 Carmen J. Zamora-Sánchez1
Carmen J. Zamora-Sánchez1 Juan Carlos González-Orozco2
Juan Carlos González-Orozco2 Jonatan Mendoza-Ortega1,3
Jonatan Mendoza-Ortega1,3 Mariana L. Villegas-Soto4
Mariana L. Villegas-Soto4 Ignacio Camacho-Arroyo4
Ignacio Camacho-Arroyo4 Guadalupe Estrada-Gutierrez1*
Guadalupe Estrada-Gutierrez1*- 1Department of Immunobiochemistry, National Institute of Perinatology, Mexico, Mexico
- 2Department of Genetics, The University of Texas MD Anderson Cancer Center, Houston, TX, United States
- 3Departamento de Inmunología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Ciudad de México, Mexico
- 4Unidad de Investigación en Reproducción Humana, Instituto Nacional de Perinatología-Facultad de Química, Universidad Nacional Autónoma de México (UNAM), Mexico, Mexico
Neuroactive steroids synthesized within the maternal-placental-fetal unit play a crucial role in fetal neurodevelopment by regulating cell proliferation, migration, and myelination, neurogenesis, gliogenesis, and synaptogenesis, ultimately shaping brain maturation. Dysregulation of neuroactive steroid metabolism, receptor signaling, and downstream pathways has been linked to neurodevelopmental and mood disorders. Maternal overweight and obesity, increasingly prevalent worldwide, induce profound metabolic and hormonal adaptations that may interfere with neuroactive steroid synthesis and function. These disturbances are associated with a higher risk of autism spectrum disorder, attention deficit hyperactivity disorder, and cognitive impairments in offspring, frequently with sex-specific effects. Despite these observations, the impact of obesity on neuroactive steroid levels and their regulatory roles during pregnancy remains poorly understood. This review synthesizes preclinical and clinical evidence on the biosynthesis, mechanisms of action, and neurodevelopmental effects of neuroactive steroids during the critical window of fetal programming. Furthermore, it highlights a current knowledge on how maternal overweight and obesity alter neuroactive steroid metabolism within the maternal–placental–fetal unit and explores their potential contribution to adverse neurodevelopmental outcomes. Addressing these knowledge gaps may uncover novel biomarkers and therapeutic targets to improve neurodevelopmental trajectories in populations increasingly exposed to maternal metabolic comorbidities.
1 Introduction
The terms neuroactive steroid and neurosteroid are frequently used interchangeably, although they differ in origin. Neurosteroids are synthesized locally within the nervous system, whereas neuroactive steroids encompass a broader group of steroids produced in peripheral tissues —including the gonads and adrenal glands— that exert regulatory effects on the central nervous system (CNS). Thus, all neurosteroids are neuroactive steroids, but not all neuroactive steroids are neurosteroids. This review focuses on neuroactive steroids in general, since, as we describe further, during pregnancy, these molecules are produced by distinct organs for both the mother and fetus.
During pregnancy, the maternal–placental–fetal unit, comprising the ovaries, placenta, and fetal adrenal zone, becomes a critical source of neuroactive steroids (1). These molecules interact with diverse receptors — including nuclear receptors, G-protein coupled receptors, membrane receptors, and ligand-gated ion channels (2–7)—modulating fundamental processes such as neurogenesis, synaptogenesis, myelination, and neuroprotection (1, 5, 7–10).
Neurodevelopment itself is a highly orchestrated process in which proliferation, migration, and differentiation of neural cells give rise to the cellular diversity and structural complexity of the CNS (11). The most vulnerable period spans “the first 1,000 days of life”, from conception through the first two years of postnatal life, during which, neuroactive steroids, such as progestogens, estrogens, androgens, and glucocorticoids are synthesized at particularly high levels by the maternal-placental-fetal unit. These steroid networks are highly sensitive to physiological, metabolic, and environmental influences. Among these, maternal overweight and obesity stand out as conditions that disrupt endocrine homeostasis, potentially altering the production and activity of neuroactive steroids during pregnancy.
Overweight and obesity are pathological conditions characterized not only by excessive fat mass but also by complex metabolic and endocrine disturbances. As adipose tissue functions as an active endocrine organ, obesity alters circulating adipokines, insulin sensitivity, and steroid hormone levels. Disruptions in these systems are known to affect fetal neurodevelopment and have been consistently associated with increased risks of autism spectrum disorder (ASD), attention deficit hyperactivity disorder (ADHD), and cognitive impairments in offspring. Importantly, alterations in neuroactive steroid levels—particularly allopregnanolone (3α-THP)—have been observed in neurodevelopmental conditions such as ASD (12, 13).
Despite these associations, direct evidence linking maternal obesity to disrupted neuroactive steroid synthesis or signaling remains scarce. This review addresses this gap by integrating mechanistic, preclinical, and clinical data to evaluate how maternal obesity modulates neuroactive steroids within the maternal–placental–fetal unit and how these alterations may shape neurodevelopmental trajectories.
2 Sources and metabolism of neuroactive steroids during prenatal and early postnatal life
Neuroactive steroids are key regulators of function and CNS homeostasis. Their influence begins in prenatal life and extends throughout the lifespan, with particularly critical roles during early neurodevelopment when neuronal circuits are formed and refined (14, 15). In mammals, neuroactive steroids reach the fetal brain via three main routes: maternal circulation, placental synthesis, and local fetal production—adrenal, hepatic, and gonadal tissues, as well as by the CNS (16). Here, we primarily focus on their synthesis within the developing brain, which originates from preexisting cholesterol, locally synthesized cholesterol, or sulfate-conjugated derivatives. As early as two weeks of gestation in humans, these steroid precursors are taken up by the placenta from the maternal circulation and transported to the fetus via various lipoprotein receptors, ATP-binding cassette transporters, and solute carrier transporters (17, 18). However, the placenta is not only involved in the uptake and transport of steroid precursors but also develops its own steroidogenic machinery. This activity becomes evident in the syncytiotrophoblast, which produces substantial placental neuroactive steroids, such as progesterone (P4), emerging between the 6th and 8th weeks of gestation in humans (19–21).
The synthesis of cholesterol in the prenatal brain (as well as in other common steroidogenic tissues) begins with two molecules of acetyl-CoA that are condensed to form acetoacetyl-CoA, which is then combined with another acetyl-CoA to produce 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA). This molecule is reduced to mevalonate by the enzyme HMG-CoA reductase, the rate-limiting step of cholesterol synthesis. Subsequently, mevalonate is modified to produce isopentenyl pyrophosphate (IPP), which is finally converted to the direct precursors of cholesterol, squalene, and lanosterol (22). Although the mammalian embryo produces most of its own cholesterol, the maternal cholesterol supply is essential during the first weeks of life (23). However, once the blood-brain barrier is well established, the emerging CNS (particularly the brain) relies solely on de novo synthesis, as it is not permeable to circulating lipoprotein-bound cholesterol (24). This occurs at the embryonic stage E11-E12 in rodents and at 14 weeks of gestation in humans (16).
Once available, cholesterol is transported to mitochondria by proteins such as the 18 kDa translocator protein (TSPO) and the steroidogenic acute regulatory protein (StAR) (25). In the mitochondria, the P450 side-chain cleavage enzyme (P450scc, also known as CYP11A1) converts cholesterol to pregnenolone, which is already considered a neurosteroid (26). Studies have shown that the mammalian CNS can start to synthesize pregnenolone from the very early stages of prenatal development. In rodents, the expression of P450scc in neural tissue is detected as early as the E9.5 embryonic stage, while in humans, its expression is noticeable around the 10th week of gestation (5, 27). Besides local synthesis in the prenatal brain, other steroidogenic organs, such as the placenta, adrenal glands, and gonads, are also important sources of pregnenolone for the prenatal and early postnatal CNS (28), as the blood-brain barrier is easily permeable to this steroid (29). Upon availability of pregnenolone, it can be further metabolized into various steroids that play important roles in neurodevelopment. Pregnenolone is directly converted to P4 by the enzyme 3β-hydroxysteroid dehydrogenase (3β-HSD) (30). Since P4 is indispensable for pregnancy maintenance in mammals, the fetal CNS is continuously exposed to high levels of this steroid hormone from the maternal placental circulation. In humans, chorionic gonadotropin is produced early in pregnancy by trophoblasts to sustain corpus luteum-mediated P4 production during the first nine weeks of gestation. Afterward, the trophoblasts of the placenta take over this function (31, 32). In contrast, in rodents, the ovarian corpus luteum maintains P4 production throughout the entire pregnancy (31, 32). In addition to its luteotropic role, human chorionic gonadotropin also modulates hypothalamic signaling, contributing to the regulation of both the hypothalamic-pituitary-adrenal (HPA) and hypothalamic-pituitary-gonadal (HPG) axes in the mother and fetus (31, 32).
Regarding the local synthesis of P4, the human CNS produces this hormone from prenatal stages through postnatal life. Expression and activity of 3β-HSD, the key enzyme involved in P4 synthesis, have been detected in the fetal brain as early as the second trimester of pregnancy (33, 34). Importantly, prenatal and early postnatal local synthesis of P4 in the CNS is also noticeable in rodents and sheep. In rodents, in particular, the cerebellum exhibits the largest production of P4 within the CNS during the first days of neonatal life (35–38).
P4 is further metabolized into several neuroactive steroid derivatives. It is commonly converted by the 5α-reductase (5α-R) enzyme into 5α-dihydroprogesterone (DHP), which is later reduced to allopregnanolone (3α-THP) by the 3α-hydroxysteroid oxidoreductases (3α-HSD). 3α-THP is a neurosteroid with key roles in neurodevelopment (39). High levels of 3α-THP are observed during prenatal development and immediately after birth in species such as guinea pigs, sheep, and rodents. During late pregnancy, the CNS exhibits an increase in P450scc and 5α-reductase levels, suggesting that its capacity for synthesizing P4 and 3α-THP peaks around the time of birth (36, 38, 40, 41).
The presence and activity of androgens and estrogens have also been observed during early neurodevelopment. Pregnenolone is also a direct source of androgens. It is converted into the androgen dehydroepiandrosterone (DHEA) by the enzyme 17α-hydroxylase (CYP17A1); moreover, CYP17A1 also catalyzes the conversion of P4 into androstenedione (30, 42). Notably, the expression of CYP17A1 has been detected in the rat brain during prenatal development, and it decreases after birth (36). However, the adrenal cortex becomes an essential source of DHEA, a testosterone (T) precursor, during the first years of postnatal life (43, 44). Moreover, peripheral DHEA can cross the blood-brain barrier and become a source of other active androgens and estrogens in the CNS (45).
Besides its role in progestogen metabolism, 3β-HSD also converts DHEA into androstenedione. Then, the activity of the different 17β-hydroxysteroid dehydrogenases (17β-HSD) catalyzes the synthesis of T from androstenedione. Notably, 17β-HSD can also catalyze the conversion of DHEA into androstenediol, which is also used for T synthesis by the enzyme 3β-HSD (41). Then, T is further metabolized by 5α-R to dihydrotestosterone (DHT), which is a more potent androgen (46). Expression of 17β-HSD can be detected in the human placenta as early as the 4th week of pregnancy, which supplies fetal circulation with T and other active estrogens (20, 32). More importantly, the human fetal brain expresses and shows the activity of 17β-HSD enzymes as early as 13 weeks of gestation (47), while in mice, its expression is noticeable along with 5α-R expression starting at E12. Furthermore, 17β-HSD and 5α-R expression levels increase in the mouse fetal brain of males and females as development progresses (48).
As mentioned before, the placenta also supplies the fetal brain with estrogens, as it has been found in humans and rodents (20, 32, 49, 50). However, the mammalian CNS is also capable of local estrogen synthesis. It expresses the enzyme aromatase (CYP19A1) during prenatal and postnatal life in both sexes. The enzyme aromatase unidirectionally converts T into estradiol (E2). Nevertheless, it also catalyzes the conversion of androstenedione into estrone, which is further metabolized to E2 by 17β-HSD (51–54).
Glucocorticoids also play a key role during early neurodevelopment. Corticosterone and cortisol are the most relevant neuroactive steroids belonging to this group, and the adrenal cortex is the main source of these neuroactive steroids in the organism, which easily cross the blood-brain barrier (55). These neurosteroids are synthesized from P4 and its metabolite 17-OH progesterone. Specifically, the enzyme 21-hydroxylase (CYP21) synthesizes 11-deoxycorticosterone and 11-deoxycortisol from P4 and 17-OH progesterone, respectively. Then, P450C11 (CYP11) produces corticosterone and cortisol, respectively, from such substrates (41). In humans, maternal cortisol plays a major role and is easily transferred to the fetus via the placental circulation during the first trimester of pregnancy (56). After that period, the fetal adrenals can produce corticosterone and cortisol from maternal P4 starting at 8–10 weeks post-conception (57). Furthermore, after birth, maternal milk provides corticosterone and cortisol to the neonate (58). Besides, the postnatal CNS is also capable of glucocorticoid synthesis (55). Importantly, some authors have proposed that steroidogenic enzymes expressed in the fetal unit or in the placenta play a critical role in shaping the offspring’s development and behavior by turning steroid hormones into more polar compounds, unable to bind to nuclear receptors and modulating, instead, other type of receptors (59), as will be discussed in the next section. Figure 1 summarizes a general scheme of steroidogenesis in the CNS as well as different tissues of the maternal-placental-fetal unit.
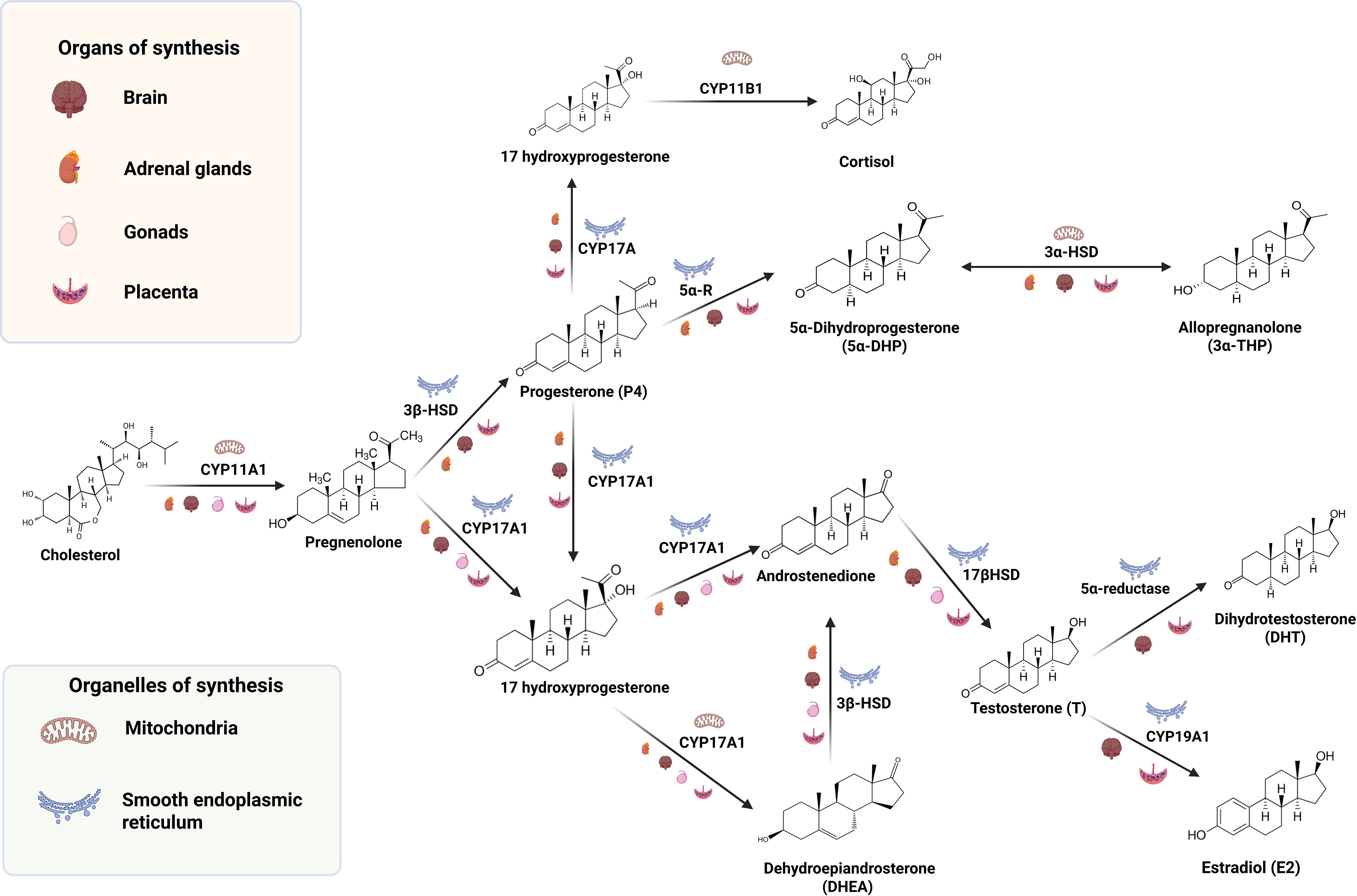
Figure 1. Schematic overview of steroidogenesis within the maternal–placental–fetal unit and developing brain, highlighting enzymes, subcellular compartments, and tissue sources relevant to prenatal and early postnatal neurodevelopment.
3 Neuroactive steroids: mechanisms of action and effects on neurodevelopment
Steroids with neuroactive effects exert their functions through very complex and diverse action pathways. These include both classical and non-classical mechanisms of steroid hormones, which are mediated by intracellular and membrane receptors. Intracellular or classical receptors act as transcription factors regulated by ligand that modulate gene expression, usually over a period of hours to days. These mechanisms influence cellular processes such as cell proliferation, differentiation, and metabolism, among others (5, 8, 42). In contrast, non-classical or non-genomic mechanisms depend on the activation of membrane-bound receptors and activate signal transduction pathways in a time span of seconds to minutes. This leads to changes in cellular functions, membrane cell polarization, calcium influx, and protein phosphorylation (60, 61). Lastly, these non-classical mechanisms can modify gene expression by regulating the activation of transcription factors downstream of signaling cascades. In addition, neuroactive steroids also modulate neurotransmitter receptors. Some of the most studied neurotransmitter receptors include GABA(A) receptors, NMDA, and AMPA receptors, which lead to changes in neural excitability. More recently, it has been proposed that, due to their lipophilic structure, neuroactive steroids regulate the fluidity of cell membranes and receptor trafficking (62). Moreover, crosstalk between steroid mechanisms could occur, as described further in this section.
Intracellular receptors involved in the neuroactive steroids signal transduction are progesterone (PR, isoforms: PR-A, and PR-B), glucocorticoid (GR, with several splice variant isoforms, such as GRα-γ), androgen (AR, with different splice variant), estrogen (ER, subtypes coded in different genes: ERα, and ERβ), and mineralocorticoid receptor (MR, with several splice variant isoforms, including MRα and MRβ). These are part of the nuclear receptor superfamily of transcription factors. Once activated by their ligands, they undergo a conformational change, promoting their release from chaperone proteins and facilitating their dimerization and translocation to the nucleus, where they bind to specific regions of DNA called hormone response elements, to recruit co-regulators, chromatin remodelers, and the transcription machinery [for review (63, 64)]. Besides its ligand activation, the activity and degradation of these proteins via the proteasome are regulated by post-translational modifications, such as phosphorylation, sumoylation, and ubiquitination (6, 65, 66). They also present non-classical mechanisms when located at the lipid rafts of the cell membrane or interacting with caveolin 1, or due to palmitoylation, which enables their polyproline-rich motif at the N-terminal domain of the receptor to interact with other proteins, such as the SRC family kinases (SFK). This interaction leads to the activation of several signaling cascades mediated by MAPK or Jak/STAT pathways (6). Such mechanisms have been described for PR (6), GR (64, 67), AR (4), ER (68), and potentially for MR (69). Although the latter has been better characterized by its interaction with receptor tyrosine kinases such as EGFR, and PDGFR, along with protein-coupled receptors like the G Protein-Coupled Estrogen Receptor 1 (GPER1, also known as GPR30) (70, 71). Steroid receptors and their mechanisms play a crucial role in differentiation, proliferation, and myelination of the nervous system, as well as brain plasticity throughout the lifespan. However, this is more relevant during the fetal and early neonatal periods as discussed forward.
In different types of tissues, including the human fetal tissues, PR-B has been widely associated with higher transcriptional activity, whereas PR-A has more transcription-inhibitory effects, particularly on the PR-B, ERβ, and GR (72). PR-A and PR-B have been identified in oligodendrocyte precursor cells derived from E14.5 mouse embryonic spinal cords, where P4 promotes proliferation in a PR-B–dependent manner. This has been demonstrated in cultured oligodendrocyte progenitor cells treated with the PR agonist R5020, in which silencing of PR-B abolishes the proliferative effect. Also, in these cells, P4 increases the expression of oligodendrocyte progenitor cell markers such as SOX9, NG2, while also inducing the expression of mature oligodendrocyte markers such as MBP and CNP1 (73), indicating that P4 induces oligodendrocyte maturation and myelination. In vitro assays show that along the differentiation process of mouse embryonic stem cells to dopaminergic neurons, they present higher expression of PR, while the expression of Erα decreases over the differentiation process (74). Moreover, the activation of ERα and dopamine D1 receptors induces the overexpression of PR-A in several brain areas of rat neonates (75, 76). This effect correlates with an increase in social play later in life, during the juvenile stage. Additionally, activation of PR and ERα promotes the differentiation of mouse embryonic cells to motoneurons, as demonstrated by in vitro experiments using ER subtype antagonists (8).
In humans, ERβ and GR are highly expressed in human undifferentiated embryonic stem cells and embryoid bodies, compared to Erα (77). GR was also highly expressed in embryoid bodies and the three developmental layers (77). Besides, they have also been identified in several brain areas of chicks, rats, and mice from embryonic to early postnatal life66. In murine models, the expression of ERα and ERβ has been demonstrated to be related to sexual differentiation of the brain. The expression of ERα has been enriched in brain areas related to sexual behavior, such as the hypothalamus, while the expression of ERβ is more broadly expressed throughout the brain [for review about ER effects on neurodevelopment and its effects on behavior, see (78)]. Besides, the estrogen, estetrol, a highly produced steroid during pregnancy, has antioxidant, antiapoptotic, and pro-myelinating effects mediated through ERα and ERβ (79).
Corticosteroids influence neurodevelopment in fetal and early postnatal life through the activation of neurogenesis in a GR-dependent manner and an MR-dependent manner. The GR expression has been reported in all cell types of human cerebral organoids from induced pluripotent stem cells. In such models, treatment with dexamethasone, a GR agonist, enriches the expression of transcription factors associated with neural differentiation like PAX6, FABP7, and NEUROD1 (80). However, it must be mentioned that cortisol at high doses could negatively impact the neurogenesis of dopaminergic neurons at the ventral tegmental nucleus in males but not females (81). Although the key role of glucocorticoids and the GR has been highlighted, it is also important to consider that excessive glucocorticoid levels, such as those seen in excessive maternal stress or obesity, can have detrimental effects on neurodevelopment (82), as we will explore in the next sections.
Regarding MR, this receptor plays a role in the antenatal development of the hippocampus. Its expression is detected in specific fetal brain regions, such as the hippocampus and hypothalamus, at embryonic day E16.5 in rats, shortly before the end of gestation at E22.5 (83). Other animal models, such as guinea pigs, display a different pattern, with MR expression decreasing in the same brain regions near delivery, highlighting a markedly species-dependent regulation of MR expression (84, 85).
Along with the ERs, the AR plays a key role in the sexual dimorphism of the brain, particularly in the brain cortex and the arcuate nucleus of the hypothalamus. In vitro studies in human neural stem cells, treatment with the potent androgen, DHT induced cell proliferation and survival when cells were exposed to nutrient-deprived conditions (86). Additionally, in these cells, DHT upregulated the expression of genes related to ASD (NRCAM, FAM107A, IGFBP5). In humans, RNA-Seq data indicate that the expression of AR in the brain cortex decreases over time from 8 weeks postconceptional to 15–17 weeks postconception (wpc) in both 46XY and 46XX fetuses. Notably, the expression of the 5α-R1 is augmented over the same development period in both males’ and females’ brain cortex. In this study, Buonocore et al. focused on this critical period because of the beginning of testicular production of T at 8 wpc in XY individuals. In such individuals, higher differences were seen in the brain cortex gene expression, attributed to chromosomal differences and Carnegie developmental stages, rather than due to steroid production by the fetal testis (87). Nonetheless, this must be further studied.
Apart from the AR, rapid effect exerted at the cell membrane, several proteins have been proposed as membrane androgen receptors, such as the transient receptor potential melastatin 8 (TRPM8), a calcium channel, or the G protein-coupled receptors GPRC6A, and the Oxoeicosanoid receptor 1 (OXER1), to which T present high affinity and exhibit effects in cancer cells (88). In animal models, their expression has been observed in hippocampal neurons and seems to be crucial for learning and memory through the activation of ERK kinases (89).
Additionally, membrane receptors mediate rapid mechanisms of neuroactive steroids, although they are less understood than those of intracellular receptors. Some of the most studied of these receptors are the five membrane P4 receptors: mPRα, mPRβ, mPRγ, mPRδ, and mPRϵ. They are located at cellular membranes and transduce signals through G-protein activation (60). Also, P4 membrane receptor components 1 and 2 (PGRMC1/2) are membrane receptors located in proximity to mPR and regulate its activity. Such receptors are activated by P4 and its metabolites with different affinities depending on the structure of each mPR type (60). These receptors are differentially expressed in fetal neural tissue, with mPRα and mPRβ being the most highly expressed (90). They mediate the activation of cAMP/PKA, PI3K/AKT, and MAP kinase/ERK1/2, by which mPRs induce neurite outgrowth (90). In zebrafish, mPRγ is involved in neural proliferation, particularly in the olfactory system and the brain shape (91). They also mediate the anti-apoptotic effects of 3α-THP and its synthetic analogs in cell models lacking intracellular receptors, by interacting with the GABA(A) receptor, the main neurotransmitter receptor regulated by neuroactive steroids, whose mechanism of action will be discussed further below (92).
Estrogens also regulate fetal neural development through the membrane receptor (GPER) (61), which is highly expressed in the rodent brain, particularly in the adult brain. However, there are very few studies about their relevance in fetal neurodevelopment, especially in humans. Similarly to mPRs, GPER activates G proteins and mediates neurite, axon, and dendrite growth in E18 hippocampal mouse neurons (93, 94). In mice, GPER participates in preventing neural cell death, astrocyte viability, and expression of inflammatory cytokines when cells are treated with high doses of glutamate (88, 95).
The GABA(A) receptor is a ligand-gated ion channel regulated by neuroactive steroids (96, 97). Its chemical structure is critical in determining the type of regulation that neuroactive steroids will exert on GABA(A) receptor. For example, 3α-steroid metabolites positively modulate it, while 3β-steroid metabolites negatively regulate it (98). The most important positive regulators of GABA(A) receptor are 3α-THP, allotetrahydrodeoxycorticosterone, androstanediol, and 17α-OH allopregnanolone (99). In human neural stem cells and rat hippocampal neuroprogenitor cells, the co-expression of GABA(A) receptor and the Na+-K+-2Cl- cotransporter SLC12A2, promotes the efflux of Cl-, which induces depolarization of the cell membrane, and the subsequent activation of voltage-dependent L-type calcium channels, the activation of PKA kinases, and the expression of cell cycle regulators, lastly inducing proliferation (98, 100, 101). Through these mechanisms, GABA(A) receptors and neuroactive steroids promote the increase in cell number in proliferative brain zones such as the embryonic cortex and the ventricular zone (101–103).
P4 plays a crucial role in sustaining pregnancy. Research involving pregnant rats shows that maternal P4 can cross the fetal bloodstream and affect the developing CNS. Also, as previously mentioned, progestogen neuroactive steroids begin to be synthesized in the placenta and locally in the fetal nervous system from the very early stages of life (5). Therefore, it has been suggested that P4 and its close derivative, 3α-THP, are crucial for proper neurodevelopment in mammals, serving specific functions in this process (1, 5). For several years, it has been noted that P4 has neuroprotective effects in the CNS, and its therapeutic use has even been clinically investigated for conditions such as traumatic brain injury (TBI), spinal cord injury (SCI), and epilepsy (104–107).
In the context of neurodevelopment, it is noteworthy that the newly formed CNS is highly vulnerable to injury from intrauterine complications, neonatal hypoxia-ischemia, and excitotoxicity, leading to neuronal death and impaired neurodevelopment. This can result in permanent neurological deficits and neonatal death (31, 106, 108). In this way, P4 and 3α-THP have been shown to mitigate CNS damage in rats and sheep shortly before and after birth by regulating cell proliferation and reducing inflammation and apoptosis during traumatic injury events and birth asphyxia (96, 97, 106, 109). Furthermore, in newborn rats, P4 treatment lessened inflammation, with fewer activated microglia in the cortex and hippocampus following hypoxia-induced brain injury. Treated pups also showed better motor and cognitive recovery by three weeks post-injury (106). Similarly, in newborn mice, P4 reduced inflammation and induced neuronal survival, suggesting that its effects involve regulating pro-inflammatory cytokines (108, 110). Interestingly, disruption of 3α-THP synthesis in sheep fetuses increases apoptosis and astrocyte proliferation in the brain (3, 96). Given that GABA is an inhibitory neurotransmitter, 3α-THP is believed to maintain a suppressive state in fetal brain neuronal activity during late gestation to prevent excitotoxicity, as demonstrated in sheep. This effect dampens fetal brain activity and helps to regulate fetal sleep-like behaviors, which are crucial for proper neurodevelopment (40, 111). Excitatory effects of GABA(A) receptor have also been characterized in neural progenitors of rats and humans, in which 3α-THP induces hyperpolarization of the cell membrane and induces cell proliferation (100, 112, 113). Besides, 3α-THP promotes the migration and maturation of Schwann cells through its modulatory effect on GABA(A) receptor and the activation of SRC and FAK kinases (114, 115).
Apart from its neuroprotective and neuromodulatory effects, 3α-THP is also strongly associated with birth outcomes in humans and with the neurodevelopment of the newborn in an indirect manner, since pregnancy is characterized by an increase in the levels of P4 and 3α-THP in the maternal plasma due to the placental synthesis (1, 9). A decrease in these neuroactive steroids after delivery has been associated with the development of the maternal psychiatric condition known as postpartum depression (116, 117), which also impacts the behavioral and cognitive development of the newborn (118). Importantly, it has also been shown that placental 3α-THP insufficiency in mice leads to cerebellar myelination impairment after birth, which correlated with autistic-like behavior in male offspring (119). P4 has been involved in neurodevelopmental myelination, particularly in the early postnatal cerebellum of rodents (120, 121). It is also reported that P4 could be involved in oligodendrocyte differentiation in the mouse embryonic spinal cord (73). Likewise, P4 increases mature oligodendrocyte survival after white matter injury induced by chronic hypoxia in neonatal rats (122).
Androgens have been largely associated with brain development, particularly in the first and second decades of life in humans. The evidence shows that androgens, especially DHEA, play a crucial role in brain cortex maturation in an age- and sex-specific manner during middle childhood and adolescence (123). Interestingly, this group of neuroactive steroids also has effects during fetal and early postnatal stages of neurodevelopment. One of the key roles in fetal neurodevelopment is its involvement in brain sexual differentiation, as high levels of this androgen contribute to brain masculinization (124). Additionally, T and its immediate metabolite, DHT, have been shown to influence the maturation of hippocampal neural circuits in a sex-specific manner during the first few postnatal days in rats. Notably, T also contributes to the masculinization of this brain structure since its neonatal administration to females masculinizes spatial memory performance, a function largely associated with the hippocampal circuitry (125–127). The effects of T and DHT on cortical and hippocampal maturation are presumed to be due to the regulation of neurogenesis, neurite growth, and synaptic density (128, 129). In support of this, it has been demonstrated that DHT upregulates the expression of genes associated with proliferation and cell survival in human embryonic neural stem cells (86).
In recent years, growing evidence has positively linked androgens to the early onset of human neurodevelopmental conditions. Particularly, ADHD and autism have been related to prenatal exposure to high levels of T, since these conditions are more frequent in males (86, 130–132). Since T and DHT are involved in synaptic density and neural circuitry maturation, it is hypothesized that their high levels in the maternal-fetal circulation dysregulate gene expression associated with the neuronal events, leading to an excitation-inhibition imbalance in the developing brain (133).
Regarding estrogens’ actions during early neurodevelopment, E2 has been shown to play a critical role in brain development and function, influencing neurogenesis, synaptogenesis, and neuroprotection (134). The enzyme aromatase is widely expressed in the forebrain of primates (including humans), and although estrogens are more associated with female functions, the expression of this enzyme in the CNS does not depend on plasma E2 levels (135, 136). Moreover, E2 levels and aromatase expression in the fetal brain have been associated with adequate cortical development in rats and mice (134, 137). Particularly, evidence from ERβ knockout mice shows that E2 influences the migration and survival of cortical neurons at late prenatal stages (138). Also, in vitro studies have shown that E2 promotes the proliferation of neural stem cells isolated from rat embryonic telencephalon and oligodendroglial cells (139). Additionally, the knockdown of ERβ in mouse embryonic neural stem cells reduces the number of progenitors of mesencephalic dopaminergic neurons that originate from them (140). Likewise, E2, through the GPER, stimulates neurite outgrowth in hippocampal neurons from the E17 mouse fetal brain (141), indicating that it also plays a role in neurogenesis and the maturation of neural circuits in different brain structures.
To make masculinization effective at the molecular and cellular levels, aromatization of T to E2 is required (142). Then, E2, through the ER, changes gene expression patterns to influence the number of neurons and shape of dendritic spines in the most sexually dimorphic brain regions, including the preoptic area and the hippocampus (143, 144). Importantly, Vacher et al. recently reviewed the significance of sex steroid levels, especially androgens, and their association with neurodevelopmental conditions such as autism in different cohorts. In such studies, higher levels of androgens in mothers with polycystic ovaries were associated with a higher risk of autism in males. Also, they remark on the relevance of neuroactive steroids synthesized by the placenta at the very early developmental stages as major regulators of neurodevelopmental processes (1).
Glucocorticoids exert key functions in the CNS and play a fundamental role in pregnancy at late stages. Maintaining glucocorticoid levels is so important that they are administered to women at risk of preterm delivery, since it favors pulmonary maturation (145, 146). Besides, premature birth is often associated with a disruption of proper neurodevelopment, leading to conditions such as cognitive impairment, cerebral palsy, and speech deficits (147). Notably, maternal cortisol and corticosterone disruption have been related to these conditions as they regulate neurogenesis, neuronal migration, plasticity, and neurotransmitter activity (147, 148). It has been reported that physiological levels of cortisol stimulate the proliferation of human hippocampal neural progenitors, while high levels associated with stress conditions can inhibit the differentiation of these cells (149). In the same way, elevated levels of corticosterone due to a stress response affect the proliferation and differentiation of neural stem cells during embryonic and adult neurogenesis (150, 151). Additionally, it has been suggested that high levels of maternal glucocorticoids associated with stress conditions during pregnancy affect proper neurodevelopment by altering the maturation of neural circuits, impacting the individual’s behavior after birth and throughout life (152–154).
All functions mediated by neuroactive steroids, highlight the relevance of these molecules in understanding how their dysregulation, regardless of the underlying cause, could compromise the proper development of the CNS. Figure 2 presents a summary of the most important biological events regulated by neuroactive steroids involved in neurodevelopment during prenatal and early postnatal life. In the following section, we will discuss in greater detail how obesity and overweight can alter neurosteroid action and, consequently, affect early CNS development.
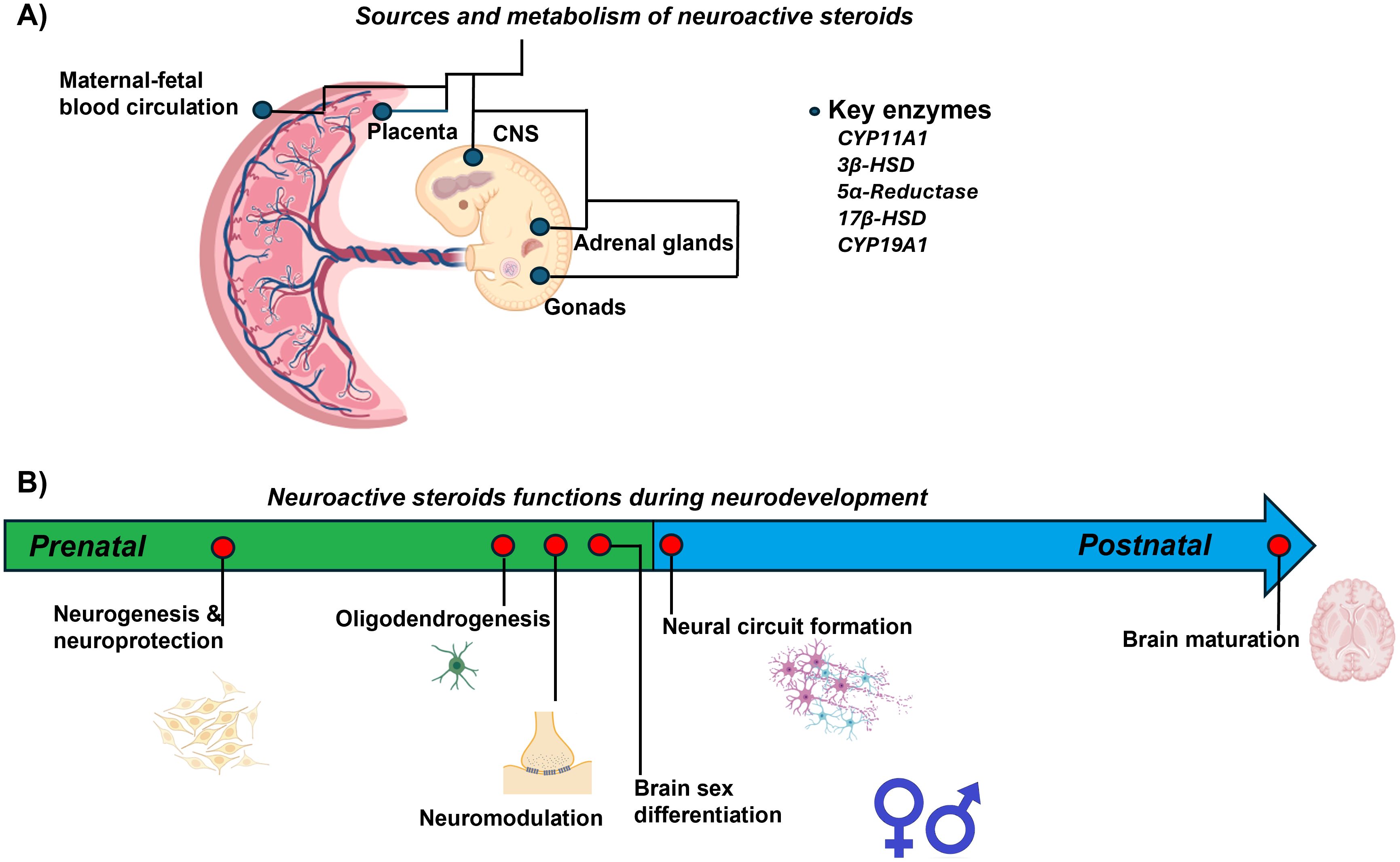
Figure 2. Summary of neuroactive steroid actions during prenatal and early postnatal neurodevelopment. (A) Sources and metabolism of neuroactive steroids in the maternal-placental-fetal unit. (B) Neuroactive steroids function during neurodevelopment. The arrow indicates events regulated by neuroactive steroids during fetal and early postnatal life.
4 Neurodevelopmental outcomes in offspring of mothers with overweight and obesity
Maternal overweight and obesity impact the neuroendocrine, metabolic, and inflammatory regulation systems in women. These conditions, particularly during pregnancy, predispose to adverse reproductive health outcomes, including infertility, gestational hypertension, preeclampsia, gestational diabetes, postpartum hemorrhage, emergency cesarean deliveries, and both intrauterine growth restriction and macrosomia. Overweight and obesity in pregnant women can influence the developing fetal CNS through various mechanisms, encompassing hormonal, metabolic, inflammatory, and epigenetic pathways, leading to early adverse CNS development outcomes and mood disorders in their offspring (155–158). Outcomes differ depending on whether the exposure is pregestational obesity or excessive gestational weight gain.
4.1 Outcomes associated with pregestational overweight and obesity
Preclinical models show that maternal obesity prior to conception alters fetal brain structure, connectivity, and behavior. Studies in mice have shown that pregestational obesity leads to a reduction in hippocampal cortical thickness and an increase in the soma and branches of astrocytes in the offspring (159). Other studies hypothesize that obesity in the offspring is predisposed by pregestational maternal obesity due to changes in the food-intake circuits innervation. In this line, offspring of pregestational obese female mice present less innervation of neuropeptide Y neurons to the paraventricular nucleus and less neurite growth in the arcuate nucleus of the hypothalamus at gestational day 17.5 compared to offspring of pregestational normal weight mothers. Such nuclei are critical hubs for adipokine signaling of hunger and satiety (160). Also, the offspring of these mothers showed a significantly increased density of GFAP-positive astrocytes in the arcuate and supraoptic nuclei of the fetal hypothalamus. This increase may indicate disrupted hypothalamic programming induced by the intrauterine inflammatory environment (161). In addition, mouse offspring of pregestational obese females present early deficits in communication and olfactory discrimination, impaired social play behavior, and reduced expression of synaptopodin, a protein involved in synaptic plasticity in the hippocampus and the prefrontal cortex (162).
In rats, maternal pregestational obesity has been associated with an increased incidence of congenital CNS malformations, including neural tube defects and microcephaly. These conditions reflect disruptions in early neurodevelopment (163). Besides, elevated glucose levels and insulin resistance, which are usual comorbidities along with obesity, have also been reported to alter brain development and neuronal maturation in rats (164). Also, in these animals, obesogenic diets have been shown to cause deficits in cognitive functions along with damage in the hippocampus and the prefrontal cortex (165). It is important to consider the time of exposure to this obesogenic diet, for example, several months before pregnancy vs only during pregnancy, also have different effects, which are explored in other reviews (165).
Pregestational overweight and obesity in humans have been more consistently linked to structural and functional alterations, including increased rates of behavioral symptoms, emotional dysregulation, and psychiatric disorders. Maternal pregestational body weight is also associated with the development of the autonomic nervous system, evaluated in utero by measuring the heart rate variability at trimesters 1 and 3. In this study, the heart rate variability was inversely correlated with the maternal body weight (166). High maternal BMI at the beginning of pregnancy has also been associated with poor inter-hemispheric communication, poor functional connectivity between the prefrontal cortex and the anterior/inferior insula of the left frontal gyrus in the fetal brain (167), and reduced cortical thickness in key neonatal brain regions, such as the left frontal lobe (168).
Pregestational maternal obesity has also been reported to impair skills such as memory, attention, inhibitory control, and problem-solving (164). Also, some studies have linked maternal pregestational overweight and obesity with lower cognitive, linguistic, and motor skill scores in children at 2 years of age (169, 170). A meta-analysis of 41 studies from 36 perinatal cohorts—including populations from the United States, the Netherlands, the United Kingdom, Denmark, Finland, Sweden, Australia, and Norway—reported that children of women with pregestational overweight had a 17% higher likelihood of neurodevelopmental disorders, while those of women with obesity showed a 51% increased risk compared to children of mothers with normal weight (171). More specifically, the meta-analysis identified elevated risks for attention-deficit/hyperactivity disorder (62%), autism spectrum disorder (36%), cognitive delays (58%), and emotional or behavioral problems (42%). Other cohort studies have linked maternal obesity before conception to lower performance in cognitive domains such as practical reasoning at 36 months of age (153) and verbal recognition at 9 years (172).
In summary, there are global and homogenous data of preclinical and clinical studies indicating that maternal pregestational overweight and obesity alter signaling, connectivity, and CNS structure during fetal development, and increase the risk of different neurodevelopmental conditions such as ADHD, autism spectrum disorders (ASD), eating disorders, anxiety, and depression in their offspring later in life (156, 164, 165). In Table 1, we present a summary of clinical studies regarding the effects of pregestational overweight and obesity on anatomical changes and poor neurodevelopmental outcomes. These findings consistently indicate that maternal pregestational overweight or obesity are associated with adverse neurodevelopmental outcomes in offspring. Overall, elevated maternal adiposity appears to affect brain structure, cognitive performance, and behavioral regulation during early life and childhood, potentially through inflammatory, metabolic, and microbiome-related pathways. However, heterogeneity in study design, sample size, and neurodevelopmental assessments, along with differences in the age of participants in these studies, limits the comparability of existing data. Future research should aim to delineate critical windows of vulnerability, elucidate the underlying biological mechanisms, and determine whether targeted nutritional or metabolic interventions during pregnancy can attenuate these adverse outcomes.
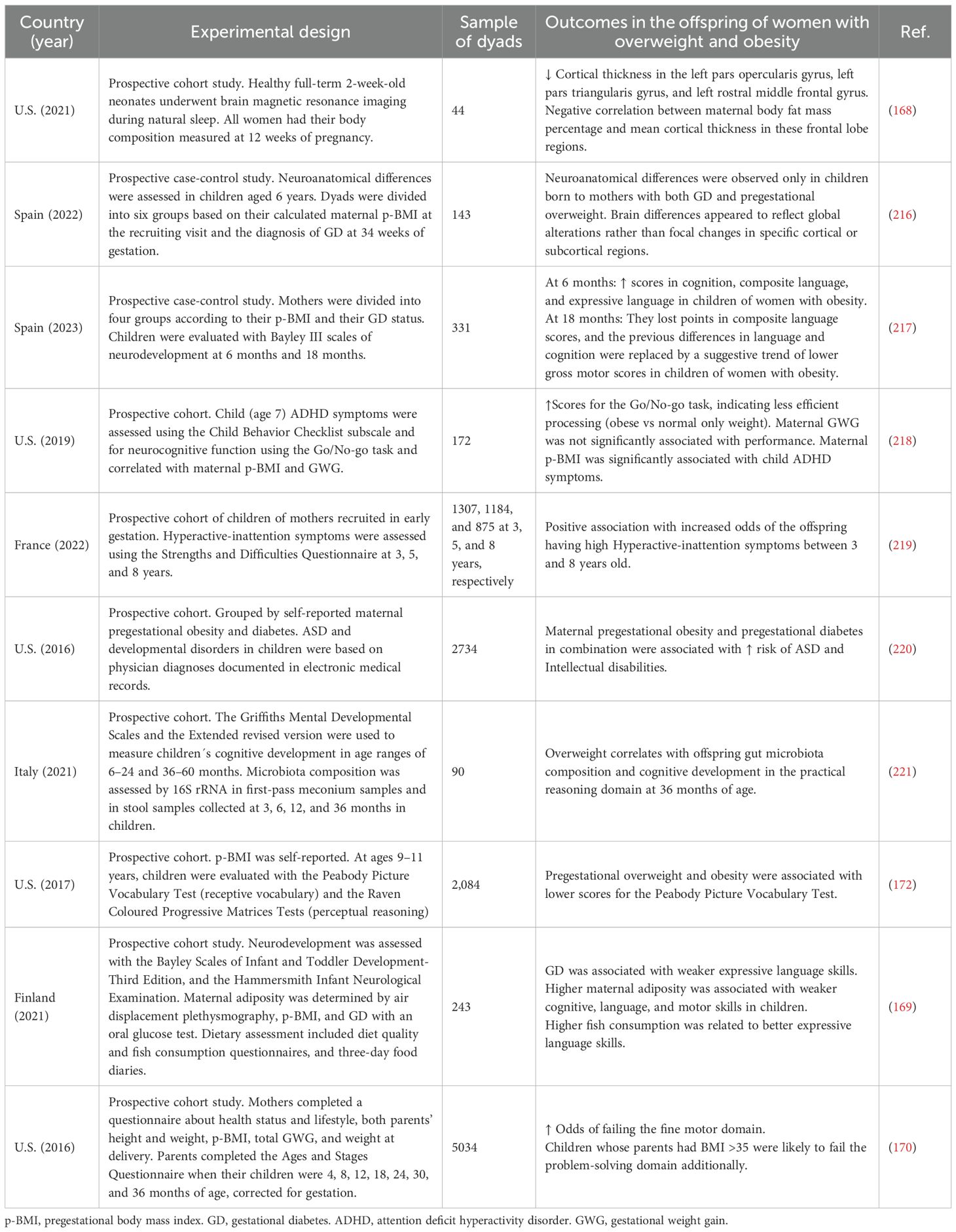
Table 1. Summary of clinical studies on neurodevelopmental outcomes in offspring of mothers with pregestational overweight or obesity.
4.2 Outcomes associated with excessive gestational weight gain
Gestational obesity is typically reflected by weight gain above Institute of Medicine guidelines. Few information has been described regarding neurodevelopmental outcomes and excessive weight gain during pregnancy, which is a complex condition, influenced not only by an increased food intake and a poor quality of diet, but also includes pregestational overweight, frequent cravings, less exercise, and psychosocial factors such as depression and body image dissatisfaction (173–175). Excessive weight has been mostly associated with outcomes related to body composition rather than neurodevelopmental disorders. Regarding body composition, the most consistent studies have demonstrated the association of excessive weight gain and the outcomes of large-for-gestational age and macrosomia (157, 176–178). About neurodevelopmental outcomes, most studies consistently show a positive association between excessive weight gain and ASD, as summarized in Table 2. Clinical evidence from multiple prospective and case–control studies suggests that excessive gestational weight gain (GWG) and maternal overweight or obesity during pregnancy are consistently associated with an increased risk of adverse neurodevelopmental outcomes in offspring, as summarized in Table 2. Reported outcomes include a higher incidence of global neurodevelopmental disorders (NDD), autism spectrum disorder (ASD), attention deficit hyperactivity disorder (ADHD), and intellectual developmental disorder (IDD), as well as subtle impairments in cognition, language, and executive functioning. Excessive GWG—particularly when superimposed on pregestational overweight or obesity—emerges as a consistent and independent risk factor for suboptimal neurodevelopmental outcomes. Although causality cannot yet be firmly established, the reproducibility of associations across diverse populations and study designs highlights the potential importance of gestational weight management as a modifiable target for the prevention of neurodevelopmental disorders. Future longitudinal and mechanistic studies integrating metabolic, inflammatory, and neuroimaging biomarkers are needed to elucidate causal pathways and identify critical windows for effective intervention.
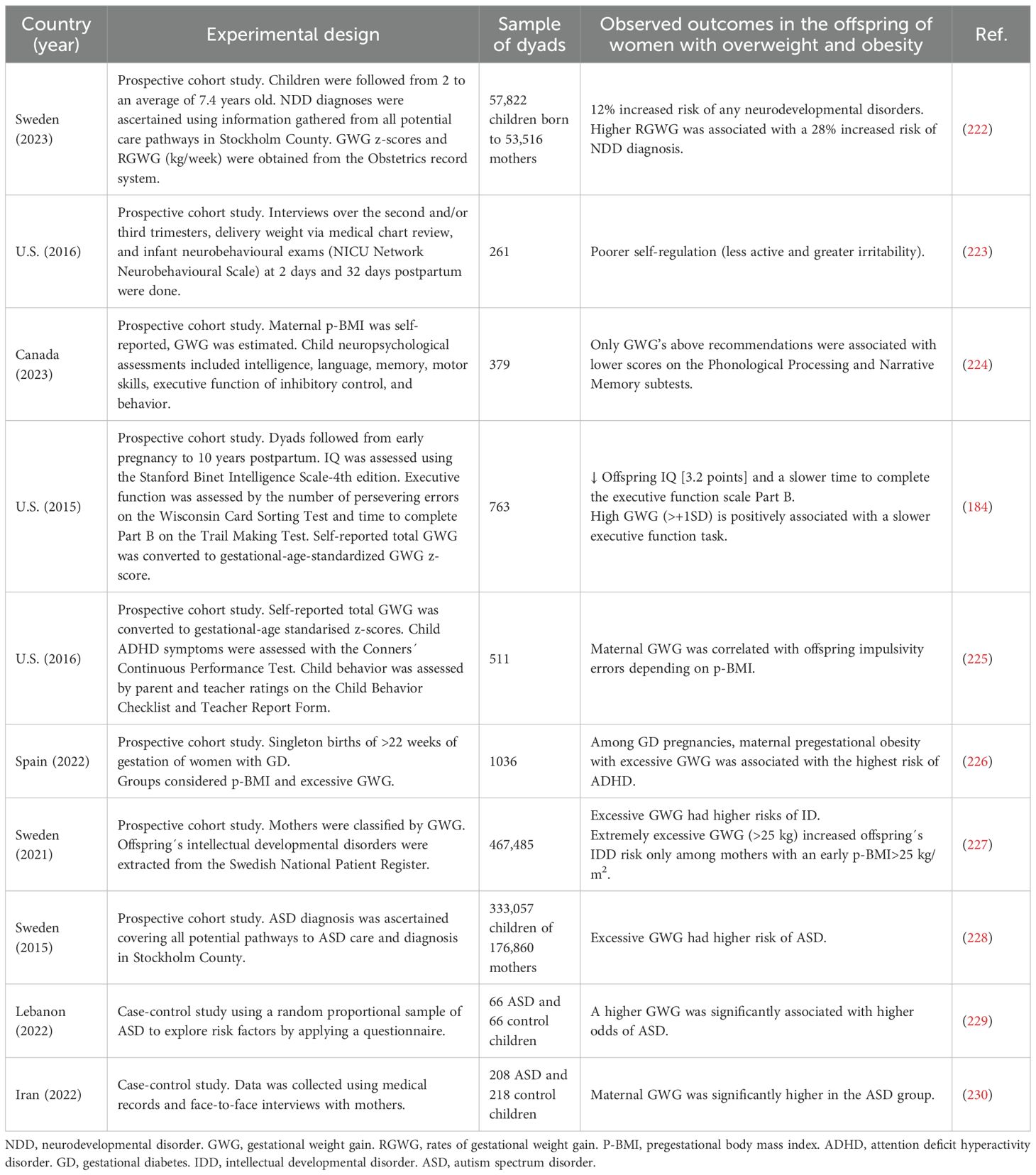
Table 2. Summary of clinical studies on neurodevelopment outcomes in the offspring of mothers with gestational overweight and obesity.
In animals, high-calorie, or high-fat diets administered only during pregnancy are often used as models of gestational obesity. In mice, a gestational obesity model fed from the preimplantation to lactation with a high-fat diet, induced a higher density of immature neurons in males and a lower density of mature neurons in the dentate gyrus with an increased density of astrocytes and microglia in the hippocampus of both male and female adult offspring (179). In rats, this type of diet induces in the offspring, atrophy in hippocampal pyramidal cells, primarily affecting basal dendrites, while apical dendrites remain largely unaffected. It is suggested that elevated leptin levels during early development contribute to the observed dendritic morphological alterations, as leptin plays a neurotrophic role in brain development (180). In this model, gestational obesity also impairs memory, as evaluated by conditioned pain aversion assays in the offspring (180).
4.3 Neurotransmitter signaling pathways affected by maternal obesity
Some of the mechanisms altered by maternal obesity are related to dopaminergic and serotonergic (5-HT) signaling systems. In mice, sex-specific responses in the regulation of dopamine-related genes in fetuses have been found. Male fetuses showed increased expression of the dopamine receptor D2 (DRD2) in the endocannabinoid system within the prefrontal cortex, without changes in dopamine metabolites. In female fetuses, DRD2 expression decreased while homovanilic acid, the final product of dopamine metabolism, increased, suggesting enhanced dopamine metabolism (181). In another study, mice exposed to a high food diet in utero demonstrated dysregulation of the dopamine reuptake transporter, which is associated with impairments in eating behaviors and reward response to food, and its association with schizophrenia, ASD, and ADHD has also been demonstrated (155).
Regarding the role of 5-HT system in offspring of obese women, it has been shown that a high-fat diet reduces 5-HT synthesis in offspring, associated with reduced serotonergic axonal density and embryonic neuronal survival in human brain regions critical for behavioral regulation, and contributes to an increased risk of neurodevelopmental pathologies such as ADHD, ASD, anxiety, and depression. In mice, it has been described that maternal high-fat diet induces an increase in the offspring hippocampal levels of the enzyme TPH2 involved in 5-HT synthesis. Besides, in humans, methylation of the 5-HT receptor 2A gene (HTR2A) promoter, has been associated con with psychiatric conditions such as schizophrenia and bipolar disorder effects in four analyzed cytosines in the promoter region, located – 166 (L1), - 1439 (L2), -1421 (L3) and –1224 (L4), particularly some specific genetic polymorphisms (rs6311 and rs6306). It has been reported that higher pregestational BMI is associated with lower HTR2A promoter methylation in placentas of male fetuses of the (182).
An additional indicator of synaptic overexcitation is glutamate levels, which are also affected by maternal obesity, with increased concentrations observed in mouse fetuses in the endocannabinoid system within the prefrontal cortex (181). Furthermore, maternal high-fat diets have been shown to alter fetal brain glutamate metabolism. In this context, levels of glutathione disulfide (GSSG)—the oxidized form of glutathione that also interacts with NMDA and AMPA glutamatergic receptors—are decreased in fetal brains of mothers that received a high-fat diet. This reduction may have behavioral implications, given that glutamate is the main excitatory neurotransmitter in the CNS (183). In the same study, the expression of Tbr1—a gene typically expressed by excitatory glutamatergic neurons in the amygdala and involved in regulating the expression of neuronal activation markers—was decreased in response to maternal high-fat diets (184). This is further supported by a mice model study, which reported that in the context of maternal high-fat diets, Mecp2 was detected in the amygdala of male offspring and was associated with upregulation of Tbr1. Tbr1 is normally repressed by Mecp2 during brain development, a regulatory mechanism that is critical for proper neuronal differentiation and neurite formation. Mutations or dysregulation of Mecp2 have been linked to several neurodevelopmental disorders. In line with this repressive role, previous studies indicate that reduced Mecp2 activity leads to increased Tbr1 transcription. Consistent with these findings, the authors observed elevated Tbr1 mRNA levels and a higher number of Tbr1-expressing cells within the basomedial (BM) nucleus of the basolateral amygdala in offspring of mothers that received a high-fat diet (185). Additionally, the expression of glutamate receptors such as GluR7 and the glutamate ionotropic receptor NMDA type subunit 2A (Grin2a), as well as scaffolding proteins such as Shank2, critical for synaptic plasticity, are altered in such model of obesity (183). The increased baseline glutamatergic activity in the absence of behavioral stimuli suggests an imbalance in neuronal excitation, which should predispose offspring to behavioral alterations, particularly anxiety, ASD, and ADHD (183, 185).
Although pregestational and gestational obesity are distinct, they are often overlapping conditions. For example, women with overweight or obesity also tend to present excessive weight gain during gestation (173–175). As mentioned in this section, both factors could independently and additively increase the likelihood of adverse outcomes. However, pregestational obesity often has a stronger and more persistent effect on offspring’s health. Moreover, research about the effects of excessive weight gain during pregnancy is needed.
5 Insights into neuroactive steroid status in the maternal-placental-fetal unit in overweight and obesity
As mentioned before, beginning pregnancy with a condition of overweight or obesity, and its associated comorbidities such as hypertension, insulin resistance, type 2 diabetes, or polycystic ovary syndrome, are associated with an increased, chronic pro-inflammatory state and an altered metabolic environment, rather than a more acute state of metabolic adaptation to gestational obesity. There is an interplay between energy homeostasis and neuroactive steroids´ functions. In this line, overweight and obesity are conditions characterized by excessive energy storage, as mentioned before, and include alterations in the local and peripheral effects of neuroactive steroids. Importantly, neuroactive steroids are modulators of the hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes, as well as mood and sexual behavior modulators. They are also regulators of energy storage in adipose depots, homeostasis of reproductive function, among others, and altered levels and signaling of neuroactive steroids in female overweight and obesity have been broadly described (186–188). They also influence alterations in steroid hormone levels in the maternal-placental-fetal unit, as described in this section. Importantly, an emerging trend in research suggests a potential distinction in the neurodevelopmental outcomes associated with maternal obesity, depending on the timing of exposure: pregestational obesity or excessive weight gain during pregnancy, as mentioned before.
Maternal obesity is increasingly recognized as a state of chronic low-grade inflammation, or metainflammation, driven by altered metabolic and immune regulation (189, 190). During pregnancy, this inflammatory milieu is further exacerbated, exposing the fetus to an intrauterine environment enriched in cytokines and metabolic stressors that can disrupt key neurodevelopmental processes. Women with obesity frequently exhibit elevated levels of IL-6, IL-8, TNF-α, IFN-γ, and CRP, along with altered adipokine profiles, which can impair placental functions such as trophoblast invasion, nutrient transport, and cytokine signaling (190). These alterations compromise the maternal–fetal interface, allowing proinflammatory mediators to influence fetal brain development.
Neuroinflammation and oxidative stress have been implicated in microglial activation, lipid peroxidation, and cytokine overexpression in offspring (189). Inflammatory mediators such as pJNK and TNF-α can disrupt brain-derived neurotrophic factor (BDNF) metabolism and tryptophan hydroxylase-2 (TPH2) expression—key regulators of hippocampal neurogenesis and serotonin synthesis. Furthermore, maternal inflammation enhances placental serotonin production, which has been linked to impaired axonal growth and abnormal neuronal connectivity (189). It has also been suggested a complex interplay between immune signaling and neuroactive steroids, since steroid hormones such as P4, E2, T, cortisol, DHEA, and 3α-THP are key immunomodulators whose effects as pro- or anti-inflammatory often depend on their concentration (191–194). Collectively, these findings indicate that the maternal proinflammatory state in obesity reshapes placental signaling and fetal brain development, predisposing offspring to long-term neurobehavioral vulnerabilities. Neuroactive steroids may exert a bidirectional modulatory role that remains to be fully elucidated.
Research about the role of steroids in fetal programming is increasing, particularly in the new field of neuroplacentology, which studies how placental factors influence both normal and altered perinatal outcomes, such as preterm birth, fetal growth restriction, and associated poor neurodevelopmental outcomes (1, 119, 195). This is crucial for species adaptation because it has been shown that maternal steroid hormones are rapidly metabolized by fetal-placental tissues to shape the fetal brain and modulate neurodevelopment and behavior as an adaptive consequence to the exposure of maternal steroid hormones, and independently of its classical effects mediated by their nuclear steroid hormone receptors (1, 59, 196). Despite this, very few studies have focused directly on the influence of maternal overweight and obesity on the maternal-placental-fetal unit neuroactive steroids metabolism, their signaling and its consequences on fetal neurodevelopment.
Pregestational maternal overweight and obesity are linked to significant changes in fetal steroid hormone synthesis. A prospective cohort study of Lassance et al., which included 144 pregnant women with obesity and 90 normal weight ones, showed decreased plasmatic levels of P4 and E2 at 38.5–40 weeks of pregnancy prior to their scheduled C-section. Importantly, in this study, the mitochondrial fraction of the placentas showed significantly lower levels of the TSPO protein, involved in the cholesterol transport to the mitochondrion, a rate-limiting step in the biosynthesis of neuroactive steroids, along with lower cholesterol levels in the pregestational obesity group compared with the normal weight one (197). A case-control study was conducted to evaluate the umbilical cord metabolome of neonates born to mothers with either normal weight (n=32) or overweight/obesity (n=33). In this study, the levels of 23 steroid metabolites were significantly lower in the umbilical cord of neonates in the pregestational overweight/obesity group compared with normal weight mothers. Among the most relevant in terms of their potential as neuroactive steroids were T, 7α-hydroxytestosterone, corticosterone, 11-deoxycortisol, and cortisol (198). Moreover, in a study of 80 women presenting gestational diabetes mellitus (BMI < 25 Kg/m2: n = 56; BMI > 25 Kg/m2: n = 24) and 79 pregnant women without diabetes (BMI < 25 Kg/m2: n = 67; BMI > 25 Kg/m2: n = 25) at 23–26 gestational weeks218, the diabetes group presented lower serum levels of pregnenolone, P4, E2, and estrone than the normal-weight group. Besides, pregnant women with obesity and diabetes mellitus present a higher ratio of E3/E2 and a decreased ratio of estrone/androstenedione (199).
Glucocorticoids, particularly cortisol, are one of the most extensively studied neuroactive steroids associated with poor neurodevelopmental and behavioral consequences in the offspring. Obesity deregulates the maternal and fetal hypothalamic-pituitary-adrenal axis, contributing to altered levels of cortisol (155). Consequently, maternal obesity influences fetal brain development by augmenting fetal cortisol levels, particularly in male offspring in baboons (200). Cortisol concentrations may vary depending on the anatomical site and the species. However, evidence indicates that women with high BMI early in pregnancy present lower concentrations of cortisol and corticosterone in umbilical cord blood. This imbalance is linked to altered activity of the enzymes 11β-HSD1, which synthesize cortisol, and 11β-HSD2, which inactivates it, potentially affecting glucocorticoid transfer and metabolism in the placenta (201). Also, a systematic review found that maternal obesity reduces the activity of the placental 11β-HSD2, which diminishes fetal cortisol metabolism into a less active metabolite, cortisone. This reduction in cortisol metabolism may increase fetal exposure to cortisol (202) and alter the programming of the hypothalamic-pituitary-adrenal axis, impacting neuroendocrine development (202, 203). In preterm birth, which is highly associated with poor neurodevelopmental outcomes, cortisol levels are augmented. In a nested case-control study within the framework of the Healthy Start Study cohort with paired healthy controls and preterm delivery cases, there was a significant association with higher cortisol levels at weeks 13 to 22, and preterm birth (204).
As mentioned before, 3α-THP is associated with neurodevelopment and child behavior (1, 59). However, its metabolism is understudied in the context of pregestational obesity and neurodevelopmental outcomes in the offspring. In a study with 9 women diagnosed with polycystic ovary syndrome and 24 age-matched controls (16 with normal weight and 8 with overweight), it was observed that women with overweight and polycystic ovary syndrome presented higher 3α-THP concentrations than controls with normal-weight (205), suggesting that obesity modifies 3α-THP metabolism in women with comorbidities associated with obesity.
It is hypothesized that 3α-THP plays a role in the pathophysiology of obesity per se since it regulates GABAergic signaling involved in feeding regulation at the hypothalamus (206). In male rodents, P4 and 3α-THP induce food intake. This aspect has not been evaluated in female rodents along the estrous cycle (207, 208). Articles published in the 90s indicated that food intake increases in women during the luteal phase, which correlates with the period of the highest P4 and 3α-THP leves (206). However, in women with obesity, there are contradictory reports regarding this hypothesis about the role of 3α-THP in the etiology of obesity. Some articles have reported that 3α-THP levels are low in women with obesity, while T is elevated when compared to normal healthy ones (209), which is also related to a higher susceptibility for these women to present mood disorders such as anxiety or depression throughout life, including postpartum (118).
Notably, one study indicates that women with very low levels of 3α-THP beginning pregnancy or with high 3α-THP levels during the third trimester and with a rapid drop near birth have been associated with higher depressive symptoms at the postpartum (210). However, this study only includes women with a normal weight, so it is important to explore this in women with obesity. Conversely, another study found a negative correlation between 3α-THP maternal blood levels and BMI in the first trimester. This effect was also observed for women who smoked in the first trimester and those who developed pregnancy-induced hypertension (211). In this study, there was no correlation between 3α-THP levels and depressive symptoms. A Swedish longitudinal study of 56 women reported that 3α-THP determined at gestational weeks 12 and 35 was slightly positively correlated with the weight increase during pregnancy. In a group with higher weight gain (≥ 11 Kg) there were significant higher 3α-THP levels compared to the < 11 Kg group (212).
These data indicate that 3α-THP imbalances are related to maternal mood disorders, especially during pregnancy and postpartum. Research in this topic has led to the development and FDA approval of brexanolone, a pharmaceutical form of 3α-THP as a treatment for postpartum depression. Besides, other studies indicate that such imbalances in 3α-THP levels could be related to the weight composition of the mother (212).
Regarding the offspring’s health, little has been explored about the role of 3α-THP in dyads of women with pregestational overweight or obesity. Since preclinical studies have indicated that 3α-THP is involved in fetal neurodevelopment, some authors have hypothesized that low levels of 3α-THP during fetal life are related to a higher risk of ADHD, ASD disorders, or anxiety during childhood and adolescence (213). Some conditions that are associated with a lower fetal exposure to 3α-THP and other neuroactive steroids are high exposure to maternal stress, increased cortisol levels, or preterm birth (204). Although this hypothesis has been broadly evaluated in models such as guinea pigs (214, 215) it has not been evaluated in clinical research nor in the context of maternal pregestational obesity.
6 Conclusions
Maternal overweight and obesity clearly increase the risk of adverse neurodevelopmental and neuropsychiatric outcomes, including ASD, ADHD, cognitive delays, and mood disorders in offspring, which can persist throughout life. Neuroactive steroids—central regulators of neurogenesis, myelination, synaptic formation, and immune modulation—represent a plausible mechanistic link between maternal metabolic status and offspring brain development.
Although animal models provide strong evidence that maternal obesity alters steroidogenesis and receptor signaling, human data remain scarce and inconsistent, particularly regarding allopregnanolone and other pregnane derivatives. Addressing these gaps requires integrative approaches combining maternal endocrine profiling, placental and cord blood metabolomics, and longitudinal neurodevelopmental follow-up.
The emerging field of neuroplacentology provides a conceptual framework to explore how maternal metabolic conditions reprogram fetal neurodevelopment via placental steroid metabolism. Advancing this field could inform novel biomarkers, preventive strategies, and even therapeutic interventions to mitigate adverse neurodevelopmental trajectories in children exposed to maternal obesity.
Author contributions
CZ-S: Conceptualization, Investigation, Writing – original draft, Writing – review & editing. JG-O: Investigation, Visualization, Writing – original draft, Writing – review & editing. JM-O: Investigation, Visualization, Writing – original draft. MV-S: Data curation, Investigation, Writing – review & editing. IC-A: Funding acquisition, Validation, Writing – review & editing. GE-G: Conceptualization, Funding acquisition, Resources, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The publication of this review was financially supported by the Instituto Nacional de Perinatología (projects 2025-1-10 and 2024-1-43). This work is also a product derived from the grant CBF2023-2024-3091 Ciencia Básica y de Frontera awarded by SECIHTI to IC-A. CZ-S was supported by the program Estancias Posdoctorales por México from SECIHTI. JM-O is a doctoral student in the Posgrado en Ciencias en Inmunología, ENCB-IPN, and was supported by a SECIHTI fellowship (No. 001537).
Acknowledgments
All figures were created with BioRender.com, accessed on September 9, 2025.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Vacher CM, Tsompanidis A, Firestein MR, and Penn AA. Neuroactive steroid exposure impacts neurodevelopment: Comparison of human and rodent placental contribution. J Neuroendocrinol. (2025), 37:e13489. doi: 10.1111/JNE.13489
2. Chintala SM, Tateiwa H, Qian M, Xu Y, Amtashar F, Chen ZW, et al. Direct measurements of neurosteroid binding to specific sites on GABAA receptors. Br J Pharmacol. (2024) 181:4229–44. doi: 10.1111/BPH.16490
3. Belelli D and Lambert JJ. Neurosteroids: endogenous regulators of the GABAA receptor. Nat Rev Neurosci. (2005) 6:565–75. doi: 10.1038/nrn1703
4. Foradori CD, Weiser MJ, and Handa RJ. Non-genomic actions of androgens. Front Neuroendocrinol. (2008) 29:169–81. doi: 10.1016/J.YFRNE.2007.10.005
5. González-Orozco JC and Camacho-Arroyo I. Progesterone actions during central nervous system development. Front Neurosci. (2019) 13:503. doi: 10.3389/fnins.2019.00503
6. Bello-Alvarez C, Zamora-Sánchez CJ, and Camacho-Arroyo I. Rapid Actions of the Nuclear Progesterone Receptor through cSrc in Cancer. Cells. (2022) 11:1964. doi: 10.3390/cells11121964
7. Camacho-Arroyo I, Hansberg-Pastor V, Vázquez-Martínez ER, and Cerbón M. Mechanism of progesterone action in the brain. In: Hormones, Brain and Behavior (Amsterdam: Elsevier) (2017). 181–214.
8. López‐González R, Camacho‐Arroyo I, and Velasco I. Progesterone and 17β‐estradiol increase differentiation of mouse embryonic stem cells to motor neurons. IUBMB Life. (2011) 63:930–9. doi: 10.1002/iub.560
9. Brunton PJ, Russell JA, and Hirst JJ. Allopregnanolone in the brain: Protecting pregnancy and birth outcomes. Prog Neurobiol. (2014) 113:106–36. doi: 10.1016/J.PNEUROBIO.2013.08.005
10. Frye CA, Hirst JJ, Brunton PJ, and Russell JA. Neurosteroids for a successful pregnancy. Stress. (2011) 14:1–5. doi: 10.3109/10253890.2010.540050
11. Bethlehem RAI, Seidlitz J, White SR, Vogel JW, Anderson KM, Adamson C, et al. Brain charts for the human lifespan. Nature. (2022) 604:525–33. doi: 10.1038/s41586-022-04554-y
12. Jansakova K, Hill M, Celusakova H, Repiska G, Bicikova M, Macova L, et al. Steroidogenic pathway in girls diagnosed with autism spectrum disorders. PLoS One. (2024) 19:e0312933. doi: 10.1371/JOURNAL.PONE.0312933
13. Chew L, Sun KL, Sun W, Wang Z, Rajadas J, Flores RE, et al. Association of serum allopregnanolone with restricted and repetitive behaviors in adult males with autism. Psychoneuroendocrinology. (2021) 123:105039. doi: 10.1016/J.PSYNEUEN.2020.105039
14. Darbra S and Pallarès M. Developmental actions of neurosteroids in rodents: focus on allopregnanolone. Curr Opin Endocr Metab Res. (2022) 23:100317. doi: 10.1016/J.COEMR.2022.100317
15. Maguire JL and Mennerick S. Neurosteroids: mechanistic considerations and clinical prospects. Neuropsychopharmacology. (2023) 49:73–82. doi: 10.1038/s41386-023-01626-z
16. Saili KS, Zurlinden TJ, Schwab AJ, Silvin A, Baker NC, Hunter ES, et al. Blood-brain barrier development: Systems modeling and predictive toxicology. Birth Defects Res. (2017) 109:1680–710. doi: 10.1002/BDR2.1180
17. Vázquez M, Sobrevia L, Yañez MJ, and Leiva A. Human placental intracellular cholesterol transport: A focus on lysosomal and mitochondrial dysfunction and oxidative stress. Antioxidants. (2022) 11:500. doi: 10.3390/ANTIOX11030500
18. Kallol S and Albrecht C. Materno-fetal cholesterol transport during pregnancy. Biochem Soc Trans. (2020) 48:775–86. doi: 10.1042/BST20190129
19. Karahoda R, Kallol S, Groessl M, Ontsouka E, Anderle P, Fluck C, et al. Revisiting steroidogenic pathways in the human placenta and primary human trophoblast cells. Int J Mol Sci. (2021) 22:1–18. doi: 10.3390/ijms22041704
20. Cao J, Wang Y, Wang S, Shen Y, Li W, Wei Z, et al. Expression of key steroidogenic enzymes in human placenta and associated adverse pregnancy outcomes. Maternal-Fetal Med. (2023) 5:163–72. doi: 10.1097/FM9.0000000000000167
21. Chatuphonprasert W, Jarukamjorn K, and Ellinger I. Physiology and pathophysiology of steroid biosynthesis, transport and metabolism in the human placenta. Front Pharmacol. (2018) 9:1027. doi: 10.3389/fphar.2018.01027
22. Petrov AM, Kasimov MR, and Zefirov AL. Brain cholesterol metabolism and its defects: linkage to neurodegenerative diseases and synaptic dysfunction. Acta Naturae. (2016) 8:58. doi: 10.32607/20758251-2016-8-1-58-73
23. Burke KT, Colvin PL, Myatt L, Graf GA, Schroeder F, and Woollett LA. Transport of maternal cholesterol to the fetus is affected by maternal plasma cholesterol concentrations in the Golden Syrian hamster. J Lipid Res. (2009) 50:1146–55. doi: 10.1194/JLR.M800538
24. Saeed AA, Genové G, Li T, Lütjohann D, Olin M, Mast N, et al. Effects of a disrupted blood-brain barrier on cholesterol homeostasis in the brain. J Biol Chem. (2014) 289:23712–22. doi: 10.1074/jbc.M114.556159
25. Selvaraj V, Stocco DM, and Tu LN. Minireview: translocator protein (TSPO) and steroidogenesis: A reappraisal. Mol Endocrinol. (2015) 29:490–501. doi: 10.1210/ME.2015-1033
26. Lin YC, Cheung G, Porter E, and Papadopoulos V. The neurosteroid pregnenolone is synthesized by a mitochondrial P450 enzyme other than CYP11A1 in human glial cells. J Biol Chem. (2022) 298:102110. doi: 10.1016/j.jbc.2022.102110
27. Schonemann MD, Muench MO, Tee MK, Miller WL, and Mellon SH. Expression of P450c17 in the human fetal nervous system. Endocrinology. (2012) 153:2494–505. doi: 10.1210/EN.2011-1545
28. Legacki EL, Ball BA, Corbin CJ, Loux SC, Scoggin KE, Stanley SD, et al. Equine fetal adrenal, gonadal and placental steroidogenesis. Reproduction. (2017) 154:445–54. doi: 10.1530/REP-17-0239
29. Qaiser MZ, Dolman DEM, Begley DJ, Abbott NJ, Cazacu-Davidescu M, Corol DI, et al. Uptake and metabolism of sulphated steroids by the blood–brain barrier in the adult male rat. J Neurochem. (2017) 142:672–85. doi: 10.1111/JNC.14117
30. Schiffer L, Barnard L, Baranowski ES, Gilligan LC, Taylor AE, Arlt W, et al. Human steroid biosynthesis, metabolism and excretion are differentially reflected by serum and urine steroid metabolomes: A comprehensive review. J Steroid Biochem Mol Biol. (2019) 194:105439. doi: 10.1016/J.JSBMB.2019.105439
31. Schumacher M, Liere P, and Ghoumari A. Progesterone and fetal-neonatal neuroprotection. Best Pract Res Clin Obstet Gynaecol. (2020) 69:50–61. doi: 10.1016/J.BPOBGYN.2020.09.001
32. Kallen CB. Steroid hormone synthesis in pregnancy. Obstet Gynecol Clin North Am. (2004) 31:795–816. doi: 10.1016/J.OGC.2004.08.009
33. Milewich L, Shaw CE, Doody KM, Rainey WE, Mason JI, and Carr BR. 3β-hydroxysteroid dehydrogenase activity in glandular and extraglandular human fetal tissues. J Clin Endocrinol Metab. (1991) 73:1134–40. doi: 10.1210/JCEM-73-5-1134
34. Guennoun R. Progesterone in the brain: hormone, neurosteroid and neuroprotectant. Int J Mol Sci. (2020) 21:5271. doi: 10.3390/IJMS21155271
35. Ukena K, Kohchi C, and Tsutsui K. Expression and activity of 3β-hydroxysteroid dehydrogenase/Δ5-Δ4-isomerase in the rat purkinje neuron during neonatal life. Endocrinology. (1999) 140:805–13. doi: 10.1210/ENDO.140.2.6516
36. Compagnone NA and Mellon SH. Neurosteroids: biosynthesis and function of these novel neuromodulators. Front Neuroendocrinol. (2000) 21:1–56. doi: 10.1006/FRNE.1999.0188
37. Sakamoto H, Ukena K, and Tsutsui K. Effects of progesterone synthesized de novo in the developing purkinje cell on its dendritic growth and synaptogenesis. J Neurosci. (2001) 21:6221–32. doi: 10.1523/JNEUROSCI.21-16-06221.2001
38. Nguyen PN, Billiards SS, Walker DW, and Hirst JJ. Changes in 5α-pregnane steroids and neurosteroidogenic enzyme expression in the perinatal sheep. Pediatr Res. (2003) 53:956–64. doi: 10.1203/01.pdr.0000064905.64688.10
39. Diviccaro S, Cioffi L, Falvo E, Giatti S, and Melcangi RC. Allopregnanolone: An overview on its synthesis and effects. J Neuroendocrinol. (2022) 34:e12996. doi: 10.1111/JNE.12996
40. Hirst JJ, Kelleher MA, Walker DW, and Palliser HK. Neuroactive steroids in pregnancy: Key regulatory and protective roles in the foetal brain. J Steroid Biochem Mol Biol. (2014) 139:144–53. doi: 10.1016/J.JSBMB.2013.04.002
41. Kelleher MA, Palliser HK, Walker DW, and Hirst JJ. Sex-dependent effect of a low neurosteroid environment and intrauterine growth restriction on foetal Guinea pig brain development. J Endocrinol. (2011) 208:301–9. doi: 10.1677/JOE-10-0248
42. Diotel N, Charlier TD, Lefebvre d’Hellencourt C, Couret D, Trudeau VL, Nicolau JC, et al. Steroid transport, local synthesis, and signaling within the brain: roles in neurogenesis, neuroprotection, and sexual behaviors. Front Neurosci. (2018) 12:84. doi: 10.3389/fnins.2018.00084
43. Greaves RF, Wudy SA, Badoer E, Zacharin M, Hirst JJ, Quinn T, et al. A tale of two steroids: The importance of the androgens DHEA and DHEAS for early neurodevelopment. J Steroid Biochem Mol Biol. (2019) 188:77–85. doi: 10.1016/J.JSBMB.2018.12.007
44. Cumberland AL, Hirst JJ, Badoer E, Wudy SA, Greaves RF, Zacharin M, et al. The enigma of the adrenarche: identifying the early life mechanisms and possible role in postnatal brain development. Int J Mol Sci. (2021) 22:4296. doi: 10.3390/IJMS22094296
45. Munley KM, Rendon NM, and Demas GE. Neural androgen synthesis and aggression: insights from a seasonally breeding rodent. Front Endocrinol (Lausanne). (2018) 9:136. doi: 10.3389/fendo.2018.00136
46. Frye CA, DaCosta D, Lembo VF, and Walf AA. Advances in knowledge of androgens: how intentional and accidental neurosteroid changes inform us of their action and role. Curr Sex Health Rep. (2020) 12:209–20. doi: 10.1007/s11930-020-00276-2
47. Takeyama J, Suzuki T, Hirasawa G, Muramatsu Y, Nagura H, Iinuma K, et al. 17β-hydroxysteroid dehydrogenase type 1 and 2 expression in the human fetus. J Clin Endocrinol Metab. (2000) 85:410–6. doi: 10.1210/JCEM.85.1.6323
48. Spanic T, Fabjan T, and Majdic G. Expression levels of mRNA for neurosteroidogenic enzymes 17β-HSD, 5α-reductase, 3α-HSD and cytochrome P450 aromatase in the fetal wild type and SF-1 knockout mouse brain. Endocr Res. (2015) 40:44–8. doi: 10.3109/07435800.2014.933974
49. Lopez-Tello J and Sferruzzi-Perri AN. Characterization of placental endocrine function and fetal brain development in a mouse model of small for gestational age. Front Endocrinol (Lausanne). (2023) 14:1116770. doi: 10.3389/fendo.2023.1116770
50. Baud O and Berkane N. Hormonal changes associated with intra-uterine growth restriction: Impact on the developing brain and future neurodevelopment. Front Endocrinol (Lausanne). (2019) 10:179/BIBTEX. doi: 10.3389/FENDO.2019.00179/BIBTEX
51. Roselli CF. Brain aromatase: Roles in reproduction and neuroprotection. J Steroid Biochem Mol Biol. (2007) 106:143–50. doi: 10.1016/J.JSBMB.2007.05.014
52. Montelli S, Peruffo A, Zambenedetti P, Rossipal E, Giacomello M, Zatta P, et al. Expression of aromatase P450AROM in the human fetal and early postnatal cerebral cortex. Brain Res. (2012) 1475:11–8. doi: 10.1016/J.BRAINRES.2012.08.010
53. Shay DA, Vieira-Potter VJ, and Rosenfeld CS. Sexually dimorphic effects of aromatase on neurobehavioral responses. Front Mol Neurosci. (2018) 11:374. doi: 10.3389/fnmol.2018.00374
54. Azcoitia I, Mendez P, and Garcia-Segura LM. Aromatase in the human brain. Androgens. (2021) 2:189. doi: 10.1089/ANDRO.2021.0007
55. Salehzadeh M and Soma KK. Glucocorticoid production in the thymus and brain: Immunosteroids and neurosteroids. Brain Behav Immun Health. (2021) 18:100352. doi: 10.1016/J.BBIH.2021.100352
56. Howland MA, Sandman CA, and Glynn LM. Developmental origins of the human hypothalamic-pituitary-adrenal axis. Expert Rev Endocrinol Metab. (2017) 12:321–39. doi: 10.1080/17446651.2017.1356222
57. Pignatti E, du Toit T, and Flück CE. Development and function of the fetal adrenal. Rev Endocrine Metab Disord. (2022) 24:5–21. doi: 10.1007/S11154-022-09756-3
58. Pundir S, Gridneva Z, Pillai A, Thorstensen EB, Wall CR, Geddes DT, et al. Human milk glucocorticoid levels are associated with infant adiposity and head circumference over the first year of life. Front Nutr. (2020) 7:166/BIBTEX. doi: 10.3389/FNUT.2020.00166/BIBTEX
59. Mouton JC and Duckworth RA. Maternally derived hormones, neurosteroids and the development of behaviour. Proc R Soc B: Biol Sci. (2021) 288:20202467. doi: 10.1098/rspb.2020.2467
60. Scarpin KM, Graham JD, Mote PA, and Clarke CL. Membrane progesterone receptors (mPRs, PAQRs): review of structural and signaling characteristics. Cells. (2022) 11:1785. doi: 10.3390/CELLS11111785
61. Prossnitz ER and Barton M. The G protein-coupled oestrogen receptor GPER in health and disease: an update. Nat Rev Endocrinol. (2023) 19:407–24. doi: 10.1038/s41574-023-00822-7
62. Vallée M. Advances in steroid research from the pioneering neurosteroid concept to metabolomics: New insights into pregnenolone function. Front Neuroendocrinol. (2024) 72:101113. doi: 10.1016/J.YFRNE.2023.101113
63. Chen Y, Anderson MT, Payne N, Santori FR, and Ivanova NB. Nuclear receptors and the hidden language of the metabolome. Cells. (2024) 13:1284. doi: 10.3390/CELLS13151284
64. Oakley RH and Cidlowski JA. The biology of the glucocorticoid receptor: New signaling mechanisms in health and disease. J Allergy Clin Immunol. (2013) 132:1033–44. doi: 10.1016/j.jaci.2013.09.007
65. Levin ER and Pietras RJ. Estrogen receptors outside the nucleus in breast cancer. Breast Cancer Res Treat. (2008) 108:351–61. doi: 10.1007/s10549-007-9618-4
66. González-Arenas A, Peña-Ortiz M, Hansberg-Pastor V, Marquina-Sánchez B, Baranda-Ávila N, Nava-Castro K, et al. PKCα and PKCδ Activation regulates transcriptional activity and degradation of progesterone receptor in human astrocytoma cells. Endocrinology. (2015) 156:1010–22. doi: 10.1210/en.2014-1137
67. Samarasinghe RA, Witchel SF, and DeFranco DB. Cooperativity and complementarity: Synergies in non-classical and classical glucocorticoid signaling. Cell Cycle. (2012) 11:2819–27. doi: 10.4161/CC.21018
68. Clusan L, Ferrière F, Flouriot G, and Pakdel F. A basic review on estrogen receptor signaling pathways in breast cancer. Int J Mol Sci. (2023) 24:6834. doi: 10.3390/IJMS24076834
69. O’Brien KT, Mooney C, Lopez C, Pollastri G, and Shields DC. Prediction of polyproline II secondary structure propensity in proteins. R Soc Open Sci. (2020) 7:191239. doi: 10.1098/rsos.191239
70. Ruhs S, Nolze A, Hübschmann R, and Grossmann C. 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: Nongenomic effects via the mineralocorticoid receptor. J Endocrinol. (2017) 234:T107–24. doi: 10.1530/JOE-16-0659
71. Grossmann C, Almeida‐Prieto B, Nolze A, and Alvarez de la Rosa D. Structural and molecular determinants of mineralocorticoid receptor signalling. Br J Pharmacol. (2022) 179:3103–18. doi: 10.1111/bph.15746
72. Inoue T, Akahira JI, Takeyama J, Suzuki T, Darnel AD, Kaneko C, et al. Spatial and topological distribution of progesterone receptor A and B isoforms during human development. Mol Cell Endocrinol. (2001) 182:83–9. doi: 10.1016/S0303-7207(01)00549-4
73. González-Orozco JC, Moral-Morales AD, and Camacho-Arroyo I. Progesterone through progesterone receptor B isoform promotes rodent embryonic oligodendrogenesis. Cells. (2020) 9. doi: 10.3390/cells9040960
74. Díaz NF, Guerra-Arraiza C, Díaz-Martínez NE, Salazar P, Molina-Hernández A, Camacho-Arroyo I, et al. Changes in the content of estrogen α and progesterone receptors during differentiation of mouse embryonic stem cells to dopamine neurons. Brain Res Bull. (2007) 73:75–80. doi: 10.1016/J.BRAINRESBULL.2007.02.002
75. Olesen KM, Jessen HM, Auger CJ, and Auger AP. Dopaminergic activation of estrogen receptors in neonatal brain alters progestin receptor expression and juvenile social play behavior. Endocrinology. (2005) 146:3705–12. doi: 10.1210/EN.2005-0498
76. Camacho-Arroyo I, Guerra-Araiza C, and Cerbón MA. Progesterone receptor isoforms are differentially regulated by sex steroids in the rat forebrain. Neuroreport. (1998) 9:3993–6. doi: 10.1097/00001756-199812210-00001
77. Hong SH, Nah HY, Lee YJ, Lee JW, Park JH, Kim SJ, et al. Expression of estrogen receptor-α and -β, glucocorticoid receptor, and progesterone receptor genes in human embryonic stem cells and embryoid bodies. Mol Cells. (2004) 18:320–5. doi: 10.1016/S1016-8478(23)13119-0
78. Varshney M and Nalvarte I. Genes, gender, environment, and novel functions of estrogen receptor beta in the susceptibility to neurodevelopmental disorders. Brain Sci. (2017) 7:24. doi: 10.3390/BRAINSCI7030024
79. Tskitishvili E, Pequeux C, Munaut C, Viellevoye R, Nisolle M, Noël A, et al. Estrogen receptors and estetrol-dependent neuroprotective actions: a pilot study. J Endocrinol. (2017) 232:85–95. doi: 10.1530/JOE-16-0434
80. Cruceanu C, Dony L, Krontira AC, Fischer DS, Roeh S, Di Giaimo R, et al. Cell-type-specific impact of glucocorticoid receptor activation on the developing brain: A cerebral organoid study. Am J Psychiatry. (2022) 179:375–87. doi: 10.1176/appi.ajp.2021.21010095
81. Gillies GE, Virdee K, Pienaar I, Al-Zaid F, and Dalley JW. Enduring, sexually dimorphic impact of in utero exposure to elevated levels of glucocorticoids on midbrain dopaminergic populations. Brain Sci. (2016) 7:5. doi: 10.3390/BRAINSCI7010005
82. Correia-Branco A, Keating E, and Martel F. Maternal undernutrition and fetal developmental programming of obesity: the glucocorticoid connection. Reprod Sci. (2015) 22:138–45. doi: 10.1177/1933719114542012
83. Diaz R, Brown RW, and Seckl JR. Distinct ontogeny of glucocorticoid and mineralocorticoid receptor and 11β-hydroxysteroid dehydrogenase types I and II mRNAs in the fetal rat brain suggest a complex control of glucocorticoid actions. J Neurosci. (1998) 18:2570–80. doi: 10.1523/JNEUROSCI.18-07-02570.1998
84. Matthews SG. Dynamic changes in glucocorticoid and mineralocorticoid receptor mRNA in the developing Guinea pig brain. Dev Brain Res. (1998) 107:123–32. doi: 10.1016/S0165-3806(98)00008-X
85. Matthews SG. Antenatal glucocorticoids and programming of the developing CNS. Pediatr Res. (2000) 47:291–300. doi: 10.1203/00006450-200003000-00003
86. Quartier A, Chatrousse L, Redin C, Keime C, Haumesser N, Maglott-Roth A, et al. Genes and pathways regulated by androgens in human neural cells, potential candidates for the male excess in autism spectrum disorder. Biol Psychiatry. (2018) 84:239–52. doi: 10.1016/J.BIOPSYCH.2018.01.002/ATTACHMENT/2811F65C-39FC-48E0-9ACF-413842FCD536/MMC2.XLSX
87. Buonocore F, Suntharalingham JP, Ogunbiyi OK, Jones A, Moreno N, Niola P, et al. Transcriptomic sex differences in early human fetal brain development. Commun Biol. (2025) 8:1–14. doi: 10.1038/s42003-025-08070-3
88. Yue J, Wang XS, Feng B, Hu LN, Yang LK, Lu L, et al. Activation of G-Protein-Coupled Receptor 30 Protects Neurons against Excitotoxicity through Inhibiting Excessive Autophagy Induced by Glutamate. ACS Chem Neurosci. (2019) 10:4227–36. doi: 10.1021/ACSCHEMNEURO.9B00287/ASSET/IMAGES/MEDIUM/CN9B00287_0010.GIF
89. Song L, Chen H, Qiao D, Zhang B, Guo F, Zhang Y, et al. ZIP9 mediates the effects of DHT on learning, memory and hippocampal synaptic plasticity of male Tfm and APP/PS1 mice. Front Endocrinol (Lausanne). (2023) 14:1139874/BIBTEX. doi: 10.3389/FENDO.2023.1139874/BIBTEX
90. Kasubuchi M, Watanabe K, Hirano K, Inoue D, Li X, Terasawa K, et al. Membrane progesterone receptor beta (mPRβ/Paqr8) promotes progesterone-dependent neurite outgrowth in PC12 neuronal cells via non-G protein-coupled receptor (GPCR) signaling. Sci Rep. (2017) 7:1–12. doi: 10.1038/s41598-017-05423-9
91. Mustary UH, Maeno A, Rahaman MM, Ali MH, and Tokumoto T. Membrane progesterone receptor γ (paqr5b) is essential for the formation of neurons in the zebrafish olfactory rosette. Sci Rep. (2024) 14:1–15. doi: 10.1038/s41598-024-74674-0
92. Thomas P and Pang Y. Anti-apoptotic actions of allopregnanolone and ganaxolone mediated through membrane progesterone receptors (PAQRs) in neuronal cells. Front Endocrinol (Lausanne). (2020) 11:417/BIBTEX. doi: 10.3389/FENDO.2020.00417/BIBTEX
93. Deleon C, Pemberton K, Green M, Kalajdzic V, Rosato M, Xu F, et al. Novel GPER agonist, CITFA, increases neurite growth in rat embryonic (E18) hippocampal neurons. ACS Chem Neurosci. (2022) 13:1119–28. doi: 10.1021/ACSCHEMNEURO.1C00811/SUPPL_FILE/CN1C00811_SI_001.PDF
94. Pemberton K, Xu F, Rosato M, Dedert C, Deleon C, and Arnatt C. Differential effects of the G-protein-coupled estrogen receptor (GPER) on rat embryonic (E18) hippocampal and cortical neurons. eNeuro. (2022) 9. doi: 10.1523/ENEURO.0475-21.2022
95. Wang XS, Yue J, Hu LN, Tian Z, Zhang K, Yang L, et al. Activation of G protein-coupled receptor 30 protects neurons by regulating autophagy in astrocytes. Glia. (2020) 68:27–43. doi: 10.1002/GLIA.23697
96. Yawno T, Hirst JJ, Castillo-Melendez M, and Walker DW. Role of neurosteroids in regulating cell death and proliferation in the late gestation fetal brain. Neuroscience. (2009) 163:838–47. doi: 10.1016/j.neuroscience.2009.07.009
97. Yawno T, Yan EB, Walker DW, and Hirst JJ. Inhibition of neurosteroid synthesis increases asphyxia-induced brain injury in the late gestation fetal sheep. Neuroscience. (2007) 146:1726–33. doi: 10.1016/j.neuroscience.2007.03.023
98. Wang L, Covey DF, Akk G, and Evers AS. Neurosteroid modulation of GABAA receptor function by independent action at multiple specific binding sites. Curr Neuropharmacol. (2021) 20:886–90. doi: 10.2174/1570159X19666211202150041
99. Tuem KB and Atey TM. Neuroactive steroids: Receptor interactions and responses. Front Neurol. (2017) 8:442/BIBTEX. doi: 10.3389/FNEUR.2017.00442/BIBTEX
100. Wang JM, Johnston PB, Ball BG, and Brinton RD. The neurosteroid allopregnanolone promotes proliferation of rodent and human neural progenitor cells and regulates cell-cycle gene and protein expression. J Neurosci. (2005) 25:4706–18. doi: 10.1523/JNEUROSCI.4520-04.2005
101. Wang DD and Kriegstein AR. Defining the role of GABA in cortical development. J Physiol. (2009) 587:1873–9. doi: 10.1113/JPHYSIOL.2008.167635
102. Akerman CJ and Cline HT. Depolarizing GABAergic conductances regulate the balance of excitation to inhibition in the developing retinotectal circuit in vivo. J Neurosci. (2006) 26:5117–30. doi: 10.1523/JNEUROSCI.0319-06.2006
103. Owens DF, Liu X, and Kriegstein AR. Changing properties of GABA(A) receptor-mediated signaling during early neocortical development. J Neurophysiol. (1999) 82:570–83. doi: 10.1152/JN.1999.82.2.570/ASSET/IMAGES/LARGE/9K0890385009.JPEG
104. Espinosa‐Garcia C, Zeleke H, and Rojas A. Impact of stress on epilepsy: Focus on neuroinflammation— a mini review. Int J Mol Sci. (2021) 22:4061. doi: 10.3390/IJMS22084061/S1
105. Pan ZY, Zhao YH, Huang WH, Xiao ZZ, and Li ZQ. Effect of progesterone administration on the prognosis of patients with severe traumatic brain injury: a meta-analysis of randomized clinical trials. Drug Des Devel Ther. (2019) 13:265–73. doi: 10.2147/DDDT.S192633
106. Singh M, Krishnamoorthy VR, Kim S, Khurana S, and LaPorte HM. Brain-derived neuerotrophic factor and related mechanisms that mediate and influence progesterone-induced neuroprotection. Front Endocrinol (Lausanne). (2024) 15:1286066/BIBTEX. doi: 10.3389/FENDO.2024.1286066/BIBTEX
107. Frye CA, Manjarrez J, and Camacho-Arroyo I. Infusion of 3alpha,5alpha-THP to the pontine reticular formation attenuates PTZ-induced seizures. Brain Res. (2000) 881:98–102. doi: 10.1016/s0006-8993(00)02897–3
108. Atif F, Yousuf S, and Stein DG. Progesterone in the treatment of neonatal arterial ischemic stroke and acute seizures: Role of BDNF/TrkB signaling. Neuropharmacology. (2016) 107:317–28. doi: 10.1016/J.NEUROPHARM.2016.03.052
109. Barha CK, Ishrat T, Epp JR, Galea LAM, and Stein DG. Progesterone treatment normalizes the levels of cell proliferation and cell death in the dentate gyrus of the hippocampus after traumatic brain injury. Exp Neurol. (2011) 231:72–81. doi: 10.1016/J.EXPNEUROL.2011.05.016
110. Dixon KJ and Sherrard RM. Brain-derived neurotrophic factor induces post-lesion transcommissural growth of olivary axons that develop normal climbing fibers on mature Purkinje cells. Exp Neurol. (2006) 202:44–56. doi: 10.1016/J.EXPNEUROL.2006.05.010
111. Nicol MB, Hirst JJ, and Walker DW. Effect of finasteride on behavioural arousal and somatosensory evoked potentials in fetal sheep. Neurosci Lett. (2001) 306:13–6. doi: 10.1016/S0304-3940(01)01861-4
112. Wang J and Brinton R. Allopregnanolone-induced rise in intracellular calcium in embryonic hippocampal neurons parallels their proliferative potential. BMC Neurosci. (2008) 9:S11. doi: 10.1186/1471-2202-9-S2-S11
113. Wang T, Chen S, Mao Z, Shang Y, and Brinton RD. Allopregnanolone pleiotropic action in neurons and astrocytes: calcium signaling as a unifying mechanism. Front Endocrinol (Lausanne). (2023) 14:1286931/BIBTEX. doi: 10.3389/FENDO.2023.1286931/BIBTEX
114. Castelnovo LF, Bonalume V, Melfi S, Ballabio M, Colleoni D, and Magnaghi V. Schwann cell development, maturation and regeneration: A focus on classic and emerging intracellular signaling pathways. Neural Regener Res. (2017) 12:1013–23. doi: 10.4103/1673-5374.211172
115. Melfi S, Montt Guevara MM, Bonalume V, Ruscica M, Colciago A, Simoncini T, et al. Src and phospho-FAK kinases are activated by allopregnanolone promoting Schwann cell motility, morphology and myelination. J Neurochem. (2017) 141:165–78. doi: 10.1111/jnc.13951
116. Reddy DS, Mbilinyi RH, and Estes E. Preclinical and clinical pharmacology of brexanolone (allopregnanolone) for postpartum depression: a landmark journey from concept to clinic in neurosteroid replacement therapy. Psychopharmacology. (2023) 240:1841–63. doi: 10.1007/S00213-023-06427-2
117. Björväng RD, Walldén Y, Fransson E, Comasco E, Sundström-Poromaa I, and Skalkidou A. Mid-pregnancy allopregnanolone levels and trajectories of perinatal depressive symptoms. Psychoneuroendocrinology. (2024) 164:107009. doi: 10.1016/J.PSYNEUEN.2024.107009
118. Pinna G, Almeida FB, and Davis JM. Allopregnanolone in postpartum depression. Front Glob Womens Health. (2022) 3:823616/BIBTEX. doi: 10.3389/FGWH.2022.823616/BIBTEX
119. Vacher CM, Lacaille H, O’Reilly JJ, Salzbank J, Bakalar D, Sebaoui S, et al. Placental endocrine function shapes cerebellar development and social behavior. Nat Neurosci. (2021) 24:1392–401. doi: 10.1038/s41593-021-00896-4
120. Ghoumari AM, Ibanez C, El-Etr M, Leclerc P, Eychenne B, O’Malley BW, et al. Progesterone and its metabolites increase myelin basic protein expression in organotypic slice cultures of rat cerebellum. J Neurochem. (2003) 86:848–59. doi: 10.1046/J.1471-4159.2003.01881.X
121. Ghoumari AM, Baulieu EE, and Schumacher M. Progesterone increases oligodendroglial cell proliferation in rat cerebellar slice cultures. Neuroscience. (2005) 135:47–58. doi: 10.1016/j.neuroscience.2005.05.023
122. Liu G, Yan Y, Shi B, Huang J, Mu H, Li C, et al. Benefits of progesterone on brain immaturity and white matter injury induced by chronic hypoxia in neonatal rats. J Thorac Cardiovasc Surg. (2020) 160:e55–66. doi: 10.1016/j.jtcvs.2020.03.057
123. Nguyen TV. Developmental effects of androgens in the human brain. J Neuroendocrinol. (2018) 30:e12486. doi: 10.1111/JNE.12486
124. Ostatníková D, Lakatošová S, Babková J, Hodosy J, and Celec P. Testosterone and the brain: from cognition to autism. Physiol Res. (2020) 69:S403–S19. doi: 10.33549/physiolres.934592
125. Isgor C and Sengelaub DR. Effects of neonatal gonadal steroids on adult CA3 pyramidal neuron dendritic morphology and spatial memory in rats. J Neurobiol. (2003) 55:179–90. doi: 10.1002/NEU.10200
126. Hatanaka Y, Mukai H, Mitsuhashi K, Hojo Y, Murakami G, Komatsuzaki Y, et al. Androgen rapidly increases dendritic thorns of CA3 neurons in male rat hippocampus. Biochem Biophys Res Commun. (2009) 381:728–32. doi: 10.1016/J.BBRC.2009.02.130
127. Kight KE, McCarthy MM, and McCarthy MM. Androgens and the developing hippocampus. Biol Sex Dif. (2020) 11:1–14. doi: 10.1186/S13293-020-00307-6
128. Hatanaka Y, Hojo Y, Mukai H, Murakami G, Komatsuzaki Y, Kim J, et al. Rapid increase of spines by dihydrotestosterone and testosterone in hippocampal neurons: Dependence on synaptic androgen receptor and kinase networks. Brain Res. (2015) 1621:121–32. doi: 10.1016/J.BRAINRES.2014.12.011
129. Di Donato M, Bilancio A, Auricchio F, Castoria G, and Migliaccio A. Androgens and NGF mediate the neurite-outgrowth through inactivation of rhoA. Cells. (2023) 12:373. doi: 10.3390/CELLS12030373
130. Kosidou K, Dalman C, Widman L, Arver S, Lee BK, Magnusson C, et al. Maternal polycystic ovary syndrome and risk for attention-deficit/hyperactivity disorder in the offspring. Biol Psychiatry. (2017) 82:651–9. doi: 10.1016/J.BIOPSYCH.2016.09.022
131. Loomes R, Hull L, and Mandy WPL. What is the male-to-female ratio in autism spectrum disorder? A systematic review and meta-analysis. J Am Acad Child Adolesc Psychiatry. (2017) 56:466–74. doi: 10.1016/J.JAAC.2017.03.013
132. Cherskov A, Pohl A, Allison C, Zhang H, Payne RA, and Baron-Cohen S. Polycystic ovary syndrome and autism: A test of the prenatal sex steroid theory. Trans Psychiatry. (2018) 8:1–10. doi: 10.1038/s41398-018-0186-7
133. Lombardo MV, Auyeung B, Pramparo T, Quartier A, Courraud J, Holt RJ, et al. Sex-specific impact of prenatal androgens on social brain default mode subsystems. Mol Psychiatry. (2018) 25:2175–88. doi: 10.1038/s41380-018-0198-y
134. Denley MCS, Gatford NJF, Sellers KJ, and Srivastava DP. Estradiol and the development of the cerebral cortex: an unexpected role? Front Neurosci. (2018) 12:245. doi: 10.3389/fnins.2018.00245
135. Yague JG, Wang ACJ, Janssen WGM, Hof PR, Garcia-Segura LM, Azcoitia I, et al. Aromatase distribution in the monkey temporal neocortex and hippocampus. Brain Res. (2008) 1209:115–27. doi: 10.1016/J.BRAINRES.2008.02.061
136. Yague JG, Azcoitia I, DeFelipe J, Garcia-Segura LM, and Muñoz A. Aromatase expression in the normal and epileptic human hippocampus. Brain Res. (2010) 1315:41–52. doi: 10.1016/J.BRAINRES.2009.09.111
137. Konkle ATM and McCarthy MM. Developmental time course of estradiol, testosterone, and dihydrotestosterone levels in discrete regions of male and female rat brain. Endocrinology. (2011) 152:223–3510.1210/EN.2010–0607. doi: 10.1210/en.2010-0607
138. Wang L, Andersson S, Warner M, and Gustafsson J. Estrogen receptor (ER)beta knockout mice reveal a role for ERbeta in migration of cortical neurons in the developing brain. Proc Natl Acad Sci U.S.A. (2003) 100:703–8. doi: 10.1073/PNAS.242735799
139. Okada M, Makino A, Nakajima M, Okuyama S, Furukawa S, and Furukawa Y. Estrogen stimulates proliferation and differentiation of neural stem/progenitor cells through different signal transduction pathways. Int J Mol Sci. (2010) 11:4114–23. doi: 10.3390/IJMS11104114
140. Varshney MK, Inzunza J, Lupu D, Ganapathy V, Antonson P, Rüegg J, et al. Role of estrogen receptor beta in neural differentiation of mouse embryonic stem cells. Proc Natl Acad Sci U.S.A. (2017) 114:E10428–37. doi: 10.1073/PNAS.1714094114
141. Ruiz-Palmero I, Hernando M, Garcia-Segura LM, and Arevalo MA. G protein-coupled estrogen receptor is required for the neuritogenic mechanism of 17β-estradiol in developing hippocampal neurons. Mol Cell Endocrinol. (2013) 372:105–15. doi: 10.1016/J.MCE.2013.03.018
143. Tsukahara S and Morishita M. Sexually dimorphic formation of the preoptic area and the bed nucleus of the stria terminalis by neuroestrogens. Front Neurosci. (2020) 14:797. doi: 10.3389/fnins.2020.00797
144. Rinaudo M, Natale F, La Greca F, Spinelli M, Farsetti A, Paciello F, et al. Hippocampal estrogen signaling mediates sex differences in retroactive interference. Biomedicines. (2022) 10. doi: 10.3390/BIOMEDICINES10061387
145. Briceño-Pérez C, Reyna-Villasmil E, and Vigil-De-Gracia P. Antenatal corticosteroid therapy: Historical and scientific basis to improve preterm birth management. Eur J Obstet Gynecol Reprod Biol. (2019) 234:32–7. doi: 10.1016/J.EJOGRB.2018.12.025
146. Bandoli G, Jelliffe-Pawlowski LL, Feuer SK, Liang L, Oltman SP, Paynter R, et al. Second trimester serum cortisol and preterm birth: an analysis by timing and subtype. J Perinatol. (2018) 38:973–81. doi: 10.1038/S41372-018-0128-5
147. Nath N, Beltrano W, Haynes L, Dewey D, and Bray S. Long-term effects of preterm birth on children’s brain structure: an analysis of the adolescent brain cognitive development (ABCD) study. eNeuro. (2023) 10. doi: 10.1523/ENEURO.0196-22.2023
148. Manuela Z, Julien P, Elodie B, Olivier B, and Jérôme M. Glucocorticosteroids effects on brain development in the preterm infant: A role for microglia? Curr Neuropharmacol. (2021) 19:2188–204. doi: 10.2174/1570159X19666210517112913
149. Anacker C, Cattaneo A, Luoni A, Musaelyan K, Zunszain PA, Milanesi E, et al. Glucocorticoid-related molecular signaling pathways regulating hippocampal neurogenesis. Neuropsychopharmacology. (2013) 38:872–83. doi: 10.1038/NPP.2012.253
150. Odaka H, Adachi N, and Numakawa T. Impact of glucocorticoid on neurogenesis. Neural Regener Res. (2017) 12:1028–35. doi: 10.4103/1673-5374.211174
151. Wang G, Cao L, Li S, Zhang M, Li Y, Duan J, et al. Corticosterone Impairs Hippocampal Neurogenesis and Behaviors through p21-Mediated ROS Accumulation. Biomolecules. (2024) 14. doi: 10.3390/BIOM14030268
152. Kim DJ, Davis EP, Sandman CA, Sporns O, O’Donnell BF, Buss C, et al. Prenatal maternal cortisol has sex-specific associations with child brain network properties. Cereb Cortex. (2017) 27:5230–41. doi: 10.1093/CERCOR/BHW303
153. Graham AM, Rasmussen JM, Entringer S, Ben Ward E, Rudolph MD, Gilmore JH, et al. Maternal cortisol concentrations during pregnancy and sex-specific associations with neonatal amygdala connectivity and emerging internalizing behaviors. Biol Psychiatry. (2019) 85:172–81. doi: 10.1016/J.BIOPSYCH.2018.06.023
154. Byg LM, Bilenberg N, Glintborg D, Andersen MS, and Wesselhoeft R. The longitudinal relationship between prenatal maternal cortisol and preschool internalising problems. Psychoneuroendocrinology. (2024) 168. doi: 10.1016/J.PSYNEUEN.2024.107141
155. Edlow AG. Maternal obesity and neurodevelopmental and psychiatric disorders in offspring. Prenat Diagn. (2017) 37:95–110. doi: 10.1002/PD.4932
156. Cirulli F, Musillo C, and Berry A. Maternal obesity as a risk factor for brain development and mental health in the offspring. Neuroscience. (2020) 447:122–35. doi: 10.1016/j.neuroscience.2020.01.023
157. Valencia-Ortega J, Castillo-Santos A, Molerés-Orduña M, Solis-Paredes JM, Saucedo R, Estrada-Gutierrez G, et al. Influence of maternal adipokines on anthropometry, adiposity, and neurodevelopmental outcomes of the offspring. Int J Mol Sci. (2024) 25:11655. doi: 10.3390/IJMS252111655
158. Moreno‐Estrada LA, Valdez‐González J, Garza‐Silva A, Fernández‐Chau IF, Garza‐Rodríguez C, Reguera‐Hernández C, et al. Impact of overweight, obesity, and gestational weight gain on fetal somatometry, birth weight, and pregnancy complications. Int J Gynecol Obstetrics. (2025). doi: 10.1002/ijgo.70359
159. Shiadeh SMJ, Goretta F, Svedin P, Jansson T, Mallard C, and Ardalan M. Long-term impact of maternal obesity on the gliovascular unit and ephrin signaling in the hippocampus of adult offspring. J Neuroinflamm. (2024) 21:39. doi: 10.1186/s12974-024-03030-w
160. Sanders TR, Kim DW, Glendining KA, and Jasoni CL. Maternal obesity and IL-6 lead to aberrant developmental gene expression and deregulated neurite growth in the fetal arcuate nucleus. Endocrinology. (2014) 155:2566–77. doi: 10.1210/EN.2013-1968
161. Kim DW, Glendining KA, Grattan DR, and Jasoni CL. Maternal obesity leads to increased proliferation and numbers of astrocytes in the developing fetal and neonatal mouse hypothalamus. Int J Dev Neurosci. (2016) 53:18–25. doi: 10.1016/J.IJDEVNEU.2016.06.005
162. De Novais CO, Batista TH, Ribeiro ACAF, Vitor-Vieira F, Rojas VCT, Ferri BG, et al. Maternal overweight induced by reduced litter size impairs the behavioral neurodevelopment of offspring. Life Sci. (2021) 277:119611. doi: 10.1016/J.LFS.2021.119611
163. Suárez-Román G, Fernández-Romero T, Perera-Calderín AJ, Rodríguez-Sosa VM, Arranz C, and Hernandez SC. Pregestational obesity-induced embryopathy. Reprod Sci. (2016) 23:1250–7. doi: 10.1177/1933719116635279
164. Flores-Dorantes MT, Díaz-López YE, and Gutiérrez-Aguilar R. Environment and gene association with obesity and their impact on neurodegenerative and neurodevelopmental diseases. Front Neurosci. (2020) 14:863. doi: 10.3389/fnins.2020.00863
165. Mitchell AJ, Dunn GA, and Sullivan EL. The influence of maternal metabolic state and nutrition on offspring neurobehavioral development: A focus on preclinical models. Biol Psychiatry Cognit Neurosci Neuroimaging. (2022) 7:450–60. doi: 10.1016/J.BPSC.2021.11.014
166. Christifano DN, Taylor MK, Carlson SE, Colombo J, and Gustafson KM. Higher maternal weight is related to poorer fetal autonomic function. J Dev Orig Health Dis. (2021) 12:354–6. doi: 10.1017/S2040174420000653
167. Norr ME, Hect JL, Lenniger CJ, Van den Heuvel M, and Thomason ME. An examination of maternal prenatal BMI and human fetal brain development. J Child Psychol Psychiatry. (2021) 62:458–69. doi: 10.1111/JCPP.13301
168. Wang G, Li DK, Williams X, Ou L, Badger CM, Glasier RR, et al. Maternal obesity during pregnancy is associated with lower cortical thickness in the neonate brain. Am J Neuroradiol. (2021) 42:2238–44. doi: 10.3174/AJNR.A7316
169. Saros L, Lind A, Setänen S, Tertti K, Koivuniemi E, Ahtola A, et al. Maternal obesity, gestational diabetes mellitus, and diet in association with neurodevelopment of 2-year-old children. Pediatr Res. (2023) 94:280–9. doi: 10.1038/s41390-022-02455-4
170. Yeung EH, Sundaram R, Ghassabian A, Xie Y, and Louis GB. Parental obesity and early childhood development. Pediatrics. (2017) 139. doi: 10.1542/PEDS.2016-1459/60230
171. Sanchez CE, Barry C, Sabhlok A, Russell K, Majors A, Kollins SH, et al. Maternal pre-pregnancy obesity and child neurodevelopmental outcomes: a meta-analysis. Obes Rev. (2018) 19:464–84. doi: 10.1111/OBR.12643;CTYPE:STRING:JOURNAL
172. Widen EM, Kahn LG, Cirillo P, Cohn B, Kezios KL, and Factor-Litvak P. Prepregnancy overweight and obesity are associated with impaired child neurodevelopment. Matern Child Nutr. (2018) 14. doi: 10.1111/MCN.12481
173. Hartley E, McPhie S, Skouteris H, Fuller-Tyszkiewicz M, and Hill B. Psychosocial risk factors for excessive gestational weight gain: A systematic review. Women Birth. (2015) 28:e99–e109. doi: 10.1016/J.WOMBI.2015.04.004
174. Vanstone M, Kandasamy S, Giacomini M, Dejean D, and Mcdonald SD. Pregnant women’s perceptions of gestational weight gain: A systematic review and meta-synthesis of qualitative research. Matern Child Nutr. (2016) 13. doi: 10.1111/mcn.12374
175. Niebrzydowska-Tatus M, Pełech A, Rekowska AK, Satora M, Masiarz A, Kabała Z, et al. Recent insights and recommendations for preventing excessive gestational weight gain. J Clin Med. (2024) 13:1461. doi: 10.3390/JCM13051461
176. Kalli E, Potiris A, Domali E, Zikopoulos A, Kathopoulis N, Drakaki E, et al. The impact of excessive gestational weight gain on adverse perinatal outcomes: A systematic review. J Clin Med. (2025) 14:1197. doi: 10.3390/JCM14041197/S1
177. Valencia-Ortega J, Galicia-Hernández V, Castillo-Santos A, Molerés-Orduña M, Arceo-Cerna C, Perichart-Perera O, et al. Maternal organokines throughout pregnancy as predictors of neonatal anthropometric characteristics and adiposity. Front Endocrinol (Lausanne). (2024) 15:1423950. doi: 10.3389/fendo.2024.1423950
178. Valencia-Ortega J, Solis-Paredes JM, Saucedo R, Estrada-Gutierrez G, and Camacho-Arroyo I. Excessive pregestational weight and maternal obstetric complications: the role of adipokines. Int J Mol Sci. (2023) 24:14678. doi: 10.3390/IJMS241914678/S1
179. Ojeda DA, Hutton O, Hopkins R, Cagampang F, Smyth NR, Fleming TP, et al. Preimplantation or gestation/lactation high-fat diet alters adult offspring metabolism and neurogenesis. Brain Commun. (2023) 5. doi: 10.1093/BRAINCOMMS/FCAD093
180. Janthakhin Y, Rincel M, Costa AM, Darnaudéry M, and Ferreira G. Maternal high-fat diet leads to hippocampal and amygdala dendritic remodeling in adult male offspring. Psychoneuroendocrinology. (2017) 83:49–57. doi: 10.1016/J.PSYNEUEN.2017.05.003
181. Da Silva Rodrigues F, Jantsch J, de Farias Fraga G, Luiza de Camargo Milczarski V, Silva Dias V, Scheid C, et al. Cannabidiol improves maternal obesity-induced behavioral, neuroinflammatory and neurochemical dysfunctions in the juvenile offspring. Brain Behav Immun. (2024) 119:301–16. doi: 10.1016/J.BBI.2024.04.010
182. Horvatiček M, Perić M, Bečeheli I, Klasić M, Žutić M, Kesić M, et al. Maternal metabolic state and fetal sex and genotype modulate methylation of the serotonin receptor type 2A gene (HTR2A) in the human placenta. Biomedicines. (2022) 10:467. doi: 10.3390/BIOMEDICINES10020467
183. Ratsika A, Codagnone MG, Bastiaanssen TFS, Hoffmann Sarda FA, Lynch CMK, Ventura-Silva AP, et al. Maternal high-fat diet-induced microbiota changes are associated with alterations in embryonic brain metabolites and adolescent behaviour. Brain Behav Immun. (2024) 121:317–30. doi: 10.1016/J.BBI.2024.07.020
184. Pugh SJ, Richardson GA, Hutcheon JA, Himes KP, Brooks MM, Day NL, et al. Maternal obesity and excessive gestational weight gain are associated with components of child cognition. J Nutr. (2015) 145:2562–9. doi: 10.3945/JN.115.215525
185. Glendining KA, Fisher LC, and Jasoni CL. Maternal high fat diet alters offspring epigenetic regulators, amygdala glutamatergic profile and anxiety. Psychoneuroendocrinology. (2018) 96:132–41. doi: 10.1016/J.PSYNEUEN.2018.06.015
186. Rasic-Markovic A, Djuric E, Skrijelj D, Bjekic-Macut J, Ignjatovic Đ, Sutulovic N, et al. Neuroactive steroids in the neuroendocrine control of food intake, metabolism, and reproduction. Endocrine. (2024) 85:1050–7. doi: 10.1007/s12020-024-03755-x
187. Wang C, Xu Y, Wang C, and Xu Y. Mechanisms for sex differences in energy homeostasis. J Mol Endocrinol. (2019) 62:R129–43. doi: 10.1530/JME-18-0165
188. Mauvais-Jarvis F. Sex differences in energy metabolism: natural selection, mechanisms and consequences. Nat Rev Nephrol. (2023) 20:56–69. doi: 10.1038/s41581-023-00781-2
189. Denizli M, Capitano ML, and Kua KL. Maternal obesity and the impact of associated early-life inflammation on long-term health of offspring. Front Cell Infect Microbiol. (2022) 12:940937. doi: 10.3389/fcimb.2022.940937
190. Shook LL, Kislal S, and Edlow AG. Fetal brain and placental programming in maternal obesity: A review of human and animal model studies. Prenat Diagn. (2020) 40:1126–37. doi: 10.1002/PD.5724
191. Lombardo G, Mondelli V, Dazzan P, and Pariante CM. Sex hormones and immune system: A possible interplay in affective disorders? A systematic review. J Affect Disord. (2021) 290:1–14. doi: 10.1016/J.JAD.2021.04.035
192. Suh E, Cho AR, Haam JH, Gil M, Lee YK, and Kim YS. Relationship between serum cortisol, dehydroepiandrosterone sulfate (DHEAS) levels, and natural killer cell activity: A cross-sectional study. J Clin Med. (2023) 12:4027. doi: 10.3390/JCM12124027/S1
193. Deng H, Chen Y, Xing J, Zhang N, and Xu L. Systematic low-grade chronic inflammation and intrinsic mechanisms in polycystic ovary syndrome. Front Immunol. (2024) 15:1470283. doi: 10.3389/fimmu.2024.1470283
194. Balan I, Aurelian L, Schleicher R, Boero G, O’Buckley T, and Morrow AL. Neurosteroid allopregnanolone (3α,5α-THP) inhibits inflammatory signals induced by activated MyD88-dependent toll-like receptors. Transl Psychiatry. (2021) 11:1–11. doi: 10.1038/S41398-021-01266-1
195. Gardella B, Dominoni M, Scatigno AL, Cesari S, Fiandrino G, Orcesi S, et al. What is known about neuroplacentology in fetal growth restriction and in preterm infants: A narrative review of literature. Front Endocrinol (Lausanne). (2022) 13:936171. doi: 10.3389/fendo.2022.936171
196. Lacaille H, Vacher CM, and Penn AA. Preterm birth alters the maturation of the GABAergic system in the human prefrontal cortex. Front Mol Neurosci. (2022) 14:827370/BIBTEX. doi: 10.3389/FNMOL.2021.827370/BIBTEX
197. Lassance L, Haghiac M, Minium J, Catalano P, and Hauguel-de Mouzon S. Obesity-induced down-regulation of the mitochondrial translocator protein (TSPO) impairs placental steroid production. J Clin Endocrinol Metab. (2015) 100:E11–8. doi: 10.1210/JC.2014-2792
198. Yuan X, Ma Y, Wang J, Zhao Y, Zheng W, Yang R, et al. The influence of maternal prepregnancy weight and gestational weight gain on the umbilical cord blood metabolome: a case-control study (2024) 24:1–13. doi: 10.1186/s12884-024-06507-x
199. Sun Y, Zhu B, Meng X, Yin B, Wu K, Liu Y, et al. Effect of maternal body mass index on the steroid profile in women with gestational diabetes mellitus. Front Endocrinol (Lausanne). (2022) 13:999154. doi: 10.3389/fendo.2022.999154
200. Li C, Jenkins S, Considine MM, Cox LA, Gerow KG, Huber HF, et al. Effect of maternal obesity on fetal and postnatal baboon (Papio species) early life phenotype. J Med Primatol. (2019) 48:90–8. doi: 10.1111/JMP.12396
201. Jin S, Sun X, Liu Q, Liang H, Li C, Mao Z, et al. Steroid hormones in cord blood mediate the association between maternal prepregnancy BMI and birth weight. Obesity. (2019) 27:1338–46. doi: 10.1002/OBY.22524
202. Volqvartz T, Andersen HHB, Pedersen LH, and Larsen A. Obesity in pregnancy—Long-term effects on offspring hypothalamic-pituitary-adrenal axis and associations with placental cortisol metabolism: A systematic review. Eur J Neurosci. (2023) 58:4393–422. doi: 10.1111/EJN.16184
203. Johns EC, Denison FC, and Reynolds RM. The impact of maternal obesity in pregnancy on placental glucocorticoid and macronutrient transport and metabolism. Biochim Biophys Acta (BBA) - Mol Basis Dis. (2020) 1866:165374. doi: 10.1016/J.BBADIS.2018.12.025
204. Mayne GB, Dewitt PE, Ringham B, Warrener AG, Christians U, Dabelea D, et al. A nested case-control study of allopregnanolone and preterm birth in the healthy start cohort. J Endocr Soc. (2022) 7:1–9. doi: 10.1210/JENDSO/BVAC179
205. Hedström H, Bäckström T, Bixo M, Nyberg S, Wang M, Gideonsson I, et al. Women with polycystic ovary syndrome have elevated serum concentrations of and altered GABAA receptor sensitivity to allopregnanolone. Clin Endocrinol (Oxf). (2015) 83:643–50. doi: 10.1111/cen.12809
206. Holmberg E, Sjöstedt J, Malinina E, Johansson M, Turkmen S, Ragagnin G, et al. Allopregnanolone involvement in feeding regulation, overeating and obesity. Front Neuroendocrinol. (2018) 48:70–7. doi: 10.1016/J.YFRNE.2017.07.002
207. Holmberg E, Bäckström T, Johansson M, Löfgren M, and Haage D. Allopregnanolone induces a diurnally dependent hyperphagic effect and alters feeding latency and duration in male Wistar rats. Acta Physiol. (2013) 208:400–9. doi: 10.1111/APHA.12100
208. Holmberg E, Johansson M, Bäckström T, Löfgren M, and Haage D. Repeated allopregnanolone exposure induces weight gain in schedule fed rats on high fat diet. Physiol Behav. (2015) 140:1–7. doi: 10.1016/J.PHYSBEH.2014.12.012
209. Dichtel LE, Lawson EA, Schorr M, Meenaghan E, Paskal ML, Eddy KT, et al. Neuroactive steroids and affective symptoms in women across the weight spectrum. Neuropsychopharmacology. (2018) 43:1436–44. doi: 10.1038/NPP.2017.269
210. Grötsch MK and Ehlert U. Allopregnanolone and mood in the peripartum: a longitudinal assessment in healthy women. Front Behav Neurosci. (2024) 18:1499416. doi: 10.3389/fnbeh.2024.1499416
211. Hellgren C, Comasco E, Skalkidou A, and Sundström-Poromaa I. Allopregnanolone levels and depressive symptoms during pregnancy in relation to single nucleotide polymorphisms in the allopregnanolone synthesis pathway. Horm Behav. (2017) 94:106–13. doi: 10.1016/J.YHBEH.2017.06.008
212. Lundqvist A, Sandström H, and Bäckström T. The relationship between weight gain during pregnancy and allopregnanolone levels: A longitudinal study. Endocr Connect. (2017) 6:253–9. doi: 10.1530/EC-17-0046
213. Hirst JJ, Palliser HK, Pavy C, Shaw JC, and Moloney RA. Neurosteroid replacement approaches for improving outcomes after compromised pregnancies and preterm birth. Front Neuroendocrinol. (2025) 76:101169. doi: 10.1016/J.YFRNE.2024.101169
214. Shaw JC, Dyson RM, Palliser HK, Gray C, Berry MJ, and Hirst JJ. Neurosteroid replacement therapy using the allopregnanolone-analogue ganaxolone following preterm birth in male Guinea pigs. Pediatr Res. (2018) 85:86–96. doi: 10.1038/s41390-018-0185-7
215. Shaw JC, Dyson RM, Palliser HK, Sixtus RP, Barnes H, Pavy CL, et al. Examining neurosteroid-analogue therapy in the preterm neonate for promoting hippocampal neurodevelopment. Front Physiol. (2022) 13:871265. doi: 10.3389/fphys.2022.871265
216. Manuello J, Verdejo-Román J, Torres Espínola F, Escudero-Marín M, Catena A, Cauda F, et al. Influence of gestational diabetes and pregestational maternal BMI on the brain of six-year-old offspring. Pediatr Neurol. (2022) 133:55–62. doi: 10.1016/j.pediatrneurol.2022.05.005
217. Torres-Espínola FJ, Berglund SK, García-Valdés LM, Segura MT, Jerez A, Campos D, et al. Maternal obesity, overweight and gestational diabetes affect the offspring neurodevelopment at 6 and 18 months of age – A follow up from the PREOBE cohort. PLoS One. (2015) 10:e0133010. doi: 10.1371/JOURNAL.PONE.0133010
218. Buss C, Entringer S, Davis EP, Hobel CJ, Swanson JM, Wadhwa PD, et al. Impaired executive function mediates the association between maternal pre-pregnancy body mass index and child ADHD symptoms. PLoS One. (2012) 7:e37758. doi: 10.1371/JOURNAL.PONE.0037758
219. Dow C, Galera C, Charles M-A, and Heude B. Maternal pre-pregnancy BMI and offspring hyperactivity–inattention trajectories from 3 to 8 years in the EDEN birth cohort study. Eur Child Adolesc Psychiatry. (2023) 32:2057–65. doi: 10.1007/s00787-022-02047-x
220. Li M, Fallin MD, Riley A, Landa R, Walker SO, Silverstein M, et al. The association of maternal obesity and diabetes with autism and other developmental disabilities. Pediatrics. (2016) 137. doi: 10.1542/PEDS.2015-2206/52698
221. Guzzardi MA, Ederveen THA, Rizzo F, Weisz A, Collado MC, Muratori F, et al. Maternal pre-pregnancy overweight and neonatal gut bacterial colonization are associated with cognitive development and gut microbiota composition in pre-school-age offspring. Brain Behav Immun. (2022) 100:311–20. doi: 10.1016/J.BBI.2021.12.009
222. Chen S, Fan M, Lee BK, Dalman C, Karlsson H, and Gardner RM. Rates of maternal weight gain over the course of pregnancy and offspring risk of neurodevelopmental disorders. BMC Med. (2023) 21:1–16. doi: 10.1186/S12916-023-02799-6/FIGURES/4
223. Aubuchon-Endsley N, Morales M, Giudice C, Bublitz MH, Lester BM, Salisbury AL, et al. Maternal pre-pregnancy obesity and gestational weight gain influence neonatal neurobehaviour. Matern Child Nutr. (2017) 13. doi: 10.1111/MCN.12317
224. England-Mason G, Anderson A, Bell RC, Subhan FB, Field CJ, Letourneau N, et al. Maternal pre-pregnancy BMI and gestational weight gain are associated with preschool children’s neuropsychological outcomes in the APrON cohort. Children. (2023) 10:1849. doi: 10.3390/CHILDREN10121849/S1
225. Pugh S, Hutcheon J, Richardson G, Brooks M, Himes K, Day N, et al. Gestational weight gain, prepregnancy body mass index and offspring attention‐deficit hyperactivity disorder symptoms and behaviour at age 10. BJOG. (2016) 123:2094–103. doi: 10.1111/1471-0528.13909
226. Perea V, Simó-Servat A, Quirós C, Alonso-Carril N, Valverde M, Urquizu X, et al. Role of excessive weight gain during gestation in the risk of ADHD in offspring of women with gestational diabetes. J Clin Endocrinol Metab. (2022) 107:e4203–11. doi: 10.1210/CLINEM/DGAC483
227. Lee P, Tse L, László K, Wei D, Yu Y, and Li J. Association of maternal gestational weight gain with intellectual developmental disorder in the offspring: a nationwide follow‐up study in Sweden. BJOG. (2022) 129:540–9. doi: 10.1111/1471-0528.16887
228. Gardner RM, Lee BK, Magnusson C, Rai D, Frisell T, Karlsson H, et al. Maternal body mass index during early pregnancy, gestational weight gain, and risk of autism spectrum disorders: Results from a Swedish total population and discordant sibling study. Int J Epidemiol. (2015) 44:870–83. doi: 10.1093/IJE/DYV081
229. Aline H, Hallit S, Ellkhatib R, Abi Haidar S, Hajj Moussa Lteif F, Hajj L, et al. Saudi Arabia. Ms. L. Hajj and Ms. Moudawar are with SESOBEL in Ain El Rihani. Innov Clin Neurosci. (2022) 19:22–7.
Keywords: neuroplacentology, neurosteroids, pregnancy, fetal programming, allopregnanolone, steroid metabolism, body mass index, maternal obesity
Citation: Zamora-Sánchez CJ, González-Orozco JC, Mendoza-Ortega J, Villegas-Soto ML, Camacho-Arroyo I and Estrada-Gutierrez G (2025) Neuroactive steroids in the programming of neurodevelopment: implications of maternal obesity. Front. Endocrinol. 16:1703103. doi: 10.3389/fendo.2025.1703103
Received: 10 September 2025; Accepted: 27 October 2025;
Published: 17 November 2025.
Edited by:
Rachida Guennoun, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Roberto Cosimo Melcangi, University of Milan, ItalyClaire-Marie Vacher, Columbia University, United States
Copyright © 2025 Zamora-Sánchez, González-Orozco, Mendoza-Ortega, Villegas-Soto, Camacho-Arroyo and Estrada-Gutierrez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guadalupe Estrada-Gutierrez, Z3Blc3RyYWRAZ21haWwuY29t