 Jhommara Bautista1,2,3
Jhommara Bautista1,2,3 Andrés López-Cortés
Andrés López-Cortés- 1Cancer Research Group (CRG), Faculty of Medicine, Universidad de Las Américas, Quito, Ecuador
- 2Facultade de Ciencias, Campus de A Zapateira, Universidade da Coruña, A Coruña, Spain
- 3Instituto de Investigación Biomédica de A Coruña (INIBIC), A Coruña, Spain
Circadian rhythms orchestrate nearly every aspect of human physiology through a hierarchical network of clocks. While the suprachiasmatic nucleus (SCN) serves as the central pacemaker, peripheral clocks within the brain, heart, liver, gut, pancreas, adipose tissue, adrenal glands, lungs, and skeletal muscle independently regulate organ-specific functions. These autonomous oscillators, governed by transcriptional–translational feedback loops of core clock genes, align with environmental and physiological zeitgebers such as light, feeding, temperature, and hormones. Disruption of this temporal organization—through shift work, genetic alterations, or lifestyle factors—drives systemic misalignment, contributing to metabolic disease, cardiovascular dysfunction, neurodegeneration, cancer, and immune imbalance. This review explores the molecular mechanisms and physiological roles of peripheral clocks across organ systems, emphasizing their interplay with the SCN and zeitgebers. We also highlight emerging chronotherapeutic strategies that exploit circadian biology to optimize treatment outcomes. Understanding inter-organ circadian communication is key to unlocking personalized interventions and restoring systemic rhythmicity for health.
Introduction
Circadian rhythms are near-24-hour endogenous cycles that govern a wide spectrum of physiological and behavioral functions, including sleep-wake cycles, hormone secretion, metabolism, and immune responses. These rhythms are controlled by a complex hierarchy of cellular clocks present in nearly every tissue, all coordinated by the central pacemaker located in the suprachiasmatic nucleus (SCN) of the hypothalamus (1, 2). The molecular architecture underlying circadian clocks is based on transcriptional-translational feedback loops (TTFLs) involving core clock genes—BMAL1, BMAL2, CLOCK, CRY1, CRY2, CSNK1D, CSNK1E, NPAS2, NR1D1, NR1D2, PER1, PER2, PER3, RORA, RORB, RORC, and TIMELESS—modulated by secondary regulators like REV-ERBs and RORs (2, 3). These feedback loops drive rhythmic gene expression that synchronizes internal physiology with external or internal zeitgebers, including light, feeding schedules, temperature fluctuations, exercise, hormonal rhythms (e.g., glucocorticoids and insulin), microbial metabolites, and even social and pharmacological cues (1) (Figures 1A, B).
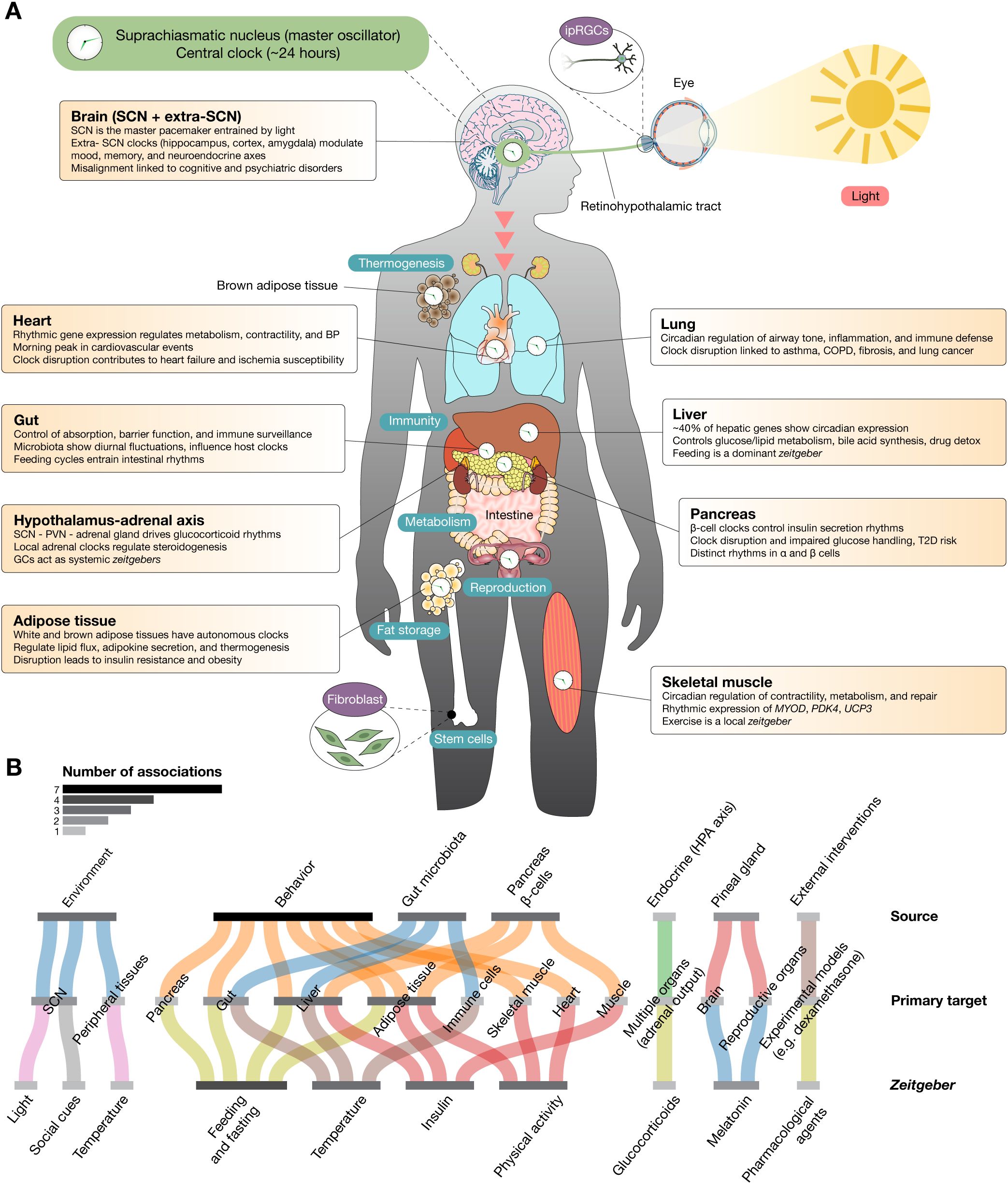
Figure 1. Multiorgan distribution of peripheral circadian clocks and their physiological functions. (A) The human body contains autonomous circadian clocks in nearly all major organs, each governed by transcriptional–translational feedback loops involving core clock genes. These peripheral clocks coordinate tissue-specific processes including metabolism, hormone secretion, immune defense, and cellular repair. Although synchronized by the central pacemaker in the suprachiasmatic nucleus via neural and hormonal signals, peripheral oscillators can also be entrained by local zeitgebers such as feeding, temperature, physical activity, and glucocorticoids. Key organ-specific functions are highlighted: brain clocks regulate cognition, mood, and neuroendocrine axes; cardiac clocks modulate metabolism and electrophysiology; gut clocks integrate host-microbiota dynamics; hepatic clocks control nutrient metabolism and xenobiotic detoxification; adipose clocks manage lipid flux and thermogenesis; pancreatic clocks govern insulin and glucagon rhythms; adrenal clocks drive glucocorticoid secretion; lung clocks influence airway physiology and immune defense; and skeletal muscle clocks regulate performance, energy use, and regeneration. Disruption of circadian alignment across these systems contributes to chronic diseases. (B) Sankey plot illustrating the associations between zeitgebers, primary targets, and their sources. The grayscale color scale represents the number of associations: lighter shades indicate fewer events, while darker shades represent a higher number of events. Zeitgebers are environmental or physiological cues that synchronize the internal biological clock (circadian rhythms) with the external environment. Primary targets refer to the main physiological systems, tissues, or molecular pathways that respond directly to zeitgebers. Sources are the origins of the zeitgeber signals or the systems delivering these cues. They can be external (environmental) or internal (systemic cues).
The SCN receives light input through intrinsically photosensitive retinal ganglion cells and synchronizes peripheral oscillators via neural, hormonal, and behavioral signals (1, 4). However, peripheral clocks possess a significant degree of autonomy and can be entrained by local cues such as feeding schedules, body temperature, and hormonal fluctuations (4–6). This decentralized and multi-oscillator structure enables organs to fine-tune their functions to specific daily demands, integrating both central and local signals to maintain systemic homeostasis (7, 8).
Disruptions in circadian synchronization are increasingly implicated in a broad array of chronic diseases, including metabolic syndrome, cardiovascular disease, neurodegenerative conditions, and cancer (1, 2, 9). Peripheral clocks in organs such as the liver, heart, pancreas, gut, adipose tissue, adrenal gland, lung, skeletal muscle, and even in non-SCN regions of the brain, independently regulate essential physiological processes. The interplay among these clocks—and between them and the SCN—provides opportunities for therapeutic interventions aimed at restoring circadian alignment. In this review, we examine the mechanisms and functions of peripheral clocks across multiple human organs and explore how their disruption contributes to disease.
Central and extra-SCN oscillators in neurocognitive and neuroendocrine regulation
The brain houses the central circadian pacemaker in the SCN and includes autonomous oscillators in other regions such as the hippocampus, amygdala, cortex, and olfactory bulb. These extra-SCN clocks support region-specific functions, including cognition, mood regulation, sensory processing, and neuroendocrine control (8, 10). All brain clocks operate via TTFLs involving CLOCK, BMAL1, PER, and CRY proteins, generating self-sustaining ~24-hour molecular cycles (8, 10). The SCN, situated in the anterior hypothalamus, receives photic signals through the retinohypothalamic tract and orchestrates circadian rhythms via neuroendocrine, autonomic, and behavioral outputs, as demonstrated predominantly in rodent models (11, 12). Its ~20,000 neurons include distinct populations such as VIP- and AVP-expressing cells, which mediate intercellular synchronization and broadcast time cues to both brain and peripheral tissues (12, 13). In the hippocampus, circadian rhythms regulate long-term potentiation and synaptic plasticity, which are critical for memory formation. Desynchronization between hippocampal and SCN rhythms has been shown in rodents to impair learning and memory (14, 15). Similarly, local clocks in the amygdala and cortex modulate emotion and cognition, with disruptions in these rhythms, observed in both animal models and human clinical studies, being associated with psychiatric and neurodegenerative disorders (8, 12, 15). The brain’s circadian influence extends to neuroendocrine axes, especially the hypothalamic-pituitary-adrenal (HPA) axis (16, 17). Rodent studies have demonstrated that the SCN modulates glucocorticoid rhythms via the PVN and adrenal gland, while glucocorticoids, in turn, serve as zeitgebers for peripheral clocks (12, 18). This bidirectional relationship is further modulated by feedback from metabolic cues and hormones like melatonin and cortisol, illustrating the integrative nature of circadian timing (18). Genetic or environmental circadian disruption—via clock gene knockouts or chronic jet lag models in rodents—can lead to altered neurogenesis, increased neuroinflammation, and cognitive decline (8, 15). Lastly, a growing body of evidence from human clinical studies and animal models links circadian misalignment to neurological and psychiatric conditions including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, amyotrophic lateral sclerosis, epilepsy, bipolar disorder, schizophrenia, autism spectrum disorders, anxiety disorders, dementia, and depression, emphasizing the critical role of circadian synchronization in mental health (19–22).
Temporal control of heart function, electrophysiology, and disease risk
The heart harbors an intrinsic circadian clock that regulates daily oscillations in cardiac metabolism, contractility, electrophysiology, and susceptibility to disease. While centrally influenced by the SCN pacemaker, the cardiac clock operates autonomously via TTFLs involving CLOCK, BMAL1, PER, CRY, and nuclear receptor proteins like REV-ERBs and RORs. These mechanisms have been extensively characterized in murine models and shown to regulate time-of-day–dependent gene expression in both rodent and human cardiac tissue (23–25). Cardiomyocytes and vascular cells exhibit circadian transcriptional programs that control approximately 6–10% of cardiac genes, including those involved in energy metabolism, contraction, redox homeostasis, and protein turnover (25–27). These rhythmic gene expressions prepare the heart for daily fluctuations in workload, with peak contractility, mitochondrial ATP production, and fatty acid oxidation occurring during the active phase (26, 28). In both rodent models and human observational studies, metabolic substrate use is temporally regulated: fatty acid oxidation dominates during the active phase, while glucose oxidation increases during rest (28). Disruption of these rhythms, such as through shift work or chronic circadian misalignment, impairs metabolic flexibility and contributes to pathological remodeling, including hypertrophy, fibrosis, and eventually heart failure (29). Experimental models demonstrate that repeated light-phase shifts—mimicking jet lag—induce diastolic dysfunction and features of heart failure with preserved ejection fraction (HFpEF). Mechanistically, this is linked to the downregulation of the CLOCK–sGC–cGMP–PKG1 axis, a signaling pathway critical for myocardial relaxation and vascular tone (29). Circadian rhythms also govern cardiovascular physiology at the systemic level. Blood pressure, heart rate, vascular tone, and cardiac output follow daily cycles, with peaks in the early active phase (23, 30). This coincides with increased sympathetic tone and renin-angiotensin activity, contributing to the observed morning surge in cardiovascular events such as myocardial infarction and stroke (23, 29, 31). Loss of synchrony between the SCN and peripheral cardiac clocks alters the timing of metabolic and electrophysiological events. Cardiomyocyte-specific deletions of BMAL1 or CLOCK result in impaired contractility, mitochondrial dysfunction, and heightened sensitivity to ischemic damage (26, 30). Moreover, circadian regulation of fibrinolysis and coagulation plays a critical role in thrombotic risk. Plasminogen activator inhibitor-1 (PAI-1), which inhibits fibrinolysis, peaks in the early morning alongside elevations in blood pressure and sympathetic activity, contributing to a transient prothrombotic state (31). Chronotherapy—aligning treatment with biological rhythms—offers a compelling strategy in cardiovascular medicine. Timing the administration of antihypertensives, antiplatelet agents, or metabolic modulators may improve outcomes and minimize adverse effects (25, 32). Pharmacological restoration of circadian pathways, such as via sGC activators like riociguat, is being explored as a potential intervention for circadian-related cardiac dysfunction (29).
Circadian regulation of gut physiology and the host–microbiota interface
The gastrointestinal (GI) tract exhibits robust circadian rhythmicity across its many functions, including nutrient absorption, barrier maintenance, immune defense, and host-microbiota interactions. These rhythms are driven by intrinsic gut clocks and modulated by external signals from the SCN, feeding behavior, and microbial activity. Most of the mechanistic evidence stems from murine models, although emerging data from human studies support similar principles (33, 34). At the molecular level, intestinal clocks operate through canonical TTFLs involving CLOCK, BMAL1, PER, and CRY proteins. These regulate circadian gene expression affecting epithelial renewal, nutrient transport, and mucosal defense (34, 35). Daily variations in stem cell proliferation, mucus secretion, and immune surveillance reflect the influence of circadian regulation and are synchronized with feeding rhythms and microbial cues (36, 37). The gut microbiota itself exhibits diurnal oscillations in both composition and function, shaped by feeding cycles and host circadian clocks (33, 35). Around 10–15% of bacterial taxa fluctuate across the day, generating time-specific production of metabolites like short-chain fatty acids (SCFAs) and bile acids. These microbial products can act as zeitgebers, synchronizing peripheral clocks and even influencing the phase of circadian genes like PER2 in the liver and colon, as shown in rodent models (38, 39). Microbiota-driven rhythms are critical for gut homeostasis. Germ-free or antibiotic-treated mice show reduced clock gene expression and diminished rhythmic chromatin accessibility in gut epithelial cells (38–40). Conversely, circadian misalignment due to behavioral or environmental disruptions alters the microbial composition and metabolite cycling, leading to dysbiosis and inflammation (35, 41). Dietary patterns strongly influence gut circadian rhythms. Time-restricted feeding restores microbial and host gene oscillations even in disrupted systems (33). In contrast, high-fat or high-sugar diets dampen both microbial and host rhythms, impairing metabolic health (34, 39, 42). Protein-rich diets, however, enhance microbial rhythmicity and metabolite diversity (33). The gut clock-microbiota axis also governs immune defense. Circadian regulation of antimicrobial peptides, leukocyte trafficking, and mucosal barrier integrity coordinates with microbial dynamics to mitigate infection risk (36, 37). Susceptibility to pathogens like Salmonella varies with the circadian phase, emphasizing the importance of synchrony between host defense and microbial activity (36). Despite recent insights, questions remain about which microbial species are rhythm drivers and how non-bacterial members like fungi or archaea fit into this regulatory network (41). Translating findings from animal models to human physiology remains a key challenge.
Metabolic coordination and chrono-pharmacology in the liver
The liver displays robust circadian rhythms tightly coupled to feeding-fasting cycles. Its peripheral clock operates through TTFLs involving CLOCK, BMAL1, PER, CRY, and nuclear protein receptors like REV-ERBs and RORs (43–45). These oscillators regulate about 40% of hepatic transcripts, including genes for glucose metabolism, lipid handling, bile acid synthesis, and xenobiotic detoxification, as shown primarily in murine transcriptomic studies (44, 46, 47). Though the SCN synchronizes liver rhythms via hormonal and neural outputs, feeding serves as a dominant zeitgeber. Restricted feeding can entrain liver clocks independently of light cues, demonstrating the organ’s metabolic sensitivity (45, 48, 49). The CLOCK: BMAL1 complex activates PER and CRY expression, which in turn inhibit CLOCK: BMAL1 in a negative feedback loop, while REV-ERBs and RORs further modulate BMAL1 transcription (45, 47, 50). Liver-specific deletion of BMAL1 disrupts glucose and lipid metabolism, leading to insulin resistance and hepatic steatosis (47, 48, 51). Circadian misalignment through jet lag or shift work, supported by both animal models and epidemiological studies in humans, exacerbates metabolic dysfunction and increases the risk of obesity and nonalcoholic fatty liver disease (NAFLD) (44, 46, 47). Chrono-pharmacology reveals time-of-day–dependent variability in drug metabolism. Hepatic expression of cytochrome P450 enzymes and inflammatory mediators like IL-6 and TNF-α fluctuates across the day, affecting responses to xenobiotics and pathogens (52). The liver also communicates with other organs: synchronized interactions with skeletal muscle and the gut optimize glucose homeostasis and energy balance. Disruption in intestinal clocks, for example, alters hepatic gluconeogenesis and lipid synthesis (49–51). Lastly, a study in clock-deficient mice shows that feeding can partially restore hepatic rhythms, though at reduced amplitude, highlighting the dual role of intrinsic clocks and behavioral cues (47).
Temporal regulation of lipid metabolism, thermogenesis, and endocrine signaling
Adipose tissue is a circadian-regulated endocrine organ comprising white (WAT), brown (BAT), and inducible beige fat. Each type contains autonomous clocks that synchronize with the SCN and respond to local cues such as feeding, temperature, and physical activity. Most functional and molecular insights derive from rodent models (53, 54). In WAT, core clock genes regulate lipid uptake and release, adipogenesis, and adipokine secretion. Genes such as PPARγ, NAMPT, and SREBF1 exhibit rhythmic expression, influencing lipid metabolism and insulin sensitivity. Adipokines like leptin and adiponectin are secreted diurnally in both mice and humans, modulating systemic energy balance (53, 55, 56). Disruption of WAT clocks—via high-fat diet or genetic ablation of BMAL1 or CLOCK—leads to impaired adipogenesis, altered hormone secretion, and increased risk of obesity and insulin resistance (55, 56). In BAT, the circadian clock governs thermogenesis via rhythmic expression of UCP1, PPARGC1A, and fatty acid oxidation genes. These processes peak before the active phase to meet energy demands. CLOCK and BMAL1 promote thermogenic gene expression, while REV-ERBα suppresses it. Sympathetic input from the SCN enhances BAT rhythmicity, which is diminished in circadian mutants or under chronic light exposure (57, 58). Beige adipocytes in WAT also exhibit circadian regulation. Their recruitment and thermogenic capacity decline with clock disruption, contributing to obesity in mouse models (58). Lastly, transcriptomic studies reveal that WAT gene expression follows intrinsic rhythms, with regulatory genes peaking in the morning and oxidative metabolism genes in the evening (59). Time-restricted feeding restores these rhythms and improves metabolic outcomes (60).
Circadian control of steroidogenesis and systemic hormonal rhythms
The adrenal gland generates circadian glucocorticoid (GC) rhythms crucial for homeostasis and stress responses. While GC secretion is SCN-regulated via the HPA axis, the adrenal cortex also contains autonomous clocks essential for steroidogenesis—findings supported primarily by rodent studies (61, 62). Within the zona fasciculata, the BMAL1:CLOCK complex drives PER and CRY expression and regulates steroidogenic genes such as STAR, which mediates cholesterol transport into mitochondria. STAR expression follows a circadian pattern and is disrupted by adrenal-specific BMAL1 deletion, impairing GC rhythms (63). The SCN entrains the adrenal clock through hormonal cues and sympathetic innervation via the splanchnic nerve. Light pulses can induce acute GC release independently of adrenal clocks in both rodent and primate models, reflecting direct SCN influence (64). Optimal GC rhythmicity requires both central and adrenal clocks. Disruption leads to blunted hormonal cycles and pathologies including hypertension, metabolic syndrome, and altered feeding behavior (62, 64, 65). GCs also serve as systemic zeitgebers, synchronizing peripheral clocks and aligning energy mobilization with activity cycles. Dysregulation impairs immune function, cognition, and cardiovascular health (66–68).
Rhythmic regulation of lung function, inflammation, and disease susceptibility
The lung features strong circadian regulation that impacts pulmonary physiology, immune defense, and response to environmental stress. Autonomous lung clocks, composed of core TTFL components and nuclear receptors like REV-ERBα and RORα, orchestrate rhythmic gene expression in airway cells and resident immune populations, primarily characterized in rodent models (69, 70). These rhythms influence airway tone, mucus secretion, inflammation, and oxidative stress. Lung clocks also respond to systemic signals such as glucocorticoids and catecholamines, aligning pulmonary functions with behavioral cycles (69, 70). Disruption of lung clocks contributes to diseases such as asthma, chronic obstructive pulmonary disease (COPD), and fibrosis. In COPD, core clock genes like BMAL1 and CLOCK are suppressed by cigarette smoke, accelerating inflammation and senescence via the MAPK pathway (71). REV-ERBα modulates fibrosis-related gene expression; its loss exacerbates fibrotic responses, while its activation offers therapeutic potential (72). Circadian rhythms also regulate pulmonary immune responses. Time-of-day–dependent variation in leukocyte trafficking and cytokine expression alters susceptibility to infection and inflammation (73–76). Lastly, chronic circadian disruption promotes lung tumorigenesis in KRAS-driven models through HSF1 hyperactivation (77).
Circadian control of islet hormone secretion and glucose homeostasis
The pancreas exhibits circadian regulation in both endocrine and exocrine compartments. Core clock genes orchestrate daily rhythms in insulin secretion, glucose sensing, and islet cell function (78–80). In β-cells, autonomous clocks control insulin release in response to feeding. BMAL1 deletion impairs glucose-stimulated insulin secretion (GSIS), reduces β-cell mass, and alters circadian gene expression (81, 82). α- and β-cells exhibit distinct rhythmic gene profiles, coordinating insulin and glucagon output across the day (81). Clock genes regulate GLUT2, GCK, and components of the exocytosis machinery such as SNAREs and calcium channels (78, 83). GSIS peaks during the active phase, optimizing nutrient handling (78). Disruption of pancreatic clocks contributes to type 2 diabetes by impairing insulin secretion and increasing resistance (80, 83). Circadian disruption also exacerbates acute pancreatitis by impairing immune timing and tissue recovery (84). Lastly, in pancreatic cancer, circadian gene expression is suppressed, and BMAL1 deficiency in murine models enhances tumor growth and chemoresistance (85).
Muscle metabolism, performance, and regeneration under circadian control
Skeletal muscle contains autonomous circadian clocks that govern metabolism, contractility, and repair. These TTFL-based clocks regulate the expression of genes like MYOD, UCP3, FBXO32, PDK4, and MYH1, affecting muscle growth, mitochondrial activity, and protein turnover (51, 86, 87). About 3–5% of the muscle transcriptome is rhythmically expressed, peaking during the active phase in mice to support performance and energy use (86–88). BMAL1 deletion leads to muscle atrophy, impaired contractility, and altered fiber composition via disrupted MYOD and WNT signaling (88, 89). Muscle clocks can be entrained by feeding and exercise independently of the SCN. Scheduled activity resets gene expression rhythms, enhancing performance and recovery (51, 86). Coordination with liver clocks is necessary for systemic glucose tolerance. Reconstitution of BMAL1 in both tissues restores metabolic homeostasis in otherwise arrhythmic mice (86). Lastly, circadian disruption impairs strength and regeneration and exacerbates conditions like Duchenne muscular dystrophy (DMD), where altered myogenesis and structure reflect underlying clock dysfunction (88).
Interplay between circadian clocks across the human body
The circadian system is a hierarchically organized and dynamically coupled network of oscillators that spans the entire human body. At its apex lies the SCN, which acts as the master pacemaker, synchronizing peripheral clocks via neural, hormonal, and behavioral cues. However, each peripheral oscillator, embedded in tissues such as the liver, heart, gut, muscle, adipose tissue, adrenal gland, and lungs, retains a remarkable degree of autonomy, allowing it to respond to tissue-specific zeitgebers like feeding, temperature, microbial metabolites, and physical activity. These principles are supported by extensive rodent data and are increasingly validated by human studies (90, 91).
Coherence among these clocks is not maintained solely by unidirectional signaling from the SCN. Instead, circadian rhythms in peripheral tissues exhibit a reciprocal interplay that is essential for systemic homeostasis. Experimental evidence reveals that peripheral clocks exhibit organ-specific phase relationships with the SCN and with each other, and disruptions in one organ’s clock can propagate misalignment across the network (92). For instance, hepatic clocks influence pancreatic insulin secretion, and gut-derived microbial metabolites modulate hepatic gene expression via enterohepatic circulation (90, 93). Coupling mechanisms include shared hormonal rhythms, autonomic nervous system outputs, and circulating metabolites that act as systemic synchronizers. Glucocorticoids, for instance, not only reflect SCN activity through the HPA axis but also entrain clocks in peripheral tissues including the lung, liver, and adipose tissue (91, 94). Moreover, peripheral clocks themselves contribute to the feedback regulation of the HPA axis and metabolic pathways, suggesting a bidirectional flow of timing information (90, 95).
Mathematical and experimental modeling of circadian networks has further supported the notion that the circadian system behaves as a coupled oscillator network, where the stability of rhythmic output depends on robust inter-oscillator communication (91). Weak or disrupted coupling, as seen in chronic jet lag, shift work, or genetic ablation of clock genes, leads to phase desynchrony and dampened rhythmicity, thereby increasing vulnerability to metabolic, inflammatory, and neuropsychiatric diseases (92, 96). Understanding the interplay among circadian clocks opens new avenues for chrono-therapeutic strategies that aim to restore inter-organ synchrony. Approaches such as timed feeding, light exposure, physical activity, and pharmacological modulation of clock components are under investigation to recalibrate misaligned circadian systems in clinical contexts.
Conclusions and future perspectives toward circadian precision medicine
Circadian rhythms are central to the temporal regulation of physiology across virtually all organs. The existence of autonomous peripheral clocks in the brain, heart, gut, liver, adipose tissue, adrenal gland, lung, pancreas, and skeletal muscle—established largely through rodent studies and increasingly supported by human transcriptomic and imaging data— highlights the widespread integration of circadian timing into cellular and systemic processes. These clocks are not only coordinated by the suprachiasmatic nucleus but are also entrainable by diverse environmental and physiological zeitgebers, such as light, feeding, temperature, and hormonal cues (3). The precision of this temporal network is critical for health, while its disruption contributes to a broad range of chronic diseases.
Misalignment of circadian rhythms, whether due to genetic mutations, lifestyle factors like shift work, or environmental stressors, is now recognized as a key contributor to pathologies including metabolic syndrome, cardiovascular disease, neurodegenerative disorders, cancer, and immune dysfunction (2, 7, 9). At the molecular level, these disruptions alter rhythmic gene expression, leading to impaired cellular metabolism, hormone secretion, immune responses, and stress resilience. The mechanistic insight gained from the study of peripheral clocks has also revealed tissue-specific vulnerabilities and interactions, such as the dependence of hepatic and skeletal muscle glucose regulation on synchronized oscillators (4).
Importantly, a growing body of evidence reveals that sex differences significantly modulate circadian rhythmicity, influencing both the amplitude and phase of biological rhythms across nearly all organ systems (97–99). These sex-specific variations arise from the interplay of genetic, hormonal, neuroanatomical, and epigenetic factors that affect the central pacemaker in the SCN as well as peripheral clocks throughout the body (100). Estrogens, androgens, and their respective receptors (ERα, ERβ, and AR), which are expressed in the SCN and its afferent and efferent pathways, contribute to sex-specific entrainment responses to photic and nonphotic zeitgebers (101). For instance, the SCN and its inputs from the retinohypothalamic tract, intergeniculate leaflet, and dorsal raphe nuclei all exhibit sexually dimorphic expression of these steroid receptors, allowing sex hormones to directly modulate the clock’s structure and output (97).
Chronotherapy—tailoring the timing of treatment to align with circadian phases—has shown promise in enhancing therapeutic efficacy and reducing toxicity. Cardiovascular drugs, chemotherapeutics, and immunomodulators can all benefit from circadian-informed administration schedules (8, 9). For example, the effectiveness of anticancer drugs has been shown to vary with the time of administration, due to circadian modulation of DNA repair, drug metabolism, and cell cycle activity (9). Furthermore, leveraging circadian biology has advanced immunotherapy strategies by identifying windows of enhanced immune responsiveness, such as time-of-day–dependent T-cell activation and cytokine production (102–104).
Moving forward, a deeper understanding of inter-organ circadian communication and zeitgeber-specific entrainment mechanisms will be essential for translating chronobiological principles into clinical practice. This includes identifying optimal timing for drug delivery, meal schedules, physical activity, and even surgical procedures. Technologies such as circadian biomarkers, wearable sensors, and computational modeling will support precision medicine approaches tailored to individual circadian profiles (1, 6). In conclusion, circadian biology offers a transformative framework for understanding human physiology and disease. Targeting peripheral clocks and synchronizing internal rhythms holds immense therapeutic potential across multiple domains, from metabolic and cardiovascular health to neurodegeneration, immunity, and cancer. As our understanding of these complex temporal networks deepens, chronobiology is poised to become a cornerstone of personalized and preventive medicine.
Author contributions
JB: Methodology, Investigation, Conceptualization, Writing – original draft, Writing – review & editing, Visualization, Formal analysis, Validation, Resources, Data curation. SO-M: Data curation, Investigation, Resources, Writing – original draft, Visualization, Conceptualization, Validation, Writing – review & editing, Methodology, Formal analysis. DO-L: Investigation, Validation, Conceptualization, Resources, Writing – review & editing, Formal analysis, Writing – original draft, Data curation, Visualization, Methodology. AL-C: Visualization, Data curation, Methodology, Validation, Conceptualization, Investigation, Supervision, Funding acquisition, Writing – review & editing, Project administration, Resources, Formal analysis, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors would like to acknowledge the support and funding provided by Universidad de Las Américas from Quito, Ecuador.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Pérez-Villa A, Echeverría-Garcés G, Ramos-Medina MJ, Prathap L, Martínez-López M, Ramírez-Sánchez D, et al. Integrated multi-omics analysis reveals the molecular interplay between circadian clocks and cancer pathogenesis. Sci Rep. (2023) 13:14198. doi: 10.1038/s41598-023-39401-1
2. de Assis LVM and Oster H. The circadian clock and metabolic homeostasis: entangled networks. Cell Mol Life Sci. (2021) 78:4563–87. doi: 10.1007/s00018-021-03800-2
3. Heyde I and Oster H. Differentiating external zeitgeber impact on peripheral circadian clock resetting. Sci Rep. (2019) 9:20114. doi: 10.1038/s41598-019-56323-z
4. Zhang S, Dai M, Wang X, Jiang S-H, Hu L-P, Zhang X-L, et al. Signalling entrains the peripheral circadian clock. Cell Signal. (2020) 69:109433. doi: 10.1016/j.cellsig.2019.109433
5. Lamia KA, Storch K-F, and Weitz CJ. Physiological significance of a peripheral tissue circadian clock. Proc Natl Acad Sci USA. (2008) 105:15172–7. doi: 10.1073/pnas.0806717105
6. Saini C, Brown SA, and Dibner C. Human peripheral clocks: applications for studying circadian phenotypes in physiology and pathophysiology. Front Neurol. (2015) 6:95. doi: 10.3389/fneur.2015.00095
7. Bass J and Lazar MA. Circadian time signatures of fitness and disease. Science. (2016) 354:994–9. doi: 10.1126/science.aah4965
8. Albrecht U. Timing to perfection: the biology of central and peripheral circadian clocks. Neuron. (2012) 74:246–60. doi: 10.1016/j.neuron.2012.04.006
9. Shafi AA and Knudsen KE. Cancer and the circadian clock. Cancer Res. (2019) 79:3806–14. doi: 10.1158/0008-5472.CAN-19-0566
10. Begemann K, Neumann A-M, and Oster H. Regulation and function of extra-SCN circadian oscillators in the brain. Acta Physiol (Oxf). (2020) 229:e13446. doi: 10.1111/apha.13446
11. Buijs FN, León-Mercado L, Guzmán-Ruiz M, Guerrero-Vargas NN, Romo-Nava F, and Buijs RM. The circadian system: A regulatory feedback network of periphery and brain. Physiol (Bethesda). (2016) 31:170–81. doi: 10.1152/physiol.00037.2015
12. Schurhoff N and Toborek M. Circadian rhythms in the blood-brain barrier: impact on neurological disorders and stress responses. Mol Brain. (2023) 16:5. doi: 10.1186/s13041-023-00997-0
13. Mendoza J. Brain circadian clocks timing the 24h rhythms of behavior. NPJ Biol Timing Sleep. (2025) 2:13. doi: 10.1038/s44323-025-00030-8
14. Hughey JJ and Butte AJ. Differential phasing between circadian clocks in the brain and peripheral organs in humans. J Biol Rhythms. (2016) 31:588–97. doi: 10.1177/0748730416668049
15. Van Drunen R and Eckel-Mahan K. Circadian rhythms as modulators of brain health during development and throughout aging. Front Neural Circuits. (2022) 16:1059229. doi: 10.3389/fncir.2022.1059229
16. Kumar A, Vaca-Dempere M, Mortimer T, Deryagin O, Smith JG, Petrus P, et al. Brain-muscle communication prevents muscle aging by maintaining daily physiology. Science. (2024) 384:563–72. doi: 10.1126/science.adj8533
17. Mortimer T, Zinna VM, Atalay M, Laudanna C, Deryagin O, Posas G, et al. The epidermal circadian clock integrates and subverts brain signals to guarantee skin homeostasis. Cell Stem Cell. (2024) 31:834–849.e4. doi: 10.1016/j.stem.2024.04.013
18. Hughes ME, Hong H-K, Chong JL, Indacochea AA, Lee SS, Han M, et al. Brain-specific rescue of Clock reveals system-driven transcriptional rhythms in peripheral tissue. PloS Genet. (2012) 8:e1002835. doi: 10.1371/journal.pgen.1002835
19. Musiek ES and Holtzman DM. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science. (2016) 354:1004–8. doi: 10.1126/science.aah4968
20. Leng Y, Musiek ES, Hu K, Cappuccio FP, and Yaffe K. Association between circadian rhythms and neurodegenerative diseases. Lancet Neurol. (2019) 18:307–18. doi: 10.1016/S1474-4422(18)30461-7
21. Logan RW and McClung CA. Rhythms of life: circadian disruption and brain disorders across the lifespan. Nat Rev Neurosci. (2019) 20:49–65. doi: 10.1038/s41583-018-0088-y
22. Tang T, Zhou Y, and Zhai X. Circadian rhythm and epilepsy: a nationally representative cross-sectional study based on actigraphy data. Front Neurol. (2024) 15:1496507. doi: 10.3389/fneur.2024.1496507
23. Reilly DF, Westgate EJ, and FitzGerald GA. Peripheral circadian clocks in the vasculature. Arterioscler Thromb Vasc Biol. (2007) 27:1694–705. doi: 10.1161/ATVBAHA.107.144923
24. Sartor F, Ferrero-Bordera B, Haspel J, Sperandio M, Holloway PM, and Merrow M. Circadian clock and hypoxia. Circ Res. (2024) 134:618–34. doi: 10.1161/CIRCRESAHA.124.323518
25. Che Y, Shimizu Y, Hayashi T, Suzuki J, Pu Z, Tsuzuki K, et al. Chronic circadian rhythm disorder induces heart failure with preserved ejection fraction-like phenotype through the Clock-sGC-cGMP-PKG1 signaling pathway. Sci Rep. (2024) 14:10777. doi: 10.1038/s41598-024-61710-2
26. Young ME. The circadian clock within the heart: potential influence on myocardial gene expression, metabolism, and function. Am J Physiol Heart Circ Physiol. (2006) 290:H1–16. doi: 10.1152/ajpheart.00582.2005
27. Ramos-Medina MJ, Echeverría-Garcés G, Kyriakidis NC, León Cáceres Á, Ortiz-Prado E, Bautista J, et al. CardiOmics signatures reveal therapeutically actionable targets and drugs for cardiovascular diseases. Heliyon. (2024) 10:e23682. doi: 10.1016/j.heliyon.2023.e23682
28. Lal H, Verma SK, Wang Y, Xie M, and Young ME. Circadian rhythms in cardiovascular metabolism. Circ Res. (2024) 134:635–58. doi: 10.1161/CIRCRESAHA.123.323520
29. Malhan D and Relógio A. A matter of timing? The influence of circadian rhythms on cardiac physiology and disease. Eur Heart J. (2024) 45:561–3. doi: 10.1093/eurheartj/ehad816
30. Young ME. The cardiac circadian clock: implications for cardiovascular disease and its treatment. JACC Basic Transl Sci. (2023) 8:1613–28. doi: 10.1016/j.jacbts.2023.03.024
31. Maemura K, Takeda N, and Nagai R. Circadian rhythms in the CNS and peripheral clock disorders: role of the biological clock in cardiovascular diseases. J Pharmacol Sci. (2007) 103:134–8. doi: 10.1254/jphs.fmj06003x2
32. Takeda N and Maemura K. Circadian clock and cardiovascular disease. J Cardiol. (2011) 57:249–56. doi: 10.1016/j.jjcc.2011.02.006
33. Heddes M, Altaha B, Niu Y, Reitmeier S, Kleigrewe K, Haller D, et al. The intestinal clock drives the microbiome to maintain gastrointestinal homeostasis. Nat Commun. (2022) 13:6068. doi: 10.1038/s41467-022-33609-x
34. Zhao E, Tait C, Minacapelli CD, Catalano C, and Rustgi VK. Circadian rhythms, the gut microbiome, and metabolic disorders. Gastro Hep Adv. (2022) 1:93–105. doi: 10.1016/j.gastha.2021.10.008
35. Schmid DW, Capilla-Lasheras P, Dominoni DM, Müller-Klein N, Sommer S, and Risely A. Circadian rhythms of hosts and their gut microbiomes: Implications for animal physiology and ecology. Funct Ecol. (2023) 37:476–87. doi: 10.1111/1365-2435.14255
36. Rosselot AE, Hong CI, and Moore SR. Rhythm and bugs: circadian clocks, gut microbiota, and enteric infections. Curr Opin Gastroenterol. (2016) 32:7–11. doi: 10.1097/MOG.0000000000000227
37. Lotti S, Dinu M, Colombini B, Amedei A, and Sofi F. Circadian rhythms, gut microbiota, and diet: Possible implications for health. Nutr Metab Cardiovasc Dis. (2023) 33:1490–500. doi: 10.1016/j.numecd.2023.05.009
38. Zhang Y, Li Y, and Barber AF. The microbiome stabilizes circadian rhythms in the gut. Proc Natl Acad Sci USA. (2023) 120:5. doi: 10.1073/pnas.2217532120
39. Gutierrez Lopez DE, Lashinger LM, Weinstock GM, and Bray MS. Circadian rhythms and the gut microbiome synchronize the host’s metabolic response to diet. Cell Metab. (2021) 33:873–87. doi: 10.1016/j.cmet.2021.03.015
40. Frazier K and Chang EB. Intersection of the gut microbiome and circadian rhythms in metabolism. Trends Endocrinol Metab. (2020) 31:25–36. doi: 10.1016/j.tem.2019.08.013
41. Wollmuth EM and Angert ER. Microbial circadian clocks: host-microbe interplay in diel cycles. BMC Microbiol. (2023) 23:124. doi: 10.1186/s12866-023-02839-4
42. Echeverría CE, Oyarzún VI, López-Cortés A, Cancino J, Sotomayor PC, Goncalves MD, et al. Biological role of fructose in the male reproductive system: Potential implications for prostate cancer. Prostate. (2024) 84:8–24. doi: 10.1002/pros.24631
43. Mukherji A, Bailey SM, Staels B, and Baumert TF. The circadian clock and liver function in health and disease. J Hepatol. (2019) 71:200–11. doi: 10.1016/j.jhep.2019.03.020
44. Reinke H and Asher G. Circadian clock control of liver metabolic functions. Gastroenterology. (2016) 150:574–80. doi: 10.1053/j.gastro.2015.11.043
45. Tahara Y and Shibata S. Circadian rhythms of liver physiology and disease: experimental and clinical evidence. Nat Rev Gastroenterol Hepatol. (2016) 13:217–26. doi: 10.1038/nrgastro.2016.8
46. Daniels LJ, Kay D, Marjot T, Hodson L, and Ray DW. Circadian regulation of liver metabolism: experimental approaches in human, rodent, and cellular models. Am J Physiol Cell Physiol. (2023) 325:C1158–77. doi: 10.1152/ajpcell.00551.2022
47. Weger BD, Gobet C, David FPA, Atger F, Martin E, Phillips NE, et al. Systematic analysis of differential rhythmic liver gene expression mediated by the circadian clock and feeding rhythms. Proc Natl Acad Sci USA. (2021) 118(3):e2015803118. doi: 10.1073/pnas.2015803118
48. Greco CM, Koronowski KB, Smith JG, Shi J, Kunderfranco P, Carriero R, et al. Integration of feeding behavior by the liver circadian clock reveals network dependency of metabolic rhythms. Sci Adv. (2021) 7:eabi7828. doi: 10.1126/sciadv.abi7828
49. Manella G, Sabath E, Aviram R, Dandavate V, Ezagouri S, Golik M, et al. The liver-clock coordinates rhythmicity of peripheral tissues in response to feeding. Nat Metab. (2021) 3:829–42. doi: 10.1038/s42255-021-00395-7
50. Fondevila MF and Schnabl B. Intestinal clock modulates rhythmic hepatic metabolism. J Hepatol. (2023) 79:589–91. doi: 10.1016/j.jhep.2023.05.021
51. Smith JG, Koronowski KB, Mortimer T, Sato T, Greco CM, Petrus P, et al. Liver and muscle circadian clocks cooperate to support glucose tolerance in mice. Cell Rep. (2023) 42:112588. doi: 10.1016/j.celrep.2023.112588
52. March S, Nerurkar N, Jain A, Andrus L, Kim D, Whittaker CA, et al. Autonomous circadian rhythms in the human hepatocyte regulate hepatic drug metabolism and inflammatory responses. Sci Adv. (2024) 10:eadm9281. doi: 10.1126/sciadv.adm9281
53. Heyde I, Begemann K, and Oster H. Contributions of white and brown adipose tissues to the circadian regulation of energy metabolism. Endocrinology. (2021) 162. doi: 10.1210/endocr/bqab009
54. Froy O and Garaulet M. The circadian clock in white and brown adipose tissue: mechanistic, endocrine, and clinical aspects. Endocr Rev. (2018) 39:261–73. doi: 10.1210/er.2017-00193
55. Ribas-Latre A, Santos RB, Fekry B, Tamim YM, Shivshankar S, Mohamed AMT, et al. Cellular and physiological circadian mechanisms drive diurnal cell proliferation and expansion of white adipose tissue. Nat Commun. (2021) 12:3482. doi: 10.1038/s41467-021-23770-0
56. Aggarwal A, Costa MJ, Rivero-Gutiérrez B, Ji L, Morgan SL, and Feldman BJ. The circadian clock regulates adipogenesis by a per3 crosstalk pathway to klf15. Cell Rep. (2017) 21:2367–75. doi: 10.1016/j.celrep.2017.11.004
57. Straat ME, Hogenboom R, Boon MR, Rensen PCN, and Kooijman S. Circadian control of brown adipose tissue. Biochim Biophys Acta Mol Cell Biol Lipids. (2021) 1866:158961. doi: 10.1016/j.bbalip.2021.158961
58. Peng X and Chen Y. The emerging role of circadian rhythms in the development and function of thermogenic fat. Front Endocrinol (Lausanne). (2023) 14:1175845. doi: 10.3389/fendo.2023.1175845
59. Christou S, Wehrens SMT, Isherwood C, Möller-Levet CS, Wu H, Revell VL, et al. Circadian regulation in human white adipose tissue revealed by transcriptome and metabolic network analysis. Sci Rep. (2019) 9:2641. doi: 10.1038/s41598-019-39668-3
60. Yamamuro D, Takahashi M, Nagashima S, Wakabayashi T, Yamazaki H, Takei A, et al. Peripheral circadian rhythms in the liver and white adipose tissue of mice are attenuated by constant light and restored by time-restricted feeding. PloS One. (2020) 15:e0234439. doi: 10.1371/journal.pone.0234439
61. Chung S, Son GH, and Kim K. Circadian rhythm of adrenal glucocorticoid: its regulation and clinical implications. Biochim Biophys Acta. (2011) 1812:581–91. doi: 10.1016/j.bbadis.2011.02.003
62. Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. (2006) 4:163–73. doi: 10.1016/j.cmet.2006.07.002
63. Son GH, Chung S, Choe HK, Kim H-D, Baik S-M, Lee H, et al. Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production. Proc Natl Acad Sci USA. (2008) 105:20970–5. doi: 10.1073/pnas.0806962106
64. Chung S, Lee EJ, Cha HK, Kim J, Kim D, Son GH, et al. Cooperative roles of the suprachiasmatic nucleus central clock and the adrenal clock in controlling circadian glucocorticoid rhythm. Sci Rep. (2017) 7:46404. doi: 10.1038/srep46404
65. Moore BN and Pluznick JL. BMAL1 in the adrenal gland: it’s about time-A perspective on “Adrenal-specific KO of the circadian clock protein BMAL1 alters blood pressure rhythm and timing of eating behavior. Funct (Oxf). (2023) 4:zqad008. doi: 10.1093/function/zqad008
66. Dumbell R, Matveeva O, and Oster H. Circadian clocks, stress, and immunity. Front Endocrinol (Lausanne). (2016) 7:37. doi: 10.3389/fendo.2016.00037
67. Leliavski A, Dumbell R, Ott V, and Oster H. Adrenal clocks and the role of adrenal hormones in the regulation of circadian physiology. J Biol Rhythms. (2015) 30:20–34. doi: 10.1177/0748730414553971
68. Dickmeis T, Lahiri K, Nica G, Vallone D, Santoriello C, Neumann CJ, et al. Glucocorticoids play a key role in circadian cell cycle rhythms. PloS Biol. (2007) 5:e78. doi: 10.1371/journal.pbio.0050078
69. Giri A, Wang Q, Rahman I, and Sundar IK. Circadian molecular clock disruption in chronic pulmonary diseases. Trends Mol Med. (2022) 28:513–27. doi: 10.1016/j.molmed.2022.04.002
70. Sundar IK, Yao H, Sellix MT, and Rahman I. Circadian molecular clock in lung pathophysiology. Am J Physiol Lung Cell Mol Physiol. (2015) 309:L1056–75. doi: 10.1152/ajplung.00152.2015
71. Li L, Zhang M, Zhao C, Cheng Y, Liu C, and Shi M. Circadian clock gene Clock-Bmal1 regulates cellular senescence in Chronic obstructive pulmonary disease. BMC Pulm Med. (2022) 22:435. doi: 10.1186/s12890-022-02237-y
72. Wang Q, Sundar IK, Lucas JH, Park J-G, Nogales A, Martinez-Sobrido L, et al. Circadian clock molecule REV-ERBα regulates lung fibrotic progression through collagen stabilization. Nat Commun. (2023) 14:1295. doi: 10.1038/s41467-023-36896-0
73. Haspel JA, Chettimada S, Shaik RS, Chu J-H, Raby BA, Cernadas M, et al. Circadian rhythm reprogramming during lung inflammation. Nat Commun. (2014) 5:4753. doi: 10.1038/ncomms5753
74. Nosal C, Ehlers A, and Haspel JA. Why lungs keep time: circadian rhythms and lung immunity. Annu Rev Physiol. (2020) 82:391–412. doi: 10.1146/annurev-physiol-021119-034602
75. López-Cortés A, Guerrero S, Ortiz-Prado E, Yumiceba V, Vera-Guapi A, León Cáceres Á, et al. Pulmonary inflammatory response in lethal COVID-19 reveals potential therapeutic targets and drugs in phases III/IV clinical trials. Front Pharmacol. (2022) 13:833174. doi: 10.3389/fphar.2022.833174
76. López-Cortés A, Guevara-Ramírez P, Kyriakidis NC, Barba-Ostria C, León Cáceres Á, Guerrero S, et al. In silico analyses of immune system protein interactome network, single-cell RNA sequencing of human tissues, and artificial neural networks reveal potential therapeutic targets for drug repurposing against COVID-19. Front Pharmacol. (2021) 12:598925. doi: 10.3389/fphar.2021.598925
77. Pariollaud M, Ibrahim LH, Irizarry E, Mello RM, Chan AB, Altman BJ, et al. Circadian disruption enhances HSF1 signaling and tumorigenesis in Kras-driven lung cancer. Sci Adv. (2022) 8:eabo1123. doi: 10.1126/sciadv.abo1123
78. Seshadri N and Doucette CA. Circadian regulation of the pancreatic beta cell. Endocrinology. (2021) 162(9):bqab089. doi: 10.1210/endocr/bqab089
79. García-Costela M, Escudero-Feliú J, Puentes-Pardo JD, San Juán SM, Morales-Santana S, Ríos-Arrabal S, et al. Circadian genes as therapeutic targets in pancreatic cancer. Front Endocrinol (Lausanne). (2020) 11:638. doi: 10.3389/fendo.2020.00638
80. Chan K, Wong FS, and Pearson JA. Circadian rhythms and pancreas physiology: A review. Front Endocrinol (Lausanne). (2022) 13:920261. doi: 10.3389/fendo.2022.920261
81. Petrenko V, Saini C, Giovannoni L, Gobet C, Sage D, Unser M, et al. Pancreatic α- and β-cellular clocks have distinct molecular properties and impact on islet hormone secretion and gene expression. Genes Dev. (2017) 31:383–98. doi: 10.1101/gad.290379.116
82. Sadacca LA, Lamia KA, deLemos AS, Blum B, and Weitz CJ. An intrinsic circadian clock of the pancreas is required for normal insulin release and glucose homeostasis in mice. Diabetologia. (2011) 54:120–4. doi: 10.1007/s00125-010-1920-8
83. Stenvers DJ, Scheer FAJL, Schrauwen P, la Fleur SE, and Kalsbeek A. Circadian clocks and insulin resistance. Nat Rev Endocrinol. (2019) 15:75–89. doi: 10.1038/s41574-018-0122-1
84. Waddell H, Stevenson TJ, and Mole DJ. The role of the circadian rhythms in critical illness with a focus on acute pancreatitis. Heliyon. (2023) 9:e15335. doi: 10.1016/j.heliyon.2023.e15335
85. Schwartz PB, Nukaya M, Berres ME, Rubinstein CD, Wu G, Hogenesch JB, et al. The circadian clock is disrupted in pancreatic cancer. PloS Genet. (2023) 19:e1010770. doi: 10.1371/journal.pgen.1010770
86. Hesketh SJ and Esser KA. The clockwork of champions: Influence of circadian biology on exercise performance. Free Radic Biol Med. (2024) 224:78–87. doi: 10.1016/j.freeradbiomed.2024.08.020
87. Dudek M and Meng Q-J. Running on time: the role of circadian clocks in the musculoskeletal system. Biochem J. (2014) 463:1–8. doi: 10.1042/BJ20140700
88. Kiperman T and Ma K. Circadian clock in muscle disease etiology and therapeutic potential for duchenne muscular dystrophy. Int J Mol Sci. (2024) 25(9):4767. doi: 10.3390/ijms25094767
89. Aoyama S and Shibata S. The role of circadian rhythms in muscular and osseous physiology and their regulation by nutrition and exercise. Front Neurosci. (2017) 11:63. doi: 10.3389/fnins.2017.00063
90. Grosjean E, Simonneaux V, and Challet E. Reciprocal interactions between circadian clocks, food intake, and energy metabolism. Biol (Basel). (2023) 12(4):539. doi: 10.3390/biology12040539
91. Finger A-M, Dibner C, and Kramer A. Coupled network of the circadian clocks: a driving force of rhythmic physiology. FEBS Lett. (2020) 594:2734–69. doi: 10.1002/1873-3468.13898
92. Roenneberg T and Merrow M. The circadian clock and human health. Curr Biol. (2016) 26:R432–43. doi: 10.1016/j.cub.2016.04.011
93. Gopalakrishnan S and Kannan NN. Only time will tell: the interplay between circadian clock and metabolism. Chronobiol Int. (2021) 38:149–67. doi: 10.1080/07420528.2020.1842436
94. Borrmann H and Rijo-Ferreira F. Crosstalk between circadian clocks and pathogen niche. PloS Pathog. (2024) 20:e1012157. doi: 10.1371/journal.ppat.1012157
95. Hergenhan S, Holtkamp S, and Scheiermann C. Molecular interactions between components of the circadian clock and the immune system. J Mol Biol. (2020) 432:3700–13. doi: 10.1016/j.jmb.2019.12.044
96. Kim P, Oster H, Lehnert H, Schmid SM, Salamat N, Barclay JL, et al. Coupling the circadian clock to homeostasis: the role of period in timing physiology. Endocr Rev. (2019) 40:66–95. doi: 10.1210/er.2018-00049
97. Walton JC, Bumgarner JR, and Nelson RJ. Sex differences in circadian rhythms. Cold Spring Harb Perspect Biol. (2022) 14(7):a039107. doi: 10.1101/cshperspect.a039107
98. Lok R, Qian J, and Chellappa SL. Sex differences in sleep, circadian rhythms, and metabolism: Implications for precision medicine. Sleep Med Rev. (2024) 75:101926. doi: 10.1016/j.smrv.2024.101926
99. Bailey M and Silver R. Sex differences in circadian timing systems: implications for disease. Front Neuroendocrinol. (2014) 35:111–39. doi: 10.1016/j.yfrne.2013.11.003
100. Ratnu VS, Emami MR, and Bredy TW. Genetic and epigenetic factors underlying sex differences in the regulation of gene expression in the brain. J Neurosci Res. (2017) 95:301–10. doi: 10.1002/jnr.23886
101. Hatcher KM, Royston SE, and Mahoney MM. Modulation of circadian rhythms through estrogen receptor signaling. Eur J Neurosci. (2020) 51:217–28. doi: 10.1111/ejn.14184
102. Karaboué A, Innominato PF, Wreglesworth NI, Duchemann B, Adam R, and Lévi FA. Why does circadian timing of administration matter for immune checkpoint inhibitors’ efficacy? Br J Cancer. (2024) 131:783–96. doi: 10.1038/s41416-024-02704-9
103. Quist M, van Os M, van Laake LW, Bovenschen N, and Crnko S. Integration of circadian rhythms and immunotherapy for enhanced precision in brain cancer treatment. EBioMedicine. (2024) 109:105395. doi: 10.1016/j.ebiom.2024.105395
Keywords: circadian rhythms, peripheral clocks, zeitgebers, chronotherapy, metabolic disease, cardiovascular dysfunction, neurodegeneration, cancer
Citation: Bautista J, Ojeda-Mosquera S, Ordóñez-Lozada D and López-Cortés A (2025) Peripheral clocks and systemic zeitgeber interactions: from molecular mechanisms to circadian precision medicine. Front. Endocrinol. 16:1606242. doi: 10.3389/fendo.2025.1606242
Received: 08 April 2025; Accepted: 13 May 2025;
Published: 29 May 2025.
Edited by:
Oliana Carnevali, Marche Polytechnic University, ItalyReviewed by:
Cristiano Bertolucci, University of Ferrara, ItalyCopyright © 2025 Bautista, Ojeda-Mosquera, Ordóñez-Lozada and López-Cortés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés López-Cortés, YWFsYzg0QGdtYWlsLmNvbQ==