 István Fodor1*
István Fodor1* Gabriel V. Markov2
Gabriel V. Markov2 Luis Alfonso Yañez-Guerra3
Luis Alfonso Yañez-Guerra3 Károly Elekes1
Károly Elekes1 Edit Pollák4
Edit Pollák4 László Molnár1
László Molnár1 Zsolt Pirger1
Zsolt Pirger1- 1Ecophysiological and Environmental Toxicological Research Group, HUN-REN Balaton Limnological Research Institute, Tihany, Hungary
- 2Sorbonne Université, CNRS, Laboratoire de Biologie Intégrative des Modèles Marins, LBI2M, Roscoff, France
- 3Institute for Life Sciences, University of Southampton, Southampton, United Kingdom
- 4Department of Neurobiology, Institute of Biology, Faculty of Natural Sciences, University of Pécs, Pécs, Hungary
Recently, critical evaluations have challenged the presence of vertebrate-type sex steroid signaling in mollusks, underlying the need of new research lines to shed light on molluscan endocrinology. The investigation of cholesterol metabolism and the potential biological role of sterols in mollusks has emerged as a growing research field in recent years. However, there is no clear consensus on several aspects of this topic and there is a great lack of relevant molecular data. In this perspective paper, we present an overview of our current knowledge on the role of cholesterol and sterols in mollusks and try to outline possible future research directions in molluscan endocrinology. Our study also provides a framework for studying cholesterol synthesis, uptake, transport, and metabolism in mollusks.
Introduction
Cholesterol is the principal sterol in all animals, playing a key role in various biological processes, including the regulation of membrane fluidity, mediation of intercellular communication, and serving as a precursor for sex steroid synthesis (1). There has been a long-standing interest in the presence and role of cholesterol in mollusks, particularly due to the belief, over many years, that they use the same sex steroids as those of vertebrates to control reproduction. However, this belief has been challenged. There is also no clear consensus on the synthesis, transport, and metabolism of cholesterol, or of the exact biological roles of cholesterol or any other sterols that have been found in mollusks. The process of cholesterol transport mechanisms in mollusks has also not been studied enough at the molecular biological level.
In this perspective paper, we present a concise overview of our current knowledge on the role of cholesterol and sterols in mollusks. Additionally, using the great pond snail (Lymnaea stagnalis), a well-established molluscan model in neuroscience and neuroendocrinology [reviewed by (2)], we try to provide a framework for studying cholesterol synthesis, uptake, transport, and metabolism in mollusks. Our aims are 1) to further support the paradigm shift away from the currently prevailing vertebrate-centric view of molluscan reproduction and 2) to outline possible future research directions in molluscan endocrinology.
Current knowledge on the role of cholesterol and sterols in molluscan endocrinology
Presence and synthesis
Cholesterol is present in molluscan tissues in quantities more than sufficient to serve as a precursor for steroid synthesis (3). In L. stagnalis, for example, its presence can be demonstrated in all neurons of the central nervous system using histochemical (Figure 1A), immunohistochemical (Figures 1B, C), or mass spectrometric (4, 5) methods. There is a relatively long-standing debate on whether molluscan species are capable of synthesizing cholesterol de novo or whether they rely on obtaining it from their diet. The origin of cholesterol formation, in the endoplasmic reticulum (ER) from acetyl-CoA by a metabolic pathway consisting of more than 20 steps, predates the protostome-deuterostome split and is thought to be broadly conserved across eukaryotes with secondary losses in cnidarians and many protostome taxa (1). Based on the findings of early studies using radiolabeled precursors (reviewed by (6)), the current idea is that the ability to produce cholesterol de novo depends on whether the given molluscan species is carnivorous or not. Of course, non-carnivorous mollusks might also rely on obtaining it from their diet: a previous study clearly demonstrated that the northern bay scallop (Argopecten irradians irradians) can convert dietary phytosterols into cholesterol through dealkylation (7) – a process well-known in phytophagous insects (8). Interestingly, a recent study showed that, in addition to cholesterol, marine annelids could synthesize sitosterol de novo using a non-canonical C-24 sterol methyltransferase (9). Building on this finding, another recent study proposed that although early eumetazoans were capable of synthesizing C28+ sterols, many animal lineages, including mollusks, independently lost their biosynthesis (10). In accordance with this, bioconversion of dietary C28+ sterols to cholesterol has been demonstrated in the larvae of the Pacific oyster (Crassostrea gigas), suggesting a contribution of well-conserved and lineage-specific enzymes (11). Importantly, future studies are needed to further characterize the enzymes involved in the bioconversion of dietary sterols and reveal the biological role of such metabolized sterols in mollusks.
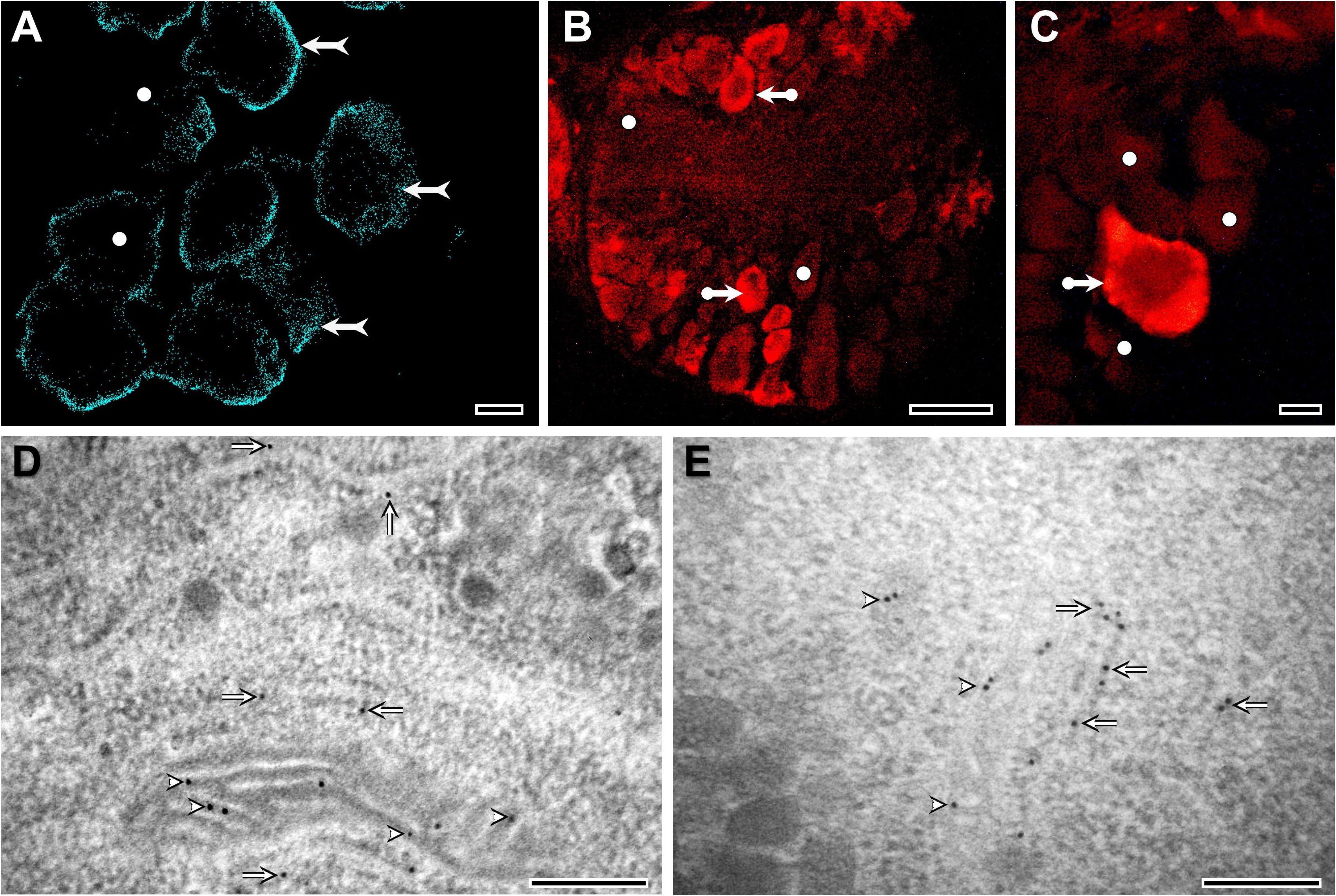
Figure 1. Distribution and localization of cholesterol in L. stagnalis neurons, visualized by Filipin III staining (A), immunohistochemistry (B, C), and immunogold labeling (D, E). At the light microscopic level, most of the staining is located close to the plasma membrane (forked arrows) (A). However, the intensity of neuronal staining varies characteristically from cell to cell. Several neurons in the ganglia show moderate or weak labeling (white dots), whereas the majority of neurons exhibit intense immunolabeling (bullet arrows), suggesting differences in cholesterol accumulation/metabolism among neurons (A-C). At the ultrastructural level, the localization of gold particles indicates the presence of cholesterol along the membranes of mitochondria (arrowheads) and rough endoplasmic reticulum (arrows) (D, E). Detailed methodology of Filipin III staining, immunohistochemistry, and immunogold electron microscopy is presented in the Supplementary Information. Bars: 10 μm (A-C) and 200 nm (D, E).
Cellular uptake and transport
Cholesterol is sparingly soluble in water and cannot thus be transported in the bloodstream (or in the case of mollusks, the hemolymph) in the sort of quantities that are needed by the cells of the body. Instead, cholesterol is first of all conjugated to a fatty acid. This is known as esterification and it involves the condensation of the hydroxyl group on the 3rd Carbon atom of cholesterol to the alcohol function of the carboxyl group from a fatty acid with the elimination of a water molecule. The ‘cholesterol ester’ thus formed is then bound up with other fat molecules into spherical particles by lipoproteins. The lipoproteins are amphiphilic and form a shell with their lipophilic ends facing inwards (where the fats are stored) and hydrophilic ends facing outwards. There are two main types of lipoproteins: low-density lipoprotein (LDL) and high-density lipoprotein (HDL).
The uptake of the spherical particles is primarily mediated by LDL membrane receptors (LDL-Rs) on the cell walls (Figure 2A) (12), which bind to LDL particles to facilitate their endocytosis into the cell. Once in the cells, they fuse with lysosomes, which contain carboxyl ester lipase 1 (CEL1) enzyme that converts cholesterol ester back into free cholesterol. The free cholesterol is then distributed by sterol carrier protein 2 (SCP2), a non-specific lipid transfer proteins which is also involved in the transport and metabolism of many lipids (13, 14), and utilized in various biological processes. A small percent of cholesterol esters, bound to HDL, can also be incorporated directly into the plasma membrane. This process is mediated by a hydrophobic channel within scavenger receptor class B type 1 (SRB1), a high-density lipoprotein receptor (i.e. without classical endocytosis). Once entering the membrane, cholesteryl esters are also hydrolyzed and then released as free cholesterol. Although SRB1 homologs have been identified in some molluscan species (15, 16), the investigation of cholesterol transporters and carriers in this taxon has somewhat been neglected. Screening both our own and publicly available L. stagnalis genome and transcriptome data revealed sequences homologous to vertebrate LDL-R, CEL1, SRB1, and SCP2 proteins (see Supplementary Information), suggesting an uptake mechanism highly similar to that of vertebrates.
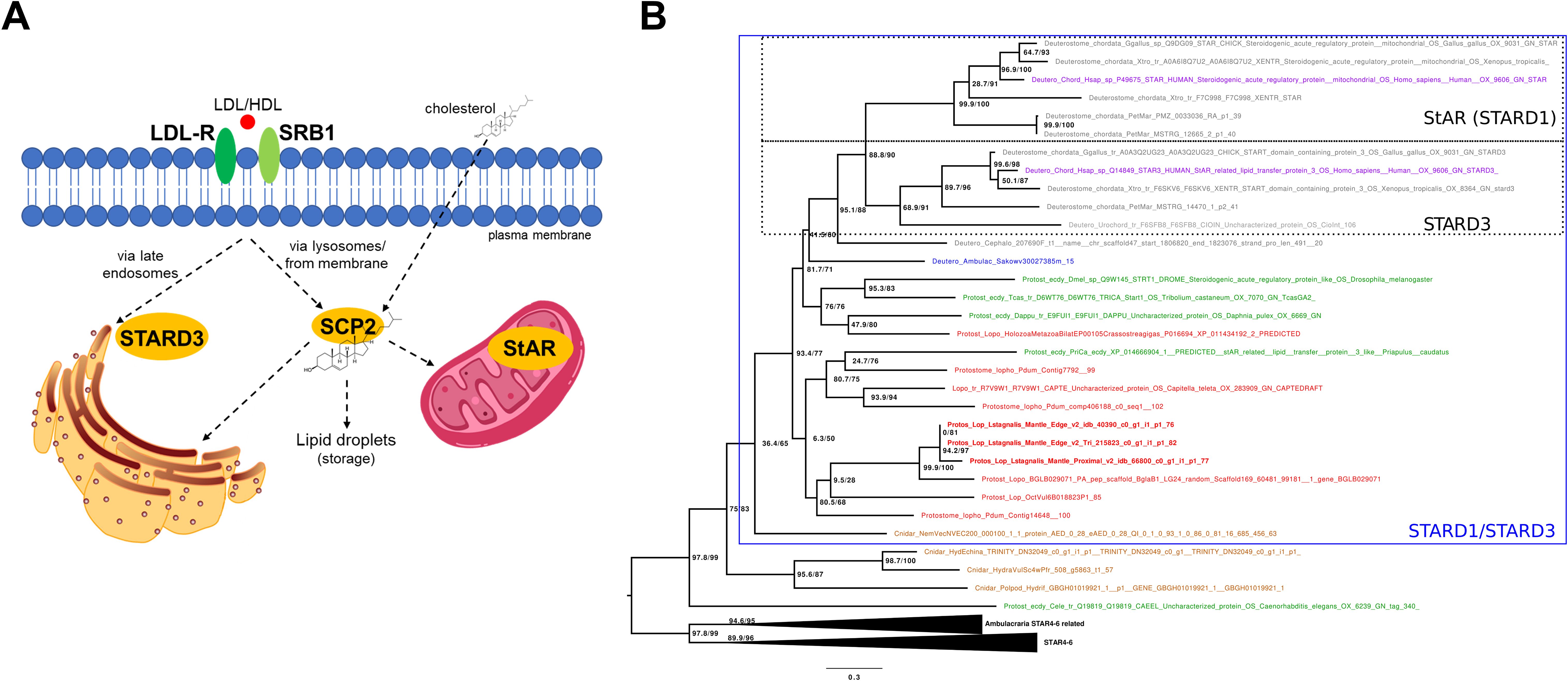
Figure 2. Cellular uptake and transport with the most relevant proteins of cholesterol. (A) Schematic overview of cellular uptake and transport of cholesterol in vertebrates. Low density lipoprotein (LDL) binds to the LDL receptor (LDL-R) and is trafficked through the membrane via the classic the endosomal pathway. LDL-containing endosomes fuse with lysosomes, where cholesterol esters are hydrolyzed into free cholesterol which is then distributed by sterol carrier protein 2 (SCP2). Cholesterol esters can also be incorporated directly into the plasma membrane through receptor scavenger receptor class B type 1 (SRB1), a high-density lipoprotein receptor. Once in the membrane, cholesteryl esters are also hydrolyzed and released as free cholesterol. A small percentage of cholesterol can also enter cells via passive diffusion across the plasma membrane. The steroidogenic acute regulatory protein (StAR; STARD1) mediates the transfer of cholesterol between the two membranes of the mitochondria. StAR related lipid transfer domain containing 3 (MLN64; STARD3) mediates the cholesterol transfer at endoplasmic reticulum-endosome contact sites. L. stagnalis sequences homologous to these proteins suggest transport mechanisms highly similar to those of vertebrates. (B) Phylogenetic tree showing the occurrence and relationships of STARD proteins. The tree shows that StAR and STARD3 represent a chordate-specific expansion. Lophotrochozoans possess many-to-many homologs to chordate StAR and STARD3 proteins. Branch support values are indicated at nodes in the format aLRT/UFBoot (e.g., 100/100). Detailed methodology of the phylogenetic analysis and the raw tree are presented in the Supplementary Information.
The intracellular transport of cholesterol within organelles has been of particular interest in molluscan research due to its presumed role as a precursor for vertebrate-type sex steroids (6, 17). In vertebrates, the crucial step in steroid synthesis takes place within the mitochondria, where Cytochrome P450 (CYP) 11A converts cholesterol to pregnenolone (17). Since cholesterol cannot simply diffuse across the aqueous phase between the two membranes, its transfer is mediated by the steroidogenic acute regulatory protein (StAR; also known as STARD1), which is the rate-limiting step in the synthesis of steroid hormones in vertebrates (Figure 2A) (13).
Approximately 30% of cholesterol taken up by LDL-Rs is directly transported to the ER (18). In the case of the LDL-containing endosomes targeting the ER, StAR related lipid transfer domain containing 3 protein (STARD3; also known as MLN64) mediates the cholesterol transfer at ER–endosome contact sites (18). Although the presence of StAR has been hypothesized in mollusks in general (1), according to our best knowledge, previous studies have identified only homologs of STARD3 (19, 20). Moreover, there is a single preliminary phylogenetic analysis providing only the information that molluscan STARD3 sequences are separated from vertebrate STARD3s (20). Using Hidden Markov Models, our thorough searches in the available genome and/or transcriptome data of deuterostome, protostome, and non-bilaterian animals revealed candidate genes for both StAR and STARD3. Then, using all-vs-all cluster-based methodologies (Supplementary Figure S1) and phylogenetic analysis (Figure 2B; Supplementary Figure S2), we clearly demonstrated that 1) StAR and STARD3 are a duplication that happened in chordates and that 2) lophotrochozoans have many-to-many homologs to chordate StAR and STARD3 proteins. In L. stagnalis, specifically, our analysis revealed the presence of three STARD1/D3 homologs (see Supplementary Information). Conserved domain analysis confirmed that these proteins contain the START/SRPBCC domain characteristic of STARD proteins (see Supplementary Information). Since our immunogold electron microscopy investigations demonstrated the presence of cholesterol in the inner mitochondrial membrane of L. stagnalis neurons (Figures 1D, E), we propose that the transport mechanism of cholesterol across the mitochondria in mollusks is similar to that in vertebrates and is mediated by at least one STARD1/D3 protein.
Given that there is no clear evidence for vertebrate-type sex steroid synthesis in mollusks (6, 17, 21, 22), the actual role of cholesterol found within the mitochondria in these species remains unclear. This is also the case for arthropods, as the cholesterol to 7-dehydrocholesterol conversion in the first step of ecdysone synthesis is implemented in the ER (23). Hence, it would also be important to investigate the proteins which mediate the cholesterol transfer at the ER–endosome contact site. In addition to STARD3, proteins such as Niemann-Pick intracellular cholesterol transporter 1 (NPC1), NPC2, Oxysterol-binding protein–related protein 5 (ORP5), ORP1, VAP proteins, Ras-related protein Rab-7a (Rab7), and Rab-interacting lysosomal protein (RILP) are known to play key roles in the process in vertebrates (18). The presence of molluscan homologs to these proteins has been scarcely studied so far (1). Screening the L. stagnalis genome and transcriptome data revealed sequences homologous to these vertebrate proteins, except for RILP (see Supplementary Information). These homologs, as well as the presence of cholesterol along the membranes of rough ER in L. stagnalis neurons (Figures 1D, E), suggest a transport mechanism sharing some similarities to that of vertebrates. Future research on other species is required to clarify the general application of cellular processes involved in cholesterol uptake and transport in mollusks.
Metabolism: a diversity of long-chained steroids as a new direction
A recent study suggested that paraestrol A, an aromatized sterol (i.e. a cholesterol metabolite that has not undergone side-chain cleavage), functions as an activator of the ancestral steroid receptor (22). Given this, investigating sterols as potential hormones in mollusks has been proposed as a new research direction that could shed light on molluscan (21).
Aromatized long-chained steroids (e.g., geodisterol) were first identified unequivocally in sponges and cnidarians (24, 25), and some of them, like the fevicordins, are also found in some land plants [cucurbids and malvales (26)]. This raises the possibility that some aromatized sterols could be present in mollusks as well – with their aromatization being catalyzed by a (still unknown) paralogous CYP enzyme. All those molecules indicate that the chemical space to be explored is larger than the one restricted to vertebrate-type steroids. Moreover, natural product chemists have already identified some specific molluscan sterols. For example, epidioxysterols were identified in Aplysia (some of them were isolated from the eggs) (27), referring to the potential role of non-aromatized steroids in mollusks, either as defense compounds or as hormones. We propose that molluscan 5α-reductases are involved in the synthesis of such sterols. 5α-reductase is an ancient enzyme present in both plants and animals and is known to use sterols (as well as testosterone and androstenedione in vertebrates) as substrates (28, 29). In plants, campesterol is a substrate for 5α-reductase (30), so it is reasonable to suppose that molluscan 5α-reductases could catalyze a similar reaction on cholesterol. This would make perfect sense when looking at the structures of the 5α,8α epidioxysterols reported in Aplysia: those molecules derive probably from cholesterol, and they must lose the delta 5–6 double bond by delta-5-reduction at some point. The potential role of sterols in molluscan endocrine processes is also supported by a recent finding that inhibition of 5α-reductase caused marked shell malformations during the embryonic development of freshwater gastropods (30). Additional to epidioxysterols, other secosteroids, which may derive from further oxidative degradation of the sterol backbone, were also identified (27), indicating that the chemical space to be explored is large, and enabling a variety of functions for all those molecules. An additional important question is in which tissues these steroids are synthesized in mollusks. Previous preliminary analyses conducted in the Aplysia genus have shown that these metabolites are not accumulated in a specific organ but distributed in the whole organism (31). However, their presence in a given tissue does not necessarily imply that synthesis occurs there. Tissue-specific gene expression studies (e.g., analysis of the tissue distribution of the 5α-reductase enzyme) are required to clearly determine the production sites. In summary, future studies, on the one hand, are needed to further characterize the diversity of molluscan sterols, and on the other hand, to identify the molluscan enzymes and tissues involved in sterol synthesis.
It is worth noting, however, that the concept of sterols as potential hormones warrants careful consideration in light of their chemical nature. Sterol-based compounds, like paraestrols, share structural similarities with cholesterol, which is characterized by its low water solubility. Due to this property, sterols are generally not freely soluble in aqueous environments such as the hemolymph and are therefore unlikely to be efficiently transported as classical circulating hormones. Instead, their bioactivity is likely confined to the cells or tissues where they are synthesized, suggesting a role more consistent with autocrine or paracrine signaling rather than endocrine function. This physicochemical constraint highlights the need for caution when applying the term “hormone” to sterol-like molecules, as their mode of action may not involve systemic transport but rather localized, cell-autonomous effects.
Finally, it is possible that some molluscan long-chain steroids bind to nuclear receptors belonging to the NR1H or NR1I/J/K families, which include oxysterol and bile acid receptors in vertebrates, the ecdysone receptor in arthropods, and the dafachronic acid receptor in nematodes. In our opinion, this is the direction where it would be reasonable to search for nuclear receptor-based steroid signaling in lophotrochozoans (see also (32)). Speculatively, long-chained steroids might also exert their effects via non-genomic signaling, through either membrane receptors or other lipid-binding domains which are not directly transcription factors. Indeed, long chained vertebrate bile acids were found to be activators of sodium channels in an acorn worm and a brachiopod, suggesting that unknown bile acid-like molecules may be the endogenous ligands for those receptors that are widespread across bilaterians (33).
Summary
Since the 1950s, a vertebrate-centered perspective has characterized molluscan endocrinology research. Starting from 2011, critical evaluations have challenged the concept of sex steroid signaling in mollusks, highlighting the absence of endogenous synthesis and nuclear receptors for vertebrate-type sex steroids (6, 17, 21, 34–37). In light of this, clearly, new research directions are needed to shed light on actual questions in molluscan endocrinology. We believe that investigation of sterols can significantly contribute to the understanding of molluscan endocrine systems, such as nuclear receptor-based steroid signaling, although not necessarily only in relation to reproduction. Future studies should aim 1) to investigate cholesterol metabolism and sterol synthesis in details, 2) to identify sterols and their receptors in different molluscan species, and 3) to investigate the role of sterols in molluscan physiology.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because Experiments on Lymnaea do not require ethical approval. However, all procedures, methods, and experiments on the specimens were carried out in accordance with the relevant guidelines and regulations approved by the Scientific Committee of Animal Experimentation of the Balaton Limnological Research Institute (VE-I-001/01890-10/2013).
Author contributions
IF: Conceptualization, Data curation, Funding acquisition, Investigation, Visualization, Writing – original draft. GM: Conceptualization, Writing – review & editing. LY-G: Data curation, Funding acquisition, Investigation, Visualization, Writing – review & editing. KE: Writing – review & editing. EP: Investigation, Visualization, Writing – review & editing. LM: Conceptualization, Investigation, Visualization, Writing – review & editing. ZP: Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the National Brain Project (#NAP2022-I-10/2022, ZP), the Hungarian Scientific Research Fund (#138039, ZP; #146787, IF), the Bolyai Foundation (#BO/00521/24, IF), the Biotechnology and Biological Sciences Research Council (#BB/W010305/2, LY-G), and the Royal Society (#RG\R1\241397, LY-G).
Acknowledgments
The authors thank Prof. Alexander P. Scott (CEFAS, Weymouth, UK) for reviewing the English of the manuscript and for providing useful suggestions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2025.1627166/full#supplementary-material
References
1. Zhang T, Yuan D, Xie J, Lei Y, Li J, Fang G, et al. Evolution of the cholesterol biosynthesis pathway in animals. Mol Biol Evol. (2019) 36:2548–56. doi: 10.1093/molbev/msz167
2. Fodor I, Hussein AA, Benjamin PR, Koene JM, and Pirger Z. The unlimited potential of the great pond snail. Lymnaea Stagnalis eLife. (2020) 9:e56962. doi: 10.7554/eLife.56962
3. Idler DR and Wiseman P. Molluscan sterols: a review. J Fish Res Board Can. (1972) 29:385–98. doi: 10.1139/f72-065
4. Altelaar AF, van Minnen J, Jimenez CR, Heeren RM, and Piersma SR. Direct molecular imaging of Lymnaea stagnalis nervous tissue at subcellular spatial resolution by mass spectrometry. Anal Chem. (2005) 77:735–41. doi: 10.1021/ac048329g
5. McDonnell LA, Piersma SR, MaartenAltelaar AF, Mize TH, Luxembourg SL, Verhaert PD, et al. Subcellular imaging mass spectrometry of brain tissue. J Mass Spectrom. (2005) 40:160–8. doi: 10.1002/jms.735
6. Scott AP. Do mollusks use vertebrate sex steroids as reproductive hormones? Part I. Critical appraisal of the evidence for the presence, biosynthesis and uptake of steroids. Steroids. (2012) 77:1450–68. doi: 10.1016/j.steroids.2012.08.009
7. Giner JL, Zhao H, Dixon MS, and Wikfors GH. Bioconversion of (13)C-labeled microalgal phytosterols to cholesterol by the Northern Bay scallop. Argopecten Irradians Irradians Comp Biochem Physiol B Biochem Mol Biol. (2016) 192:1–8. doi: 10.1016/j.cbpb.2015.11.003
8. Ikekawa N, Morisaki M, and Fujimoto Y. Sterol-metabolism in insects - dealkylation of phytosterol to cholesterol. Accounts Chem Res. (1993) 26:139–46. doi: 10.1021/ar00028a002
9. Michellod D, Bien T, Birgel D, Violette M, Kleiner M, Fearn S, et al. De novo phytosterol synthesis in animals. Science. (2023) 380:520–6. doi: 10.1126/science.add7830
10. Brunoir T, Mulligan C, Sistiaga A, Vuu KM, Shih PM, O’Reilly SS, et al. Common origin of sterol biosynthesis points to a feeding strategy shift in Neoproterozoic animals. Nat Comm. (2023) 14:7941. doi: 10.1038/s41467-023-43545-z
11. da Costa F, Mathieu-Resuge M, Le Grand F, Quéré C, Markov GV, Wikfors GH, et al. Sterol Biosynthesis and Phytosterol Bioconversion in Crassostrea Gigas larvae: new evidence from mass-balance feeding studies. Biochimie. (2025) 16:S0300–9084(25)00095-1. doi: 10.1016/j.biochi.2025.05.005
12. Gallo-Payet N and Battista MC. Steroidogenesis-adrenal cell signal transduction. Compr Physiol. (2014) 4:889–964. doi: 10.1002/cphy.c130050
13. Galano M, Venugopal S, and Papadopoulos V. Role of STAR and SCP2/SCPx in the transport of cholesterol and other lipids. Int J Mol Sci. (2022) 23:12115. doi: 10.3390/ijms232012115
14. Schroeder F, Atshaves BP, McIntosh AL, Gallegos AM, Storey SM, Parr RD, et al. Sterol carrier protein-2: new roles in regulating lipid rafts and signaling. Biochim Biophys Acta. (2007) 1771:700–18. doi: 10.1016/j.bbalip.2007.04.005
15. Wei X, Zhao T, Ai K, Li H, Jiang X, Li C, et al. Role of scavenger receptor from Octopus ocellatus as a co-receptor of Toll-like receptor in initiation of TLR-NF-kappaB signaling during anti-bacterial response. Dev Comp Immun. (2018) 84:14–27. doi: 10.1016/j.dci.2018.01.023
16. Yang D, Han Y, Chen L, Liu Y, Cao R, Wang Q, et al. Scavenger receptor class B type I (SR-BI) in Ruditapes philippinarum: A versatile receptor with multiple functions. Fish Shellfish Immun. (2019) 88:328–34. doi: 10.1016/j.fsi.2019.03.009
17. Fodor I, Urban P, Scott AP, and Pirger Z. A critical evaluation of some of the recent so-called ‘evidence’ for the involvement of vertebrate-type sex steroids in the reproduction of mollusks. Mol Cell Endocrinol. (2020) 516:110949. doi: 10.1016/j.mce.2020.110949
18. Raiborg C, Wenzel EM, and Stenmark H. ER-endosome contact sites: molecular compositions and functions. EMBO J. (2015) 34:1848–58. doi: 10.15252/embj.201591481
19. Fodor I, Koene JM, and Pirger Z. Neuronal transcriptome analysis of a widely recognised molluscan model organism highlights the absence of key proteins involved in the de novo synthesis and receptor-mediation of sex steroids in vertebrates. Malacologia. (2021) 64:69–77. doi: 10.4002/040.064.0103
20. Thitiphuree T, Nagasawa K, and Osada M. Molecular identification of steroidogenesis-related genes in scallops and their potential roles in gametogenesis. J Steroid Biochem Mol Biol. (2019) 186:22–33. doi: 10.1016/j.jsbmb.2018.09.004
21. Fodor I and Pirger Z. From dark to light - an overview of over 70 years of endocrine disruption research on marine mollusks. Front Endocrinol. (2022) 13:903575. doi: 10.3389/fendo.2022.903575
22. Markov GV, Gutierreze-Mazariegos J, Pitrat D, Billas IML, Bonneton F, Moras D, et al. Origin of an ancient hormone/receptor couple revealed by resurrection of an ancestral estrogen. Sci Adv. (2017) 3:e1601778. doi: 10.1126/sciadv.1601778
23. Yoshiyama T, Namiki T, Mita K, Kataoka H, and Niwa R. Neverland is an evolutionally conserved Rieske-domain protein that is essential for ecdysone synthesis and insect growth. Development. (2006) 133:2565–74. doi: 10.1242/dev.02428
24. Wang G-Y-S and Crews P. Geodisterol, a novel polyoxygenated sterol with an aromatic A ring from the tropical marine sponge Geodia sp. Tetrahedron Lett. (1996) 37:8145–6. doi: 10.1016/0040-4039(96)01894-1
25. Yan XH, Liu HL, Huang H, Li XB, and Guo YW. Steroids with aromatic A-rings from the Hainan soft coral. Dendronephthya Studeri Ridley J Nat Prod. (2011) 74:175–80. doi: 10.1021/np100562n
26. Markov GV, Girard J, Laudet V, and Leblanc C. Hormonally active phytochemicals from macroalgae: A largely untapped source of ligands to deorphanize nuclear receptors in emerging marine animal models. Gen Comp Endocrinol. (2018) 265:41–5. doi: 10.1016/j.ygcen.2018.06.004
27. Pereira RB, Andrade PB, and Valentao P. Chemical diversity and biological properties of secondary metabolites from sea hares of aplysia genus. Mar Drugs. (2016) 14:39. doi: 10.3390/md14020039
28. Rosati F, Danza G, Guarna A, Cini N, Racchi ML, and Serio M. New evidence of similarity between human and plant steroid metabolism: 5alpha-reductase activity in. Solanum Malacoxylon Endocrinol. (2003) 144:220–9. doi: 10.1210/en.2002-220649
29. Ohnishi T. Recent advances in brassinosteroid biosynthetic pathway: insight into novel brassinosteroid shortcut pathway. J Pestic Sci. (2018) 43:159–67. doi: 10.1584/jpestics.D18-040
30. Baynes A, Montagut Pino G, Duong GH, Lockyer AE, McDougall C, Jobling S, et al. Early embryonic exposure of freshwater gastropods to pharmaceutical 5-alpha-reductase inhibitors results in a surprising open-coiled “banana-shaped” shell. Sci Rep. (2019) 9:16439. doi: 10.1038/s41598-019-52850-x
31. Jimenez C, Quinoa E, Castedo L, and Riguera R. Epidioxy Sterols from the Tunicates Dendrodoa grossularia and Ascidiella aspersa and the Gastropoda Aplysia depilans and Aplysia punctata. J Nat Prod. (1986) 49:905–9. doi: 10.1021/np50047a023
32. Markov GV and Laudet V. Origin and evolution of the ligand-binding ability of nuclear receptors. Mol Cell Endocrinol. (2011) 334:21–30. doi: 10.1016/j.mce.2010.10.017
33. Marti-Solans J, Borve A, Vevle L, Hejnol A, and Lynagh T. Invertebrate bile acid-sensitive ion channels and their emergence in bilateria. FASEB J. (2025) 39:e70526. doi: 10.1096/fj.202403216R
34. Scott AP. Do mollusks use vertebrate sex steroids as reproductive hormones? II. Critical review of the evidence that steroids have biological effects. Steroids. (2013) 78:268–81. doi: 10.1016/j.steroids.2012.11.006
35. Scott AP. Is there any value in measuring vertebrate steroids in invertebrates? Gen Comp Endocrinol. (2018) 265:77–82. doi: 10.1016/j.ygcen.2018.04.005
36. Balbi T, Ciacci C, and Canesi L. Estrogenic compounds as exogenous modulators of physiological functions in molluscs: Signaling pathways and biological responses. Comp Biochem Physiol C. (2019) 222:135–44. doi: 10.1016/j.cbpc.2019.05.004
Keywords: mollusks, cholesterol, sterols, synthesis, transport, metabolism, sex steroids
Citation: Fodor I, Markov GV, Yañez-Guerra LA, Elekes K, Pollák E, Molnár L and Pirger Z (2025) Cholesterol and sterols in molluscan endocrinology: past, present, future. Front. Endocrinol. 16:1627166. doi: 10.3389/fendo.2025.1627166
Received: 12 May 2025; Accepted: 14 July 2025;
Published: 31 July 2025.
Edited by:
Toshio Sekiguchi, Kanazawa University, JapanReviewed by:
Takashi Yazawa, Asahikawa Medical University, JapanCopyright © 2025 Fodor, Markov, Yañez-Guerra, Elekes, Pollák, Molnár and Pirger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: István Fodor, Zm9kb3IuaXN0dmFuQGJsa2kuaHU=