 Andrés González
Andrés González Javier Casado
Javier Casado Ángel Lanas
Ángel Lanas- 1Group of Translational Research in Digestive Diseases, Institute for Health Research Aragón (IIS Aragón), Zaragoza, Spain
- 2Department of Medicine, Psychiatry and Dermatology, University of Zaragoza, Zaragoza, Spain
- 3Biomedical Research Networking Center in Hepatic and Digestive Diseases (CIBERehd), Madrid, Spain
- 4Department of Biochemistry and Molecular & Cellular Biology, University of Zaragoza, Zaragoza, Spain
- 5Digestive Diseases Service, University Clinic Hospital Lozano Blesa, Zaragoza, Spain
Over half of the world’s population is estimated to be infected with Helicobacter pylori. Chronic infection with this microbial class I carcinogen is considered the most important risk factor for developing gastric cancer. The increasing antimicrobial resistance to first-line antibiotics mainly causes the failure of current eradication therapies, inducing refractory infections. The alarming increase in multidrug resistance in H. pylori isolates worldwide is already beginning to limit the efficacy of existing treatments. Consequently, the World Health Organization (WHO) has included H. pylori in its list of “priority pathogens” for which new antibiotics are urgently needed. Novel strategies must be followed to fight this antibiotic crisis, including properly exploiting the proven therapeutic potential of medicinal plants and plant-derived phytochemicals. In this mini-review, we overview the impressive properties of naturally occurring flavonoids as effective antimicrobial agents against H. pylori, which support the use of these plant-derived bioactive compounds as promising drug candidates for inclusion in novel and personalized combinatory therapies against H. pylori infection.
Introduction
Helicobacter pylori inhabits the gastric mucosa of almost 4.4 billion people worldwide (Hooi et al., 2017). Without effective eradication therapy, infection usually persists lifelong, causing gastric mucosal inflammation, which may gradually progress to peptic ulcer disease, gastric adenocarcinoma, and mucosa-associated lymphoid-tissue (MALT) lymphoma (Kusters et al., 2006; Yamaoka, 2010). Presently, the efficacy of one-week standard triple therapy containing clarithromycin (CLR) and either metronidazole (MTZ) or amoxicillin (AMX) combined with a proton-pump inhibitor (PPI) has dramatically dropped, showing eradication rates as low as 50% to 70% (Fallone et al., 2016). CLR-containing regimens are no longer suitable for unconditional empiric use because of commonly high levels of antimicrobial resistance and inadequate eradication rates, while the efficacy of the other alternative treatments varies greatly, which usually causes refractory infections. Given the rate at which clinically relevant pathogens, such as H. pylori, are acquiring multidrug resistance, the feared possibility that we cannot effectively treat these human bacterial infections is becoming a reality (Boyanova et al., 2016). In 2017, the World Health Organization (WHO) included H. pylori in its first list of antibiotic-resistant “priority pathogens”, a catalogue of 12 families of bacteria that presently pose the greatest threat to human health (Tacconelli et al., 2018). Nowadays, effective novel therapy against H. pylori is mandatory to increase eradication rates and minimize both antimicrobial resistance and side effects on normal microbiota.
Long before H. pylori infection was recognized as causing chronic gastritis and peptic ulcers in 1982 (Marshall and Warren, 1984), natural products have been used by physicians and healers to combat these illnesses based on empirical knowledge (Yesilada et al., 1997). Today, over 240 plant species have demonstrated anti-H. pylori activity (Salehi et al., 2018; Baker, 2020). With the pressing need for novel therapeutic options to face the current antibiotic crisis, the scientific community’s interest in traditional medicine and the use of natural products as sources of novel antibacterial drugs have been reinforced (Cheesman et al., 2017; Anand et al., 2019). In this mini-review, we overview the impressive findings obtained due to various studies that focused on the anti-H. pylori properties of flavonoids. We also discuss the promising roles of these natural products as potential drug candidates. Finally, we revise the current strategies to improve the bioavailability and efficacy of these phytochemicals.
Flavonoids: A Large Family of Naturally Occurring Bioactive Compounds
Flavonoids are polyphenolic, low-molecular-weight bioactive compounds ubiquitous in plants (Buer et al., 2010). The flavonoid family comprises over 9,000 species of molecules, which mostly share a chemical structure based on a fifteen-carbon (C6-C3-C6) skeleton comprising two benzene rings denoted as A and B, linked through a heterocyclic pyran ring referred to as ring C. The C6-C3-C6 skeleton is often hydroxylated in positions 2, 3, 5, 7, 3´, 4´, and 5´. Methyl ethers and acetyl esters of the alcohol groups are frequent, although a plethora of other derivative groups, including different alkyls, isoprenoids, and carboxylic groups, also contribute to the vast diversity of these compounds (Kumar and Pandey, 2013). Based on the oxidation state of the central pyran ring, its degree of hydroxylation, and the connection position of benzene ring B, flavonoids could be divided into seven major classes: flavones, flavonols, flavanones, flavanonols, flavanols (also known as flavan-3-ols), anthocyanidins, and isoflavones (Table 1).
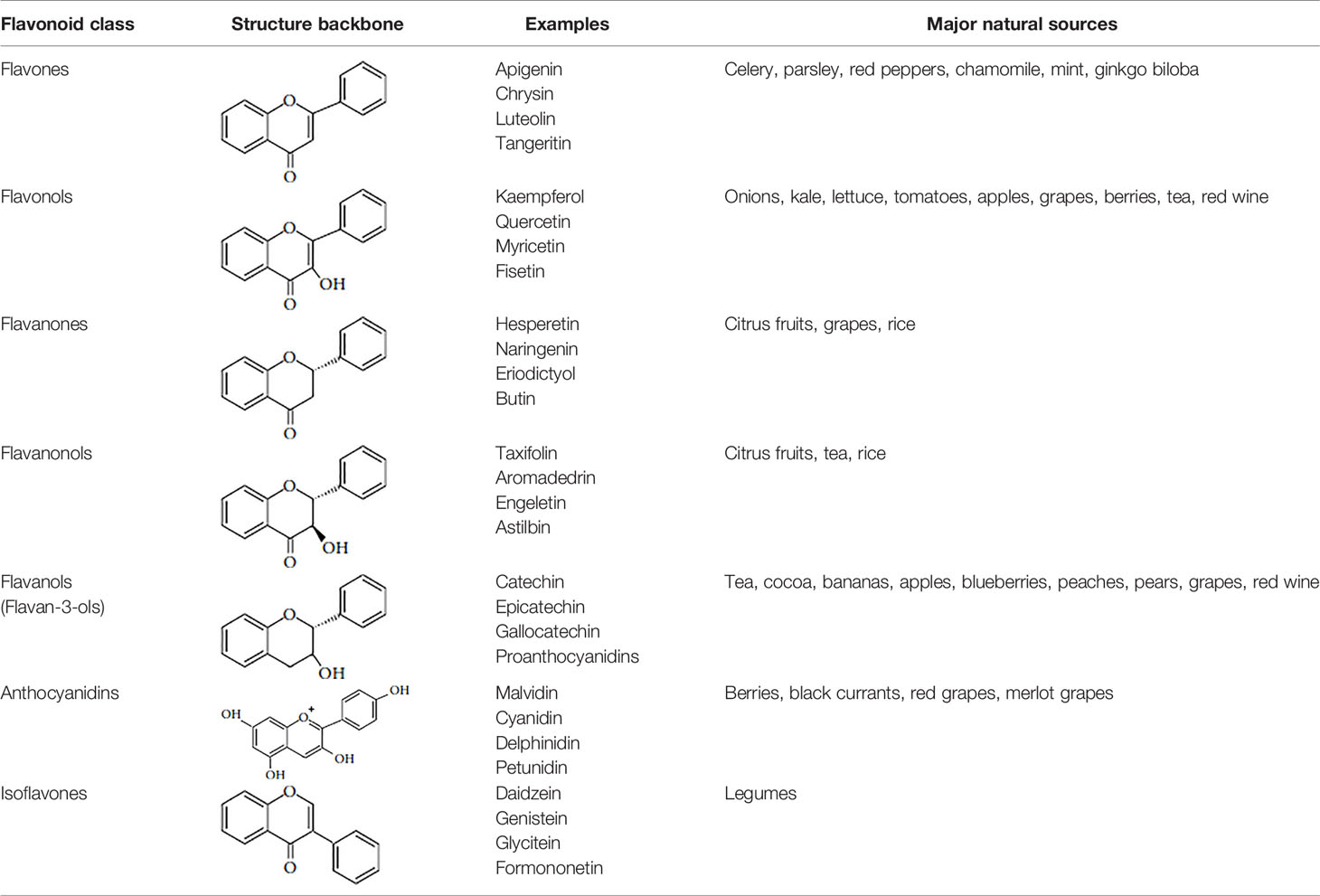
Table 1 Flavonoid classes and their major natural sources.
Flavonoids are synthesized as secondary metabolites by all plant organs. These natural polyphenols are critical in plants’ interaction with other organisms, such as microorganisms, animals, and other plants, but they also participate in responding to different abiotic stresses, including UV radiation, extreme temperatures, heavy metals, and droughts (Mierziak et al., 2014). A major role of flavonoids in plants is their function as a second line of defense against oxidative stress. Flavonoids can inhibit the generation of reactive oxygen species (ROS) by several mechanisms (Kumar and Pandey, 2013), but they also quench ROS once they are produced. Some environmental stresses, such as drought, salinity, extreme temperatures, and nutrient scarcity, may significantly reduce the activity of ROS-detoxifying enzymes in chloroplasts. As an adaptive response, plants upregulate the biosynthesis of ROS-scavenging flavonoids. These polyphenols will not only absorb the most energetic solar wavelengths (i.e., UV-B and UV-A) but will also scavenge free metal ions, peroxyl, superoxide, and peroxynitrite radicals, thereby avoiding lipid peroxidation and oxidative damage to other biomolecules (Kumar and Pandey, 2013; Del Valle et al., 2020).
Mechanisms of Antimicrobial Action—A Close Relationship Between Structure and Activity
As with other phytochemicals, the antimicrobial activity of flavonoids appears multifactorial while acting against different molecular targets in the pathogen instead of having one specific action site. However, the presence of certain structural features in the flavonoid molecule enhances its pharmacological effects, reinforcing one or another action mechanism, suggesting a relationship between the flavonoid structure and its antiviral and/or antimicrobial activities (Cushnie and Lamb, 2005; Kumar and Pandey, 2013; Wang et al., 2018; Farhadi et al., 2019; Górniak et al., 2019). Thus, while a greater abundance in hydroxyl groups increases the antioxidant effects of flavonoids due to a higher number of functional sites for scavenging free radicals and chelating metal ions, this high degree of hydroxylation diminishes simultaneously with flavonoid lipophilicity, thereby limiting the influx of these molecules across the pathogen cell membranes. Hence, lipophilic flavonoids, such as herperetin, naringenin, sophoraflavanone G, and catechins with gallate groups, could penetrate the lipid bilayer membrane up to the zone under phosphate groups and laterally diffuse into the bilayer plane, causing alterations in membrane fluidity and permeability (Tsuchiya and Iinuma, 2000; Tarahovsky et al., 2014). Other flavonoids such as quercetin cause a decrease in the proton-motive force impairing the production of adenosine triphosphate (ATP), while apigenin and morin induce destabilization of the membrane structure by the disordering and disorientation of membrane lipids (Górniak et al., 2019).
Also, hydrophilic flavonoids could interact at the membrane surface and/or in the cytosol with proteins involved in different essential functions, including adhesins, cell envelope transporters, transcriptional regulators, enzymes, and toxins, inactivating these biomolecules by forming flavonoid-protein complexes through hydrophobic interactions, hydrogen, and/or covalent bonds (Kumar and Pandey, 2013; Górniak et al., 2019). Notably, little difference in the molecular structure of two flavonoids could be responsible for different effectivities in their capability to inhibit the biological activity of the same protein target. For instance, both apigenin and quercetin inhibited the function of D-alanine:D-alanine ligase (Ddl) using the same inhibition mechanism as competing with the substrate ATP (Wu D. et al., 2008). Although these two flavonoids only differ in the two additional hydroxyl groups that quercetin possesses at positions 3 and 3´, this little difference induces a substantial increase in the affinity of quercetin by the active site of the enzyme, resulting in at least a three-fold increase in its inhibitory activity against Ddl regarding apigenin. However, quercetin exhibited lower antibacterial activity than apigenin, which could be a consequence of poorer transport across cell membranes due to lower lipophilicity (Wu D. et al., 2008).
Natural Flavonoids Against Helicobacter pylori Infection
Although many published studies have described the therapeutic potential of different plant extracts and other flavonoid-rich natural products (Ankolekar et al., 2011; Njume et al., 2011; Takeuchi et al., 2014; Wang, 2014; Boyanova et al., 2015; Salehi et al., 2018; Baker, 2020; Mendonca et al., 2020), we focused this mini-review on the advances in the knowledge of the antimicrobial activities of natural purified flavonoids against H. pylori (Table 2). Notably, several flavonoids have exhibited potent antimicrobial activities (MIC ≤ 8 µg/mL) against H. pylori. These in vitro antimicrobial potencies are comparable with those exhibited by some conventional antibiotics traditionally used in anti-H. pylori therapies, such as metronidazole, against sensitive strains (Loo et al., 1997). Although the anti-H. pylori activity exhibited by these flavonoids is probably multifactorial, an increasing number of studies have successfully identified specific molecular targets of these bioactive compounds in H. pylori, unravelling both antimicrobial and antivirulence mechanisms. Thus, several bactericidal flavonoids noticeably inhibited the essential function of HsrA (González et al., 2019), an OmpR-like orphan response regulator (Lee et al., 2006), which acts as a global homeostatic regulator synchronizing metabolic functions and virulence with the availability of nutrients and cell division, also mediating the response to oxidative stress (Olekhnovich et al., 2013; Olekhnovich et al., 2014; Pelliciari et al., 2017). Isothermal titration calorimetry studies indicated that chrysin, apigenin, kaempferol, and hesperetin bind to HsrA with dissociation constants in the micromolar range, showing a 1:1 stoichiometry. Molecular docking analyses suggest that interactions between these flavonoids and HsrA preferably occur by the C-terminal effector domain of the response regulator, thereby blocking its interaction with DNA (González et al., 2019). Notably, apigenin, kaempferol, and hesperetin also affected other recognized molecular targets in H. pylori, including enzymes (Wu D. et al., 2008; Zhang et al., 2008), secretion systems (Yeon et al., 2019), and cell membranes (Moon et al., 2013).

Table 2 Natural flavonoids with antimicrobial activities against H. pylori.
Many naturally occurring flavonoids exhibit anti-urease activity (Table 2). Molecular docking studies and structure–activity relationship analyses proved that 3-OH, 5-OH, and 3′,4′-dihydroxyl groups of quercetin generate hydrogen bonds with amino acid residues of H. pylori urease, which appear essential for the inhibitory activity exerted by this flavonoid. Removing or substituting any of these functional hydroxyl groups from the quercetin structure significantly decreases its urease inhibitory activity (Xiao et al., 2012). The critical impact of OH groups on the affinity and half maximal inhibitory concentration (IC50) of flavonoids against H. pylori enzymes has also been demonstrated in other studies (Wu D. et al., 2008; Yu et al., 2015). Flavonoids’ value as effective anti-H. pylori therapeutic drugs is not only supported by their proven bactericidal effect but also due to their antivirulence actions, which in many cases reduce damage to the host and alleviate associated diseases. Some H. pylori virulence factors, including cytotoxin-associated gene A (CagA) and vacuolating cytotoxin A (VacA), are critical in the inflammation process associated with infection with this pathogen. Cytotoxin CagA (Ansari and Yamaoka, 2020), encoded by the cag pathogenicity island, is translocated to host cells via the type IV secretion system (T4SS), a sophisticated transmembrane protein complex that directly injects the toxin into gastric epithelial cells (Backert et al., 2017). Once into the target cells, CagA activates NF-κB, a master regulator of immune and inflammatory responses that modulates the gene expression of pro-inflammatory cytokines, such as IL-8, TNF-α, and IL-1β (Lamb and Chen, 2013). Additionally, the cytotoxin VacA (Palframan et al., 2012), which is secreted from H. pylori via the type V secretion system (T5SS), acts on the host cell, inducing vacuolation and apoptosis, and also increases IL-8 production by activating the p38 MAPK via intracellular Ca2+ release, thereby activating the transcription factors, ATF-2, CREB, and NF-κB (Hisatsune et al., 2008). Several flavonoids, including apigenin (Wang and Huang, 2013), kaempferol (Yeon et al., 2019), quercetin (Gonzalez-Segovia et al., 2008; Zhang et al., 2017), nobiletin (Ouyang et al., 2020), baicalin, baicalein (Chen et al., 2018), galangin (Skiba et al., 2016), and genistein (Siriviriyakul et al., 2020) have shown protection against gastric inflammation associated with H. pylori infection by reducing pro-inflammatory cytokine expression. Thus, kaempferol decreased the mRNA levels of IL-8, TNF-α, and IL-1β in gastric adenocarcinoma cells infected with H. pylori by inhibiting vacA expression and suppressing CagA and VacA translocation to target cells by inhibiting the expression of several T4SS and T5SS components (Yeon et al., 2019). Quercetin significantly reduced in vivo gastric inflammation in H. pylori-infected mice by reducing IL-8 secretion and downregulating the p38 MAPK signaling pathway (Zhang et al., 2017). Apigenin decreased the levels of IL-8 by inhibiting the activation of NF-κB (Wang and Huang, 2013). Adding to their anti-inflammatory actions, several flavonoids have been demonstrated to protect against vacuolation, apoptosis, and lipid peroxidation induced by H. pylori in gastric mucosa (Shin et al., 2005; Gonzalez-Segovia et al., 2008; Zhang et al., 2017).
A further benefit of flavonoids as antimicrobials against H. pylori infections lies in their capacity for synergism combined with anti-H. pylori first-line antibiotics, including CLR, MTZ, or AMX (Table 2). Notably, chrysin induced an eight-fold decrease in the MIC value of CLR (FIC = 0.125), and caused a 16-fold decrease in the MIC value of MTZ (FIC = 0.0625) (González et al., 2019). Likewise, hesperetin led to a 4-fold increase in the inhibitory activity of MTZ and a two-fold increase in the CLR anti-H. pylori in vitro activity (González et al., 2019). Although the molecular mechanisms by which flavonoids enhance the antimicrobial activities of conventional antibiotics remain poorly understood, some experimental evidence unravels putative synergistic interactions. Thus, the increased inhibitory activities of AMX and TET in multidrug-resistant strains of H. pylori after baicalin action appeared to be associated with a decrease in the expression of the efflux pump gene hefA (Huang et al., 2015). hefA encodes a TolC-like outer membrane channel tunnel protein that interacts with different inner-membrane translocases to form efflux systems involved in drug resistance (Liu et al., 2008). Myricetin strongly inhibited the expression of genes involved in the morphological transition of H. pylori from spiral to coccoid forms, thereby avoiding the increase in antimicrobial resistance associated with cell shape transformation, which has been observed in this pathogen. Consequently, myricetin induced a 4–16-fold reduction in the MIC values of CLR, MTZ, LVX, TET, and AMX (Krzyzek et al., 2021). In addition, several authors suggest that the proven damage triggered by certain flavonoids in the cytoplasmic membrane and/or cell wall could enhance the susceptibility of bacterial pathogens to the action of antibiotics (Amin et al., 2015; Sanhueza et al., 2017).
Challenges and Perspectives in Using Flavonoids as Antimicrobials
Low solubility, poor permeability, relative chemical instability, rapid release, and susceptibility to environmental influences, but mainly low bioavailability, contribute to the fact that the in vivo exposure levels of flavonoids are usually inconsistent and much below the effective concentrations observed in the in vitro studies. Most flavonoids undergo sulfation, methylation, and glucuronidation in the small intestine and liver due to phase 2 metabolism reactions, resulting in more hydrophilic conjugated metabolites, which show reduced bioactivity compared to parent compounds (D’Archivio et al., 2010; Thilakarathna and Rupasinghe, 2013; Hu et al., 2017; Yang et al., 2020).
In recent years, many advanced nanoparticles have been developed not only to improve polyphenol bioavailability but also to control/target their release. Liposomes, phospholipid complexes, niosomes, protein-based nanoparticles, micelles, emulsions, and metal nanoparticles have been demonstrated to significantly increase bioavailability and improve the pharmacokinetics of polyphenols, becoming promising options for flavonoid delivery systems (Chen et al., 2020; Riaz et al., 2020; Yang et al., 2020). Another approach to improving the bioavailability of flavonoids is the rational modification of their molecular structures to bypass phase 2 metabolism during absorption. Some chemical modifications of bioactive molecules generate inactive forms, known as prodrugs, which can usually be transformed into their active forms by a single-step reaction after ingestion. Designing different synthetic prodrugs of polyphenols by capping phenolic hydroxyls with different protecting groups could increase bioavailability and reinforce the therapeutic properties of these bioactive compounds (Biasutto and Zoratti, 2014; Biasutto et al., 2017). Finally, phyto-phospholipid complexes have emerged as a promising strategy for enhancing the bioavailability of bioactive polyphenols. Phytosomes are obtained by complexing phytochemicals with phospholipids at defined molar ratios and under certain conditions. The resulting complexes are more readily absorbed and exhibit higher bioavailability than free bioactive compounds (Lu et al., 2019; Riva et al., 2019).
Discussion
Antibiotic resistance is among the greatest threats to global health in this century. An impressive accumulation of antibiotic resistance genes by clinically relevant bacterial pathogens, jointly with the present slowdown in developing new antibiotics, is inducing untreatable infections worldwide. H. pylori is a carcinogenic bacterium that infects over half of the global population, causing chronic progressive gastric inflammation and various diseases, including gastric and duodenal ulcers and gastric cancer. This malignancy constitutes the fifth most common cancer and the third leading cause of cancer-related mortality globally, representing 9% of all cancer-related deaths worldwide. Although the eradication of H. pylori infection has been proven to significantly reduce gastric cancer incidence, the efficacy of current eradication therapies has dramatically decreased, mainly because of an increasing development of antibiotic resistance. In this context, different R&D strategies must be urgently considered to fast-track novel, effective therapeutic options against H. pylori infection. Hence, validating novel therapeutic targets, repurposing the existing drugs, using synergistic combinatory therapies, and properly exploiting the proven therapeutic potential of medicinal plants and other natural products could accelerate the delivery of new antimicrobials and the design of novel and personalized treatments against H. pylori refractory infections.
Since most of the currently prescribed antibiotics have been obtained from microbial sources or derivatives thereof, using bioactive phytochemicals to develop new antibiotics could overcome the circulating resistome and slowdown the emergence and dissemination of novel antibiotic resistance mechanisms. Among these naturally occurring substances, flavonoids stand out for their multiple and remarkable beneficial effects on human health. Several flavonoids have revealed potent antimicrobial activities against H. pylori, in some cases, at the level achieved by first-line antibiotics, such as metronidazole. In addition, some of these polyphenolic molecules exhibited synergistic effects combined with conventional antibiotics, thereby reverting antibiotic resistant phenotypes. Furthermore, the antivirulence actions of these compounds against H. pylori contribute to protecting against gastric inflammation, vacuolation, apoptosis, and lipid peroxidation, reducing the damage exerted by the pathogen to the host cells, and decreasing the progression of associated diseases. Although low bioavailability contributes to decreasing the in vivo effectivity of natural flavonoids, the current development of novel delivery systems, such as prodrugs, phytosomes, and several nanotechnology approaches, enables the inclusion of flavonoids as novel therapeutic tools against H. pylori infection.
Author Contributions
AG, JC, and ÁL wrote the review. All authors contributed to the article and approved the submitted version.
Funding
This work has been supported by the Government of Aragon, Spain (B25_17R) and University of Zaragoza (2018/0420).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Amin, M. U., Khurram, M., Khattak, B., Khan, J. (2015). Antibiotic Additive and Synergistic Action of Rutin, Morin and Quercetin Against Methicillin Resistant Staphylococcus aureus. BMC Complement Altern. Med. 15, 59. doi: 10.1186/s12906-015-0580-0
Anand, U., Jacobo-Herrera, N., Altemimi, A., Lakhssassi, N. (2019). A Comprehensive Review on Medicinal Plants as Antimicrobial Therapeutics: Potential Avenues of Biocompatible Drug Discovery. Metabolites 9 (11), 258. doi: 10.3390/metabo9110258
Ankolekar, C., Johnson, D., Pinto Mda, S., Johnson, K., Labbe, R., Shetty, K. (2011). Inhibitory Potential of Tea Polyphenolics and Influence of Extraction Time Against Helicobacter pylori and Lack of Inhibition of Beneficial Lactic Acid Bacteria. J. Med. Food 14 (11), 1321–1329. doi: 10.1089/jmf.2010.0237
Ansari, S., Yamaoka, Y. (2020). Helicobacter pylori Virulence Factor Cytotoxin-Associated Gene A (CagA)-Mediated Gastric Pathogenicity. Int. J. Mol. Sci. 21 (19), 7430. doi: 10.3390/ijms21197430
Backert, S., Haas, R., Gerhard, M., Naumann, M. (2017). The Helicobacter pylori Type IV Secretion System Encoded by the Cag Pathogenicity Island: Architecture, Function, and Signaling. Curr. Top. Microbiol. Immunol. 413, 187–220. doi: 10.1007/978-3-319-75241-9_8
Baker, D. A. (2020). Plants Against Helicobacter pylori to Combat Resistance: An Ethnopharmacological Review. Biotechnol. Rep. (Amst.) 26, e00470. doi: 10.1016/j.btre.2020.e00470
Biasutto, L., Mattarei, A., Azzolini, M., La Spina, M., Sassi, N., Romio, M., et al. (2017). Resveratrol Derivatives as a Pharmacological Tool. Ann. N. Y. Acad. Sci. 1403 (1), 27–37. doi: 10.1111/nyas.13401
Biasutto, L., Zoratti, M. (2014). Prodrugs of Quercetin and Resveratrol: A Strategy Under Development. Curr. Drug Metab. 15 (1), 77–95. doi: 10.2174/1389200214666131211160005
Bisignano, C., Filocamo, A., La Camera, E., Zummo, S., Fera, M. T., Mandalari, G. (2013). Antibacterial Activities of Almond Skins on cagA-Positive and-Negative Clinical Isolates of Helicobacter pylori. BMC Microbiol. 13, 103. doi: 10.1186/1471-2180-13-103
Boyanova, L., Evstatiev, I., Yordanov, D., Markovska, R., Mitov, I. (2016). Three Unsuccessful Treatments of Helicobacter pylori Infection by a Highly Virulent Strain With Quadruple Antibiotic Resistance. Folia Microbiol. (Praha) 61 (4), 307–310. doi: 10.1007/s12223-015-0439-2
Boyanova, L., Ilieva, J., Gergova, G., Vladimirov, B., Nikolov, R., Mitov, I. (2015). Honey and Green/Black Tea Consumption may Reduce the Risk of Helicobacter pylori Infection. Diagn. Microbiol. Infect. Dis. 82 (1), 85–86. doi: 10.1016/j.diagmicrobio.2015.03.001
Brown, J. C., Jiang, X. (2013). Activities of Muscadine Grape Skin and Polyphenolic Constituents Against Helicobacter pylori. J. Appl. Microbiol. 114 (4), 982–991. doi: 10.1111/jam.12129
Brown, J. C., Wang, J., Kasman, L., Jiang, X., Haley-Zitlin, V. (2011). Activities of Muscadine Grape Skin and Quercetin Against Helicobacter pylori Infection in Mice. J. Appl. Microbiol. 110 (1), 139–146. doi: 10.1111/j.1365-2672.2010.04870.x
Buer, C. S., Imin, N., Djordjevic, M. A. (2010). Flavonoids: New Roles for Old Molecules. J. Integr. Plant Biol. 52 (1), 98–111. doi: 10.1111/j.1744-7909.2010.00905.x
Cheesman, M. J., Ilanko, A., Blonk, B., Cock, I. E. (2017). Developing New Antimicrobial Therapies: Are Synergistic Combinations of Plant Extracts/Compounds With Conventional Antibiotics the Solution? Pharmacogn. Rev. 11 (22), 57–72. doi: 10.4103/phrev.phrev_21_17
Chen, K. T. J., Anantha, M., Leung, A. W. Y., Kulkarni, J. A., Militao, G. G. C., Wehbe, M., et al. (2020). Characterization of a Liposomal Copper(II)-Quercetin Formulation Suitable for Parenteral Use. Drug Delivery Transl. Res. 10 (1), 202–215. doi: 10.1007/s13346-019-00674-7
Chen, M. E., Su, C. H., Yang, J. S., Lu, C. C., Hou, Y. C., Wu, J. B., et al. (2018). Baicalin, Baicalein, and Lactobacillus Rhamnosus JB3 Alleviated Helicobacter pylori Infections In Vitro and In Vivo. J. Food Sci. 83 (12), 3118–3125. doi: 10.1111/1750-3841.14372
Chledzik, S., Strawa, J., Matuszek, K., Nazaruk, J. (2018). Pharmacological Effects of Scutellarin, an Active Component of Genus Scutellaria and Erigeron: A Systematic Review. Am. J. Chin. Med. 46 (2), 319–337. doi: 10.1142/S0192415X18500167
Chung, J. G., Hsia, T. C., Kuo, H. M., Li, Y. C., Lee, Y. M., Lin, S. S., et al. (2001). Inhibitory Actions of Luteolin on the Growth and Arylamine N-Acetyltransferase Activity in Strains of Helicobacter pylori From Ulcer Patients. Toxicol. In Vitro 15 (3), 191–198. doi: 10.1016/S0887-2333(01)00015-7
Cushnie, T. P., Lamb, A. J. (2005). Antimicrobial Activity of Flavonoids. Int. J. Antimicrob. Agents 26 (5), 343–356. doi: 10.1016/j.ijantimicag.2005.09.002
D’Archivio, M., Filesi, C., Vari, R., Scazzocchio, B., Masella, R. (2010). Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 11 (4), 1321–1342. doi: 10.3390/ijms11041321
Del Valle, J. C., Buide, M. L., Whittall, J. B., Valladares, F., Narbona, E. (2020). UV Radiation Increases Phenolic Compound Protection But Decreases Reproduction in Silene littorea. PloS One 15 (6), e0231611. doi: 10.1371/journal.pone.0231611
Fallone, C. A., Chiba, N., van Zanten, S. V., Fischbach, L., Gisbert, J. P., Hunt, R. H., et al. (2016). The Toronto Consensus for the Treatment of Helicobacter pylori Infection in Adults. Gastroenterology 151 (1), 51–69.e14. doi: 10.1053/j.gastro.2016.04.006
Farhadi, F., Khameneh, B., Iranshahi, M., Iranshahy, M. (2019). Antibacterial Activity of Flavonoids and Their Structure-Activity Relationship: An Update Review. Phytother. Res. 33 (1), 13–40. doi: 10.1002/ptr.6208
Fong, P., Hao, C. H., Io, C. C., Sin, P. I., Meng, L. R. (2019). In Silico and In Vitro Anti-Helicobacter pylori Effects of Combinations of Phytochemicals and Antibiotics. Molecules 24 (19), 3608. doi: 10.3390/molecules24193608
Gomez-Chang, E., Uribe-Estanislao, G. V., Martinez-Martinez, M., Galvez-Mariscal, A., Romero, I. (2018). Anti-Helicobacter pylori Potential of Three Edible Plants Known as Quelites in Mexico. J. Med. Food 21 (11), 1150–1157. doi: 10.1089/jmf.2017.0137
González, A., Salillas, S., Velázquez-Campoy, A., Espinosa Angarica, V., Fillat, M. F., Sancho, J., et al. (2019). Identifying Potential Novel Drugs Against Helicobacter pylori by Targeting the Essential Response Regulator HsrA. Sci. Rep. 9 (1), 11294. doi: 10.1038/s41598-019-47746-9
Gonzalez-Segovia, R., Quintanar, J. L., Salinas, E., Ceballos-Salazar, R., Aviles-Jimenez, F., Torres-Lopez, J. (2008). Effect of the Flavonoid Quercetin on Inflammation and Lipid Peroxidation Induced by Helicobacter pylori in Gastric Mucosa of Guinea Pig. J. Gastroenterol. 43 (6), 441–447. doi: 10.1007/s00535-008-2184-7
Górniak, I., Bartoszewski, R., Króliczewski, J. (2019). Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 18 (1), 241–272. doi: 10.1007/s11101-018-9591-z
Haghi, A., Azimi, H., Rahimi, R. (2017). A Comprehensive Review on Pharmacotherapeutics of Three Phytochemicals, Curcumin, Quercetin, and Allicin, in the Treatment of Gastric Cancer. J. Gastrointest. Cancer 48 (4), 314–320. doi: 10.1007/s12029-017-9997-7
Hisatsune, J., Nakayama, M., Isomoto, H., Kurazono, H., Mukaida, N., Mukhopadhyay, A. K., et al. (2008). Molecular Characterization of Helicobacter pylori VacA Induction of IL-8 in U937 Cells Reveals a Prominent Role for p38MAPK in Activating Transcription Factor-2, cAMP Response Element Binding Protein, and NF-kappaB Activation. J. Immunol. 180 (7), 5017–5027. doi: 10.4049/jimmunol.180.7.5017
Hooi, J. K. Y., Lai, W. Y., Ng, W. K., Suen, M. M. Y., Underwood, F. E., Tanyingoh, D., et al. (2017). Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 153 (2), 420–429. doi: 10.1053/j.gastro.2017.04.022
Hribova, P., Khazneh, E., Zemlicka, M., Svajdlenka, E., Ghoneim, M. M., Elokely, K. M., et al. (2014). Antiurease Activity of Plants Growing in the Czech Republic. Nat. Prod. Res. 28 (12), 868–873. doi: 10.1080/14786419.2014.888553
Huang, Y. Q., Huang, G. R., Wu, M. H., Tang, H. Y., Huang, Z. S., Zhou, X. H., et al. (2015). Inhibitory Effects of Emodin, Baicalin, Schizandrin and Berberine on hefA Gene: Treatment of Helicobacter pylori-Induced Multidrug Resistance. World J. Gastroenterol. 21 (14), 4225–4231. doi: 10.3748/wjg.v21.i14.4225
Hu, M., Wu, B., Liu, Z. (2017). Bioavailability of Polyphenols and Flavonoids in the Era of Precision Medicine. Mol. Pharm. 14 (9), 2861–2863. doi: 10.1021/acs.molpharmaceut.7b00545
Isobe, T., Doe, M., Morimoto, Y., Nagata, K., Ohsaki, A. (2006). The Anti-Helicobacter pylori Flavones in a Brazilian Plant, Hyptis Fasciculata, and the Activity of Methoxyflavones. Biol. Pharm. Bull. 29 (5), 1039–1041. doi: 10.1248/bpb.29.1039
Kataria, R., Khatkar, A. (2019a). In-Silico Designing, ADMET Analysis, Synthesis and Biological Evaluation of Novel Derivatives of Diosmin Against Urease Protein and Helicobacter pylori Bacterium. Curr. Top. Med. Chem. 19 (29), 2658–2675. doi: 10.2174/1568026619666191114123452
Kataria, R., Khatkar, A. (2019b). Molecular Docking, Synthesis, Kinetics Study, Structure-Activity Relationship and ADMET Analysis of Morin Analogous as Helicobacter pylori Urease Inhibitors. BMC Chem. 13 (1), 45. doi: 10.1186/s13065-019-0562-2
Kim, S. H., Lee, M. H., Park, M., Woo, H. J., Kim, Y. S., Tharmalingam, N., et al. (2018). Regulatory Effects of Black Rice Extract on Helicobacter pylori Infection-Induced Apoptosis. Mol. Nutr. Food Res. 62 (3), 1700586. doi: 10.1002/mnfr.201700586
Kim, S. H., Park, M., Woo, H., Tharmalingam, N., Lee, G., Rhee, K. J., et al. (2012). Inhibitory Effects of Anthocyanins on Secretion of Helicobacter pylori CagA and VacA Toxins. Int. J. Med. Sci. 9 (10), 838–842. doi: 10.7150/ijms.5094
Kim, S. H., Woo, H., Park, M., Rhee, K. J., Moon, C., Lee, D., et al. (2014). Cyanidin 3-O-Glucoside Reduces Helicobacter pylori VacA-Induced Cell Death of Gastric KATO III Cells Through Inhibition of the SecA Pathway. Int. J. Med. Sci. 11 (7), 742–747. doi: 10.7150/ijms.7167
Krzyzek, P., Migdal, P., Paluch, E., Karwanska, M., Wieliczko, A., Gosciniak, G. (2021). Myricetin as an Antivirulence Compound Interfering With a Morphological Transformation Into Coccoid Forms and Potentiating Activity of Antibiotics Against Helicobacter pylori. Int. J. Mol. Sci. 22 (5), 2695. doi: 10.3390/ijms22052695
Kumar, S., Pandey, A. K. (2013). Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013:162750. doi: 10.1155/2013/162750
Kuo, C. H., Weng, B. C., Wu, C. C., Yang, S. F., Wu, D. C., Wang, Y. C. (2014). Apigenin has Anti-Atrophic Gastritis and Anti-Gastric Cancer Progression Effects in Helicobacter pylori-Infected Mongolian Gerbils. J. Ethnopharmacol. 151 (3), 1031–1039. doi: 10.1016/j.jep.2013.11.040
Kusters, J. G., van Vliet, A. H., Kuipers, E. J. (2006). Pathogenesis of Helicobacter pylori Infection. Clin. Microbiol. Rev. 19 (3), 449–490. doi: 10.1128/CMR.00054-05
Lamb, A., Chen, L. F. (2013). Role of the Helicobacter pylori-Induced Inflammatory Response in the Development of Gastric Cancer. J. Cell Biochem. 114 (3), 491–497. doi: 10.1002/jcb.24389
Lee, H. M., Hong, E., Jeon, B. Y., Kim, D. U., Byun, J. S., Lee, W., et al. (2006). Crystallization and Preliminary X-Ray Crystallographic Study of HP1043, a Helicobacter pylori Orphan Response Regulator. Biochim. Biophys. Acta 1764 (5), 989–991. doi: 10.1016/j.bbapap.2005.10.024
Lee, Y. S., Huh, J. Y., Nam, S. H., Moon, S. K., Lee, S. B. (2012). Enzymatic Bioconversion of Citrus Hesperidin by Aspergillus Sojae Naringinase: Enhanced Solubility of Hesperetin-7-O-Glucoside With In Vitro Inhibition of Human Intestinal Maltase, HMG-CoA Reductase, and Growth of Helicobacter pylori. Food Chem. 135 (4), 2253–2259. doi: 10.1016/j.foodchem.2012.07.007
Liu, Z. Q., Zheng, P. Y., Yang, P. C. (2008). Efflux Pump Gene hefA of Helicobacter pylori Plays an Important Role in Multidrug Resistance. World J. Gastroenterol. 14 (33), 5217–5222. doi: 10.3748/wjg.14.5217
Loo, V. G., Fallone, C. A., De Souza, E., Lavallee, J., Barkun, A. N. (1997). In-Vitro Susceptibility of Helicobacter pylori to Ampicillin, Clarithromycin, Metronidazole and Omeprazole. J. Antimicrob. Chemother. 40 (6), 881–883. doi: 10.1093/jac/40.6.881
Lu, M., Qiu, Q., Luo, X., Liu, X., Sun, J., Wang, C., et al. (2019). Phyto-Phospholipid Complexes (Phytosomes): A Novel Strategy to Improve the Bioavailability of Active Constituents. Asian J. Pharm. Sci. 14 (3), 265–274. doi: 10.1016/j.ajps.2018.05.011
Macomber, L., Minkara, M. S., Hausinger, R. P., Merz, K. M., Jr. (2015). Reduction of Urease Activity by Interaction With the Flap Covering the Active Site. J. Chem. Inf. Model 55 (2), 354–361. doi: 10.1021/ci500562t
Marshall, B. J., Warren, J. R. (1984). Unidentified Curved Bacilli in the Stomach of Patients With Gastritis and Peptic Ulceration. Lancet 1 (8390), 1311–1315. doi: 10.1016/s0140-6736(84)91816-6
Martini, S., D’Addario, C., Colacevich, A., Focardi, S., Borghini, F., Santucci, A., et al. (2009). Antimicrobial Activity Against Helicobacter pylori Strains and Antioxidant Properties of Blackberry Leaves (Rubus ulmifolius) and Isolated Compounds. Int. J. Antimicrob. Agents 34 (1), 50–59. doi: 10.1016/j.ijantimicag.2009.01.010
Matsumoto, T., Takahashi, T., Yamada, H. (2008). A Novel Approach for Screening of New Anti-Helicobacter pylori Substances. Biol. Pharm. Bull. 31 (1), 143–145. doi: 10.1248/bpb.31.143
Mendonca, M. A. A., Ribeiro, A. R. S., Lima, A. K., Bezerra, G. B., Pinheiro, M. S., Albuquerque-Junior, R. L. C., et al. (2020). Red Propolis and its Dyslipidemic Regulator Formononetin: Evaluation of Antioxidant Activity and Gastroprotective Effects in Rat Model of Gastric Ulcer. Nutrients 12 (10), 2951. doi: 10.3390/nu12102951
Mierziak, J., Kostyn, K., Kulma, A. (2014). Flavonoids as Important Molecules of Plant Interactions With the Environment. Molecules 19 (10), 16240–16265. doi: 10.3390/molecules191016240
Moon, S. H., Lee, J. H., Kim, K. T., Park, Y. S., Nah, S. Y., Ahn, D. U., et al. (2013). Antimicrobial Effect of 7-O-Butylnaringenin, a Novel Flavonoid, and Various Natural Flavonoids Against Helicobacter pylori Strains. Int. J. Environ. Res. Public Health 10 (11), 5459–5469. doi: 10.3390/ijerph10115459
Nakagawa, H., Takaishi, Y., Tanaka, N., Tsuchiya, K., Shibata, H., Higuti, T. (2006). Chemical Constituents From the Peels of Citrus sudachi. J. Nat. Prod. 69 (8), 1177–1179. doi: 10.1021/np060217s
Ndemangou, B., Sielinou, V. T., Vardamides, J. C., Ali, M. S., Lateef, M., Iqbal, L., et al. (2013). Urease Inhibitory Isoflavonoids From Different Parts of Calopogonium Mucunoides (Fabaceae). J. Enzyme Inhib. Med. Chem. 28 (6), 1156–1161. doi: 10.3109/14756366.2012.719025
Njume, C., Jide, A. A., Ndip, R. N. (2011). Aqueous and Organic Solvent-Extracts of Selected South African Medicinal Plants Possess Antimicrobial Activity Against Drug-Resistant Strains of Helicobacter pylori: Inhibitory and Bactericidal Potential. Int. J. Mol. Sci. 12 (9), 5652–5665. doi: 10.3390/ijms12095652
Olekhnovich, I. N., Vitko, S., Chertihin, O., Hontecillas, R., Viladomiu, M., Bassaganya-Riera, J., et al. (2013). Mutations to Essential Orphan Response Regulator HP1043 of Helicobacter pylori Result in Growth-Stage Regulatory Defects. Infect. Immun. 81 (5), 1439–1449. doi: 10.1128/IAI.01193-12
Olekhnovich, I. N., Vitko, S., Valliere, M., Hoffman, P. S. (2014). Response to Metronidazole and Oxidative Stress Is Mediated Through Homeostatic Regulator HsrA (HP1043) in Helicobacter pylori. J. Bacteriol. 196 (4), 729–739. doi: 10.1128/JB.01047-13
Ouyang, Y., Li, L., Ling, P. (2020). Nobiletin Inhibits Helicobacterium Pylori Infection-Induced Gastric Carcinogenic Signaling by Blocking Inflammation, Apoptosis, and Mitogen-Activated Protein Kinase Events in Gastric Epithelial-1 Cells. J. Environ. Pathol. Toxicol. Oncol. 39 (1), 77–88. doi: 10.1615/JEnvironPatholToxicolOncol.2020031272
Palframan, S. L., Kwok, T., Gabriel, K. (2012). Vacuolating Cytotoxin A (VacA), a Key Toxin for Helicobacter pylori Pathogenesis. Front. Cell Infect. Microbiol. 2, 92. doi: 10.3389/fcimb.2012.00092
Pastene, E., Parada, V., Avello, M., Ruiz, A., Garcia, A. (2014). Catechin-Based Procyanidins From Peumus boldus Mol. Aqueous Extract Inhibit Helicobacter pylori Urease and Adherence to Adenocarcinoma Gastric Cells. Phytother. Res. 28 (11), 1637–1645. doi: 10.1002/ptr.5176
Pelliciari, S., Pinatel, E., Vannini, A., Peano, C., Puccio, S., De Bellis, G., et al. (2017). Insight Into the Essential Role of the Helicobacter pylori HP1043 Orphan Response Regulator: Genome-Wide Identification and Characterization of the DNA-Binding Sites. Sci. Rep. 7, 41063. doi: 10.1038/srep41063
Rajesh, K. P., Manjunatha, H., Bharath, B. R. (2013). Simulated Screening of Flavonoids as Probable Anti-Helicobacter pylori Drug. Med. Chem. Res. 22, 4537–4546. doi: 10.1007/s00044-012-0426-y
Riaz, S., Fatima Rana, N., Hussain, I., Tanweer, T., Nawaz, A., Menaa, F., et al. (2020). Effect of Flavonoid-Coated Gold Nanoparticles on Bacterial Colonization in Mice Organs. Nanomaterials (Basel) 10 (9), 1769. doi: 10.3390/nano10091769
Riva, A., Ronchi, M., Petrangolini, G., Bosisio, S., Allegrini, P. (2019). Improved Oral Absorption of Quercetin From Quercetin Phytosome™, a New Delivery System Based on Food Grade Lecithin. Eur. J. Drug Metab. Pharmacokinet. 44 (2), 169–177. doi: 10.1007/s13318-018-0517-3
Salehi, B., Sharopov, F., Martorell, M., Rajkovic, J., Ademiluyi, A. O., Sharifi-Rad, M., et al. (2018). Phytochemicals in Helicobacter pylori Infections: What Are We Doing Now? Int. J. Mol. Sci. 19 (8), 2361. doi: 10.3390/ijms19082361
Sanhueza, L., Melo, R., Montero, R., Maisey, K., Mendoza, L., Wilkens, M. (2017). Synergistic Interactions Between Phenolic Compounds Identified in Grape Pomace Extract With Antibiotics of Different Classes Against Staphylococcus Aureus and Escherichia Coli. PloS One 12 (2), e0172273. doi: 10.1371/journal.pone.0172273
Shin, J. E., Kim, J. M., Bae, E. A., Hyun, Y. J., Kim, D. H. (2005). In Vitro Inhibitory Effect of Flavonoids on Growth, Infection and Vacuolation of Helicobacter pylori. Planta Med. 71 (3), 197–201. doi: 10.1055/s-2005-837816
Silvan, J. M., Gutierrez-Docio, A., Moreno-Fernandez, S., Alarcon-Cavero, T., Prodanov, M., Martinez-Rodriguez, A. J. (2020). Procyanidin-Rich Extract From Grape Seeds as a Putative Tool Against Helicobacter pylori. Foods 9 (10), 1370. doi: 10.3390/foods9101370
Siriviriyakul, P., Werawatganon, D., Phetnoo, N., Somanawat, K., Chatsuwan, T., Klaikeaw, N., et al. (2020). Genistein Attenuated Gastric Inflammation and Apoptosis in Helicobacter pylori-Induced Gastropathy in Rats. BMC Gastroenterol. 20 (1), 410. doi: 10.1186/s12876-020-01555-x
Skiba, M. A., Szendzielorz, K., Mazur, B., Krol, W. (2016). The Inhibitory Effect of Flavonoids on Interleukin-8 Release by Human Gastric Adenocarcinoma (AGS) Cells Infected With cag PAI (+) Helicobacter pylori. Cent. Eur. J. Immunol. 41 (3), 229–235. doi: 10.5114/ceji.2016.63119
Stompor, M. (2020). A Review on Sources and Pharmacological Aspects of Sakuranetin. Nutrients 12 (2), 513. doi: 10.3390/nu12020513
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 18 (3), 318–327. doi: 10.1016/S1473-3099(17)30753-3
Takeuchi, H., Trang, V. T., Morimoto, N., Nishida, Y., Matsumura, Y., Sugiura, T. (2014). Natural Products and Food Components With Anti-Helicobacter pylori Activities. World J. Gastroenterol. 20 (27), 8971–8978. doi: 10.3748/wjg.v20.i27.8971
Tarahovsky, Y. S., Kim, Y. A., Yagolnik, E. A., Muzafarov, E. N. (2014). Flavonoid-Membrane Interactions: Involvement of Flavonoid-Metal Complexes in Raft Signaling. Biochim. Biophys. Acta 1838 (5), 1235–1246. doi: 10.1016/j.bbamem.2014.01.021
The European Committee on Antimicrobial Susceptibility Testing. (2020). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0. Available at: http://www.eucast.org.
Thilakarathna, S. H., Rupasinghe, H. P. (2013). Flavonoid Bioavailability and Attempts for Bioavailability Enhancement. Nutrients 5 (9), 3367–3387. doi: 10.3390/nu5093367
Tran Trung, H., Truong Thi Huynh, H., Nguyen Thi Thuy, L., Nguyen Van Minh, H., Thi Nguyen, M. N., Luong Thi, M. N. (2020). Growth-Inhibiting, Bactericidal, Antibiofilm, and Urease Inhibitory Activities of Hibiscus rosa sinensis L. Flower Constituents Toward Antibiotic Sensitive- and Resistant-Strains of Helicobacter pylori. ACS Omega 5 (32), 20080–20089. doi: 10.1021/acsomega.0c01640
Tsuchiya, H., Iinuma, M. (2000). Reduction of Membrane Fluidity by Antibacterial Sophoraflavanone G Isolated From Sophora exigua. Phytomedicine 7 (2), 161–165. doi: 10.1016/S0944-7113(00)80089-6
Ustun, O., Ozcelik, B., Akyon, Y., Abbasoglu, U., Yesilada, E. (2006). Flavonoids With Anti-Helicobacter pylori Activity From Cistus laurifolius Leaves. J. Ethnopharmacol. 108 (3), 457–461. doi: 10.1016/j.jep.2006.06.001
Wang, Y. C. (2014). Medicinal Plant Activity on Helicobacter pylori Related Diseases. World J. Gastroenterol. 20 (30), 10368–10382. doi: 10.3748/wjg.v20.i30.10368
Wang, Y. C., Huang, K. M. (2013). In Vitro Anti-Inflammatory Effect of Apigenin in the Helicobacter pylori-Infected Gastric Adenocarcinoma Cells. Food Chem. Toxicol. 53, 376–383. doi: 10.1016/j.fct.2012.12.018
Wang, T., Li, Q., Bi, K. (2018). Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 13 (1), 12–23. doi: 10.1016/j.ajps.2017.08.004
White, R. L., Burgess, D. S., Manduru, M., Bosso, J. A. (1996). Comparison of Three Different In Vitro Methods of Detecting Synergy: Time-Kill, Checkerboard, and E Test. Antimicrob. Agents Chemother. 40 (8), 1914–1918. doi: 10.1128/AAC.40.8.1914
Wittschier, N., Lengsfeld, C., Vorthems, S., Stratmann, U., Ernst, J. F., Verspohl, E. J., et al. (2007). Large Molecules as Anti-Adhesive Compounds Against Pathogens. J. Pharm. Pharmacol. 59 (6), 777–786. doi: 10.1211/jpp.59.6.0004
Wu, J., Hu, D., Wang, K. X. (2008). Study of Scutellaria baicalensis and Baicalin Against Antimicrobial Susceptibility of Helicobacter pylori Strains In Vitro. Zhong Yao Cai 31 (5), 707–710.
Wu, D., Kong, Y., Han, C., Chen, J., Hu, L., Jiang, H., et al. (2008). D-Alanine:D-Alanine Ligase as a New Target for the Flavonoids Quercetin and Apigenin. Int. J. Antimicrob. Agents 32 (5), 421–426. doi: 10.1016/j.ijantimicag.2008.06.010
Xiao, Z. P., Wang, X. D., Peng, Z. Y., Huang, S., Yang, P., Li, Q. S., et al. (2012). Molecular Docking, Kinetics Study, and Structure-Activity Relationship Analysis of Quercetin and Its Analogous as Helicobacter pylori Urease Inhibitors. J. Agric. Food Chem. 60 (42), 10572–10577. doi: 10.1021/jf303393n
Yamaoka, Y. (2010). Mechanisms of Disease: Helicobacter pylori Virulence Factors. Nat. Rev. Gastroenterol. Hepatol. 7 (11), 629–641. doi: 10.1038/nrgastro.2010.154
Yang, B., Dong, Y., Wang, F., Zhang, Y. (2020). Nanoformulations to Enhance the Bioavailability and Physiological Functions of Polyphenols. Molecules 25 (20), 4613. doi: 10.3390/molecules25204613
Yeon, M. J., Lee, M. H., Kim, D. H., Yang, J. Y., Woo, H. J., Kwon, H. J., et al. (2019). Anti-Inflammatory Effects of Kaempferol on Helicobacter pylori-Induced Inflammation. Biosci. Biotechnol. Biochem. 83 (1), 166–173. doi: 10.1080/09168451.2018.1528140
Yesilada, E., Gurbuz, I., Ergun, E. (1997). Effects of Cistus Laurifolius L. Flowers on Gastric and Duodenal Lesions. J. Ethnopharmacol. 55 (3), 201–211. doi: 10.1016/s0378-8741(96)01502-4
Yuan, L., Li, X., He, S., Gao, C., Wang, C., Shao, Y. (2018). Effects of Natural Flavonoid Isoorientin on Growth Performance and Gut Microbiota of Mice. J. Agric. Food Chem. 66 (37), 9777–9784. doi: 10.1021/acs.jafc.8b03568
Yu, X. D., Zheng, R. B., Xie, J. H., Su, J. Y., Huang, X. Q., Wang, Y. H., et al. (2015). Biological Evaluation and Molecular Docking of Baicalin and Scutellarin as Helicobacter pylori Urease Inhibitors. J. Ethnopharmacol. 162, 69–78. doi: 10.1016/j.jep.2014.12.041
Zhang, S., Huang, J., Xie, X., He, Y., Mo, F., Luo, Z. (2017). Quercetin From Polygonum capitatum Protects Against Gastric Inflammation and Apoptosis Associated With Helicobacter pylori Infection by Affecting the Levels of P38mapk, BCL-2 and BAX. Molecules 22 (5), 744. doi: 10.3390/molecules22050744
Zhang, L., Kong, Y., Wu, D., Zhang, H., Wu, J., Chen, J., et al. (2008). Three Flavonoids Targeting the Beta-Hydroxyacyl-Acyl Carrier Protein Dehydratase From Helicobacter pylori: Crystal Structure Characterization With Enzymatic Inhibition Assay. Protein Sci. 17 (11), 1971–1978. doi: 10.1110/ps.036186.108
Keywords: Helicobacter pylori, flavonoids, plant-derived antimicrobials, antibiotic resistance, natural products
Citation: González A, Casado J and Lanas Á (2021) Fighting the Antibiotic Crisis: Flavonoids as Promising Antibacterial Drugs Against Helicobacter pylori Infection. Front. Cell. Infect. Microbiol. 11:709749. doi: 10.3389/fcimb.2021.709749
Received: 14 May 2021; Accepted: 05 July 2021;
Published: 20 July 2021.
Edited by:
Sinéad Marian Smith, Trinity College Dublin, IrelandReviewed by:
Jianzhong Zhang, Chinese Center For Disease Control and Prevention, ChinaAbbas Yadegar, Shahid Beheshti University of Medical Sciences, Iran
Copyright © 2021 González, Casado and Lanas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés González, YW5kcmVzZ2xlenJvZEBnbWFpbC5jb20=