 Payal Patel
Payal Patel Joy Ku
Joy Ku Ike Uzoaru2
Ike Uzoaru2 Jeffery A. Goldstein
Jeffery A. Goldstein- 1Carle Illinois College of Medicine, Urbana, IL, United States
- 2Department of Pathology, Carle Foundation Hospital, Urbana, IL, United States
- 3Department of Pathology, Northwestern University Feinberg School of Medicine and Northwestern Memorial Hospital, Chicago, IL, United States
Neurodevelopment shapes how children think, move, and engage with their surroundings. Understanding the pathways underlying neurodevelopmental pathophysiology in the perinatal stage can inform intervention strategies to mitigate or reduce the severity and extent of developmental brain injuries. Early risk stratification enables timely therapies and resource planning during a critical period for the developing brain. Over the past decade, attention has turned to the placenta as a uniquely informative vantage point for the identification of pregnancies at high risk for adverse neurodevelopmental outcomes. Situated at the maternal-fetal interface, the placenta functions as a dynamic record of intrauterine conditions, integrating genetic and environmental signals into distinct and quantifiable biomarkers. Emerging evidence indicates these placental biomarkers may predict later neurodevelopmental outcomes, highlighting the organ’s value in precision perinatal care. With this in mind, the objective of this scoping review will be to investigate the current use of placental biomarkers as predictors of neurodevelopmental outcomes in clinical practice, particularly the trisomies (T13, T18, T21). In the second section of this paper, we will focus on recent advancements and improvements in the use of placental biomarkers for diagnostic and prognostic purposes in other neurodevelopmental outcomes. Finally, this article concludes with a discussion of the impact of neuroplacentology in protocol development, risk stratification, and psychosocial wellness of pregnant women.
1 Background
As the first fetal organ to develop, the placenta defines the intrauterine environment and regulates maternal-fetal communication through nutrient exchange, immune modulation and endocrine signaling (Burton and Fowden, 2015). The Developmental Origins of Health and Disease (DOHaD) theory emphasizes the pivotal role of the placenta in shaping long-term outcomes for childhood health development (Ganguly et al., 2020).
Within this framework, neuroplacentology is an emerging field focused on exploring how placental structure and function influence fetal and neonatal neurodevelopment (Kratimenos et al., 2019). Abnormal placental signaling due to hypoxia, inflammation, or disrupted hormone regulation has been implicated in perinatal brain injuries and is associated with increased risk of neurodevelopmental disorders (Gall et al., 2022; Espinoza et al., 2022; Zhou et al., 2023; Vacher et al., 2021).
Given this connection, perinatal biomarkers have been established for the testing of fetal diseases. In one example, the American College of Medical Genetics and Genomics (ACMG) recommends the use of noninvasive prenatal testing (NIPT) for fetal chromosome abnormalities (Trisomy 13, 18, and 21) (Page et al., 2017). NIPT utilizes cell free fetal DNA (cffDNA) extracted from maternal plasma, which undergoes whole genome sequencing to screen for fetal aneuploidy (Nelissen et al., 2011).
In neuroplacentology, researchers are studying epigenetic markers and autoantibodies to develop minimally invasive liquid biopsy methods to determine a fetus’s risk for neurodevelopmental conditions.
Within the scope of this review, we will look at specific types of biomarkers that are used for the prediction and prognosis of neurodevelopmental outcomes. While the field also focuses on physiological factors that affect fetal development, such as maternal preeclampsia, our discussion centers around the communication systems themselves and how they may induce pathological developmental patterns.
2 Methods
We conducted a scoping review in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses for Scoping Reviews (PRISMA-ScR) reporting guidelines. To identify potentially relevant documents, the following bibliographic databases were searched from April 1990 to 14 March 2025: PubMed and Scopus. The final search strategy can be found in Supplementary Table 1, and the list of papers included within the final review found in Supplementary Data Sheet 1.
2.1 Eligibility criteria
Studies that examined the relationship between placenta-derived biomarkers in the context of adverse neurodevelopmental outcomes during the perinatal period in human subjects and written in English were included. Any type of review article, case reports, or editorials were excluded.
As defined by the U.S. Food and Drug Administration and National Institutes of Health (FDA-NIH) Joint Leadership Council, biomarkers are any measured biological end-point which, depending on their source, can be molecular, histologic, radiographic, or physiologic (FDA-NIH Biomarker Working Group, 2016). Biomarkers are used for a myriad of purposes, including diagnostics, disease prediction, and prognostics; thus, we kept our definition of biomarkers open to allow for a wide range of investigations within neuroplacentology.
Maternal conditions (e.g., preeclampsia, diet, stress), delivery complications, and environmental toxicants (e.g., lead, mercury) are exposures or risk factors that may drive placental dysfunction, but are not biomarkers themselves. Studies focused solely on these exposures were excluded in order to reduce confounding factors impacting neurodevelopmental outcomes in the neonate.
Several placental biomarkers associated with maternal conditions (e.g., PIGF for preeclampsia) have been clinically implemented, but diagnostic biomarkers specifically predictive of neurodevelopmental delay remain largely unexplored. Additionally, the goal is to examine signals originating from the placenta, so studies that directly tested for infectious organisms or environmental pollutants were excluded as well.
2.2 Data extraction
The final search results were exported into Zotero and duplicates were removed by the application. Titles, abstracts and full texts were screened by two investigators (PP and JK). In an effort to increase consistency among reviewers, both reviewers screened the same records. Sequential evaluation of titles, abstracts, and full texts of all identified publications was conducted, with any disagreements on study selection resolved by consensus between the two reviewers.
3 Results
The initial search criteria outlined above yielded 535 studies, which were reduced to 479 after removing duplicates, 221 after title and abstract screening, and ultimately 106 following review of full texts for study methods. A detailed breakdown is reported in the PRISMA flow diagram Figure 1. We grouped the studies by the type of neurodevelopmental disorder (NDD) they analyzed, organized by date of publication, and summarized the studied biomarker, outcome measures, and key results for each study.
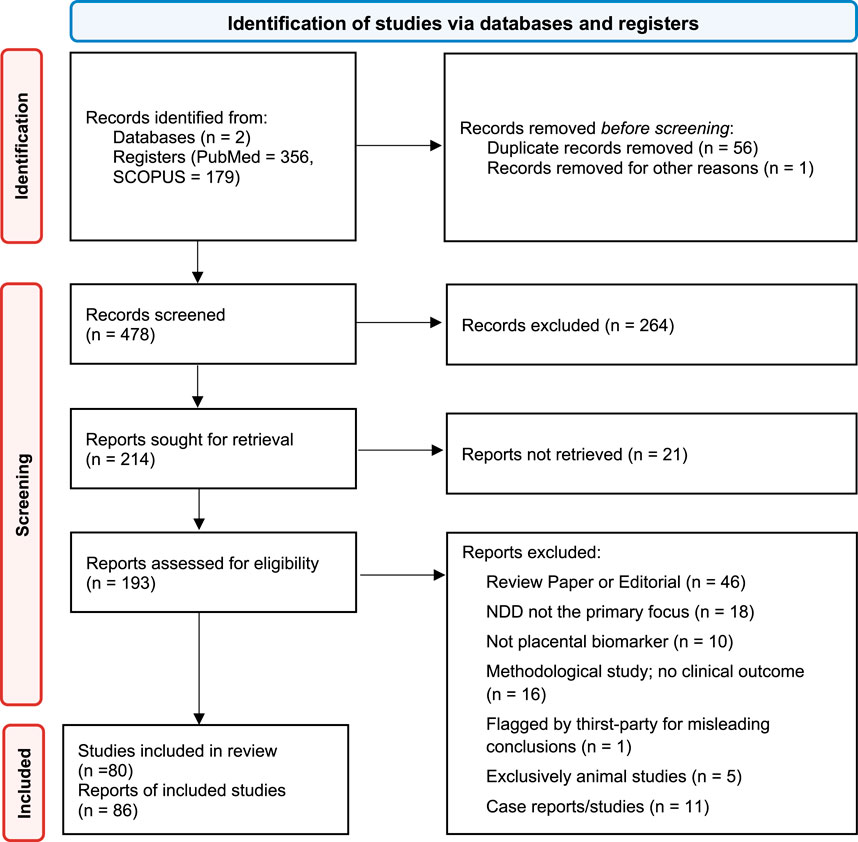
Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analyses for Scoping Reviews (PRISMA-ScR) flow diagram.
3.1 Trisomy 21 - A case study for placental biomarker-based screening
Trisomy 21 (T21) is one of the most commonly screened prenatal conditions, with the American College of Obstetricians and Gynecologists recommending that all pregnant individuals, regardless of age or risk factors, be offered screening and diagnostic testing for T21 (Mj et al., 2022). With over 40 years of collaboration by teams across the world, prenatal testing for T21 can be seen as a model for the progression and development of prenatal screening for NDDs. In addition, almost half of the studies identified in this review are pertaining to T21, with a few also describing the use for other trisomies (T13, T18) (Pertl et al., 1999; Jauniaux et al., 2000; Metzenbauer et al., 2002). As such, we will focus on describing developments within biomarkers for T21 separately from other neurodevelopmental disorders.
Beginning in 1984, initial trials identified several markers with abnormal expression levels in T21 pregnancies that could be captured via urine or maternal serum, including alpha-fetoprotein (AFP), PAPP-A (pregnancy associated plasma protein A), unconjugated estriol 3 (UE3), human chorionic gonadotropin (CG) and free β-hCG (CGB) (Brock et al., 1990; Brizot et al., 1995; David et al., 1996; Newby et al., 1996; Cuckle et al., 1999). Other markers that were investigated can be found in Table 1 (Brock et al., 1990; Newby et al., 2000; Christiansen et al., 1999). Teams also recognized the importance of distinguishing between two prenatal screening periods: first trimester (8–14 weeks) and second trimester (14–18 weeks). Many earlier studies focused on evaluating biomarkers in the second trimester as expression differences between T21 and normal pregnancies were more detectable.
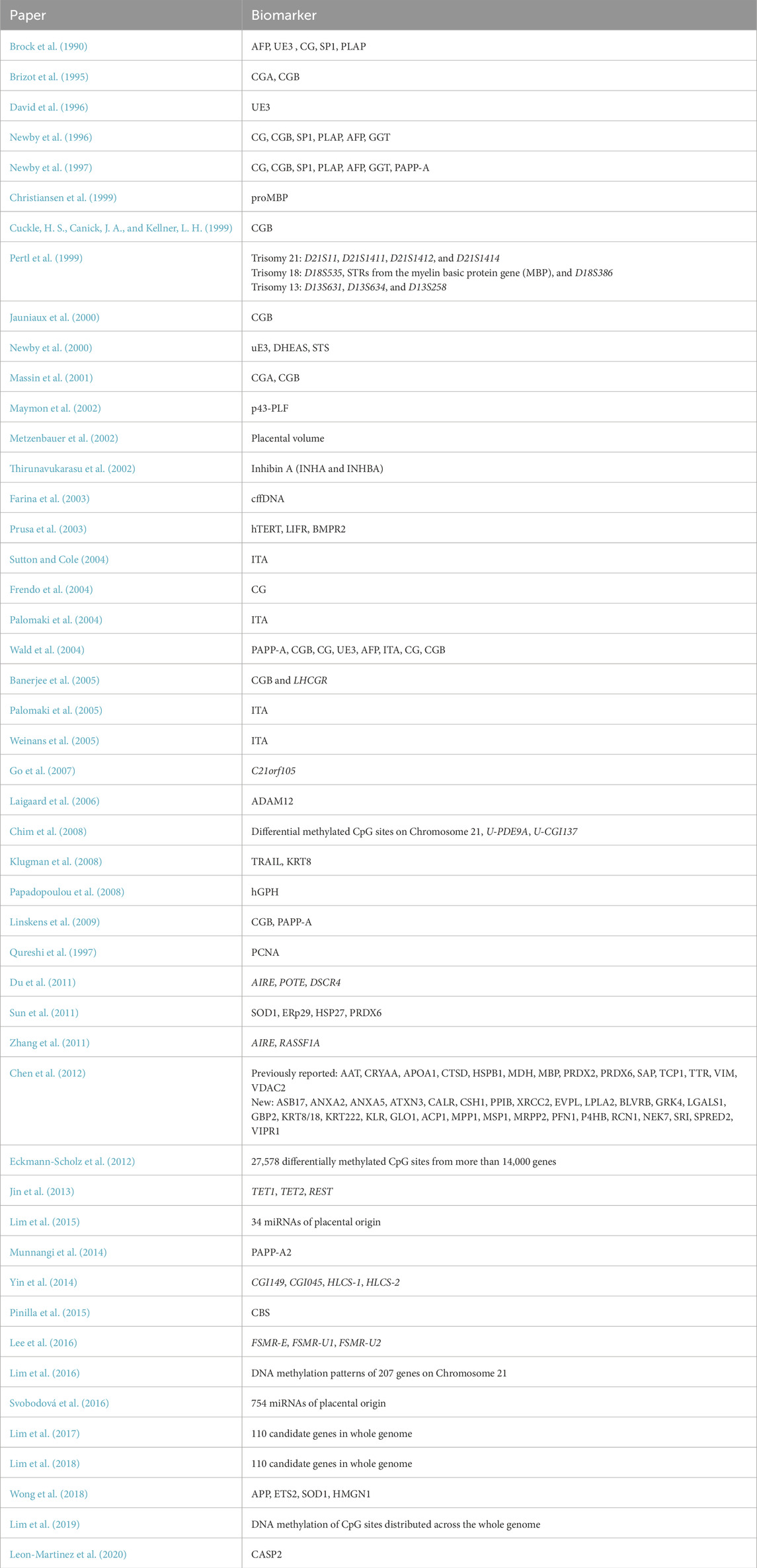
Table 1. Summary of biomarkers discussed for use in T21 screening (Mj et al., 2022; Pertl et al., 1999; Jauniaux et al., 2000; Metzenbauer et al., 2002; Brock et al., 1990; Brizot et al., 1995; David et al., 1996; Newby et al., 1996; Cuckle et al., 1999; Newby et al., 2000; Christiansen et al., 1999), (Palomaki et al., 2004; Wald et al., 2004; Weinans et al., 2005; Palomaki et al., 2005; Thirunavukarasu et al., 2002; Chim et al., 2008; Du et al., 2011; Zhang et al., 2011; Eckmann-Scholz et al., 2012; Jin et al., 2013; Yin et al., 2014; Lee et al., 2016; Lim et al., 2016; Lim et al., 2017; Lim et al., 2018), (Gordevičius et al., 2020; Lim et al., 2015; Svobodov et al., 2016; Newby et al., 1997; Massin et al., 2001; Maymon et al., 2002; Farina et al., 2003; Prusa et al., 2003; Frendo et al., 2004; Banerjee et al., 2005; Go et al., 2007; Laigaard et al., 2006; Klugman et al., 2008; Papadopoulou et al., 2008; Linskens et al., 2009; Qureshi et al., 1997; Sun et al., 2011; Chen et al., 2012; Munnangi et al., 2014; Pinilla et al., 2015; Wong et al., 2018).
Entering the new millennium, invasive trophoblast antigen (ITA) and inhibin A were introduced as a new potential biomarker for T21, reflecting the ongoing efforts at the time in improving the sensitivity and specificity of prenatal T21 screening (Sutton and Cole, 2004; Palomaki et al., 2004; Wald et al., 2004; Weinans et al., 2005; Palomaki et al., 2005; Thirunavukarasu et al., 2002). At the same time, there was an increasing focus on the optimization of screening protocols, with Wald et al. as seen in Table 1 comparing false positive rates for several combinations of biomarkers that have since helped to guide current screening procedures as will be described below (Wald et al., 2004).
Since then, investigations have shifted from protein-based methods towards epigenetic-based methods to improve noninvasive T21 prenatal screening (Chim et al., 2008; Du et al., 2011; Zhang et al., 2011; Eckmann-Scholz et al., 2012; Jin et al., 2013; Yin et al., 2014; Lee et al., 2016; Lim et al., 2016; Lim et al., 2017; Lim et al., 2018; Lim et al., 2019; Gordevičius et al., 2020). Differentially methylated DNA is used in the identification and isolation of cffDNA from maternal serum samples, with teams identifying several CpG sites which were specifically hyper- or hypomethylated in fetal samples, allowing for noninvasive prenatal screening using cffDNA. The upregulation of certain microRNAs (miRNAs) was also highlighted by several teams as potential screening tools for T21 (Lim et al., 2015; Svobodov et al., 2016).
At this time, prenatal T21 testing includes noninvasive screening and invasive diagnostic tests as illustrated in Figure 2 (MedlinePlus Medical Test, 2025). These are typically performed for expectant mothers with risk factors such as older maternal age (>35 years of age at time of pregnancy), family history of T21, having a previous child or pregnancy diagnosed with T21, and a fetal ultrasound with possible signs of T21. Looking to the future, it continues to be important to improve the cost-effectiveness of testing while reducing the rates of false positives. This demonstrates the process of developing minimally invasive and high specificity diagnostic tools to improve neurological outcomes in fetuses and neonates–a process that will continue to evolve for other neurodevelopmental outcomes described below.
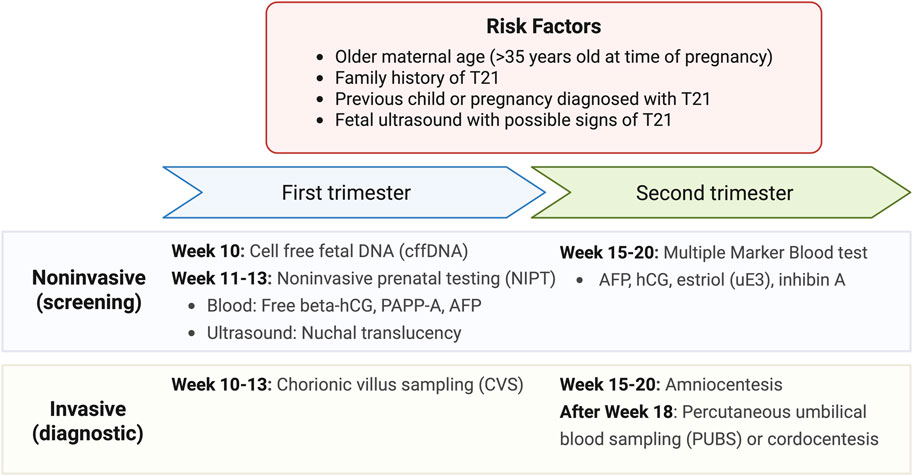
Figure 2. Current tests available for Down Syndrome screening and diagnosis. Results from noninvasive screening from first and second trimesters can also be integrated into the combined testing method which provides a more robust estimate of the risk of prenatal T21 (MedlinePlus Medical Test, 2025).
3.2 Current investigations
As discussed, the placenta also modulates numerous interactions across the maternal-fetal axis. Current studies on placental biomarkers highlight their diverse roles in inflammation, hypoxic signaling, timing of delivery, hormone and metabolic regulation, and various omics-related pathways as listed in Table 2. Beyond established use in T21 and select genetic disorders (e.g. T13, T18), placental biomarkers for other neurodevelopmental outcomes remain exploratory and have yet to enter routine clinical practice. Figure 3 highlights the promise of an integrated biomarker approach in this expanding area of research.
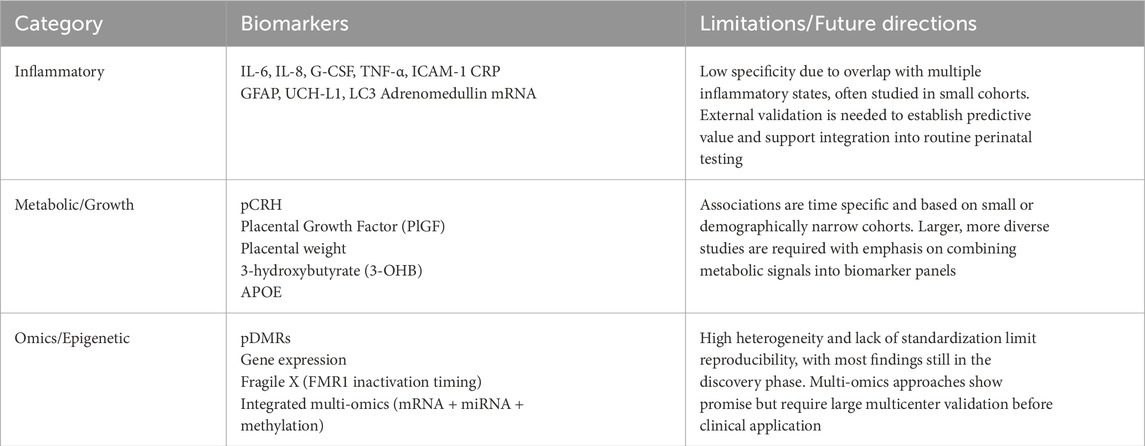
Table 2. Summary of exploratory placental biomarkers with current limitations and future directions.
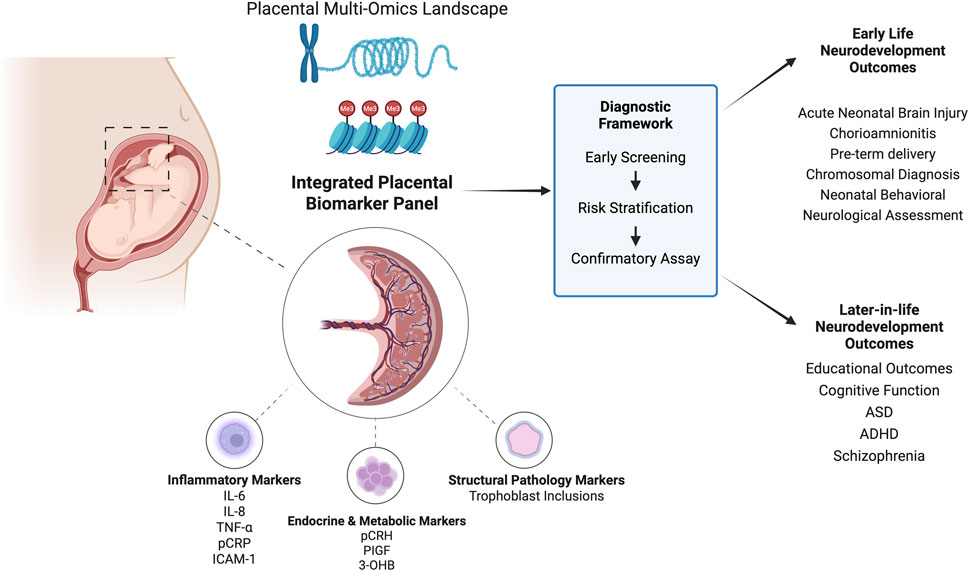
Figure 3. Landscape of placental biomarkers for early screening of neurodevelopmental outcomes. Review shows potential of an integrated placental biomarker panel to develop a diagnostic framework similar to Trisomy 21.
3.2.1 Inflammatory markers
Over the past few decades, studies have made significant strides from identifying inflammatory markers during pregnancy to linking distinct inflammatory pathways with specific NDDs. In 1992, Saito et al. identified elevations in cytokines (IL-6 and IL-8) in cytotrophoblasts and decidual stromal cells in cases of chorioamnionitis and preterm births, implicating intrauterine infection in neonatal outcome (Saito et al., 1993). These early observations prompted researchers to ask whether inflammatory markers implicated in preterm birth might also be involved in fetal brain injury, thereby linking placental inflammation to the pathogenesis of neonatal encephalopathy (NE). NE typically manifests with difficulty initiating breathing, low muscle tone, decreased consciousness, and seizures. It often occurs as a complication of perinatal hypoxia and/or pro-inflammatory cytokines, which impair gas and nutrient exchange and cause oxidative stress. It is associated with increased risk of death and long-term neurodevelopmental impairment, thus is a significant area of perinatal research (Shankaran et al., 1991; Kurinczuk et al., 2010).
Early explorations into genetic susceptibility identified HLA allele subtypes, namely, HLA-B, -DR, and -DQ, as a potential risk factor for NE (Cowan et al., 1994). Expansion of the placental landscape has identified glial fibrillary acidic protein (GFAP), an intermediate filament protein in astrocytes; ubiquitin carboxy-terminal hydrolase L1 (UCH-L1), a neuroendocrine specific enzyme; and increased LC3 expression in syncytiotrophoblasts, a marker of autophagy, as promising biomarkers for detecting the incidence and severity of NE. (Mir and Chalak, 2014; Douglas-Escobar et al., 2010; Avagliano et al., 2013).
Several studies have examined the timing and maternal context of inflammatory responses, including distinct oxidative stress signatures in fetal brain versus placental samples and adrenomedullin mRNA levels in the placenta to pinpoint the timing of hypoxic-ischemic encephalopathy associated death (Baldari et al., 2023). The expression of peptide adrenomedullin by fetal trophoblasts has been studied as a potential biomarker and therapeutic agent for preeclampsia primarily in animal models due to its role in the maternal-fetal perfusion response (Zhang et al., 2023; Li et al., 2013). Adrenomedullin mRNA in humans provides a window into timing that expands upon well-established and routine markers like pH and lactate levels to differentiate between acute and chronic intrauterine hypoxia; however, its potential as a prognostic factor needs to be explored (Trollmann et al., 2002; Gea et al., 2007).
Timing of inflammatory responses has brought about the two-hit hypothesis which, as shown in Figure 4, proposes that sequential antenatal and postnatal inflammatory insults (‘hits’) contribute to neurodevelopmental injury. Mapping risk profiles across the placenta began with the umbilical cord using new screening tools and imaging techniques. Studies reported that isolated placental perfusion insufficiency, detected antenatally by elevated pulsatility indices in the umbilical artery, ductus venosus, and inferior vena cava, predicted suboptimal neurodevelopmental outcomes at 1 year even when histological inflammation was absent (Kaukola et al., 2005).
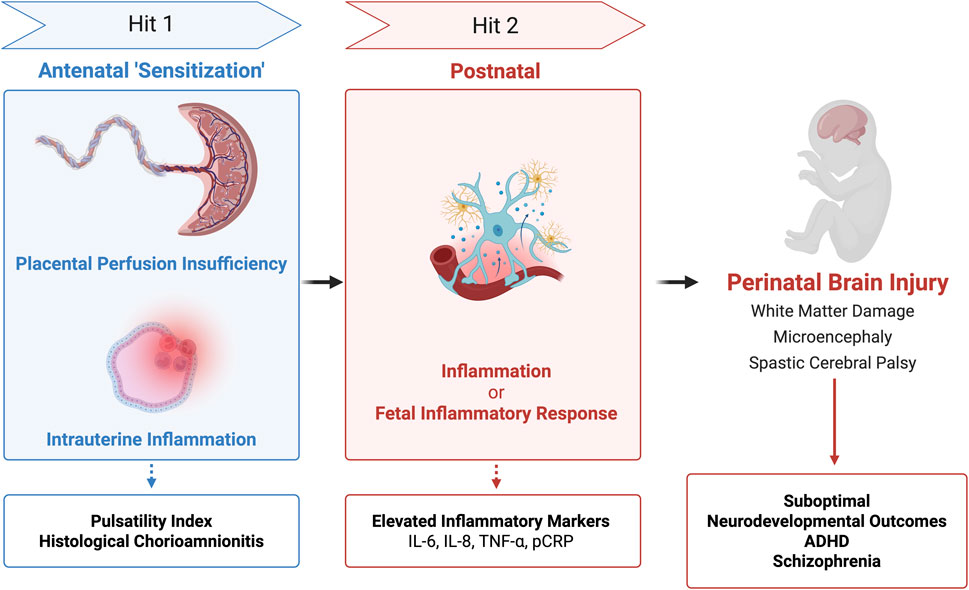
Figure 4. Two Hit Hypothesis of Perinatal Brain Injury. Integration of placental testing (pulsatility index, histology, and inflammatory markers) based on timing of injury (‘hits’) may have associations with brain injury and long-term neurodevelopmental outcomes.
Subsequent work broadened the model to intrauterine inflammation. Some cases of histological chorioamnionitis (HCA) progress to fetal inflammatory response (FIR) and at its extreme to fetal inflammatory response syndrome (FIRS). Conversely, non-infectious stressors like hypoxia-ischemia can trigger FIR/FIRS in the absence of HCA. These infectious or sterile pathways describe two related intrauterine ‘hits’ in preterm infants. Regardless of the initiating pathway, elevated cytokine levels (IL-6, IL-8, G-CSF) in the umbilical cord correlated with increased severity of brain injury (Lu et al., 2016). Some researchers speculated that the first ‘hit’ may be sensitizing the fetal brain to subsequent perinatal or postnatal stressors.
Yanni et al. explicitly applied the two-hit framework and demonstrated that antenatal inflammation (perfusion insufficiency or HCA) followed by postnatal inflammation (elevated inflammatory proteins) significantly increased the risk of adverse neurodevelopmental outcomes. Specifically, elevated placental C-reactive protein (pCRP) and ICAM-1 were associated with white matter damage; TNF-α and IL-8 with spastic cerebral palsy; and IL-6, TNF-α, or ICAM-1 with microcephaly. These findings suggest cytokines are not merely messengers of inflammation, but may drive or even directly mediate damage to neural tissue, making them potential candidates for perinatal screening or diagnostic biomarkers (Yanni et al., 2017). However, further research is needed to improve their specificity and diagnostic accuracy. Other investigations have linked pregnancy-related anxiety to attention-deficit/hyperactivity disorder (ADHD) symptoms at 36 months of age with sex-specific associations, particularly in boys, between pCRP mRNA expression, TNF-α and ADHD symptoms (Shao et al., 2020). Larger cohorts have expanded into neuropsychiatry, namely, schizophrenia, in which each 1 mg/L increase in maternal CRP was associated with a 28% higher risk of schizophrenia (Canetta et al., 2014). Collectively, these findings support that the placenta archives fetal exposure to infection, hypoxia, and oxidative stress. Integrating cytokines, acute phase reactants, placenta-derived peptides and autophagy markers into a biomarker panel could enable early risk stratification of infants at risk and shift perinatal clinical practice toward precision medicine.
3.2.2 Metabolic markers
Placental biomarker research has gone beyond acute injury to investigating placenta as a predictive organ for long-term neurodevelopmental outcomes. Endocrine and growth-related signals are emerging as especially informative. Elevations in placental corticotropin-releasing hormone (pCRH) have been associated with poor motor outcomes and ventriculomegaly with a study finding that the timing of pCRH exposure at 19 weeks and 31 weeks altered cortical development and spatial attention, respectively (Sandman et al., 2018; Leviton et al., 2016).
Studies found that fetal growth restriction (FGR) and placental growth factor (PlGF) in the second trimester were predictive of lower educational outcomes at 5–7 years and neurodivergent behaviors, respectively (Leon-Martinez et al., 2020; Tsompanidis et al., 2023). The study also found that distinct PlGF patterns were associated with male sex, potentially explaining the higher prevalence of neurodivergent behaviors observed in males.
Decreased placental weight has been connected to increased risk of FGR for several years. More recently, the opposite end has been explored with one study reporting a possible link between increased placental weight and risk of Autism Spectrum Disorder (ASD). However, placental weight is a nonspecific biomarker and the study relied on a small subset of single nucleotide polymorphisms; thus, placental weight may be a potential proxy for intrauterine conditions rather than a causal mechanism (Liu et al., 2021; Liu, 2024).
In parallel, emerging work expands the scope of placental biomarkers to metabolic signals, namely, 3-hydroxybutyrate (3-OHB). 3-OHB, a maternally produced ketone body, crosses the placenta and is a signal of metabolic stress. The included study reported that elevated umbilical cord 3-OHB levels were linked to atypical development on the Mullen Scales of Early Learning, but showed no association with ASD (Parenti et al., 2024). Further exploration into metabolomics as a diagnostic tool in addition to growth factors and placental weight may be a powerful combination for risk stratification of long term neurodevelopmental outcomes.
3.2.3 Vast omics of the placenta
This expanding view reflects a broader shift toward the omics of the placenta to create a more integrated picture of fetal programming as shown in Figure 3.
Analysis of ‘omics’ emphasizes the placenta as a ‘record-keeper’ of the fetal environment with research progressing from broad measures like differentially methylated regions in the placenta (pDMRs) and isolated gene expression to an integrated, multi-omics approach.
Earlier studies focused on imprinted gene expression with outcomes showing distinct placental expression profiles. In Fragile X syndrome, the Fragile X Messenger Ribonucleoprotein 1 (FMR1) gene in chorionic villi samples showed absence of FMR1 in cytotrophoblasts and connected gestational age to the timing of inactivation of the FMR1 allele with DNA methylation found at specific CpG islands (Willemsen et al., 2002; Luo et al., 1993). These studies demonstrated a new method for prenatal diagnosis of Fragile X syndrome by leveraging placental expression and epigenetic timing; however, it has yet to be clinically implemented.
Building on this foundation, research has increasingly focused on the prenatal origins of ASD, linking it to maternal immune activation, gestational diabetes, and metabolic conditions (Liu, 2024; Li et al., 2016; Meltzer and Van de Water, 2017; Gardner et al., 2015; Xiang et al., 2015). Structural abnormalities in the placenta, such as trophoblast inclusions, have been associated with altered neurodevelopment, though causative relationships in placental structure continue to be explored (Anderson et al., 2007). Epigenetic studies further support placental involvement, with both hyper- and hypomethylated CpG regions linked to ASD, attention deficits, and cognitive delays. Hypermethylated CpG regions associated with reduced expression of GPR135, ITGBL1, FHIT, IRS2, DLL1, and LRRFIP1 suggest disruptions in synapse quantity, neuritogenesis, abnormal morphology, immune regulation, and GABAergic neuronal signaling, while hypomethylated regions affecting CYP2E1 and 22q13.33 imply dysregulation in oxidative stress and synaptic development/function (Bahado-Singh et al., 2021; Paquette et al., 2016; Tilley et al., 2018; Zhu et al., 2019; Zhu et al., 2022; Schroeder et al., 2016; Freedman et al., 2023).
As the understanding of the perinatal environment develops, research has moved beyond broad methylation profiles toward gene-specific signals. Several placental proteins and epigenetic marks have emerged as early indicators of later neurodevelopment. One team found Apolipoprotein E (APOE) to be upregulated in the placenta of preterm infants who experienced developmental delays at 6 months (Zhu et al., 2023). APOE, a key mediator of lipid transport and synaptic plasticity, complements the metabolomic finding of 3-OHB by underscoring the notion that placental lipid handling shapes early neurodevelopment and offers promises as a screening biomarker (Lane-Donovan and Herz, 2017).
Single-cohort reports show that methylation of neuronal growth regulator 1 (NEGR1) and serotonin 2A receptor (HTR2A) correlated with adverse developmental outcomes, but many of these findings await confirmation in subsequent studies (Breton et al., 2020; Paquette et al., 2013). Integrating multiple omic layers can markedly enhance biomarker performance. Santos et al. found integration of methylation, mRNA, miRNA data improved accuracy of predicting ASD compared to single platform model (Santos et al., 2020).
4 Discussion
Neuroplacentology is a rapidly growing field and stands to provide incredible improvements in risk stratification for pregnant women and neonates. For instance, a team from the University of Texas Southwestern’s Parkland Hospital has implemented a targeted hospital-based protocol for triaging placentas from high-risk deliveries for histological evaluations in order to inform and facilitate timely treatment intervention strategies (Mir et al., 2021). This demonstrates the promising future of neuroplacentology and its possible incorporation into hospital-based practices in order to better address adverse neurodevelopmental outcomes that are amenable to early intervention, such as neonatal encephalopathy and chorioamnionitis.
As screening methods within neuroplacentology continue to improve, the psychosocial welfare of mothers must continue to be a main consideration. In 2017, the Netherlands launched a nationwide study known as the TRIDENT-2 study which offered NIPT to all pregnant women (van der Meij et al., 2019). The study focused on screening for T13, T18, T21, with additional findings outside of these three conditions being provided only upon request of the mother. Six years later, results on the psychological impacts of the testing’s “additional findings” were discussed by Bakkeren et al. (2024). Over 92% of women stated that receiving the additional finding was an unexpected shock and 85% reported that it caused significant amounts of worry. The study also looked at measures of anxiety and distress, highlighting that women who gave birth to an affected child were most impacted.
Currently, the American College of Obstetricians and Gynecologists (ACOG) recommends that all pregnant women be offered the option of screening and diagnostic testing for Down syndrome. As NIPT becomes more accessible to mothers across the globe, healthcare providers must remain vigilant in supporting women to understand the implications of prenatal testing (Screening for Fetal Chromosomal Abnormalities, 2020). This includes the potential need for and risks of follow-up invasive diagnostic tests. Counseling plays an important role in managing expectations and concerns regarding testing results, and should be strongly recommended to mentally prepare women for the possible outcomes from these screenings.
Additionally, it would be remiss not to mention that the literature examined in this review comes predominantly from high-income countries. As such, prediction models built upon data generated by these studies may not be validated in low- to middle-income countries. Antwi et al. share a possible method of addressing this challenge, such as updating the models to align with clinical characteristics observed within that population (Antwi et al., 2020). It is also important to recognize that serum-based biomarker screenings may not be as readily available in these countries as well, which impacts the generalizability of results and outcomes regarding the clinical implementation of placental biomarker-based screenings.
Author contributions
PP: Conceptualization, Writing – original draft, Writing – review and editing, Data curation. JK: Conceptualization, Writing – original draft, Writing – review and editing. IU: Resources, Supervision, Writing – review and editing. JG: Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Carle Illinois College of Medicine and Carle Foundation Hospital, and Feinberg School of Medicine.
Acknowledgments
All schematics were created with https://BioRender.com
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2025.1663960/full#supplementary-material
References
Anderson, G. M., Jacobs-Stannard, A., Chawarska, K., Volkmar, F. R., and Kliman, H. J. (2007). Placental trophoblast inclusions in autism spectrum disorder. Biol. Psychiatry 61 (4), 487–491. doi:10.1016/j.biopsych.2006.03.068
Antwi, E., Amoakoh-Coleman, M., Vieira, D. L., Madhavaram, S., Koram, K. A., Grobbee, D. E., et al. (2020). Systematic review of prediction models for gestational hypertension and preeclampsia. PLoS ONE 15 (4), e0230955. doi:10.1371/journal.pone.0230955
Avagliano, L., Danti, L., Doi, P., Felis, S., Guala, M., Locatelli, A., et al. (2013). Autophagy in placentas from acidotic newborns: an immunohistochemical study of LC3 expression. Placenta 34 (11), 1091–1094. doi:10.1016/j.placenta.2013.09.004
Bahado-Singh, R. O., Vishweswaraiah, S., Aydas, B., and Radhakrishna, U. (2021). “Placental DNA methylation changes and the early prediction of autism in full-term newborns,” 16. doi:10.1371/journal.pone.0253340
Bakkeren, I. M., Henneman, L., van Vliet-Lachotzki, E. H., Martin, L., Gitsels-van der Wal, J. T., Polak, M. G., et al. (2024). Psychological impact of additional findings detected by genome-wide Non-Invasive Prenatal Testing (NIPT): TRIDENT-2 study. Eur. J. Hum. Genet. 32 (3), 302–308. doi:10.1038/s41431-023-01504-8
Baldari, B., De Simone, S., Cipolloni, L., Frisoni, P., Alfieri, L., D'Errico, S., et al. (2023). Oxidative stress markers in human brain and placenta may reveal the timing of hypoxic-ischemic injury: evidence from an immunohistochemical study. Int. J. Mol. Sci. 24 (15), 12221. doi:10.3390/ijms241512221
Banerjee, S., Smallwood, A., Chambers, A. E., Papageorghiou, A., Loosfelt, H., Spencer, K., et al. (2005). A link between high serum levels of human chorionic gonadotrophin and chorionic expression of its mature functional receptor (LHCGR) in Down’s syndrome pregnancies. Reprod. Biol. Endocrinol. 3 (1), 25. doi:10.1186/1477-7827-3-25
Breton, E., Gagné-Ouellet, V., Thibeault, K., Guérin, R., Van Lieshout, R., Perron, P., et al. (2020). Placental NEGR1 DNA methylation is associated with BMI and neurodevelopment in preschool-age children. Epigenetics 15 (3), 323–335. doi:10.1080/15592294.2019.1666653
Brizot, M. L., Jauniaux, E., Mckie, A. T., Farzaneh, F., and Nicolaides, K. H. (1995). Molecular interactions during pregnancy: placental expression of and subunits of human chorionic gonadotrophin in early pregnancies with Down’s syndrome. Hum. Reprod. 10 (9), 2506–2509. doi:10.1093/oxfordjournals.humrep.a136331
Brock, D. J. H., Barron, L., Holloway, S., Liston, W. A., Hillier, S. G., and Seppala, M. (1990). First-trimester maternal serum biochemical indicators in Down syndrome. Prenat. Diagn 10 (4), 245–251. doi:10.1002/pd.1970100406
Burton, G. J., and Fowden, A. L. (2015). The placenta: a multifaceted, transient organ. Philos. Trans. R. Soc. B Biol. Sci. 370 (1663), 20140066. doi:10.1098/rstb.2014.0066
Canetta, S., Sourander, A., Surcel, H. M., Hinkka-Yli-Salomäki, S., Leiviskä, J., Kellendonk, C., et al. (2014). Elevated maternal C-reactive protein and increased risk of schizophrenia in a national birth cohort. Am. J. Psychiatry 171 (9), 960–968. doi:10.1176/appi.ajp.2014.13121579
Chen, C. P., Chen, Y. H., Chern, S. R., Chang, S. J., Tsai, T. L., Li, S. H., et al. (2012). Placenta proteome analysis from Down syndrome pregnancies for biomarker discovery. Mol. Biosyst. 8 (9), 2360–2372. doi:10.1039/c2mb25081k
Chim, S. S. C., Jin, S., Lee, T. Y. H., Lun, F. M. F., Lee, W. S., Chan, L. Y. S., et al. (2008). Systematic search for placental DNA-methylation markers on chromosome 21: toward a maternal plasma-based epigenetic test for fetal trisomy 21. Clin. Chem. 54 (3), 500–511. doi:10.1373/clinchem.2007.098731
Christiansen, M., Oxvig, C., Wagner, J. M., Qin, Q. P., Nguyen, T. H., Overgaard, M. T., et al. (1999). The proform of eosinophil major basic protein: a new maternal serum marker for Down syndrome. Prenat. Diagn 19 (10), 905–910. doi:10.1002/(sici)1097-0223(199910)19:10<905::aid-pd658>3.0.co;2-q
Cowan, L. D., Hudson, L., Bobele, G., Chancellor, I., and Baker, J. (1994). Maternal-fetal HLA sharing and risk of newborn encephalopathy and Seizures: a pilot study. J. Child. Neurol. 9 (2), 173–177. doi:10.1177/088307389400900214
Cuckle, H. S., Canick, J. A., and Kellner, L. H. (1999). Collaborative study of maternal urine β -core human chorionic gonadotrophin screening for Down syndrome. Prenat. Diagn 19 (10), 911–917. doi:10.1002/(sici)1097-0223(199910)19:10<911::aid-pd666>3.0.co;2-w
David, M., Merksamer, R., Israel †, N., and Dar, H. (1996). Unconjugated estriol as maternal serum marker for the detection of down syndrome pregnancies. Fetal Diagn Ther. 11 (2), 99–105. doi:10.1159/000264287
Douglas-Escobar, M., Yang, C., Bennett, J., Shuster, J., Theriaque, D., Leibovici, A., et al. (2010). A pilot study of novel biomarkers in neonates with hypoxic-ischemic encephalopathy. Pediatr. Res. 68 (6), 531–536. doi:10.1203/PDR.0b013e3181f85a03
Du, Y., Zhang, J., Wang, H., Yan, X., Yang, Y., Yang, L., et al. (2011). Hypomethylated DSCR4 is a placenta-derived epigenetic marker for trisomy 21. Prenat. Diagn 31 (2), 207–214. doi:10.1002/pd.2684
Eckmann-Scholz, C., Bens, S., Kolarova, J., Schneppenheim, S., Caliebe, A., Heidemann, S., et al. (2012). DNA-methylation profiling of fetal tissues reveals marked epigenetic differences between chorionic and amniotic samples. PLoS ONE 7 (6), e39014. doi:10.1371/journal.pone.0039014
Espinoza, M. L., Brundler, M. A., Hasan, S. U., Mohammad, K., Momin, S., Al Shaikh, B., et al. (2022). Placental pathology as a marker of brain injury in infants with hypoxic ischemic encephalopathy. Early Hum. Dev. 174, 105683. doi:10.1016/j.earlhumdev.2022.105683
Farina, A., LeShane, E. S., Lambert-Messerlian, G. M., Canick, J. A., Lee, T., Neveux, L. M., et al. (2003). Evaluation of cell-free fetal DNA as a second-trimester maternal serum marker of down syndrome pregnancy. Clin. Chem. 49 (2), 239–242. doi:10.1373/49.2.239
FDA-NIH Biomarker Working Group (2016). BEST (biomarkers, EndpointS, and other tools) resource. Food Drug Adm. (US). Available online at: http://www.ncbi.nlm.nih.gov/books/NBK326791/(Accessed June 10, 2025).
Freedman, A. N., Clark, J., Eaves, L. A., Roell, K., Oran, A., Koval, L., et al. (2023). A multi-omic approach identifies an autism spectrum disorder (ASD) regulatory complex of functional epimutations in placentas from children born preterm. Autism Res. 16 (5), 918–934. doi:10.1002/aur.2915
Frendo, J. L., Guibourdenche, J., Pidoux, G., Vidaud, M., Luton, D., Giovangrandi, Y., et al. (2004). Trophoblast production of a Weakly Bioactive human chorionic gonadotropin in trisomy 21-affected pregnancy. J. Clin. Endocrinol. Metab. 89 (2), 727–732. doi:10.1210/jc.2003-030668
Gall, A. R., Amoah, S. K., Kitase, Y., and Jantzie, L. L. (2022). Placental mediated mechanisms of perinatal brain injury: Evolving inflammation and exosomes. Exp. Neurol. 347, 113914. doi:10.1016/j.expneurol.2021.113914
Ganguly, E., Hula, N., Spaans, F., Cooke, C. L. M., and Davidge, S. T. (2020). Placenta-targeted treatment strategies: an opportunity to impact fetal development and improve offspring health later in life. Pharmacol. Res. 157, 104836. doi:10.1016/j.phrs.2020.104836
Gardner, R. M., Lee, B. K., Magnusson, C., Rai, D., Frisell, T., Karlsson, H., et al. (2015). Maternal body mass index during early pregnancy, gestational weight gain, and risk of autism spectrum disorders: results from a Swedish total population and discordant sibling study. Int. J. Epidemiol. 44 (3), 870–883. doi:10.1093/ije/dyv081
Gea, Y., Araujo, O., and Silva, L. V. R. (2007). Clinical value of lactate measurement and nucleated red blood cell counts in the placental segment of the umbilical vein of premature newborns for diagnosis of hypoxia-ischemia. J. Pediatr. Rio J. 0 (0), 186–190. doi:10.2223/JPED.1597
Go, A. T. J. I., Visser, A., Mulders, M. A. M., Twisk, J. W. R., Blankenstein, M. A., van Vugt, J. M. G., et al. (2007). C21ORF105, A chromosome 21-encoded mRNA, is not a discriminative marker gene for prediction of Down syndrome in maternal plasma. Prenat. Diagn 27 (2), 146–149. doi:10.1002/pd.1637
Gordevičius, J., Narmontė, M., Gibas, P., Kvederavičiūtė, K., Tomkutė, V., Paluoja, P., et al. (2020). Identification of fetal unmodified and 5-hydroxymethylated CG sites in maternal cell-free DNA for non-invasive prenatal testing. Clin. Epigenetics 12 (1), 153. doi:10.1186/s13148-020-00938-x
Jauniaux, E., Bao, S., Eblen, A., Li, X., Lei, Z. M., Meuris, S., et al. (2000). HCG concentration and receptor gene expression in placental tissue from trisomy 18 and 21. Mol. Hum. Reprod. 6 (1), 5–10. doi:10.1093/molehr/6.1.5
Jin, S., Lee, Y. K., Lim, Y. C., Zheng, Z., Lin, X. M., Ng, D. P. Y., et al. (2013). “Global DNA hypermethylation in down syndrome placenta,” 9. doi:10.1371/journal.pgen.1003515
Kratimenos, P., and Penn, A. A. (2019). Placental programming of neuropsychiatric disease. Pediatr. Res. 86 (2), 157–164. doi:10.1038/s41390-019-0405-9
Kaukola, T., Räsänen, J., Herva, R., Patel, D. D., and Hallman, M. (2005). Suboptimal neurodevelopment in very preterm infants is related to fetal cardiovascular compromise in placental insufficiency. Am. J. Obstet. Gynecol. 193 (2), 414–420. doi:10.1016/j.ajog.2004.12.005
Klugman, S. D., Gross, S. J., Khabele, D., Livne, K., Gross, B., et al. (2008). Expression of keratin 8 and TNF-related apoptosis-I inducing ligand (TRAIL) in down syndrome placentas. Placenta 29 (4), 382–384. doi:10.1016/j.placenta.2008.01.013
Kurinczuk, J. J., White-Koning, M., and Badawi, N. (2010). Epidemiology of neonatal encephalopathy and hypoxic–ischaemic encephalopathy. Early Hum. Dev. 86 (6), 329–338. doi:10.1016/j.earlhumdev.2010.05.010
Laigaard, J., Spencer, K., Christiansen, M., Cowans, N. J., Larsen, S. O., Pedersen, B. N., et al. (2006). ADAM 12 as a first-trimester maternal serum marker in screening for Down syndrome. Prenat. Diagn 26 (10), 973–979. doi:10.1002/pd.1540
Lane-Donovan, C., and Herz, J. (2017). ApoE, ApoE receptors, and the synapse in alzheimer’s disease. Trends Endocrinol. Metab. Tem. 28 (4), 273–284. doi:10.1016/j.tem.2016.12.001
Lee, D. E., Lim, J. H., Kim, M. H., Park, S. Y., and Ryu, H. M. (2016). Novel epigenetic markers on chromosome 21 for noninvasive prenatal testing of fetal trisomy 21. J. Mol. Diagn 18 (3), 378–387. doi:10.1016/j.jmoldx.2015.12.002
Leon-Martinez, D., Robinson, J. F., Zdravkovic, T., Genbacev, O., Gormley, M., Mcmaster, M., et al. (2020). Trisomy 21 is associated with caspase-2 upregulation in cytotrophoblasts at the maternal-fetal interface. Reprod. Sci. 27 (1), 100–109. doi:10.1007/s43032-019-00002-x
Leviton, A., Allred, E. N., Kuban, K. C. K., O'Shea, T. M., Paneth, N., Majzoub, J., et al. (2016). Brain disorders associated with corticotropin-releasing hormone expression in the placenta among children born before the 28th week of gestation. Acta Paediatr. 105 (1), e7–e11. doi:10.1111/apa.13174
Li, M., Schwerbrock, N. M. J., Lenhart, P. M., Fritz-Six, K. L., Kadmiel, M., Christine, K. S., et al. (2013). Fetal-derived adrenomedullin mediates the innate immune milieu of the placenta. J. Clin. Invest 123 (6), 2408–2420. doi:10.1172/JCI67039
Li, M., Fallin, M. D., Riley, A., Landa, R., Walker, S. O., Silverstein, M., et al. (2016). The association of maternal obesity and diabetes with autism and other developmental disabilities. Pediatrics 137 (2), e20152206. doi:10.1542/peds.2015-2206
Lim, J. H., Kim, D. J., Lee, D. E., Han, J. Y., Chung, J. H., Ahn, H. K., et al. (2015). Genome-wide microRNA expression profiling in placentas of fetuses with Down syndrome. Placenta 36 (3), 322–328. doi:10.1016/j.placenta.2014.12.020
Lim, J. H., Kim, S. Y., Han, J. Y., Kim, M. Y., Park, S. Y., and Ryu, H. M. (2016). Comprehensive investigation of DNA methylation and gene expression in trisomy 21 placenta. Placenta 42, 17–24. doi:10.1016/j.placenta.2016.03.012
Lim, J. H., Han, Y. J., Kim, H. J., Kwak, D. W., Park, S. Y., Chun, S. H., et al. (2017). Genome-wide gene expression analysis in the placenta from fetus with trisomy 21. BMC Genomics 18 (1), 720. doi:10.1186/s12864-017-3993-y
Lim, J. H., Han, Y. J., Kim, H. J., Kim, M. Y., Park, S. Y., Cho, Y. H., et al. (2018). Integrative analyses of genes and microRNA expressions in human trisomy 21 placentas. BMC Med. Genomics 11 (1), 46. doi:10.1186/s12920-018-0361-y
Lim, J. H., Kang, Y. J., Lee, B. Y., Han, Y. J., Chung, J. H., Kim, M. Y., et al. (2019). Epigenome-wide base-resolution profiling of DNA methylation in chorionic villi of fetuses with Down syndrome by methyl-capture sequencing. Clin. Epigenetics 11 (1), 180. doi:10.1186/s13148-019-0756-4
Linskens, I. H., Spreeuwenberg, M. D., Blankenstein, M. A., and Van Vugt, J. M. G. (2009). Early first-trimester free β-hCG and PAPP-A serum distributions in monochorionic and dichorionic twins. Prenat. Diagn 29 (1), 74–78. doi:10.1002/pd.2184
Liu, Z. (2024). Genetic causal relationship between placental weight and autism spectrum disorder: a two-sample Mendelian randomization study. J. Psychosom. Res. 184, 111857. doi:10.1016/j.jpsychores.2024.111857
Liu, H. J., Liu, P. C., Hua, J., Zhao, Y., and Cao, J. (2021). Placental weight and size in relation to fetal growth restriction: a case-control study. J. Matern-Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obstet. 34 (9), 1356–1360. doi:10.1080/14767058.2019.1636371
Lu, H. -Y., Zhang, Q., xia, W. Q., and Lu, J. -Y. (2016). Contribution of histologic chorioamnionitis and fetal inflammatory response syndrome to increased risk of brain injury in infants with preterm premature Rupture of Membranes. Pediatr. Neurol. 61, 94–98.e1. doi:10.1016/j.pediatrneurol.2016.05.001
Luo, S., Robinson, J. C., Reiss, A. L., and Migeon, B. R. (1993). DNA methylation of the fragile X locus in somatic and germ cells during fetal development: relevance to the Fragile X syndrome and X inactivation. Somat. Cell Mol. Genet. 19 (4), 393–404. doi:10.1007/BF01232750
Massin, N., Frendo, J. L., Guibourdenche, J., Luton, D., Giovangrandi, Y., Muller, F., et al. (2001). Defect of syncytiotrophoblast formation and human chorionic gonadotropin expression in Down’s syndrome. Placenta 22, S93–S97. doi:10.1053/plac.2001.0658
Maymon, R., Jauniaux, E., and Moroz, C. (2002). Enhanced expression of the immunoregulator, p43-placental isoferritin, in Down’s syndrome placentae and fetal kidneys. Mol. Hum. Reprod. 8 (12), 1125–1128. doi:10.1093/molehr/8.12.1125
MedlinePlus Medical Test (2025). Down syndrome tests: MedlinePlus medical test. Available online at: https://medlineplus.gov/lab-tests/down-syndrome-tests/(Accessed June 25, 2025).
Meltzer, A., and Van de Water, J. (2017). The role of the immune system in autism spectrum disorder. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 42 (1), 284–298. doi:10.1038/npp.2016.158
Metzenbauer, M., Hafner, E., Schuchter, K., and Philipp, K. (2002). First-trimester placental volume as a marker for chromosomal anomalies: preliminary results from an unselected population. Ultrasound Obstet. Gynecol. 19 (3), 240–242. doi:10.1046/j.1469-0705.2002.00655.x
Mir, I. N., and Chalak, L. F. (2014). Serum biomarkers to evaluate the integrity of the neurovascular unit. Early Hum. Dev. 90 (10), 707–711. doi:10.1016/j.earlhumdev.2014.06.010
Mir, I. N., Leon, R., and Chalak, L. F. (2021). Placental origins of neonatal diseases: toward a precision medicine approach. Pediatr. Res. 89 (2), 377–383. doi:10.1038/s41390-020-01293-6
Mj, B., T, T., Sl, S., Christensen, C., and Grout, R. W. (2022). Health supervision for children and adolescents with down syndrome. Pediatrics 149 (5), e2022057010. doi:10.1542/peds.2022-057010
Munnangi, S., Gross, S. J., Madankumar, R., Salcedo, G., and Reznik, S. E. (2014). Pregnancy associated plasma protein-A2: a novel biomarker for down syndrome. Placenta 35 (11), 900–906. doi:10.1016/j.placenta.2014.08.001
Nelissen, E. C. M., van Montfoort, A. P. A., Dumoulin, J. C. M., and Evers, J. L. H. (2011). Epigenetics and the placenta. Hum. Reprod. Update 17 (3), 397–417. doi:10.1093/humupd/dmq052
Newby, D., Aitken, D. A., Crossley, J. A., Howatson, A. G., and Connor, J. M. (1996). The pathophysiology of Down’s syndrome pregnancies. Early Hum. Dev. 47, S67–S68. doi:10.1016/S0378-3782(96)01824-5
Newby, D., Aitken, D. A., Crossley, J. A., Howatson, A. G., Macri, J. N., and Connor, J. M. (1997). Biochemical markers of trisomy 21 and the pathophysiology of Down’s syndrome pregnancies. Prenat. Diagn 17 (10), 941–951. doi:10.1002/(sici)1097-0223(199710)17:10<941::aid-pd183>3.0.co;2-g
Newby, D., Aitken, D. A., Howatson, A. G., and Connor, J. M. (2000). Placental synthesis of oestriol in down’s syndrome pregnancies. Placenta 21 (2-3), 263–267. doi:10.1053/plac.1999.0469
Page, J. M., Christiansen-Lindquist, L., Thorsten, V., Parker, C. B., Reddy, U. M., Dudley, D. J., et al. (2017). Diagnostic tests for evaluation of stillbirth: results from the stillbirth collaborative research network. Obstet. Gynecol. 129 (4), 699–706. doi:10.1097/AOG.0000000000001937
Palomaki, G. E., Knight, G. J., Roberson, M. M., Cunningham, G. C., Lee, J. E., Strom, C. M., et al. (2004). Invasive trophoblast antigen (hyperglycosylated human chorionic gonadotropin) in second-trimester maternal urine as a marker for down syndrome: preliminary results of an observational study on fresh samples. Clin. Chem. 50 (1), 182–189. doi:10.1373/clinchem.2003.023986
Palomaki, G. E., Knight, G. J., Neveux, L. M., Pandian, R., and Haddow, J. E. (2005). Maternal serum invasive trophoblast antigen and first-trimester down syndrome screening. Clin. Chem. 51 (8), 1499–1504. doi:10.1373/clinchem.2005.050567
Papadopoulou, E., Sifakis, S., Giahnakis, E., Fragouli, Y., Karkavitsas, N., Koumantakis, E., et al. (2008). Human placental growth hormone is increased in maternal serum in pregnancies affected by down syndrome. Fetal Diagn Ther. 23 (3), 211–216. doi:10.1159/000116743
Paquette, A. G., Lesseur, C., Armstrong, D. A., Koestler, D. C., Appleton, A. A., Lester, B. M., et al. (2013). Placental HTR2A methylation is associated with infant neurobehavioral outcomes. Epigenetics 8 (8), 796–801. doi:10.4161/epi.25358
Paquette, A. G., Houseman, E. A., Green, B. B., Lesseur, C., Armstrong, D. A., Lester, B., et al. (2016). Regions of variable DNA methylation in human placenta associated with newborn neurobehavior. Epigenetics 11 (8), 603–613. doi:10.1080/15592294.2016.1195534
Parenti, M., Schmidt, R. J., Tancredi, D. J., Hertz-Picciotto, I., Walker, C. K., and Slupsky, C. M. (2024). Neurodevelopment and metabolism in the maternal-placental-fetal unit. JAMA Netw. Open 7 (5), e2413399. doi:10.1001/jamanetworkopen.2024.13399
Pertl, B., Kopp, S., Kroisel, P., Tului, L., Brambati, B., and Adinolfi, M. (1999). Rapid detection of chromosome aneuploidies by quantitative fluorescence PCR: first application on 247 chorionic villus samples. J. Med. Genet. 36 (4), 300–303. Available online at https://pubmed.ncbi.nlm.nih.gov/10227397/.
Pinilla, J. M., Ayala-Ramírez, P., García-Robles, R., and Bermúdez, M. (2015). Expression of cystathionine beta-synthase and histopathological observations in placentas of patients with Down syndrome. J. Neonatal-Perinat Med. 8 (2), 77–84. doi:10.3233/NPM-15814092
Prusa, A. R., Marton, E., Rosner, M., Freilinger, A., Bernaschek, G., and Hengstschläger, M. (2003). Stem cell marker expression in human trisomy 21 amniotic fluid cells and trophoblasts. In: J. Neural Transm. Suppl. 67 235–242. doi:10.1007/978-3-7091-6721-2_21
Qureshi, F., Jacques, S. M., Johnson, M. P., Hume, R. F., Kramer, R. L., Yaron, Y., et al. (1997). Trisomy 21 placentas: histopathological and immunohistochemical findings using proliferating cell nuclear antigen. Fetal Diagn Ther. 12 (4), 210–215. doi:10.1159/000264470
Saito, S., Kasahara, T., Kato, Y., Ishihara, Y., and Ichijo, M. (1993). Elevation of amniotic fluid interleukin 6 (IL-6), IL-8 and granulocyte colony stimulating factor (G-CSF) in term and preterm parturition. Cytokine 5 (1), 81–88. doi:10.1016/1043-4666(93)90027-3
Sandman, C. A., Curran, M. M., Davis, E. P., Glynn, L. M., Head, K., and Baram, T. Z. (2018). Cortical thinning and neuropsychiatric outcomes in children exposed to prenatal adversity: a role for placental crh? Am. J. Psychiatry 175 (5), 471–479. doi:10.1176/appi.ajp.2017.16121433
Santos, Jr H. P., Bhattacharya, A., Joseph, R. M., Smeester, L., Kuban, K. C. K., Marsit, C. J., et al. (2020). Evidence for the placenta-brain axis: multi-omic kernel aggregation predicts intellectual and social impairment in children born extremely preterm. Mol. Autism 11 (1), 97. doi:10.1186/s13229-020-00402-w
Schroeder, D. I., Schmidt, R. J., Crary-Dooley, F. K., Walker, C. K., Ozonoff, S., Tancredi, D. J., et al. (2016). Placental methylome analysis from a prospective autism study. Mol. Autism 7 (1), 51. doi:10.1186/s13229-016-0114-8
Screening for Fetal Chromosomal Abnormalities (2020). Screening for fetal chromosomal abnormalities: ACOG practice bulletin summary, number 226. Obstet. Gynecol. 136 (4), 859–867. doi:10.1097/AOG.0000000000004107
Shankaran, S., Woldt, E., Koepke, T., Bedard, M. P., and Nandyal, R. (1991). Acute neonatal morbidity and long-term central nervous system sequelae of perinatal asphyxia in term infants. Early Hum. Dev. 25 (2), 135–148. doi:10.1016/0378-3782(91)90191-5
Shao, S., Wang, J., Huang, K., and Liu, H. (2020). Prenatal pregnancy-related anxiety predicts boys’ ADHD symptoms via placental C-reactive protein. Psychoneuroendocrinology 120, 104797. doi:10.1016/j.psyneuen.2020.104797
Sun, C. juan, yu, Y. L., Wang, W., Yu, S., Wang, X., and Zhang, W. yuan (2011). Proteomic analysis of the alteration of protein expression in the placenta of Down syndrome. Chin. Med. J. Engl. 124 (22), 3738–3745. doi:10.3760/cma.j.issn.0366-6999.2011.22.025
Sutton, J. M., and Cole, L. A. (2004). Sialic acid–deficient invasive trophoblast antigen (sd-ITA): a new urinary variant for gestational Down syndrome screening. Prenat. Diagn 24 (3), 194–197. doi:10.1002/pd.836
Svobodová, I., Korabečná, M., Calda, P., Břešťák, M., Pazourková, E., Pospíšilová, Š., et al. (2016). Differentially expressed miRNAs in trisomy 21 placentas: miRNAs in Down syndrome placenta. Prenat. Diagn 36 (8), 775–784. doi:10.1002/pd.4861
Thirunavukarasu, P. P., Lambert-Messerlian, G., Robertson, D. M., Dawson, G., Canick, J., and Wallace, E. M. (2002). Molecular weight forms of inhibin A, inhibin B and pro-αC in maternal serum, amniotic fluid and placental extracts of normal and Down syndrome pregnancies. Prenat. Diagn 22 (12), 1086–1092. doi:10.1002/pd.478
Tilley, S. K., Martin, E. M., Smeester, L., Joseph, R. M., Kuban, K. C. K., Heeren, T. C., et al. (2018). “Placental CpG methylation of infants born extremely preterm predicts cognitive impairment later in life,” 13. doi:10.1371/journal.pone.0193271
Trollmann, R., Schoof, E., Beinder, E., Wenzel, D., Rascher, W., and Dotsch, J. (2002). Adrenomedullin gene expression in human placental tIssue and leukocytes: a potential marker of severe tIssue hypoxia in neonates with birth asphyxia. Eur. J. Endocrinol. 147, 711–716. doi:10.1530/eje.0.1470711
Tsompanidis, A., Blanken, L., Broere-Brown, Z. A., Van Rijn, B. B., Baron-Cohen, S., and Tiemeier, H. (2023). Sex differences in placenta-derived markers and later autistic traits in children. Transl. Psychiatry 13 (1), 256. doi:10.1038/s41398-023-02552-w
Vacher, C. M., Lacaille, H., O’Reilly, J. J., Salzbank, J., Bakalar, D., Sebaoui, S., et al. (2021). Placental endocrine function shapes cerebellar development and social behavior. Nat. Neurosci. 24 (10), 1392–1401. doi:10.1038/s41593-021-00896-4
van der Meij, K. R. M., Sistermans, E. A., Macville, M. V. E., Stevens, S. J. C., Bax, C. J., Bekker, M. N., et al. (2019). TRIDENT-2: national implementation of genome-wide non-invasive prenatal testing as a first-tier screening test in The Netherlands. Am. J. Hum. Genet. 105 (6), 1091–1101. doi:10.1016/j.ajhg.2019.10.005
Wald, N. J., Rodeck, C., Hackshaw, A. K., and Rudnicka, A. (2004). SURUSS in perspective. BJOG Int. J. Obstet. Gynaecol. 111 (6), 521–531. doi:10.1111/j.1471-0528.2004.00193.x
Weinans, M. J., Sancken, U., Pandian, R., van de Ouweland, J. M. W., de Bruijn, H. W. A., Holm, J. P., et al. (2005). Invasive trophoblast antigen (hyperglycosylated human chorionic gonadotropin) as a first-trimester serum marker for down syndrome. Clin. Chem. 51 (7), 1276–1279. doi:10.1373/clinchem.2005.048751
Willemsen, R., Bontekoe, C. J., Severijnen, L. A., and Oostra, B. A. (2002). Timing of the absence of FMR1 expression in full mutation chorionic villi. Hum. Genet. 110 (6), 601–605. doi:10.1007/s00439-002-0723-5
Wong, O. G. W., Cheung, C. L. Y., Ip, P. P. C., Ngan, H. Y. S., and Cheung, A. N. Y. (2018). Amyloid precursor protein overexpression in down syndrome trophoblast reduces cell invasiveness and interferes with syncytialization. Am. J. Pathol. 188 (10), 2307–2317. doi:10.1016/j.ajpath.2018.07.004
Xiang, A. H., Wang, X., Martinez, M. P., Walthall, J. C., Curry, E. S., Page, K., et al. (2015). Association of maternal diabetes with autism in offspring. JAMA 313 (14), 1425–1434. doi:10.1001/jama.2015.2707
Yanni, D., Allred, E. N., Fichorova, R. N., O'Shea, T. M., Kuban, K., et al. (2017). Both antenatal and postnatal inflammation contribute information about the risk of brain damage in extremely preterm newborns. Pediatr. Res. 82 (4), 691–696. doi:10.1038/pr.2017.128
Yin, Y. Z., She, Q., Zhang, J., Zhang, P. Z., Zhang, Y., Lin, J. W., et al. (2014). Placental methylation markers in normal and trisomy 21 tissues: DNA methylation in trisomy 21 placental tissue. Prenat. Diagn 34 (1), 63–70. doi:10.1002/pd.4256
Zhang, M., Li, T., Chen, J., Li, L., Zhou, C., Wang, Y., et al. (2011). Non-invasive prenatal diagnosis of trisomy 21 by dosage ratio of fetal chromosome-specific epigenetic markers in maternal plasma. J. Huazhong Univ. Sci. Technol. Med. Sci. 31 (5), 687–692. doi:10.1007/s11596-011-0583-0
Zhang, Q., Lee, C. L., Yang, T., Li, J., Zeng, Q., Liu, X., et al. (2023). Adrenomedullin has a pivotal role in trophoblast differentiation: a promising nanotechnology-based therapeutic target for early-onset preeclampsia. Sci. Adv. 9 (44), eadi4777. doi:10.1126/sciadv.adi4777
Zhou, H., Zhao, C., Wang, P., Yang, W., Zhu, H., and Zhang, S. (2023). Regulators involved in trophoblast syncytialization in the placenta of intrauterine growth restriction. Front. Endocrinol. 14, 1107182. doi:10.3389/fendo.2023.1107182
Zhu, Y., Mordaunt, C. E., Yasui, D. H., Marathe, R., Coulson, R. L., Dunaway, K. W., et al. (2019). Placental DNA methylation levels at CYP2E1 and IRS2 are associated with child outcome in a prospective autism study. Hum. Mol. Genet. 28 (16), 2659–2674. doi:10.1093/hmg/ddz084
Zhu, Y., Gomez, J. A., Laufer, B. I., Mordaunt, C. E., Mouat, J. S., Soto, D. C., et al. (2022). Placental methylome reveals a 22q13.33 brain regulatory gene locus associated with autism. Genome Biol. 23 (1), 46. doi:10.1186/s13059-022-02613-1
Keywords: placental biomarkers, neuroplacentology, proteomics, trisomies, hypoxic-ischemic encephalopathy
Citation: Patel P, Ku J, Uzoaru I and Goldstein JA (2025) Placental biomarkers for the prediction of neurodevelopmental disorders. Front. Cell Dev. Biol. 13:1663960. doi: 10.3389/fcell.2025.1663960
Received: 11 July 2025; Accepted: 22 September 2025;
Published: 07 October 2025.
Edited by:
Clara Limback, Oxford University Hospitals, United KingdomReviewed by:
Clarissa Lim Velayo, University of the Philippines Manila, PhilippinesAndrea González Rojas, Pontificia Universidad Católica de Valparaíso, Chile
Copyright © 2025 Patel, Ku, Uzoaru and Goldstein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Payal Patel, cGF5YWx2cDJAaWxsaW5vaXMuZWR1; Joy Ku, am95bGsyQGlsbGlub2lzLmVkdQ==
†These authors have contributed equally to this work