 Grazieli Maboni1
Grazieli Maboni1 Rebecca Davenport1
Rebecca Davenport1 Kate Sessford1
Kate Sessford1 Kerstin Baiker1
Kerstin Baiker1 Tim K. Jensen2
Tim K. Jensen2 Adam M. Blanchard1
Adam M. Blanchard1 Sean Wattegedera3
Sean Wattegedera3 Gary Entrican3
Gary Entrican3 Sabine Tötemeyer1*
Sabine Tötemeyer1*- 1School of Veterinary Medicine and Science, University of Nottingham, Nottingham, United Kingdom
- 2National Veterinary Institute, Technical University of Denmark, Copenhagen, Denmark
- 3Moredun Research Institute, Penicuik, United Kingdom
Skin infection studies are often limited by financial and ethical constraints, and alternatives, such as monolayer cell culture, do not reflect many cellular processes limiting their application. For a more functional replacement, 3D skin culture models offer many advantages such as the maintenance of the tissue structure and the cell types present in the host environment. A 3D skin culture model can be set up using tissues acquired from surgical procedures or post slaughter, making it a cost effective and attractive alternative to animal experimentation. The majority of 3D culture models have been established for aerobic pathogens, but currently there are no models for anaerobic skin infections. Footrot is an anaerobic bacterial infection which affects the ovine interdigital skin causing a substantial animal welfare and financial impact worldwide. Dichelobacter nodosus is a Gram-negative anaerobic bacterium and the causative agent of footrot. The mechanism of infection and host immune response to D. nodosus is poorly understood. Here we present a novel 3D skin ex vivo model to study anaerobic bacterial infections using ovine skin explants infected with D. nodosus. Our results demonstrate that D. nodosus can invade the skin explant, and that altered expression of key inflammatory markers could be quantified in the culture media. The viability of explants was assessed by tissue integrity (histopathological features) and cell death (DNA fragmentation) over 76 h showing the model was stable for 28 h. D. nodosus was quantified in all infected skin explants by qPCR and the bacterium was visualized invading the epidermis by Fluorescent in situ Hybridization. Measurement of pro-inflammatory cytokines/chemokines in the culture media revealed that the explants released IL1β in response to bacteria. In contrast, levels of CXCL8 production were no different to mock-infected explants. The 3D skin model realistically simulates the interdigital skin and has demonstrated that D. nodosus invades the skin and triggered an early cellular inflammatory response to this bacterium. This novel model is the first of its kind for investigating an anaerobic bacterial infection.
Introduction
Skin infection studies are often limited by cost prohibitive experiments and ethical concerns, leading to a dependence on in vitro models utilizing traditional monolayer cell culture. The limitations of monolayer cell culture have been recently recognized as they do not reflect the in vivo cellular processes, due to the lack of multicellular interaction (Ren et al., 2006; MacNeil, 2007; Edmondson et al., 2014). Single cell models consequently may differ in gene and protein expression from in vivo models (Gurski et al., 2010; Price et al., 2012). Therefore, alternative in vitro methods are needed to provide physiologically relevant conditions in a system that can realistically simulate mechanisms of skin infections. Three-dimensional (3D) organ culture can be used to investigate bacterial infections based on in vitro culture of skin explants. These tissues can be acquired from surgical procedures or post slaughter (Sanders et al., 2002; Smijs et al., 2007; Steinstraesser et al., 2010; Sidgwick et al., 2016; Wang et al., 2016). Since the multicellular interaction and tissue cytoarchitecture are preserved, 3D models are considered to be phenotypically and histologically similar to the organs and tissues in vivo and their importance in the development of relevant models have been widely recognized (Ren et al., 2006; Edmondson et al., 2014). Research on bacterial infections using 3D skin culture has been mainly developed for facultative anaerobic and aerobic bacteria such as Staphylococcus aureus, Pseudomonas aeruginosa, and Acinetobacter spp. (Steinstraesser et al., 2010; de Breij et al., 2012; Popov et al., 2014). In contrast, there is no established 3D skin model to study anaerobic bacterial infections.
Dichelobacter nodosus is an anaerobic Gram-negative bacterium and the causative agent of footrot, an interdigital skin infection of sheep (Beveridge, 1941; Egerton et al., 1969; Kennan et al., 2010). The disease is characterized by the separation of the hoof from the underlying soft tissues and causes a substantial animal welfare issue leading to a significant financial impact for farmers worldwide (Hickford et al., 2005; Wassink et al., 2010; Rather et al., 2011). The etiology is complex and virulent strains of D. nodosus are important in the initiation of the disease (Beveridge, 1941; Kennan et al., 2010). Virulent and benign D. nodosus strains differ in their ability to degrade the extracellular matrix of host tissues using extracellular proteases, whereby the virulent AprV2 protease differs from its benign counterpart AprB2 by a single amino acid change (Tyr92Arg) (Riffkin et al., 1995; Kennan et al., 2010). A recent whole genome analysis of 103 D. nodosus strains, isolated from eight different countries, identified that D. nodosus has a global conserved bimodal population with two distinct clades. Clade I was generally more associated with virulent D. nodosus and Clade II with benign D. nodosus strains (Kennan et al., 2014). It is important to highlight that these clades do not always correlate with severity of clinical presentations, as virulent D. nodosus can be identified in sheep without any clinical signs (Stäuble et al., 2014) and benign D. nodosus strains have been isolated from underrunning footrot lesions of Swedish sheep (Frosth et al., 2015). The severity of footrot is thought to be exacerbated by the intense inflammatory response to the infection. However, little is known about the host immune response to this disease. IL1β was suggested to be involved in the inflammatory response to D. nodosus with high levels of expression, significantly associated with abundance in naturally infected skin biopsies (Davenport et al., 2014; Maboni et al., 2017). Conversely, high expression of other cytokines/chemokines, such as CXCL8, IL6 and IL17, were not correlated with D. nodosus abundance (Maboni et al., 2017). Additional evidence of the role played by IL1β was obtained using a single cell type model, where increased expression was measured in ovine fibroblasts stimulated with D. nodosus (Davenport et al., 2014). To date, this monolayer fibroblast model is the only in vitro cell-based method available to investigate footrot. In this context, we hypothesized that a 3D skin model could more realistically simulate the ovine interdigital skin microenvironment and D. nodosus could penetrate the skin, triggering an early local cellular inflammatory response.
In this study we describe the development of a novel 3D ex vivo skin culture model to study the infection caused by D. nodosus using ovine interdigital skin explants. We demonstrate that D. nodosus can invade the skin and alter the expression of key inflammatory markers. Tissue and cell viability was assessed and the early cellular response to bacteria was investigated by quantification of IL1β and CXCL8 protein release in the medium. In summary, this method is simple, robust and can be applied to investigate other anaerobic bacterial infections.
Materials and Methods
Sample Collection
Healthy ovine feet were collected from sheep at a local abattoir post-slaughter. Feet with good hoof conformation and with apparently healthy interdigital skin were selected. Environmental contaminants were removed from the foot and interdigital space using 70% ethanol and antimicrobial skin cleanser (Hibiscrub®) before removal of the hair using scissors. The entire ovine interdigital skin was removed using a sterile scalpel. Skin explants were immediately immersed in transport media after collection [DMEM-HAM'S F12 1:1 (Sigma Aldrich®), Penicillin+Streptomycin 0.01 μg/ml (Gibco®), Amphotericin B 0.01 μg/ml (Lonza®), Gentamicin 5 μg/ml (Sigma Aldrich®), L-glutamine 0.01 μg/ml (Gibco®)]. Biopsies were collected aseptically from each interdigital skin fragment using an 8 mm punch biopsy (National Veterinary Service, UK). The surface of the biopsy was gently incised 4 times with a scalpel blade to simulate naturally occurring damage to the skin (required for the invasion of D. nodosus).
Assembling the 3D Skin Explant Model
Biopsies were immersed in wash medium [DMEM-HAM'S F12 1:1, Penicillin+Streptomycin 0.01 μg/ml (Gibco®), Amphotericin B 0.01 μg/ml (Lonza®), Gentamicin 2 μg/mL (Sigma Aldrich®)], pre-heated to 37°C, and incubated at room temperature for 15 min. The media change and incubation steps were repeated 3 times.
To provide a physical support for the biopsies, an agarose pedestal (500 μl of 1.2% w/v agarose (Thermo Scientific®) in DMEM-HAM'S F12 1:1 (Sigma-Aldrich®) covered with 0.5 cm2 sterile surgical gauze was assembled in a 12 well-plate (Thermo Scientific®). The skin biopsy was placed on the top of the surgical gauze and agarose pedestal (Figure 1B).
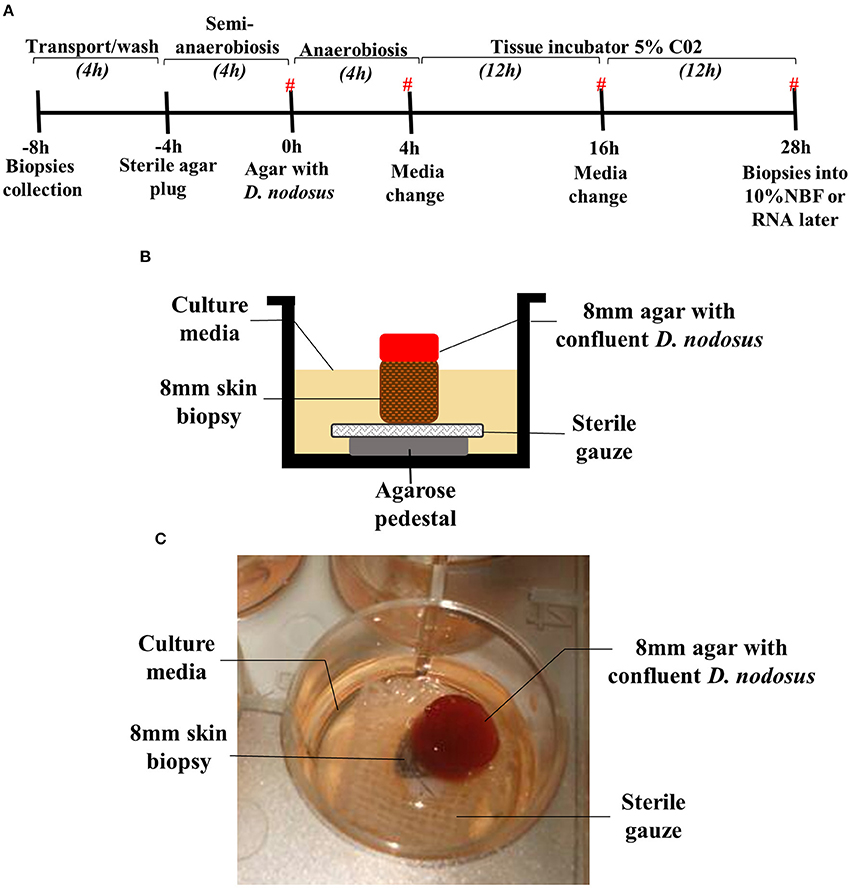
Figure 1. Assembled 3D skin explant model for anaerobic bacterial infection. (A) Timeline of the model infection highlighting the time points where biopsies were placed into RNAlater or 10% NBF. (#) Time points where the culture media were collected for cytokine measurement. Culture media without antibiotics was used between −4 and 4 h. Culture media containing antibiotics was used between 4 and 28 h after bacterial exposure. (B) Schematic overview of the assembled model at 0–28 h (exposure to Dichelobacter nodosus). (C) Photo of the assembled model in a 12 well-plate.
For the initial assessment of model viability, five time points were evaluated for up to 76 h of mock infection: biopsy collection −8 h, mock infection 0, 28, 52, and 76 h post mock infection. Two biopsies were collected at each time point; one placed into RNAlater (Sigma-Aldrich®) for DNA extraction and the other one was fixed in 10% v/v Neutral Buffer Formalin (NBF) for histological investigation. The model viability was analyzed assessing at each time point the tissue integrity and architecture (scoring of 2 H&E stained sections), and cell viability by TUNEL stain (5 non-overlapping images for each, epidermis and dermis).
Skin Explants Infection with D. nodosus
Four experiments using explants from different sheep collected in different occasions were performed. A microenvironment with restricted oxygen supply was produced by placing a sterile agarose plug (300 μl of 0.8% w/v agarose in DMEM-HAM'S F12 1:1) on the top of the biopsy, which was placed on the top of the agarose pedestal and surgical gauze, and incubated in a humidified tissue incubator at 37°C (Heracell 150i, Thermo Fisher Scientific®) with 5% CO2 for 4 h. 1 mL of culture media without antibiotics [DMEM high glucose (Gibco®) + DMEM HAM'S F12 1:1 (Sigma Aldrich®) in a proportion 3:1, Amphotericin B 0.25 μg/ml (Lonza®), L-glutamine 0.01 μg/ml (Gibco®), 10% Fetal Bovine Serum (Gibco®)] was placed in each well. The infection of the explants with D. nodosus was performed using an 8 mm punch biopsy to aseptically collect a Fastidious Anaerobic Agar plug confluent with D. nodosus strain MM261 (aprV2, Clade I, which correlates with virulent strains) or strain MM277 (aprB2, Clade II, which correlates with benign strains). The agar plug with D. nodosus was placed on the top of the biopsy and 1 mL of culture media without antibiotics was added in each well. The plate was incubated anaerobically (Oxoid AnaeroGen, Thermo Scientific®) at 37°C for 4 h. Post 4 h, maintenance of the model included culture media containing antibiotics [DMEM High Glucose (Gibco®) + DMEM HAM'S F12 1:1 (Sigma Aldrich®) in a proportion 3:1, Penicillin+Streptomycin 0.01 μg/ml (Gibco®), Gentamicin 5 μg/ml (Sigma Aldrich®), Amphotericin B 0.25 μg/ml (Lonza®), L-glutamine 0.01 μg/ml (Gibco®), 10% v/v Fetal Bovine Serum (Gibco®)] in a humidified tissue incubator with 5% C02for 24 h. The media were changed 4 h post infection and then every 12 h keeping the same agar plug with D. nodosus placed on the top of the biopsy. Culture media from all time points were collected and stored at −80°C for further cytokine measurement. After 28 h of infection with D. nodosus, biopsies were placed into RNAlater for further DNA extraction or 10% v/v NBF for histology. For DNA extraction each tissue was cut into approximately 4 × 4 mm pieces and incubated with 180 μl of tissue lysis buffer (ATL) with the addition of 20 μl of proteinase K (20 mg/ml) (Qiagen) at 56°C for 3 h. DNA was isolated using the QIAamp cador kit according to manufacturer's recommendations and eluted in 50 μl AVE buffer (Qiagen). The DNA was used for D. nodosus quantification by qPCR, targeting the 16S rRNA gene (Forward primer: 5′-CGGGGTTATGTAGCTTGCTATG-3′, Reverse primer: 5′-TACGTTGTCCCCCACCATAA-3′, probe: 5′FAM-TGGCGGACGGGTGAGTAATATATAGGAATC-TAMRA-3′) (Frosth et al., 2012). qPCR data were normalized to pg of D. nodosus DNA present in the total DNA extracted from each biopsy. The cut off of 0.1pg was assigned for negative qPCR results. The D. nodosus load present on an 8 mm agar area was estimated using the total DNA extracted from D. nodosus confluent on the 8 mm surface and confirmed by qPCR.
Skin Tissue Fixing, Processing, and Staining
Interdigital skin biopsies were fixed for 48 h at 4°C using an extended tissue processing protocol (60 min in dH2O, 4 h in 50% ethanol, 4 h in 70% ethanol, 16 h in 90% ethanol, 4 h in 100% ethanol, 4 h in xylene, 2 h of wax immersion at 60°C). Paraffin wax embedded tissues were soaked in 10% (v/v) ammoniated water while 3 and 6 μm thick sections were cut from each block by microtome (Leica RM2255®) and placed on polystyrene microscope slides (Thermo Scientific®). H&E stain (Sigma-Aldrich®) was used to allow visual assessment of the 6 μm tissue sections (see Supplementary Table 1 for H&E protocol). All slides were mounted with Distyrene Plasticizer Xylene (DPX) (Sigma-Aldrich®) and were analyzed by microscopy (Leica DM 2500®). A board certified veterinary pathologist, blinded to slide identity, evaluated 54 H&E-stained biopsy sections to assess tissue integrity and architecture. A qualitative, semi-quantitative scoring system was developed to grade the tissue integrity and architecture according to histopathological features (Table 1 and Supplementary Figure 2).
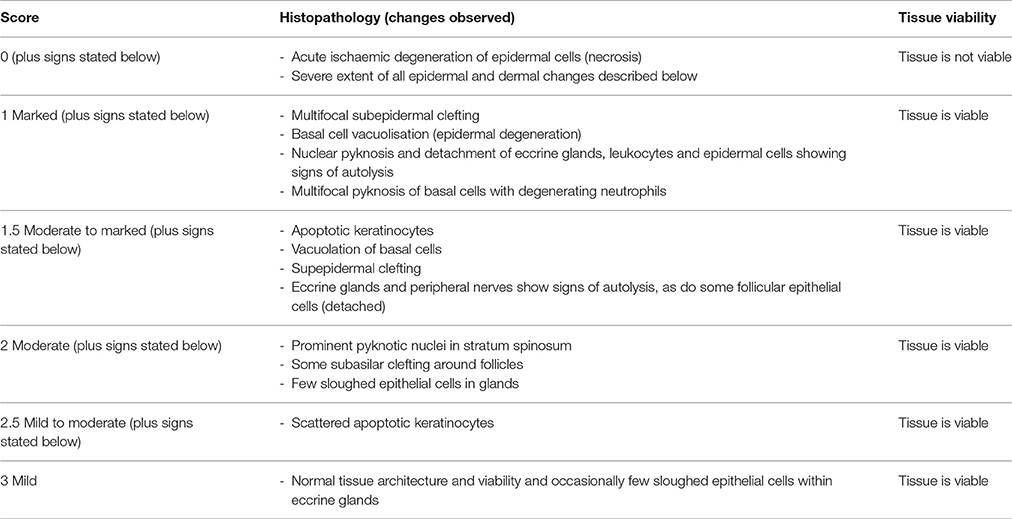
Table 1. Qualitative, semi-quantitative scoring system used to evaluate the tissue integrity and viability of the ovine skin explants from the 3D culture model.
The modified DeadEnd Colorimetric TUNEL System (Promega®) was used to detect DNA fragmentation. Assays were performed according to the manufacturer's instructions using 6 μm tissue sections. Cells were stripped of proteins and made permeable by incubation with 20 μg/ml of proteinase K solution for 20 min and DAB 20x chromogen solution incubation was performed for 3 min. The sections were counterstained with haematoxylin solution for 5 s, rinsed in tap and deionized water. All slides were mounted with DPX. Negative controls were obtained by omitting the TdT enzyme from the TdT mix in the reaction (according to manufacturer's instructions). Positive controls were generated by treating a skin section with DNAse (Promega®). In order to assess the number of dead and live cells, 5 non-overlapping images from each, the epidermis and the dermis of each biopsy tissue section, were used (200x total magnification). Images were captured and analyzed by microscopy (CTR500 microscope, Leica Microsystems®). All cells from all images (epidermis and dermis) from each sample were counted using Fiji/ImageJ-win64 software (https://fiji.sc/, October 2016) and the percentage of dead and live cells were calculated for all 10 images/sample based on the total number of dead and live cells obtained. Brown stained nuclei were assessed as dead cells, whereas blue stained nuclei were assessed as live cells due to counterstain with haematoxylin.
Fluorescent in Situ Hybridization (FISH) for D. nodosus and Eubacteria Detection And Localization
FISH analysis was performed as previously described (Rasmussen et al., 2012). The 16S rRNA-targeting oligonucleotide probes for D. nodosus (S-S-D.nodosus-443 5′-CATGCACCGTTCTTCACT-3') and eubacteria domain (S-D-eub-338-alexa 5′-GTCATTCCATCGAAACATA-3′) labeled with fluorescein isothiocyanate (FITC) or Cy3 were used on 3 μm tissue sections. Hybridization was carried out at 46°C with hybridization buffer (100 mM Tris, pH 7.2, 0.9 M NaCl, 0.1% sodium dodecyl sulfate) containing 5 ng/ml of each applied oligonucleotide probe (usually double hybridizations with a FITC and CY3 labeled probe) for 16 h in a Sequenza slide rack (Thermo Shandon, Cheshire, United Kingdom). Slides were then washed with prewarmed (46°C) hybridisation buffer for 3 × 3 min followed by wash with prewarmed (46°C) washing buffer (100 mM Tris, pH 7.2, 0.9 M NaCl) for 3 × 3 min. Finally they were rinsed in water, air dried and mounted with Vectashield (Vector Laboratories Inc.®) for epifluorescense microscopy using an Axioimager M1 epifluorescense microscope. Images were obtained using an AxioCAM MRm version 3 FireWiremonochrome camera (Carl Zeiss®).
Cytokine/Chemokine Measurement by Enzyme-Linked Immunosorbent Assay (ELISA)
IL1β and CXCL8 were measured in the tissue culture media collected from −4 to 0 h, 0 to 4 h, 4 to 16 h, and 16 to 28 h post infection, using specific sandwich ELISAs as previously described (Doull et al., 2015). Recombinant ovine IL1β (Kingfisher®) and recombinant ovine CXCL8 (Doull et al., 2015) were used as quantifiable ELISA standards. Cytokine concentrations were calculated using a polynomial quadratic regression in GraphPad Prism version 7b.
Statistical Analysis
One-way analysis of variance followed by Dunn's multiple comparisons test was applied for D. nodosus load as well as for percentage of TUNEL positive cells. Mann Whitney (non-parametric test) was used for IL1β release between infected and mock-infected biopsies across the time course. Mean, media, standard deviation (SD) and all other analysis were performed using GraphPad Prism version 7b. A P ≥ 0.05 was considered significant.
Results
Assembling the 3D Skin Model
The assembled 3D skin infection model consisted of an agarose pedestal overlaid with surgical gauze to provide support for the biopsy in the well of a 12 well-plate (Figure 1B). The skin biopsy was then placed upon the surgical gauze. The skin explants were initially maintained during 4 h under atmospheric oxygen, but the biopsy surface area was sealed with a sterile agar plug to simulate the natural ovine feet microenvironment with a moist surface and restricted oxygen supply (Figures 1A,B). For the infection experiment, the sterile agar plug was replaced by an agar plug with confluent D. nodosus placed on the top of the biopsy and incubated under anaerobiosis for 4 h, followed by 24 h of aerobic incubation to ensure skin tissue survival (Figure 1A). For the mock-infection viability experiment, the sterile agar plug was replaced by another sterile agar plug and the model was maintained under the same temperature and incubation conditions as described above. Up to 1 ml of medium was added up to the top level of the explant to avoid flooding of the explant or removal of the agar plug.
Tissue Integrity and Cell Viability Maintenance during 76 h of Model Incubation
A 76 h time course mock-infection experiment was performed to identify optimal conditions for oxygen dependent tissue to remain viable, while also allowing transient anaerobic incubation essential for the pathogen (Supplementary Figure 1A). The viability of explants was assessed by tissue integrity and cell death at five time points: immediately after sample collection −8 h, mock-infection 0, 28 h, 52 and 76 h post mock infection. Using the scoring system developed to grade the tissue integrity and architecture according to histopathological features (Table 1 and Supplementary Figure 2), it was visualized that skin explants maintained the normal tissue architecture for up to 28 h of incubation with early signs of tissue degeneration in biopsies incubated for more than 52 h (Supplementary Figure 1B). The early signs of tissue degeneration included basal cell vacuolisation, subepidermal clefting, sloughed cells in glands and follicular epithelial cells (Supplementary Figure 1B). Additionally, cell viability was investigated through the quantification of DNA fragmentation using the TUNEL stain. The percentage of live cells in the epidermis decreased from −8 h [99% (2497/2524)) to 76 h of incubation (45% (296/660)] (Supplementary Figures 1C, 3). A high percentage of live cells was found in the dermis in all time points, with a slight decrease from −8 h [99.3% (2092/2105)] to 76 h [84% (952/1131)] (Supplementary Figure 1C). As expected, biopsies fixed at the abattoir post slaughter had a low level of dead cells. The cell viability data confirmed the tissue integrity data as skin explants presented a high percentage of live cells (TUNEL) and well preserved tissue architecture (H&E tissue integrity scores) in both epidermis and dermis (Supplementary Figure 1C). As the 28 h incubation showed more than 50% of the epidermal and dermal cells were alive and the tissue architecture was viable (score 1.5) (Supplementary Figure 1C), this time point was used as the implemented cut-off for further infection experiments.
Ovine Skin Explant as an Infection Model for Anaerobic Bacteria
The D. nodosus of the 8 mm agar plug was quantified by qPCR to estimate the copy number used to infect each biopsy (1.13 × 104 of aprV2 D. nodosus DNA, strain MM261, and 7.16 × 104 of aprB2 D. nodosus DNA, strain MM277). The presence of D. nodosus DNA was confirmed in all infected biopsies with aprV2 (2.13 × 104 D. nodosus/biopsy, n = 6) and aprB2 D. nodosus (1.11 × 105 D. nodosus/biopsy, n = 6) using qPCR. In contrast, all mock-infected biopsies (n = 8) were negative for D. nodosus (Figure 2A).
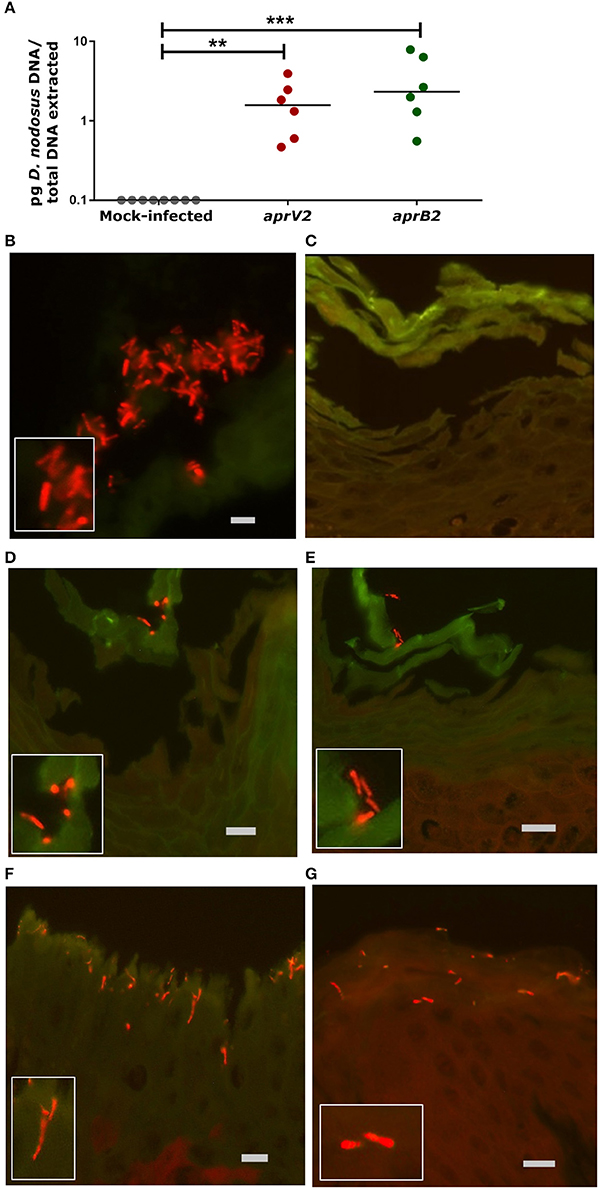
Figure 2. Infection of the 3D skin explant model with Dichelobacter nodosus. (A) Detection of D. nodosus aprV2 and aprB2 strains in the skin explants using quantitative PCR after 28 h of infection. Each point indicates a single biopsy. 0.1 = results below of the limit of detection. Mean is represented by black bars. Data were analyzed by Dunn's multiple comparisons test using GraphPad Prism **P ≤ 0.01, ***P ≤ 0.001. (B–G) Fluorescent in situ Hybridization on ovine interdigital skin explants after 28 h of infection with D. nodosus. (B) Positive tissue control with D. nodosus reference strain (CCUG 27824) (red/orange); (C) Uninfected negative control; (D,E) Demonstration of aprB2 and (F,G) aprV2 D. nodosus (red/orange) on the surface or migrating within the epidermal layers. (C–E) were hybridized with the Cy3 labeled D. nodosus probe only, while (F,G) were hybridized with both, the D nodosus and the eubacteria probe. Squares located on the bottom/left side show the zoomed image. Scale bars (gray): (B), 5 μm, (C–G), 10 μm.
Fluorescent in situ hybridization (FISH) was used to localize D. nodosus within the skin explant. D. nodosus bacterial cells were identified in the positive porcine lung tissue control (Figure 2B) and in all biopsies infected with either aprV2 (n = 4) or aprB2 D. nodosus (n = 4; Figures 2D–G). Negative tissue control (n = 1; Figure 2C) and all mock-infected skin control biopsies (n = 5) were negative for D. nodosus (Table 2). D. nodosus was identified on the skin surface (Figures 2D–E) and also invading the superficial layers of the epidermis (Figures 2F,G). D. nodosus cells were visualized throughout these layers suggesting that the bacteria were moving away from the initial skin laceration site. D. nodosus was not identified in the dermis. A general eubacterial domain probe did not detect the presence of other bacteria in any infected or mock-infected explants. This showed that both aprV2 and aprB2 strains of D. nodosus have the ability to migrate into the damaged skin layers during 28 h of incubation.

Table 2. Fluorescent in situ Hybridization of targeted Dichelobacter nodosus on ovine interdigital skin tissue samples from the 3D skin culture model.
Bacterial Infection Had Minimal Impact on Skin Explants Viability Over 28 h
The tissue integrity and cell viability from infected and mock-infected skin explants were assessed to investigate the histological effects of bacterial infection on the tissue viability and maintenance. D. nodosus infection had little impact on tissue integrity/architecture or on cell viability after 28 h (Figure 3). Tissue viability of all mock-infected control biopsies was maintained over 28 h of incubation (median histopathology score 2.5), with the exception of one biopsy (score 1.5) (Figure 3A; See Table 1 for description of histopathology scores). Similarly, infected biopsies maintained the tissue integrity (median histopathology score 2), with the exception of two biopsies (scores 1 and 1.5) (Figure 3A). All mock-infected controls and infected explants presented more than 50% (69–98.9%) of epidermal and dermal cells alive (TUNEL stain) after 28 h of bacterial exposure, with the exception of one biopsy infected with the aprV2 D. nodosus strain (Figure 3B). There was no statistically significant difference between infected and mock-infected skin explants for the percentage of live cells.
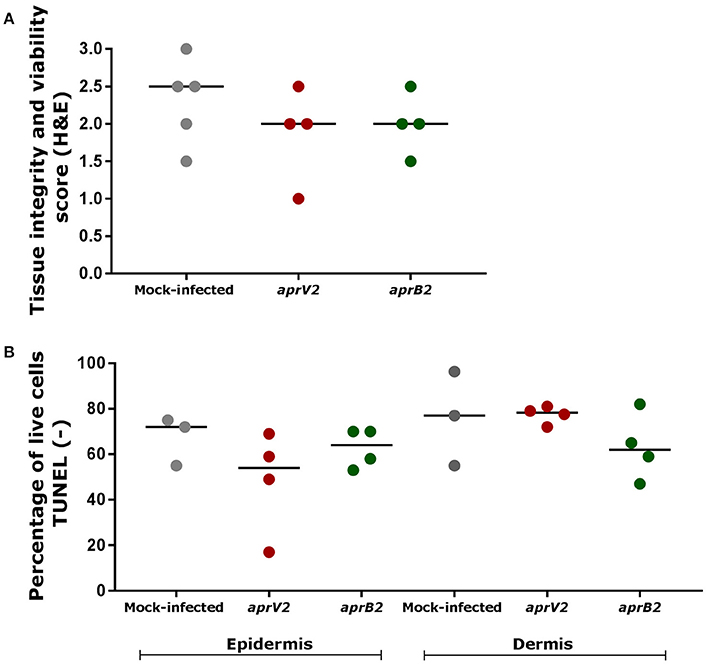
Figure 3. Tissue integrity and cell viability assessment of skin explants cultured in the 3D model after 28 h of infection with aprV2 and aprB2 Dichelobacter nodosus. (A) Tissue integrity and viability score (H&E) after 28 h of bacterial infection for both epidermis and dermis. Histological score 0: tissue is not viable; score 1: viable tissue, but showing marked signs of tissue degeneration; score 1.5: viable tissue, but showing moderate to marked signs of tissue degeneration; score 2: viable tissue, but showing moderate signs of tissue degeneration; score 2.5: viable tissue, but showing mild to moderate signs of tissue degeneration; score 3: tissue is viable and shows only few mild signs of tissue degeneration. (B) Percentage of live cells (TUNEL negative, no DNA fragmentation) in the epidermis and dermis of skin explants after 28 h of bacterial infection. Each point indicates a single explant. Black bars indicate median. TUNEL data were analyzed by Dunn's multiple comparisons test using GraphPad Prism.
Pro-inflammatory Cytokine/Chemokine Released in the Culture Media
Skin explants infected with aprV2 and aprB2 D. nodosus released IL1β in the culture media, which accumulated between 4–16 h and 16–28 h post infection (Figure 4A). At 4–16 h, the concentration of IL1β in infected biopsies (600 ± 492 IL1β pg/mL, n = 8) was statistically significantly greater than in mock-infected controls (10 ± 20 IL1β pg/mL, n = 4) (P ≤ 0.01). All media samples from mock-infected and infected biopsies collected at −4 h mock infection and 4 h after infection (4 h of accumulation) were negative or showed very low concentrations of IL1β (Figure 4A). Mock-infected controls and infected skin explants released similar concentrations of CXCL8 in the culture media that were accumulated between 0–4 h, 4–16 h and 16–28h post infection (Figure 4B). CXCL8 was not released in the culture media between −4 and 0 h of mock infection time (Figure 4B).
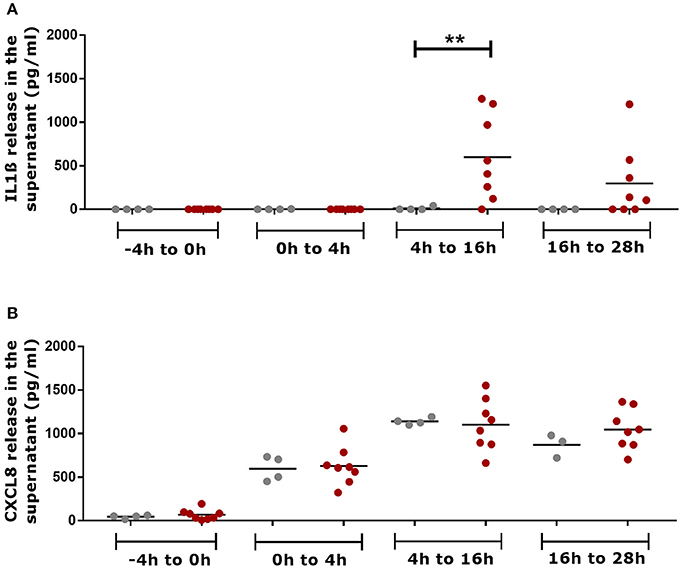
Figure 4. Measurement of pro-inflammatory mediators in the culture media supernatants of the 3D skin explant model using IL1β and CXCL8 specific ELISAs. (A) Accumulative detection of IL1β in mock-infected and infected explants with aprV2 and aprB2 Dichelobacter nodosus at different time points. (B) Accumulative detection of CXCL8 in mock-infected and infected explants. −4–0 h = total accumulation in the media supernatant during 4 h of semi-anaerobic conditions (mock infection); 0–4 h = total accumulation in the media supernatant 4 h post infection with D. nodosus; 4–16 h total accumulation in the media 4–16 h post infection with D. nodosus (12 h) and 16–28 h = total accumulation in the media 16–28 h post infection with D. nodosus (12 h). Mann Whitney test (non-parametric) was performed for comparison between mock-infected and infected explants using GraphPad Prism **P ≤ 0.01. Each point indicates a single explant. Gray points indicate mock-infected and red points indicate D. nodosus infected explants. Black bars represent mean.
Discussion
This study presents a novel 3D skin model culture using ex vivo explants for infection with anaerobic bacteria. The key findings include that both, aprV2 and aprB2 strains of D. nodosus, were able to migrate from the agar plug into tissues after 28 h of exposure, and those tissues responded to bacterial infection with the release of IL1β into the culture supernatant. To our knowledge, this is the first study to establish a 3D culture model using ovine skin explants to investigate an anaerobic bacterial infection. The use of ex vivo ovine skin in 3D culture has the advantage that explants are easily available from abattoirs, where they are by-products of the slaughter process.
Migration of D. nodosus into the skin explants was confirmed by qPCR and FISH. D. nodosus was clearly visualized, not only on the skin surface, but also invading the stratum corneum and superficially in the stratum granulosum in all infected biopsies. These findings correspond with an early report that D. nodosus was restricted to the superficial epidermal layers of the ovine skin (Egerton et al., 1969), and those by Witcomb et al. (2015), who also localized D. nodosus by FISH within the superficial epidermal layer of ovine biopsies naturally affected by footrot. We further investigated the initial inflammatory response after bacterial challenge to determine the immunological functionality of the cultured skin. Importantly, all mock-infected controls did not release IL1β suggesting this cytokine was primarily released in response to D. nodosus infection. IL1β has a range of stimulatory effects on immune cells playing a key role in the innate immunity of the skin (Arend et al., 2008). It has been reported elsewhere that the mRNA expression levels of IL1β and CXCL8 increase after human skin is exposed to Acinetobacter spp., while IL1β protein is undetectable in the culture media (de Breij et al., 2012). We have shown previously that IL1β mRNA is expressed in footrot samples (Davenport et al., 2014; Maboni et al., 2017). mRNA expression of TLR4 and TLR2 was also increased in these samples (Davenport et al., 2014), which likely resulted in the activation of signaling pathways culminating in the transcription and translation of the inactive precursors of IL1β. Processing of the IL1ß precursor and release into the medium demonstrates that the inflammasome multiprotein complex is involved in the inflammatory response triggered by D. nodosus. The inflammasome comprises of the activation of caspase-1 protease, which is responsible for the proteolytic processing of the IL1β precursor hence catalyzing the posttranslational mechanism that is required for the secretion of the bioactive form of this cytokine (Fantuzzi et al., 1997; Dinarello, 2006).
CXCL8 plays a role as a potent chemoattractant for neutrophils, basophils and T cells as well as having an effect on the proliferation of keratinocytes (Tuschil et al., 1992). CXCL8 in the skin is mainly produced by keratinocytes, which increases the expression of this chemokine in response to inflammatory stimuli (Cataisson et al., 2006). Here we found similar concentrations of CXCL8 obtained from mock-infected and infected explants with an increased release from 0 to 28 h. The high levels of CXCL8 observed in the mock-infected controls in response to the model incubation conditions may have masked any response to D. nodosus. However, we have found previously that high mRNA expression levels of CXCL8 were not associated with high D. nodosus load in naturally infected skin biopsies (Maboni et al., 2017). In contrast, CXCL8 has been reported to be significantly more expressed in response to Staphylococcus aureus, Pseudomonas aeruginosa and Acinetobacter spp. in human skin infection models (Steinstraesser et al., 2010; de Breij et al., 2012). In the context of anaerobic bacterial infections, mRNA expression of IL1α, CXCL8, TNFα and human beta defensin (hBD)-2 were stimulated in keratinocytes by Propionibacterium acnes, an anaerobic bacterium (Lee et al., 2010). Also, IL17 was shown to be essential in the host defense against Staphylococcus aureus, a facultative anaerobic bacterium (Cho et al., 2010). The skin model developed in this study could be used to investigate the release of these molecules in response to other anaerobic bacteria.
Natural occurrence of footrot requires a damp and damaged skin microenvironment to allow D. nodosus to establish in the interdigital skin and to initiate under-running footrot lesions (Beveridge, 1941; Kennan et al., 2010). In vivo infection trials simulated the natural occurrence of footrot maintaining the sheep feet under wet conditions that allowed maceration of the interdigital skin prior to bacterial infection (Beveridge, 1941; Egerton et al., 1969; Kennan et al., 2001, 2010; Knappe-Poindecker et al., 2014). In order to replicate these conditions in vitro, the epidermis of each explant was lacerated before infection with D. nodosus. Skin explants were never allowed to dry out during transport and kept hydrated with restricted oxygen supply through the use of an agar plug, throughout the experiment. In addition, footrot has been reported to be a localized foot infection with little or unknown involvement of the ovine systemic response (Bhardwaj et al., 2014; Davenport et al., 2014). This model therefore represents a suitable approach to investigate footrot in the context of local skin architecture and cellular components.
Tissue integrity and cell viability of the skin explants were maintained over 28 h of incubation, with more marked signs of tissue degeneration in biopsies incubated for more than 52 h. Aspects that may have affected tissue survivability include animal age and breed, equal distribution of nutrients within the tissue, culture media pH and antimicrobials toxicity in the tissue explants. In this study, a heterogeneous population of sheep sampled at the abattoir from different breeds, ages and underlying subclinical disease may have impacted on tissue viability, as observed in mock-infected biopsies presenting variable integrity scores and percentage of live cells (Figure 3).
We showed that FISH targeting a general eubacterial domain did not detect the presence of other bacteria in any infected or mock-infected explants, which were likely eliminated by the antimicrobial agents of the washing medium. Although antibiotics and antifungal agents were essential in the medium to prevent contamination, they may have had a detrimental effect on tissue survivability. Since 3D models lack circulatory and excretion systems, it is likely that cytotoxic effects may happen due to the accumulation of antimicrobials in the tissue explants (Levy, 2000; Gibson et al., 2008; Costa et al., 2016). Considering that D. nodosus and other microorganisms may have a synergistic relationship in the pathogenesis of footrot (Maboni et al., 2016, 2017), the clearance of skin commensal microorganisms may also have an impact on the mechanism of D. nodosus establishment in the skin explants as well as the immune response. As expected, DNA fragmentation investigated through the TUNEL stain revealed that the percentage of live cells in the epidermis decreased from −8 h (99%) to 76 h of incubation (45%). Part of the cell death detected in the skin explants might be associated with the normal life cycle of keratinocytes, whereby DNA fragmentation for apoptosis and differentiation mechanisms occur to allow cornification of the keratinocytes that establish a tight barrier of dead cells protecting the skin (Lippens et al., 2005).
The model developed in this study could be used to generate new insights into the pathogenesis of ovine footrot as well as other bacterial infections. We envision this model as an in vitro alternative to investigate the synergistic role of D. nodosus and other bacterial species involved in footrot lesions such as F. necrophorum, Treponema spp. and Mycoplasma spp. (Beveridge, 1941; Frosth et al., 2015; Maboni et al., 2017). Further studies could investigate the role of twitching motility in the early stages of infection. Cytokine arrays could be applied to investigate a wider range of inflammatory molecules triggered in response to bacterial exposure. Importantly, the skin explant model could be improved in terms of incubation period. A time longer than 28 h of model exposure to bacteria would inform whether this model is conducive to bacterial replication as well as bacterial invasion studies. Extending the viability of the explant model could be achieved by developing a perfusion system to optimize the nutrient supply or developing a set up where more oxygen can be supplied to the tissues.
In summary, a novel skin explant model was developed using ovine ex vivo interdigital skin biopsies. Both, aprV2 and aprB2 D. nodosus migrated into the skin layers and IL1β and CXCL8 were released in the culture media indicating the tissues were alive after 28 h of bacterial exposure. IL1β in particular was shown to be released in response to D. nodosus challenge. We demonstrated a proof of principle that the anaerobic bacterium D. nodosus could invade the 3D skin explant model and that the expression of key inflammatory markers could be quantified.
Author Contributions
Conception and design of study: ST and GM. Acquisition and analysis of data: GM, RD, KS, KB, TJ, AB, SW, GE, and ST. Drafting of article and/or critical revision: GM, RD, KB, TJ, AB, SW, GE, and ST. All authors approved the final article.
Funding
This work was supported by the Biotechnology and Biological Sciences Research Council [grant numbers BB/M012085/1, BB/M011941/1] (BBSRC) Animal Health Research Club 2014 (University of Nottingham, AB, ST and Moredun Research Institute, GE, SW respectively). GE and SW were also supported by the Scottish Government Rural and Environment Science and Analytical Services (RESAS) Division. RD, KS, and KB was supported by the University of Nottingham and GM was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) and the University of Nottingham.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Dr. Catrin Rutland, Aziza Alibhai and Ceri Staley for the technical support provided in the preparation of histological samples.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fcimb.2017.00404/full#supplementary-material
Supplementary Figure 1. Viability of the interdigital skin explants over a time course of 76 h (mock-infected explants). (A) Timeline of viability experiment where one biopsies was placed into RNAlater and one into 10% NBF at each time point. −8h (biopsy fixed at the abattoir a few minutes after sheep slaughter); 0 h (biopsy fixed at MOCK infection time); 28 h (biopsy fixed 28 h after MOCK infection time); 52 h (biopsy fixed 52 h after MOCK infection time); 76 h (biopsy fixed 76 h after MOCK infection time). (B) H&E images illustrating the overall maintenance of the tissue structure and viability over 76 h of incubation. Early signs of tissue degeneration were more marked at 52 and 76 h of incubation including apoptotic keratinocytes (1), basal cell vacuolisation (2), subepidermal clefting (3) (scale bars: 100 μm). (C) Comparison of cell viability (TUNEL) and tissue integrity (H&E scores) in the epidermis and in the dermis of ovine interdigital skin. Histological score 0: tissue is not viable; score 1: viable tissue, but showing marked signs of tissue degeneration; score 1.5: viable tissue, but showing moderate to marked signs of tissue degeneration; score 2: viable tissue, but showing moderate signs of tissue degeneration; score 2.5: viable tissue, but showing mild to moderate signs of tissue degeneration; score 3: tissue is viable and shows only few mild signs of tissue degeneration. Mean and standard deviation of live cells from 5 non-overlapping images from the epidermis are shown. For H&E analysis, 2 slides were analysed for each single biopsy; for all time points, both slides received the same histopathological score. Black horizontal line indicates histopathology score 1, the minimal score for a tissue to be deemed viable. Blue bars show proportion of TUNEL-negative cells. Pink bars indicate tissue integrity and viability score (H&E). Each bar indicates single biopsies.
Supplementary Figure 2. H&E images illustrating examples of the qualitative, semi-quantitative scoring system used to evaluate skin explants viability (mock-infected explants). (A) Histological score 0 (tissue not viable) 1 = subepidermal clefting, 2 = basal cell vacuolisation, 3 = necrotic area. (B) Histological score 1 (viable tissue), marked 1 = subepidermal clefting, 2 = basal cell vacuolisation. (C) Histological score 2 (viable tissue), mild to moderate, 4 = apoptotic keratinocyte. (D) Histological score 3 (viable tissue), mild, 5 = (single) few sloughed cells within eccrine gland. Scale bars: (A) 200 μm, (B–D) 100 μm.
Supplementary Figure 3. Demonstration of DNA fragmentation (TUNEL) in epidermal cells of the skin explants over a time course of 76 h (viability experiment with mock-infected explants). (A) Negative control slide of the TUNEL assay (tissue section not treated with DNAse); (B) Positive control slide of the TUNEL assay (tissue section treated with DNAse); (C) Skin explant fixed at −8 h (at the abattoir few minutes after animal slaughter); (D) 0 h (biopsy fixed at MOCK infection time); (E) 28 h (biopsy fixed 28 h after MOCK infection time); (F) 52 h (biopsy fixed 52 h after MOCK infection time); (G) 76 h (biopsy fixed 76 h after MOCK infection time). Purple nuclei: live cell (TUNEL negative suggesting there was no DNA fragmentation); Brown nuclei: dead cell (TUNEL positive revealing DNA fragmentation). (Scale bar = 50 μm).
References
Arend, W. P., Palmer, G., and Gabay, C. (2008). IL-1, IL-18, and IL-33 families of cytokines. Immunol. Rev. 223, 20–38. doi: 10.1111/j.1600-065X.2008.00624.x
Beveridge, W. I. B. (1941). Foot-rot in sheep: a transmissible disease due to infection with Fusiformis nodosus (n. sp.): studies on its cause, epidemiology and control. CSIRO Aust. Bull. 140, 1–56.
Bhardwaj, V., Dhungyel, O., de Silva, K., and Whittington, R. J. (2014). Investigation of immunity in sheep following footrot infection and vaccination. Vaccine 32, 6979–6985. doi: 10.1016/j.vaccine.2014.10.031
Cataisson, C., Pearson, A. J., Tsien, M. Z., Mascia, F., Gao, J. L., Pastore, S., et al. (2006). CXCR2 ligands and G-CSF mediate PKCα-induced intraepidermal inflammation. J. Clin. Invest. 116, 2757–2766. doi: 10.1172/JCI27514
Cho, J. S., Pietras, E. M., Garcia, N. C., Ramos, R. I., Farzam, D. M., Monroe, H. R., et al. (2010). IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Invest. 120:1762. doi: 10.1172/JCI40891
Costa, M. O., Harding, J. C. S., and Hill, J. E. (2016). Development and evaluation of a porcine in vitro colon organ culture technique. Vitr. Cell. Dev. Biol. Anim. 52, 942–952. doi: 10.1007/s11626-016-0060-y
Davenport, R., Heawood, C., Sessford, K., Baker, M., Baiker, K., Blacklaws, B., et al. (2014). Differential expression of toll-like receptors and inflammatory cytokines in ovine interdigital dermatitis and footrot. Vet. Immunol. Immunopathol. 161, 90–98. doi: 10.1016/j.vetimm.2014.07.007
de Breij, A., Eveillard, M., Dijkshoorn, L., van den Broek, P. J., Nibbering, P. H., and Joly-Guillou, M. L. (2012). Differences in Acinetobacter baumannii strains and host innate immune response determine morbidity and mortality in experimental pneumonia. PLoS ONE 7, 1–8. doi: 10.1371/journal.pone.0030673
Dinarello, C. A. (2006). Interleukin 1 and interleukin 18 as mediators of inflammation and the aging process. Am. J. Clin. Nutr. 83, 447S–455S. Available online at: http://ajcn.nutrition.org/content/83/2/447S.full.pdf+html
Doull, L., Wattegedera, S. R., Longbottom, D., Mwangi, D., Nath, M., Glass, E. J., et al. (2015). Late production of CXCL8 in ruminant oro-nasal turbinate cells in response to Chlamydia abortus infection. Vet. Immunol. Immunopathol. 168, 97–102. doi: 10.1016/j.vetimm.2015.08.011
Edmondson, R., Broglie, J. J., Adcock, A. F., and Yang, L. (2014). Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 12, 207–218. doi: 10.1089/adt.2014.573
Egerton, J. R., Roberts, D. S., and Parsonson, I. M. (1969). The aetiology and pathogenesis of ovine foot-rot. J. Comp. Pathol. 79, 217–227. doi: 10.1016/0021-9975(69)90007-3
Fantuzzi, G., Sacco, S., Ghezzi, P., and Dinarello, C. A. (1997). Physiological and cytokine responses in IL-1 beta-deficient mice after zymosan-induced inflammation. Am. J. Physiol. 273, R400–R406.
Frosth, S., König, U., Nyman, A. K., Pringle, M., and Aspán, A. (2015). Characterisation of Dichelobacter nodosus and detection of Fusobacterium necrophorum and Treponema spp. in sheep with different clinical manifestations of footrot. Vet. Microbiol. 179, 82–90. doi: 10.1016/j.vetmic.2015.02.034
Frosth, S., Slettemeas, J. S., Jorgensen, H. J., Angen, O., and Aspan, A. (2012). Development and comparison of a real-time PCR assay for detection of Dichelobacter nodosus with culturing and conventional PCR: harmonisation between three laboratories. Acta Vet. Scand. 54:6. doi: 10.1186/1751-0147-54-6
Gibson, A. L., Schurr, M. J., Schlosser, S. J., Comer, A. R., and Allen-Hoffmann, B. L. (2008). Comparison of therapeutic antibiotic treatments on tissue-engineered human skin substitutes. Tissue Eng. A 14, 629–638. doi: 10.1089/tea.2007.0126
Gurski, L. A., Petrelli, N. J., Jia, X., and Farach-carson, M. C. (2010). 3D Matrices for anti-cancer drug testing and development. Oncol. Issues 25, 20–25. Available online at: http://www.tandfonline.com/doi/abs/10.1080/10463356.2010.11883480
Hickford, J. G. H., Davies, S., Zhou, H., and Gudex, B. W. (2005). A survey of the control and financial impact of footrot in the New Zealand Merino industry. Proc. New Zeal. Soc. An. 65, 117–122. Available online at: http://www.nzsap.org/system/files/proceedings/2005/ab05026.pdf
Kennan, R. M., Dhungyel, O. P., Whittington, R. J., Egerton, J. R., and Rood, J. I. (2001). The type IV Fimbrial subunit gene (fimA) of Dichelobacter nodosus is essential for virulence, protease secretion, and natural competence. J. Bacteriol. 183, 4451–4458. doi: 10.1128/JB.183.15.4451-4458.2001
Kennan, R. M., Gilhuus, M., Frosth, S., Seemann, T., Dhungyel, O. P., Whittington, R. J., et al. (2014). Genomic evidence for a globally distributed, bimodal population in the ovine footrot pathogen Dichelobacter nodosus. MBio 5:e01821–e01814. doi: 10.1128/mBio.01821-14
Kennan, R. M., Wong, W., Dhungyel, O. P., Han, X., Wong, D., Parker, D., et al. (2010). The subtilisin-like protease AprV2 is required for virulence and uses a novel disulphide-tethered exosite to bind substrates. PLoS Pathog. 6:e1001210. doi: 10.1371/journal.ppat.1001210
Knappe-Poindecker, M., Jørgensen, H. J., Jensen, T. K., Tesfamichael, B., Ulvund, M. J., Vatn, S., et al. (2014). Experimental infection of sheep with ovine and bovine Dichelobacter nodosus isolates. Small Rumin. Res. 121, 411–417. doi: 10.1016/j.smallrumres.2014.07.021
Lee, S. E., Kim, J. M., Jeong, S. K., Jeon, J. E., Yoon, H. J., Jeong, M. K., et al. (2010). Protease-activated receptor-2 mediates the expression of inflammatory cytokines, antimicrobial peptides, and matrix metalloproteinases in keratinocytes in response to Propionibacterium acnes. Arch. Dermatol. Res. 302, 745–756. doi: 10.1007/s00403-010-1074-z
Levy, J. (2000). The effects of antibiotic use on gastrointestinal function. Am. J. Gastroenterol. 95, S8–S10. doi: 10.1016/S0002-9270(99)00808-4
Lippens, S., Denecker, G., Ovaere, P., Vandenabeele, P., and Declercq, W. (2005). Death penalty for keratinocytes: apoptosis versus cornification. Cell Death Differ. 12, 1497–1508. doi: 10.1038/sj.cdd.4401722
Maboni, G., Blanchard, A., Frosth, S., Stewart, C., Emes, R., and Tötemeyer, S. (2017). A distinct bacterial dysbiosis associated skin inflammation in ovine footrot. Sci. Rep. 7:45220. doi: 10.1038/srep45220
Maboni, G., Frosth, S., Aspán, A., and Tötemeyer, S. (2016). Ovine footrot: new insights into bacterial colonisation. Vet. Rec. 179:228. doi: 10.1136/vr.103610
MacNeil, S. (2007). Progress and opportunities for tissue-engineered skin. Nature 445, 874–880. doi: 10.1038/nature05664
Popov, L., Kovalski, J., Grandi, G., Bagnoli, F., and Amieva, M. R. (2014). Three-dimensional human skin models to understand Staphylococcus aureus skin colonization and infection. Front. Immunol. 5, 1–6. doi: 10.3389/fimmu.2014.00041
Price, K. J., Tsykin, A., Giles, K. M., Sladic, R. T., Epis, M. R., Ganss, R., et al. (2012). Matrigel basement membrane matrix influences expression of microRNAs in cancer cell lines. Biochem. Biophys. Res. Commun. 427, 343–348. doi: 10.1016/j.bbrc.2012.09.059
Rasmussen, M., Capion, N., Klitgaard, K., Rogdo, T., Fjeldaas, T., Boye, M., et al. (2012). Bovine digital dermatitis: possible pathogenic consortium consisting of Dichelobacter nodosus and multiple Treponema species. Vet. Microbiol. 160, 151–161. doi: 10.1016/j.vetmic.2012.05.018
Rather, M. A., Wani, S. A., Hussain, I., Bhat, M. A., Kabli, Z. A., and Magray, S. N. (2011). Determination of prevalence and economic impact of ovine footrot in central Kashmir India with isolation and molecular characterization of Dichelobacter nodosus. Anaerobe 17, 73–77. doi: 10.1016/j.anaerobe.2011.02.003
Ren, Y., Chan, H. M., Fan, J., Xie, Y., Chen, Y. X., Li, W., et al. (2006). Inhibition of tumor growth and metastasis in vitro and in vivo by targeting macrophage migration inhibitory factor in human neuroblastoma. Oncogene 25, 3501–3508. doi: 10.1038/sj.onc.1209395
Riffkin, M. C., Wang, L. F., Kortt, A. A., and Stewart, D. J. (1995). A single amino-acid change between the antigenically different extracellular serine proteases V2 and B2 from Dichelobacter nodosus. Gene 167, 279–283. doi: 10.1016/0378-1119(95)00664-8
Sanders, J. E., Mitchell, S. B., Wang, Y. N., and Wu, K. (2002). An explant model for the investigation of skin adaptation to mechanical stress. IEEE Trans. Biomed. Eng. 49, 1626–1631. doi: 10.1109/TBME.2002.805469
Sidgwick, G. P., McGeorge, D., and Bayat, A. (2016). Functional testing of topical skin formulations using an optimised ex vivo skin organ culture model. Arch. Dermatol. Res. 308, 297–308. doi: 10.1007/s00403-016-1645-8
Smijs, T. G. M., Bouwstra, J. A., Schuitmaker, H. J., Talebi, M., and Pavel, S. (2007). A novel ex vivo skin model to study the susceptibility of the dermatophyte Trichophyton rubrum to photodynamic treatment in different growth phases. J. Antimicrob. Chemother. 59, 433–440. doi: 10.1093/jac/dkl490
Stäuble, A., Steiner, A., Normand, L., Kuhnert, P., and Frey, J. (2014). Molecular genetic analysis of Dichelobacter nodosus proteases AprV2/B2, AprV5/ B5 and BprV/B in clinical material from European sheep flocks. Vet. Microb. 168, 177–184. doi: 10.1016/j.vetmic.2013.11.013
Steinstraesser, L., Sorkin, M., Niederbichler, A. D., Becerikli, M., Stupka, J., Daigeler, A., et al. (2010). A novel human skin chamber model to study wound infection ex vivo. Arch. Dermatol. Res. 302, 357–365. doi: 10.1007/s00403-009-1009-8
Tuschil, A., Lam, C., Haslberger, A., and Lindley, I. (1992). Interleukin-8 stimulates calcium transients and promotes epidermal cell proliferation. J. Invest. Dermatol. 99, 294–298. doi: 10.1111/1523-1747.ep12616634
Wang, Y., Gutierrez-Herrera, E., Ortega-Martinez, A., Anderson, R. R., and Franco, W. (2016). UV fluorescence excitation imaging of healing of wounds in skin: evaluation of wound closure in organ culture model. Lasers Surg. Med. 48, 678–685. doi: 10.1002/lsm.22523
Wassink, G. J., George, T. R., Kaler, J., and Green, L. E. (2010). Footrot and interdigital dermatitis in sheep: farmer satisfaction with current management, their ideal management and sources used to adopt new strategies. Prev. Vet. Med. 96, 65–73. doi: 10.1016/j.prevetmed.2010.06.002
Keywords: skin culture, ex vivo model, footrot, bacterial infection, Dichelobacter nodosus, ovine pro-inflammatory cytokines
Citation: Maboni G, Davenport R, Sessford K, Baiker K, Jensen TK, Blanchard AM, Wattegedera S, Entrican G and Tötemeyer S (2017) A Novel 3D Skin Explant Model to Study Anaerobic Bacterial Infection. Front. Cell. Infect. Microbiol. 7:404. doi: 10.3389/fcimb.2017.00404
Received: 22 June 2017; Accepted: 31 August 2017;
Published: 14 September 2017.
Edited by:
Georgios N. Belibasakis, Karolinska Institute (KI), SwedenReviewed by:
Kai Bao, University of Zurich, SwitzerlandKerstin Honer Zu Bentrup, Tulane University, United States
Julian I. Rood, Monash University, Australia
Copyright © 2017 Maboni, Davenport, Sessford, Baiker, Jensen, Blanchard, Wattegedera, Entrican and Tötemeyer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sabine Tötemeyer, c2FiaW5lLnRvdGVtZXllckBub3R0aW5naGFtLmFjLnVr