 Sara Sobreviela Sánchez
Sara Sobreviela Sánchez Ravi Giusfredi Quevedo
Ravi Giusfredi Quevedo Tiago Fernandes
Tiago Fernandes Guilherme Wesley Peixoto da Fonseca
Guilherme Wesley Peixoto da Fonseca- School of Physical Education and Sport, University of São Paulo, São Paulo, SP, Brazil
Obesity is recognized as a systemic disease characterized by chronic, low-grade inflammation. The persistent inflammatory state can be driven by secretion of cytokines from adipose tissue, also known as adipokines. In patients with obesity, adipose tissue releases elevated levels of pro-inflammatory cytokines that can have an autocrine and paracrine function. The main cytokines involved in this process include tumor necrosis factor alpha (TNF-α), interleukin-6, and interleukin-1 beta (IL-1β). These molecules actively contribute to metabolic dysregulation by interfering with insulin signaling pathways and facilitating the infiltration of immune cells into adipose tissue. Thus, a vicious cycle can be established in which inflammation perpetuates metabolic disturbances, increasing the risk of developing cardiovascular disease, type II diabetes mellitus, and other chronic conditions. On the other hand, physical exercise can release myokines with anti-inflammatory properties, such as interleukin-6 and irisin, which can positively modulate immune response. Regular physical activity and healthy eating patterns emerge as essential tools to counteract low-grade inflammation. A diet rich in bioactive compounds, such as antioxidants and polyunsaturated fatty acids, may also regulate cytokine expression, reinforcing the role of nutrition as a therapeutic strategy for obesity management. In conclusion, the role of inflammatory cytokines in obesity is central and managing their activity through non-pharmacological interventions, combining exercise and nutrition, represents a powerful tool to prevent long-term complications. However, more studies are needed to elucidate the exact molecular mechanisms by which nutrition and exercise modulate inflammation in obesity, in order to develop more effective interventions.
Introduction
Adipose tissue was historically considered a passive reservoir for energy storage; however, it is now recognized as an active endocrine organ. Adipsin (1), one of the first adipokines discovered, was shown to modulate the immune system and energy metabolism, suggesting that the secretory capacity of adipose tissue could influence human health (2). The regulation and signaling of adipose tissue, as an endocrine organ, has recently gained significant attention for the management of obesity, a condition marked by chronic low-grade inflammation and altered adipokine secretion.
Obesity is a condition characterized by excessive accumulation of adipose tissue, affecting approximately 16% of the global adult population as of 2022, according to World Health Organization (WHO) data (3). The risk associated with obesity may be related to the function of adipose tissue via releasing pro-inflammatory cytokines. Moreover, secretion of several pro-inflammatory cytokines contributes to the development of obesity-associated comorbidities, such as insulin resistance, metabolic syndrome, cardiovascular diseases, and sarcopenia (4).
Regular physical activity has been proposed as an effective strategy to mitigate these adverse effects of obesity. Exercise training, beyond promoting weight loss, stimulates the release of myokines by skeletal muscle, such as interleukin-6 (IL-6), which engage in bidirectional crosstalk with adipose tissue and other organs modulating systemic inflammation (5, 6). Thus, physical exercise is an important tool to effectively manage obesity in clinical practice.
On the other hand, several nutrients, when incorporated into the diet, have been shown to modulate inflammation, including omega-3 fatty acids, polyphenols, and dietary fiber, which modulate immune responses through gut microbiota and inflammatory pathways (7). Dietary patterns can also provide health benefits, as reported with the Mediterranean diet (8). Furthermore, these benefits can be particularly important for obesity management, as obese patients may present elevated levels of inflammation, increasing the risk of metabolic diseases (9). However, few studies integrate exercise and nutritional interventions to modulate inflammatory cytokines activity.
Therefore, this narrative review aims to examine the interplay between cytokines, physical exercise, and nutrition in the context of obesity, with a focus on their mechanistic roles in modulating inflammation and their practical implications for clinical management. For this purpose, a comprehensive literature search was conducted using PubMed, covering publications from 1995 to 2025.
Obesity and inflammation
Obesity is defined as the excessive accumulation of adipose tissue, identified in patients with a body mass index (BMI) equal or higher than 30 kg/m2 (10). Although BMI can be an easy measurement to acquire in clinical practice using only body weight and height, BMI lacks sensitivity for detecting excess adiposity, particularly in individuals with high muscle mass (11). However, despite normal BMI, patients may have excessive adipose tissue, a phenomenon known as normal weight obesity, and yet present an increased risk of obesity-related comorbidities (12). Thus, assessing body composition, especially percentage of fat mass, can assist in analyzing health risks related to obesity. Moreover, obesity is not always accompanied by other comorbidities. Preclinical obesity refers to an early stage of obesity where mild metabolic disturbance can occur, whereas clinical obesity can be related to organ dysfunction (13).
Adipose tissue is an active endocrine organ that secretes adipokine, cytokines and hormone-like substances involved in metabolic and inflammatory regulation (14). The transition from lean to obese is accompanied by a chronic low-grade inflammation and immune dysregulation, as well as enhanced release of pro-inflammatory cytokines, which can consequently promote obesity-induced comorbidities. Moreover, pro-inflammatory cytokines can contribute to skeletal muscle disorders, such as sarcopenia (15), presenting similar elevated inflammatory markers as obesity, like interleukin-1β (IL-1β), IL-6 and tumor necrosis factor alpha (TNF-α) (16). Obesity can also lead to the infiltration of immune cells, primarily macrophages, into skeletal muscle, reducing muscle density and impairing muscle function (17).
Regulation of gut microbiota has been proposed as an additional mechanism through which obesity can promote low-grade inflammation (18). The microbiota of patients with obesity seems to exhibit specific characteristics related to impaired diversity and microbial composition, differing from individuals with normal weight (19). In addition, there may be differences in the microbiota depending on associated-comorbidities in patients with obesity, suggesting an interplay between metabolic disorders and obesity meditated by gut microbiota (20). It appears that the microbiota of obese patients may provide more energy to the host, possibly through enhanced fermentation of dietary substrates and altered production of short-chain fatty acids (SCFAs), which influence energy balance and appetite regulation (21).
Low-grade inflammation has been widely associated with disruptions in glucose metabolism, commonly reported in patients with obesity and type II diabetes mellitus (22). Additionally, inflammation can increase the risk of cardiovascular diseases, especially when other risk factors coexist (23). Therefore, reducing low-grade inflammation caused by excessive adipose tissue accumulation can be a strategy to manage obesity. In this context, specific pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 have emerged as key players in the pathophysiology of obesity and its complications.
Tumor necrosis factor alpha
Tumor necrosis factor alpha (TNF-α), a pro-inflammatory cytokine secreted primarily by macrophages, has been associated with the pathogenesis of autoimmune diseases (24). TNF-α plays a central role in mediating inflammation by promoting apoptosis of damaged cells and increasing oxidative stress, contributing to metabolic dysfunction when chronically elevated (25). Glucose metabolism is also impaired with elevated TNF-α causing insulin resistance (26). Increased level of TNF-α has shown to impair intracellular insulin receptor signaling in adipocytes, hepatocytes, and skeletal muscle cells by inhibiting insulin receptor substrate-1 (IRS-1) (27). Additionally, it has been reported that the administration of TNF-α in human adipocytes reduces mRNA expression of glucose transporter type 4 (GLUT-4) (28). GLUT-4, a member of the glucose transporter family, translocates to the cell membrane and allows influx of monosaccharide into the cell cytoplasm, showing that the regulation of insulin signaling pathway can be mediated by inflammation promoted by high levels of adipose tissue (29).
Excessive release of TNF-α can also affect lipid metabolism. Patients with dyslipidemia have shown to have higher levels of this cytokine, which is associated with increased concentrations of triglycerides (TG) and low-density lipoprotein (LDL) (30). TNF-α can also inhibit the activity of lipoprotein lipase (LPL) in adipose tissue, an enzyme responsible for hydrolysis of triacylglycerol present in chylomicrons and very low-density lipoprotein (VLDL) (28). Although anti-TNF-α antibody therapy may play a role in obesity-related comorbidities, the inflammatory response observed in obesity is more likely a consequence of excess adiposity rather than its primary cause (31). On the other hand, if TNF-α inhibitors may cause an increase in body weight by impairing lipolysis, blocking TNF-α signaling could become a new strategy to treat skeletal muscle disorders with severely reduced weight loss, such as cancer cachexia (32).
Elevated TNF-α levels have been also associated with endothelial dysfunction and increased arterial stiffness, both key contributors to hypertension and atherosclerosis (33). Endothelin-1 can be involved in vasoconstriction of vascular smooth muscle cells, reducing the lumen of blood vessels that may lead chronically to persistent increases in blood pressure. These changes in vascular response through elevated secretion of TNF-α may become more severe with aging (34).
Exercise has shown to improve TNF-α levels in mice in a high-fat diet, suggesting that exercise may be a strong regulators of TNF-α (35). Several types of exercise have shown to modulate TNF-α levels, such as light-intensity walking (36), even though other studies have not demonstrated significant impact of exercise on reducing TNF-α (37, 38). These conflicting results may be related to the maintenance or transitory increase in pro-inflammatory cytokines in response to muscle damage caused by intense and prolonged bout of exercise (39, 40).
Nutritional strategies also play a crucial role in modulating TNF-α. The consumption of natural soy products seems to have positive effects on reducing plasma levels of TNF-α (41), which may be due to the effect of isoflavones as antioxidants (42). The combination of omega-3 fatty acids with curcumin supplementation has shown to reduce TNF-α expression and serum levels in patients with migraines (43). In patients with sarcopenia, protein supplementation, whether soy or whey protein, was also capable of decreasing serum TNF-α (Table 1) (44). Moreover, other compounds, such as coenzyme Q10 (45), or tannins (46), also seem to have an anti-inflammatory effect and significantly reduce TNF-α levels, due to its antioxidant capacity.
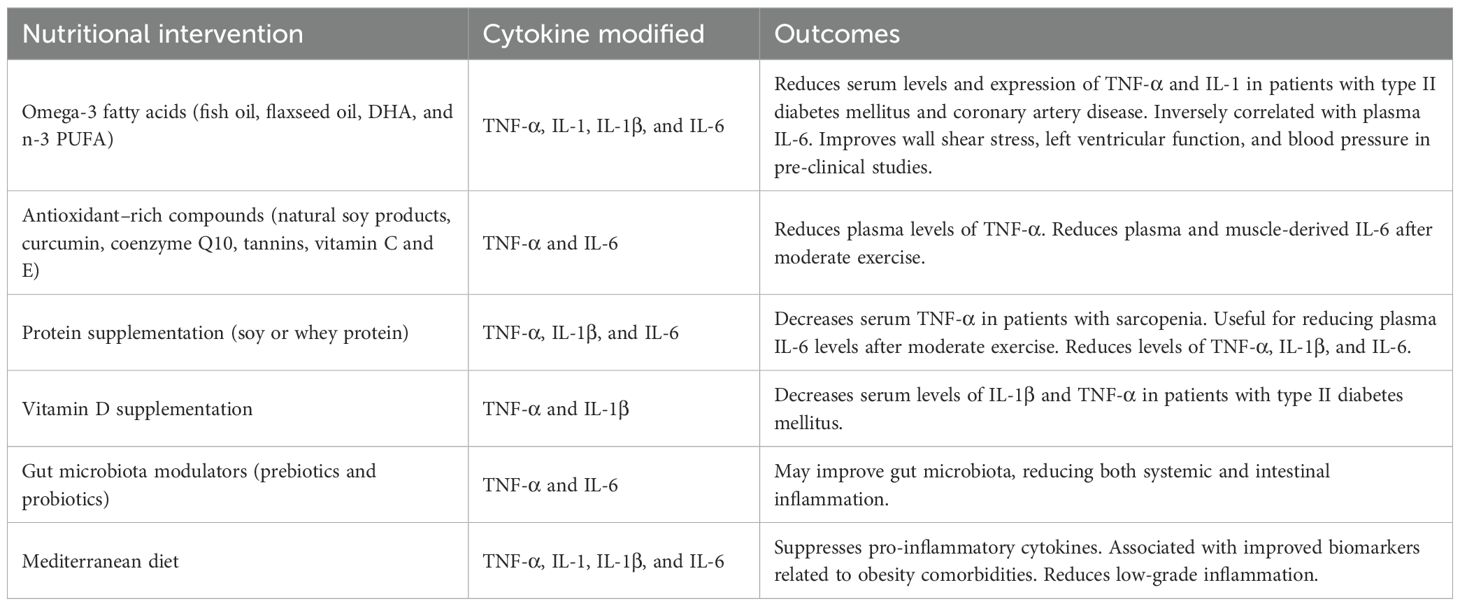
Table 1. Effect of nutritional interventions on inflammatory profile and obesity-related comorbidities.
In summary, TNF-α is a pro-inflammatory cytokine secreted by macrophages in a healthy environment, but it can also be secreted by adipose tissue in obesity, showing strong association with health issues such as insulin resistance, dyslipidemia, sarcopenia and cardiovascular conditions. Low-intensity exercise and several bioactive compounds, including omega-3 fatty acids and antioxidants, can modulate inflammation through TNF-α (Figure 1). However, more studies are necessary to determine the impact of different training variables (i.e., intensity and volume), as well as the combination of exercise with nutritional strategies, on modulating TNF-α in patients with obesity.
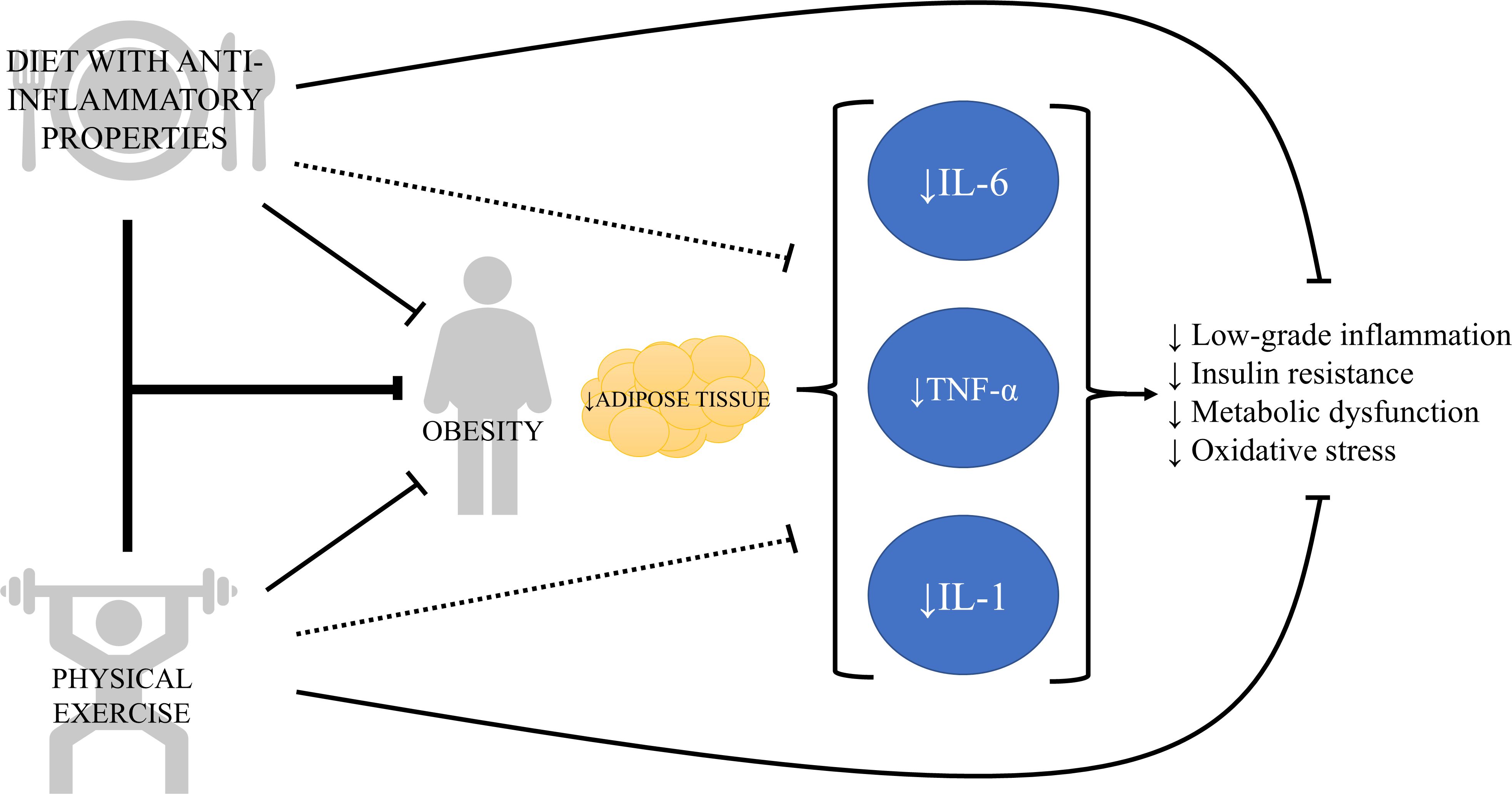
Figure 1. The vicious cycle of chronic inflammation in obesity and its modulation by lifestyle strategies. This diagram illustrates that obesity increases adipose tissue, which secretes pro-inflammatory cytokines such as interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α), and interleukin-1 (IL-1). These cytokines, in turn, contribute to insulin resistance, metabolic dysfunction, oxidative stress, and tissue damage, which physical exercise and diet with anti-inflammatory components can work synergistically or independently to counteract the chronic inflammation and obesity-related complications.
Interleukin-1
The interleukin-1 family comprises 11 cytokines that play a central role in regulating immune and inflammatory responses. Interleukin-1 (IL-1) is a pro-inflammatory cytokine essential for regulating immune response against infections. IL-1α and IL-1β are the most studied members as they possess strong pro-inflammatory effects. IL-1 works via innate immune response activating macrophages and neutrophils, while can enhance activation of T and B cells in adaptive immune response (47). Moreover, IL-1 induces local histamine release from mast cells, stimulating early vasodilation and increasing vascular permeability (48). IL-1 has shown to act synergistically with TNF-α through promoting cell death (49). As TNF-α, IL-1 can also be secreted by adipose tissue under pathological conditions.
Glucose metabolism is also affected by IL-1. It has been reported that blocking IL-1 can preserve pancreatic cell mass and function, while IL-1β can promote apoptosis of pancreatic beta cells in hyperglycemic conditions (50). The risk of atherosclerosis may also be increased in the presence of IL-1, since IL-1 is involved in vascular wall inflammation, via activation of monocytes and expression of adhesion molecules (51). Additionally, IL-1 in adipose tissue can promote macrophage infiltration and release of other pro-inflammatory cytokines, which creates a vicious cycle in the progression of obesity (52).
Interleukin-18 (IL-18), another pro-inflammatory cytokine belonging to the IL-1 family, can be considered essential in the inflammatory state of patients with obesity. Plasma levels of IL-18 are positively correlated with insulin resistance and may be related to the development of type II diabetes mellitus (53). Moreover, the contribution of IL-18 to chronic inflammation can be associated with facilitating formation of atherosclerotic plaques and endothelial dysfunction that can promote the development of cardiovascular diseases (54).
Interestingly, physical exercise can be a key strategy when it comes to reducing IL-1 family cytokine levels (Figure 1). It has been shown that physical exercise reduces IL-1β gene expression in patients with heart failure (55), which in turn may improve exercise capacity of these patients (56). In experimental studies with diabetic rats, swimming significantly reduced the expression of inflammatory cytokines, such as IL-1β (57). However, as of TNF-α, plasma levels of IL-1 may increase after intense exercise as a physiological response to strenuous muscle effort (58), making it difficult to assess the isolated impact of physical activity on IL-1.
Vitamin D and calcium supplementation through a drinkable yogurt has been associated with a decrease in serum levels of IL-1β and TNF-α in patients with type II diabetes mellitus (59). In addition, when supplemented only calcium no significant effects on inflammation were reported, suggesting that the anti-inflammatory effect may be specific to vitamin D (Table 1). Likewise, supplementation with flaxseed oil, a source of linolenic acid, significantly improved the expression levels of IL-1 and TNF-α in patients with diabetes and coronary artery disease (Table 1) (60). Dietary intake of docosahexaenoic acid (DHA) in mice has been shown to reduce wall shear stress, improve left ventricular function, and lower blood pressure compared to controls, suggesting that reduced local IL-1β expression can mediate these adaptations (61). These last two studies reaffirm the anti-inflammatory capabilities of omega-3 fatty acids and their benefits on obesity-related comorbidities.
The IL-1 family plays a critical role in regulating both innate and adaptive immune responses. Despite triggering low-grade inflammation, IL-1 has been linked to obesity-related comorbidities such as atherosclerosis and endothelial dysfunction. Supplementation of omega-3 fatty acids stands again as clinical strategy to modulate inflammation, while low-intensity exercise can modulate several isoforms of IL-1. Nonetheless, more studies are needed to understand the role of IL-1 family in the context of exercise and nutritional interventions in patients with obesity.
Interleukin-6
Interleukin-6 (IL-6) is a cytokine with both pro-inflammatory and anti-inflammatory properties (Figure 2) (62). Under normal conditions, IL-6 is required not only for inflammatory processes, but also for hematopoiesis, bone metabolism and coagulation (63), as well as for facilitating glycemic control and providing cross-talk, linking tissues such as intestinal L cells and pancreatic islets (5).
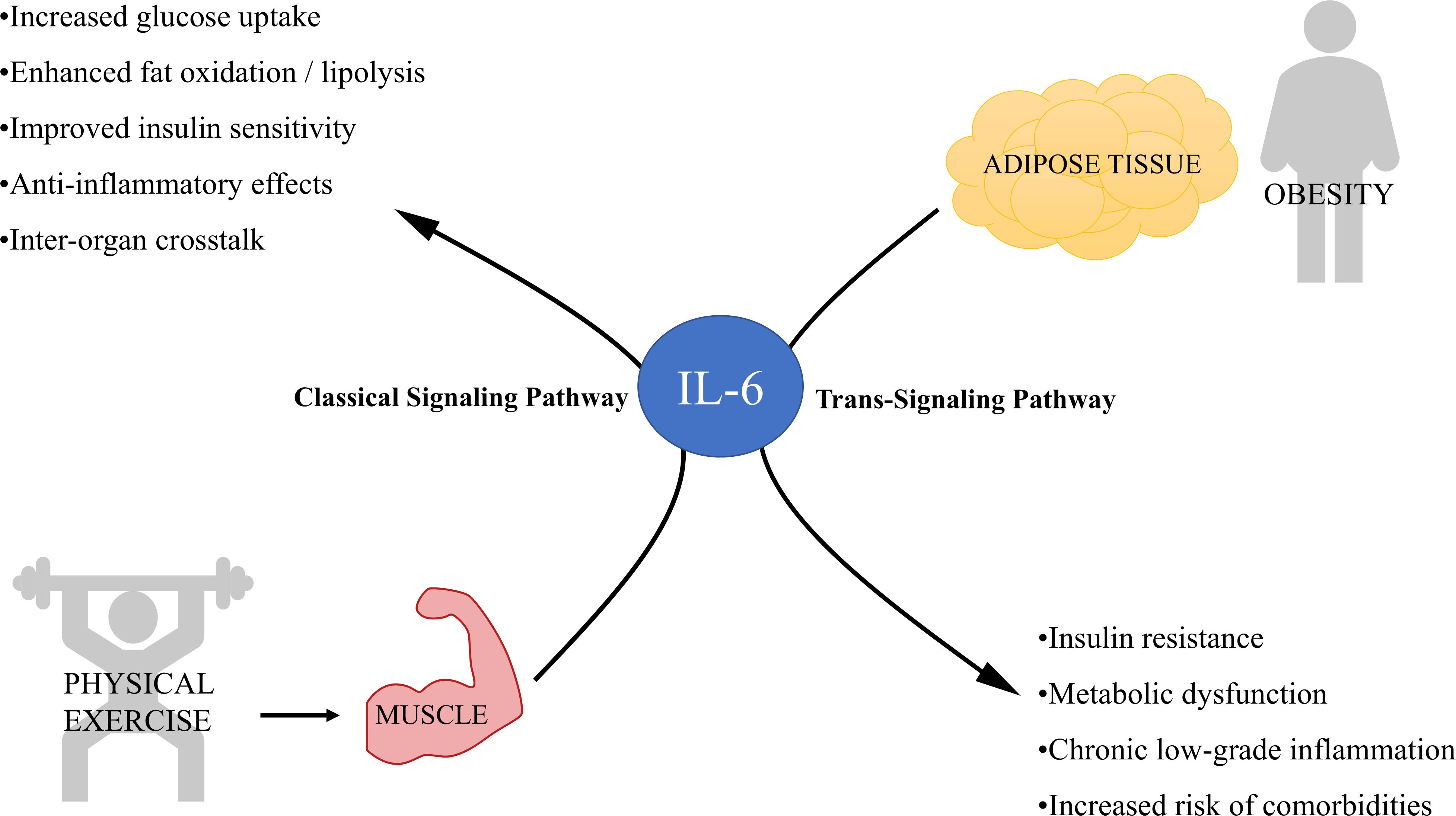
Figure 2. Dual role of interleukin-6 (IL-6) released from skeletal muscle and adipose tissue in metabolism and inflammation. This diagram illustrates the contrasting effects of IL-6 depending on its source and signaling pathway involved. On the left, physical exercise stimulates skeletal muscle to release IL-6, which primarily signals through the classical signaling pathway, producing anti-inflammatory effects. On the right, obesity leads to increased IL-6 secretion from adipose tissue, predominantly activating the trans-signaling pathway that is associated with chronic inflammation and metabolic dysregulation.
Mechanistically, two main signaling pathways can be activated by IL-6. In the classical signaling pathway, IL-6 binds to its membrane receptor (IL-6R), followed by dimerization of glycoprotein 130 (gp130) and activation of the JAK/STAT, MAPK and PI3K/AKT pathways (64). Through the trans-signaling pathway, IL-6 can also bind to molecules of the soluble IL-6 receptor (sIL-6R), which are generated through ectodomain release by metalloproteases (ADAM-10 and ADAM-17) (65). The expression levels of IL-6 receptor subunits vary among cell types and determine whether IL-6 classical signaling pathway, which primarily induces anti-inflammatory responses, or IL-6 trans-signaling pathway, generally associated with pro-inflammatory responses, is triggered (66). Moreover, the presence of other cytokines may modulate IL-6 response (67).
As mentioned before, low-grade inflammation in obese patients impacts negatively in insulin signaling pathways and causes insulin resistance, with IL-6 playing an important role in this process (68). In pre-clinical studies, IL-6 knockout mice developed obesity, associated with a disturbed carbohydrate and lipid metabolism (69). Indeed, these data were subsequently supported indicating increased body weight, impaired glucose tolerance and exacerbated insulin resistance in IL-6 knockout mice (70). In regards to lipid metabolism, IL-6 has been linked to the development of dyslipidemia and a massive mobilization of fatty acids that may lead to lipid accumulation in the myocardium, potentially causing cardiac lipotoxicity (71). In addition, IL-6 has been associated with increase risk of several types of cancer, including breast, liver, and colon cancer (72).
However, IL-6 can be somewhat paradoxical, as both harmful and beneficial effects have been reported. For instance, muscle-derived IL-6 has beneficial effects unlike that secreted during inflammation, despite being the same molecule. One explanation for this paradox lies in the duration of IL-6 exposure. Although chronic elevations are associated with insulin resistance, acute elevations may enhance insulin sensitivity (73). IL-6 also has regenerative, anti-inflammatory and anti-diabetogenic functions, when secreted as myokine by skeletal muscles during physical exercise. IL-6 released from skeletal muscle during exercise has been associated with improved glucose uptake and fat oxidation (74), suggesting that IL-6 has a crucial role in energy mobilization, even though IL-6 release from adipose tissue may not have the same effect (Figure 2).
Muscle contraction has been linked to an increase in plasma IL-6 levels, which is related to exercise intensity (75). IL-6 is one of the pathways through which physical activity improves insulin sensitivity, as it enhances GLUT-4 expression (76). Moreover, IL-6 can stimulate lipolysis and fat oxidation in humans without causing hypertriglyceridemia, which can have significant benefits for patients with metabolic syndrome (77). However, administering IL-6 exogenously as a mimetic for physical exercise is not an appropriate strategy, since elevated blood levels of IL-6 are still associated with inflammation (78), despite anti-inflammatory properties of muscle-derived IL-6.
The concentration of IL-6 has been inversely associated with 25-OH-D levels in older adults (79), suggesting that maintaining adequate 25-OH-D levels may reduce inflammation. A cohort study of over 5000 women and men found that n-3 PUFA intake was inversely correlated with plasma IL-6 levels (80). Two experimental studies examined the effect of whey protein or vitamin C and E supplementation on post-exercise inflammation. Both strategies were found to be useful in reducing plasma and muscle derived IL-6 levels, respectively, after moderate exercise (81, 82). Therefore, these results may be primarily due to a reduction in muscle rather than systemic inflammation.
IL-6 may present pro-inflammatory properties when derived from adipose tissue, but also anti-inflammatory effect derived from skeletal muscle. More studies are necessary to understand this paradoxal relationship of IL-6 and its impact on obesity management.
System impact of physical exercise on inflammation
Physical exercise can be extremely helpful in managing inflammation and comorbidities associated with obesity. Exercise can significantly alleviate type II diabetes mellitus, as it promotes insulin sensitivity through various pathways, such as the reduction of plasma ceramides (83), enhancement of pancreatic β-cell function (84), and increased skeletal muscle capillarization (85). Exercise also improves lipid profile and cardiovascular health (86). Moreover, like adipose tissue, muscle tissue also acts as a secretory organ, releasing myokines to counteract inflammation and obesity-related comorbidities. Aerobic exercise can significantly decrease the level of inflammatory cytokines, when performed from moderate to high intensity with session lasting between 30 to 60 minutes in a frequency of 2 to 3 times per week (87).
Muscle-derived IL-6 can inhibit the production of inflammatory cytokines such as TNF-α and IL-1β, while promoting secretion of anti-inflammatory cytokines like interleukin-10 (IL-10) (88), thereby improving inflammatory profile in patients with sarcopenic obesity. The improved profile can attenuate age-related inflammation (inflammaging) and, consequently, improve glucose metabolism in skeletal muscle (89), as well as suppressing activation of macrophages, IL-2, and interferon-gamma (IFN-γ) (88).
Other myokine, such as irisin, are also associated with improvement of obesity-related conditions. Irisin is secreted by muscle and can induce white adipose tissue browning, which regulates energy metabolism (90). In addition, irisin has also been linked to beneficial effects on inflammation, neurodegeneration, oxidative stress, diabetes mellitus, and sarcopenia (91, 92). Strength training can play a role in activating mTOR and AMPK pathways for the secretion of these myokines (93). Moreover, physical exercise can also modify the composition of the gut microbiota that may improve systemic inflammation (94). Therefore, through several mechanisms, exercise can be used as an effective intervention to improve chronic inflammation in patients with obesity (95).
Systemic impact of nutrition on inflammation
Dietary patterns are also essential for managing chronic low-grade inflammation in patients with obesity. It has been reported that high-fat diets, also described as Western diet, can be correlated with increased inflammatory profile and gut dysbiosis (96). These high-fat diets are mainly composed of ultra-processed foods, which appear to promote a chronic pro-inflammatory state (97), whilst diets with a higher content of whole, natural foods may reduce low-grade inflammation in obesity, placing particular emphasis on omega-3 polyunsaturated fatty acids, vitamins from vegetables and fruits (98), and dietary fiber (99). Providing prebiotics along with probiotics could also improve gut microbiota, reducing both systemic and intestinal inflammation (100). Other alternative supplements include branched-chain amino acids, calcium, and vitamin D3, which may reduce levels of TNF-α, IL-1β, and IL-6 (101).
The Mediterranean diet has been shown to improve several aspects for the management and prevention of obesity (102). The Mediterranean diet is characterized by increased consumption of fruits, vegetables, legumes, whole grains, olive oil, moderate intake of fish and dairy, and low intake of red meat. Moreover, the Mediterranean diet has been associated with improved biomarkers related to obesity comorbidities, such as glucose metabolism, plasma lipids, and cardiovascular diseases (103, 104). Therefore, following dietary patterns like the Mediterranean diet is currently recommended for reducing low-grade inflammation associated with obesity and sarcopenia (105, 106), as many of its components individually exhibit anti-inflammatory effects.
Future perspective
This narrative review reinforces the view that obesity is a complex, multifactorial disease, characterized not only by excessive fat accumulation but also by chronic low-grade inflammation mediated by dysregulated cytokine secretion. Additionally, pro-inflammatory mediators such as IL-6, IL-1, and TNF-α contribute to the pathogenesis of obesity-related comorbidities including insulin resistance, cardiovascular disease, sarcopenia, and dyslipidemia.
Both physical exercise and evidence-based nutritional strategies can improve the inflammatory profile of patients with obesity, and consequently reduce the incidence of comorbidities. Exercise-induced myokines such as IL-6, IL-10, and irisin promote anti-inflammatory effects and improve metabolic outcomes. Moreover, certain dietary patterns, such as the Mediterranean diet and increased intake of nutrients with anti-inflammatory properties, including omega-3 fatty acids, polyphenols and antioxidants, can significantly reduce the secretion of pro-inflammatory cytokines and improve the metabolic profile of patients with obesity. Nevertheless, further research is needed to confirm the combined or isolated effect of physical exercise and nutrition on the management of obesity, especially longitudinal studies. Moreover, an important limitation of this review is the heterogeneity among studies, varying in outcomes assessed, intervention protocols, study design and duration.
In conclusion, addressing inflammation as a key component of obesity provides a more comprehensive approach to the treatment. Strategies combining regular physical exercise with appropriate nutritional interventions may offer a powerful tool to reduce inflammatory complications caused by obesity and obesity-related comorbidities. Investigations incorporating molecular biomarkers, microbiota profiling and individualized responses to interventions will be crucial for the development of precision therapies. Ultimately, treating inflammation as a core component of obesity may enhance current management strategies and improve long-term patient outcomes.
Author contributions
SSS: Writing – original draft, Formal analysis, Methodology, Visualization. RGQ: Writing – review & editing, Methodology, Visualization. TF: Writing – review & editing, Conceptualization, Supervision. GWPF: Writing – review & editing, Conceptualization, Supervision, Visualization, Funding aquisition, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by Pró-Reitoria de Pesquisa e Inovaçao at the University of São Paulo (Process number: 22.1.09345.01.2).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Cook KS, Min HY, Johnson D, Chaplinsky RJ, Flier JS, Hunt CR, et al. Adipsin: a circulating serine protease homolog secreted by adipose tissue and sciatic nerve. Science. (1987) 237:402–5. doi: 10.1126/science.3299705
2. Fantuzzi G. Adipose tissue, adipokines, and inflammation. J Allergy Clin Immunol. (2005) 115:911–9; quiz 920. doi: 10.1016/j.jaci.2005.02.023
3. Collaborators GRF. Global burden and strength of evidence for 88 risk factors in 204 countries and 811 subnational locations, 1990-2021: a systematic analysis for the Global Burden of Disease Study 2021. Lancet. (2024) 403:2162–203. doi: 10.1016/S0140-6736(24)00933-4
4. Colleluori G and Villareal DT. Aging, obesity, sarcopenia and the effect of diet and exercise intervention. Exp Gerontol. (2021) 155:111561. doi: 10.1016/j.exger.2021.111561
5. Pedersen BK and Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat Rev Endocrinol. (2012) 8:457–65. doi: 10.1038/nrendo.2012.49
6. Pedersen BK and Febbraio MA. Muscle as an endocrine organ: focus on muscle-derived interleukin-6. Physiol Rev. (2008) 88:1379–406. doi: 10.1152/physrev.90100.2007
7. Calder PC. Omega-3 fatty acids and inflammatory processes: from molecules to man. Biochem Soc Trans. (2017) 45:1105–15. doi: 10.1042/BST20160474
8. Estruch R, Ros E, Salas-Salvadó J, Covas MI, Corella D, Arós F, et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N Engl J Med. (2018) 378:e34. doi: 10.1056/NEJMoa1800389
9. Lumeng CN and Saltiel AR. Inflammatory links between obesity and metabolic disease. J Clin Invest Jun. (2011) 121:2111–7. doi: 10.1172/JCI57132
10. Gaskin CJ, Cooper K, Stephens LD, Peeters A, Salmon J, and Porter J. Clinical practice guidelines for the management of overweight and obesity published internationally: A scoping review. Obes Rev May. (2024) 25:e13700. doi: 10.1111/obr.13700
11. Baile JI. Is it useful to use the body mass index to assess obesity in muscular people]? Nutr Hosp. (2015) 32:2353. doi: 10.3305/nh.2015.32.5.9598
12. Oliveros E, Somers VK, Sochor O, Goel K, and Lopez-Jimenez F. The concept of normal weight obesity. Prog Cardiovasc Dis. (2014) 56:426–33. doi: 10.1016/j.pcad.2013.10.003
13. Rubino F, Cummings DE, Eckel RH, Cohen RV, Wilding JPH, Brown WA, et al. Definition and diagnostic criteria of clinical obesity. Lancet Diabetes Endocrinol. (2025) 13:221–62. doi: 10.1016/S2213-8587(24)00316-4
14. Wang T and He C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. (2018) 44:38–50. doi: 10.1016/j.cytogfr.2018.10.002
15. Liang Z, Zhang T, Liu H, Li Z, Peng L, Wang C, et al. Inflammaging: The ground for sarcopenia? Exp Gerontol. (2022) 168:111931. doi: 10.1016/j.exger.2022.111931
16. Jimenez-Gutierrez GE, Martínez-Gómez LE, Martínez-Armenta C, Pineda C, Martínez-Nava GA, and Lopez-Reyes A. Molecular Mechanisms of Inflammation in Sarcopenia: Diagnosis and Therapeutic Update. Cells. (2022) 11:2359. doi: 10.3390/cells11152359
17. Lee YS, Wollam J, and Olefsky JM. An Integrated View of Immunometabolism. Cell. (2018) 172:22–40. doi: 10.1016/j.cell.2017.12.025
18. Vetrani C, Di Nisio A, Paschou SA, Barrea L, Muscogiuri G, Graziadio C, et al. From Gut Microbiota through Low-Grade Inflammation to Obesity: Key Players and Potential Targets. Nutrients. (2022) 14:2103. doi: 10.3390/nu14102103
19. Schwiertz A, Taras D, Schäfer K, Beijer S, Bos NA, Donus C, et al. Microbiota and SCFA in lean and overweight healthy subjects. Obes (Silver Spring). (2010) 18:190–5. doi: 10.1038/oby.2009.167
20. Pitocco D, Di Leo M, Tartaglione L, De Leva F, Petruzziello C, Saviano A, et al. The role of gut microbiota in mediating obesity and diabetes mellitus. Eur Rev Med Pharmacol Sci. (2020) 24:1548–62. doi: 10.26355/eurrev_202002_20213
21. Asadi A, Shadab Mehr N, Mohamadi MH, Shokri F, Heidary M, Sadeghifard N, et al. Obesity and gut-microbiota-brain axis: A narrative review. J Clin Lab Anal. (2022) 36:e24420. doi: 10.1002/jcla.24420
22. Gregor MF and Hotamisligil GS. Inflammatory mechanisms in obesity. Annu Rev Immunol. (2011) 29:415–45. doi: 10.1146/annurev-immunol-031210-101322
23. Elías-López D, Doi T, Nordestgaard BG, and Kobylecki CJ. Remnant cholesterol and low-grade inflammation jointly in atherosclerotic cardiovascular disease: implications for clinical trials. Curr Opin Clin Nutr Metab Care. (2024) 27:125–35. doi: 10.1097/MCO.0000000000000999
24. Jang DI, Lee AH, Shin HY, Song HR, Park JH, Kang TB, et al. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int J Mol Sci. (2021) 22:2719. doi: 10.3390/ijms22052719
25. Gul A and Yilmaz R. Determination of inflammation by TNF-alpha and IL-10 levels in obese children and adolescents. Nutr Hosp. (2024) 41:788–92. doi: 10.20960/nh.05064
26. Hotamisligil GS. Mechanisms of TNF-alpha-induced insulin resistance. Exp Clin Endocrinol Diabetes. (1999) 107:119–25. doi: 10.1055/s-0029-1212086
27. Ramírez Alvarado MM and Sánchez Roitz C. Tumor necrosis factor-α, insulin resistance, the lipoprotein metabolism and obesity in humans. Nutr Hosp. (2012) 27:1751–7. doi: 10.3305/nh.2012.27.6.6004
28. Hauner H, Petruschke T, Russ M, Röhrig K, and Eckel J. Effects of tumour necrosis factor alpha (TNF alpha) on glucose transport and lipid metabolism of newly-differentiated human fat cells in cell culture. Diabetologia. (1995) 38:764–71. doi: 10.1007/s001250050350
29. Wang T, Wang J, Hu X, Huang XJ, and Chen GX. Current understanding of glucose transporter 4 expression and functional mechanisms. World J Biol Chem. (2020) 11:76–98. doi: 10.4331/wjbc.v11.i3.76
30. Jovinge S, Hamsten A, Tornvall P, Proudler A, Båvenholm P, Ericsson CG, et al. Evidence for a role of tumor necrosis factor alpha in disturbances of triglyceride and glucose metabolism predisposing to coronary heart disease. Metabolism. (1998) 47:113–8. doi: 10.1016/s0026-0495(98)90203-7
31. Peluso I and Palmery M. The relationship between body weight and inflammation: Lesson from anti-TNF-α antibody therapy. Hum Immunol. (2016) 77:47–53. doi: 10.1016/j.humimm.2015.10.008
32. Patsalos O, Dalton B, Leppanen J, Ibrahim MAA, and Himmerich H. Impact of TNF-α Inhibitors on Body Weight and BMI: A Systematic Review and Meta-Analysis. Front Pharmacol. (2020) 11:481. doi: 10.3389/fphar.2020.00481
33. Hu H, Huang J, Zhang S, Zhang B, Li W, and Sun K. Tumor necrosis factor-α stimulation endothelial-to-mesenchymal transition during cardiac fibrosis via endothelin-1 signaling. J Biochem Mol Toxicol. (2023) 37:e23411. doi: 10.1002/jbt.23411
34. Bruunsgaard H, Ladelund S, Pedersen AN, Schroll M, Jørgensen T, and Pedersen BK. Predicting death from tumour necrosis factor-alpha and interleukin-6 in 80-year-old people. Clin Exp Immunol Apr. (2003) 132:24–31. doi: 10.1046/j.1365-2249.2003.02137.x
35. Feng L, Huang F, Ma Y, and Tang J. The Effect of High-Fat Diet and Exercise Intervention on the TNF-α Level in Rat Spleen. Front Immunol. (2021) 12:671167. doi: 10.3389/fimmu.2021.671167
36. Safdar NZ, Alobaid AM, Hopkins M, Dempsey PC, Pearson SM, Kietsiriroje N, et al. Short, frequent, light-intensity walking activity improves postprandial vascular-inflammatory biomarkers in people with type 1 diabetes: The SIT-LESS randomized controlled trial. Diabetes Obes Metab Jun. (2024) 26:2439–45. doi: 10.1111/dom.15564
37. Libardi CA, De Souza GV, Cavaglieri CR, Madruga VA, and Chacon-Mikahil MP. Effect of resistance, endurance, and concurrent training on TNF-α, IL-6, and CRP. Med Sci Sports Exerc Jan. (2012) 44:50–6. doi: 10.1249/MSS.0b013e318229d2e9
38. Chen HT, Wu HJ, Chen YJ, Ho SY, and Chung YC. Effects of 8-week kettlebell training on body composition, muscle strength, pulmonary function, and chronic low-grade inflammation in elderly women with sarcopenia. Exp Gerontol. (2018) 112:112–8. doi: 10.1016/j.exger.2018.09.015
39. Bernecker C, Scherr J, Schinner S, Braun S, Scherbaum WA, and Halle M. Evidence for an exercise induced increase of TNF-α and IL-6 in marathon runners. Scand J Med Sci Sports. (2013) 23:207–14. doi: 10.1111/j.1600-0838.2011.01372.x
40. Reihmane D, Jurka A, Tretjakovs P, and Dela F. Increase in IL-6, TNF-α, and MMP-9, but not sICAM-1, concentrations depends on exercise duration. Eur J Appl Physiol. (2013) 113:851–8. doi: 10.1007/s00421-012-2491-9
41. Khodarahmi M, Foroumandi E, and Asghari Jafarabadi M. Effects of soy intake on circulating levels of TNF-α and interleukin-6: a systematic review and meta-analysis of randomized controlled trials. Eur J Nutr. (2021) 60:581–601. doi: 10.1007/s00394-020-02458-z
42. Patel RP, Boersma BJ, Crawford JH, Hogg N, Kirk M, Kalyanaraman B, et al. Antioxidant mechanisms of isoflavones in lipid systems: paradoxical effects of peroxyl radical scavenging. Free Radic Biol Med. (2001) 31:1570–81. doi: 10.1016/s0891-5849(01)00737-7
43. Abdolahi M, Tafakhori A, Togha M, Okhovat AA, Siassi F, Eshraghian MR, et al. The synergistic effects of ω-3 fatty acids and nano-curcumin supplementation on tumor necrosis factor (TNF)-α gene expression and serum level in migraine patients. Immunogenetics. (2017) 69:371–8. doi: 10.1007/s00251-017-0992-8
44. Prokopidis K, Mazidi M, Sankaranarayanan R, Tajik B, McArdle A, and Isanejad M. Effects of whey and soy protein supplementation on inflammatory cytokines in older adults: a systematic review and meta-analysis. Br J Nutr. (2023) 129:759–70. doi: 10.1017/S0007114522001787
45. Farsi F, Heshmati J, Keshtkar A, Irandoost P, Alamdari NM, Akbari A, et al. Can coenzyme Q10 supplementation effectively reduce human tumor necrosis factor-α and interleukin-6 levels in chronic inflammatory diseases? A systematic review and meta-analysis of randomized controlled trials. Pharmacol Res. (2019) 148:104290. doi: 10.1016/j.phrs.2019.104290
46. Machado KN, Paula Barbosa A, de Freitas AA, Alvarenga LF, Pádua RM, Gomes Faraco AA, et al. TNF-α inhibition, antioxidant effects and chemical analysis of extracts and fraction from Brazilian guaraná seed powder. Food Chem. (2021) 355:129563. doi: 10.1016/j.foodchem.2021.129563
47. Mantovani A, Dinarello CA, Molgora M, and Garlanda C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity. (2019) 50:778–95. doi: 10.1016/j.immuni.2019.03.012
48. Theoharides TC, Kempuraj D, Tagen M, Conti P, and Kalogeromitros D. Differential release of mast cell mediators and the pathogenesis of inflammation. Immunol Rev. (2007) 217:65–78. doi: 10.1111/j.1600-065X.2007.00519.x
49. Banerjee M and Saxena M. Interleukin-1 (IL-1) family of cytokines: role in type 2 diabetes. Clin Chim Acta. (2012) 413:1163–70. doi: 10.1016/j.cca.2012.03.021
50. Larsen CM, Faulenbach M, Vaag A, Vølund A, Ehses JA, Seifert B, et al. Interleukin-1-receptor antagonist in type 2 diabetes mellitus. N Engl J Med. (2007) 356:1517–26. doi: 10.1056/NEJMoa065213
51. Fragoso-Lona JM, Ramírez-Bello J, Cruz-Robles D, Pérez-Méndez O, de la Peña A, and Vargas-Alarcón G. Pro-inflammatory and anti-inflammatory markers in coronary artery disease and acute ischemic coronary syndrome. Arch Cardiol Mex. (2009) 79:54–62.
52. Marcos-Gómez B, Bustos M, Prieto J, Martínez JA, and Moreno-Aliaga MJ. Obesity, inflammation and insulin resistance: role of gp 130 receptor ligands. Sist Sanit Navar. (2008) 31:113–23. doi: 10.4321/s1137-66272008000300002
53. Martinez-Hervas S, Martínez-Barquero V, Nuñez Savall E, Lendínez V, Olivares L, Benito E, et al. Plasma IL-18 levels are related to insulin and are modulated by IL-18 gene polymorphisms. Clin Investig Arterioscler. (2015) 27:265–71. doi: 10.1016/j.arteri.2015.04.004
54. Dawood A, Alkafrawy N, Saleh S, Noreldin R, and Zewain S. The relationship between IL-18 and atherosclerotic cardiovascular risk in Egyptian lean women with polycystic ovary syndrome. Gynecol Endocrinol. (2018) 34:294–7. doi: 10.1080/09513590.2017.1395835
55. Butts B, Butler J, Dunbar SB, Corwin E, and Gary RA. Effects of Exercise on ASC Methylation and IL-1 Cytokines in Heart Failure. Med Sci Sports Exerc. (2018) 50:1757–66. doi: 10.1249/MSS.0000000000001641
56. Moroni F, Golino M, Carbone S, Trankle C, Del Buono MG, Talasaz A, et al. Interleukin-1 blockade in heart failure: an on-treatment and off-treatment cardiorespiratory fitness analysis. ESC Heart Fail. (2023) 10:3199–202. doi: 10.1002/ehf2.14460
57. Habibi P, Ahmadiasl N, Nourazarian A, and Yousefi H. Swimming exercise improves SIRT1, NF-κB, and IL-1β protein levels and pancreatic tissue injury in ovariectomized diabetic rats. Horm Mol Biol Clin Investig. (2022) 43:345–52. doi: 10.1515/hmbci-2021-0069
58. Shephard RJ, Rhind S, and Shek PN. Exercise and training: influences on cytotoxicity, interleukin-1, interleukin-2 and receptor structures. Int J Sports Med. (1994) 15:S154–66. doi: 10.1055/s-2007-1021131
59. Neyestani TR, Nikooyeh B, Alavi-Majd H, Shariatzadeh N, Kalayi A, Tayebinejad N, et al. Improvement of vitamin D status via daily intake of fortified yogurt drink either with or without extra calcium ameliorates systemic inflammatory biomarkers, including adipokines, in the subjects with type 2 diabetes. J Clin Endocrinol Metab. (2012) 97:2005–11. doi: 10.1210/jc.2011-3465
60. Hashemzadeh AA, Nasoohi N, Raygan F, Aghadavod E, Akbari E, Taghizadeh M, et al. Flaxseed Oil Supplementation Improve Gene Expression Levels of PPAR-γ, LP(a), IL-1 and TNF-α in Type 2 Diabetic Patients with Coronary Heart Disease. Lipids. (2017) 52:907–15. doi: 10.1007/s11745-017-4295-5
61. Alfaidi MA, Chamberlain J, Rothman A, Crossman D, Villa-Uriol MC, Hadoke P, et al. Dietary Docosahexaenoic Acid Reduces Oscillatory Wall Shear Stress, Atherosclerosis, and Hypertension, Most Likely Mediated via an IL-1-Mediated Mechanism. J Am Heart Assoc. (2018) 7:e008757. doi: 10.1161/JAHA.118.008757
62. Kang S, Narazaki M, Metwally H, and Kishimoto T. Historical overview of the interleukin-6 family cytokine. J Exp Med. (2020) 217:e20190347. doi: 10.1084/jem.20190347
63. Tanaka T, Narazaki M, and Kishimoto T. Interleukin (IL-6) Immunotherapy. Cold Spring Harb Perspect Biol. (2018) 10:a028456. doi: 10.1101/cshperspect.a028456
64. Garbers C, Hermanns HM, Schaper F, Müller-Newen G, Grötzinger J, Rose-John S, et al. Plasticity and cross-talk of interleukin 6-type cytokines. Cytokine Growth Factor Rev. (2012) 23:85–97. doi: 10.1016/j.cytogfr.2012.04.001
65. Scheller J, Chalaris A, Schmidt-Arras D, and Rose-John S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta. (2011) 1813:878–88. doi: 10.1016/j.bbamcr.2011.01.034
66. Reeh H, Rudolph N, Billing U, Christen H, Streif S, Bullinger E, et al. Response to IL-6 trans- and IL-6 classic signalling is determined by the ratio of the IL-6 receptor α to gp130 expression: fusing experimental insights and dynamic modelling. Cell Commun Signal. (2019) 17:46. doi: 10.1186/s12964-019-0356-0
67. Orange ST, Leslie J, Ross M, Mann DA, and Wackerhage H. The exercise IL-6 enigma in cancer. Trends Endocrinol Metab. (2023) 34:749–63. doi: 10.1016/j.tem.2023.08.001
68. León-Pedroza JI, González-Tapia LA, del Olmo-Gil E, Castellanos-Rodríguez D, Escobedo G, and González-Chávez A. Low-grade systemic inflammation and the development of metabolic diseases: from the molecular evidence to the clinical practice. Cir Cir. (2015) 83:543–51. doi: 10.1016/j.circir.2015.05.041
69. Wallenius V, Wallenius K, Ahrén B, Rudling M, Carlsten H, Dickson SL, et al. Interleukin-6-deficient mice develop mature-onset obesity. Nat Med Jan. (2002) 8:75–9. doi: 10.1038/nm0102-75
70. Matthews VB, Allen TL, Risis S, Chan MH, Henstridge DC, Watson N, et al. Interleukin-6-deficient mice develop hepatic inflammation and systemic insulin resistance. Diabetologia. (2010) 53:2431–41. doi: 10.1007/s00125-010-1865-y
71. Xu Y, Zhang Y, and Ye J. IL-6: A Potential Role in Cardiac Metabolic Homeostasis. Int J Mol Sci. (2018) 19:2474. doi: 10.3390/ijms19092474
72. Ghosh S and Ashcraft K. An IL-6 link between obesity and cancer. Front Biosci (Elite Ed). (2013) 5:461–78. doi: 10.2741/e628
73. Holmes AG, Mesa JL, Neill BA, Chung J, Carey AL, Steinberg GR, et al. Prolonged interleukin-6 administration enhances glucose tolerance and increases skeletal muscle PPARalpha and UCP2 expression in rats. J Endocrinol Aug. (2008) 198:367–74. doi: 10.1677/JOE-08-0113
74. Al-Khalili L, Bouzakri K, Glund S, Lönnqvist F, Koistinen HA, and Krook A. Signaling specificity of interleukin-6 action on glucose and lipid metabolism in skeletal muscle. Mol Endocrinol. (2006) 20:3364–75. doi: 10.1210/me.2005-0490
75. Ostrowski K, Schjerling P, and Pedersen BK. Physical activity and plasma interleukin-6 in humans–effect of intensity of exercise. Eur J Appl Physiol. (2000) 83:512–5. doi: 10.1007/s004210000312
76. Ikeda SI, Tamura Y, Kakehi S, Sanada H, Kawamori R, and Watada H. Exercise-induced increase in IL-6 level enhances GLUT4 expression and insulin sensitivity in mouse skeletal muscle. Biochem Biophys Res Commun. (2016) 473:947–52. doi: 10.1016/j.bbrc.2016.03.159
77. van Hall G, Steensberg A, Sacchetti M, Fischer C, Keller C, Schjerling P, et al. Interleukin-6 stimulates lipolysis and fat oxidation in humans. J Clin Endocrinol Metab. (2003) 88:3005–10. doi: 10.1210/jc.2002-021687
78. Tanaka T, Narazaki M, and Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. (2014) 6:a016295. doi: 10.1101/cshperspect.a016295
79. Laird E, McNulty H, Ward M, Hoey L, McSorley E, Wallace JM, et al. Vitamin D deficiency is associated with inflammation in older Irish adults. J Clin Endocrinol Metab. (2014) 99:1807–15. doi: 10.1210/jc.2013-3507
80. He K, Liu K, Daviglus ML, Jenny NS, Mayer-Davis E, Jiang R, et al. Associations of dietary long-chain n-3 polyunsaturated fatty acids and fish with biomarkers of inflammation and endothelial activation (from the Multi-Ethnic Study of Atherosclerosis [MESA]). Am J Cardiol. (2009) 103:1238–43. doi: 10.1016/j.amjcard.2009.01.016
81. Kerasioti E, Stagos D, Jamurtas A, Kiskini A, Koutedakis Y, Goutzourelas N, et al. Anti-inflammatory effects of a special carbohydrate-whey protein cake after exhaustive cycling in humans. Food Chem Toxicol. (2013) 61:42–6. doi: 10.1016/j.fct.2013.01.023
82. Fischer CP, Hiscock NJ, Penkowa M, Basu S, Vessby B, Kallner A, et al. Supplementation with vitamins C and E inhibits the release of interleukin-6 from contracting human skeletal muscle. J Physiol. (2004) 558:633–45. doi: 10.1113/jphysiol.2004.066779
83. Kasumov T, Solomon TP, Hwang C, Huang H, Haus JM, Zhang R, et al. Improved insulin sensitivity after exercise training is linked to reduced plasma C14:0 ceramide in obesity and type 2 diabetes. Obes (Silver Spring). (2015) 23:1414–21. doi: 10.1002/oby.21117
84. Malin SK, Solomon TP, Blaszczak A, Finnegan S, Filion J, and Kirwan JP. Pancreatic β-cell function increases in a linear dose-response manner following exercise training in adults with prediabetes. Am J Physiol Endocrinol Metab. (2013) 305:E1248–54. doi: 10.1152/ajpendo.00260.2013
85. Prior SJ, Blumenthal JB, Katzel LI, Goldberg AP, and Ryan AS. Increased skeletal muscle capillarization after aerobic exercise training and weight loss improves insulin sensitivity in adults with IGT. Diabetes Care. (2014) 37:1469–75. doi: 10.2337/dc13-2358
86. Delgado-Floody P, Caamaño-Navarrete F, Jerez-Mayorga D, Martínez-Salazar C, García-Pinillos F, and Latorre-Román P. Adaptaciones al ejercicio físico en el perfil lipídico y la salud cardiovascular de obesos mórbidos. Gac Med Mex. (2017) 153:781–6. doi: 10.24875/GMM.17002894
87. Tayebi SM, Poorhabibi H, Heidary D, Amini MA, and Sadeghi A. Impact of aerobic exercise on chronic inflammation in older adults: a systematic review and meta-analysis. BMC Sports Sci Med Rehabil. (2025) 17:229. doi: 10.1186/s13102-025-01279-z
88. Alizadeh Pahlavani H. Exercise Therapy for People With Sarcopenic Obesity: Myokines and Adipokines as Effective Actors. Front Endocrinol (Lausanne). (2022) 13:811751. doi: 10.3389/fendo.2022.811751
89. Dagdeviren S, Jung DY, Friedline RH, Noh HL, Kim JH, Patel PR, et al. IL-10 prevents aging-associated inflammation and insulin resistance in skeletal muscle. FASEB J. (2017) 31:701–10. doi: 10.1096/fj.201600832R
90. Boström P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. (2012) 481:463–8. doi: 10.1038/nature10777
91. Trettel CDS, Pelozin BRA, Barros MP, Bachi ALL, Braga PGS, Momesso CM, et al. Irisin: An anti-inflammatory exerkine in aging and redox-mediated comorbidities. Front Endocrinol (Lausanne). (2023) 14:1106529. doi: 10.3389/fendo.2023.1106529
92. Guo M, Yao J, Li J, Zhang J, Wang D, Zuo H, et al. Irisin ameliorates age-associated sarcopenia and metabolic dysfunction. J Cachexia Sarcopenia Muscle. (2023) 14:391–405. doi: 10.1002/jcsm.13141
93. Zunner BEM, Wachsmuth NB, Eckstein ML, Scherl L, Schierbauer JR, Haupt S, et al. Myokines and Resistance Training: A Narrative Review. Int J Mol Sci. (2022) 23:3501. doi: 10.3390/ijms23073501
94. Aragón-Vela J, Solis-Urra P, Ruiz-Ojeda FJ, Álvarez-Mercado AI, Olivares-Arancibia J, and Plaza-Diaz J. Impact of Exercise on Gut Microbiota in Obesity. Nutrients. (2021) 13:3999. doi: 10.3390/nu13113999
95. Han Y, Liu Y, Zhao Z, Zhen S, Chen J, Ding N, et al. Does Physical Activity-Based Intervention Improve Systemic Proinflammatory Cytokine Levels in Overweight or Obese Children and Adolescents? Insights from a Meta-Analysis of Randomized Control Trials. Obes Facts. (2019) 12:653–68. doi: 10.1159/000501970
96. Malesza IJ, Malesza M, Walkowiak J, Mussin N, Walkowiak D, Aringazina R, et al. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells. (2021) 10:3164. doi: 10.3390/cells10113164
97. Tristan Asensi M, Napoletano A, Sofi F, and Dinu M. Low-Grade Inflammation and Ultra-Processed Foods Consumption: A Review. Nutrients. (2023) 15:1546. doi: 10.3390/nu15061546
98. Holt EM, Steffen LM, Moran A, Basu S, Steinberger J, Ross JA, et al. Fruit and vegetable consumption and its relation to markers of inflammation and oxidative stress in adolescents. J Am Diet Assoc. (2009) 109:414–21. doi: 10.1016/j.jada.2008.11.036
99. Miller SJ, Batra AK, Shearrer GE, House BT, Cook LT, Pont SJ, et al. Dietary fibre linked to decreased inflammation in overweight minority youth. Pediatr Obes. (2016) 11:33–9. doi: 10.1111/ijpo.12017
100. Herrera-Rocha KM, Manjarrez-Juanes MM, Larrosa M, Barrios-Payán JA, Rocha-Guzmán NE, Macías-Salas A, et al. The Synergistic Effect of Quince Fruit and Probiotics. Nutrients. (2023) 15:4764. doi: 10.3390/nu15224764
101. Chang K, Wu W, Chen Y, Chen L, Hsu W, Lin Y, et al. Enhanced serum levels of tumor necrosis factor-α, interleukin-1β, and -6 in sarcopenia: alleviation through exercise and nutrition intervention. Aging (Albany NY). (2023) 15:13471–85. doi: 10.18632/aging.205254
102. Dominguez LJ, Veronese N, Di Bella G, Cusumano C, Parisi A, Tagliaferri F, et al. Mediterranean diet in the management and prevention of obesity. Exp Gerontol. (2023) 174:112121. doi: 10.1016/j.exger.2023.112121
103. Martínez-González MA, Salas-Salvadó J, Estruch R, Corella D, Fitó M, Ros E, et al. Benefits of the Mediterranean Diet: Insights From the PREDIMED Study. Prog Cardiovasc Dis. (2015) 58:50–60. doi: 10.1016/j.pcad.2015.04.003
104. Vidal Damasceno J, Garcez A, Anelo Alves A, da Mata IR, Morelo Dal Bosco S, and Garavaglia J. Effects of daily extra virgin olive oil consumption on biomarkers of inflammation and oxidative stress: a systematic review and meta-analysis. Crit Rev Food Sci Nutr. (2025), 1–17. doi: 10.1080/10408398.2025.2525446
105. Grosso G, Laudisio D, Frias-Toral E, Barrea L, Muscogiuri G, Savastano S, et al. Anti-Inflammatory Nutrients and Obesity-Associated Metabolic-Inflammation: State of the Art and Future Direction. Nutrients. (2022) 14:1137. doi: 10.3390/nu14061137
Keywords: exercise, nutrition, cytokines, inflammation, obesity
Citation: Sobreviela Sánchez S, Giusfredi Quevedo R, Fernandes T and da Fonseca GWP (2025) The role of exercise and nutrition in modulating inflammatory cytokines activity for obesity management. Front. Endocrinol. 16:1618360. doi: 10.3389/fendo.2025.1618360
Received: 25 April 2025; Accepted: 16 September 2025;
Published: 09 October 2025.
Edited by:
Bing Shen, Macau University of Science and Technology, Macao SAR, ChinaReviewed by:
Ana Carolina Martinez-Torres, Autonomous University of Nuevo León, MexicoMaría Antonia Parra Rizo, Miguel Hernández University of Elche, Spain
Copyright © 2025 Sobreviela Sánchez, Giusfredi Quevedo, Fernandes and da Fonseca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guilherme Wesley Peixoto da Fonseca, Z3VpbGhlcm1lZm9uc2VjYUB1c3AuYnI=