentially control mediator release from allergen-triggered RBL-2H3 cells. Inflamm. Res.72, 639–649. 10.1007/s00011-023-01698-z
3
Akula S. Paivandy A. Fu Z. Thorpe M. Pejler G. Hellman L. (2020). Quantitative in-depth analysis of the mouse mast cell transcriptome reveals organ-specific mast cell heterogeneity. Cells9, 211. 10.3390/cells9010211
4
Ali H. (2021). Revisiting the role of MRGPRX2 on hypersensitivity reactions to neuromuscular blocking drugs. Curr. Opin. Immunol.72, 65–71. 10.1016/j.coi.2021.03.011
5
Azouz N. P. Matsui T. Fukuda M. Sagi-Eisenberg R. (2012). Decoding the regulation of mast cell exocytosis by networks of Rab GTPases. J. Immunol.189, 2169–2180. 10.4049/jimmunol.1200542
6
Azouz N. P. Zur N. Efergan A. Ohbayashi N. Fukuda M. Amihai D. et al (2014). Rab5 is a novel regulator of mast cell secretory granules: impact on size, cargo, and exocytosis. J. Immunol.192, 4043–4053. 10.4049/jimmunol.1302196
7
Banushi B. Simpson F. (2022). Overlapping machinery in lysosome-related organelle trafficking: a lesson from rare multisystem disorders. Cells11, 3702. 10.3390/cells11223702
8
Bao C. Abraham S. N. (2024). Mast cell-sensory neuron crosstalk in allergic diseases. J. Allergy Clin. Immunol.153, 939–953. 10.1016/j.jaci.2024.02.005
9
Baram D. Adachi R. Medalia O. Tuvim M. Dickey B. F. Mekori Y. A. et al (1999). Synaptotagmin II negatively regulates Ca2+-triggered exocytosis of lysosomes in mast cells. J. Exp. Med.189, 1649–1658. 10.1084/jem.189.10.1649
10
Begley M. Aragon M. Baker R. W. (2024). A structure-based mechanism for initiation of AP-3 coated vesicle formation. Proc. Natl. Acad. Sci. U. S. A.121, e2411974121. 10.1073/pnas.2411974121
11
Bel S. Pendse M. Wang Y. Li Y. Ruhn K. A. Hassell B. et al (2017). Paneth cells secrete lysozyme via secretory autophagy during bacterial infection of the intestine. Science357, 1047–1052. 10.1126/science.aal4677
12
Blank U. Huang H. Kawakami T. (2021). The high affinity IgE receptor: a signaling update. Curr. Opin. Immunol.72, 51–58. 10.1016/j.coi.2021.03.015
13
Blank U. Madera-Salcedo I. K. Danelli L. Claver J. Tiwari N. SáNCHEZ-Miranda E. et al (2014). Vesicular trafficking and signaling for cytokine and chemokine secretion in mast cells. Front. Immunol.5, 453. 10.3389/fimmu.2014.00453
14
Bonifacino J. S. Yuan L. Sandoval I. V. (1989). Internalization and recycling to serotonin-containing granules of the 80K integral membrane protein exposed on the surface of secreting rat basophilic leukaemia cells. J. Cell Sci.92 (Pt 4), 701–712. 10.1242/jcs.92.4.701
15
Bowman S. L. Bi-Karchin J. Le L. Marks M. S. (2019). The road to lysosome-related organelles: insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic20, 404–435. 10.1111/tra.12646
16
Bultema J. J. Di Pietro S. M. (2013). Cell type-specific Rab32 and Rab38 cooperate with the ubiquitous lysosome biogenesis machinery to synthesize specialized lysosome-related organelles. Small GTPases4, 16–21. 10.4161/sgtp.22349
17
Chatterjee V. Gashev A. A. (2012). Aging-associated shifts in functional status of mast cells located by adult and aged mesenteric lymphatic vessels. Am. J. Physiol. Heart Circ. Physiol.303, H693–H702. 10.1152/ajpheart.00378.2012
18
Chia S. L. Kapoor S. Carvalho C. Bajénoff M. Gentek R. (2023). Mast cell ontogeny: from fetal development to life-long health and disease. Immunol. Rev.315, 31–53. 10.1111/imr.13191
19
Combs J. W. (1971). An electron microscope study of mouse mast cells arising in vivo and in vitro. J. Cell Biol.48, 676–684. 10.1083/jcb.48.3.676
20
Dacks J. B. Robinson M. S. (2017). Outerwear through the ages: evolutionary cell biology of vesicle coats. Curr. Opin. Cell Biol.47, 108–116. 10.1016/j.ceb.2017.04.001
21
Dahlin J. S. Maurer M. Metcalfe D. D. Pejler G. Sagi-Eisenberg R. Nilsson G. (2022). The ingenious mast cell: contemporary insights into mast cell behavior and function. Allergy77, 83–99. 10.1111/all.14881
22
Daniele T. Hackmann Y. Ritter A. T. Wenham M. Booth S. Bossi G. et al (2011). A role for Rab7 in the movement of secretory granules in cytotoxic T lymphocytes. Traffic12, 902–911. 10.1111/j.1600-0854.2011.01194.x
23
Da Silva E. Z. Freitas-Filho E. G. De Souza-Júnior D. A. Dasilva L. L. Jamur M. C. Oliver C. (2017). Adaptor protein-3: a key player in RBL-2H3 mast cell mediator release. PLoS One12, e0173462. 10.1371/journal.pone.0173462
24
Delevoye C. Marks M. S. Raposo G. (2019). Lysosome-related organelles as functional adaptations of the endolysosomal system. Curr. Opin. Cell Biol.59, 147–158. 10.1016/j.ceb.2019.05.003
25
Dell'Angelica E. C. (2009). AP-3-dependent trafficking and disease: the first decade. Curr. Opin. Cell Biol.21, 552–559. 10.1016/j.ceb.2009.04.014
26
Dell'Angelica E. C. Mullins C. Caplan S. Bonifacino J. S. (2000). Lysosome-related organelles. Faseb J.14, 1265–1278. 10.1096/fj.14.10.1265
27
Dema B. Suzuki R. Rivera J. (2014). Rethinking the role of immunoglobulin E and its high-affinity receptor: new insights into allergy and beyond. Int. Arch. Allergy Immunol.164, 271–279. 10.1159/000365633
28
Efergan A. Azouz N. P. Klein O. Noguchi K. Rothenberg M. E. Fukuda M. et al (2016). Rab12 regulates retrograde transport of mast cell secretory granules by interacting with the RILP-dynein complex. J. Immunol.196, 1091–1101. 10.4049/jimmunol.1500731
29
Falcone F. H. Wan D. Barwary N. Sagi-Eisenberg R. (2018). RBL cells as models for in vitro studies of mast cells and basophils. Immunol. Rev.282, 47–57. 10.1111/imr.12628
30
Flašker A. Jorgačevski J. Calejo A. I. Kreft M. Zorec R. (2013). Vesicle size determines unitary exocytic properties and their sensitivity to sphingosine. Mol. Cell Endocrinol.376, 136–147. 10.1016/j.mce.2013.06.012
31
Gordon J. R. Galli S. J. (1990). Mast cells as a source of both preformed and immunologically inducible TNF-alpha/cachectin. Nature346, 274–276. 10.1038/346274a0
32
Gri G. Frossi B. D'Inca F. Danelli L. Betto E. Mion F. et al (2012). Mast cell: an emerging partner in immune interaction. Front. Immunol.3, 120. 10.3389/fimmu.2012.00120
33
Grimberg E. Peng Z. Hammel I. Sagi-Eisenberg R. (2003). Synaptotagmin III is a critical factor for the formation of the perinuclear endocytic recycling compartment and determination of secretory granules size. J. Cell Sci.116, 145–154. 10.1242/jcs.00186
34
Grujic M. Calounova G. Eriksson I. Feyerabend T. Rodewald H. R. Tchougounova E. et al (2013). Distorted secretory granule composition in mast cells with multiple protease deficiency. J. Immunol.191, 3931–3938. 10.4049/jimmunol.1301441
35
Haberman Y. Grimberg E. Fukuda M. Sagi-Eisenberg R. (2003). Synaptotagmin IX, a possible linker between the perinuclear endocytic recycling compartment and the microtubules. J. Cell Sci.116, 4307–4318. 10.1242/jcs.00719
36
Haberman Y. Ziv I. Gorzalczany Y. Hirschberg K. Mittleman L. Fukuda M. et al (2007). Synaptotagmin (Syt) IX is an essential determinant for protein sorting to secretory granules in mast cells. Blood109, 3385–3392. 10.1182/blood-2006-07-033126
37
Hammel I. Lagunoff D. Galli S. J. (2010). Regulation of secretory granule size by the precise generation and fusion of unit granules. J. Cell Mol. Med.14, 1904–1916. 10.1111/j.1582-4934.2010.01071.x
38
Hendriksen E. Van Bergeijk D. Oosting R. S. Redegeld F. A. (2017). Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav Rev.79, 119–133. 10.1016/j.neubiorev.2017.05.001
39
Huang X. Lan Z. Hu Z. (2024). Role and mechanisms of mast cells in brain disorders. Front. Immunol.15, 1445867. 10.3389/fimmu.2024.1445867
40
Hui K. Y. Fernandez-Hernandez H. Hu J. Schaffner A. Pankratz N. Hsu N. Y. et al (2018). Functional variants in the LRRK2 gene confer shared effects on risk for Crohn's disease and Parkinson's disease. Sci. Transl. Med.10, eaai7795. 10.1126/scitranslmed.aai7795
41
Jamur M. C. Vugman I. (1990). Cytochemical demonstration of acid phosphatase, trimetaphosphatase and basic protein in rat peritoneal mast cells during 48/80 induced exocytosis. Cell Mol. Biol.36 (5), 509–514.
42
Jiménez M. Cervantes-García D. Córdova-Dávalos L. E. Pérez-Rodríguez M. J. Gonzalez-Espinosa C. Salinas E. (2021). Responses of mast cells to pathogens: beneficial and detrimental roles. Front. Immunol.12, 685865. 10.3389/fimmu.2021.685865
43
Johnson R. G. Carty S. E. Fingerhood B. J. Scarpa A. (1980). The internal pH of mast cell granules. FEBS Lett.120, 75–79. 10.1016/0014-5793(80)81050-7
44
Kirshenbaum A. S. Cruse G. Desai A. Bandara G. Leerkes M. Lee C. C. et al (2016). Immunophenotypic and ultrastructural analysis of mast cells in hermansky-pudlak syndrome type-1: a possible connection to pulmonary fibrosis. PLoS One11, e0159177. 10.1371/journal.pone.0159177
45
Kirshenbaum A. S. Goff J. P. Semere T. Foster B. Scott L. M. Metcalfe D. D. (1999). Demonstration that human mast cells arise from a progenitor cell population that is CD34(+), c-kit(+), and expresses aminopeptidase N (CD13). Blood94, 2333–2342. 10.1182/blood.v94.7.2333.419k30_2333_2342
46
Kiyoi T. Liu S. Sahid M. N. A. Shudou M. Ogasawara M. Mogi M. et al (2019). Morphological and functional analysis of beige (Chèdiak-Higashi syndrome) mouse mast cells with giant granules. Int. Immunopharmacol.69, 202–212. 10.1016/j.intimp.2019.01.053
47
Kleeblatt E. Lazki-Hagenbach P. Nabet E. Cohen R. Bahri R. Rogers N. et al (2024). p16(Ink4a)-induced senescence in cultured mast cells as a model for ageing reveals significant morphological and functional changes. Immun. Ageing21, 77. 10.1186/s12979-024-00478-5
48
Kruger J. M. Fukushima T. Cherepanov V. Borregaard N. Loeve C. Shek C. et al (2002). Protein-tyrosine phosphatase MEG2 is expressed by human neutrophils. Localization to the phagosome and activation by polyphosphoinositides. J. Biol. Chem.277, 2620–2628. 10.1074/jbc.M104550200
49
Kurowska M. Goudin N. Nehme N. T. Court M. Garin J. Fischer A. et al (2012). Terminal transport of lytic granules to the immune synapse is mediated by the kinesin-1/Slp3/Rab27a complex. Blood119, 3879–3889. 10.1182/blood-2011-09-382556
50
Lazki-Hagenbach P. Kleeblatt E. Ali H. Sagi-Eisenberg R. (2022). Spatiotemporal patterns of substance P-bound MRGPRX2 reveal a novel connection between macropinosome resolution and secretory granule regeneration in mast cells. Front. Immunol.13, 892239. 10.3389/fimmu.2022.892239
51
Lazki-Hagenbach P. Kleeblatt E. Fukuda M. Ali H. Sagi-Eisenberg R. (2024). The underlying Rab network of MRGPRX2-stimulated secretion unveils the impact of receptor trafficking on secretory granule biogenesis and secretion. Cells13, 93. 10.3390/cells13010093
52
Lin C. J. Herisson F. Le H. Jaafar N. Chetal K. Oram M. K. et al (2023). Mast cell deficiency improves cognition and enhances disease-associated microglia in 5XFAD mice. Cell Rep.42, 113141. 10.1016/j.celrep.2023.113141
53
Li X. Wang L. Hao J. Zhu Q. Guo M. Wu C. et al (2022). The role of autophagy in lamellar body formation and surfactant production in type 2 alveolar epithelial cells. Int. J. Biol. Sci.18, 1107–1119. 10.7150/ijbs.64285
54
Longé C. Bratti M. Kurowska M. Vibhushan S. David P. Desmeure V. et al (2022). Rab44 regulates murine mast cell-driven anaphylaxis through kinesin-1-dependent secretory granule translocation. J. Allergy Clin. Immunol.150, 676–689. 10.1016/j.jaci.2022.04.009
55
Lorentz A. Baumann A. Vitte J. Blank U. (2012). The SNARE machinery in mast cell secretion. Front. Immunol.3, 143. 10.3389/fimmu.2012.00143
56
Luzio J. P. Hackmann Y. Dieckmann N. M. Griffiths G. M. (2014). The biogenesis of lysosomes and lysosome-related organelles. Cold Spring Harb. Perspect. Biol.6, a016840. 10.1101/cshperspect.a016840
57
Marks M. S. Heijnen H. F. Raposo G. (2013). Lysosome-related organelles: unusual compartments become mainstream. Curr. Opin. Cell Biol.25, 495–505. 10.1016/j.ceb.2013.04.008
58
Matsui T. Ohbayashi N. Fukuda M. (2012). The Rab interacting lysosomal protein (RILP) homology domain functions as a novel effector domain for small GTPase Rab36: Rab36 regulates retrograde melanosome transport in melanocytes. J. Biol. Chem.287, 28619–28631. 10.1074/jbc.M112.370544
59
Mcneil B. D. Pundir P. Meeker S. Han L. Undem B. J. Kulka M. et al (2015). Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature519, 237–241. 10.1038/nature14022
60
Mekori Y. A. Oh C. K. Metcalfe D. D. (1995). The role of c-Kit and its ligand, stem cell factor, in mast cell apoptosis. Int. Arch. Allergy Immunol.107, 136–138. 10.1159/000236955
61
Mencarelli A. Bist P. Choi H. W. Khameneh H. J. Mortellaro A. Abraham S. N. (2024). Anaphylactic degranulation by mast cells requires the mobilization of inflammasome components. Nat. Immunol.25, 693–702. 10.1038/s41590-024-01788-y
62
Metcalfe D. D. Baram D. Mekori Y. A. (1997). Mast cells. Physiol. Rev.77, 1033–1079. 10.1152/physrev.1997.77.4.1033
63
Moon T. C. Befus A. D. Kulka M. (2014). Mast cell mediators: their differential release and the secretory pathways involved. Front. Immunol.5, 569. 10.3389/fimmu.2014.00569
64
Morishita H. Kanda Y. Kaizuka T. Chino H. Nakao K. Miki Y. et al (2020). Autophagy is required for maturation of surfactant-containing lamellar bodies in the lung and swim bladder. Cell Rep.33, 108477. 10.1016/j.celrep.2020.108477
65
Munoz I. Danelli L. Claver J. Goudin N. Kurowska M. Madera-Salcedo I. K. et al (2016). Kinesin-1 controls mast cell degranulation and anaphylaxis through PI3K-dependent recruitment to the granular Slp3/Rab27b complex. J. Cell Biol.215, 203–216. 10.1083/jcb.201605073
66
Nagata Y. Suzuki R. (2022). FcεRI: a master regulator of mast cell functions. Cells11, 622. 10.3390/cells11040622
67
Nakano H. Ushio H. (2011). An unexpected role for autophagy in degranulation of mast cells. Autophagy7, 657–659. 10.4161/auto.7.6.15384
68
Omari S. Roded A. Eisenberg M. Ali H. Fukuda M. Galli S. J. et al (2024). Mast cell secretory granule fusion with amphisomes coordinates their homotypic fusion and release of exosomes. Cell Rep.43, 114482. 10.1016/j.celrep.2024.114482
69
Patel K. K. Miyoshi H. Beatty W. L. Head R. D. Malvin N. P. Cadwell K. et al (2013). Autophagy proteins control goblet cell function by potentiating reactive oxygen species production. Embo J.32, 3130–3144. 10.1038/emboj.2013.233
70
Pejler G. Hu Frisk J. M. Sjöström D. Paivandy A. Öhrvik H. (2017). Acidic pH is essential for maintaining mast cell secretory granule homeostasis. Cell Death Dis.8, e2785. 10.1038/cddis.2017.206
71
Pilkington S. M. Barron M. J. Watson R. E. B. Griffiths C. E. M. Bulfone-Paus S. (2019). Aged human skin accumulates mast cells with altered functionality that localize to macrophages and vasoactive intestinal peptide-positive nerve fibres. Br. J. Dermatol180, 849–858. 10.1111/bjd.17268
72
Prussin C. Metcalfe D. D. (2006). 5. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol.117, S450–S456. 10.1016/j.jaci.2005.11.016
73
Puri N. Roche P. A. (2008). Mast cells possess distinct secretory granule subsets whose exocytosis is regulated by different SNARE isoforms. Proc. Natl. Acad. Sci. U. S. A.105, 2580–2585. 10.1073/pnas.0707854105
74
Raposo G. Tenza D. Mecheri S. Peronet R. Bonnerot C. Desaymard C. (1997). Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol. Biol. Cell8, 2631–2645. 10.1091/mbc.8.12.2631
75
Reber L. L. Sibilano R. Mukai K. Galli S. J. (2015). Potential effector and immunoregulatory functions of mast cells in mucosal immunity. Mucosal Immunol.8, 444–463. 10.1038/mi.2014.131
76
Rocha N. Kuijl C. Van Der Kant R. Janssen L. Houben D. Janssen H. et al (2009). Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning. J. Cell Biol.185, 1209–1225. 10.1083/jcb.200811005
77
Rönnberg E. Melo F. R. Pejler G. (2012). Mast cell proteoglycans. J. Histochem Cytochem60, 950–962. 10.1369/0022155412458927
78
Sandhu J. K. Kulka M. (2021). Decoding mast cell-microglia communication in neurodegenerative diseases. Int. J. Mol. Sci.22, 1093. 10.3390/ijms22031093
79
Sannes P. L. Spicer S. S. (1979). The heterophagic granules of mast cells: dipeptidyl aminopeptidase II activity and resistance to exocytosis. Am. J. Pathol.94 (3), 447–458.
80
Schwartz L. B. Austen K. F. (1980). Enzymes of the mast cell granule. J. Invest Dermatol74, 349–353. 10.1111/1523-1747.ep12543620
81
Segura-Villalobos D. Ramírez-Moreno I. G. Martínez-Aguilar M. Ibarra-Sánchez A. Muñoz-Bello J. O. Anaya-Rubio I. et al (2022). Mast cell-tumor interactions: molecular mechanisms of recruitment, intratumoral communication and potential therapeutic targets for tumor growth. Cells11, 349. 10.3390/cells11030349
82
Serra-Vinardell J. Sandler M. B. De Pace R. Manzella-Lapeira J. Cougnoux A. Keyvanfar K. et al (2023). LYST deficiency impairs autophagic lysosome reformation in neurons and alters lysosome number and size. Cell Mol. Life Sci.80, 53. 10.1007/s00018-023-04695-x
83
Shiflett S. L. Kaplan J. Ward D. M. (2002). Chediak-Higashi Syndrome: a rare disorder of lysosomes and lysosome related organelles. Pigment. Cell Res.15, 251–257. 10.1034/j.1600-0749.2002.02038.x
84
Smith A. J. Pfeiffer J. R. Zhang J. Martinez A. M. Griffiths G. M. Wilson B. S. (2003). Microtubule-dependent transport of secretory vesicles in RBL-2H3 cells. Traffic4, 302–312. 10.1034/j.1600-0854.2003.00084.x
85
Starcevic M. Nazarian R. Dell'Angelica E. C. (2002). The molecular machinery for the biogenesis of lysosome-related organelles: lessons from Hermansky-Pudlak syndrome. Semin. Cell Dev. Biol.13, 271–278. 10.1016/s1084952102000563
86
St John A. L. Abraham S. N. (2013). Innate immunity and its regulation by mast cells. J. Immunol.190, 4458–4463. 10.4049/jimmunol.1203420
87
Suárez-Quian C. A. (1987). The distribution of four lysosomal integral membrane proteins (LIMPs) in rat basophilic leukemia cells. Tissue Cell19, 495–504. 10.1016/0040-8166(87)90043-7
88
Subramanian H. Gupta K. Ali H. (2016). Roles of Mas-related G protein-coupled receptor X2 on mast cell-mediated host defense, pseudoallergic drug reactions, and chronic inflammatory diseases. J. Allergy Clin. Immunol.138, 700–710. 10.1016/j.jaci.2016.04.051
89
Tauber M. Basso L. Martin J. Bostan L. Pinto M. M. Thierry G. R. et al (2023). Landscape of mast cell populations across organs in mice and humans. J. Exp. Med.220, e20230570. 10.1084/jem.20230570
90
Theoharides T. C. Petra A. I. Taracanova A. Panagiotidou S. Conti P. (2015). Targeting IL-33 in autoimmunity and inflammation. J. Pharmacol. Exp. Ther.354, 24–31. 10.1124/jpet.114.222505
91
Theoharides T. C. Twahir A. Kempuraj D. (2024). Mast cells in the autonomic nervous system and potential role in disorders with dysautonomia and neuroinflammation. Ann. Allergy Asthma Immunol.132, 440–454. 10.1016/j.anai.2023.10.032
92
Toyoshima S. Okayama Y. (2022). Neuro-allergology: mast cell-nerve cross-talk. Allergol. Int.71, 288–293. 10.1016/j.alit.2022.04.002
93
Trojani M. C. Santucci-Darmanin S. Breuil V. Carle G. F. Pierrefite-Carle V. (2024). Lysosomal exocytosis: from cell protection to protumoral functions. Cancer Lett.597, 217024. 10.1016/j.canlet.2024.217024
94
Turner M. E. Che J. Mirhaidari G. J. M. Kennedy C. C. Blum K. M. Rajesh S. et al (2024). The lysosomal trafficking regulator “LYST”: an 80-year traffic jam. Front. Immunol.15, 1404846. 10.3389/fimmu.2024.1404846
95
Vitte J. Vibhushan S. Bratti M. Montero-Hernandez J. E. Blank U. (2022). Allergy, anaphylaxis, and nonallergic hypersensitivity: IgE, mast cells, and beyond. Med. Princ. Pract.31, 501–515. 10.1159/000527481
96
Vukoja A. Rey U. Petzoldt A. G. Ott C. Vollweiter D. Quentin C. et al (2018). Presynaptic biogenesis requires axonal transport of lysosome-related vesicles. Neuron99, 1216–1232.e7. 10.1016/j.neuron.2018.08.004
97
Wang X. Zhang P. Tang Y. Chen Y. Zhou E. Gao K. (2024). Mast cells: a double-edged sword in inflammation and fibrosis. Front. Cell Dev. Biol.12, 1466491. 10.3389/fcell.2024.1466491
98
Wei M. L. (2006). Hermansky-Pudlak syndrome: a disease of protein trafficking and organelle function. Pigment. Cell Res.19, 19–42. 10.1111/j.1600-0749.2005.00289.x
99
Wernersson S. Pejler G. (2014). Mast cell secretory granules: armed for battle. Nat. Rev. Immunol.14, 478–494. 10.1038/nri3690
100
West P. W. Bulfone-Paus S. (2022). Mast cell tissue heterogeneity and specificity of immune cell recruitment. Front. Immunol.13, 932090. 10.3389/fimmu.2022.932090
101
Wolfes A. C. Dean C. (2020). The diversity of synaptotagmin isoforms. Curr. Opin. Neurobiol.63, 198–209. 10.1016/j.conb.2020.04.006
102
Xu H. Bin N. R. Sugita S. (2018). Diverse exocytic pathways for mast cell mediators. Biochem. Soc. Trans.46, 235–247. 10.1042/BST20170450
103
Xu K. Williams R. M. Holowka D. Baird B. (1998). Stimulated release of fluorescently labeled IgE fragments that efficiently accumulate in secretory granules after endocytosis in RBL-2H3 mast cells. J. Cell Sci.111 (Pt 16), 2385–2396. 10.1242/jcs.111.16.2385
104
Zhang B. Asadi S. Weng Z. Sismanopoulos N. Theoharides T. C. (2012). Stimulated human mast cells secrete mitochondrial components that have autocrine and paracrine inflammatory actions. PLoS One7, e49767. 10.1371/journal.pone.0049767
Summary
Keywords
mast cells, lysosome related organelles (LRO), secretory granules, amphisomes, fusion, fission
Citation
Sagi-Eisenberg R (2025) Biogenesis and homeostasis of mast cell lysosome related secretory granules. Front. Cell Dev. Biol. 13:1603999. doi: 10.3389/fcell.2025.1603999
Received
01 April 2025
Accepted
12 May 2025
Published
23 May 2025
Volume
13 - 2025
Edited by
Subba Rao Gangi Setty, Indian Institute of Science (IISc), India
Reviewed by
Gael Ménasché, Institut National de la Santé et de la Recherche Médicale (INSERM), France
Michel Simon, Université de Toulouse, France
Updates
Copyright
© 2025 Sagi-Eisenberg.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ronit Sagi-Eisenberg, histol3@tauex.tau.ac.il
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.